Characteristics and influencing factors of Bowen ratio variation in typical terrestrial ecosystems in China
Song-Yu HUANG1,2, Xin JIA,1,2,3,*, Jia-Jia ZHENG1,2, Rui-Zhi YANG1,2, Yu MU1,2, He-Di YUAN11School of Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China 2Yanchi Ecology Research Station of MU US Desert, Beijing 100083, China 3Key Laboratory of State Forestry and Grassland Administration on Soil and Water Conservation, Beijing Forestry University, Beijing 100083, China
National Natural Science Foundation of China(31670708) National Natural Science Foundation of China(31670710) National Natural Science Foundation of China(31901366) Fundamental Research Funds for the Central Universities(2015ZCQ-SB-02)
Abstract Aims Bowen ratio (β) is an important parameter in land-surface processes. It affects the energy exchange between the surface and the atmosphere. This paper used integrated analyses to investigate the spatial variability and influencing factors of β. Methods We collected the published literature on the measurement of surface energy balance by the Eddy Covariance (EC) method carried out in different ecosystem types in China, constructed the database of β and meteorological environment factors and analyzed the difference of β among ecosystems, the spatial variation characteristics of β and its influencing factors. Important findings (1) The variation of β follows a lognormal distribution. The average β in all ecosystems was 0.95 ± 0.64, the coefficient of variation of β was 67%, the skewness was 1.58, and the kurtosis was 3.07. The shrub ecosystem has the highest mean value (1.26) and the wetland ecosystem has the lowest (0.49). (2) β is significantly different among ecosystems: β of shrub ecosystems is significantly higher than those in grassland, forest and wetland ecosystems, and β of croplands is between grassland ecosystems and forest with wetland ecosystems. (3) β increases with increasing latitude and does not change with longitude and altitude. For every 1° increase in latitude,β increases by 0.038. (4) β decreases with increase in mean annual precipitation (MAP), mean annual temperature (MAT), net radiation (Rn), precipitation of the studied year (PPT), mean temperature of the studied year (Ta), and leaf area index (LAI). (5) There are significant differences in the response of β to biotic and abiotic factors in different ecosystems: β of grassland, forest and shrub ecosystems are sensitive to changes in biotic and abiotic factors, while β of croplands and wetland ecosystems have no correlations with biotic and abiotic factors. (6) MAPand Rn are the direct factors influencing β. MAT affects βindirectly by affecting MAP, Rn and LAI. LAI affects β indirectly by affectingRn. Our results indicate significant effect of the interaction between vegetation types and climatic factors on β. The most important factor affecting energy distribution is precipitation, and the regulation of leaf area on energy distribution is not significant. Keywords:Bowen ratio;latent heat flux;sensible heat flux;ecosystems;energy partitioning;climate;Leaf Area Index
PDF (1578KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 黄松宇, 贾昕, 郑甲佳, 杨睿智, 牟钰, 袁和第. 中国典型陆地生态系统波文比特征及影响因素. 植物生态学报, 2021, 45(2): 119-130. DOI: 10.17521/cjpe.2019.0301 HUANG Song-Yu, JIA Xin, ZHENG Jia-Jia, YANG Rui-Zhi, MU Yu, YUAN He-Di. Characteristics and influencing factors of Bowen ratio variation in typical terrestrial ecosystems in China. Chinese Journal of Plant Ecology, 2021, 45(2): 119-130. DOI: 10.17521/cjpe.2019.0301
地表与大气之间的能量平衡影响生物地球化学循环(如碳、氮、水循环)过程, 并调控区域气候(原文文等, 2015; 张翔等, 2017)。作为表面能量平衡的重要组分, 显热通量(H)和潜热通量(LE)通过对温度、水分传输和植被的生长发育的调控驱动区域气候的变化(Chen et al., 2009; 岳平等, 2011)。波文比(β)是H和LE的比值(H/LE), 它能够反映地表能量交换特征(Bowen, 1926; 陈云飞等, 2013; 夏露和张强, 2014; 葛骏等, 2016), 在众多陆面过程相关分析的研究中有所涉及, 在天气预报和气候预测中也有诸多应用(Jo et al., 2004; 何学敏等, 2017)。了解β的变化特征及影响因素, 有助于深入理解陆-气相互作用模式, 准确预测区域或全球尺度的水文循环、能量平衡和气候变化(Chen et al., 2009; Tang et al., 2014; Morwal et al., 2017)。
已有研究表明β受到地理(纬度)、气候(温度、降水、净辐射)和生物(叶面积指数)等因素的影响(Matsumoto et al., 2008; Launiainen, 2010; 张强等, 2014; Launiainen et al., 2016; 王佩等, 2017)。但是, 就目前的研究来说, 地理因素和气候因素与β的关系存在较大不确定性。例如, 孙成(2014)在对亚热带毛竹(Phyllostachys edulis)林能量通量的研究中认为, β的变异受降水量及其他因素的综合影响; Grünwald和Bernhofer (2007)在对德国温带云杉(Picea abies)林长达10年的通量观测研究中也指出, 年β主要受到年降水量的控制。而很多****在研究之后发现, 年降水量和年β之间并不存在显著的相关关系(Ryu et al., 2008; Liu et al., 2009; Tang et al., 2014; Yue et al., 2018)。在空间尺度上, β的研究相对较少, 目前仅张强等(2014)在黄土高原地表能量通量的研究中表明, β随降水的增加而降低, 但没有明确β随温度的变化。Burakowski等(2017)在研究β对美国东部生态系统能量平衡的作用时提及, β会根据生物群落和纬度而变化, 但是并未明确给出β是如何在生物群落间和纬度上变化的。另外, 由于目前的研究大部分仅在单个的生态系统内进行, 因此生物和非生物因素对β变异的影响以及β在不同生态系统间的差异都不明确, 需要进一步的研究。
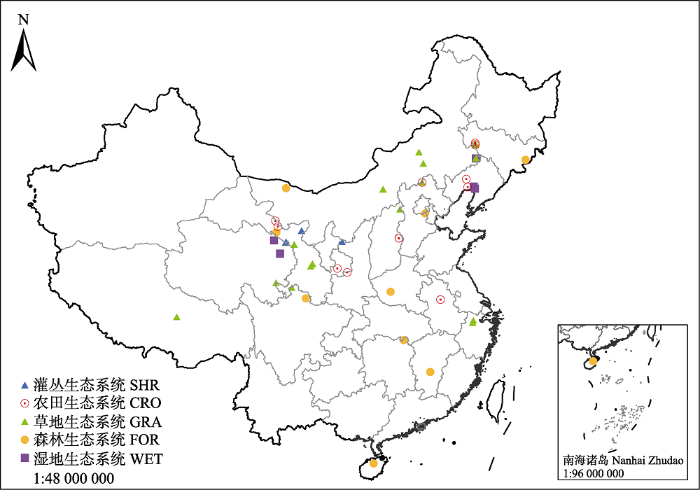
Fig. 1Distribution of the data resources of Bowen ratio in typical terrestrial ecosystems in China. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems.
1.2 数据分析
数据分析时发现, FOR中的离群值(Zhu et al., 2014)会影响分析结果, 即该点会改变FOR中β的整体趋势: 该点使本无显著相关关系的β和Alt显著相关, 使本该有显著相关关系的β和Lat、Lon、MAP、PPT之间无显著相关关系。Zhu等(2014)的研究地处青藏高原, 海拔较高(2 800 m), 地面获得的太阳辐射和净辐射都非常高(高于97%的站点数据), 但是该处降水量却相对较少(平均年降水量435 mm, 研究期当年降水量428 mm)。水分亏缺导致蒸散发不足, 因此用于驱动蒸散发的能量较少, 大部分净辐射分配给显热通量, 导致该地区波文比值异常地高。在数据分析中去掉了此异常值点。
Fig. 3Boxing diagram of the Bowen ratio (β) of different ecosystem types. Lowercase letters indicate the results of multiple comparisons. The same letter indicates that the difference of β between ecosystems is not significant (p > 0.5), while different letters indicate significant differences in β between ecosystems (p < 0.5). n is the sample size. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems.
Fig. 4Relationships between the Bowen ratio (β), sensible heat flux (H), latent heat flux (LE) and latitude, longitude and altitude of terrestrial ecosystems in China. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems.
Table 2 表2 表2波文比(β)与地理、气候及生物因素在不同生态系统中的线性回归结果 Table 2Linear regressions between Bowen ratio (β) and geographical, climatic and biological factors in different ecosystems
影响因素 Influencing factor
生态系统类型 Ecosystem type
回归方程 Regression equation
样本数 Sample size
R2
纬度 Lat
FOR
β= 0.016Lat - 0.02
12
0.57**
经度 Lon
FOR
β= 0.014Lon -1.13
12
0.32*
年降水量 MAP
GRA
β= -0.00076MAP + 1.46
22
0.22*
FOR
β= -0.00021MAP + 0.73
12
0.36*
SHR
β= -0.0046MAP + 3.73
4
0.91*
当年降水量 PPT
GRA
β= -0.00082PPT + 1.43
15
0.41**
FOR
β= -0.00025PPT + 0.77
10
0.42*
年平均气温 MAT
FOR
β= -0.017MAT + 0.74
11
0.45*
当年平均气温 Ta
FOR
β= -0.020Ta + 0.77
8
0.48*
净辐射 Rn
GRA
β= -000063Rn + 2.54
20
0.34***
SHR
β= -0.0021Rn + 7.09
4
0.97*
叶面积指数 LAI
GRA
β= -0.20LAI + 1.45
12
0.34*
FOR, 森林生态系统; GRA, 草地生态系统; SHR, 灌丛生态系统。*,p < 0.05; **, p < 0.01; ***, p < 0.001。 LAI, Leaf Area Index;Lat, latitude; Lon, longitude; MAP, mean annual precipitation; MAT, mean annual air temperature; PPT, annual precipitation of the studied year; Rn, net radiation; Ta, mean annual air temperature of the studied year. FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Table 3 表3 表3显热通量(H)与地理、气候及生物因素在不同生态系统中的线性回归结果 Table 3Linear regressions between sensible heat flux (H) and geographical, climatic and biological factors in different ecosystems
影响因素 Influencing factor
生态系统类型 Ecosystem type
回归方程 Regression equation
样本数 Sample size
R2
纬度 Lat
SHR
H =-373.96Lat + 15579.69
4
0.90*
当年降水量 PPT
FOR
H =-0.20PPT + 1015.80
10
0.41*
净辐射 Rn
SHR
H= 0.36Rn + 521.99
4
0.99**
FOR, 森林生态系统; SHR, 灌丛生态系统。*,p < 0.05; **, p < 0.01。 Lat, latitude; PPT, annual precipitation of the studied year; Rn, net radiation. FOR, forest ecosystems; SHR, shrubland ecosystems. *, p < 0.05; **, p < 0.01.
Table 4 表4 表4潜热通量(LE)与地理、气候及生物因素在不同生态系统中的线性回归结果 Table 4Linear regressions between latent heat flux (LE) and geographical, climatic and biological factors in different ecosystems
影响因素 Influencing factor
生态系统类型 Ecosystem type
回归方程 Regression equation
样本数 Sample size
R2
纬度 Lat
GRA
LE= -45.40Lat +2767.46
22
0.36**
CRO
LE= -57.21Lat + 3424.40
12
0.48**
FOR
LE= -40.80Lat + 3032.86
12
0.62**
经度 Lon
CRO
LE= -21.36Lon -3604.25
12
0.30*
FOR
LE =-36.92Lon +5944.80
12
0.39*
海拔 Alt
SHR
LE= 0.43Alt -116.08
4
0.89*
年降水量 MAP
GRA
LE =0.64MAP +683.66
22
0.34**
CRO
LE =1.18MAP +500.76
10
0.52*
FOR
LE= 0.50MAP + 1203.54
12
0.36*
SHR
LE= 2.19MAP - 78.90
4
0.97**
当年降水量 PPT
GRA
LE= 0.65PPT + 725.23
15
0.44**
年平均气温 MAT
FOR
LE =45.30MAT +1122.30
11
0.61**
SHR
LE= -80.99MAT + 1200.72
4
0.90*
当年平均气温 Ta
FOR
LE= 54.34Ta + 1024.19
8
0.62*
净辐射 Rn
GRA
LE= 0.47Rn - 53.39
20
0.51***
FOR
LE= 0.58Rn + 39.06
12
0.61**
WET
LE= 0.69Rn - 441.67
5
0.77*
CRO, 农田生态系统; FOR, 森林生态系统; GRA, 草地生态系统; SHR, 灌丛生态系统; WET, 湿地生态系统。*,p < 0.05; **, p < 0.01; ***, p < 0.001。 Lat, latitude; Lon, longitude; MAP, mean annual precipitation; MAT, mean annual air temperature; PPT, annual precipitation of the studied year; Rn, net radiation; Ta, mean annual air temperature of the studied year. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
Fig. 5Relationships between Bowen ratio (β) and mean annual precipitation (MAP), annual precipitation of the studied year (PPT), mean annual air temperature (MAT), mean annual air temperature of the studied year (Ta), net radiation (Rn) and Leaf Area Index (LAI) of China's terrestrial ecosystems. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems.
Fig. 6Relationships between sensible heat flux (β) and mean annual precipitation (MAP), annual precipitation of the studied year (PPT), mean annual air temperature (MAT), mean annual air temperature of the studied year (Ta), net radiation (Rn) and Leaf Area Index (LAI) of China's terrestrial ecosystems. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems.
Fig. 7The relationships between latent heat flux (LE) and mean annual precipitation (MAP), annual precipitation of the studied year (PPT), mean annual air temperature (MAT), mean annual air temperature of the studied year (Ta), net radiation (Rn) and Leaf Area Index (LAI) of China's terrestrial ecosystems. CRO, cropland ecosystems; FOR, forest ecosystems; GRA, grassland ecosystems; SHR, shrubland ecosystems; WET, wetland ecosystems.
气候因素对β的影响因生态系统中的植被类型而异。生态系统植被的类型和MAP对β存在交互影响(MAP, F1,41 = 46.77, p < 0.01; 植被类型, F5,41 = 8.18, p < 0.01; MAP ×植被类型, F4,41 = 6.35, p < 0.01)。草地、灌木和森林生态系统中 β随MAP的增加而显著降低, 农田和湿地生态系统中β与MAP无相关关系(表2)。其中, 灌木生态系统MAP对β影响的解释度最高(R2= 0.91), β对的变化MAP也最为敏感(MAP每增加100 mm, 灌木生态系统β降低0.46, 而草地和森林生态系统分别降低0.076和0.021)。相似地, 生态系统类型和PPT对β也存在交互效应(PPT, F1,30= 31.17, p < 0.01; 植被类型, F4,30 = 13.82, p < 0.01; PPT ×植被类型, F3,30 = 3.72, p < 0.05)。 PPT增加时, 草地和森林生态系统中β显著降低, 但PPT对农田、灌木和湿地生态系统β没有影响(表2)。相较于森林生态系统, 草地生态系统β对PPT的变化更敏感。
不同生态系统类型植被类型中的β对MAT的响应存在明显差异(MAT, F1,40 = 13.58, p < 0.01; 植被类型, F4,40 = 6.30, p < 0.01; MAT ×植被类型, F4,40 = 4.62, p < 0.01)。森林生态系统 β随MAT的增加而降低, 而其他生态系统中β不随MAT的变化而变化(表2)。而Ta, 仅森林生态系统中β对其变化有显著响应(表2), Ta和生态系统类型对β的影响不存在交互作用(p = 0.85)。
不同生态系统的植被类型和Rn对β同样存在交互影响(Rn, F1,41 = 54.31, p < 0.01; 植被类型, F4,41 = 11.59, p < 0.01; Rn ×植被类型, F4,41 = 5.61, p < 0.01): 草地和灌木生态系统 β随Rn增加而降低, 农田、森林和湿地生态系统β与Rn无显著相关关系(表2)。其中, 灌木生态系统中Rn对β变化的解释度高达97%, β对Rn变化的敏感性也较高(Rn每增加100 MJ·m-2·a-1, 草地生态系统β降低0.063, 而灌木生态系统β降低0.21)。
Fig. 8Path diagrams for the effects of mean annual precipitation (MAP), mean annual air temperature (MAT), net radiation (Rn) and Leaf Area Index (LAI). The statistical results of the structural equation model (SEM) are χ2 = 1.75, df = 2, p = 0.093, RMSEA = 0.038. Solid and dashed arrows represent positive and negative correlations, respectively. The figure for each arrow is the standardized path coefficient. The bold numbers are the explained variance (R2) of LAI, Rn and β, respectively.
BoisierJP, deNoblet-Ducoudré N, PitmanAJ, CruzFT, DelireC, vanden Hurk BJJM, vander Molen MK, MüllerC, VoldoireA (2012). Attributing the impacts of land-cover changes in temperate regions on surface temperature and heat fluxes to specific causes: results from the first LUCID set of simulations Journal of Gerphysical Research, 117,D12116. DOI: 10.1029/2011JD017106. DOI:10.1029/2011JD017106 [本文引用: 1]
BonanGB (2008). Forests and climate change: forcings, feedbacks, and the climate benefits of forests Science, 320,1444-1449. [本文引用: 2]
BowenIS (1926). The ratio of heat losses by conduction and by evaporation from any water surface Physical Review, 27,779-787. DOI:10.1103/PhysRev.27.779URL [本文引用: 1]
BurakowskiE, TawfikA, OuimetteA, LepineL, NovickD, OllingerS, ZarzyckiC, BonanG (2017). The role of surface roughness, albedo, and Bowen ratio on ecosystem energy balance in the Eastern United States Agricultural and Forest Meteorology, 249,367-376. DOI:10.1016/j.agrformet.2017.11.030URL [本文引用: 1]
ChenL, DirmeyerPA (2016). Adapting observationally based metrics of biogeophysical feedbacks from land cover/land use change to climate modeling Environmental Research Letters, 11,034002. DOI: 10.1088/1748-9326/11/3/034002. DOI:10.1088/1748-9326/11/3/034002URL [本文引用: 1]
ChenSP, ChenJQ, LinGH, ZhangWL, MiaoHX, WeiL, HuangJH, HanXG (2009). Energy balance and partition in Inner Mongolia steppe ecosystems with different land use types Agricultural and Forest Meteorology, 149,1800-1809. DOI:10.1016/j.agrformet.2009.06.009URL [本文引用: 2]
ChenYF, JiangH, ZhouGM, SunC, ChenJ (2013). Energy flux and energy balance closure of intensively managed Lei bamboo forest ecosystem Chinese Journal of Applied Ecology, 24,1063-1069. [本文引用: 1]
GeJ, YuY, LiZC, XieJ, LiuC, ZanBL (2016). Impacts of freeze/thaw processes on land surface energy fluxes in the permafrost region of Qinghai-Xizang Plateau Plateau Meteorology, 35,608-620. [本文引用: 1]
GrünwaldT, BernhoferC (2007). A decade of carbon, water and energy flux measurements of an old spruce forest at the Anchor Station Tharandt Tellus, 59(B),387-396. [本文引用: 1]
HanB, LüSH, LiRQ, WangX, ZhaoL, ZhaoCL, WangDY, MengXH (2017). Global land surface climate analysis based on the calculation of a modified Bowen ratio Advances in Atmospheric Sciences, 34,663-678. DOI:10.1007/s00376-016-6175-yURL [本文引用: 2]
HeXM, QinL, LüGH, YangJJ, GongYM, YangXD (2017). Surface energy balance of an arid desert wetland in Ebinur Lake basin, Xinjiang, China Chinese Journal of Ecology, 36,309-317. [本文引用: 1]
JiaX, ZhaTS, WuB, ZhangYQ, QinSG, ChenGP, FengW, Kellom?kiS, PeltolaH (2016). Energy partitioning over a semi-arid shrubland in northern China Hydrological Processes, 30,972-985. DOI:10.1002/hyp.v30.6URL [本文引用: 3]
JoYH, YanXH, PanJY, LiuWT, HeMX (2004). Sensible and latent heat flux in the tropical Pacific from satellite multi-sensor data Remote Sensing of Environment, 90,166-177. DOI:10.1016/j.rse.2003.12.003URL [本文引用: 1]
JuangJY, KatulG, SiqueiraM, StoyP, NovickK (2007). Separating the effects of albedo from eco-physiological changes on surface temperature along a successional chronosequence in the southeastern United States Geophysical Research Letters, 34,L21408. DOI: 10.1029/2007GL031296. DOI:10.1029/2007GL031296URL [本文引用: 1]
LauniainenS (2010). Seasonal and inter-annual variability of energy exchange above a boreal Scots pine forest Biogeosciences, 7,3921-3940. DOI:10.5194/bg-7-3921-2010URL [本文引用: 1]
LauniainenS, KatulGG, KolariP, LindrothA, LohilaA, AurelaM, VarlagindA, GrelleA, VesalaT (2016). Do the energy fluxes and surface conductance of boreal coniferous forests in Europe scale with leaf area Global Change Biology, 22,4096-4113. DOI:10.1111/gcb.13497PMID:27614117 [本文引用: 2] Earth observing systems are now routinely used to infer leaf area index (LAI) given its significance in spatial aggregation of land surface fluxes. Whether LAI is an appropriate scaling parameter for daytime growing season energy budget, surface conductance (G ), water- and light-use efficiency and surface-atmosphere coupling of European boreal coniferous forests was explored using eddy-covariance (EC) energy and CO fluxes. The observed scaling relations were then explained using a biophysical multilayer soil-vegetation-atmosphere transfer model as well as by a bulk G representation. The LAI variations significantly alter radiation regime, within-canopy microclimate, sink/source distributions of CO , H O and heat, and forest floor fluxes. The contribution of forest floor to ecosystem-scale energy exchange is shown to decrease asymptotically with increased LAI, as expected. Compared with other energy budget components, dry-canopy evapotranspiration (ET) was reasonably 'conservative' over the studied LAI range 0.5-7.0?m m . Both ET and G experienced a minimum in the LAI range 1-2?m m caused by opposing nonproportional response of stomatally controlled transpiration and 'free' forest floor evaporation to changes in canopy density. The young forests had strongest coupling with the atmosphere while stomatal control of energy partitioning was strongest in relatively sparse (LAI ~2?m m ) pine stands growing on mineral soils. The data analysis and model results suggest that LAI may be an effective scaling parameter for net radiation and its partitioning but only in sparse stands (LAI <3?m m ). This finding emphasizes the significance of stand-replacing disturbances on the controls of surface energy exchange. In denser forests, any LAI dependency varies with physiological traits such as light-saturated water-use efficiency. The results suggest that incorporating species traits and site conditions are necessary when LAI is used in upscaling energy exchanges of boreal coniferous forests.? 2016 John Wiley & Sons Ltd.
LiY, ZhaoM, MotesharreiS, MuQ, KalnayE, LiS (2015). Local cooling and warming effects of forests based on satellite observations Nature Communications, 6,6603. DOI: 10.1038/ncomms7603. DOI:10.1038/ncomms7603URL [本文引用: 1]
LiuS, LiSG, YuGR, SunXM, ZhangLM, HuZM, LiYN, ZhangXZ (2009). Surface energy exchanges above two grassland ecosystems on the Qinghai-Tibetan Plateau Biogeosciences Discussions, 6,9161-9192. [本文引用: 1]
MatsumotoK, OhtaT, NakaiT, KuwadaT, DaikokuK, IidaS, YabukiH, KononovAV, MolenMK, KodamaY, MaximovTC, DolmanAJ, HattoriS (2008). Energy consumption and evapotranspiration at several boreal and temperate forests in the Far East Agricultural and Forest Meteorology, 148,1978-1989. DOI:10.1016/j.agrformet.2008.09.008URL [本文引用: 1]
MorwalSB, NarkhedkarSG, PadmakumariB, MaheskumarRS, DeshpandeCG, KulkarniJP (2017). Intra-seasonal and inter-annual variability of Bowen ratio over rain- shadow region of north peninsular India Theoretical and Applied Climatology, 128,835-844. DOI:10.1007/s00704-016-1745-6URL [本文引用: 1]
RyuY, BaldocchiDD, MaS, HehnT (2008). Interannual variability of evapotranspiration and energy exchange over an annual grassland in California Journal of Geophysical Research, 113,D09104. DOI: 10.1029/2007JD009263. DOI:10.1029/2007JD009263 [本文引用: 1]
SunC (2014). The Study on the CO2 Flux and Energy Balance Variations in a Phyllostachys edulis Forest Ecosystem PhD dissertation, Zhejiang A&F University, Lin'an, Zhejiang. [本文引用: 1]
TangYK, WenXF, SunXM, WangHM (2014). Interannual variation of the Bowen ratio in a subtropical coniferous plantation in southeast China, 2003-2012 PLOS ONE, 9, e88267. DOI: 10.1371/journal.pone.0088267. DOI:10.1371/journal.pone.0088267 [本文引用: 3]
WangP, MaQS, WangJQ, HuangJY, LiW, ZhangCC (2017). Comparison of evapotranspiration and Bowen ratio method by eddy correlation and Bowen ratio system in a temperate grassland Acta Agrestia Sinica, 25,453-459. [本文引用: 2]
YuanWW, TongXJ, ZhangJS, MengP, LiJ, ZhengN (2015). Characteristics of energy balance of a mixed plantation in the Xiaolangdi area in the growing season Acta Ecologica Sinica, 35,4492-4499. [本文引用: 1]
YueP, ZhangQ, YangY, ZhangL, ZhangHL, HaoXC, SunXY (2018). Seasonal and inter-annual variability of the Bowen smith ratio over a semi-arid grassland in the Chinese Loess Plateau Agricultural and Forest Meteorology, 252,99-108. DOI:10.1016/j.agrformet.2018.01.006URL [本文引用: 1]
ZhangQ, ZhangL, HuangJ, ZhangLY, WangWY, ShaS (2014). Spatial distribution of surface energy fluxes over the Loess Plateau in China and its relationship with climate and the environment Science China: Earth Sciences (Chinese Version), 44,2062-2076. [本文引用: 8]
ZhangX, LiuXQ, ZhangLF, NiuB, ZhaoL, GuS (2017). Energy balance of an artificial grassland in the Three-River Source Region of the Qinghai-Tibet Plateau Acta Ecologica Sinica, 37,4973-4983. [本文引用: 1]
ZhuGf, LuL, SuYH, WangXF, CuiX, MaJZ, HeJH, ZhangK, LiCB (2014). Energy flux partitioning and evapotranspiration in a sub-alpine spruce forest ecosystem Hydrological Processes, 28,5093-5104. DOI:10.1002/hyp.9995URL [本文引用: 3]
YuGR, ZhuXJ, FuYL, HeHL, WangQF, WenXF, LiXR, ZhangLM, ZhangL, SuW, LiSG, SunXM, ZhangYP, ZhangJH, YanJH, et al. (2013). Spatial patterns and climate drivers of carbon fluxes in terrestrial ecosystems of China Global Change Biology, 19,798-810. DOI:10.1111/gcb.2013.19.issue-3URL
Attributing the impacts of land-cover changes in temperate regions on surface temperature and heat fluxes to specific causes: results from the first LUCID set of simulations 1 2012
... 不同生态系统类型间的β的差异表现为灌木林的β高于草地高于森林和湿地, 而农田介于草地和森林与湿地之间, 这种差异来源于生态系统间冠层结构和水分条件的不同.冠层结构能够通过影响LAI、地面粗糙度和地表反照率影响能量分配(Bonan, 2008; Boisier et al., 2012), 与冠层结构简单的生态系统相比, 冠层结构复杂的生态系统具有更强的蒸散发和冷却效应(即更低的β)(Juang et al., 2007; Kueppers et al., 2007; Li et al., 2015).而水分直接影响生态系统的蒸散发(即LE).我们的研究结果也证实了这一点(图5).相对其他生态系统来说, 森林和湿地具有更复杂的冠层结构和更充足的水分条件, 因此β显著较低.灌木生态系统由于分布在干旱地区, 水分稀缺, 且植被覆盖度低, 冠层结构简单, 因此β均值最高.草地生态系统的水分条件介于灌木生态系统与森林和湿地生态系统之间, 其植被覆盖度远低于森林和湿地生态系统.由于数据样本点问题, 草地和灌木生态系统难以进行LAI的比较, 但从结构方程的结果可知, 水分的影响远高于叶面积指数对β的影响, 所以在无法比较植被覆盖时, 认为水分对β的影响占主导.因此, 草地生态系统的β均值高于森林和湿地生态系统而低于灌木生态系统.而与草地和森林、湿地差异都不显著的农田生态系统, 其β变异主要受人类活动影响. ...
Forests and climate change: forcings, feedbacks, and the climate benefits of forests 2 2008
... 不同生态系统类型间的β的差异表现为灌木林的β高于草地高于森林和湿地, 而农田介于草地和森林与湿地之间, 这种差异来源于生态系统间冠层结构和水分条件的不同.冠层结构能够通过影响LAI、地面粗糙度和地表反照率影响能量分配(Bonan, 2008; Boisier et al., 2012), 与冠层结构简单的生态系统相比, 冠层结构复杂的生态系统具有更强的蒸散发和冷却效应(即更低的β)(Juang et al., 2007; Kueppers et al., 2007; Li et al., 2015).而水分直接影响生态系统的蒸散发(即LE).我们的研究结果也证实了这一点(图5).相对其他生态系统来说, 森林和湿地具有更复杂的冠层结构和更充足的水分条件, 因此β显著较低.灌木生态系统由于分布在干旱地区, 水分稀缺, 且植被覆盖度低, 冠层结构简单, 因此β均值最高.草地生态系统的水分条件介于灌木生态系统与森林和湿地生态系统之间, 其植被覆盖度远低于森林和湿地生态系统.由于数据样本点问题, 草地和灌木生态系统难以进行LAI的比较, 但从结构方程的结果可知, 水分的影响远高于叶面积指数对β的影响, 所以在无法比较植被覆盖时, 认为水分对β的影响占主导.因此, 草地生态系统的β均值高于森林和湿地生态系统而低于灌木生态系统.而与草地和森林、湿地差异都不显著的农田生态系统, 其β变异主要受人类活动影响. ...
Sensible and latent heat flux in the tropical Pacific from satellite multi-sensor data 1 2004
... 地表与大气之间的能量平衡影响生物地球化学循环(如碳、氮、水循环)过程, 并调控区域气候(原文文等, 2015; 张翔等, 2017).作为表面能量平衡的重要组分, 显热通量(H)和潜热通量(LE)通过对温度、水分传输和植被的生长发育的调控驱动区域气候的变化(Chen et al., 2009; 岳平等, 2011).波文比(β)是H和LE的比值(H/LE), 它能够反映地表能量交换特征(Bowen, 1926; 陈云飞等, 2013; 夏露和张强, 2014; 葛骏等, 2016), 在众多陆面过程相关分析的研究中有所涉及, 在天气预报和气候预测中也有诸多应用(Jo et al., 2004; 何学敏等, 2017).了解β的变化特征及影响因素, 有助于深入理解陆-气相互作用模式, 准确预测区域或全球尺度的水文循环、能量平衡和气候变化(Chen et al., 2009; Tang et al., 2014; Morwal et al., 2017). ...
Separating the effects of albedo from eco-physiological changes on surface temperature along a successional chronosequence in the southeastern United States 1 2007
... 不同生态系统类型间的β的差异表现为灌木林的β高于草地高于森林和湿地, 而农田介于草地和森林与湿地之间, 这种差异来源于生态系统间冠层结构和水分条件的不同.冠层结构能够通过影响LAI、地面粗糙度和地表反照率影响能量分配(Bonan, 2008; Boisier et al., 2012), 与冠层结构简单的生态系统相比, 冠层结构复杂的生态系统具有更强的蒸散发和冷却效应(即更低的β)(Juang et al., 2007; Kueppers et al., 2007; Li et al., 2015).而水分直接影响生态系统的蒸散发(即LE).我们的研究结果也证实了这一点(图5).相对其他生态系统来说, 森林和湿地具有更复杂的冠层结构和更充足的水分条件, 因此β显著较低.灌木生态系统由于分布在干旱地区, 水分稀缺, 且植被覆盖度低, 冠层结构简单, 因此β均值最高.草地生态系统的水分条件介于灌木生态系统与森林和湿地生态系统之间, 其植被覆盖度远低于森林和湿地生态系统.由于数据样本点问题, 草地和灌木生态系统难以进行LAI的比较, 但从结构方程的结果可知, 水分的影响远高于叶面积指数对β的影响, 所以在无法比较植被覆盖时, 认为水分对β的影响占主导.因此, 草地生态系统的β均值高于森林和湿地生态系统而低于灌木生态系统.而与草地和森林、湿地差异都不显著的农田生态系统, 其β变异主要受人类活动影响. ...
 ,1,2,3,*, 郑甲佳1,2, 杨睿智1,2, 牟钰1,2, 袁和第1
,1,2,3,*, 郑甲佳1,2, 杨睿智1,2, 牟钰1,2, 袁和第1

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT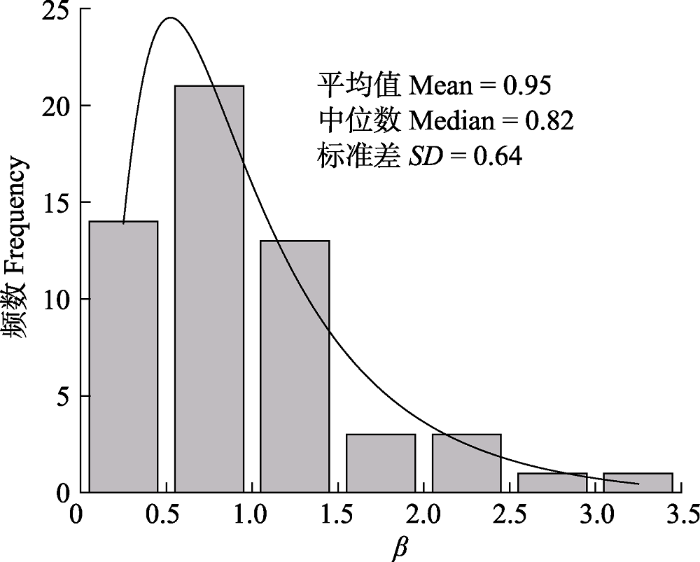 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT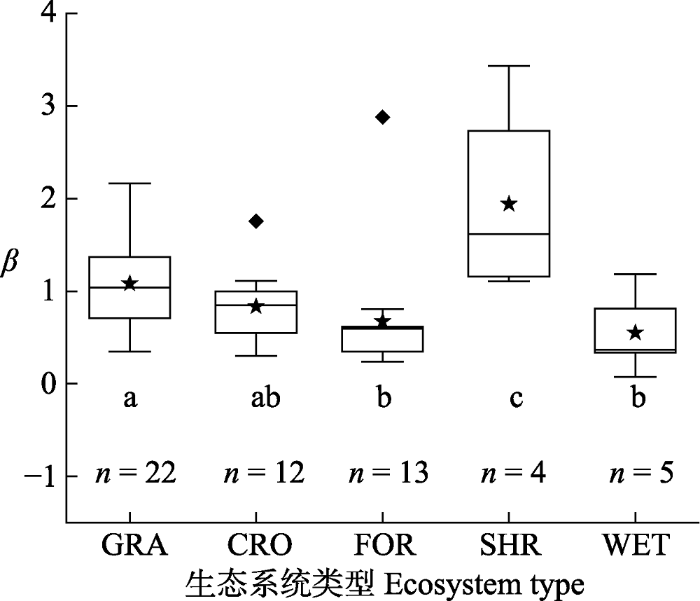 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT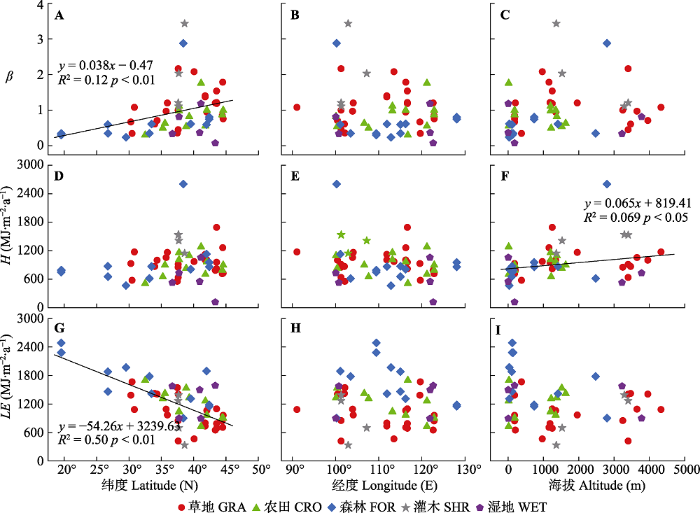 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT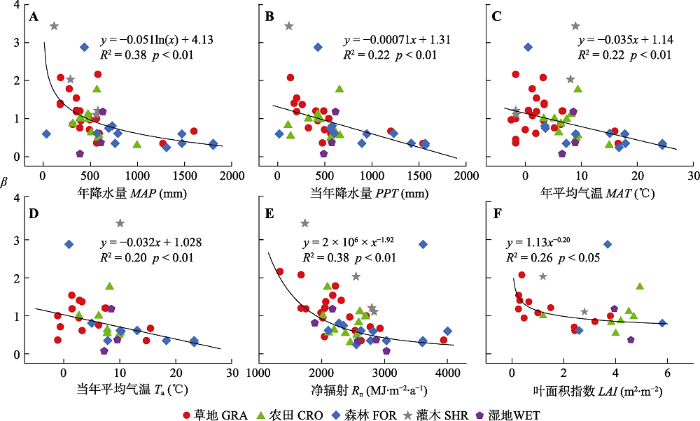 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT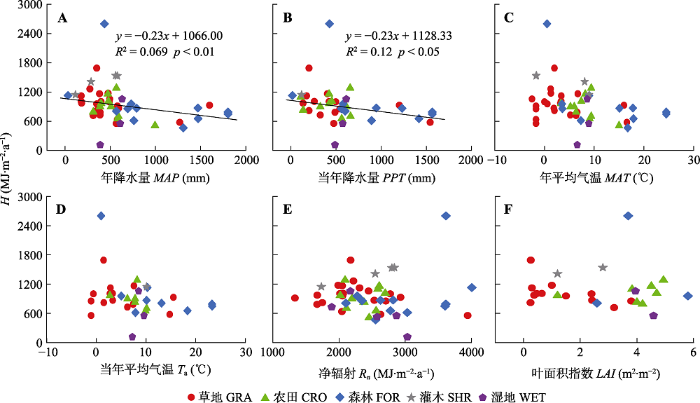 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT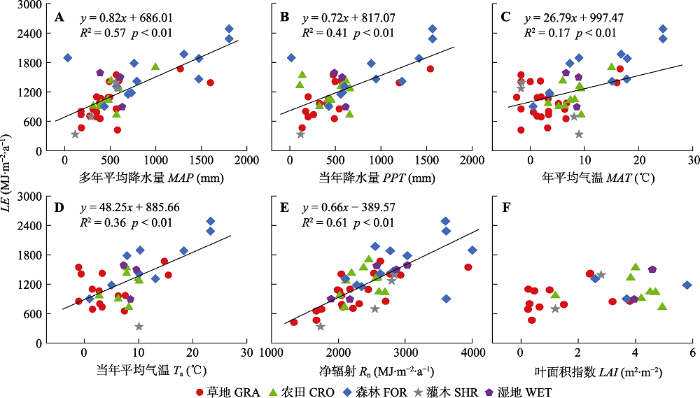 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT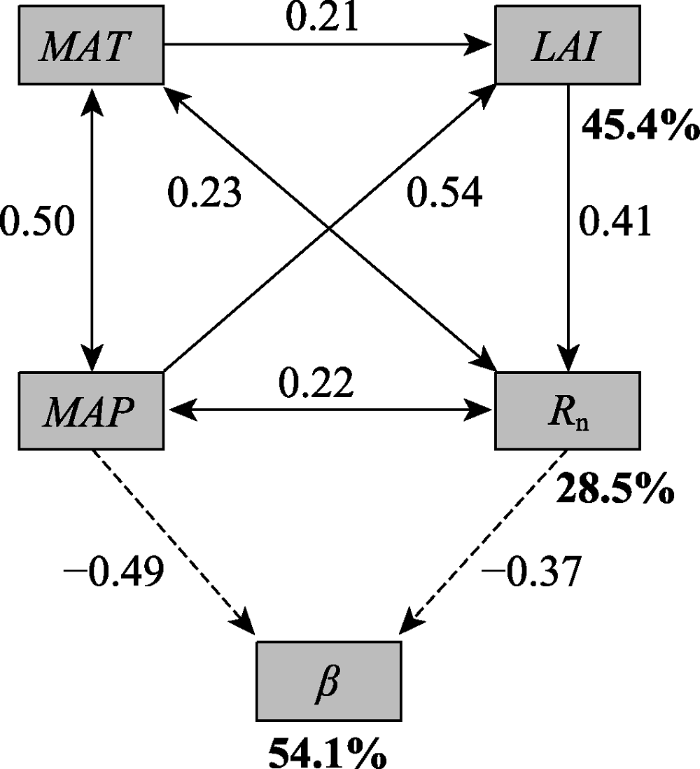 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}