Abstract Aims Altitude has prominent effects on many environmental factors, such as atmospheric pressure, temperature, precipitation, soil moisture and wind velocity. The relationship between plant functional traits and altitude are critical for predicting the effects of climate change on montane plants. Our objective is to examine the effect of altitude on community-level plant functional traits in the Qinghai Lake Basin, China. Methods Five sites were selected with 200 m increase in altitude (3 400-4 200 m) in the Qinghai Lake Basin, China. Community structure, plant functional traits, soil property and atmospheric factors were surveyed and analyzed in this study. Community-weighted mean functional traits (CWM) was calculated according to the relative abundance of species. Important findings The results showed that: (1) Community-weighted mean plant height (H), leaf dry matter content (LDMC), leaf C:N ratio (C:N) and leaf N:P ratio (N:P) decreased significantly along altitude, while specific root surface area (SRA) fluctuated with altitude. Specific leaf area (SLA), leaf nitrogen content (LNC) and leaf phosphorus content (LPC) increased significantly along altitude, while altitude had no significant effect on leaf carbon content (LCC), root tissue density (RTD) and specific root length (SRL). (2) The variation in CWM along altitude could be explained by species turnover more rather than intraspecific variability. N:P and LPC had a positive covariation, other CWM had a negative covariation. (3) Precipitation and 0-10 cm depth soil nutrients content explained the largest proportion change of SLA. Temperature and 10-20 cm depth soil nutrients content explained the largest proportion change of other CWM along altitude. Overall, these findings suggested that the plant communities in our study adapted to altitude through species turnover, and the non-dominant species tended to occupy opposite trait spaces to the dominant species in the Qinghai Lake Basin, China. Temperature and deeper soil nutrients content had significant effects on CWM along altitude. Keywords:altitude;community-level trait;leaf stoichiometry;intraspecific variability;species turnover
PDF (3328KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 向响, 黄永梅, 杨崇曜, 李泽卿, 陈慧颖, 潘莹萍, 霍佳璇, 任梁. 海拔对青海湖流域群落水平植物功能性状的影响. 植物生态学报, 2021, 45(5): 456-466. DOI: 10.17521/cjpe.2020.0140 XIANG Xiang, HUANG Yong-Mei, YANG Chong-Yao, LI Ze-Qing, CHEN Hui-Ying, PAN Ying-Ping, HUO Jia-Xuan, REN Liang. Effect of altitude on community-level plant functional traits in the Qinghai Lake Basin, China. Chinese Journal of Plant Ecology, 2021, 45(5): 456-466. DOI: 10.17521/cjpe.2020.0140
Table 3 表3 表3青海湖流域各样地的气候和土壤营养元素含量特征(平均值±标准误, n = 3) Table 3Climate and soil nutrient content characteristics of each site in the Qinghai Lake Basin, China (mean ± SE, n = 3)
序号 Serial number
年平均气温 Mean annual air temperature (℃)
年降水量 Mean annual precipitation (mm)
0-10 cm土壤全氮含量 Soil total nitrogen content in 0-10 cm layer (mg·g-1)
0-10 cm土壤全磷含量 Soil total phosphorus content in 0-10 cm layer (mg·g-1)
10-20 cm土壤全氮含量 Soil total nitrogen content in 10-20 cm layer (mg·g-1)
10-20 cm土壤全磷含量 Soil total phosphorus content in 10-20 cm layer (mg·g-1)
1
1
348
0.39 ± 0.003c
0.66 ± 0.164a
0.34 ± 0.033b
0.62 ± 0.001ab
2
-1
434
0.39 ± 0.003c
0.62 ± 0.008a
0.31 ± 0.033c
0.63 ± 0.004a
3
-2
430
0.43 ± 0.003b
0.65 ± 0.004a
0.26 ± 0.000d
0.57 ± 0.015bc
4
-4
401
0.49 ± 0.001a
0.59 ± 0.004a
0.38 ± 0.000a
0.59 ± 0.002b
5
-6
371
0.32 ± 0.003d
0.53 ± 0.005a
0.18 ± 0.033e
0.53 ± 0.007d
同一列不同小写字母表示各样地间差异显著(p < 0.05)。 Different lowercase letters in the same column indicate significant differences among sites (p < 0.05).
新窗口打开|下载原图ZIP|生成PPT 图3群落加权平均性状变异来源分解。竖线与方框顶部(种内性状变异+物种组成变化)的距离代表协变效应量, 竖线与方框相交时协变效应为负, 不相交时协变效应为正。*, p < 0.05; **, p < 0.01。性状简写见表2。
Fig. 3Decomposition of total variability in community- weighted mean functional traits. The space between the top of the column and the bar corresponds to the effect of covariation. If the bar is above the column, the covariation is positive, if the bar crosses the column, the covariation is negative. *, p < 0.05; **, p < 0.01. See Table 2 for abbreviation of traits.
Fig. 4Principal component analysis (PCA) of six environmental factors. MAP, mean annual precipitation (mm); N10, soil total nitrogen content in 0-10 cm layer (mg·g-1); N20, soil total nitrogen content in 10-20 cm layer (mg·g-1); P10, soil total phosphorus content in 0-10 cm layer (mg·g-1); P20, soil total phosphorus content in 10-20 cm layer (mg·g-1); t, mean annual air temperature (°C).
Fig. 5Proportion of variance in community-weighted mean functional traits explained by PC1 and PC2. PC1, temperature and deep soil nutrient content; PC2, precipitation and surface soil nutrient content. See Table 2 for abbreviation of traits.
3 讨论
3.1 群落水平植物功能性状随海拔的变化特征
在青海湖流域, 随着海拔升高群落H显著降低(图2F), 可能因为随着海拔升高, 植物受到低温、大风的环境胁迫, 植物群落逐渐低矮化, 直至变成垫状。植株高度对纬度梯度也存在类似的响应, 即: 随着纬度升高, 温度和降水发生显著变化, 纬度越高群落植株高度越低(Moles et al., 2009; de Frenne et al., 2013)。SLA呈现出随海拔升高先上升后下降的现象(图2C), 可能是因为随着海拔上升, 植物对水分的可获得性增加(表3), 而4 200 m处唐古红景天高山垫状植被样地位于垭口, 气温低, 降水量少, 常年大风导致了较高的土壤水分蒸发量, 植物对水分的可获得性极低, 即干旱条件下SLA显著低于湿润条件下, 这一结果与Campetella等(2019)的研究结果一致。干旱环境下的植物叶片往往会有较厚的细胞壁和叶肉层(Poorter et al., 2009), 植物叶组织相对营养留存高, 导致SLA较小、LDMC较大。在本研究中, LDMC与SLA呈现显著的负相关关系, 这与以往的认知是一致的, 然而海拔3 400和4 200 m处较其他海拔相对干旱, LDMC却较小, 可能是因为不同性状随海拔变化的主导因子有差异, LDMC还受到温度、辐射强度等因素的影响, 如: 有研究发现SLA和LDMC受到温度的影响较大(Roche et al., 2004)。本研究所选的根系形态性状发生波动变化或不随海拔发生变化(图2I-2K), 以往有研究提到: 较大的SRL与更高的养分和水分吸收率、更快的根系周转率有关, 是表征环境变化的较为敏感的性状(Geng et al., 2014), 而在本研究中, SRL的变化始终不显著, 这是令人意外的。
AsnerGP,MartinRE(2016).Convergent elevation trends in canopy chemical traits of tropical forests .Global Change Biology,22, 2216-2227. DOI:10.1111/gcb.13164URL [本文引用: 1]
BothS,RiuttaT,Timothy PaineCE,EliasDMO,CruzRS,JainA,JohnsonD,KritzlerUH,KuntzM,Majalap-LeeN,MielkeN,Montoya PillcoMX,OstleNJ,TehYA,MalhiY,BurslemDFRP(2019).Logging and soil nutrients independently explain plant trait expression in tropical forests .New Phytologist,221, 1853-1865. DOI:10.1111/nph.2019.221.issue-4URL [本文引用: 2]
CampetellaG,ChelliS,WellsteinC,FarrisE,CalviaG,SimonettiE,BorsukiewiczL,VanderplankS,MarignaniM(2019).Contrasting patterns in leaf traits of Mediterranean shrub communities along an elevation gradient: measurements matter .Plant Ecology,220, 765-776. DOI:10.1007/s11258-019-00951-y [本文引用: 3] We assessed the changes in community-weighted mean (CWM) and variability of specific leaf area (SLA) and leaf area (LA) of different Mediterranean shrub communities along an elevation gradient in the island of Sardinia (Italy). Furthermore, we explored the relative contribution of species turnover and intraspecific variation to shifts in CWM values along the gradient. Forty sampling units (5x5m) were selected in a probabilistic way along a 1300m elevation gradient which crossed four thermotypes (thermometric belts). Leaf traits were measured in each sampling unit. ANOVA and a trend test for monotonic changes in variance were used to assess, respectively, CWM differences and variability in both the leaf traits across thermotypes. Variance decomposition of CWM values was used to identify the role of inter- and intraspecific variation. SLA and LA responded differently along the studied gradient in terms of abundance-weighted mean values and variability: CWM of SLA showed the lowest values in the driest thermotype, while LA in the more humid one; SLA variability showed a significant increasing trend with increased water availability, while LA variability did not show any pattern. The contribution of intraspecific trait variation was significant for both the leaf traits, but higher for SLA, where negative covariation between inter- and intraspecific variation was detected. We highlight the importance of simultaneously considering measurements of both leaf traits to understand the functional response of communities in Mediterranean environments. Moreover, neglecting intraspecific variation in leaf traits, even along steep gradients with relevant species compositional changes, can result in the underestimation of the amount of trait variation in response to environmental changes.
CingolaniAM,CabidoM,GurvichDE,RenisonD,DíazS(2007).Filtering processes in the assembly of plant communities: Are species presence and abundance driven by the same traits? Journal of Vegetation Science,18, 911-920. DOI:10.1111/j.1654-1103.2007.tb02607.xURL [本文引用: 1]
CornelissenJHC,LavorelS,GarnierE,DíazS,BuchmannN,GurvichDE,ReichPB,ter SteegeH,MorganHD,van der HeijdenMGA,PausasJG,PoorterH(2003).A handbook of protocols for standardised and easy measurement of plant functional traits worldwide .Australian Journal of Botany,51, 335-380. DOI:10.1071/BT02124URL [本文引用: 2]
de BelloF,LavorelS,DíazS,HarringtonR,CornelissenJHC,BardgettRD,BergMP,CipriottiP,FeldCK,HeringD,da SilvaPM,PottsSG,SandinL,SousaJP,StorkeyJ,WardleDA,HarrisonPA(2010).Towards an assessment of multiple ecosystem processes and services via functional traits .Biodiversity and Conservation,19, 2873-2893. DOI:10.1007/s10531-010-9850-9URL [本文引用: 1]
de BelloF,LavorelS,AlbertCH,ThuillerW,GrigulisK,DolezalJ,JanečekŠ,LepšJ(2011).Quantifying the relevance of intraspecific trait variability for functional diversity .Methods in Ecology and Evolution,2, 163-174. DOI:10.1111/j.2041-210X.2010.00071.xURL [本文引用: 1]
de FrenneP,GraaeBJ,Rodríguez-SánchezF,KolbA,ChabrerieO,DecocqG,de KortH,De SchrijverA,DiekmannM,ErikssonO,GruwezR,HermyM,LenoirJ,PlueJ,CoomesDA,VerheyenK(2013).Latitudinal gradients as natural laboratories to infer speciesʼ responses to temperature .Journal of Ecology,101, 784-795. DOI:10.1111/jec.2013.101.issue-3URL [本文引用: 1]
de la RivaE,Pérez-RamosIM,TostoA,Navarro-FernándezC,OlmoM,MarañónT,VillarR(2016).Disentangling the relative importance of species occurrence, abundance and intraspecific variability in community assembly: a trait- based approach at the whole-plant level in Mediterranean forests .Oikos,125, 354-363. DOI:10.1111/oik.2016.v125.i3URL [本文引用: 1]
de OliveiraACP,NunesA,RodriguesRG,BranquinhoC(2020).The response of plant functional traits to aridity in a tropical dry forest .Science of the Total Environment,747, 141177. DOI:10.1016/j.scitotenv.2020.141177. DOI:10.1016/j.scitotenv.2020.141177URL [本文引用: 1]
DíazS,LavorelS,de BelloF,QuétierF,GrigulisK,RobsonTM(2007).Incorporating plant functional diversity effects in ecosystem service assessments .Proceedings of the National Academy of Sciences of the United States of America,104, 20684-20689. [本文引用: 1]
GengY,WangL,JinDM,LiuHY,HeJS(2014).Alpine climate alters the relationships between leaf and root morphological traits but not chemical traits .Oecologia,175, 445-455. DOI:10.1007/s00442-014-2919-5PMID:24633995 [本文引用: 1] Leaves and fine roots are among the most important and dynamic components of terrestrial ecosystems. To what extent plants synchronize their resource capture strategies above- and belowground remains uncertain. Existing results of trait relationships between leaf and root showed great inconsistency, which may be partly due to the differences in abiotic environmental conditions such as climate and soil. Moreover, there is currently little evidence on whether and how the stringent environments of high-altitude alpine ecosystems alter the coordination between above- and belowground. Here we measured six sets of analogous traits for both leaves and fine roots of 139 species collected from Tibetan alpine grassland and Mongolian temperate grassland. N, P and N:P ratio of leaves and fine roots were positively correlated, independent of biogeographic regions, phylogenetic affiliation or climate. In contrast, leaves and fine roots seem to regulate morphological traits more independently. The specific leaf area (SLA)-specific root length (SRL) correlation shifted from negative at sites under low temperature to positive at warmer sites. The cold climate of alpine regions may impose different constraints on shoots and roots, selecting simultaneously for high SLA leaves for rapid C assimilation during the short growing season, but low SRL roots with high physical robustness to withstand soil freezing. In addition, there might be more community heterogeneity in cold soils, resulting in multidirectional strategies of root in resource acquisition. Thus our results demonstrated that alpine climate alters the relationships between leaf and root morphological but not chemical traits.
GraaeBJ,de FrenneP,KolbA,BrunetJ,ChabrerieO,VerheyenK,PepinN,HeinkenT,ZobelM,ShevtsovaA,NijsI,MilbauA(2012).On the use of weather data in ecological studies along altitudinal and latitudinal gradients .Oikos,121, 3-19. DOI:10.1111/more.2012.121.issue-1URL [本文引用: 1]
GüsewellS(2004).N:P ratios in terrestrial plants: variation and functional significance .New Phytologist,164, 243-266. DOI:10.1111/nph.2004.164.issue-2URL [本文引用: 1]
HeJ,YangK(2011).China Meteorological Forcing Dataset .Cold and Arid Regions Science Data Center,Lanzhou. [本文引用: 1]
HuH,BaoWK,LiFL(2020).Differencial vertical distribuction of functional traits of fine roots of four cultivated tree species in the upper reaches of Minjiang River .Chinese Journal of Ecology,39, 46-56. [本文引用: 1]
HuangJJ,WangXH(2003).Leaf nutrient and structural characteristics of 32 evergreen broad-leaved species .Journal of East China Normal University (Natural Science),1, 92-97. [本文引用: 1]
JungV,AlbertCH,ViolleC,KunstlerG,LoucougarayG,SpiegelbergerT(2014).Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events .Journal of Ecology,102, 45-53. DOI:10.1111/jec.2013.102.issue-1URL [本文引用: 1]
KergunteuilA,DescombesP,GlauserG,PellissierL,RasmannS(2018).Plant physical and chemical defence variation along elevation gradients: a functional trait-based approach .Oecologia,187, 561-571. DOI:10.1007/s00442-018-4162-yPMID:29774426 [本文引用: 1] Predicting variation in plant functional traits related to anti-herbivore defences remains a major challenge in ecological research, considering that multiple traits have evolved in response to both abiotic and biotic conditions. Therefore, understanding variation in plant anti-herbivore defence traits requires studying their expression along steep environmental gradients, such as along elevation, where multiple biotic and abiotic factors co-vary. We expand on plant defence theory and propose a novel conceptual framework to address the sources of variations of plant resistance traits at the community level. We analysed elevation patterns of within-community trait dissimilarity using the RaoQ index, and the community-weighted-mean (CWM) index, on several plant functional traits: plant height, specific leaf area (SLA), leaf-dry-matter-content (LDMC), silicium content, presence of trichomes, carbon-to-nitrogen ratio (CN) and total secondary metabolite richness. We found that at high elevation, where harsh environmental conditions persist, community functional convergence is dictated by traits relating to plant growth (plant height and SLA), while divergence arises for traits relating resource-use (LDMC). At low elevation, where greater biotic pressure occurs, we found a combination of random (plant height), convergence (metabolite richness) and divergence patterns (silicium content). This framework thus combines community assembly rules of ecological filtering and niche partition with plant defence hypotheses to unravel the relationship between environmental variations, biotic pressure and the average phenotype of plants within a community.
KicheninE,WardleDA,PeltzerDA,MorseCW,FreschetGT(2013).Contrasting effects of plant inter- and intraspecific variation on community-level trait measures along an environmental gradient .Functional Ecology,27, 1254-1261. DOI:10.1111/fec.2013.27.issue-5URL [本文引用: 1]
KlimešováJ,de BelloF(2009).CLO-PLA: the database of clonal and bud bank traits of Central European flora .Journal of Vegetation Science,20, 511-516. DOI:10.1111/jvs.2009.20.issue-3URL [本文引用: 1]
KleyerM,BekkerRM,KnevelIC,BakkerJP,ThompsonK,SonnenscheinM,PoschlodP,van GroenendaelJM,KlimešL,KlimešováJ,KlotzS,RuschGM,HermyM,AdriaensD,BoedeltjeG,et al.(2008).The LEDA Traitbase: a database of life-history traits of the Northwest European flora .Journal of Ecology,96, 1266-1274. DOI:10.1111/jec.2008.96.issue-6URL [本文引用: 1]
LalibertéE,LegendreP(2010).A distance-based framework for measuring functional diversity from multiple traits .Ecology,91, 299-305. PMID:20380219 [本文引用: 1] A new framework for measuring functional diversity (FD) from multiple traits has recently been proposed. This framework was mostly limited to quantitative traits without missing values and to situations in which there are more species than traits, although the authors had suggested a way to extend their framework to other trait types. The main purpose of this note is to further develop this suggestion. We describe a highly flexible distance-based framework to measure different facets of FD in multidimensional trait space from any distance or dissimilarity measure, any number of traits, and from different trait types (i.e., quantitative, semi-quantitative, and qualitative). This new approach allows for missing trait values and the weighting of individual traits. We also present a new multidimensional FD index, called functional dispersion (FDis), which is closely related to Rao's quadratic entropy. FDis is the multivariate analogue of the weighted mean absolute deviation (MAD), in which the weights are species relative abundances. For unweighted presence-absence data, FDis can be used for a formal statistical test of differences in FD. We provide the "FD" R language package to easily implement our distance-based FD framework.
LamarqueP,LavorelS,MouchetM,QuétierF(2014).Plant trait-based models identify direct and indirect effects of climate change on bundles of grassland ecosystem services .Proceedings of the National Academy of Sciences of the United States of America,111, 13751-13756. [本文引用: 1]
LepšJ,de BelloF,ŠmilauerP,DoležalJ(2011).Community trait response to environment: disentangling species turnover vs intraspecific trait variability effects .Ecography,34, 856-863. DOI:10.1111/ecog.2011.34.issue-5URL [本文引用: 4]
MidoloG,de FrenneP,HölzelN,WellsteinC(2019).Global patterns of intraspecific leaf trait responses to elevation .Global Change Biology,25, 2485-2498. DOI:10.1111/gcb.2019.25.issue-7URL [本文引用: 1]
MitchellRM,AmesGM,WrightJP(2020).Intraspecific trait variability shapes leaf trait response to altered fire regimes .Annals of Botany,126, mcaa179. DOI:10.1093/aob/mcaa179. DOI:10.1093/aob/mcaa179 [本文引用: 1]
MolesAT,WartonDI,WarmanL,SwensonNG,LaffanSW,ZanneAE,PitmanA,HemmingsFA,LeishmanMR(2009).Global patterns in plant height .Journal of Ecology,97, 923-932. DOI:10.1111/jec.2009.97.issue-5URL [本文引用: 1]
MooneyHA,BillingsWD(1961).Comparative physiological ecology of arctic and alpine populations of Oxyria digyna .Ecological Monographs,31, 1-29. DOI:10.2307/1950744URL [本文引用: 1]
NianK,ZhangDS,ZhangYS,ChenJF,HuangM(1997).Distribution characteristics of plant communities in Qinghai Lake Basin .Science and Technology of Qinghai Agriculture and Forestry, (4), 9-12. [本文引用: 1]
NiuKC,HeJS,LechowiczMJ(2016).Grazing-induced shifts in community functional composition and soil nutrient availability in Tibetan alpine meadows .Journal of Applied Ecology,53, 1554-1564. DOI:10.1111/1365-2664.12727URL [本文引用: 1]
NunesA,KöbelM,PinhoP,MatosP,de BelloF,CorreiaO,BranquinhoC(2017).Which plant traits respond to aridity? A critical step to assess functional diversity in Mediterranean drylands .Agricultural and Forest Meteorology,239, 176-184. DOI:10.1016/j.agrformet.2017.03.007URL [本文引用: 1]
PfennigwerthAA,BaileyJK,SchweitzerJA(2017).Trait variation along elevation gradients in a dominant woody shrub is population-specific and driven by plasticity .AoB Plants,9, plx027. DOI:10.1093/aobpla/plx027. DOI:10.1093/aobpla/plx027 [本文引用: 1]
PoorterH,NiinemetsÜ,PoorterL,WrightIJ,VillarR(2009).Causes and consequences of variation in leaf mass per area (LMA): a meta-analysis .New Phytologist,182, 565-588. DOI:10.1111/nph.2009.182.issue-3URL [本文引用: 1]
R Core Team(2019).R: a language and environment for statistical computing [2020-05-11]. https://www.R-project.org/. URL [本文引用: 1]
ReichPB,OleksynJ(2004).Global patterns of plant leaf N and P in relation to temperature and latitude .Proceedings of the National Academy of Sciences of the United States of America,101, 11001-11006. [本文引用: 2]
ReichPB,RichRL,LuX,WangYP,OleksynJ(2014).Biogeographic variation in evergreen conifer needle longevity and impacts on boreal forest carbon cycle projections .Proceedings of the National Academy of Sciences of the United States of America,111, 13703-13708. [本文引用: 1]
RocheP,Díaz-BurlinsonN,GachetS(2004).Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? Plant Ecology,174, 37-48. DOI:10.1023/B:VEGE.0000046056.94523.57URL [本文引用: 1]
SiefertA,RitchieME(2016).Intraspecific trait variation drives functional responses of old-field plant communities to nutrient enrichment .Oecologia,181, 245-255. DOI:10.1007/s00442-016-3563-zPMID:26826004 [本文引用: 1] Environmental changes are expected to shift the distribution of functional trait values in plant communities through a combination of species turnover and intraspecific variation. The strength of these shifts may depend on the availability of individuals with trait values adapted to new environmental conditions, represented by the functional diversity (FD) of existing community residents or dispersal from the regional species pool. We conducted a 3-year nutrient- and seed-addition experiment in old-field plant communities to examine the contributions of species turnover and intraspecific variation to community trait shifts, focusing on four key plant functional traits: vegetative height, leaf area, specific leaf area (SLA), and leaf dry matter content (LDMC). We further examined the influence of initial FD and seed availability on the strength of these shifts. Community mean height, leaf area, and SLA increased in response to fertilization, and these shifts were driven almost entirely by intraspecific variation. The strength of intraspecific shifts in height and leaf area was positively related to initial intraspecific FD in these traits. Intraspecific trait responses to fertilization varied among species, with species of short stature displaying stronger shifts in SLA and LDMC but weaker shifts in leaf area. Trait shifts due to species turnover were generally weak and opposed intraspecific responses. Seed addition altered community taxonomic composition but had little effect on community trait shifts. These results highlight the importance of intraspecific variation for short-term community functional responses and demonstrate that the strength of these responses may be mediated by community FD.
van WijkMT,WilliamsM,GoughL,HobbieSE,ShaverGR(2003).Luxury consumption of soil nutrients: a possible competitive strategy in above-ground and below-ground biomass allocation and root morphology for slow-growing arctic vegetation? Journal of Ecology,91, 664-676. DOI:10.1046/j.1365-2745.2003.00788.xURL [本文引用: 1]
ViolleC,NavasML,VileD,KazakouE,FortunelC,HummelI,GarnierE(2007).Let the concept of trait be functional! Oikos,116, 882-892. DOI:10.1111/oik.2007.116.issue-5URL [本文引用: 1]
ViolleC,ReichPB,PacalaSW,EnquistBJ,KattgeJ(2014).The emergence and promise of functional biogeography .Proceedings of the National Academy of Sciences of the United States of America,111, 13690-13696. [本文引用: 2]
VolfM,RedmondC,AlbertÁJ,Le Bagousse-PinguetY,BiellaP,GötzenbergerL,HrázskýZ,JanečekŠ,KlimešováJ,LepšJ,ŠebelíkováL,VlasatáT,de BelloF(2016).Effects of long- and short-term management on the functional structure of meadows through species turnover and intraspecific trait variability .Oecologia,180, 941-950. DOI:10.1007/s00442-016-3548-yURL [本文引用: 1]
WangRL,YuGR,HeNP,WangQF,ZhaoN,XuZW(2016).Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China .Journal of Geographical Sciences,26,15-26. DOI:10.1007/s11442-016-1251-xURL [本文引用: 1]
ZhouXL,GuoZ,ZhangPF,DuGZ(2018).Shift in community functional composition following nitrogen fertilization in an alpine meadow through intraspecific trait variation and community composition change .Plant and Soil,431, 289-302. DOI:10.1007/s11104-018-3771-xURL [本文引用: 1]
Convergent elevation trends in canopy chemical traits of tropical forests 1 2016
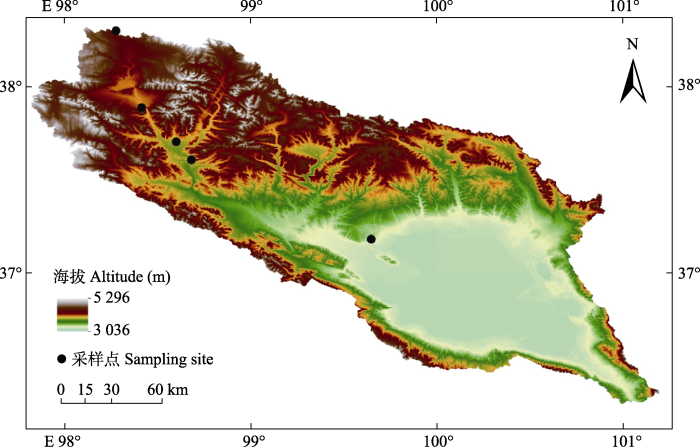
... 青海湖流域位于青藏高原东北部(36.25°- 38.33° N, 97.83°-101.33° E, 海拔3 194-5 170 m), 属于高寒大陆性气候, 年平均气温-1.1-4.0 ℃, 年降水量291-597 mm, 年蒸发量1 300-2 000 mm (年奎等, 1997).随着海拔升高, 植被类型依次为温性草原、高寒草原、高寒草甸、高寒荒漠和高山稀疏植被等(陈桂琛和彭敏, 1993), 建群种分别为西北针茅(Stipa sareptana)、紫花针茅(Stipa purpurea)、高山嵩草(Kobresia pygmaea)、唐古红景天(Rhodiola tangutica)和四蕊山莓草(Sibbaldia tetrandra)等. ...
青海湖地区植被及其分布规律 1 1993
... 青海湖流域位于青藏高原东北部(36.25°- 38.33° N, 97.83°-101.33° E, 海拔3 194-5 170 m), 属于高寒大陆性气候, 年平均气温-1.1-4.0 ℃, 年降水量291-597 mm, 年蒸发量1 300-2 000 mm (年奎等, 1997).随着海拔升高, 植被类型依次为温性草原、高寒草原、高寒草甸、高寒荒漠和高山稀疏植被等(陈桂琛和彭敏, 1993), 建群种分别为西北针茅(Stipa sareptana)、紫花针茅(Stipa purpurea)、高山嵩草(Kobresia pygmaea)、唐古红景天(Rhodiola tangutica)和四蕊山莓草(Sibbaldia tetrandra)等. ...
Temporal intraspecific trait variability drives responses of functional diversity to interannual aridity variation in grasslands 1 2019
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
Hierarchical partitioning 1 1991
... 对各个CWM进行K-W正态性检验, SLA、LDMC、SRL、RTD、C:N和N:P的群落加权平均值均符合正态分布, H、LNC、LCC、LPC和SRA的群落加权平均值进行lg转换后符合正态分布, 然后进行单因素方差分析, 采用最小显著差异(LSD)法进行差异性检验.基于嵌套方差分析和平方和分解的方法(Lepš et al., 2011), 将所有CWM分离并量化为物种组成变化和种内性状变异.为了更好地量化环境因子对各个CWM的影响, 本研究先将6个环境因子数据标准化, 利用主成分分析(PCA), 提取前2个主成分PC1和PC2, 后使用基于分层划分法的多元回归分析(Chevan & Sutherland, 1991; Both et al., 2019)量化PC1和PC2对CWM变化的解释率. ...
Filtering processes in the assembly of plant communities: Are species presence and abundance driven by the same traits? 1 2007
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide 2 2003
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
Towards an assessment of multiple ecosystem processes and services via functional traits 1 2010
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Quantifying the relevance of intraspecific trait variability for functional diversity 1 2011
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
Latitudinal gradients as natural laboratories to infer speciesʼ responses to temperature 1 2013
... 在青海湖流域, 随着海拔升高群落H显著降低(图2F), 可能因为随着海拔升高, 植物受到低温、大风的环境胁迫, 植物群落逐渐低矮化, 直至变成垫状.植株高度对纬度梯度也存在类似的响应, 即: 随着纬度升高, 温度和降水发生显著变化, 纬度越高群落植株高度越低(Moles et al., 2009; de Frenne et al., 2013).SLA呈现出随海拔升高先上升后下降的现象(图2C), 可能是因为随着海拔上升, 植物对水分的可获得性增加(表3), 而4 200 m处唐古红景天高山垫状植被样地位于垭口, 气温低, 降水量少, 常年大风导致了较高的土壤水分蒸发量, 植物对水分的可获得性极低, 即干旱条件下SLA显著低于湿润条件下, 这一结果与Campetella等(2019)的研究结果一致.干旱环境下的植物叶片往往会有较厚的细胞壁和叶肉层(Poorter et al., 2009), 植物叶组织相对营养留存高, 导致SLA较小、LDMC较大.在本研究中, LDMC与SLA呈现显著的负相关关系, 这与以往的认知是一致的, 然而海拔3 400和4 200 m处较其他海拔相对干旱, LDMC却较小, 可能是因为不同性状随海拔变化的主导因子有差异, LDMC还受到温度、辐射强度等因素的影响, 如: 有研究发现SLA和LDMC受到温度的影响较大(Roche et al., 2004).本研究所选的根系形态性状发生波动变化或不随海拔发生变化(图2I-2K), 以往有研究提到: 较大的SRL与更高的养分和水分吸收率、更快的根系周转率有关, 是表征环境变化的较为敏感的性状(Geng et al., 2014), 而在本研究中, SRL的变化始终不显著, 这是令人意外的. ...
Disentangling the relative importance of species occurrence, abundance and intraspecific variability in community assembly: a trait- based approach at the whole-plant level in Mediterranean forests 1 2016
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
The response of plant functional traits to aridity in a tropical dry forest 1 2020
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Incorporating plant functional diversity effects in ecosystem service assessments 1 2007
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
植物属性地理的研究进展与展望 1 2018
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Intraspecific trait variability mediates the response of subalpine grassland communities to extreme drought events 1 2014
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
Plant physical and chemical defence variation along elevation gradients: a functional trait-based approach 1 2018
CLO-PLA: the database of clonal and bud bank traits of Central European flora 1 2009
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
The LEDA Traitbase: a database of life-history traits of the Northwest European flora 1 2008
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
The use of “altitude” in ecological research 1 2007
A distance-based framework for measuring functional diversity from multiple traits 1 2010
... 在R 3.4.3中使用“FD”包计算各个CWM (Laliberté & Legendre, 2010; R Core Team, 2019), 使用Lepš等(2011)提出的函数分离和量化CWM, 使用“heir.part”包实现分层划分法的计算(Both et al., 2019).数据的正态性检验、数据的标准化、以及ANOVA和PCA在SPSS 20.0中进行.所有的图形均利用OriginPro 2017绘制. ...
Plant trait-based models identify direct and indirect effects of climate change on bundles of grassland ecosystem services 1 2014
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Community trait response to environment: disentangling species turnover vs intraspecific trait variability effects 4 2011
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
... 对各个CWM进行K-W正态性检验, SLA、LDMC、SRL、RTD、C:N和N:P的群落加权平均值均符合正态分布, H、LNC、LCC、LPC和SRA的群落加权平均值进行lg转换后符合正态分布, 然后进行单因素方差分析, 采用最小显著差异(LSD)法进行差异性检验.基于嵌套方差分析和平方和分解的方法(Lepš et al., 2011), 将所有CWM分离并量化为物种组成变化和种内性状变异.为了更好地量化环境因子对各个CWM的影响, 本研究先将6个环境因子数据标准化, 利用主成分分析(PCA), 提取前2个主成分PC1和PC2, 后使用基于分层划分法的多元回归分析(Chevan & Sutherland, 1991; Both et al., 2019)量化PC1和PC2对CWM变化的解释率. ...
... 在R 3.4.3中使用“FD”包计算各个CWM (Laliberté & Legendre, 2010; R Core Team, 2019), 使用Lepš等(2011)提出的函数分离和量化CWM, 使用“heir.part”包实现分层划分法的计算(Both et al., 2019).数据的正态性检验、数据的标准化、以及ANOVA和PCA在SPSS 20.0中进行.所有的图形均利用OriginPro 2017绘制. ...
Intraspecific trait variability shapes leaf trait response to altered fire regimes 1 2020
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
Global patterns in plant height 1 2009
... 在青海湖流域, 随着海拔升高群落H显著降低(图2F), 可能因为随着海拔升高, 植物受到低温、大风的环境胁迫, 植物群落逐渐低矮化, 直至变成垫状.植株高度对纬度梯度也存在类似的响应, 即: 随着纬度升高, 温度和降水发生显著变化, 纬度越高群落植株高度越低(Moles et al., 2009; de Frenne et al., 2013).SLA呈现出随海拔升高先上升后下降的现象(图2C), 可能是因为随着海拔上升, 植物对水分的可获得性增加(表3), 而4 200 m处唐古红景天高山垫状植被样地位于垭口, 气温低, 降水量少, 常年大风导致了较高的土壤水分蒸发量, 植物对水分的可获得性极低, 即干旱条件下SLA显著低于湿润条件下, 这一结果与Campetella等(2019)的研究结果一致.干旱环境下的植物叶片往往会有较厚的细胞壁和叶肉层(Poorter et al., 2009), 植物叶组织相对营养留存高, 导致SLA较小、LDMC较大.在本研究中, LDMC与SLA呈现显著的负相关关系, 这与以往的认知是一致的, 然而海拔3 400和4 200 m处较其他海拔相对干旱, LDMC却较小, 可能是因为不同性状随海拔变化的主导因子有差异, LDMC还受到温度、辐射强度等因素的影响, 如: 有研究发现SLA和LDMC受到温度的影响较大(Roche et al., 2004).本研究所选的根系形态性状发生波动变化或不随海拔发生变化(图2I-2K), 以往有研究提到: 较大的SRL与更高的养分和水分吸收率、更快的根系周转率有关, 是表征环境变化的较为敏感的性状(Geng et al., 2014), 而在本研究中, SRL的变化始终不显著, 这是令人意外的. ...
Comparative physiological ecology of arctic and alpine populations of Oxyria digyna 1 1961
Biogeographic variation in evergreen conifer needle longevity and impacts on boreal forest carbon cycle projections 1 2014
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Congruency analysis of species ranking based on leaf traits: Which traits are the more reliable? 1 2004
... 在青海湖流域, 随着海拔升高群落H显著降低(图2F), 可能因为随着海拔升高, 植物受到低温、大风的环境胁迫, 植物群落逐渐低矮化, 直至变成垫状.植株高度对纬度梯度也存在类似的响应, 即: 随着纬度升高, 温度和降水发生显著变化, 纬度越高群落植株高度越低(Moles et al., 2009; de Frenne et al., 2013).SLA呈现出随海拔升高先上升后下降的现象(图2C), 可能是因为随着海拔上升, 植物对水分的可获得性增加(表3), 而4 200 m处唐古红景天高山垫状植被样地位于垭口, 气温低, 降水量少, 常年大风导致了较高的土壤水分蒸发量, 植物对水分的可获得性极低, 即干旱条件下SLA显著低于湿润条件下, 这一结果与Campetella等(2019)的研究结果一致.干旱环境下的植物叶片往往会有较厚的细胞壁和叶肉层(Poorter et al., 2009), 植物叶组织相对营养留存高, 导致SLA较小、LDMC较大.在本研究中, LDMC与SLA呈现显著的负相关关系, 这与以往的认知是一致的, 然而海拔3 400和4 200 m处较其他海拔相对干旱, LDMC却较小, 可能是因为不同性状随海拔变化的主导因子有差异, LDMC还受到温度、辐射强度等因素的影响, 如: 有研究发现SLA和LDMC受到温度的影响较大(Roche et al., 2004).本研究所选的根系形态性状发生波动变化或不随海拔发生变化(图2I-2K), 以往有研究提到: 较大的SRL与更高的养分和水分吸收率、更快的根系周转率有关, 是表征环境变化的较为敏感的性状(Geng et al., 2014), 而在本研究中, SRL的变化始终不显著, 这是令人意外的. ...
Intraspecific trait variation drives functional responses of old-field plant communities to nutrient enrichment 1 2016
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
A global meta-analysis of the relative extent of intraspecific trait variation in plant communities 1 2015
Functional diversity enhances the resistance of ecosystem multifunctionality to aridity in Mediterranean drylands 1 2015
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Luxury consumption of soil nutrients: a possible competitive strategy in above-ground and below-ground biomass allocation and root morphology for slow-growing arctic vegetation? 1 2003
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
The emergence and promise of functional biogeography 2 2014
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
... ).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Effects of long- and short-term management on the functional structure of meadows through species turnover and intraspecific trait variability 1 2016
... 群落水平植物功能性状的变化来源包括物种组成变化和种内性状变异, 早期的“群落加权平均性状-环境”关系研究通常假设物种组成变化大于种内性状变异, 最为典型的例子是许多研究从较为广泛的性状数据库中提取数据进行分析(Cornelissen et al., 2003; Kleyer et al., 2008; Klimešová & de Bello, 2009).然而, 大量研究表明, 种内性状变异对于物种存活以及群落动态发挥着重要作用, 因此, 分离并量化植物种内性状变异、物种组成变化及二者的协变效应至关重要(de Bello et al., 2011; Lepš et al., 2011; Mitchell et al., 2020).大量研究基于自然或半自然的环境梯度, 将群落水平植物功能性状进行了分离和量化, 如: 长时间定点观测生态系统中群落水平植物功能性状对年际气候变化的响应特征(Chen et al., 2019); 或依据土壤水分梯度、人为管理模式和营养元素添加梯度等进行CWM的分离和量化(de la Riva et al., 2016; Siefert & Ritchie, 2016; Volf et al., 2016), 这些相关研究有助于更好地了解哪些生态过程调控着植物群落的集散以及植物群落对环境的适应性(Cingolani et al., 2007; Jung et al., 2014). ...
Latitudinal variation of leaf morphological traits from species to communities along a forest transect in eastern China 1 2016
... 植物功能性状是指植物的生理、生态和物候特征, 这些特征通常在个体水平上可测量, 并通过影响其生长、繁殖和存活来影响植物的行为和适应性(Violle et al., 2007).为了更好地描述植物群落对环境变化的响应, Garnier等(2004)根据质量比假说, 将个体水平植物功能性状与群落物种相对丰富度结合, 计算得到群落加权平均性状(CWM), 代表群落水平植物功能性状.全球气候变化背景下基于植物功能性状分析植被对环境变化的响应备受关注(Lamarque et al., 2014; Violle et al., 2014; 黄永梅等, 2018).大量研究在全球、大洲等大尺度上探究“性状-环境”关系, 但是这些研究多是基于个体水平植物功能性状开展, 如: Reich和Oleksyn (2004)研究了叶片氮磷含量、叶片氮磷比与年平均气温和纬度的关系; Reich等(2014)在研究北美洲针叶林叶寿命对碳循环的影响时, 分析了叶寿命对温度的响应规律.随着研究的深入, 群落水平植物功能性状, 尤其是群落加权平均植物功能性状被证实能够对环境过滤作用(如: 气候、扰动机制或生物相互作用)作出响应, 并影响主要的生态系统过程(如初级生产力或凋落物分解速率)(Díaz et al., 2007; de Bello et al., 2010; Mouillot et al., 2013; Valencia et al., 2015; Nunes et al., 2017).群落水平植物功能性状也被证实可以很好地与环境变化、生态系统服务相联系, 正成为全球变化生态学的一个重要研究工具(Violle et al., 2014).在区域尺度和局地尺度上, 植物群落调查数据更易获得, 因此“群落水平性状-环境”关系研究目前多在区域和局地尺度进行, 如: de Oliveira等(2020)在巴西研究了13个CWM对干旱的响应特征; Niu等(2016)在青藏高原东部研究了放牧强度对CWM的影响; Wang等(2016)在中国东部森林样带研究发现叶片CWM与纬度具有很好的相关性等. ...
Shift in community functional composition following nitrogen fertilization in an alpine meadow through intraspecific trait variation and community composition change 1 2018
 ,*, 杨崇曜, 李泽卿, 陈慧颖, 潘莹萍, 霍佳璇, 任梁北京师范大学地表过程与资源生态国家重点实验室, 北京师范大学地理科学学部, 北京 100875
,*, 杨崇曜, 李泽卿, 陈慧颖, 潘莹萍, 霍佳璇, 任梁北京师范大学地表过程与资源生态国家重点实验室, 北京师范大学地理科学学部, 北京 100875

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT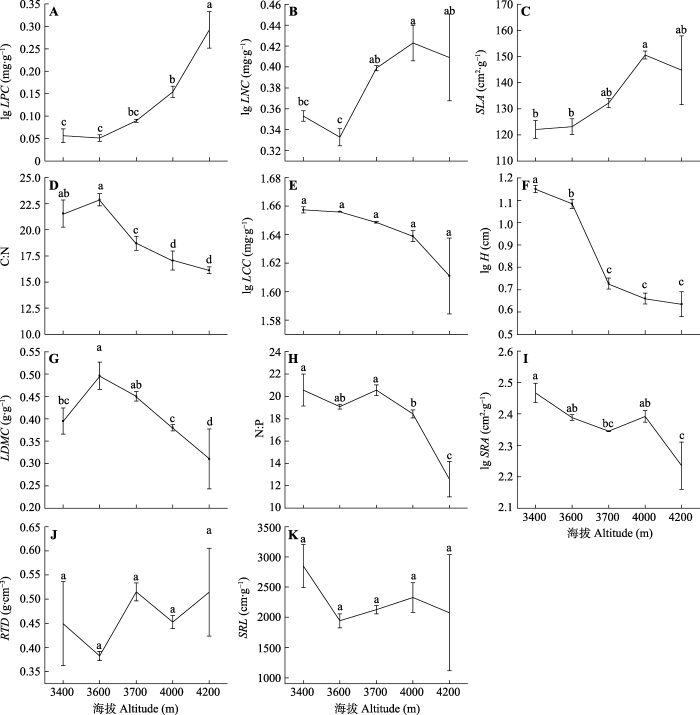 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT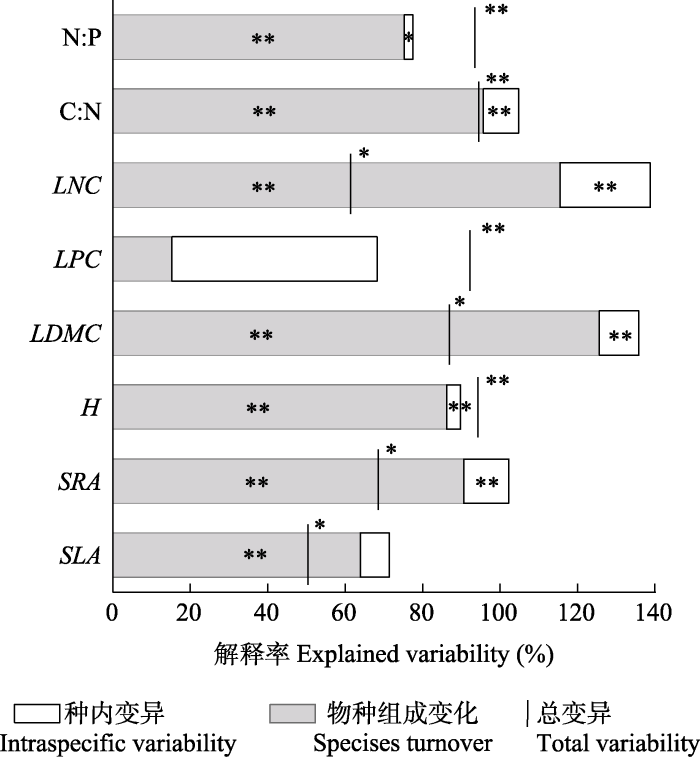 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT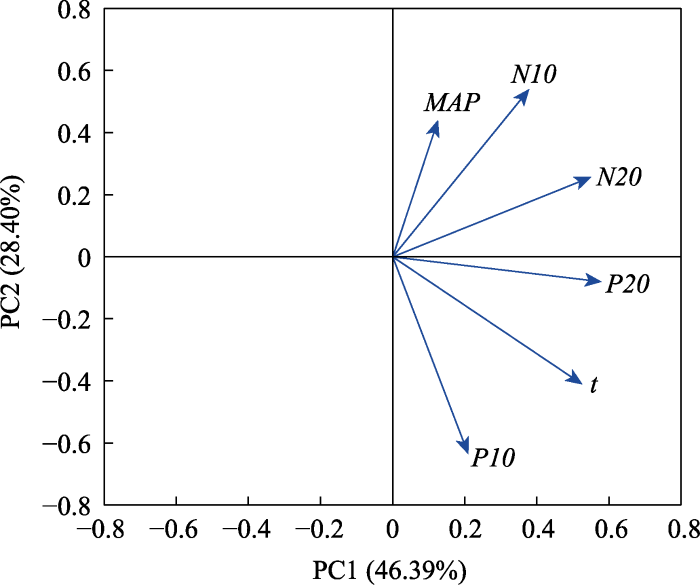 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT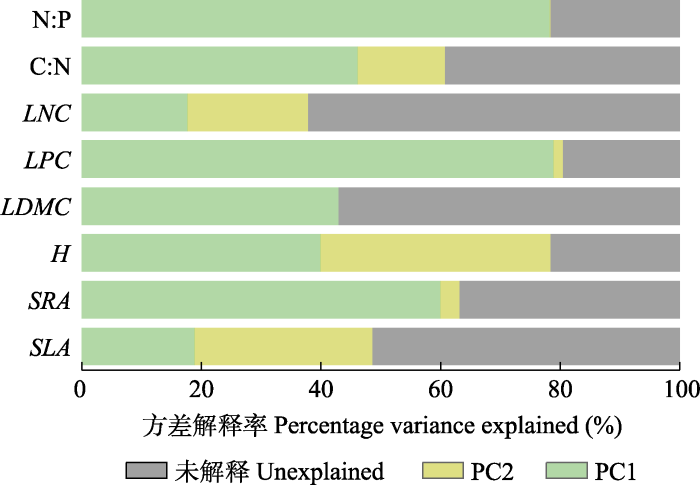 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}