Abstract Aims How soil microbial diversity assembly, maintain and change is a key topic of ecology. A large number of studies show that soil microbial biodiversity is controlled not only by soil environment but also by plant species. However, due to strong covariation between the two factors in the field, it remains a challenge to isolate and clarify the role of plant diversity in regulating soil microbial biodiversity. Hence, here, we aim to clarify how plant diversity affects soil microbial diversity in environment-consistent artificial communities. Methods In this study, we examined differences in species diversity of soil bacteria and fungi among plots subjected single- and mixed-sowing of three grass species with fertilization treatments after 13 years’ experiment on the eastern Qingzang (Tibetan) Plateau. We also analyzed the relationships between soil microbial diversity and edaphic factors as well as plant community attributes. Important findings (1) The species richness and diversity of soil bacteria, not including soil fungi, significantly and consistently decreased in mixed-sowing plots relative to single-sowing plots, with higher relative abundances in proteobacteria and actinobacteria but lower in acidobacteria, bacteroidetes and planctomycetes in the mixed- sowing plots. (2) Soil pH and total nitrogen content significantly decreased while soil total phosphorus content increased in mixed-sowing plots relative to single-sowing plots. Fertilization significantly increased soil available phosphorus while decreased soil pH and soil humidity. However, variations in these edaphic factors contributed little in variation of soil microbial diversity. (3) Fertilization significantly increased plant aboveground biomass while decreasing richness of present plant species, which was also negatively associated with soil bacterial diversity. In short, this long-term field experiment clearly showed that mixed-sowing of common grass species did not promote diversity of soil microbes. This study provides new insight into management of grasses mixed-sowing artificial grasslands. Keywords:plant diversity;soil microbial diversity;mixed-sowing;diversity begets diversity
Table 1 表1 表1广义线性混合模型检验混播和施肥对土壤微生物多样性的影响 Table 1Summary of the effect of mix-sowing and fertilization on variation in soil microbial diversity tested by generalized linear mixed effect model
变量 Variable
变异度 Variability (%) | F
混播 Mixed- Sowing
施肥 Fertilized
单播/混播 Single-sowing/Mixed-sowing (df = 1)
施肥/不施肥 Fertilized/Unfertilized (df = 2)
单播 Single-sowing
混播 Mixed-sowing
土壤微生物多样性 Soil microbial diversity
细菌丰富度(Chao1) Bacterial richness (Chao1)
38.33 | 37.87*
2.72 | 0.69
-96.92*
-30.44
-13.61
细菌Shannon多样性 Bacterial Shannon diversity
44.35 | 41.51*
0.15 | 0.04
-0.60*
-0.02
-0.05
真菌丰富度(Chao1) Fungal richness (Chao1)
16.03 | 3.10
2.18 | 0.50
-28.22
19.30
-12.65
真菌Shannon多样性 Fungal Shannon diversity
8.16 | 3.36
7.14 | 1.33
-0.15
-0.04
-0.50
土壤理化因子 Soil edaphic factor
土壤pH Soil pH
18.17 | 55.48*
27.47 | 7.25*
-0.22*
-0.24*
-0.12*
土壤湿度 Soil humidity
10.95 | 3.49
9.01 | 1.87
0.02
-0.01
-0.02
土壤全氮含量 Soil total nitrogen content
68.81 | 51.36*
2.46 | 1.47
-0.21*
0.06
0.00
土壤全磷含量 Soil total phosphorus content
41.15 | 16.46*
5.23 | 1.68
1.23*
0.18
0.46
土壤有效氮含量 Soil available nitrogen content
0.38 | 0.77
0.80 | 0.12
-0.00
-0.18
0.00
土壤速效磷含量 Soil available phosphorus content
8.25 | 4.43
31.78 | 8.47*
0.04
0.08*
0.05*
土壤有机碳含量 Soil organic carbon content
0.52 | 0.26
9.07 | 1.54
1.20
3.01
-0.42
植物群落 Plant community
植物地上生物量 Plant aboveground biomass
1.45 | 0.44
27.98 | 6.83*
-11.20
14.36
26.93*
植物物种丰富度 Plant species richness
0.84 | 1.01
34.08 | 7.69*
1.33
-1.11
-4.89*
表中前2列数值为固定因子(单播/混播、施肥/不施肥)对各变量变异度的解释量及显著性(解释量(%) | F); 后3列数值为固定因子对各变量的影响大小(回归估计斜率), 其中牧草混播影响以单播为对照, 施肥影响以不施肥为对照; 正、负值表示相对于单播, 混播处理下的增加和降低, 同理相对于不施肥, 施肥处理下的增加和降低; *代表影响显著(p < 0.05)。 Values in first two and last three columns indicate explaining variability (%) | F and effect size (i.e. estimated slopes) of fixed factors (single-sowing/mix-sowing, fertilized/unfertilized treatments) on variation of individual variables, respectively. Positive and negative values of estimated slopes indicate increase or decrease in mixed-sowing relative to single-sowing, as well as fertilized relative to unfertilized treatment, with significant effect (p < 0.05) highlighted by *.
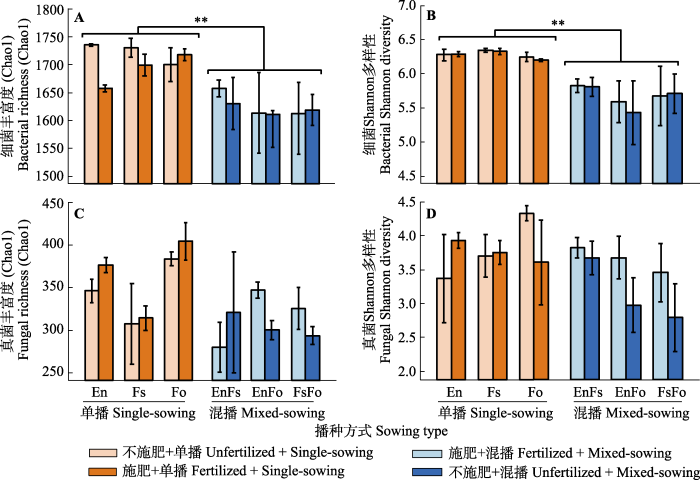
Fig. 2Relative abundance of dominant phyla of soil bacteria and fungi in single- and mixed-sowing plots, with significant difference between them (p < 0.05) indicated by *. A, Dominant phyla of bacteria. B, Dominant phyla of fungi. En, Elymus nutans; Fo, Festuca ovina; Fs, F. sinensis. Others of bacteria include Armatimonadetes, BRC1, Candidatus_Saccharibacteria, Chlamydiae, Chloroflexi, Cyanobacteria, Elusimicrobia, Firmicutes, Gemmatimonadetes, Ignavibacteriae, Latescibacteria, Nitrospirae, Parcubacteria, Unclassified_Bacteria, candidate_division_WPS-1 and candidate_division_WPS-2. Others of fungi include Chytridiomycota, Glomeromycota and three uclassified fungi phyla.
Table 2 表2 表2广义线性混合模型检验混播和施肥对土壤微生物优势菌群(门水平)的影响 Table 2Summary of the effect of mix-sowing and fertilization on variation in dominant phyla of soil microbes tested by generalized linear mixed effect model
微生物类群 Microbes taxon
变异度 Variability (%) | F
混播 Mixed- Sowing
施肥 Fertilized
单播/混播 Single-sowing/Mixed-sowing (df = 1)
施肥/不施肥 Fertilized/Unfertilized (df = 2)
单播 Single- sowing
混播 Mixed- sowing
细菌门水平优势类群 Dominant phylum of bacteria
酸杆菌门 Acidobacteria
32.84 | 11.81*
2.48 | 0.65
-0.05*
0.00
0.02
放线菌门 Actinobacteria
12.42 | 9.13*
13.84 | 2.83
0.01*
0.00
-0.01*
拟杆菌门 Bacteroidetes
34.68 | 13.62*
0.93 | 0.24
-0.03*
0.01
0.00
浮霉菌门 Planctomycetes
21.33 | 5.59
0.73 | 0.16
-0.02
0.00
0.00
变形菌门 Proteobacteria
32.67 | 28.11*
0.47 | 0.11
0.08*
-0.01
-0.01
疣微菌门 Verrucomicrobia
0.23 | 0.04
8.19 | 1.77
0.00
0.01
0.01
真菌门水平优势菌群 Dominant phylum of fungi
子囊菌门 Ascomycota
1.07 | 0.85
1.25 | 0.19
-0.05
0.01
0.05
担子菌门 Basidiomycota
2.03 | 1.82
1.14 | 0.18
0.02
-0.04
-0.01
接合菌门 Zygomycota
1.51 | 0.82
4.47 | 0.72
0.04
0.04
0.06
表中前2列数值为固定因子(单播/混播、施肥/不施肥)对各变量变异度的解释量及其显著性(解释量(%) | F); 后3列数值为固定因子对各变量的影响大小(回归估计斜率), 其中牧草混播影响以单播为对照, 施肥影响以不施肥为对照; 正、负值表示相对于单播, 混播处理下的增加和降低, 同理相对于不施肥, 施肥处理下的增加和降低; *代表影响显著(p < 0.05), 加粗字体表示影响边缘显著(p < 0.1)。 Values in first two and last three columns indicate explaining variability (%) | F and effect size (i.e. estimated slopes) of fixed factors (single-sowing/mix-sowing, fertilized/unfertilized treatments) on variation of individual variables, respectively. Positive and negative values of estimated slopes indicate increase or decrease in mixed-sowing relative to single-sowing, as well as fertilized relative to unfertilized treatment, with significant effect (p < 0.05) highlighted by * and marginally significant (p < 0.1) highlighted by bold font.
Fig. 3Difference in soil microbial community compositions in single- and mixed-sowing plots. A, Soil bacterial community composition. B, Soil fungal community composition. En, Elymus nutans; Fo, Festuca ovina; Fs, F. sinensis. PAB, plant aboveground biomass; PSR, plant species richness; SAN, soil available nitrogen; SAP, soil available phosphorus; SOC, soil organic carbon; STN, soil total nitrogen; STP, soil total phosphorus.
Fig. 4Generalized canonical discriminant analysis for links among soil bacterial and fungal diversities, edaphic factors and plant community. A, Bacteria. B, Fungal. En, Elymus nutans; Fo, Festuca ovina; Fs, F. sinensis. There are significant difference between single- and mixed-sowing plots tested by a Wilks’ lambda multivariate test. PAB, plant aboveground biomass; PSR, plant species richness; SAN, soil available nitrogen; SAP, soil available phosphorus; SOC, soil organic carbon; STN, soil total nitrogen; STP, soil total phosphorus.
Table 3 表3 表3广义线性混合模型检验土壤微生物多样性与各土壤理化因子及植物群落的相关关系(平均值±标准误) Table 3Association of soil microbial diversity with edaphic factors and plant community tested by generalized linear mixed effect model (mean ± SE)
表中R2表示回归模型的拟合程度, 固定因子(土壤理化因子、植物群落、混播以及施肥)对各微生物多样性指标的影响大小以回归估计斜率值及其标准误表示, 正、负值分别表示固定因子与各微生物多样性指标的正相关和负相关, *代表影响显著(p < 0.05)。 R2 indicates the fitting degree of regression model; the effect size of fixed factors (edaphic factors, plant community, mixed-sowing and fertilization) on various microbial diversity index are indicated by estimated slopes and its standard error. Positive and negative values of estimated slopes indicate the positive and negative correlations between fixed factors and various microbial diversity index with significant (p < 0.05) highlighted by *.
BahramM,HildebrandF,ForslundSK,AndersonJL,SoudzilovskaiaNA,BodegomPM,Bengtsson-PalmeJ,AnslanS,CoelhoLP,HarendH,Huerta-CepasJ,MedemaMH,MaltzMR,MundraS,OlssonPA,PentM,PõlmeS,SunagawaS,RybergM,TedersooL,BorkP(2018).Structure and function of the global topsoil microbiome .Nature,560, 233-237. DOI:10.1038/s41586-018-0386-6URL [本文引用: 1]
BardgettRD,van der PuttenWH(2014).Belowground biodiversity and ecosystem functioning .Nature,515, 505-511. DOI:10.1038/nature13855URL [本文引用: 1]
BoyerAG,JetzW(2014).Extinctions and the loss of ecological function in island bird communities .Global Ecology and Biogeography,23, 679-688. DOI:10.1111/geb.2014.23.issue-6URL [本文引用: 1]
CallaghanTV,JonassonS,NicholsH,HeywoodRB,WookeyPA,WadhamsP,DowdeswellJA,SchofieldAN(1995).Arctic terrestrial ecosystems and environmental change .Philosophical Transactions: Physical and Engineering Sciences,352, 259-276. [本文引用: 1]
ChenC,ChenHYH,ChenXL,HuangZQ(2019).Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration .Nature Communications,10, 1332. DOI:10.1038/s41467-019-09258-y. DOI:10.1038/s41467-019-09258-yURL [本文引用: 1]
ChenY,LüGH,LiY(2018).Soil microbial functional diversity of rhizosphere and non-rhizosphere of three dominant herbaceous plants in the Dushanzi District .Acta Ecologica Sinica,38, 3110-3117. [本文引用: 1]
CivitelloDJ,CohenJ,FatimaH,HalsteadNT,LirianoJ,McMahonTA,OrtegaCN,SauerEL,SehgalT,YoungS,RohrJR(2015).Biodiversity inhibits parasites: broad evidence for the dilution effect .Proceedings of the National Academy of Sciences of the United States of America,112, 8667-8671. DOI:10.1073/pnas.1506279112PMID:26069208 [本文引用: 1] Infectious diseases of humans, wildlife, and domesticated species are increasing worldwide, driving the need to understand the mechanisms that shape outbreaks. Simultaneously, human activities are drastically reducing biodiversity. These concurrent patterns have prompted repeated suggestions that biodiversity and disease are linked. For example, the dilution effect hypothesis posits that these patterns are causally related; diverse host communities inhibit the spread of parasites via several mechanisms, such as by regulating populations of susceptible hosts or interfering with parasite transmission. However, the generality of the dilution effect hypothesis remains controversial, especially for zoonotic diseases of humans. Here we provide broad evidence that host diversity inhibits parasite abundance using a meta-analysis of 202 effect sizes on 61 parasite species. The magnitude of these effects was independent of host density, study design, and type and specialization of parasites, indicating that dilution was robust across all ecological contexts examined. However, the magnitude of dilution was more closely related to the frequency, rather than density, of focal host species. Importantly, observational studies overwhelmingly documented dilution effects, and there was also significant evidence for dilution effects of zoonotic parasites of humans. Thus, dilution effects occur commonly in nature, and they may modulate human disease risk. A second analysis identified similar effects of diversity in plant-herbivore systems. Thus, although there can be exceptions, our results indicate that biodiversity generally decreases parasitism and herbivory. Consequently, anthropogenic declines in biodiversity could increase human and wildlife diseases and decrease crop and forest production.
de VriesFT,HofflandE,van EekerenN,BrussaardL,BloemJ(2006).Fungal/bacterial ratios in grasslands with contrasting nitrogen management .Soil Biology & Biochemistry,38, 2092-2103. DOI:10.1016/j.soilbio.2006.01.008URL [本文引用: 1]
Delegado-BaquerizoM,BardgettRD,VitousekPM,MaestreFT,WilliamsMA,EldridgeDJ,LambersH,NeuhauserS,GallardoA,Garcia-VelázquezL,SalaOE,AbadesSR,AlfaroFD,BerheAA,BowkerMA,et al.(2019).Changes in belowground biodiversity during ecosystem development .Proceedings of the National Academy of Sciences of the United States of America,116, 6891-6896. [本文引用: 1]
FangJY,JingHC,ZhangWH,GaoSQ,DuanZY,WangHS,ZhongJ,PanQM,ZhaoK,BaiWM,LiLH,BaiYF,JiangGM,HuangJH,HuangZY(2018).The concept of “grass-based livestock husbandry” and its practice in Hulun Buir, Inner Mongolia .Chinese Science Bulletin,63, 1619-1631. [本文引用: 2]
FieldE,CastagneyrolB,GibbsM,JactelH,BarsoumN,SchönroggeK,HectorA(2020).Associational resistance to both insect and pathogen damage in mixed forests is modulated by tree neighbour identity and drought .Journal of Ecology,108, 1511-1522. DOI:10.1111/jec.v108.4URL [本文引用: 1]
FiererN,JacksonRB(2006).The diversity and biogeography of soil bacterial communities .Proceedings of the National Academy of Sciences of the United States of America,103, 626-631. PMID:16407148 [本文引用: 1] For centuries, biologists have studied patterns of plant and animal diversity at continental scales. Until recently, similar studies were impossible for microorganisms, arguably the most diverse and abundant group of organisms on Earth. Here, we present a continental-scale description of soil bacterial communities and the environmental factors influencing their biodiversity. We collected 98 soil samples from across North and South America and used a ribosomal DNA-fingerprinting method to compare bacterial community composition and diversity quantitatively across sites. Bacterial diversity was unrelated to site temperature, latitude, and other variables that typically predict plant and animal diversity, and community composition was largely independent of geographic distance. The diversity and richness of soil bacterial communities differed by ecosystem type, and these differences could largely be explained by soil pH (r(2) = 0.70 and r(2) = 0.58, respectively; P < 0.0001 in both cases). Bacterial diversity was highest in neutral soils and lower in acidic soils, with soils from the Peruvian Amazon the most acidic and least diverse in our study. Our results suggest that microbial biogeography is controlled primarily by edaphic variables and differs fundamentally from the biogeography of "macro" organisms.
FriendlyM,FoxJ(2017).Candisc: Visualizing Generalized Canonical Discriminant and Canonical correlation analysis .R package version 0.8-0. [2017-09-19]. https://CRAN.R-project.org/package=candisc. URL [本文引用: 1]
HaoYQ,ZhaoXF,ZhangDY(2016).Field experimental evidence that stochastic processes predominate in the initial assembly of bacterial communities .Environmental Microbiology,18, 1730-1739. DOI:10.1111/1462-2920.12858URL [本文引用: 1]
HectorA,SchmidB,BeierkuhnleinC,CaldeiraMC,DiemerM,DimitrakopoulosPG,FinnJA,FreitasH,GillerPS,GoodJ,HarrisR,HogbergP,Huss-DanellK,JoshiJ,JumpponenA,et al.(1999).Plant diversity and productivity experiments in European grasslands .Science,286, 1123-1127. PMID:10550043 [本文引用: 1] At eight European field sites, the impact of loss of plant diversity on primary productivity was simulated by synthesizing grassland communities with different numbers of plant species. Results differed in detail at each location, but there was an overall log-linear reduction of average aboveground biomass with loss of species. For a given number of species, communities with fewer functional groups were less productive. These diversity effects occurred along with differences associated with species composition and geographic location. Niche complementarity and positive species interactions appear to play a role in generating diversity-productivity relationships within sites in addition to sampling from the species pool.
JohnsonD,CampbellCD,LeeJA,CallaghanTV,Gwynn-JonesD(2002).Arctic microorganisms respond more to elevated UV-B radiation than CO2 .Nature,416, 82-83. DOI:10.1038/416082aURL [本文引用: 1]
KongWD(2013).A review of microbial diversity in polar terrestrial environments .Biodiversity Science,21, 457-468. [本文引用: 1]
LeffJW,BardgettRD,WilkinsonA,JacksonBG,PritchardWJ,de LongJR,OakleyS,MasonKE,OstleNJ,JohnsonD,BaggsEM,FiererN(2018).Predicting the structure of soil communities from plant community taxonomy, phylogeny, and traits .The ISME Journal,12, 1794-1805. DOI:10.1038/s41396-018-0089-xURL [本文引用: 2]
LiA,NiuKC,DuGZ(2011).Resource availability, species composition and sown density effects on productivity of experimental plant communities .Plant and Soil,344, 177-186. DOI:10.1007/s11104-011-0738-6URL [本文引用: 5]
LiL(2016).Intercropping enhances agroecosystem services and functioning: current knowledge and perspectives .Chinese Journal of Eco-Agriculture,24, 403-415. [本文引用: 1]
LiuL,ZhuK,WurzburgerN,ZhangJ(2020).Relationships between plant diversity and soil microbial diversity vary across taxonomic groups and spatial scales .Ecosphere,11, e02999. DOI:10.1016/j.soilbio.2021.108143. DOI:10.1016/j.soilbio.2021.108143 [本文引用: 1]
MulderC,van WijnenHJ,van WezelAP(2005).Numerical abundance and biodiversity of below-ground taxocenes along a pH gradient across the Netherlands .Journal of Biogeography,32, 1775-1790. DOI:10.1111/jbi.2005.32.issue-10URL [本文引用: 1]
NelsonDW,SommersLE(1983).Total carbon, organic carbon, and organic matter //Page A, Miller R, Keeney D. Methods of Soil Analysis. American Society of Agronomy,Madison. 539-579. [本文引用: 1]
NiuKC,HeJS,LechowiczMJ(2016a).Grazing-induced shifts in community functional composition and soil nutrient availability in Tibetan alpine meadows .Journal of Applied Ecology,53, 1554-1564. DOI:10.1111/1365-2664.12727URL [本文引用: 2]
NiuKC,HeJS,ZhangST,LechowiczMJ(2016b).Tradeoffs between forage quality and soil fertility: lessons from Himalayan rangelands .Agriculture, Ecosystems & Environment,234, 31-39. DOI:10.1016/j.agee.2016.04.023URL [本文引用: 1]
NiuKC,LiuYN,ShenZH,HeFL,FangJY(2009).Community assembly: the relative importance of neutral theory and niche theory .Biodiversity Science,17, 579-593. DOI:10.3724/SP.J.1003.2009.09142URL [本文引用: 1]
PardoLH,FennME,GoodaleCL,GeiserLH,DriscollCT,AllenEB,BaronJS,BobbinkR,BowmanWD,ClarkCM,EmmettB,GilliamFS,GreaverTL,HallSJ,LilleskovEA,et al.(2011).Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States .Ecological Applications,21, 3049-3082. DOI:10.1890/10-2341.1URL [本文引用: 1]
PinheiroJ,BatesD,DebRoyS,SarkarD,R CoreTeam(2017).nlme: Linear and nonlinear mixed effects models R package version3 .1-131. [2021-02-03]. https://CRAN.R-project.org/package=nlme. URL [本文引用: 1]
ProberSM,LeffJW,BatesST,BorerET,FirnJ,HarpoleWS,LindEM,SeabloomEW,AdlerPB,BakkerJD,ClelandEE,DeCrappeoNM,DeLorenzeE,HagenahN,HautierY,et al.(2015).Plant diversity predicts beta but not alpha diversity of soil microbes across grasslands worldwide .Ecology Letters,18, 85-95. DOI:10.1111/ele.2014.18.issue-1URL [本文引用: 2]
RakowskiC,CardinaleBJ(2016).Herbivores control effects of algal species richness on community biomass and stability in a laboratory microcosm experiment .Oikos,125, 1627-1635. DOI:10.1111/oik.2016.v125.i11URL [本文引用: 1]
RamusAP,LongZT(2016).Producer diversity enhances consumer stability in a benthic marine community .Journal of Ecology,104, 572-579. DOI:10.1111/1365-2745.12509URL [本文引用: 1]
SoliveresS,van der PlasF,ManningP,PratiD,GossnerMM,RennerSC,AltF,ArndtH,BaumgartnerV,BinkensteinJ,BirkhoferK,BlaserS,BlüthgenN,BochS,BöhmS,BörschigC,BuscotF,DiekötterT,HeinzeJ,HölzelN,JungK,KlausVH,KleinebeckerT,KlemmerS,et al.(2016).Biodiversity at multiple trophic levels is needed for ecosystem multifunctionality .Nature,536, 456-459. DOI:10.1038/nature19092URL [本文引用: 1]
SommersLE,NelsonDW(1972).Determination of total phosphorus in soils: a rapid perchloric acid digestion procedure .Soil Science Society of America Journal,36, 902-904. DOI:10.2136/sssaj1972.03615995003600060020xURL [本文引用: 2]
WaggC,BenderSF,WidmerF,van der HeijdenMGA(2014).Soil biodiversity and soil community composition determine ecosystem multifunctionality .Proceedings of the National Academy of Sciences of the United States of America,111, 5266-5270. [本文引用: 1]
WangX,ZengZH,HuYG,ZhuB(2007).Progress and prospect on mixture of Gramineae herbage and Leguminosae herbage .Chinese Journal of Grassland,29, 92-98. [本文引用: 1]
WangXB,LüXT,YaoJ,WangZW,DengY,ChengWX,ZhouJZ,HanXG(2017).Habitat-specific patterns and drivers of bacterial β-diversity in China’s drylands .The ISME Journal,11, 1345-1358. DOI:10.1038/ismej.2017.11URL [本文引用: 1]
WangYT,NiuKC(2020).Effect of soil environment on functional diversity of soil nematodes in Tibetan alpine meadows .Biodiversity Science,28, 707-717. DOI:10.17520/biods.2020042URL [本文引用: 3]
WardleDA(2016).Do experiments exploring plant diversity- ecosystem functioning relationships inform how biodiversity loss impacts natural ecosystems? Journal of Vegetation Science,27, 646-653. DOI:10.1111/jvs.12399URL [本文引用: 1]
WardleDA,BardgettRD(2010).Aboveground-Belowground Linkages. Biotic Interactions, Ecosystem Processes, and Global Change.Biotic Interactions, Ecosystem Processes, and Global Change. Oxford University Press, Oxford,UK. [本文引用: 1]
WardleDA,BardgettRD,KlironomosJN,SetäläH,van der PuttenWH,WallDH(2004).Ecological linkages between aboveground and belowground biota .Science,304, 1629-1633. PMID:15192218 [本文引用: 1] All terrestrial ecosystems consist of aboveground and belowground components that interact to influence community- and ecosystem-level processes and properties. Here we show how these components are closely interlinked at the community level, reinforced by a greater degree of specificity between plants and soil organisms than has been previously supposed. As such, aboveground and belowground communities can be powerful mutual drivers, with both positive and negative feedbacks. A combined aboveground-belowground approach to community and ecosystem ecology is enhancing our understanding of the regulation and functional significance of biodiversity and of the environmental impacts of human-induced global change phenomena.
WardleDA,JonssonM,BansalS,BardgettRD,GundaleMJ,MetcalfeDB(2012).Linking vegetation change, carbon sequestration and biodiversity: insights from island ecosystems in a long-term natural experiment .Journal of Ecology,100, 16-30. DOI:10.1111/jec.2011.100.issue-1URL [本文引用: 1]
WeisserWW,RoscherC,MeyerST,EbelingA,LuoGJ,AllanE,BeβlerH,BarnardRL,BuchmannN,BuscotF,EngelsC,FischerC,FischerM,GesslerA,GleixnerG,et al.(2017).Biodiversity effects on ecosystem functioning in a 15-year grassland experiment: patterns, mechanisms, and open questions .Basic and Applied Ecology,23, 1-73. DOI:10.1016/j.baae.2017.06.002URL [本文引用: 3]
YangF,NiuKC,CollinsCG,YanXB,JiYG,LingN,ZhouXH,DuGZ,GuoH,HuSJ(2019a).Grazing practices affect the soil microbial community composition in a Tibetan alpine meadow .Land Degradation & Development,30, 49-59. DOI:10.1002/ldr.v30.1URL [本文引用: 1]
YangW,JeelaniN,ZhuZH,LuoYQ,ChengXL,AnSQ(2019c).Alterations in soil bacterial community in relation to Spartina alterniflora Loisel. invasion chronosequence in the eastern Chinese coastal wetlands .Applied Soil Ecology,135, 38-43. DOI:10.1016/j.apsoil.2018.11.009 [本文引用: 1] In order to better understand the variations in soil bacterial community and associated drivers following plant invasion, we investigated changes in soil bacterial community along with 9-, 13-, 20- and 23-year-old Spartina alterniflora Loisel. (SA) invasion in comparison with bare flat (BF) in the eastern Chinese coastal wetlands, based on analyses of quantitative polymerase chain reaction (qPCR) and Illumina MiSeq DNA sequencing of 16S rRNA gene. The SA invasion significantly elevated soil bacterial abundance and diversity relative to BF, with the highest levels in 9-year-old SA soil, which gradually decreased with SA invasion from 9 to 23 years. The abundance of copiotrophic Proteobacteria, beta-proteobacteria, and Bacteroidetes generally diminished along with SA invasion chronosequence. While, changes in abundance of oligotrophic Chloroflexi, Acidobacteria, Nitrospirae and Planctomycetes exhibited opposite trends. Our data suggest that soil nutrient substrates, and physiochemical properties (soil pH and/or moisture) primarily drive the shifts in soil bacterial abundance, diversity, and community composition along with SA invasion chronosequence in the costal wetlands of eastern China. Overall, soil bacterial abundance and diversity peaked in 9-year-old SA community, with soil bacterial community composition changing from copiotrophic to oligotrophic groups along with SA invasion chronosequence.
ZhangST,LiuJP,BaoXH,NiuKC(2011).Seed-to-seed potential allelopathic effects between Ligularia virgaurea and native grass species of Tibetan alpine grasslands .Ecological Research,26, 47-52. DOI:10.1007/s11284-010-0751-xURL [本文引用: 1]
ZhangXM,HanXG(2012).Nitrogen deposition alters soil chemical properties and bacterial communities in the Inner Mongolia grassland .Journal of Environmental Sciences,24, 1483-1491. DOI:10.1016/S1001-0742(11)60900-5URL [本文引用: 1]
ZhaoXG,ZhangST,NiuKC(2020a).Association of soil bacterial diversity with plant community functional attributes in alpine meadows . Scientia Sinica (Vitae),50, 70-80. [本文引用: 2]
ZhaoXG,ZhangST,NiuKC(2020b).Relationships between soil fungal diversity, plant community functional traits, and soil attributes in Tibetan alpine meadows .Chinese Journal of Applied and Environmental Biology,26, 1-9. [本文引用: 2]
ZhouZH,WangCK(2016).Changes of the relationships between soil and microbes in carbon, nitrogen and phosphorus stoichiometry during ecosystem succession .Chinese Journal of Plant Ecology,40, 1257-1266. DOI:10.17521/cjpe.2016.0218URL [本文引用: 1]
ZhouZH,WangCK,ZhengMH,JiangLF,LuoYQ(2017).Patterns and mechanisms of responses by soil microbial communities to nitrogen addition .Soil Biology & Biochemistry,115, 433-441. DOI:10.1016/j.soilbio.2017.09.015URL [本文引用: 1]
Structure and function of the global topsoil microbiome 1 2018
... 其次, 虽然混播和施肥改变了土壤理化因子或植物生产力, 但这些变化不足以解释土壤微生物多样性在处理间的差异.例如, 混播处理显著降低了土壤pH和土壤全氮含量、增加了土壤全磷含量, 但无论是土壤pH还是全氮、全磷都与土壤微生物多样性无显著相关性; 施肥显著增加了植物地上生物量, 但植物地上生物量改变与细菌和真菌的多样性变化没有显著相关.这并不能说明土壤pH和土壤养分对土壤微生物多样性没有影响, 更可能是由于其影响错综复杂, 如pH减小不仅会直接影响土壤细菌多样性(Fierer & Jackson, 2006), 也会通过影响食细菌土壤线虫多度等多种间接途径影响土壤细菌多样性(Mulder et al., 2005; 王宇彤和牛克昌, 2020), 使pH对微生物的直接影响和间接影响相互抵消, 二者关系总体不显著.同理, 虽然很多研究报道了土壤微生物多样性和土壤肥力相关(周正虎和王传宽, 2016; Delgado-Baquerizo et al., 2017; Bahram et al., 2018; Yang et al., 2019a, 2019c), 但我们并不清楚土壤全氮和全磷含量与土壤微生物之间的确切关系(Delgado-Baquerizo et al., 2017); 土壤氮磷含量, 尤其是速效氮磷含量, 与土壤微生物多样性的相互影响更有可能通过植物介导(Hacker et al., 2015; Chen et al., 2019).如, 在青藏高原高寒草地, 在土壤氮较充足的情况下, 土壤速效磷含量的增加显著促进植物生产力和功能多样性(Niu et al., 2016a, 2016b), 为土壤生物提供丰度的资源和微环境, 使高寒草地群落中土壤速效磷含量与土壤微生物(赵兴鸽等, 2020a, 2020b)及土壤线虫(王宇彤和牛克昌, 2020)多样性都有明显正相关关系.本研究中, 我们也发现施肥增加土壤速效磷含量, 速效磷含量与土壤细菌的丰富度有显著正相关关系.作为土壤微生物最重要的碳源, 植物生产力和土壤有机质含量往往和土壤细菌多样性正相关而与土壤真菌多样性负相关(Wardle et al., 2004; de Vries et al., 2006; Delgado- Baquerizo et al., 2017); 我们的研究中也发现土壤有机碳含量与土壤真菌丰富度负相关, 但其与土壤细菌多样性并无显著相关性.此外, 混播和施肥对土壤有机碳含量影响不显著, 我们也难以澄清土壤有机碳含量如何影响土壤微生物多样性. ...
Belowground biodiversity and ecosystem functioning 1 2014
... 随着人类干扰增加及全球气候剧变, 生物多样性改变、生态系统退化等生态环境问题成为当今科学研究的核心(Hector et al., 1999; Pardo et al., 2011; Boyer & Jetz, 2014).大量研究详实阐述了生物多样性变化对生态系统功能及服务的影响(Soliveres et al., 2016; 陈悦等, 2018), 越来越多的研究关注各类生物多样性之间的相互影响及其在生态系统能量流动和物质循环中的作用(Ramus & Long, 2016).作为生态系统中主要的生产者和分解者, 植物和土壤微生物多样性协同影响生态系统功能和服务(Wall & Moore, 1999; Bardgett & van der Putten, 2014).但我们对自然群落中, 植物多样性与微生物多样性之间的相互影响机制知之甚少(Rakowski & Cardinale, 2016), 甚至难以确定植物多样性本身是促进还是抑制土壤微生物多样性. ...
Extinctions and the loss of ecological function in island bird communities 1 2014
... 随着人类干扰增加及全球气候剧变, 生物多样性改变、生态系统退化等生态环境问题成为当今科学研究的核心(Hector et al., 1999; Pardo et al., 2011; Boyer & Jetz, 2014).大量研究详实阐述了生物多样性变化对生态系统功能及服务的影响(Soliveres et al., 2016; 陈悦等, 2018), 越来越多的研究关注各类生物多样性之间的相互影响及其在生态系统能量流动和物质循环中的作用(Ramus & Long, 2016).作为生态系统中主要的生产者和分解者, 植物和土壤微生物多样性协同影响生态系统功能和服务(Wall & Moore, 1999; Bardgett & van der Putten, 2014).但我们对自然群落中, 植物多样性与微生物多样性之间的相互影响机制知之甚少(Rakowski & Cardinale, 2016), 甚至难以确定植物多样性本身是促进还是抑制土壤微生物多样性. ...
Arctic terrestrial ecosystems and environmental change 1 1995
... 总而言之, 在全球变化加剧背景下, 研究植物多样性与土壤微生物多样性的相互作用对澄清生态系统功能动态极其重要.澄清土壤-植物-微生物的关系不仅需要结合野外调查研究和长期控制实验, 也需要整合考虑环境变化下植物-微生物-土壤线虫等多营养级作用.基于长期的牧草混播和施肥控制实验, 我们发现在青藏高原高寒草地牧草混播会抑制土壤微生物多样性, 尤其是土壤细菌丰富度, 这一结果与我们在该地区的野外调查和控制实验结果基本一致.目前我们并不能厘清其中缘由, 国内外其他地区也少有如此长期的控制实验研究; 青藏高原草地土壤微生物分布和多样性的特殊性(Callaghan et al., 1995; Johnson et al., 2002; 孔维栋, 2013), 也使得我们难以从其他地区的调查研究中获取很多的启示.青藏高原急需发展草牧业的现实, 迫切需要我们积极开展相关的研究, 厘清符合当地现状的优势牧草单播和混播各自的利弊(方精云等, 2018).为此, 我们认为本研究结果不仅对厘清植物多样性和土壤微生物多样性相互作用提出了新挑战, 而且为青藏高原高寒草地草牧业的发展提供了参考资料. ...
Meta-analysis shows positive effects of plant diversity on microbial biomass and respiration 1 2019
... 其次, 虽然混播和施肥改变了土壤理化因子或植物生产力, 但这些变化不足以解释土壤微生物多样性在处理间的差异.例如, 混播处理显著降低了土壤pH和土壤全氮含量、增加了土壤全磷含量, 但无论是土壤pH还是全氮、全磷都与土壤微生物多样性无显著相关性; 施肥显著增加了植物地上生物量, 但植物地上生物量改变与细菌和真菌的多样性变化没有显著相关.这并不能说明土壤pH和土壤养分对土壤微生物多样性没有影响, 更可能是由于其影响错综复杂, 如pH减小不仅会直接影响土壤细菌多样性(Fierer & Jackson, 2006), 也会通过影响食细菌土壤线虫多度等多种间接途径影响土壤细菌多样性(Mulder et al., 2005; 王宇彤和牛克昌, 2020), 使pH对微生物的直接影响和间接影响相互抵消, 二者关系总体不显著.同理, 虽然很多研究报道了土壤微生物多样性和土壤肥力相关(周正虎和王传宽, 2016; Delgado-Baquerizo et al., 2017; Bahram et al., 2018; Yang et al., 2019a, 2019c), 但我们并不清楚土壤全氮和全磷含量与土壤微生物之间的确切关系(Delgado-Baquerizo et al., 2017); 土壤氮磷含量, 尤其是速效氮磷含量, 与土壤微生物多样性的相互影响更有可能通过植物介导(Hacker et al., 2015; Chen et al., 2019).如, 在青藏高原高寒草地, 在土壤氮较充足的情况下, 土壤速效磷含量的增加显著促进植物生产力和功能多样性(Niu et al., 2016a, 2016b), 为土壤生物提供丰度的资源和微环境, 使高寒草地群落中土壤速效磷含量与土壤微生物(赵兴鸽等, 2020a, 2020b)及土壤线虫(王宇彤和牛克昌, 2020)多样性都有明显正相关关系.本研究中, 我们也发现施肥增加土壤速效磷含量, 速效磷含量与土壤细菌的丰富度有显著正相关关系.作为土壤微生物最重要的碳源, 植物生产力和土壤有机质含量往往和土壤细菌多样性正相关而与土壤真菌多样性负相关(Wardle et al., 2004; de Vries et al., 2006; Delgado- Baquerizo et al., 2017); 我们的研究中也发现土壤有机碳含量与土壤真菌丰富度负相关, 但其与土壤细菌多样性并无显著相关性.此外, 混播和施肥对土壤有机碳含量影响不显著, 我们也难以澄清土壤有机碳含量如何影响土壤微生物多样性. ...
独山子区优势草本植物根际与非根际土壤微生物功能多样性 1 2018
... 随着人类干扰增加及全球气候剧变, 生物多样性改变、生态系统退化等生态环境问题成为当今科学研究的核心(Hector et al., 1999; Pardo et al., 2011; Boyer & Jetz, 2014).大量研究详实阐述了生物多样性变化对生态系统功能及服务的影响(Soliveres et al., 2016; 陈悦等, 2018), 越来越多的研究关注各类生物多样性之间的相互影响及其在生态系统能量流动和物质循环中的作用(Ramus & Long, 2016).作为生态系统中主要的生产者和分解者, 植物和土壤微生物多样性协同影响生态系统功能和服务(Wall & Moore, 1999; Bardgett & van der Putten, 2014).但我们对自然群落中, 植物多样性与微生物多样性之间的相互影响机制知之甚少(Rakowski & Cardinale, 2016), 甚至难以确定植物多样性本身是促进还是抑制土壤微生物多样性. ...
独山子区优势草本植物根际与非根际土壤微生物功能多样性 1 2018
... 随着人类干扰增加及全球气候剧变, 生物多样性改变、生态系统退化等生态环境问题成为当今科学研究的核心(Hector et al., 1999; Pardo et al., 2011; Boyer & Jetz, 2014).大量研究详实阐述了生物多样性变化对生态系统功能及服务的影响(Soliveres et al., 2016; 陈悦等, 2018), 越来越多的研究关注各类生物多样性之间的相互影响及其在生态系统能量流动和物质循环中的作用(Ramus & Long, 2016).作为生态系统中主要的生产者和分解者, 植物和土壤微生物多样性协同影响生态系统功能和服务(Wall & Moore, 1999; Bardgett & van der Putten, 2014).但我们对自然群落中, 植物多样性与微生物多样性之间的相互影响机制知之甚少(Rakowski & Cardinale, 2016), 甚至难以确定植物多样性本身是促进还是抑制土壤微生物多样性. ...
Biodiversity inhibits parasites: broad evidence for the dilution effect 1 2015
... 再次, 植物物种丰富度与土壤细菌多样性显著负相关, 这与混播降低土壤细菌多样性的结果一致.我们发现人工草地群落构建13年后, 植物物种丰富度与土壤细菌的丰富度和Shannon多样性之间都有显著的负相关关系, 而且对丰富度的影响更大.与单播和物种较少的群落相比, 混播和物种更多的群落中土壤细菌多样性较低可能是高的植物多样性抑制了土壤细菌中多度较少的稀有物种(Civitello et al., 2015), 致使丰富度的降低比Shannon多样性(稀有种影响较小)降低更明显(Field et al., 2020).另外, 有研究表明土壤中变形菌门的相对丰度的降低与土壤退化相关(李世雄等, 2020), 我们发现相对于单播处理, 混播处理中土壤变形菌门相对丰度显著升高, 单一细菌类群相对丰度的升高可能会导致土壤细菌多样性整体降低.同时微生物群落结构分析也发现, 与单播相比, 不同的混播群落间土壤细菌群落组成差异更大, 这一结果也指示混播影响土壤细菌某些类群的相对丰度, 从而改变了土壤细菌群落结构及多样性.然而, 我们也难以明确植物物种多样性的增加如何抑制影响土壤微生物多样性.从该实验及与之相关的研究(王宇彤和牛克昌, 2020; 赵兴鸽等, 2020a, 2020b)中, 我们只能确定植物多样性对土壤细菌的影响比对土壤真菌的影响更显著(Liu et al., 2020), 对丰富度的影响比对Shannon多样性的影响更显著, 对少见种的影响比优势种的影响更加显著; 并且植物多样性对土壤微生物的影响只是部分通过改变土壤理化因子和有机质, 更多可能是以直接作用或其他未知的途径影响.将来的研究不仅需要进一步验证这些现象的普适性, 而且更应关注一些未知的影响途径.例如, 多样性的植物群落是否会产生更加多样性的化学物质以抑制土壤微生物多样性, 尤其是那些随机漂泊的少见种(Hao et al., 2016; Wang et al., 2017). ...
Fungal/bacterial ratios in grasslands with contrasting nitrogen management 1 2006
... 其次, 虽然混播和施肥改变了土壤理化因子或植物生产力, 但这些变化不足以解释土壤微生物多样性在处理间的差异.例如, 混播处理显著降低了土壤pH和土壤全氮含量、增加了土壤全磷含量, 但无论是土壤pH还是全氮、全磷都与土壤微生物多样性无显著相关性; 施肥显著增加了植物地上生物量, 但植物地上生物量改变与细菌和真菌的多样性变化没有显著相关.这并不能说明土壤pH和土壤养分对土壤微生物多样性没有影响, 更可能是由于其影响错综复杂, 如pH减小不仅会直接影响土壤细菌多样性(Fierer & Jackson, 2006), 也会通过影响食细菌土壤线虫多度等多种间接途径影响土壤细菌多样性(Mulder et al., 2005; 王宇彤和牛克昌, 2020), 使pH对微生物的直接影响和间接影响相互抵消, 二者关系总体不显著.同理, 虽然很多研究报道了土壤微生物多样性和土壤肥力相关(周正虎和王传宽, 2016; Delgado-Baquerizo et al., 2017; Bahram et al., 2018; Yang et al., 2019a, 2019c), 但我们并不清楚土壤全氮和全磷含量与土壤微生物之间的确切关系(Delgado-Baquerizo et al., 2017); 土壤氮磷含量, 尤其是速效氮磷含量, 与土壤微生物多样性的相互影响更有可能通过植物介导(Hacker et al., 2015; Chen et al., 2019).如, 在青藏高原高寒草地, 在土壤氮较充足的情况下, 土壤速效磷含量的增加显著促进植物生产力和功能多样性(Niu et al., 2016a, 2016b), 为土壤生物提供丰度的资源和微环境, 使高寒草地群落中土壤速效磷含量与土壤微生物(赵兴鸽等, 2020a, 2020b)及土壤线虫(王宇彤和牛克昌, 2020)多样性都有明显正相关关系.本研究中, 我们也发现施肥增加土壤速效磷含量, 速效磷含量与土壤细菌的丰富度有显著正相关关系.作为土壤微生物最重要的碳源, 植物生产力和土壤有机质含量往往和土壤细菌多样性正相关而与土壤真菌多样性负相关(Wardle et al., 2004; de Vries et al., 2006; Delgado- Baquerizo et al., 2017); 我们的研究中也发现土壤有机碳含量与土壤真菌丰富度负相关, 但其与土壤细菌多样性并无显著相关性.此外, 混播和施肥对土壤有机碳含量影响不显著, 我们也难以澄清土壤有机碳含量如何影响土壤微生物多样性. ...
Changes in belowground biodiversity during ecosystem development 1 2019
... 土壤微生物多样性不仅受土壤环境因子深刻影响(Zhang & Han, 2012; Zhou et al., 2017), 而且与地上植物生产力和多样性紧密相关(Tilman et al., 2014; Zhang et al., 2017; Delegado-Baquerizo et al., 2019).大量的调查研究报道了自然群落中, 高的植物多样性和生产力为微生物提供更丰富的资源等, 从而促进土壤微生物多样性, 即多样性促进多样性假说(Leff et al., 2018).如前所述, 由于自然群落中植物多样性和生产力与环境变量协变, 我们难以分离植物多样性的作用, 确定植物多样性本身是否会促进土壤微生物多样性(Weisser et al., 2017).本研究中, 我们基于环境一致下的长期牧草单播和混播野外控制实验, 发现总体上相对于单播处理, 各组合的混播处理并未显著增加土壤细菌和真菌多样性, 甚至混播显著降低了土壤细菌多样性; 而施肥虽然显著地增加了植物生产力等资源, 但并没有明显增加土壤微生物多样性.这一结果与自然群落中的调查研究并不相符, 其中缘由值得深究. ...
It is elemental: soil nutrient stoichiometry drives bacterial diversity 5 2017
... 根据多样性促进多样性假说(diversity begets diversity), 各种植物“特异性”为土壤微生物提供基质, 如根系及根系分泌物、凋落物等(Leff et al., 2018), 因此多样性高的植物群落为微生物提供更丰富和多样性的食物资源, 从而促进土壤微生物多样性; 大量且多样的微生物又反馈调控凋落物分解等营养循环过程, 促进植物生产力和多样性(Wardle & Bardgett, 2010).与理论预测一致, 很多的调查研究发现植物物种或功能多样性高的地方, 土壤微生物多样性也高(Tilman et al., 2014; Delgado- Baquerizo et al., 2019).然而, 这些野外调查研究中, 土壤环境因子和植物多样性往往联系紧密, 甚至协同共变、交互影响土壤微生物多样性, 我们难以区分和阐明植物多样性和土壤因子的各自作用.我们不明确特定群落中土壤微生物多样性较高是由于环境更适宜、非生物因子筛选作用较小、区域物种库中物种较丰富所致(牛克昌等, 2009), 还是由于生产力高、凋落物等食物资源更丰富或是由植物多样性影响所致(Tilman et al., 2014; Delgado-Baquerizo et al., 2019).虽然有些控制实验中也发现植物物种丰富度较高的处理中土壤微生物多样性较高(Wagg et al., 2014).但这些实验中, 植物物种丰富度往往与生产力、凋落物和土壤肥力都有很强的正相关关系(Lange et al., 2015), 也难证实植物多样性本身促进土壤微生物多样性(Weisser et al., 2017).更多的控制实验和野外调查实验发现植物多样性与土壤微生物之间的关系并不显著, 其中机理可能与多个影响因素相互抵消有关(Prober et al., 2015).例如, Wardle等(2012)在火烧演替序列的长期实验中发现土壤微生物多样性与植物多样性和土壤碳储量并不相关, 可能是由于植物群落组成和土壤肥力对土壤微生物的影响相反, 且相互抵消.近年来也有理论和实验认为, 自然群落中很多物种在功能上是冗余的, 即多种物种共同维持一种或几种生态系统功能, 为此也就不难理解为什么我们观测到群落物种相对较少时, 植物物种多样性与土壤微生物多样性关系密切, 而物种相对较多时二者并没有很显著的相关性(Weisser et al., 2017).同样, Porazinska等(2018)在高寒生态系统中也发现, 群落演替早期, 由于植物生长对土壤碳氮输入及酶活性的促进, 植物多样性与土壤微生物有很好的正相关关系; 而在演替晚期二者并不相关.总之, 大量的证据表明植物群落对土壤微生物多样性的影响主要是通过影响土壤环境和肥力等属性, 而由于地上地下的复杂关系, 植物多样性本身和地下土壤微生物多样性可能并不相关(Prober et al., 2015; Wardle, 2016). ...
... ; Delgado-Baquerizo et al., 2019).虽然有些控制实验中也发现植物物种丰富度较高的处理中土壤微生物多样性较高(Wagg et al., 2014).但这些实验中, 植物物种丰富度往往与生产力、凋落物和土壤肥力都有很强的正相关关系(Lange et al., 2015), 也难证实植物多样性本身促进土壤微生物多样性(Weisser et al., 2017).更多的控制实验和野外调查实验发现植物多样性与土壤微生物之间的关系并不显著, 其中机理可能与多个影响因素相互抵消有关(Prober et al., 2015).例如, Wardle等(2012)在火烧演替序列的长期实验中发现土壤微生物多样性与植物多样性和土壤碳储量并不相关, 可能是由于植物群落组成和土壤肥力对土壤微生物的影响相反, 且相互抵消.近年来也有理论和实验认为, 自然群落中很多物种在功能上是冗余的, 即多种物种共同维持一种或几种生态系统功能, 为此也就不难理解为什么我们观测到群落物种相对较少时, 植物物种多样性与土壤微生物多样性关系密切, 而物种相对较多时二者并没有很显著的相关性(Weisser et al., 2017).同样, Porazinska等(2018)在高寒生态系统中也发现, 群落演替早期, 由于植物生长对土壤碳氮输入及酶活性的促进, 植物多样性与土壤微生物有很好的正相关关系; 而在演替晚期二者并不相关.总之, 大量的证据表明植物群落对土壤微生物多样性的影响主要是通过影响土壤环境和肥力等属性, 而由于地上地下的复杂关系, 植物多样性本身和地下土壤微生物多样性可能并不相关(Prober et al., 2015; Wardle, 2016). ...
... 其次, 虽然混播和施肥改变了土壤理化因子或植物生产力, 但这些变化不足以解释土壤微生物多样性在处理间的差异.例如, 混播处理显著降低了土壤pH和土壤全氮含量、增加了土壤全磷含量, 但无论是土壤pH还是全氮、全磷都与土壤微生物多样性无显著相关性; 施肥显著增加了植物地上生物量, 但植物地上生物量改变与细菌和真菌的多样性变化没有显著相关.这并不能说明土壤pH和土壤养分对土壤微生物多样性没有影响, 更可能是由于其影响错综复杂, 如pH减小不仅会直接影响土壤细菌多样性(Fierer & Jackson, 2006), 也会通过影响食细菌土壤线虫多度等多种间接途径影响土壤细菌多样性(Mulder et al., 2005; 王宇彤和牛克昌, 2020), 使pH对微生物的直接影响和间接影响相互抵消, 二者关系总体不显著.同理, 虽然很多研究报道了土壤微生物多样性和土壤肥力相关(周正虎和王传宽, 2016; Delgado-Baquerizo et al., 2017; Bahram et al., 2018; Yang et al., 2019a, 2019c), 但我们并不清楚土壤全氮和全磷含量与土壤微生物之间的确切关系(Delgado-Baquerizo et al., 2017); 土壤氮磷含量, 尤其是速效氮磷含量, 与土壤微生物多样性的相互影响更有可能通过植物介导(Hacker et al., 2015; Chen et al., 2019).如, 在青藏高原高寒草地, 在土壤氮较充足的情况下, 土壤速效磷含量的增加显著促进植物生产力和功能多样性(Niu et al., 2016a, 2016b), 为土壤生物提供丰度的资源和微环境, 使高寒草地群落中土壤速效磷含量与土壤微生物(赵兴鸽等, 2020a, 2020b)及土壤线虫(王宇彤和牛克昌, 2020)多样性都有明显正相关关系.本研究中, 我们也发现施肥增加土壤速效磷含量, 速效磷含量与土壤细菌的丰富度有显著正相关关系.作为土壤微生物最重要的碳源, 植物生产力和土壤有机质含量往往和土壤细菌多样性正相关而与土壤真菌多样性负相关(Wardle et al., 2004; de Vries et al., 2006; Delgado- Baquerizo et al., 2017); 我们的研究中也发现土壤有机碳含量与土壤真菌丰富度负相关, 但其与土壤细菌多样性并无显著相关性.此外, 混播和施肥对土壤有机碳含量影响不显著, 我们也难以澄清土壤有机碳含量如何影响土壤微生物多样性. ...
... ), 但我们并不清楚土壤全氮和全磷含量与土壤微生物之间的确切关系(Delgado-Baquerizo et al., 2017); 土壤氮磷含量, 尤其是速效氮磷含量, 与土壤微生物多样性的相互影响更有可能通过植物介导(Hacker et al., 2015; Chen et al., 2019).如, 在青藏高原高寒草地, 在土壤氮较充足的情况下, 土壤速效磷含量的增加显著促进植物生产力和功能多样性(Niu et al., 2016a, 2016b), 为土壤生物提供丰度的资源和微环境, 使高寒草地群落中土壤速效磷含量与土壤微生物(赵兴鸽等, 2020a, 2020b)及土壤线虫(王宇彤和牛克昌, 2020)多样性都有明显正相关关系.本研究中, 我们也发现施肥增加土壤速效磷含量, 速效磷含量与土壤细菌的丰富度有显著正相关关系.作为土壤微生物最重要的碳源, 植物生产力和土壤有机质含量往往和土壤细菌多样性正相关而与土壤真菌多样性负相关(Wardle et al., 2004; de Vries et al., 2006; Delgado- Baquerizo et al., 2017); 我们的研究中也发现土壤有机碳含量与土壤真菌丰富度负相关, 但其与土壤细菌多样性并无显著相关性.此外, 混播和施肥对土壤有机碳含量影响不显著, 我们也难以澄清土壤有机碳含量如何影响土壤微生物多样性. ...
Alterations in soil bacterial community in relation to Spartina alterniflora Loisel. invasion chronosequence in the eastern Chinese coastal wetlands 1 2019
... 其次, 虽然混播和施肥改变了土壤理化因子或植物生产力, 但这些变化不足以解释土壤微生物多样性在处理间的差异.例如, 混播处理显著降低了土壤pH和土壤全氮含量、增加了土壤全磷含量, 但无论是土壤pH还是全氮、全磷都与土壤微生物多样性无显著相关性; 施肥显著增加了植物地上生物量, 但植物地上生物量改变与细菌和真菌的多样性变化没有显著相关.这并不能说明土壤pH和土壤养分对土壤微生物多样性没有影响, 更可能是由于其影响错综复杂, 如pH减小不仅会直接影响土壤细菌多样性(Fierer & Jackson, 2006), 也会通过影响食细菌土壤线虫多度等多种间接途径影响土壤细菌多样性(Mulder et al., 2005; 王宇彤和牛克昌, 2020), 使pH对微生物的直接影响和间接影响相互抵消, 二者关系总体不显著.同理, 虽然很多研究报道了土壤微生物多样性和土壤肥力相关(周正虎和王传宽, 2016; Delgado-Baquerizo et al., 2017; Bahram et al., 2018; Yang et al., 2019a, 2019c), 但我们并不清楚土壤全氮和全磷含量与土壤微生物之间的确切关系(Delgado-Baquerizo et al., 2017); 土壤氮磷含量, 尤其是速效氮磷含量, 与土壤微生物多样性的相互影响更有可能通过植物介导(Hacker et al., 2015; Chen et al., 2019).如, 在青藏高原高寒草地, 在土壤氮较充足的情况下, 土壤速效磷含量的增加显著促进植物生产力和功能多样性(Niu et al., 2016a, 2016b), 为土壤生物提供丰度的资源和微环境, 使高寒草地群落中土壤速效磷含量与土壤微生物(赵兴鸽等, 2020a, 2020b)及土壤线虫(王宇彤和牛克昌, 2020)多样性都有明显正相关关系.本研究中, 我们也发现施肥增加土壤速效磷含量, 速效磷含量与土壤细菌的丰富度有显著正相关关系.作为土壤微生物最重要的碳源, 植物生产力和土壤有机质含量往往和土壤细菌多样性正相关而与土壤真菌多样性负相关(Wardle et al., 2004; de Vries et al., 2006; Delgado- Baquerizo et al., 2017); 我们的研究中也发现土壤有机碳含量与土壤真菌丰富度负相关, 但其与土壤细菌多样性并无显著相关性.此外, 混播和施肥对土壤有机碳含量影响不显著, 我们也难以澄清土壤有机碳含量如何影响土壤微生物多样性. ...
Seed-to-seed potential allelopathic effects between Ligularia virgaurea and native grass species of Tibetan alpine grasslands 1 2011
 ,*南京大学生命科学学院, 南京 210023
,*南京大学生命科学学院, 南京 210023

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT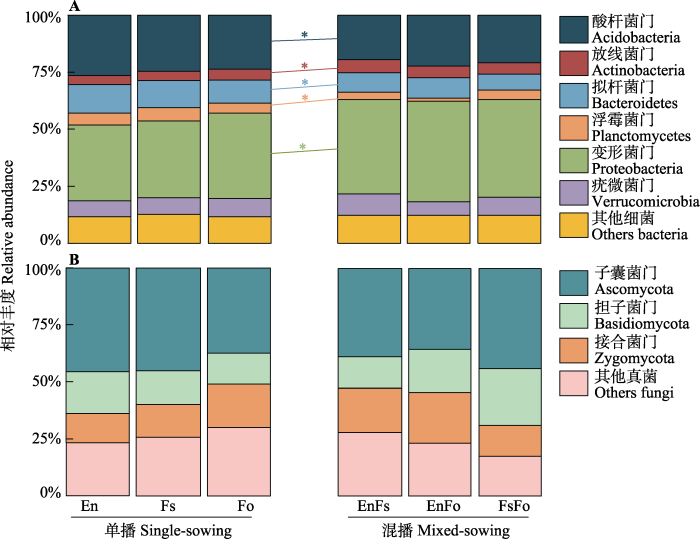 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT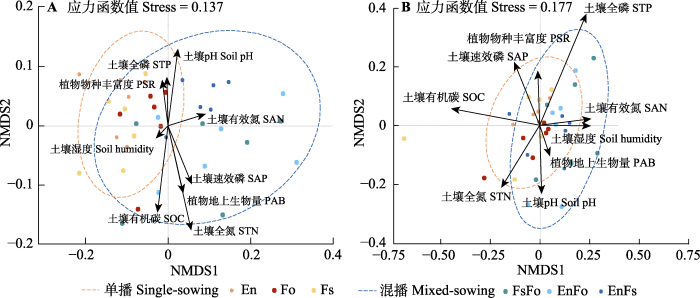 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT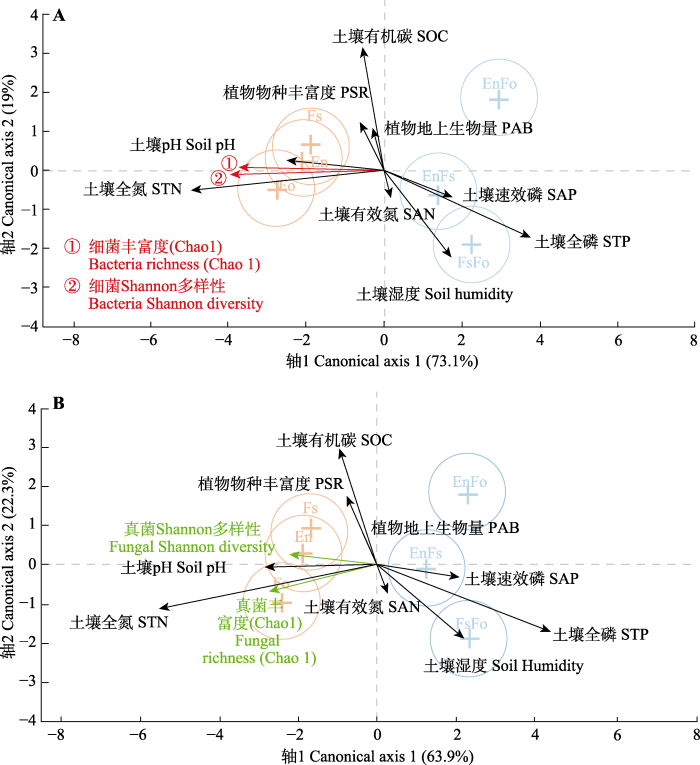 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}