Effect of soil organic matter chemical compositions on soil protease and urease activity in alpine grassland soils in Northern Xizang, China
Shu-Qin MA1, Zi-Wei WANG2,3, You-Chao CHEN4, Xu-Yang LU,2,*1College of Tourism, Henan Normal University, Xinxiang, Henan 453007, China 2Key Laboratory of Mountain Surface Processes and Ecological Regulation, Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China 3University of Chinese Academy of Science, Beijing 100049, China 4Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan 430074, China
Abstract Aims Soil enzymes play an important role in the process of soil nutrient transformation. The main purpose of this study is to explore the chemical composition of soil organic matter and its effect on soil protease and urease activity in alpine grasslands in Northern Xizang, China. Methods The pyrolysis Gas Chromatograph/Mass Spectrometer (Py-GC/MS) was used to obtain the chemical compositions of soil organic matter, and to analyze the relationships between soil chemical compositions and soil enzyme activity in five alpine grasslands, including alpine meadow, alpine steppe, alpine meadow steppe, alpine desert steppe and alpine desert ecosystems. Important findings The results showed that the enzyme activities among five alpine grassland soils (0-15 cm) were different. Soil urease activity was significantly higher than soil protease activity in the alpine desert steppe, while the difference between the urease and protease activities was not significant in other types of alpine grasslands. Soil protease activity was significantly different among five alpine grassland types, but soil urease activity was not. The correlation analysis showed that soil protease activity was closely related to the relative abundance of alkanes, alkenes and aromatics in soil organic matter and the ratio of furfural to pyrrole. However, the correlation relationships between urease activity and soil organic matter chemical compositions were not significant. The results indicated that alpine grassland type and soil organic matter chemistry were the key factors affecting soil protease activity, but their effects on soil urease activity was non-significant, which calls for further study on the influencing factors on soil urease activity. Keywords:alpine meadow;alpine steppe;soil organic matter chemical composition;enzyme activity;Pyrolysis Gas Chromatograph/Mass Spectrometer (Py-GC/MS)
PDF (5315KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 马书琴, 汪子微, 陈有超, 鲁旭阳. 藏北高寒草地土壤有机质化学组成对土壤蛋白酶和脲酶活性的影响. 植物生态学报, 2021, 45(5): 516-527. DOI: 10.17521/cjpe.2020.0169 MA Shu-Qin, WANG Zi-Wei, CHEN You-Chao, LU Xu-Yang. Effect of soil organic matter chemical compositions on soil protease and urease activity in alpine grassland soils in Northern Xizang, China. Chinese Journal of Plant Ecology, 2021, 45(5): 516-527. DOI: 10.17521/cjpe.2020.0169
土壤酶是土壤中具有生物活性的蛋白质, 主要来自土壤中动植物和微生物活动以及有机残体的分解物, 在生态系统物质循环和能量流动中扮演重要角色(Rao et al., 2016)。土壤酶和微生物共同推动着土壤的生物化学过程, 对有机残体分解和无机化合物的转化以及土壤肥力的演化具有重要作用(刘琳等, 2011; Kuypers et al., 2018)。土壤蛋白酶和脲酶是土壤中参与有机态氮转化的两种关键酶, 蛋白酶作用于有机氮的矿化, 将土壤中植物残体中的蛋白质和肽类物质分解为氨基酸(Watanabe, 2009); 脲酶则是将土壤中有机氮转化为植物和微生物可以利用的无机氮, 也是唯一可以水解土壤中尿素的水解酶, 其分解尿素的速度是尿素自然分解速度的1 014倍(余燕玲等, 2008; Fraser et al., 2013)。因此, 土壤蛋白酶和脲酶的活性直接影响着土壤中氮转化的强度和土壤的供氮能力(Du et al., 2020)。王学霞等(2018)研究青藏高原退化高寒草地土壤氮矿化影响因素发现, 土壤硝化速率和氨化速率与土壤蛋白酶和脲酶活性密切相关; 而在一个39年长期施肥的热带稻田中, 研究不同季节施肥对土壤温室气体释放的影响时发现, 冬季脲酶活性与土壤N2O的排放密切相关(Bhattacharyya et al., 2013); 但在挪威的欧洲云杉(Picea abies)林土壤有机层的研究发现, 土壤蛋白酶并未有效地降解较为顽固的单宁蛋白复合物中的蛋白质(Adamczyk et al., 2009)。因此, 土壤蛋白酶和脲酶对氮的作用方式在不同生态系统中存在一定差异, 其活性可能与基质中的养分浓度和有效性密切相关。
利用Py-GC/MS技术测定土壤有机质化学成分图谱, 用质谱数据库(National Institute of Standards and Technology, NIST 2014)比对获得化学成分。根据已有的研究结果(Grandy et al., 2007; Yassir & Buurman, 2012; Oliveira et al., 2016; Ma et al., 2018)和这些成分的化学结构相似性, 将这些成分归类为烷烃、烯烃、芳香烃、脂肪酸、木质素、含氮化合物、酚类、多环芳烃、多糖、萜烯类和其他成分(表1), 此外, 将未被识别出的12个峰归类为未知物质。土壤有机质化学成分相对丰度用各成分的相对面积表示。另外, 糠醛和吡咯的相对丰度之比、吡咯和酚的相对丰度之比分别被认为是有机质易矿化组分和难矿化组分的矿化指数; 苯和甲苯相对丰度之比与土壤腐殖化强度相关, 是由于苯来源于缩聚芳香族物质, 甲苯则主要来源于非缩聚芳香族链状物质(Aranda et al., 2015)。
Table 1 表1 表1藏北高寒草地土壤有机质化学成分 Table 1Soil organic matter chemical compositions in Northern Xizang
Table 2 表2 表2藏北不同高寒草地类型土壤酶活性(平均值±标准误) Table 2Enzyme activity in different alpine grassland soils in Northern Xizang, China (mean ± SE)
高寒草地类型 Alpine grassland type
蛋白酶 Protease (μg·g-1·h-1)
脲酶 Urease (mg·kg-1·h-1)
高寒草甸 Alpine meadow
170.20 ± 55.28Aa
96.25 ± 52.08Aa
高寒草原 Alpine steppe
117.84 ± 36.68Aab
130.20 ± 41.59Aa
高寒荒漠 Alpine desert
85.60 ± 10.02Abc
80.07 ± 1.97Aa
高寒草甸草原 Alpine meadow steppe
79.97 ± 43.97Abc
68.53 ± 6.22Aa
高寒荒漠草原 Alpine desert steppe
31.00 ± 20.81Bc
85.86 ± 14.31Aa
不同大写字母表示同一草地类型土壤不同酶的活性之间差异显著(p < 0.05), 不同小写字母表示土壤酶在不同草地类型之间的差异显著(p < 0.05)。 Different uppercase letters indicate significant difference between the two enzymes in the same grassland type (p < 0.05), while different lowercase letters indicate significant difference among different grassland types for the same type of enzyme (p < 0.05).
Table 3 表3 表3藏北不同高寒草地类型土壤净氮矿化速率、净氨化速率和净硝化速率(平均值±标准误) Table 3Net nitrogen (N) mineralization rate, net ammonification rate and net nitrification rate in different alpine grassland soils in Northern Xizang, China (mean ± SE)
高寒草地类型 Alpine grassland type
净氮矿化速率 Net N mineralization rate (mg·g-1·d-1)
净氨化速率 Net ammonification rate (mg·g-1·d-1)
净硝化速率 Net nitrification rate (mg·g-1·d-1)
高寒草甸 Alpine meadow
0.33 ± 0.17a
0.010 ± 0.004a
0.32 ± 0.16a
高寒草原 Alpine steppe
0.15 ± 0.06b
0.008 ± 0.002b
0.14 ± 0.06b
高寒荒漠 Alpine desert
0.05 ± 0.02b
0.003 ± 0.002c
0.05 ± 0.01b
高寒草甸草原 Alpine meadow steppe
0.12 ± 0.05b
0.004 ± 0.001c
0.12 ± 0.05b
高寒荒漠草原 Alpine desert steppe
0.05 ± 0.02b
0.004 ± 0.002c
0.04 ± 0.02b
不同小写字母表示不同草地类型之间的差异性(p < 0.05)。 Different lowercase letters indicate significant difference among different grassland types (p < 0.05).
Table 4 表4 表4藏北高寒草地土壤酶活性和土壤氮转化速率的关系 Table 4Relationship between soil enzyme activity and soil nitrogen (N) transformation rate in alpine grasslands in Northern Xizang, China
Table 5 表5 表5藏北高寒草地土壤有机质化学组成和酶活性的相关性 Table 5Relationship between soil organic matter compositions and enzyme activity in alpine grasslands in Northern Xizang, China
AdamczykB,KitunenV,SmolanderA(2009).Polyphenol oxidase, tannase and proteolytic activity in relation to tannin concentration in the soil organic horizon under silver birch and Norway spruce .Soil Biology & Biochemistry,41, 2085-2093. DOI:10.1016/j.soilbio.2009.07.018URL [本文引用: 1]
AndreettaA,MacciC,GiansoldatiV,MasciandaroG,CarnicelliS(2013).Microbial activity and organic matter composition in Mediterranean humus forms .Geoderma,209, 198-208. [本文引用: 1]
AvazpoorZ,MoradiM,BasiriR,MirzaeiJ,Taghizadeh-MehrjardiR,KerryR(2019).Soil enzyme activity variations in riparian forests in relation to plant species and soil depth .Arabian Journal of Geosciences,12, 708. DOI:10.1007/s12517-019-4910-2. DOI:10.1007/s12517-019-4910-2URL [本文引用: 3]
BaiJB,XuXL,FuG,SongMH,HeYT,JiangJ(2011).Effects of temperature and nitrogen input on nitrogen mineralization in alpine soils on the Tibetan Plateau .Journal of Anhui Agricultural Sciences,39, 14698-14700. [本文引用: 2]
BaligarVC,WrightRJ,FageriaNK,PittaGVE(1999).Enzyme activities in Cerrado soils of Brazil .Communications in Soil Science and Plant Analysis,30, 1551-1560. DOI:10.1080/00103629909370306URL [本文引用: 1]
BaoY,GaoY,ZengXM,YuanP,SiYT,ChenYM,ChenYY(2018).Relationships between carbon and nitrogen contents and enzyme activities in soil of three typical subtropical forests in China .Chinese Journal of Plant Ecology,42, 508-516. DOI:10.17521/cjpe.2017.0311URL [本文引用: 1]
BhattacharyyaP,NayakAK,MohantyS,TripathiR,ShahidM,KumarA,RajaR,PandaBB,RoyKS,NeogiS,DashPK,ShuklaAK,RaoKS(2013).Greenhouse gas emission in relation to labile soil C, N pools and functional microbial diversity as influenced by 39 years long-term fertilizer management in tropical rice .Soil & Tillage Research,129, 93-105. [本文引用: 1]
CaiH,ShenRF(2005).Determination of soil protease activity with modified ninhydrin colorimetry .Acta Pedologica Sinica,42, 306-313. [本文引用: 1]
CaiYJ,WangXD,DingWX,TianLL,ZhaoH,LuXY(2013).Potential short-term effects of yak and Tibetan sheep dung on greenhouse gas emissions in two alpine grassland soils under laboratory conditions .Biology and Fertility of Soils,49, 1215-1226. DOI:10.1007/s00374-013-0821-7URL [本文引用: 2]
ChenQY,NiuB,HuYL,LuoTX,ZhangGX(2020).Warming and increased precipitation indirectly affect the composition and turnover of labile-fraction soil organic matter by directly affecting vegetation and microorganisms .Science of the Total Environment,714, 136787. DOI:10.1016/j.scitotenv.2020.136787. DOI:10.1016/j.scitotenv.2020.136787URL [本文引用: 1]
ChenYC,MaSQ,SunJ,WangXD,ChengGW,LuXY(2017).Chemical diversity and incubation time affect non-additive responses of soil carbon and nitrogen cycling to litter mixtures from an alpine steppe soil .Soil Biology & Biochemistry,109, 124-134. DOI:10.1016/j.soilbio.2017.02.007URL [本文引用: 1]
CuiHX,WeiNH(1997).The geographical distribution of grassland types in Xizang, China .Foreign Animal Husbandry—Grassland and Pastures, (4), 11-17. [本文引用: 1]
CuiJF,HoldenNM(2015).The relationship between soil microbial activity and microbial biomass, soil structure and grassland management .Soil and Tillage Research,146, 32-38. DOI:10.1016/j.still.2014.07.005URL [本文引用: 1]
DerenneS,QuénéaK(2015).Analytical pyrolysis as a tool to probe soil organic matter .Journal of Analytical and Applied Pyrolysis,111, 108-120. DOI:10.1016/j.jaap.2014.12.001URL [本文引用: 1]
DineshR,ShomeBR,ShomeR,BandyopadhyayAK(1998).Soil enzymes in the mangroves: activities and their relation to relevant soil properties .Current Science,75, 510-512. [本文引用: 1]
DuEZ,TerrerC,PellegriniAFA,AhlstromA,van LissaCJ,ZhaoX,XiaN,WuXH,JacksonRB(2020).Global patterns of terrestrial nitrogen and phosphorus limitation .Nature Geoscience,13, 221-226. DOI:10.1038/s41561-019-0530-4URL [本文引用: 1]
FraserFC,HallettPD,WookeyPA,HartleyIP,HopkinsDW(2013).How do enzymes catalysing soil nitrogen transformations respond to changing temperatures? Biology and Fertility of Soils,49, 99-103. DOI:10.1007/s00374-012-0722-1URL [本文引用: 1]
GaoQZ,LiY,WanYF,QinXB,JiangcunWZ,LiuYH(2009).Dynamics of alpine grassland NPP and its response to climate change in Northern Tibet .Climatic Change,97, 515-528. DOI:10.1007/s10584-009-9617-zURL [本文引用: 1]
GrandyAS,NeffJC,WeintraubMN(2007).Carbon structure and enzyme activities in alpine and forest ecosytems .Soil Biology & Biochemistry,39, 2701-2711. DOI:10.1016/j.soilbio.2007.05.009URL [本文引用: 2]
GongSW,ZhangT,GuoR,CaoHB,ShiLX,GuoJX,SunW(2015).Response of soil enzyme activity to warming and nitrogen addition in a meadow steppe .Soil Research,53, 242-252. DOI:10.1071/SR14140URL [本文引用: 1]
HuangHL,ZongN,HeNP,TianJ(2019).Characteristics of soil enzyme stoichiometry along an altitude gradient on Qinghai-Tibet Plateau alpine meadow, China .Chinese Journal of Applied Ecology,30, 3689-3696. [本文引用: 1]
HuangJ,LiZ,ZhangJ(2012).Improvement of indophenol blue colorimetric method on activity of urease in soil .Journal of Civil, Architectural & Environment Engineering,34, 102-107. [本文引用: 1]
HuangYF,ShuYG,XiaoSY,ChenMJ(2020).Quantification of soil nutrient levels and enzyme activities in different grassland categories in karst mountains .Acta Prataculturae Sinica,29, 93-104. [本文引用: 1]
LiQ,SunYN,LiL,LiYK,DuYG,GuoXW,YangYS,CaoGM(2019).Changes of soil enzyme activities and nutrients across different succession stages of grazing alpine Kobresia grassland .Chinese Journal of Applied Ecology,30, 2267-2274. [本文引用: 3]
LiYY,DongSK,LiuSL,ZhouHK,GaoQZ,CaoGM,WangXX,SuXK,ZhangY,TangL,ZhaoHD,WuXY(2015).Seasonal changes of CO2, CH4 and N2O fluxes in different types of alpine grassland in the Qinghai-Tibetan Plateau of China .Soil Biology & Biochemistry,80, 306-314. DOI:10.1016/j.soilbio.2014.10.026URL [本文引用: 1]
LiZL,ZengZQ,TianDS,WangJS,FuZ,ZhangFY,ZhangRY,ChenWN,LuoYQ,NiuSL(2020).Global patterns and controlling factors of soil nitrification rate .Global Change Biology,26, 4147-4157. DOI:10.1111/gcb.v26.7URL [本文引用: 2]
LiangB,LiJL,YangXY,ZhouJB(2016).Effect of fertilization on extractable organic nitrogen in wheat monoculture cropping systems .Acta Ecologica Sinica,36, 4430-4437. [本文引用: 1]
LiuL,ZhuX,SunG,LuoP,WangB(2011).Effects of simulated warming and fertilization on activities of soil enzymes in alpine meadow .Pratacultural Science,28, 1405-1410. [本文引用: 1]
LiuSQ,GaoLL,PuYL,DengLJ,ZhangSR(2004).Analysis on status of soil organic matter and N nutrient and their influencing factors in Tibet .Journal of Soil and Water Conservation,18, 54-57. [本文引用: 1]
LuXY,YanY,FanJH,WangXD(2012).Gross nitrification and denitrification in alpine grassland ecosystems on the Tibetan Plateau .Arctic, Antarctic, and Alpine Research,44, 188-196. DOI:10.1657/1938-4246-44.2.188URL [本文引用: 1]
LuoRY,LuoJF,FanJL,LiuDY,HeJS,PerveenN,DingWX(2020).Responses of soil microbial communities and functions associated with organic carbon mineralization to nitrogen addition in a Tibetan grassland .Pedosphere,30, 214-225. DOI:10.1016/S1002-0160(19)60832-5URL [本文引用: 1]
MaSQ,ChenYC,LuXY,WangXD(2018).Soil organic matter chemistry: based on pyrolysis-gas chromatography mass spectrometry (Py-GC/MS) .Mini-reviews in Organic Chemistry,15, 389-403. DOI:10.2174/1570193X15666180108152845URL [本文引用: 2]
MaXX,YanY,LuXY,WangXD(2016).Dynamics of belowground biomass and its relationship with soil moisture in alpine grassland on the North Tibetan Plateau .Ecology and Environment,25, 189-195. [本文引用: 1]
MaY,YangJ,ZhangDG,ZhouH,ZhouHC,ChenJG(2020).Effects of alpine meadow degradation on soil microbial biomass and nitrogen mineralization rate in the Qilian Mountains .Acta Ecologica Sinica,40, 2680-2690. [本文引用: 1]
MehrabanianM(2013).Molecular geochemistry of soil organic matter by pyrolysis gas chromatography/mass spectrometry (GC/MS) technique: a review .Journal of Soil Science and Environmental Management,4, 11-16. DOI:10.5897/JSSEMURL [本文引用: 1]
NiuB,ZengCX,ZhangXZ,HeYT,ShiPL,TianY,FengYF,LiM,WangZP,WangXT,CaoYN(2019).High below- ground productivity allocation of alpine grasslands on the northern Tibet .Plants,8, 535. DOI:10.3390/plants8120535. DOI:10.3390/plants8120535URL [本文引用: 1]
OliveiraDMDS,SchellekensJ,CerriCEP(2016).Molecular characterization of soil organic matter from native vegetation-pasture-sugarcane transitions in Brazil .Science of the Total Environment,548, 450-462. [本文引用: 2]
RaoCS,GroverM,KunduS,DesaiS(2016).Soil enzymes//Lal R. Encyclopedia of Soil Science. 3rd ed.CRC Press, Boca Raton,USA. 2100-2107. [本文引用: 1]
SchimelJP,BecerraCA,BlankinshipJC(2017).Estimating decay dynamics for enzyme activities in soils from different ecosystems .Soil Biology & Biochemistry,114, 5-11. DOI:10.1016/j.soilbio.2017.06.023URL [本文引用: 2]
SenthilkumarK,ManianS,UdaiyanK(1997).The effect of burning on soil enzyme activities in natural grasslands in southern India .Ecological Research,12, 21-25. DOI:10.1007/BF02523606URL [本文引用: 1]
SunHL,ZhengD,YaoTD,ZhangYL(2012).Protection and construction of the national ecological security shelter zone on Tibetan Plateau .Acta Geographica Sinica,67, 3-12. [本文引用: 1]
SunJ,HouG,LiuM,FuG,ZhanTY,ZhouHK,TsunekawaA,HaregeweynN(2019).Effects of climatic and grazing changes on desertification of alpine grasslands, Northern Tibet .Ecological Indicators,107, 105647. DOI:10.1016/j.ecolind.2019.105647. DOI:10.1016/j.ecolind.2019.105647URL [本文引用: 1]
SuoNJ,TanYR,ZhuWX,GuZK,DuGZ(2012).A study on soil enzyme activity in four different grasslands of the eastern Tibetan Plateau .Acta Prataculturae Sinica,21, 10-15. [本文引用: 1]
UtoboEB,TewariL(2015).Soil enzymes as bioindicators of soil ecosystem status .Applied Ecology and Environmental Research,13, 147169. DOI:10.15666/aeer/1301_147169. DOI:10.15666/aeer/1301_147169 [本文引用: 1]
WallensteinMD,McMahonSK,SchimelJP(2009).Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils .Global Change Biology,15, 1631-1639. DOI:10.1111/gcb.2009.15.issue-7URL [本文引用: 1]
WangLD,WangFL,GuoCF,HanFG,WeiLY,LiFM(2016).Review: progress of soil enzymology .Soils,48, 12-21. [本文引用: 1]
WangXX,DongSK,GaoQZ,ZhangY,HuGZ,LuoWR(2018).The rate of soil nitrogen transformation decreased by the degradation of alpine grasslands in the Qinghai Tibet Plateau .Acta Prataculturae Sinica,27, 1-9. [本文引用: 1]
WatanabeK(2009).Detection of protease genes in field soil applied with liquid livestock feces and speculation on their function and origin .Soil Science and Plant Nutrition,55, 42-52. DOI:10.1111/j.1747-0765.2008.00323.xURL [本文引用: 1]
XuHY,LiuGM,WuXD,SmoakJM,MuCC,MaXL,ZhangXL,LiHQ,HuGL(2018).Soil enzyme response to permafrost collapse in the Northern Qinghai-Tibetan Plateau .Ecological Indicators,85, 585-593. DOI:10.1016/j.ecolind.2017.11.013URL [本文引用: 1]
YanZQ,QiYC,PengQ,DongYS,GuoSF,HeYL,WangLQ,LiZL(2017).Effects of increased precipitation and nitrogen deposition on soil enzyme activities .Acta Ecologica Sinica,37, 3019-3027. [本文引用: 1]
YassirI,BuurmanP(2012).Soil organic matter chemistry changes upon secondary succession in Imperata grasslands, Indonesia: a pyrolysis-GC/MS study .Geoderma,173, 94-103. [本文引用: 2]
YuCJ,JiangDQ,TianMY,ChenZH,ZhangYL,WangJ,JiangH,ChenLJ(2020).Soil intracellular and extracellular urease activities and their response mechanisms to carbon additions in chernozem .Chinese Journal of Applied Ecology,31, 1957-1962. [本文引用: 1]
ZhuGL,LiJ,WeiXH,HeNP(2017).Longitudinal patterns of productivity and plant diversity in Tibetan alpine grasslands .Journal of Natural Resources,32, 210-222. [本文引用: 1]
Polyphenol oxidase, tannase and proteolytic activity in relation to tannin concentration in the soil organic horizon under silver birch and Norway spruce 1 2009
... 土壤酶是土壤中具有生物活性的蛋白质, 主要来自土壤中动植物和微生物活动以及有机残体的分解物, 在生态系统物质循环和能量流动中扮演重要角色(Rao et al., 2016).土壤酶和微生物共同推动着土壤的生物化学过程, 对有机残体分解和无机化合物的转化以及土壤肥力的演化具有重要作用(刘琳等, 2011; Kuypers et al., 2018).土壤蛋白酶和脲酶是土壤中参与有机态氮转化的两种关键酶, 蛋白酶作用于有机氮的矿化, 将土壤中植物残体中的蛋白质和肽类物质分解为氨基酸(Watanabe, 2009); 脲酶则是将土壤中有机氮转化为植物和微生物可以利用的无机氮, 也是唯一可以水解土壤中尿素的水解酶, 其分解尿素的速度是尿素自然分解速度的1 014倍(余燕玲等, 2008; Fraser et al., 2013).因此, 土壤蛋白酶和脲酶的活性直接影响着土壤中氮转化的强度和土壤的供氮能力(Du et al., 2020).王学霞等(2018)研究青藏高原退化高寒草地土壤氮矿化影响因素发现, 土壤硝化速率和氨化速率与土壤蛋白酶和脲酶活性密切相关; 而在一个39年长期施肥的热带稻田中, 研究不同季节施肥对土壤温室气体释放的影响时发现, 冬季脲酶活性与土壤N2O的排放密切相关(Bhattacharyya et al., 2013); 但在挪威的欧洲云杉(Picea abies)林土壤有机层的研究发现, 土壤蛋白酶并未有效地降解较为顽固的单宁蛋白复合物中的蛋白质(Adamczyk et al., 2009).因此, 土壤蛋白酶和脲酶对氮的作用方式在不同生态系统中存在一定差异, 其活性可能与基质中的养分浓度和有效性密切相关. ...
Microbial activity and organic matter composition in Mediterranean humus forms 1 2013
Biochemical activity and chemical-structural properties of soil organic matter after 17 years of amendments with olive-mill pomace co-compost 3 2015
... 土壤酶在调节催化反应的过程中受土壤结构稳定性、有机质降解和矿化、养分循环、能量传输及其他环境条件的影响, 另外, 土壤类型(Baligar et al., 1999)、植被类型(Schimel et al., 2017)、取样季节(Wallenstein et al., 2009)和火烧(Senthilkumar et al., 1997)等环境因子也都可能影响土壤酶活性(Avazpoor et al., 2019).通常在有机养分相对丰富的土壤中, 土壤酶活性较高, 对土壤中营养元素的转化作用强度较大, 更有利于系统内物质的循环(Dinesh et al., 1998).土壤有机质是影响土壤酶活性的关键因子(Avazpoor et al., 2019; 于春甲等, 2020), 一方面, 土壤有机质和有机残体在土壤微生物和酶的作用下分解成不同的产物, 同时为微生物和酶提供能源和物质基础, 从而直接或间接地影响着土壤微生物的生长发育和土壤酶活性的消长(Aranda et al., 2015); 另一方面, 土壤有机质可以通过改善土壤物理结构(孔隙度、团粒结构和土壤保水能力等)影响土壤微生物繁殖和酶的分泌(Cui & Holden, 2015).然而, 土壤有机质是由土壤中植物、动物和微生物等有机残体混合而成的复杂物质, 其组成一直是土壤学界研究的难点之一(Derenne & Quénéa, 2015).近年来, 热裂解气质联用技术(Pyrolysis Gas Chromatography/Mass Spectrum, Py-GC/MS)被广泛用于研究草地、森林、农田和苔原等不同生态系统土壤有机质化学成分和化学组成, 对土壤有机质成分从分子水平上有了新的认识(Ma et al., 2018).Py-GC/MS通过测定土壤有机质中苯、D-丙氨酸、呋喃等化学成分的相对丰度研究土壤有机质化学组成和判断其与前体物质的关系, 从而为研究气候变化、土地利用方式和土壤管理方式下有机质的变化特征, 分析土壤过程和功能等提供了新的工具(Mehrabanian, 2013). ...
... 利用Py-GC/MS技术测定土壤有机质化学成分图谱, 用质谱数据库(National Institute of Standards and Technology, NIST 2014)比对获得化学成分.根据已有的研究结果(Grandy et al., 2007; Yassir & Buurman, 2012; Oliveira et al., 2016; Ma et al., 2018)和这些成分的化学结构相似性, 将这些成分归类为烷烃、烯烃、芳香烃、脂肪酸、木质素、含氮化合物、酚类、多环芳烃、多糖、萜烯类和其他成分(表1), 此外, 将未被识别出的12个峰归类为未知物质.土壤有机质化学成分相对丰度用各成分的相对面积表示.另外, 糠醛和吡咯的相对丰度之比、吡咯和酚的相对丰度之比分别被认为是有机质易矿化组分和难矿化组分的矿化指数; 苯和甲苯相对丰度之比与土壤腐殖化强度相关, 是由于苯来源于缩聚芳香族物质, 甲苯则主要来源于非缩聚芳香族链状物质(Aranda et al., 2015). ...
... 通常情况下, 土壤酶活性受植被类型、土地利用方式、施肥、放牧、种植年限和季节变化等因素的影响, 与土壤水分、养分、容重和微生物数量等密切相关(Aranda et al., 2015; Schimel et al., 2017).土壤有机质是影响土壤酶活性的关键因子, 这是因为在微生物的作用下, 土壤有机物质和有机残体分解成不同的中间产物和最终产物, 为微生物和植物提供了营养物质和能量, 同时有机物质通过诱导微生物生成胞外酶进而调节土壤酶活性(Avazpoor et al., 2019). ...
Soil enzyme activity variations in riparian forests in relation to plant species and soil depth 3 2019
... 土壤酶在调节催化反应的过程中受土壤结构稳定性、有机质降解和矿化、养分循环、能量传输及其他环境条件的影响, 另外, 土壤类型(Baligar et al., 1999)、植被类型(Schimel et al., 2017)、取样季节(Wallenstein et al., 2009)和火烧(Senthilkumar et al., 1997)等环境因子也都可能影响土壤酶活性(Avazpoor et al., 2019).通常在有机养分相对丰富的土壤中, 土壤酶活性较高, 对土壤中营养元素的转化作用强度较大, 更有利于系统内物质的循环(Dinesh et al., 1998).土壤有机质是影响土壤酶活性的关键因子(Avazpoor et al., 2019; 于春甲等, 2020), 一方面, 土壤有机质和有机残体在土壤微生物和酶的作用下分解成不同的产物, 同时为微生物和酶提供能源和物质基础, 从而直接或间接地影响着土壤微生物的生长发育和土壤酶活性的消长(Aranda et al., 2015); 另一方面, 土壤有机质可以通过改善土壤物理结构(孔隙度、团粒结构和土壤保水能力等)影响土壤微生物繁殖和酶的分泌(Cui & Holden, 2015).然而, 土壤有机质是由土壤中植物、动物和微生物等有机残体混合而成的复杂物质, 其组成一直是土壤学界研究的难点之一(Derenne & Quénéa, 2015).近年来, 热裂解气质联用技术(Pyrolysis Gas Chromatography/Mass Spectrum, Py-GC/MS)被广泛用于研究草地、森林、农田和苔原等不同生态系统土壤有机质化学成分和化学组成, 对土壤有机质成分从分子水平上有了新的认识(Ma et al., 2018).Py-GC/MS通过测定土壤有机质中苯、D-丙氨酸、呋喃等化学成分的相对丰度研究土壤有机质化学组成和判断其与前体物质的关系, 从而为研究气候变化、土地利用方式和土壤管理方式下有机质的变化特征, 分析土壤过程和功能等提供了新的工具(Mehrabanian, 2013). ...
... ).土壤有机质是影响土壤酶活性的关键因子(Avazpoor et al., 2019; 于春甲等, 2020), 一方面, 土壤有机质和有机残体在土壤微生物和酶的作用下分解成不同的产物, 同时为微生物和酶提供能源和物质基础, 从而直接或间接地影响着土壤微生物的生长发育和土壤酶活性的消长(Aranda et al., 2015); 另一方面, 土壤有机质可以通过改善土壤物理结构(孔隙度、团粒结构和土壤保水能力等)影响土壤微生物繁殖和酶的分泌(Cui & Holden, 2015).然而, 土壤有机质是由土壤中植物、动物和微生物等有机残体混合而成的复杂物质, 其组成一直是土壤学界研究的难点之一(Derenne & Quénéa, 2015).近年来, 热裂解气质联用技术(Pyrolysis Gas Chromatography/Mass Spectrum, Py-GC/MS)被广泛用于研究草地、森林、农田和苔原等不同生态系统土壤有机质化学成分和化学组成, 对土壤有机质成分从分子水平上有了新的认识(Ma et al., 2018).Py-GC/MS通过测定土壤有机质中苯、D-丙氨酸、呋喃等化学成分的相对丰度研究土壤有机质化学组成和判断其与前体物质的关系, 从而为研究气候变化、土地利用方式和土壤管理方式下有机质的变化特征, 分析土壤过程和功能等提供了新的工具(Mehrabanian, 2013). ...
... 通常情况下, 土壤酶活性受植被类型、土地利用方式、施肥、放牧、种植年限和季节变化等因素的影响, 与土壤水分、养分、容重和微生物数量等密切相关(Aranda et al., 2015; Schimel et al., 2017).土壤有机质是影响土壤酶活性的关键因子, 这是因为在微生物的作用下, 土壤有机物质和有机残体分解成不同的中间产物和最终产物, 为微生物和植物提供了营养物质和能量, 同时有机物质通过诱导微生物生成胞外酶进而调节土壤酶活性(Avazpoor et al., 2019). ...
Greenhouse gas emission in relation to labile soil C, N pools and functional microbial diversity as influenced by 39 years long-term fertilizer management in tropical rice 1 2013
... 土壤酶是土壤中具有生物活性的蛋白质, 主要来自土壤中动植物和微生物活动以及有机残体的分解物, 在生态系统物质循环和能量流动中扮演重要角色(Rao et al., 2016).土壤酶和微生物共同推动着土壤的生物化学过程, 对有机残体分解和无机化合物的转化以及土壤肥力的演化具有重要作用(刘琳等, 2011; Kuypers et al., 2018).土壤蛋白酶和脲酶是土壤中参与有机态氮转化的两种关键酶, 蛋白酶作用于有机氮的矿化, 将土壤中植物残体中的蛋白质和肽类物质分解为氨基酸(Watanabe, 2009); 脲酶则是将土壤中有机氮转化为植物和微生物可以利用的无机氮, 也是唯一可以水解土壤中尿素的水解酶, 其分解尿素的速度是尿素自然分解速度的1 014倍(余燕玲等, 2008; Fraser et al., 2013).因此, 土壤蛋白酶和脲酶的活性直接影响着土壤中氮转化的强度和土壤的供氮能力(Du et al., 2020).王学霞等(2018)研究青藏高原退化高寒草地土壤氮矿化影响因素发现, 土壤硝化速率和氨化速率与土壤蛋白酶和脲酶活性密切相关; 而在一个39年长期施肥的热带稻田中, 研究不同季节施肥对土壤温室气体释放的影响时发现, 冬季脲酶活性与土壤N2O的排放密切相关(Bhattacharyya et al., 2013); 但在挪威的欧洲云杉(Picea abies)林土壤有机层的研究发现, 土壤蛋白酶并未有效地降解较为顽固的单宁蛋白复合物中的蛋白质(Adamczyk et al., 2009).因此, 土壤蛋白酶和脲酶对氮的作用方式在不同生态系统中存在一定差异, 其活性可能与基质中的养分浓度和有效性密切相关. ...
Potential short-term effects of yak and Tibetan sheep dung on greenhouse gas emissions in two alpine grassland soils under laboratory conditions 2 2013
... 藏北高原主要位于冈底斯山和念青唐古拉山之间, 平均海拔4 500 m以上, 是长江、怒江等的发源地, 孕育着纳木错、色林错等湖泊.研究区属亚寒带气候, 干燥寒冷, 大部分地区年平均气温低于0 ℃, 最暖月平均气温不及14 ℃, 最冷月平均气温在-10 ℃以下, 65%-85%的降水量集中在植物生长旺盛的季节(6-8月).高寒草地是该区域面积最大和最重要的生态系统类型, 主要包括高寒草甸、高寒草原、高寒草甸草原、高寒荒漠草原和高寒荒漠5种草地类型.其中高寒草甸、高寒草原和高寒荒漠草原是西藏自治区主要的草地类型.高寒草甸在全区广泛分布, 占全区草地面积的34.14%, 以矮生嵩草(Kobresia humilis)为主, 草层高度为20-30 cm, 植被盖度明显高于其余4种草地类型, 为70%-90%.高寒草原是全区面积最大的草地类型, 占全区草地面积的37.36%, 以紫花针茅(Stipa purpurea)为主, 草层高度10-15 cm, 植被盖度20%-45%.高寒草甸草原是高寒草原和高寒草甸之间的过渡类型, 占全区草地面积的7.33%, 以青藏薹草(Carex moorcroftii)和紫花针茅为主, 草层高度约10 cm, 植被盖度约40%.高寒荒漠草原主要分布在羌塘高原西部和藏西北高原, 占全区草地面积的9.88%, 以青藏薹草、垫状驼绒藜(Ceratoides compacta)和沙生针茅(Stipa glareosa)、固沙草(Orinus thoroldii)为代表, 草层高度5-10 cm, 植被盖度5%-10%.高寒荒漠是在寒冷和极为干旱的气候条件下发育而成的草地类型, 占全区总面积的5.92%, 以垫状驼绒藜、高原芥(Christolea crassifolia)为主, 草层高度为5-15 cm, 植被盖度为5%-10% (图1)(崔恒心和维纳汉, 1997; Lu et al., 2012; Cai et al., 2013). ...
Enzyme activities as a component of soil biodiversity: a review 1 2005
... 土壤酶是土壤中有机体代谢的主要动力, 是反映生态系统土壤肥力和养分转化过程的重要指标(王理德等, 2016; Luo et al., 2020).土壤脲酶和蛋白酶是土壤中氮转化的重要催化剂, 对促进土壤氮循环和提高土壤氮的利用率有重要意义(Caldwell, 2005).脲酶促进土壤中酰胺肽键的水解生成植物根系可吸收利用的氨, 指示土壤的供氮水平(Gong et al., 2015); 蛋白酶则是参与土壤中氨基酸、蛋白质和其他含蛋白质氮的有机化合物的转化, 可用于指示土壤氮的矿化(黄玙璠等, 2020).藏北高寒草地土壤酶活性研究结果显示, 不同草地类型土壤酶活性大小存在一定差异, 其中, 高寒荒漠草原土壤脲酶活性显著高于蛋白酶活性(p < 0.05), 这说明高寒荒漠草原土壤中虽然可供微生物利用的有机氮含量较低, 但对土壤氮的利用率却相对较高. ...
Warming and increased precipitation indirectly affect the composition and turnover of labile-fraction soil organic matter by directly affecting vegetation and microorganisms 1 2020
Chemical diversity and incubation time affect non-additive responses of soil carbon and nitrogen cycling to litter mixtures from an alpine steppe soil 1 2017
Carbon structure and enzyme activities in alpine and forest ecosytems 2 2007
... 利用Py-GC/MS技术测定土壤有机质化学成分图谱, 用质谱数据库(National Institute of Standards and Technology, NIST 2014)比对获得化学成分.根据已有的研究结果(Grandy et al., 2007; Yassir & Buurman, 2012; Oliveira et al., 2016; Ma et al., 2018)和这些成分的化学结构相似性, 将这些成分归类为烷烃、烯烃、芳香烃、脂肪酸、木质素、含氮化合物、酚类、多环芳烃、多糖、萜烯类和其他成分(表1), 此外, 将未被识别出的12个峰归类为未知物质.土壤有机质化学成分相对丰度用各成分的相对面积表示.另外, 糠醛和吡咯的相对丰度之比、吡咯和酚的相对丰度之比分别被认为是有机质易矿化组分和难矿化组分的矿化指数; 苯和甲苯相对丰度之比与土壤腐殖化强度相关, 是由于苯来源于缩聚芳香族物质, 甲苯则主要来源于非缩聚芳香族链状物质(Aranda et al., 2015). ...
Gross nitrification and denitrification in alpine grassland ecosystems on the Tibetan Plateau 1 2012
... 藏北高原主要位于冈底斯山和念青唐古拉山之间, 平均海拔4 500 m以上, 是长江、怒江等的发源地, 孕育着纳木错、色林错等湖泊.研究区属亚寒带气候, 干燥寒冷, 大部分地区年平均气温低于0 ℃, 最暖月平均气温不及14 ℃, 最冷月平均气温在-10 ℃以下, 65%-85%的降水量集中在植物生长旺盛的季节(6-8月).高寒草地是该区域面积最大和最重要的生态系统类型, 主要包括高寒草甸、高寒草原、高寒草甸草原、高寒荒漠草原和高寒荒漠5种草地类型.其中高寒草甸、高寒草原和高寒荒漠草原是西藏自治区主要的草地类型.高寒草甸在全区广泛分布, 占全区草地面积的34.14%, 以矮生嵩草(Kobresia humilis)为主, 草层高度为20-30 cm, 植被盖度明显高于其余4种草地类型, 为70%-90%.高寒草原是全区面积最大的草地类型, 占全区草地面积的37.36%, 以紫花针茅(Stipa purpurea)为主, 草层高度10-15 cm, 植被盖度20%-45%.高寒草甸草原是高寒草原和高寒草甸之间的过渡类型, 占全区草地面积的7.33%, 以青藏薹草(Carex moorcroftii)和紫花针茅为主, 草层高度约10 cm, 植被盖度约40%.高寒荒漠草原主要分布在羌塘高原西部和藏西北高原, 占全区草地面积的9.88%, 以青藏薹草、垫状驼绒藜(Ceratoides compacta)和沙生针茅(Stipa glareosa)、固沙草(Orinus thoroldii)为代表, 草层高度5-10 cm, 植被盖度5%-10%.高寒荒漠是在寒冷和极为干旱的气候条件下发育而成的草地类型, 占全区总面积的5.92%, 以垫状驼绒藜、高原芥(Christolea crassifolia)为主, 草层高度为5-15 cm, 植被盖度为5%-10% (图1)(崔恒心和维纳汉, 1997; Lu et al., 2012; Cai et al., 2013). ...
Responses of soil microbial communities and functions associated with organic carbon mineralization to nitrogen addition in a Tibetan grassland 1 2020
... 土壤酶是土壤中有机体代谢的主要动力, 是反映生态系统土壤肥力和养分转化过程的重要指标(王理德等, 2016; Luo et al., 2020).土壤脲酶和蛋白酶是土壤中氮转化的重要催化剂, 对促进土壤氮循环和提高土壤氮的利用率有重要意义(Caldwell, 2005).脲酶促进土壤中酰胺肽键的水解生成植物根系可吸收利用的氨, 指示土壤的供氮水平(Gong et al., 2015); 蛋白酶则是参与土壤中氨基酸、蛋白质和其他含蛋白质氮的有机化合物的转化, 可用于指示土壤氮的矿化(黄玙璠等, 2020).藏北高寒草地土壤酶活性研究结果显示, 不同草地类型土壤酶活性大小存在一定差异, 其中, 高寒荒漠草原土壤脲酶活性显著高于蛋白酶活性(p < 0.05), 这说明高寒荒漠草原土壤中虽然可供微生物利用的有机氮含量较低, 但对土壤氮的利用率却相对较高. ...
Soil organic matter chemistry: based on pyrolysis-gas chromatography mass spectrometry (Py-GC/MS) 2 2018
... 土壤酶在调节催化反应的过程中受土壤结构稳定性、有机质降解和矿化、养分循环、能量传输及其他环境条件的影响, 另外, 土壤类型(Baligar et al., 1999)、植被类型(Schimel et al., 2017)、取样季节(Wallenstein et al., 2009)和火烧(Senthilkumar et al., 1997)等环境因子也都可能影响土壤酶活性(Avazpoor et al., 2019).通常在有机养分相对丰富的土壤中, 土壤酶活性较高, 对土壤中营养元素的转化作用强度较大, 更有利于系统内物质的循环(Dinesh et al., 1998).土壤有机质是影响土壤酶活性的关键因子(Avazpoor et al., 2019; 于春甲等, 2020), 一方面, 土壤有机质和有机残体在土壤微生物和酶的作用下分解成不同的产物, 同时为微生物和酶提供能源和物质基础, 从而直接或间接地影响着土壤微生物的生长发育和土壤酶活性的消长(Aranda et al., 2015); 另一方面, 土壤有机质可以通过改善土壤物理结构(孔隙度、团粒结构和土壤保水能力等)影响土壤微生物繁殖和酶的分泌(Cui & Holden, 2015).然而, 土壤有机质是由土壤中植物、动物和微生物等有机残体混合而成的复杂物质, 其组成一直是土壤学界研究的难点之一(Derenne & Quénéa, 2015).近年来, 热裂解气质联用技术(Pyrolysis Gas Chromatography/Mass Spectrum, Py-GC/MS)被广泛用于研究草地、森林、农田和苔原等不同生态系统土壤有机质化学成分和化学组成, 对土壤有机质成分从分子水平上有了新的认识(Ma et al., 2018).Py-GC/MS通过测定土壤有机质中苯、D-丙氨酸、呋喃等化学成分的相对丰度研究土壤有机质化学组成和判断其与前体物质的关系, 从而为研究气候变化、土地利用方式和土壤管理方式下有机质的变化特征, 分析土壤过程和功能等提供了新的工具(Mehrabanian, 2013). ...
... 利用Py-GC/MS技术测定土壤有机质化学成分图谱, 用质谱数据库(National Institute of Standards and Technology, NIST 2014)比对获得化学成分.根据已有的研究结果(Grandy et al., 2007; Yassir & Buurman, 2012; Oliveira et al., 2016; Ma et al., 2018)和这些成分的化学结构相似性, 将这些成分归类为烷烃、烯烃、芳香烃、脂肪酸、木质素、含氮化合物、酚类、多环芳烃、多糖、萜烯类和其他成分(表1), 此外, 将未被识别出的12个峰归类为未知物质.土壤有机质化学成分相对丰度用各成分的相对面积表示.另外, 糠醛和吡咯的相对丰度之比、吡咯和酚的相对丰度之比分别被认为是有机质易矿化组分和难矿化组分的矿化指数; 苯和甲苯相对丰度之比与土壤腐殖化强度相关, 是由于苯来源于缩聚芳香族物质, 甲苯则主要来源于非缩聚芳香族链状物质(Aranda et al., 2015). ...
Molecular characterization of soil organic matter from native vegetation-pasture-sugarcane transitions in Brazil 2 2016
... 利用Py-GC/MS技术测定土壤有机质化学成分图谱, 用质谱数据库(National Institute of Standards and Technology, NIST 2014)比对获得化学成分.根据已有的研究结果(Grandy et al., 2007; Yassir & Buurman, 2012; Oliveira et al., 2016; Ma et al., 2018)和这些成分的化学结构相似性, 将这些成分归类为烷烃、烯烃、芳香烃、脂肪酸、木质素、含氮化合物、酚类、多环芳烃、多糖、萜烯类和其他成分(表1), 此外, 将未被识别出的12个峰归类为未知物质.土壤有机质化学成分相对丰度用各成分的相对面积表示.另外, 糠醛和吡咯的相对丰度之比、吡咯和酚的相对丰度之比分别被认为是有机质易矿化组分和难矿化组分的矿化指数; 苯和甲苯相对丰度之比与土壤腐殖化强度相关, 是由于苯来源于缩聚芳香族物质, 甲苯则主要来源于非缩聚芳香族链状物质(Aranda et al., 2015). ...
Soil organic matter chemistry changes upon secondary succession in Imperata grasslands, Indonesia: a pyrolysis-GC/MS study 2 2012
... 利用Py-GC/MS技术测定土壤有机质化学成分图谱, 用质谱数据库(National Institute of Standards and Technology, NIST 2014)比对获得化学成分.根据已有的研究结果(Grandy et al., 2007; Yassir & Buurman, 2012; Oliveira et al., 2016; Ma et al., 2018)和这些成分的化学结构相似性, 将这些成分归类为烷烃、烯烃、芳香烃、脂肪酸、木质素、含氮化合物、酚类、多环芳烃、多糖、萜烯类和其他成分(表1), 此外, 将未被识别出的12个峰归类为未知物质.土壤有机质化学成分相对丰度用各成分的相对面积表示.另外, 糠醛和吡咯的相对丰度之比、吡咯和酚的相对丰度之比分别被认为是有机质易矿化组分和难矿化组分的矿化指数; 苯和甲苯相对丰度之比与土壤腐殖化强度相关, 是由于苯来源于缩聚芳香族物质, 甲苯则主要来源于非缩聚芳香族链状物质(Aranda et al., 2015). ...
 ,2,*
,2,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT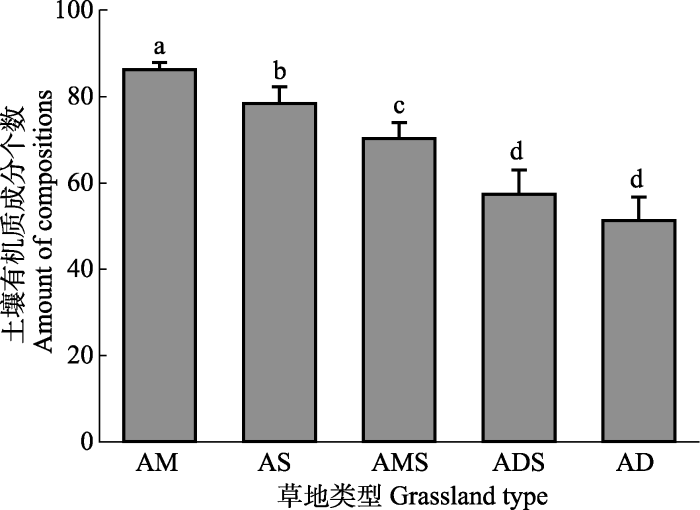 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT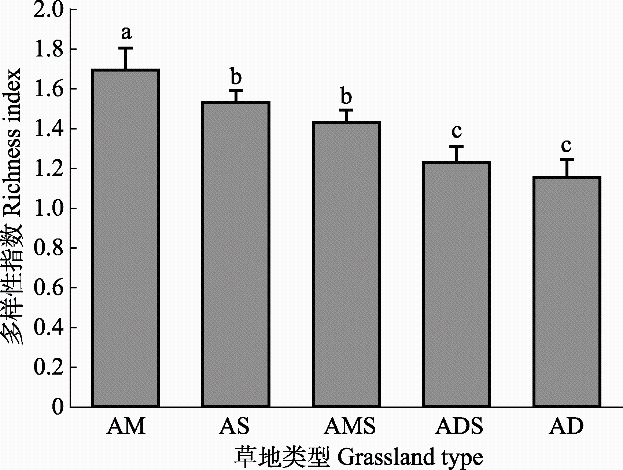 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT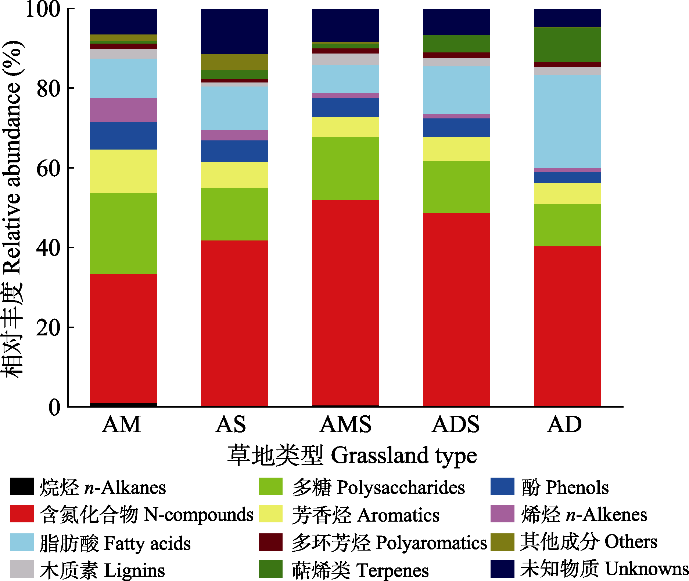 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT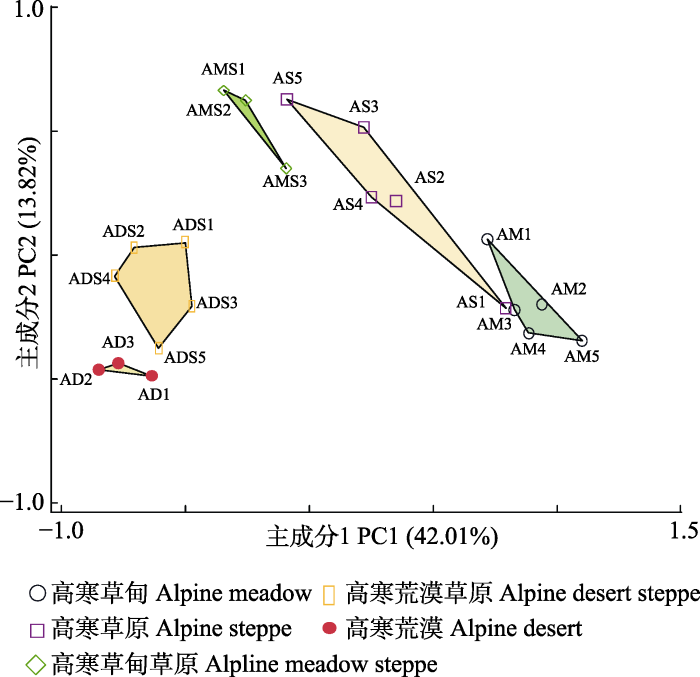 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}