Effects of long-term simulated acid rain on soil microbial community structure in a monsoon evergreen broad-leaved forest in southern China
Yuan-Liu HU1,2, Guo-Yin CHEN1,2, Jing-Wen CHEN1,2, Lian-Wei SUN1,2, Jian-Ling LI1, Ning DOU1, De-Qiang ZHANG1, Qi DENG,1,*1South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Aims Soil microorganisms are an important component of terrestrial ecosystems and play a critical role in regulating multiple ecological processes such as nutrient acquisition, carbon cycle, and soil formation, especially in the tropical forests where soils are highly weathered with poor nutrients. The objective of this study was to examine the response of soil microbial community under long-term simulated acid rain (SAR) and investigate the most important factors influencing microbial community structure. Methods Based on a long-term (10-year) field SAR experiment, we investigate the response of soil microbial community structure to soil acidification in the south subtropical monsoon evergreen broad-leaved forest of Dinghushan National Nature Reserve. Four levels of SAR treatments were set by adding the following amount of H+: 0 (CK), 9.6, 32 and 96 mol·hm-2·a-1. Important findings 1) The SAR treatment significantly reduced the pH value of soil (i.e., increased soil acidification). 2) Soil acidification did not significantly influence microbial carbon (C) content, but changed microbial nitrogen (N) and phosphorus (P) contents, leading to significant increases in microbial C:P and N:P in topsoil (0-10 cm). This result indicated that soil acidification might aggravate microbial P limitation. 3) Soil acidification also altered the microbial community structure and significantly increased the fungal/bacterial ratio in the subsoil (10-20 cm). Further analysis showed that soil pH and available P content were the most important factors affecting the soil microbial communities under the SAR treatment. Keywords:soil acidification;microbial stoichiometry;fungal:bacterial;phosphorus limitation
PDF (552KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 胡苑柳, 陈国茵, 陈静文, 孙连伟, 李健陵, 窦宁, 张德强, 邓琦. 模拟酸沉降对南亚热带季风常绿阔叶林土壤微生物群落结构的长期影响. 植物生态学报, 2021, 45(3): 298-308. DOI: 10.17521/cjpe.2020.0217 HU Yuan-Liu, CHEN Guo-Yin, CHEN Jing-Wen, SUN Lian-Wei, LI Jian-Ling, DOU Ning, ZHANG De-Qiang, DENG Qi. Effects of long-term simulated acid rain on soil microbial community structure in a monsoon evergreen broad-leaved forest in southern China. Chinese Journal of Plant Ecology, 2021, 45(3): 298-308. DOI: 10.17521/cjpe.2020.0217
自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988)。研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013)。目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5)。长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019)。农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015)。与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019)。当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018)。土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013)。
土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999)。例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016)。微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007)。此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018)。相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013)。细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017)。真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力。例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998)。随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004)。以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008)。因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019)。
土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007)。酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016)。绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019)。例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003)。此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007)。基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力。
热带森林是陆地生态系统中最重要的碳库, 储存了近1/4的陆地碳, 并且每年能够吸收大量的CO2 (Bonan, 2008)。但与此同时, 热带森林土壤较好的水热条件使得地质发育较为成熟, 土壤中养分十分贫瘠(Vitousek et al., 2010)。而长期酸雨导致土壤酸化加剧, 养分流失愈发严重(Qiao et al., 2015), 微生物活性和群落组成发生变化, 从而进一步影响了土壤中养分的循环, 进而导致植物多样性和生产力受到限制(Bouwman et al., 2002)。目前, 由于受到野外条件的限制, 热带森林生态系统中微生物对土壤酸化响应的研究主要集中在比较自然状态下pH不同的土壤其微生物结构的差异(Hu et al., 2019), 以及通过室内培养实验探究各个类群微生物对土壤响应的差异(Liu et al., 2020), 而缺乏直接的长期原位控制实验。本研究通过模拟酸沉降实验探究土壤理化性质的变化如何影响土壤微生物活性及其群落结构, 有利于揭示土壤养分有效性的变化机制, 为未来气候变化背景下森林生态系统生物多样性稳定及其可持续经营提供科学依据。
1 材料和方法
1.1 研究区概况
研究地点位于中国广东省肇庆市鼎湖山国家自然保护区内(112.50°-112.55° E, 23.15°-23.18° N), 处于南亚热带湿润森林地区, 属于典型季风气候, 雨热同期, 年降水量为1 929 mm, 年平均气温21 ℃。干湿季节交替明显, 其中4-9月为雨季, 降水量占年降水量的80%, 10月至次年3月为旱季(Lv et al., 2014)。近20多年来, 研究区域降水pH均低于4.5, 酸雨成分主要为硫酸和硝酸混合, 土壤pH逐年下降(Jiang et al., 2018)。
Table 2 表2 表2模拟酸沉降对南亚热带季风常绿阔叶林土壤理化性质的影响 Table 2Soil properties in the four simulated acid rain treatments in a monsoon evergreen broad-leaved forest in southern China
土层 Soil layer (cm)
处理 Treatment
土壤pH Soil pH
含水量 SWC (%)
有机碳含量 SOC content (g·kg-1)
氨态氮含量 NH4+-N content (mg·kg-1)
硝态氮含量 NO3--N content (mg·kg-1)
总氮含量 TN content (g·kg-1)
有效磷含量 AP content (mg·kg-1)
总磷含量 TP content (g·kg-1)
总氮:总磷 TC:TN
0-10
CK
4.18a (0.03)
43.76a (1.20)
35.25 (1.22)
1.36 (0.16)
15.85a (1.02)
2.37 (0.01)
1.68a (0.12)
0.24a (0.01)
14.93bc (0.25)
T1
4.05b (0.03)
41.68a (1.14)
37.71 (2.60)
1.16 (0.25)
10.94ab (0.90)
2.66 (0.13)
1.47ab (0.15)
0.19b (0.01)
14.11c (0.40)
T2
3.89c (0.06)
35.42b (1.15)
40.43 (3.90)
1.68 (0.24)
9.82b (1.12)
2.50 (0.19)
0.95b (0.16)
0.19b (0.01)
16.00ab (0.42)
T3
3.90c (0.03)
37.34b (2.17)
41.26 (2.32)
1.94 (0.30)
12.55ab (1.94)
2.44 (0.14)
1.20ab (0.07)
0.16c (0.01)
16.94a (0.23)
10-20
CK
4.22a (0.01)
36.31a (0.85)
25.36 (0.93)
1.18 (0.14)
8.85 (0.61)
1.68 (0.06)
0.69 (0.05)
0.19a (0.01)
15.13b (0.31)
T1
4.14a (0.03)
36.19a (0.89)
24.60 (1.98)
0.75 (0.09)
6.57 (0.66)
1.76 (0.11)
0.79 (0.12)
0.15bc (0.01)
13.95b (0.36)
T2
3.98b (0.04)
31.06b (0.68)
26.64 (2.13)
1.58 (0.41)
6.87 (0.77)
1.83 (0.14)
0.47 (0.11)
0.16b (0.01)
14.57b (0.44)
T3
4.03b (0.02)
32.26b (1.25)
28.66 (1.67)
1.56 (0.30)
7.31 (0.87)
1.74 (0.11)
0.72 (0.08)
0.13c (0.01)
16.60a (0.47)
表中数值为平均值(标准误)。CK, 对照, pH = 4.5; T1, pH = 4.0; T2, pH = 3.5; T3, pH = 3.0。不同小写字母表示相同土层不同处理下差异显著(p< 0.05)。 Data are mean (SE). CK, control, pH = 4.5; T1, pH = 4.0; T2, pH = 3.5; T3, pH = 3.0. AP, soil available phosphorus; SOC, soil organic carbon; SWC,soil water content; TC, soil total carbon; TN, soil total nitrogen; TP, soil total phosphorus. Different lowercase letters indicate significant difference between treatments in the same soil layer (p< 0.05).
Fig. 3Redundant analysis (RDA) of the relationship between soil properties and microbial community structure in soil layers 0-10 (A) and 10-20 cm (B). AP, soil available phosphorus content; NH4+, soil ammoniacal nitrogen content; NO3-, soil nitrate nitrogen content; SOC, soil organic carbon content; SWC, soil water content; TC, soil total carbon content; TN, soil total nitrogen content; TP, soil total phosphorus content.
3 讨论
3.1 模拟酸沉降对微生物生物量及其化学计量的影响
本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致。酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020)。例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018)。南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020)。此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014)。
AchatDL, MorelC, BakkerMR, AugustoL, PellerinS, Gallet-Budynek A, GonzalezM (2010). Assessing turnover of microbial biomass phosphorus: combination of an isotopic dilution method with a mass balance model Soil Biology & Biochemistry, 42, 2231-2240. DOI:10.1016/j.soilbio.2010.08.023URL [本文引用: 1]
AllisonVJ, CondronLM, PeltzerDA, RichardsonSJ, TurnerBL (2007). Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand Soil Biology & Biochemistry, 39, 1770-1781. DOI:10.1016/j.soilbio.2007.02.006URL [本文引用: 1]
B??thE, AndersonTH (2003). Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA- based techniques Soil Biology & Biochemistry, 35, 955-963. DOI:10.1016/S0038-0717(03)00154-8URL [本文引用: 2]
BardgettRD, McAlisterE (1999). The measurement of soil fungal:bacterial biomass ratios as an indicator of ecosystem self-regulation in temperate meadow grasslands Biology and Fertility of Soils, 29, 282-290. DOI:10.1007/s003740050554URL [本文引用: 2]
BonanGB (2008). Forests and Forests and climate change: forcings, feedbacks, and the climate benefits of forests Science, 320, 1444-1449. [本文引用: 1]
BouwmanAF, vanVuuren DP, DerwentRG, PoschM (2002). A global analysis of acidification and eutrophication of terrestrial ecosystems Water, Air, and Soil Pollution, 141, 349-382. DOI:10.1023/A:1021398008726URL [本文引用: 1]
ChenD, LanZ, BaiX, GraceJB, BaiY (2013). Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe Journal of Ecology, 101, 1322-1334. DOI:10.1111/jec.2013.101.issue-5URL [本文引用: 1]
ClevelandCC, HoultonBZ, SmithWK, MarkleinAR, ReedSC, PartonW, delGrosso SJ, RunningSW (2013). Patterns of new versus recycled primary production in the terrestrial biosphere Proceedings of the National Academy of Sciences of the United States of America, 110, 12733-12737. DOI:10.1073/pnas.1302768110PMID:23861492 [本文引用: 1] Nitrogen (N) and phosphorus (P) availability regulate plant productivity throughout the terrestrial biosphere, influencing the patterns and magnitude of net primary production (NPP) by land plants both now and into the future. These nutrients enter ecosystems via geologic and atmospheric pathways and are recycled to varying degrees through the plant-soil-microbe system via organic matter decay processes. However, the proportion of global NPP that can be attributed to new nutrient inputs versus recycled nutrients is unresolved, as are the large-scale patterns of variation across terrestrial ecosystems. Here, we combined satellite imagery, biogeochemical modeling, and empirical observations to identify previously unrecognized patterns of new versus recycled nutrient (N and P) productivity on land. Our analysis points to tropical forests as a hotspot of new NPP fueled by new N (accounting for 45% of total new NPP globally), much higher than previous estimates from temperate and high-latitude regions. The large fraction of tropical forest NPP resulting from new N is driven by the high capacity for N fixation, although this varies considerably within this diverse biome; N deposition explains a much smaller proportion of new NPP. By contrast, the contribution of new N to primary productivity is lower outside the tropics, and worldwide, new P inputs are uniformly low relative to plant demands. These results imply that new N inputs have the greatest capacity to fuel additional NPP by terrestrial plants, whereas low P availability may ultimately constrain NPP across much of the terrestrial biosphere.
ClevelandCC, LiptzinD (2007). C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass Biogeochemistry, 85, 235-252. DOI:10.1007/s10533-007-9132-0URL [本文引用: 3]
CuiJ, ZhouJ, PengY, HeYQ, YangH, MaoJD (2014). Atmospheric wet deposition of nitrogen and sulfur to a typical red soil agroecosystem in Southeast China during the ten-year monsoon seasons (2003-2012) Atmospheric Environment, 82, 121-129. DOI:10.1016/j.atmosenv.2013.10.023URL [本文引用: 1]
DeForestJL, ScottLG (2010). Available organic soil phosphorus has an important influence on microbial community composition Soil Science Society of America Journal, 74, 2059-2066. DOI:10.2136/sssaj2009.0426URL [本文引用: 1]
Delgado-BaquerizoM, ReichPB, KhachaneAN, CampbellCD, ThomasN, FreitagTE, AbuAl-Soud W, S?rensenS, BardgettRD, SinghBK (2017). It is elemental: soil nutrient stoichiometry drives bacterial diversity Environmental Microbiology, 19, 1176-1188. DOI:10.1111/1462-2920.13642PMID:27943556 [本文引用: 2] It is well established that resource quantity and elemental stoichiometry play major roles in shaping below and aboveground plant biodiversity, but their importance for shaping microbial diversity in soil remains unclear. Here, we used statistical modeling on a regional database covering 179 locations and six ecosystem types across Scotland to evaluate the roles of total carbon (C), nitrogen (N) and phosphorus (P) availabilities and ratios, together with land use, climate and biotic and abiotic factors, in determining regional scale patterns of soil bacterial diversity. We found that bacterial diversity and composition were primarily driven by variation in soil resource stoichiometry (total C:N:P ratios), itself linked to different land uses, and secondarily driven by other important biodiversity drivers such as climate, soil spatial heterogeneity, soil pH, root influence (plant-soil microbe interactions) and microbial biomass (soil microbe-microbe interactions). In aggregate, these findings provide evidence that nutrient stoichiometry is a strong predictor of bacterial diversity and composition at a regional scale.? 2016 Society for Applied Microbiology and John Wiley & Sons Ltd.
DengQ, HuiDF, DennisS, DennisS, ReddyKC (2017). Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: a meta-analysis Global Ecology and Biogeography, 26, 713-728. DOI:10.1111/geb.2017.26.issue-6URL [本文引用: 1]
Frosteg?rdA, B??thE (1996). The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil Biology and Fertility of Soils, 22, 59-65. DOI:10.1007/BF00384433URL [本文引用: 1]
Frosteg?rdA, B??thE, TunlioA (1993). Shifts in the structure of soil microbial communities in limed forests as revealed by phospholipid fatty acid analysis Soil Biology & Biochemistry, 25, 723-730. DOI:10.1016/0038-0717(93)90113-PURL [本文引用: 1]
GraystonSJ, GriffithGS, MawdsleyJL, CampbellCD, BardgettRD (2001). Accounting for variability in soil microbial communities of temperate upland grassland ecosystems Soil Biology & Biochemistry, 33, 533-551. DOI:10.1016/S0038-0717(00)00194-2URL [本文引用: 1]
GuoQ, YanL, KorpelainenH, Niinemetsü, LiC (2019). Plant-plant interactions and N fertilization shape soil bacterial and fungal communities Soil Biology & Biochemistry, 128, 127-138. DOI:10.1016/j.soilbio.2018.10.018URL [本文引用: 1]
H?gbergMN, H?gbergP, MyroldDD (2007). Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three Oecologia, 150, 590-601. PMID:17033802 [本文引用: 3] In Fennoscandian boreal forests, soil pH and N supply generally increase downhill as a result of water transport of base cations and N, respectively. Simultaneously, forest productivity increases, the understory changes from ericaceous dwarf shrubs to tall herbs; in the soil, fungi decrease whereas bacteria increase. The composition of the soil microbial community is mainly thought to be controlled by the pH and C-to-N ratio of the substrate. However, the latter also determines the N supply to plants, the plant community composition, and should also affect plant allocation of C below ground to roots and a major functional group of microbes, mycorrhizal fungi. We used phospholipid fatty acids (PLFAs) to analyze the potential importance of mycorrhizal fungi by comparing the microbial community composition in a tree-girdling experiment, where tree belowground C allocation was terminated, and in a long-term (34 years) N loading experiment, with the shifts across a natural pH and N supply gradient. Both tree girdling and N loading caused a decline of ca. 45% of the fungal biomarker PLFA 18:2omega6,9, suggesting a common mechanism, i.e., that N loading caused a decrease in the C supply to ectomycorrhizal fungi just as tree girdling did. The total abundance of bacterial PLFAs did not respond to tree girdling or to N loading, in which cases the pH (of the mor layer) did not change appreciably, but bacterial PLFAs increased considerably when pH increased across the natural gradient. Fungal biomass was high only in acid soil (pH < 4.1) with a high C-to-N ratio (>38). According to a principal component analysis, the soil C-to-N ratio was as good as predictor of microbial community structure as pH. Our study thus indicated the soil C-to-N ratio, and the response of trees to this ratio, as important factors that together with soil pH influence soil microbial community composition.
HouE, WenD, KuangY, CongJ, ChenC, HeX, HeenanM, LuH, ZhangY (2018). Soil pH predominantly controls the forms of organic phosphorus in topsoils under natural broadleaved forests along a 2500 km latitudinal gradient Geoderma, 315, 65-74. DOI:10.1016/j.geoderma.2017.11.041URL [本文引用: 1]
HuL, ZiH, WuP, WangY, LerdauM, WuX, WangC (2019). Soil bacterial communities in grasslands revegetated using Elymus nutans are largely influenced by soil pH and total phosphorus across restoration time Land Degradation & Development, 30, 2243-2256. DOI:10.1002/ldr.v30.18URL [本文引用: 1]
JiangJ, WangYP, YuMX, CaoNN, YanJH (2018). Soil organic matter is important for acid buffering and reducing aluminum leaching from acidic forest soils Chemical Geology, 501, 86-94. DOI:10.1016/j.chemgeo.2018.10.009URL [本文引用: 4]
JohnsonAH, FrizanoJ, VannDR (2003). Biogeochemical implications of labile phosphorus in forest soils determined by the Hedley fractionation procedure Oecologia, 135, 487-499. PMID:12695899 [本文引用: 1] Forest ecologists and biogeochemists have used a variety of extraction techniques to assess labile vs. non-labile soil P pools in chronosequences, the balance between biological vs. geochemical control of P transformations across a wide range of soil orders, the role of plants with either N-fixing or mycorrhizal symbionts in controlling soil P fractions, and to make inferences about plant-available P. Currently, variants of the sequential extraction procedure developed by M. J. Hedley and co-workers afford the greatest discrimination among labile and non-labile organic and inorganic P pools. Results of recent studies that used this technique to evaluate P fractions in forest soils indicate the following: (1) in intact, highly weathered forest soils of the humid tropics, Hedley-labile P values are several times larger than extractable P values resulting from mildly acidic extracting solutions which were commonly used in the past 2 decades; (2) pools of Hedley-labile P are several times larger than the annual forest P requirement and P required from the soil annually in both temperate and tropical forests; (3) long-term trends in non-labile P pools during pedogenesis are adequately represented by the Walker and Syers' model of changes in P fractionation during soil development. However, to better represent trends in pools that can supply plant-available P across forest soils of different age and weathering status, the paradigm should be modified; and (4) across a wide range of tropical and temperate forest soils, organic matter content is an important determinant of Hedley-labile P.
JonesDL, OburgerE (2011). Phosphorus in Action: Biological Processes in Soil Phosphorus Cycling. Springer, Berlin.169-198. [本文引用: 1]
KaiserEA, MuellerT, JoergensenRG, InsamH, HeinemeyerO (1992). Evaluation of methods to estimate the soil microbial biomass and the relationship with soil texture and organic matter Soil Biology & Biochemistry, 24, 675-683. DOI:10.1016/0038-0717(92)90046-ZURL [本文引用: 1]
KangHZ, GaoHH, YuWJ, YiY, WangY, NingML (2018). Changes in soil microbial community structure and function after afforestation depend on species and age: case study in a subtropical alluvial island Science of the Total Environment, 625, 1423-1432. DOI:10.1016/j.scitotenv.2017.12.180URL [本文引用: 1]
KillhamK, FirestoneMK, McColl JG (1983). Acid rain and soil microbial activity: effects and their mechanisms Journal of Environmental Quality, 12, 133-137. [本文引用: 1]
KwakJH, NaethMA, ChangSX (2018). Microbial activities and gross nitrogen transformation unaffected by ten-year nitrogen and sulfur addition Soil Science Society of America Journal, 82, 362-370. DOI:10.2136/sssaj2017.11.0384URL [本文引用: 1]
LambersH, MougelC, JaillardB, HinsingerP (2009). Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective Plant and Soil, 321, 83-115. DOI:10.1007/s11104-009-0042-xURL [本文引用: 1]
LauberCL, StricklandMS, BradfordMA, FiererN (2008). The influence of soil properties on the structure of bacterial and fungal communities across land-use types Soil Biology & Biochemistry, 40, 2407-2415. DOI:10.1016/j.soilbio.2008.05.021URL [本文引用: 1]
LiCY, WangY, BrookesP, DangTH, WangWZ (2013). Effect of soil pH on soil microbial carbon phosphorus ratio Scientia Agricultura Sinica, 46, 2709-2716. [本文引用: 1]
LiW, ShengH, EkawatiD, JiangY, YangH (2019). Variations in the compositions of soil bacterial and fungal communities due to microhabitat effects induced by simulated nitrogen deposition of a bamboo forest in wetland Forests, 10, 1098. DOI: 10.3390/f10121098. DOI:10.3390/f10121098 [本文引用: 1]
LiY, SunJ, TianD, TianDS, WangJS, HaDL, QuYX, JingGW, NiuSL (2018). Soil acid cations induced reduction in soil respiration under nitrogen enrichment and soil acidification Science of the Total Environment, 615, 1535-1546. DOI:10.1016/j.scitotenv.2017.09.131URL [本文引用: 2]
LiangGH, WuJP, XiongX, WuXY, ChuGW, ZhouGY, ZengRS, ZhangDQ (2015). Responses of soil pH value and soil microbial biomass carbon and nitrogen to simulated acid rain in three successional subtropical forests at Dinghushan nature reserve Ecology and Environmental Sciences, 24, 911-918. [本文引用: 1]
LiuKH, PengSL, MoJM, HuangZL, FangYT (2005). The process and mechanism of rain deposition upon forest plants Ecology and Environment, 14, 953-960. [本文引用: 1]
LiuX, ZhangB, ZhaoWR, WangL, XieDJ, HuoWT, WuYW, ZhangJC (2017). Comparative effects of sulfuric and nitric acid rain on litter decomposition and soil microbial community in subtropical plantation of Yangtze River Delta region Science of the Total Environment, 601, 669-678. [本文引用: 3]
LiuXC, ZhangST (2019). Nitrogen addition shapes soil enzyme activity patterns by changing pH rather than the composition of the plant and microbial communities in an alpine meadow soil Plant and Soil, 440, 11-24. DOI:10.1007/s11104-019-04054-5URL [本文引用: 1]
LiuZ, LiD, ZhangJ, SaleemM, ZhangY, MaR, HeY, YangJ, XiangH, WeiH (2020). Effect of simulated acid rain on soil CO2, CH 4 and N2O emissions and microbial communities in an agricultural soil Geoderma, 366, 114222. DOI: 10.1016/j.geoderma.2020.114222. DOI:10.1016/j.geoderma.2020.114222 [本文引用: 3]
LvYN, WangCY, JiaYY, WangWW, MaX, DuJJ, PuGZ, TianXJ (2014). Effects of sulfuric, nitric, and mixed acid rain on litter decomposition, soil microbial biomass, and enzyme activities in subtropical forests of China Applied Soil Ecology, 79, 1-9. DOI:10.1016/j.apsoil.2013.12.002URL [本文引用: 2]
MaH, ZouW, YangJ, HoganJA, XuH, ChenJ (2019). Dominant tree species shape soil microbial community via regulating assembly processes in planted subtropical forests Forests, 10, 978. DOI: 10.3390/f10110978. DOI:10.3390/f10110978URL [本文引用: 1]
MaltzMR, ChenZ, CaoJ, ArogyaswamyK, ShulmanH, AronsonEL (2019). Inoculation with Pisolithus tinctorius may ameliorate acid rain impacts on soil microbial communities associated with Pinus massoniana seedlings Fungal Ecology, 40, 50-61. DOI:10.1016/j.funeco.2018.11.011 [本文引用: 1] Human activities accelerate acidification, particularly as acid rain, which may have lasting impacts on soil abiotic and biotic parameters. However, the effects of acidification on aboveground vegetation, below-ground communities, and carbon cycling remains unresolved. We examined the effects of long-term acidic treatments and Pisolithus tinctorius inoculation on plants, soils, and microbial communities in pine (Pinus) plantations and found that exposure to severely-acidic treatments diminished plant performance, altered microbial communities, and decreased organic matter, nitrate, and available phosphorus. Although we did not detect any benefits of P. tinctorius inoculation for Pinus seedlings impacted by severely-acidic treatments, when these severe treatments were inoculated with P. tinctorius, both soil properties and microbial community composition shifted. We posit that inoculation with P. tinctorius may alleviate stressful environmental conditions, and change the structure of mycorrhizal fungal communities. Although acidification may alter biogeochemical cycles and constrain aboveground and belowground communities, P. tinctorius inoculation may provide benefits to some components of forested ecosystems. (C) 2018 Elsevier Ltd and British Mycological Society.
MengC, TianDS, ZengH, LiZL, YiCX, NiuSL (2019). Global soil acidification impacts on belowground processes Environmental Research Letters, 14, 074003. DOI: 10.1088/1748-9326/ab239c. DOI:10.1088/1748-9326/ab239cURL [本文引用: 2]
MoJ, ZhangW, ZhuW, GundersenP, FangY, LiD, WangH (2008). Nitrogen addition reduces soil respiration in a mature tropical forest in southern China Global Change Biology, 14, 403-412. DOI:10.1111/j.1365-2486.2007.01503.xURL [本文引用: 1]
NottinghamAT, HicksLC, CcahuanaAJQ, SalinasN, B??thE, MeirP (2018). Nutrient limitations to bacterial and fungal growth during cellulose decomposition in tropical forest soils Biology and Fertility of Soils, 54, 219-228. DOI:10.1007/s00374-017-1247-4URL [本文引用: 1]
ObersonA, FriesenDK, MorelC, TiessenH (1997). Determination of phosphorus released by chloroform fumigation from microbial biomass in high P sorbing tropical soils Soil Biology & Biochemistry, 29, 1579-1583. DOI:10.1016/S0038-0717(97)00049-7URL [本文引用: 1]
OksanenJF, BlanchetG, FriendlyM, KindtR, LegendreP, McGlinnD, MinchinPR, O?HaraRB, SimpsonGL, SolymosP, StevensMHH, SzoecsE, WagnerH (2018). vegan: Community ecology package. R package version 2.5-2. [2020-03-06]. https://CRAN.R-project.org/package=vegan. URL [本文引用: 1]
PanGX (1990). Soil chemical analysis on the process of soil acidification Journal of Ecology, 9(6), 48-52. [本文引用: 1]
PennanenT, FritzeH, VanhalaP, Kiikkil?O, NeuvonenS, B??thE (1998). Structure of a microbial community in soil after prolonged addition of low levels of simulated acid rain Applied and Environmental Microbiology, 64, 2173-2180. PMID:9603831 [本文引用: 1] Humus samples were collected 12 growing seasons after the start of a simulated acid rain experiment situated in the subarctic environment. The acid rain was simulated with H2SO4, a combination of H2SO4 and HNO3, and HNO3 at two levels of moderate acidic loads close to the natural anthropogenic pollution levels of southern Scandinavia. The higher levels of acid applications resulted in acidification, as defined by humus chemistry. The concentrations of base cations decreased, while the concentrations of exchangeable H+, Al, and Fe increased. Humus pH decreased from 3.83 to 3.65. Basal respiration decreased with decreasing humus pH, and total microbial biomass, measured by substrate-induced respiration and total amount of phospholipid fatty acids (PLFA), decreased slightly. An altered PLFA pattern indicated a change in the microbial community structure at the higher levels of acid applications. In general, branched fatty acids, typical of gram-positive bacteria, increased in the acid plots. PLFA analysis performed on the bacterial community growing on agar plates also showed that the relative amount of PLFA specific for gram-positive bacteria increased due to the acidification. The changed bacterial community was adapted to the more acidic environment in the acid-treated plots, even though bacterial growth rates, estimated by thymidine and leucine incorporation, decreased with pH. Fungal activity (measured as acetate incorporation into ergosterol) was not affected. This result indicates that bacteria were more affected than fungi by the acidification. The capacity of the bacterial community to utilize 95 different carbon sources was variable and only showed weak correlations to pH. Differences in the toxicities of H2SO4 and HNO3 for the microbial community were not found.
QiaoX, XiaoW, JaffeD, KotaSH, YingQ, TangY (2015). Atmospheric wet deposition of sulfur and nitrogen in Jiuzhaigou National Nature Reserve, Sichuan Province, China Science of the Total Environment, 511, 28-36. DOI:10.1016/j.scitotenv.2014.12.028URL [本文引用: 2]
RCore Team (2016). R: a Language and Environment for Statistical Computing R Foundation for Statistical Computing, Vienna, Austria. [本文引用: 1]
TresederKK (2008). Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies Ecology Letters, 11, 1111-1120. DOI:10.1111/ele.2008.11.issue-10URL [本文引用: 1]
van der Heijden MGA, BardgettRD, vanStraalen NM (2008). The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems Ecology Letters, 11, 296-310. PMID:18047587 Microbes are the unseen majority in soil and comprise a large portion of life's genetic diversity. Despite their abundance, the impact of soil microbes on ecosystem processes is still poorly understood. Here we explore the various roles that soil microbes play in terrestrial ecosystems with special emphasis on their contribution to plant productivity and diversity. Soil microbes are important regulators of plant productivity, especially in nutrient poor ecosystems where plant symbionts are responsible for the acquisition of limiting nutrients. Mycorrhizal fungi and nitrogen-fixing bacteria are responsible for c. 5-20% (grassland and savannah) to 80% (temperate and boreal forests) of all nitrogen, and up to 75% of phosphorus, that is acquired by plants annually. Free-living microbes also strongly regulate plant productivity, through the mineralization of, and competition for, nutrients that sustain plant productivity. Soil microbes, including microbial pathogens, are also important regulators of plant community dynamics and plant diversity, determining plant abundance and, in some cases, facilitating invasion by exotic plants. Conservative estimates suggest that c. 20 000 plant species are completely dependent on microbial symbionts for growth and survival pointing to the importance of soil microbes as regulators of plant species richness on Earth. Overall, this review shows that soil microbes must be considered as important drivers of plant diversity and productivity in terrestrial ecosystems.
van der HeijdenMGA, KlironomosJN, UrsicM, MoutoglisP, Streitwolf-EngelR, BollerT, WiemkenA, SandersIR (1998). Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity Nature, 396, 69-72. DOI:10.1038/23932URL
WareGW, NiggsHN, BevenueA (1990). Reviews of Environmental Contamination and Toxicology: Continuation of Residue Reviews. Springer, New York. [本文引用: 2]
WuJP, LiangGH, HuiDF, DengQ, XiongX, QiuQY, LiuJX, ChuGW, ZhouGY, ZhangDQ (2016). Prolonged acid rain facilitates soil organic carbon accumulation in a mature forest in Southern China Science of the Total Environment, 544, 94-102. DOI:10.1016/j.scitotenv.2015.11.025URL [本文引用: 1]
XieSY, WangRB, ZhengHH (2012). Analysis on the acid rain from 2005 to 2011 in China Environmental Monitoring and Forewarning, 4(5), 33-37. [本文引用: 1]
XuX, ThorntonPE, PostWM (2013). A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems Global Ecology and Biogeography, 22, 737-749. DOI:10.1111/geb.12029URL [本文引用: 1]
YangJL, FuXL, MaZQ, DiYB, LiuQJ, WangHM (2015). Characteristics of soil microbial community in five forest types in mid-subtropical China Research of Environmental Sciences, 28, 720-727. [本文引用: 1]
ZhangHL, WuJP, XiongX, ChuGW, ZhouGY, ZhangDQ (2018). Effects of simulated acid rain on soil labile organic carbon and carbon management index in subtropical forests of China Acta Ecologica Sinica, 38, 657-667. [本文引用: 1]
ZhangXM, ChaiFH, WangSL, SunXZ, HanM (2010). Research progress of acid precipitation in China Research of Environmental Sciences, 23, 527-532. [本文引用: 1]
ZhaoD, XiongJ, XuY, ChanWH (1988). Acid rain in southwestern China Atmospheric Environment, 22, 349-358. [本文引用: 1]
ZhengK, ZhaoTL, ZhangL, ZengN, ZhengXB, YangQJ (2019). Characteristics of wet deposition of sulfate and nitrate in three typical cities in China in 2001-2017 Ecology and Environmental Sciences, 28, 2390-2397. [本文引用: 1]
ZhengS, BianHF, QuanQ, XuL, ChenZ, HeNP (2018). Effect of nitrogen and acid deposition on soil respiration in a temperate forest in China Geoderma, 329, 82-90. DOI:10.1016/j.geoderma.2018.05.022URL [本文引用: 2]
ZhouGY, PengCH, LiYL, LiuSZ, ZhangQM, TangXL, LiuJX, YanJH, ZhangDQ, ChuGW (2013). A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China Global Change Biology, 19, 1197-1210. DOI:10.1111/gcb.12128URL [本文引用: 1]
ZhouXD, XuZF, LiuWJ, WuY, ZhaoT, JiangH, ZhangX, ZhangJY, ZhouL, WangYC (2019). Chemical composition of precipitation in Shenzhen, a coastal mega-city in South China: influence of urbanization and anthropogenic activities on acidity and ionic composition Science of the Total Environment, 662, 218-226. DOI:10.1016/j.scitotenv.2019.01.096URL [本文引用: 1]
ZhuQ, deVries W, LiuX, ZengM, HaoT, DuE, ZhangF, ShenJ (2016). The contribution of atmospheric deposition and forest harvesting to forest soil acidification in China since 1980 Atmospheric Environment, 146, 215-222. DOI:10.1016/j.atmosenv.2016.04.023URL [本文引用: 1]
ZieglerSE, BillingsSA, LaneCS, LiJW, FogelML (2013). Warming alters routing of labile and slower-turnover carbon through distinct microbial groups in boreal forest organic soils Soil Biology & Biochemistry, 60, 23-32. DOI:10.1016/j.soilbio.2013.01.001URL [本文引用: 1]
ZouS, ZhouGY, ZhangQM, XuS, XiongX, XiaYJ, LiuSZ, MengZ, ChuGW (2018). Long-term (1992-2015) dynamics of community composition and structure in a monsoon evergreen broad-leaved forest in Dinghushan Biosphere Reserve Chinese Journal of Plant Ecology, 42, 442-452. DOI:10.17521/cjpe.2017.0171URL [本文引用: 1]
[邹顺, 周国逸, 张倩媚, 徐姗, 熊鑫, 夏艳菊, 刘世忠, 孟泽, 褚国伟 (2018). 1992-2015年鼎湖山季风常绿阔叶林群落结构动态 植物生态学报, 42, 442-452.] DOI:10.17521/cjpe.2017.0171 [本文引用: 1] 季风常绿阔叶林是中国南亚热带地区的地带性植被, 其群落结构及其动态变化直接影响着该森林能为该地区提供的生态系统服务功能质量。该文利用1992-2015年共24年的长期定点监测数据, 从群落种类组成、生物量、径级、密度等数量特征方面研究了鼎湖山季风常绿阔叶林群落结构的动态变化。结果表明: 1)到2015年该森林群落林分个体密度增加了42.7%, 总生物量减少了5.1%; 24年间基于生物量的β多样性指数群落差异为37.4%, 基于个体数的差异则高达65.6%; 2)灌木和小乔木的个体数剧增, 生物量增大, 中乔木和大乔木的个体数变化虽不显著, 但生物量显著降低; 3)小径级(胸径Aidia canthioides)、鼎湖血桐(Macaranga sampsonii)、柏拉木(Blastus cochinchinensis)等物种个体数急剧增加, 锥(Castanopsis chinensis)、木荷(Schima superba)、黄果厚壳桂(Cryptocarya concinna)等树种生物量大量减少, 白颜树(Gironniera subaequalis)、窄叶半枫荷(Pterospermum lanceifolium)等树种生物量增加, 这些物种是群落结构变化的主要贡献者。鼎湖山季风常绿阔叶林群落结构在1992-2015年发生了巨大变化, 与演替、虫害和气候变化等影响有关。 Assessing turnover of microbial biomass phosphorus: combination of an isotopic dilution method with a mass balance model 1 2010
Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence, New Zealand 1 2007
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA- based techniques 2 2003
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
The measurement of soil fungal:bacterial biomass ratios as an indicator of ecosystem self-regulation in temperate meadow grasslands 2 1999
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
... ., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
Forests and climate change: forcings, feedbacks, and the climate benefits of forests 1 2008
... 热带森林是陆地生态系统中最重要的碳库, 储存了近1/4的陆地碳, 并且每年能够吸收大量的CO2 (Bonan, 2008).但与此同时, 热带森林土壤较好的水热条件使得地质发育较为成熟, 土壤中养分十分贫瘠(Vitousek et al., 2010).而长期酸雨导致土壤酸化加剧, 养分流失愈发严重(Qiao et al., 2015), 微生物活性和群落组成发生变化, 从而进一步影响了土壤中养分的循环, 进而导致植物多样性和生产力受到限制(Bouwman et al., 2002).目前, 由于受到野外条件的限制, 热带森林生态系统中微生物对土壤酸化响应的研究主要集中在比较自然状态下pH不同的土壤其微生物结构的差异(Hu et al., 2019), 以及通过室内培养实验探究各个类群微生物对土壤响应的差异(Liu et al., 2020), 而缺乏直接的长期原位控制实验.本研究通过模拟酸沉降实验探究土壤理化性质的变化如何影响土壤微生物活性及其群落结构, 有利于揭示土壤养分有效性的变化机制, 为未来气候变化背景下森林生态系统生物多样性稳定及其可持续经营提供科学依据. ...
A global analysis of acidification and eutrophication of terrestrial ecosystems 1 2002
... 热带森林是陆地生态系统中最重要的碳库, 储存了近1/4的陆地碳, 并且每年能够吸收大量的CO2 (Bonan, 2008).但与此同时, 热带森林土壤较好的水热条件使得地质发育较为成熟, 土壤中养分十分贫瘠(Vitousek et al., 2010).而长期酸雨导致土壤酸化加剧, 养分流失愈发严重(Qiao et al., 2015), 微生物活性和群落组成发生变化, 从而进一步影响了土壤中养分的循环, 进而导致植物多样性和生产力受到限制(Bouwman et al., 2002).目前, 由于受到野外条件的限制, 热带森林生态系统中微生物对土壤酸化响应的研究主要集中在比较自然状态下pH不同的土壤其微生物结构的差异(Hu et al., 2019), 以及通过室内培养实验探究各个类群微生物对土壤响应的差异(Liu et al., 2020), 而缺乏直接的长期原位控制实验.本研究通过模拟酸沉降实验探究土壤理化性质的变化如何影响土壤微生物活性及其群落结构, 有利于揭示土壤养分有效性的变化机制, 为未来气候变化背景下森林生态系统生物多样性稳定及其可持续经营提供科学依据. ...
Evidence that acidification-induced declines in plant diversity and productivity are mediated by changes in below-ground communities and soil properties in a semi-arid steppe 1 2013
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Patterns of new versus recycled primary production in the terrestrial biosphere 1 2013
... 本研究的对象为南亚热带季风常绿阔叶林, 其土壤高度风化, 并且在长期受到不平衡氮磷沉降后, 植物生产力和土壤微生物活性的主要限制性养分由氮转为了磷(Mo et al., 2008; Cleveland et al., 2013; Deng et al., 2017).因此, 长达10年的酸沉降实验虽然显著改变了SWC、土壤硝态氮含量以及C:N (表2), 但这并未成为影响土壤微生物群落结构的主要因素(图3).此外, 在酸化加剧背景下, 土壤AP、TP以及MBP含量均呈显著的下降趋势, 这意味着土壤酸化确实有可能导致热带森林磷限制的加剧, 从而减少微生物对磷的同化作用. ...
C:N:P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass 3 2007
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
Atmospheric wet deposition of nitrogen and sulfur to a typical red soil agroecosystem in Southeast China during the ten-year monsoon seasons (2003-2012) 1 2014
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Available organic soil phosphorus has an important influence on microbial community composition 1 2010
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
It is elemental: soil nutrient stoichiometry drives bacterial diversity 2 2017
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Responses of terrestrial ecosystem phosphorus cycling to nitrogen addition: a meta-analysis 1 2017
... 本研究的对象为南亚热带季风常绿阔叶林, 其土壤高度风化, 并且在长期受到不平衡氮磷沉降后, 植物生产力和土壤微生物活性的主要限制性养分由氮转为了磷(Mo et al., 2008; Cleveland et al., 2013; Deng et al., 2017).因此, 长达10年的酸沉降实验虽然显著改变了SWC、土壤硝态氮含量以及C:N (表2), 但这并未成为影响土壤微生物群落结构的主要因素(图3).此外, 在酸化加剧背景下, 土壤AP、TP以及MBP含量均呈显著的下降趋势, 这意味着土壤酸化确实有可能导致热带森林磷限制的加剧, 从而减少微生物对磷的同化作用. ...
The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil 1 1996
Accounting for variability in soil microbial communities of temperate upland grassland ecosystems 1 2001
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Plant-plant interactions and N fertilization shape soil bacterial and fungal communities 1 2019
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
Is microbial community composition in boreal forest soils determined by pH, C-to-N ratio, the trees, or all three 3 2007
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
... ., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
Soil pH predominantly controls the forms of organic phosphorus in topsoils under natural broadleaved forests along a 2500 km latitudinal gradient 1 2018
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
Soil bacterial communities in grasslands revegetated using Elymus nutans are largely influenced by soil pH and total phosphorus across restoration time 1 2019
... 热带森林是陆地生态系统中最重要的碳库, 储存了近1/4的陆地碳, 并且每年能够吸收大量的CO2 (Bonan, 2008).但与此同时, 热带森林土壤较好的水热条件使得地质发育较为成熟, 土壤中养分十分贫瘠(Vitousek et al., 2010).而长期酸雨导致土壤酸化加剧, 养分流失愈发严重(Qiao et al., 2015), 微生物活性和群落组成发生变化, 从而进一步影响了土壤中养分的循环, 进而导致植物多样性和生产力受到限制(Bouwman et al., 2002).目前, 由于受到野外条件的限制, 热带森林生态系统中微生物对土壤酸化响应的研究主要集中在比较自然状态下pH不同的土壤其微生物结构的差异(Hu et al., 2019), 以及通过室内培养实验探究各个类群微生物对土壤响应的差异(Liu et al., 2020), 而缺乏直接的长期原位控制实验.本研究通过模拟酸沉降实验探究土壤理化性质的变化如何影响土壤微生物活性及其群落结构, 有利于揭示土壤养分有效性的变化机制, 为未来气候变化背景下森林生态系统生物多样性稳定及其可持续经营提供科学依据. ...
Soil microbiomes and climate change 2 2020
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Soil organic matter is important for acid buffering and reducing aluminum leaching from acidic forest soils 4 2018
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
... ., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
... 研究地点位于中国广东省肇庆市鼎湖山国家自然保护区内(112.50°-112.55° E, 23.15°-23.18° N), 处于南亚热带湿润森林地区, 属于典型季风气候, 雨热同期, 年降水量为1 929 mm, 年平均气温21 ℃.干湿季节交替明显, 其中4-9月为雨季, 降水量占年降水量的80%, 10月至次年3月为旱季(Lv et al., 2014).近20多年来, 研究区域降水pH均低于4.5, 酸雨成分主要为硫酸和硝酸混合, 土壤pH逐年下降(Jiang et al., 2018). ...
Changes in soil microbial community structure and function after afforestation depend on species and age: case study in a subtropical alluvial island 1 2018
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
Acid rain and soil microbial activity: effects and their mechanisms 1 1983
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
Microbial activities and gross nitrogen transformation unaffected by ten-year nitrogen and sulfur addition 1 2018
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
Plant-microbe-soil interactions in the rhizosphere: an evolutionary perspective 1 2009
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
The influence of soil properties on the structure of bacterial and fungal communities across land-use types 1 2008
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
Variations in the compositions of soil bacterial and fungal communities due to microhabitat effects induced by simulated nitrogen deposition of a bamboo forest in wetland 1 2019
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
Soil acid cations induced reduction in soil respiration under nitrogen enrichment and soil acidification 2 2018
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
... ).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
酸沉降对森林植物影响过程和机理 1 2005
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Comparative effects of sulfuric and nitric acid rain on litter decomposition and soil microbial community in subtropical plantation of Yangtze River Delta region 3 2017
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
... )发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
Nitrogen addition shapes soil enzyme activity patterns by changing pH rather than the composition of the plant and microbial communities in an alpine meadow soil 1 2019
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
Enhanced nitrogen deposition over China 2 2013
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
... ., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Effect of simulated acid rain on soil CO2, CH 4 and N2O emissions and microbial communities in an agricultural soil 3 2020
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
... 热带森林是陆地生态系统中最重要的碳库, 储存了近1/4的陆地碳, 并且每年能够吸收大量的CO2 (Bonan, 2008).但与此同时, 热带森林土壤较好的水热条件使得地质发育较为成熟, 土壤中养分十分贫瘠(Vitousek et al., 2010).而长期酸雨导致土壤酸化加剧, 养分流失愈发严重(Qiao et al., 2015), 微生物活性和群落组成发生变化, 从而进一步影响了土壤中养分的循环, 进而导致植物多样性和生产力受到限制(Bouwman et al., 2002).目前, 由于受到野外条件的限制, 热带森林生态系统中微生物对土壤酸化响应的研究主要集中在比较自然状态下pH不同的土壤其微生物结构的差异(Hu et al., 2019), 以及通过室内培养实验探究各个类群微生物对土壤响应的差异(Liu et al., 2020), 而缺乏直接的长期原位控制实验.本研究通过模拟酸沉降实验探究土壤理化性质的变化如何影响土壤微生物活性及其群落结构, 有利于揭示土壤养分有效性的变化机制, 为未来气候变化背景下森林生态系统生物多样性稳定及其可持续经营提供科学依据. ...
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
Effects of sulfuric, nitric, and mixed acid rain on litter decomposition, soil microbial biomass, and enzyme activities in subtropical forests of China 2 2014
... 研究地点位于中国广东省肇庆市鼎湖山国家自然保护区内(112.50°-112.55° E, 23.15°-23.18° N), 处于南亚热带湿润森林地区, 属于典型季风气候, 雨热同期, 年降水量为1 929 mm, 年平均气温21 ℃.干湿季节交替明显, 其中4-9月为雨季, 降水量占年降水量的80%, 10月至次年3月为旱季(Lv et al., 2014).近20多年来, 研究区域降水pH均低于4.5, 酸雨成分主要为硫酸和硝酸混合, 土壤pH逐年下降(Jiang et al., 2018). ...
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
Dominant tree species shape soil microbial community via regulating assembly processes in planted subtropical forests 1 2019
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Inoculation with Pisolithus tinctorius may ameliorate acid rain impacts on soil microbial communities associated with Pinus massoniana seedlings 1 2019
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
Global soil acidification impacts on belowground processes 2 2019
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
Nitrogen addition reduces soil respiration in a mature tropical forest in southern China 1 2008
... 本研究的对象为南亚热带季风常绿阔叶林, 其土壤高度风化, 并且在长期受到不平衡氮磷沉降后, 植物生产力和土壤微生物活性的主要限制性养分由氮转为了磷(Mo et al., 2008; Cleveland et al., 2013; Deng et al., 2017).因此, 长达10年的酸沉降实验虽然显著改变了SWC、土壤硝态氮含量以及C:N (表2), 但这并未成为影响土壤微生物群落结构的主要因素(图3).此外, 在酸化加剧背景下, 土壤AP、TP以及MBP含量均呈显著的下降趋势, 这意味着土壤酸化确实有可能导致热带森林磷限制的加剧, 从而减少微生物对磷的同化作用. ...
Nutrient limitations to bacterial and fungal growth during cellulose decomposition in tropical forest soils 1 2018
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
Determination of phosphorus released by chloroform fumigation from microbial biomass in high P sorbing tropical soils 1 1997
Microbial carbon use efficiency and biomass turnover times depending on soil depth—Implications for carbon cycling 1 2016
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
Considering fungal:bacterial dominance in soils—Methods, controls, and ecosystem implications 1 2010
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
Acidic deposition, nutrient leaching and forest growth 1 2003
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Nitrogen additions and microbial biomass: a meta-analysis of ecosystem studies 1 2008
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems 2008
Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen-phosphorus interactions 1 2010
... 热带森林是陆地生态系统中最重要的碳库, 储存了近1/4的陆地碳, 并且每年能够吸收大量的CO2 (Bonan, 2008).但与此同时, 热带森林土壤较好的水热条件使得地质发育较为成熟, 土壤中养分十分贫瘠(Vitousek et al., 2010).而长期酸雨导致土壤酸化加剧, 养分流失愈发严重(Qiao et al., 2015), 微生物活性和群落组成发生变化, 从而进一步影响了土壤中养分的循环, 进而导致植物多样性和生产力受到限制(Bouwman et al., 2002).目前, 由于受到野外条件的限制, 热带森林生态系统中微生物对土壤酸化响应的研究主要集中在比较自然状态下pH不同的土壤其微生物结构的差异(Hu et al., 2019), 以及通过室内培养实验探究各个类群微生物对土壤响应的差异(Liu et al., 2020), 而缺乏直接的长期原位控制实验.本研究通过模拟酸沉降实验探究土壤理化性质的变化如何影响土壤微生物活性及其群落结构, 有利于揭示土壤养分有效性的变化机制, 为未来气候变化背景下森林生态系统生物多样性稳定及其可持续经营提供科学依据. ...
Fungi to bacteria ratio: historical misinterpretations and potential implications 1 2019
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
广州酸雨现状及影响因素分析 1 2011
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
广州酸雨现状及影响因素分析 1 2011
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Ecological linkages between aboveground and belowground biota 5 2004
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
... ., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
... ., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
... 除土壤pH外, 土壤磷含量是导致本研究中土壤微生物群落结构发生变化的另一个主要原因(图3), 即土壤酸化导致的土壤磷限制加剧可能进一步促进土壤微生物群落向以真菌为主的微生物群落的转变(表2, 表3).大量研究表明, 在土壤磷限制严重的热带森林生态系统中, 微生物真菌和细菌的比值显著高于农田和草地土壤, 且真菌丰度与土壤磷含量和C:N具有很强的相关关系(Lauber et al., 2008), 而这可能主要是真菌在养分贫瘠环境中较高的适应性, 以及在酸性条件下对磷具有更强的获取能力所导致(DeForest & Scott, 2010; Jones & Oburger, 2011).例如, 在磷有效性过低的情况下, 为了维持生长所需的足够养分, 植物将更多的光合作用产物投入地下, 通过增加其根系上菌根真菌的定殖以扩大土壤磷的获取范围和增加吸收面积, 增加对养分的获取能力(van der Heijden et al., 1998; Wardle et al., 2004), 或者通过养分的重吸收从而减少养分的消耗等途径适应养分贫瘠的土壤, 而这些变化最终也将导致土壤中微生物的群落结构发生改变(Strickland & Rousk, 2010).此外, 在热带森林土壤纤维素分解过程的研究中发现, 真菌和细菌生长的限制性养分存在差异, 细菌由于较大的比表面积对磷脂的需求较高, 使得土壤磷含量的改变也可能进一步抑制了细菌的活性(Nottingham et al., 2018).不同的是, 温带森林生态系统中, 模拟酸沉降导致的微生物群落结构的变化主要是土壤中碳和氮的有效性改变所导致的(Killham et al., 1983), 在氮有效性较低的条件下, 氮的添加可以通过解除土壤中微生物的碳和氮的限制, 从而缓解氮添加导致的土壤pH下降对微生物生长和群落结构的影响(Treseder, 2008). ...
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
2 1990
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
... ., 1990), 更能够适应未来的土壤酸化. ...
Prolonged acid rain facilitates soil organic carbon accumulation in a mature forest in Southern China 1 2016
... 土壤pH是影响土壤微生物活性和群落结构的重要因素(H?gberget al., 2007).酸雨则主要是通过加剧土壤的酸化以及影响土壤中的养分状态等直接改变土壤中的微生物的活动和群落组成(Wu et al., 2016).绝大多数土壤细菌较适应中性或弱酸性土壤, 而真菌则能够在更低的土壤pH中生存(H?gberget al., 2007), 甚至随着土壤pH的下降, 真菌的活性可能增加, 降解作用呈现出增加的趋势, 有利于缓解土壤中的养分限制(Maltz et al., 2019).例如, Liu和Zhang (2019)研究表明, 土壤pH是导致土壤中微生物活性以及群落结构发生变化的决定因素; 酸雨可以显著改变土壤微生物生物量(Liu et al., 2020), 导致F:B随着土壤pH的增加呈现出显著的下降趋势(B??th & Anderson,2003).此外, 土壤pH还显著改变了土壤中的养分状态和含量(Hou et al., 2018), 导致了土壤微生物化学计量(Cleveland & Liptzin, 2007)和群落功能发生转变(Allison et al., 2007).基于此, 我们推测森林生态系统长期的模拟酸雨实验可能会通过加剧土壤的酸化程度以及影响土壤中的理化性质和养分状况等途径, 引起土壤中微生物活性的变化, 并通过改变微生物群落的组成, 尤其是F:B, 来适应土壤环境的变化, 从而维持地上部分的生产力. ...
2005-2011年全国酸雨状况分析 1 2012
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
2005-2011年全国酸雨状况分析 1 2012
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems 1 2013
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
中亚热带5种类型森林土壤微生物群落特征 1 2015
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
中亚热带5种类型森林土壤微生物群落特征 1 2015
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
南亚热带森林土壤碳库稳定性与碳库管理指数对模拟酸雨的响应 1 2018
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
南亚热带森林土壤碳库稳定性与碳库管理指数对模拟酸雨的响应 1 2018
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
中国酸雨研究现状 1 2010
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
中国酸雨研究现状 1 2010
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Acid rain in southwestern China 1 1988
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
2001-2017年中国3个典型城市硫酸盐和硝酸盐湿沉降特征 1 2019
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
2001-2017年中国3个典型城市硫酸盐和硝酸盐湿沉降特征 1 2019
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Effect of nitrogen and acid deposition on soil respiration in a temperate forest in China 2 2018
... 本研究中, 长期的模拟酸沉降实验并未显著改变土壤中MBC含量(图1A), 这与本样地的早期结果(张慧玲等, 2018)一致.酸沉降对土壤微生物活性的影响取决于生态系统类型(Jansson & Hofmockel, 2020).例如, 在副极地生态系统的长期模拟酸沉降实验中发现, 土壤酸化显著降低土壤MBC含量(Pennanen et al., 1998), 而在中国温带森林生态系统的中性偏碱性土壤中, 酸雨并未显著改变土壤微生物生物量(Kwak et al., 2018; Zheng et al., 2018).南亚热带室内模拟酸雨淋洗土柱的研究则发现, 在短期内含氮酸雨输入带来的“施肥效应”以及对土壤中有效性养分的活化等抵消了酸雨导致的负面效应, 反而会刺激土壤微生物的活性(Liuet al., 2020).此外, 在初始pH较低的森林土壤中, 微生物活性可能对外界酸的输入有着更高的抗性(Meng et al., 2019), 而凋落物质量的差异等还使得针叶林中微生物对土壤pH的敏感程度远高于阔叶林(Lv et al., 2014). ...
... 在本研究中, 土壤酸化的加剧导致土壤微生物群落结构发生显著变化(图2A), 导致10-20 cm土壤中F:B显著增加(表3), 其中土壤pH是导致微生物群落结构发生变化的主要原因(图3).这些发现与先前亚热带森林生态系统中的研究结果相符(Liu et al., 2017; Li et al., 2018), 即在一定的pH范围内外源酸的输入对细菌和真菌有着不同的影响(Meng et al., 2019), 真菌对土壤pH的下降具有较强的适应性(B??th & Anderson,2003).在亚热带阔叶落叶林中, Li等(2018)发现土壤酸化虽然对真菌和细菌均没有显著的影响, 但显著增加F:B.Liu等(2017)在亚热带人工林中则发现土壤酸化显著减少细菌的相对丰度, 但对真菌没有显著影响, 导致以细菌为主的微生物群落转变为以真菌为主的微生物群落.在北方森林生态系统中, 随着土壤pH的增加, 细菌相对丰度显著增加, 而真菌则在酸性土壤中表现出较高的活性(H?gberget al., 2007).湿地森林生态系统中, 模拟氮沉降导致土壤pH显著下降也会使得土壤微生物群落结构发生显著的变化(Li et al., 2019).而在土壤中性偏碱性的温带森林生态系统中, 两年的模拟酸雨处理导致土壤pH下降则并未显著改变土壤中的F:B (Zheng et al., 2018).通常认为, 真菌将过剩的H+储存在液泡中的能力及其异于细菌的细胞壁结构是真菌和细菌具有不同的最适pH范围的主要原因(Ware et al., 1990).因此相较于细菌, 真菌的最适pH范围更低且更广(Ware et al., 1990), 更能够适应未来的土壤酸化. ...
A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China 1 2013
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
Chemical composition of precipitation in Shenzhen, a coastal mega-city in South China: influence of urbanization and anthropogenic activities on acidity and ionic composition 1 2019
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
The contribution of atmospheric deposition and forest harvesting to forest soil acidification in China since 1980 1 2016
... 自酸雨被列为国际性环境问题, 并引起普遍重视以来, 我国一直进行着大规模的酸雨监测和研究(Zhao et al., 1988).研究结果表明我国酸雨范围广, 对社会的经济发展和生态系统的功能影响逐年加剧, 带来了严重的经济和生态损失(Liu et al., 2013).目前我国已经成为继欧洲和北美之后的世界三大酸雨区之一, 并具有最大的强酸雨区(pH < 4.5).长江以南地区更是全球的酸雨中心(张新民等, 2010), 降雨酸化程度显著增加(Zhou et al., 2019).农业化肥的施用和化石燃料燃烧等人为排放的SO2、NOx和NHx等污染物质的增加是近年来酸沉降加剧的主要原因(Liu et al., 2013; Qiao et al., 2015).与此同时, 我国酸雨中的硫化物占比逐年减少(解淑艳等, 2012), NO3-含量增加(Cui et al., 2014), 南亚热带地区的酸雨已转变为硫酸-硝酸混合型酸雨(王志春等, 2011; Jiang et al., 2018; 郑珂等, 2019).当SO2和NOx等大气主要的污染物长期以降雨、降尘等形式进入土壤溶液中时, 将不可避免地造成土壤的酸化(潘根兴, 1990; Zhu et al., 2016; Jiang et al., 2018).土壤酸化的加剧往往会使生态系统变得更加脆弱, 并且通过抑制土壤微生物活性等影响土壤养分的循环(Liu et al., 2017), 从而影响生态系统物种的多样性和生产力(Chen et al., 2013). ...
Warming alters routing of labile and slower-turnover carbon through distinct microbial groups in boreal forest organic soils 1 2013
... 土壤微生物是土壤中最主要和活跃的生命形式(van der Heijden et al., 2008), 并且作为各种土壤过程的重要参与者, 不仅在维持植物生产力和多样性以及生态系统功能等方面有着重要的作用(van der Heijden et al., 1998; Wardle et al., 2004), 其活性和群落组成还能够作为生态系统中各种土壤过程的重要指标(Bardgett & McAlister, 1999).例如, 可用于表征土壤中微生物活性的土壤微生物生物量碳含量不仅是土壤中重要的活性有机碳库(Xu et al., 2013), 并且由于具有相较于土壤有机质而言更快的周转速率和对环境变化更为敏感等特性使其成为探究并预测当前全球变化背景下生态系统过程变化的重要指标(Spohn et al., 2016).微生物的化学计量比与环境中的养分状态存在很强的相关关系, 微生物生物量碳、氮和磷含量的比值对土壤中养分的限制种类及程度等也有很好的指示效果(Cleveland & Liptzin, 2007).此外, 由于不同微生物类群对环境变化的适应性存在差异, 随着土壤环境的变化, 其群落结构也会发生明显的改变(Kang et al., 2018).相反, 微生物群落结构揭示了微生物的生态功能差异(Guo et al., 2019), 并能在很大程度上反映并决定其生境的理化性质和养分状况(Wardle et al., 2004; Ziegler et al., 2013).细菌是土壤微生物群落中最活跃的组分, 体积小、生长迅速以及繁殖快等特性使其对养分质量具有较高的需求, 因此以细菌为主要微生物的土壤一般较为肥沃(Delgado-Baquerizo et al., 2017).真菌对环境的变化有着较高的适应性, 具有更强的养分获取能力.例如, 真菌的孢子和菌核等对环境有着较强的忍耐力, 而菌丝则可以保证其在养分贫瘠的环境下获取远处的养分(van der Heijden et al., 1998).随着土壤微生物群落组成的变化, 生态系统功能也会发生一定的变化(Wardle et al., 2004).以细菌为主要微生物的食物网一般具有较高的矿化速率, 养分周转效率较快, 而以真菌为主要微生物的食物网中养分的循环缓慢且高度保守(van der Heijden et al., 2008).因此, 土壤中真菌:细菌(F:B)变化作为微生物群落结构动态的指标, 也能够很好地反映生态系统中环境以及养分的变化情况, 具有重要的生态意义(Bardgett & McAlister, 1999; Wang et al., 2019). ...
1992-2015年鼎湖山季风常绿阔叶林群落结构动态 1 2018
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
1992-2015年鼎湖山季风常绿阔叶林群落结构动态 1 2018
... 研究还发现土壤理化性质和养分状态仅能部分解释土壤微生物群落的变化(两个土层分别为: 40.8%和47.8%), 这表明土壤酸化的加剧还可能通过其他途径影响地下部分的结构和功能, 如改变植物群落结构以及养分的输入等(Wardle et al., 2004; Jansson & Hofmockel, 2020).已有研究表明长期酸沉降对植被群落的生长和多样性有着重要的影响(Tomlinson, 2003; 刘可慧等, 2005), 而地上部分植物则可以通过改变根际周围环境以及进入到土壤中的凋落物数量和质量等对土壤酸化的适应性变化进一步改变地下部分的结构和功能, 从而直接或间接影响着土壤微生物的活性和组成(Lambers et al., 2009; Delgado-Baquerizo et al., 2017).例如, 优势树种不同的亚热带森林土壤中具有其特有的微生物群落结构和网络(Ma et al., 2019), 林下灌层丰富度与土壤地下部分群落结构的丰富度也存在显著的正相关关系(杨君珑等, 2015).在温带山地草原的研究中也发现, 植物群落组成的变化对该草原生态系统养分循环的影响, 是微生物群落结构重要的影响因子(Grayston et al., 2001).以前的研究发现, 亚热带森林生态系统对长期的气候变化缺乏抵抗能力(Zhou et al., 2013).邹顺等(2018)对本研究区域季风常绿阔叶林1 hm2永久样地过去近30年地上植被群落变化的调查发现, 其中灌木和小乔木树种个体数量显著增加, 而锥、木荷、黄果厚壳桂等优势乔木树种生物量大量减少.因此, 热带森林生态系统长期酸沉降是否会进一步影响植被的群落结构, 对地下部分的群落结构和功能将产生怎样的影响及其作用途径可能是接下来研究的重点. ...
 ,1,*
,1,*

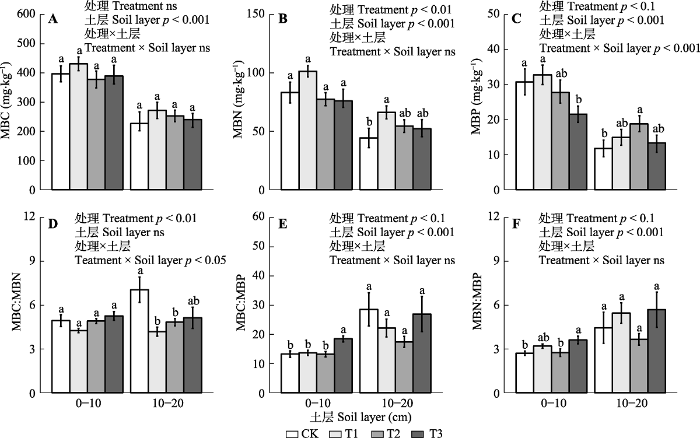 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT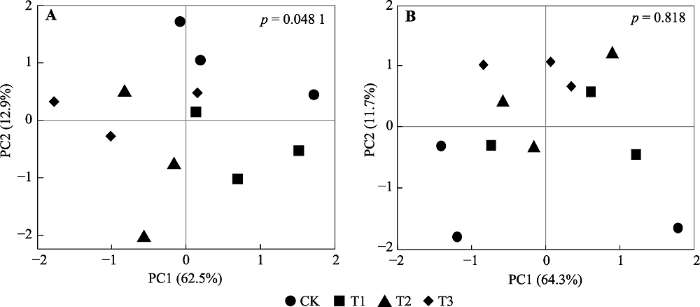 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT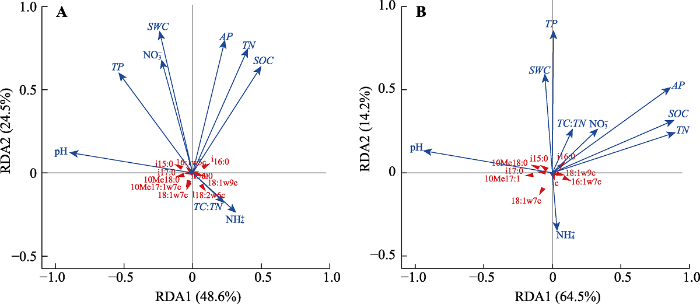 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}