Photodamage to photosystem in a typically shade-tolerant species Panax notoginseng exposed to a sudden increase in light intensity
Hong-Min WU,1,2, Sheng-Pu SHUANG1,2, Jin-Yan ZHANG1,2, Zhu CUN1,2, Zhen-Gui MENG1,2, Long-Gen LI1,2, Ben-Cai SHA1, Jun-Wen CHEN,1,2,*1National & Local Joint Engineering Research Center on Germplasm Innovation & Utilization of Chinese Medicinal Materials in Southwestern China, Key Laboratory of Medicinal Plant Biology of Yunnan Proince, Yunnan Agricultural University, Kunming 650201, China 2College of Agronomy and Biotechnology, Yunnan Agricultural University, Kunming 650201, China
National Natural Science Foundation of China(81860676) National Natural Science Foundation of China(81360609) Key Science and Technology Project of Yunnan Province(2016ZF001) Key Science and Technology Project of Yunnan Province(2017ZF001)
Abstract Aims Photodamage to a shade-tolerant species is common due to a sudden increase in growth light intensity. However, it is unknown about the underlying mechanism of the sensitivity of the shade-tolerant species to high light. The objective of the present study was to elucidate the mechanisms involved in the inability of the typically shade-tolerant species Panax notoginseng to survive under natural full-light condition. Methods The relative chlorophyll content (SPAD), photosynthetic parameters and chlorophyll fluorescence parameters were continuously examined in P. notoginseng when transferred from shade (10% of full sunlight) to full sunlight for three days. Important findings The net photosynthetic rate (Pn) of P. notoginseng exposed to full sunlight condition showed a “double-peaked” diurnal curve, and Pn decreased with the prolonged days of full light treatment. The SPAD value, water utilization efficiency and light use efficiency were significantly decreased under full sunlight condition. Furthermore, the maximum fluorescence signal of the P700 reaction center, electron transfer rate of photosystem II (PSII), the maximum quantum efficiency of PSII under dark adaptation and maximum quantum efficiency of PSII under light adaptation were significantly lower in full sunlight than those under shading condition, while the fraction of energy passively dissipated in the forms of heat and fluorescence, energy dissipation due to acceptor side limitation of PSI, and cyclic electron flow were significantly higher under the full light condition. Moreover, the sudden increase in growth light intensity caused a significant change in the fluorescence induction kinetic curve and significantly increased the fluorescence yield on the donor and acceptor side of PSII. The oxygen-evolving complex activity in the donor side of PSII was impaired under full sunlight. Furthermore, the electron transfer in the acceptor side of PSII was inhibited and the over-reduction of the acceptor side of PSI was caused by PSI photoinhibition. The study reveals that the full sunlight might induce the irreversible damage to PSII and the moderate photoinhibition to PSI in shade-tolerant species, and it may be an important underlying mechanisms why the shade-tolerant speciesP. notoginseng cannot survive under full sunlight. Keywords:shade-tolerant species;full sunlight;photosynthesis;photosystem;photoprotection;Panax notoginseng
PDF (1348KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 武洪敏, 双升普, 张金燕, 寸竹, 孟珍贵, 李龙根, 沙本才, 陈军文. 短期生长环境光强骤增导致典型阴生植物三七光系统受损的机制. 植物生态学报, 2021, 45(4): 404-419. DOI: 10.17521/cjpe.2021.0013 WU Hong-Min, SHUANG Sheng-Pu, ZHANG Jin-Yan, CUN Zhu, MENG Zhen-Gui, LI Long-Gen, SHA Ben-Cai, CHEN Jun-Wen. Photodamage to photosystem in a typically shade-tolerant species Panax notoginseng exposed to a sudden increase in light intensity . Chinese Journal of Plant Ecology, 2021, 45(4): 404-419. DOI: 10.17521/cjpe.2021.0013
Fig. 3Diurnal pattern of relative chlorophyll content (SPAD value) in shade treatment and full sunlight treatment (mean ± SE, n = 7). *, significant difference between the two treatments at same measurement time ( p< 0.05).
Fig. 4Diurnal pattern of net photosynthetic rate (A), intercellular CO2 concentration (B) and stomatal conductance (C) in shade treatment and full sunlight treatment (mean ± SE, n = 7). *, significant difference in gas exchange parameters between the two treatments at same measurement time ( p< 0.05).
Fig. 5Water use efficiency (A) and light use efficiency (B) of Panax notoginseng exposed to shade and full sunlight conditions on the first, second and third day of the experiment, respectively (mean ± SE, n = 7). Different lowercase letters between shade and full sunlight conditions indicate significant difference (p< 0.05).
Fig. 6Diurnal pattern of maximum quantum efficiency under light adaptation (A), maximum quantum efficiency under dark adaptation (B) and non-photochemical quenching of PSII (C) in shade treatment and full sunlight treatment (mean ± SE, n = 7). *, significant difference between the two treatments at same measurement time ( p< 0.05).
Fig. 7The maximum quantum efficiency under dark adaptation (A), potential activity of PSII reaction center (B) and the maximum fluorescence signal of the P700 reaction center (C) of Panax notoginseng exposed to shade and full sunlight conditions on the first, second and third day of the experiment, respectively (mean ± SE, n = 7). *, significant difference between the two treatments at same measurement day ( p< 0.05).
Fig. 8Changes in rapid fluorescence induction kinetic curves (WO-J and Δ WO-J) in leaves of Panax notoginseng transferred from a shade environment with 10% transmittance to full sunlight. The abscissa is plotted on a linear time scale. W,the characteristic point at 100 μs (K point). Day 0, Sunlight-day 1, Sunlight-day 2, Sunlight-day 3 represent the exposure to low light, and the transfer to full sunlight for one, two and three days. Different lowercase letters indicate significant differences ( p< 0.05).
Fig. 9Daily pattern of parameters related to the rapid fluorescence induction kinetic curve in leaves of Panax notoginsengtransferred from a shade environment with 10% transmittance to full sunlight (mean ± SE, n = 7). Different letters indicate significant difference between shade and full sunlight conditions (p< 0.05).
Fig. 10Effect of shade and full sunlight treatment on electron transfer and photosystem energy partitioning (mean ± SE, n = 7). LL, 10% transmittance-grown Panax notoginseng (230 μmol·m -2·s-1, 13:00); HL, full sunlight-grown Panax notoginseng (2 300 μmol·m -2·s-1, 13:00). Y(I), effective quantum yield of PSI; Y(ND), heat dissipation efficiency at the donors quantum yield of PSI; Y(NA), quantum yield of PSI non-photochemical energy dissipation due to acceptor; Y(II), effective quantum yield of PSII; Y(NPQ), quantum yield of energy dissipation in PSII; Y(NO), fraction of energy passively dissipated in forms of heat and fluorescence. ETR(I), electron transport rate of PSI; ETR(II), electron transport rate of PSII;CEF, cyclic electron flow around PSI. *, significant difference between the two treatments at same measurement day ( p< 0.05).
Fig. 11Changes in electron transfer ratio in leaves of Panax notoginseng exposed to shade and full sunlight conditions on the first, second and third day of the experiment, respectively (mean ± SE, n = 7). ETR(I), electron transport rate of PSI; ETR(II), electron transport rate of PSII;CEF, cyclic electron flow around PSI. *, significant difference between the two treatments ( p< 0.05).
AllakhverdievSI, NishiyamaY, TakahashiS, MiyairiS, SuzukiI, MurataN (2005). Systematic analysis of the relation of electron transport and ATP synthesis to the photodamage and repair of photosystem II in Synechocystis Plant Physiology, 137, 263-273. PMID:15618415 [本文引用: 1] The photosynthetic machinery and, in particular, the photosystem II (PSII) complex are susceptible to strong light, and the effects of strong light are referred to as photodamage or photoinhibition. In living organisms, photodamaged PSII is rapidly repaired and, as a result, the extent of photoinhibition represents a balance between rates of photodamage and the repair of PSII. In this study, we examined the roles of electron transport and ATP synthesis in these two processes by monitoring them separately and systematically in the cyanobacterium Synechocystis sp. PCC 6803. We found that the rate of photodamage, which was proportional to light intensity, was unaffected by inhibition of the electron transport in PSII, by acceleration of electron transport in PSI, and by inhibition of ATP synthesis. By contrast, the rate of repair was reduced upon inhibition of the synthesis of ATP either via PSI or PSII. Northern blotting and radiolabeling analysis with [(35)S]Met revealed that synthesis of the D1 protein was enhanced by the synthesis of ATP. Our observations suggest that ATP synthesis might regulate the repair of PSII, in particular, at the level of translation of the psbA genes for the precursor to the D1 protein, whereas neither electron transport nor the synthesis of ATP affects the extent of photodamage.
BaiKD, LiaoDB, JiangDB, CaoKF (2008). Photosynthetic induction in leaves of co-occurring Fagus lucida and Castanopsis lamontii saplings grown in contrasting light environments Trees, 22, 449-462. DOI:10.1007/s00468-007-0205-4URL [本文引用: 2]
BarthC, KrauseGH, WinterK (2001). Responses of photosystem I compared with photosystem II to high-light stress in tropical shade and sun leaves Plant, Cell & Environment, 24, 163-176. [本文引用: 2]
ChenJW, KuangSB, LongGQ, MengZG, LiLG, ChenZJ, ZhangGH, YangSC (2014). Steady-state and dynamic photosynthetic performance and nitrogen partitioning in the shade-demanding plant Panax notoginseng under different levels of growth irradiance c Acta Physiologiae Plantarum, 36, 2409-2420. DOI:10.1007/s11738-014-1614-9URL [本文引用: 2]
ChenJW, KuangSB, LongGQ, YangSC, MengZG, LiLG, ChenZJ, ZhangGH (2016). Photosynthesis, light energy partitioning, and photoprotection in the shade-demanding species Panax notoginseng under high and low level of growth irradiance Functional Plant Biology, 43, 479-491. DOI:10.1071/FP15283URL [本文引用: 1]
DongLH, HanQH, YangY, YuanM ( 2015). Photosynthetic characteristics of Chlorophytum capense var. medio- pictum under short duration high light intensity Acta Prataculturae Sinica, 24, 245-252. [本文引用: 3]
HuWH, ZhangSS, XiaoYA, YanXH ( 2015). Physiological responses and photo-protective mechanisms of two Rhododendron plants to natural sunlight after long term shading Chinese Journal of Plant Ecology, 39, 1093-1100. DOI:10.17521/cjpe.2015.0106URL [本文引用: 1]
HuangW ( 2012). The Significant Role of Cyclic Electron Flow in Plants Adaptation to Environmental Stresses PhD dissertation, University of Science and Technology of China, Hefei. [本文引用: 6]
HuangW, TikkanenM, ZhangSB (2018a). Photoinhibition of photosystem I in Nephrolepis falciformis depends on reactive oxygen species generated in the chloroplast stroma Photosynthesis Research, 137, 129-140. DOI:10.1007/s11120-018-0484-1URL [本文引用: 2]
HuangW, YangYJ, ZhangJL, HuH, ZhangSB (2017). Superoxide generated in the chloroplast stroma causes photoinhibition of photosystem I in the shade-establishing tree species Psychotria henryi Photosynthesis Research, 132, 293-303. DOI:10.1007/s11120-017-0389-4PMID:28432538 [本文引用: 1] Our previous studies indicated that high light induced significant photoinhibition of photosystem I (PSI) in the shade-establishing tree species Psychotria henryi. However, the underlying mechanism has not been fully clarified. In the present study, in order to investigate the mechanism of PSI photoinhibition in P. henryi, we treated detached leaves with constant high light in the presence of methyl viologen (MV) or a soluble α-tocopherol analog, 2,2,5,7,8-pentamethyl-6-chromanol (PMC). We found that MV significantly depressed photochemical quantum yields in PSI and PSII when compared to PMC. On condition that no PSI photoinhibition happened, although cyclic electron flow (CEF) was abolished in the MV-treated samples, P700 oxidation ratio was maintain at higher levels than the PMC-treated samples. In the presence of PMC, PSI photoinhibition little changed but PSII photoinhibition was significantly alleviated. Importantly, PSI photoinhibition was largely accelerated in the presence of MV, which stimulates the production of superoxide and subsequently other reactive oxygen species at the chloroplast stroma by accepting electrons from PSI. Furthermore, MV largely aggravated PSII photoinhibition when compared to control. These results suggest that high P700 oxidation ratio cannot prevent PSI photoinhibition in P. henryi. Furthermore, the superoxide produced in the chloroplast stroma is critical for PSI photoinhibition in the higher plant P. henryi, which is opposite to the mechanism underlying PSI photoinhibition in Arabidopsis thaliana and spinach. These findings highlight a new mechanism of PSI photoinhibition in higher plants.
HuangW, YangYJ, ZhangSB (2019). Photoinhibition of photosystem I under fluctuating light is linked to the insufficient ΔpH upon a sudden transition from low to high light Environmental and Experimental Botany, 160, 112-119. DOI:10.1016/j.envexpbot.2019.01.012 [本文引用: 2] Despite photosystem I (PSI) is susceptible to photoinhibition under fluctuating light in wild-type angiosperms, the underlying mechanism is not well known. Because proton gradient (Delta pH) across the thylakoid membranes plays a crucial role in protecting PSI, we hypothesized that PSI photoinhibition under fluctuating light may be linked to the formation of Delta pH. To test this hypothesis, we measured chlorophyll fluorescence, PSI redox state, and the electrochromic shift signal during transition from low to high light in two angiosperms Bletilla striate (Orchidaceae) and Arabidopsis thaliana. The measurement of fast P700 redox kinetics indicated that the fast re oxidation of P700 mediated by photo-reduction of O-2 was absent in B. striate. Furthermore, the redox state of PSI is highly determined by Delta pH in both species. For the first 20 s after transition from low to high light, both species could not build up a sufficient Delta pH, which was accompanied by the over-reduction of P700. During prolonged illumination at high light, the sufficient Delta pH made PSI to be highly oxidized. These results demonstrated that PSI photoinhibition under fluctuating light in wild-type angiosperms occurred mainly at the first 20s after an increase in light intensity, which was caused by the insufficient Delta pH.
KimJH, KimSJ, ChoSH, ChowWS, LeeCH (2005). Photosystem I acceptor side limitation is a prerequisite for the reversible decrease in the maximum extent of P700 oxidation after short-term chilling in the light in four plant species with different chilling sensitivities Physiologia Plantarum, 123, 100-107. DOI:10.1111/ppl.2005.123.issue-1URL [本文引用: 1]
KitajimaK (1994). Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees Oecologia, 98, 419-428. DOI:10.1007/BF00324232PMID:28313920 [本文引用: 1] Among 13 tropical tree species on Barro Colorado Island, species with high seedling mortality rates during the first year in shade had higher reltive growth rates (RGR) from germination to 2 months in both sun (23% full sun) and shade [2%, with and without lowered red: far red (R:FR) ratio] than shade tolerant species. Species with higher RGR in sun also had higher RGR in shade. These interspecific trends could be explained by differences in morphological traits and allocation paterns among species. Within each light regime, seedlings of shade-intolerant species had lower root: shoot ratios, higher leaf mass per unit area, and higher leaf area ratios (LAR) than shade tolerant species. In contrast, leaf gas exchange characteristics, or acclimation potential in these traits, had no relationship with seedling mortality rates in shade. In both shade tolerant and intolerant species, light saturated photosynthesis rates, dark respiration, and light compensation points were higher for sungrown seedlings than for shade-grown seedlings. Differences in R:FR ratio in shade did not affect gas exchange, allocation patterns, or growth rates of any species. Survival of young tree seedlings in shade did not depend on higher net photosynthesis or biomass accumulation rates in shade. Rather, species with higher RGR died faster in shade than species with lower RGR. This trend could be explained if survival depends on morphological characteristics likely to enhance defense against herbivores and pathogens, such as dense and tough leaves, a well-established root system, and high wood density. High construction costs for these traits, and low LAR as a consequence of these traits, should result in lower rates of whole-plant carbon gain and RGR for shade tolerant species than shade-intolerant species in shade as well as in sun.
LiPM, GaoHY, StrasserR ( 2005). Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study Acta Photophysiologica Sinica, 31, 559-566. [本文引用: 2]
LiXX, LiuBX, GuoZT, ChangYX, HeL, ChenF, LuBS ( 2013). Effects of NaCl stress on photosynthesis characteristics and fast chlorophyll fluorescence induction dynamics of Pistacia chinensisleaves Chinese Journal of Applied Ecology, 24, 2479-2484. [本文引用: 1]
LiZZ, LiuDH, ZhaoSW, JiangCD, ShiL ( 2014). Mechanisms of photoinhibition induced by high light in Hosta grown outdoors Chinese Journal of Plant Ecology, 38, 720-728. DOI:10.3724/SP.J.1258.2014.00067 [本文引用: 6] <p><em>Aims</em> It has long been recognized that photoinhibition of photosynthesis is induced by high light. However, our recent studies are not consistent with this traditional view. Therefore, the objective of this study is to explore the induction of photoinhibition and its mechanisms under full sunlight outdoors.</br> <em>Methods</em> Changes of leaf morphology, gas exchange, and chlorophyll a fluorescence were measured to investigate the induction and mechanisms of photoinhibition under high light in <em>Hosta</em>, which is a typical shade-tolerant plant.</br> <em>Important findings</em> Hosta plants grown under full sunlight (HT) and low light (LT) developed sun- and shade-type leaf morphological characteristics, respectively. Under a full sunlight, <em>Hosta</em> plants had lower photosynthetic rate and chlorophyll content than under the LT; whereas, there were only slight difference in the maximum quantum yield of photosystem II (<em>F</em><sub>v</sub>/<em>F</em><sub>m</sub>) between the two treatments, suggesting that <em>Hosta</em> plants could grow normally under full sunlight without severe photoinhibition. After transition from the low to a high light (LHT), the photosynthetic rate and maximum quantum yield of photosystem II decreased sharply, reflecting that the LHT treatment led to irreversibly inactivation of photosystem II. Additionally, the shape of chlorophyll a fluorescence transients also changed significantly; the relative fluorescence yield of the K and J steps were reduced by 24.3% and 34.2%, respectively, indicating that the acceptor side of photosystem II was damaged more severely than the donor side. Consequently, we postulate that photoinhibition in <em>Hosta</em> leaves is mainly induced by the sudden enhancement of light intensity outdoors. <em>Hosta</em> can acclimate to high irradiance through leaf development outdoors. Our finding is of great significance in understanding the acclimation of plants to high light and cultivation of shade-tolerant plants in field.</p> [ 李志真, 刘东焕, 赵世伟, 姜闯道, 石雷 ( 2014). 环境强光诱导玉簪叶片光抑制的机制 植物生态学报, 38, 720-728.] [本文引用: 6]
LiangY, LiYT, CheXK, LiYN, LuoJ, ZhangKH, ZhangZS, ZhaoSJ ( 2018). Effect of PSI photoinhibition on photosynthetic electron transport chain in wheat ( Triticum aestivum) leaves. Plant Physiology Journal, 54, 1426-1432. [本文引用: 2]
LiuSL, MaMD, PanYZ, WeiLL, HeCX, YangKM ( 2012). Effects of light regimes on photosynthetic characteristics and antioxidant system in seedlings of two alder species Chinese Journal of Plant Ecology, 36, 1062-1074. DOI:10.3724/SP.J.1258.2012.01062URL [本文引用: 1]
LuT ( 2016). The Response Mechanisms of Photoinhibiton and Photoprotection Intomato (Solaunm Lycopersicum L.) Leaves to Sub-high Temperature and High Light Intensity Stress. PhD dissertation, Shenyang Agricultural University Shenyang. [本文引用: 1]
MaZY, BehlingS, FordED (2014). The contribution of dynamic changes in photosynthesis to shade tolerance of two conifer species Tree Physiology, 34, 730-743. DOI:10.1093/treephys/tpu054URL [本文引用: 1]
MaxwellK, JohnsonGN (2000). Chlorophyll florescence—A practical guide Journal of Experimental Botany, 51, 659-668. PMID:10938857 [本文引用: 1] Chlorophyll fluorescence analysis has become one of the most powerful and widely used techniques available to plant physiologists and ecophysiologists. This review aims to provide an introduction for the novice into the methodology and applications of chlorophyll fluorescence. After a brief introduction into the theoretical background of the technique, the methodology and some of the technical pitfalls that can be encountered are explained. A selection of examples is then used to illustrate the types of information that fluorescence can provide.
MiyakeC, MiyataM, ShinzakiY, TomizawaKI (2005). CO2 response of cyclic electron flow around PSI (CEF-PSI) in tobacco leaves—Relative electron fluxes through PSI and PSII determine the magnitude of non-photochemical quenching (NPQ) of chl fluorescence Plant and Cell Physiology, 46, 629-637. DOI:10.1093/pcp/pci067URL [本文引用: 1]
MurataN, TakahashiS, NishiyamaY, AllakhverdievSI (2007). Photoinhibition of photosystem II under environmental stress Biochimica et Biophysica Acta (BBA): Bioenergetics, 1767, 414-421. [本文引用: 1]
OliveiraMG, OliosiG, PartelliFL, RamalhoJC (2018). Physiological responses of photosynthesis in black pepper plants under different shade levels promoted by intercropping with rubber trees Ciência e Agrotecnologia, 42, 513-526. DOI:10.1590/1413-70542018425020418URL [本文引用: 1]
ParkerWC, MohammedGH (2000). Photosynthetic acclimation of shade-grown red pine (Pinus resinosaAit.) seedlings to a high light environment New Forests, 19, 1-11. DOI:10.1023/A:1006668928091URL [本文引用: 1]
RascherU, NedbalL (2006). Dynamics of photosynthesis in fluctuating light Current Opinion in Plant Biology, 9, 671-678. PMID:17011815 [本文引用: 1] Our understanding of the molecular mechanisms of plant photosynthesis is expanding from insights into static fluxes in constant irradiance to an understanding of complex dynamic patterns in fluctuating light. Knowledge about regulatory interactions, information about relevant biological features that emerge in fluctuating light, and the new standards for sharing biological models allow world-wide consortia aimed at the comprehensive modeling of photosynthetic dynamics.
ShimizuM, IshidaA, HogetsuT (2005). Root hydraulic conductivity and whole-plant water balance in tropical saplings following a shade-to-sun transfer Oecologia, 143, 189-197. DOI:10.1007/s00442-004-1797-7URL [本文引用: 1]
SrivastavaA, GuisséB, GreppinH, StrasserRJ (1997). Regulation of antenna structure and electron transport in Photosystem II of Pisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP Biochimica et Biophysica Acta (BBA): Bioenergetics, 1320, 95-106. [本文引用: 1]
StrasserRJ, SrivastavaA, Tsimilli-MichaelM (2000). The fluorescence transient as a tool to characterize and screen photosynthetic samples//Yunus M, Pathre U, Mohanty P. Probing Photosynthesis: Mechanism, Regulation and Adaptation. Taylor and Francis Press,London. 445-480. [本文引用: 1]
StrasserRJ, Tsimilli-MichaelM, SrivastavaA (2004). Analysis of the Chlorophyll a Fluorescence Transient. Chlorophyll a Fluorescence Springer,Netherlands. 321-362. [本文引用: 1]
SunGC, ZhaoP, ZengXP ( 2004). Photosynthetic acclimation to growth-irradiance in two tree species of Magnoliaceae Acta Ecologica Sinica, 24, 1111-1117. [本文引用: 1]
ValladaresF, PearcyRW (1998). The functional ecology of shoot architecture in sun and shade plants of Heteromeles arbutifolia M. Roem., a Californian chaparral shrub Oecologia, 114, 1-10. DOI:10.1007/s004420050413PMID:28307546 [本文引用: 1] The functional roles of the contrasting morphologies of sun and shade shoots of the evergreen shrub Heteromeles arbutifolia were investigated in chaparral and understory habitats by applying a three-dimensional plant architecture simulation model, YPLANT. The simulations were shown to accurately predict the measured frequency distribution of photosynthetic photon flux density (PFD) on both the leaves and a horizontal surface in the open, and gave reasonably good agreement for the more complex light environment in the shade. The sun shoot architecture was orthotropic and characterized by steeply inclined (mean?=?71) leaves in a spiral phyllotaxy with short internodes. This architecture resulted in relatively low light absorption efficiencies (E ) for both diffuse and direct PFD, especially during the summer when solar elevation angles were high. Shade shoots were more plagiotropic with longer internodes and a pseudo-distichous phyllotaxis caused by bending of the petioles that positioned the leaves in a nearly horizontal plane (mean?=?5). This shade-shoot architecture resulted in higher E values for both direct and diffuse PFD as compared to those of the sun shoots. Differences in E between sun and shade shoots and between summer and winter were related to differences in projection efficiencies as determined by leaf and solar angles, and by differences in self shading resulting from leaf overlap. The leaves exhibited photosynthetic acclimation to the sun and the shade, with the sun leaves having higher photosynthetic capacities per unit area, higher leaf mass per unit area and lower respiration rates per unit area than shade leaves. Despite having 7 times greater available PFD, sun shoots absorbed only 3 times more and had daily carbon gains only double of those of shade shoots. Simulations showed that sun and shade plants performed similarly in the open light environment, but that shade shoots substantially outperformed sun shoots in the shade light environment. The shoot architecture observed in sun plants appears to achieve an efficient compromise between maximizing carbon gain while minimizing the time that the leaf surfaces are exposed to PFDs in excess of those required for light saturation of photosynthesis and therefore potentially photoinhibitory.
van HeerdenPDR, StrasserRJ, KrügerGHJ (2004). Reduction of dark chilling stress in N2-fixing soybean by nitrate as indicated by chlorophyll a fluorescence kinetics Physiologia Plantarum, 121, 239-249. DOI:10.1111/ppl.2004.121.issue-2URL [本文引用: 1]
WayDA, PearcyRW (2012). Sunflecks in trees and forests: from photosynthetic physiology to global change biology Tree Physiology, 32, 1066-1081. DOI:10.1093/treephys/tps064URL [本文引用: 1]
XieHT, YuMK, ChengXR ( 2017). Effects of light intensity variation on nitrogen and phosphorus contents, allocation and limitation in five shade-enduring plants Chinese Journal of Plant Ecology, 41, 559-569. DOI:10.17521/cjpe.2016.0248URL [本文引用: 1]
XuXZ, ZhangJY, ZhangGH, LongGQ, YangSC, ChenZJ, WeiFG, ChenJW ( 2018). Effects of light intensity on photosynthetic capacity and light energy allocation in Panax notoginseng Chinese Journal of Applied Ecology, 29, 193-204. [本文引用: 1]
YuHX ( 2017). The Effects on the Growth of Hosta Cultivar under Shading and Interactive Effects of Nitrogen and Light Condition and Comprehensive Evaluation about Hosta Cultivar Master degree dissertation, Shenyang Agricultural University,Shenyang. [本文引用: 1]
ZhangQ, ZhangTJ, ChowWS, XieX, ChenYJ, PengCL (2015). Photosynthetic characteristics and light energy conversions under different light environments in five tree species occupying dominant status at different stages of subtropical forest succession Functional Plant Biology, 42, 609-619. DOI:10.1071/FP14355URL [本文引用: 1]
ZhangSP, SchellerHV (2004). Photoinhibition of photosystem I at chilling temperature and subsequent recovery in Arabidopsis thaliana Plant and Cell Physiology, 45, 1595-1602. DOI:10.1093/pcp/pch180URL [本文引用: 1]
ZhengF, LiZJ, QiuZJ, ZhaoHB, ZhouGY ( 2020). Effects of understory light on functional traits of evergreen broad- leaved forest saplings in Nanling Mountains, Guangdong Province Acta Ecologica Sinica, 40, 4516-4527. [本文引用: 1]
ZivcakM, BresticM, KunderlikovaK, SytarO, AllakhverdievSI (2015). Repetitive light pulse-induced photoinhibition of photosystem I severely affects CO2 assimilation and photoprotection in wheat leaves Photosynthesis Research, 126, 449-463. DOI:10.1007/s11120-015-0121-1PMID:25829027 [本文引用: 1] It was previously found that photosystem I (PSI) photoinhibition represents mostly irreversible damage with a slow recovery; however, its physiological significance has not been sufficiently characterized. The aim of the study was to assess the effect of PSI photoinhibition on photosynthesis in vivo. The inactivation of PSI was done by a series of short light saturation pulses applied by fluorimeter in darkness (every 10 s for 15 min), which led to decrease of both PSI (~60 %) and photosystem II (PSII) (~15 %) photochemical activity. No PSI recovery was observed within 2 days, whereas the PSII was fully recovered. Strongly limited PSI electron transport led to an imbalance between PSII and PSI photochemistry, with a high excitation pressure on PSII acceptor side and low oxidation of the PSI donor side. Low and delayed light-induced NPQ and P700(+) rise in inactivated samples indicated a decrease in formation of transthylakoid proton gradient (ΔpH), which was confirmed also by analysis of electrochromic bandshift (ECSt) records. In parallel with photochemical parameters, the CO2 assimilation was also strongly inhibited, more in low light (~70 %) than in high light (~45 %); the decrease was not caused by stomatal closure. PSI electron transport limited the CO2 assimilation at low to moderate light intensities, but it seems not to be directly responsible for a low CO2 assimilation at high light. In this regard, the possible effects of PSI photoinhibition on the redox signaling in chloroplast and its role in downregulation of Calvin cycle activity are discussed. Systematic analysis of the relation of electron transport and ATP synthesis to the photodamage and repair of photosystem II in Synechocystis 1 2005
... 自然环境中生长的植物, 其PSI通常对高光环境不敏感(Barth et al., 2001 ).一般只有在低温弱光、常温频闪光或是饱和脉冲处理植物时, PSI才会发生光抑制(Zhang & Scheller, 2004; 梁英等, 2018; Yamamoto & Shikanai, 2019).而滇南九节(Psychotria henryi)和镰叶肾蕨(Nephrolepis falciformis)等喜阴植物叶片的PSI却在恒定高光处理下发生光抑制(Huang et al., 2017 , 2018a).PSI发生光抑制的显著特征是PSI活性下降(Huang et al., 2018b ; 梁英等, 2018).Huang等(2018b)研究发现, 三七在高光下处理30和60 min时, Pm不受影响, 说明三七的PSI对高光胁迫不敏感.但本研究中, 短时环境强光导致三七的Pm显著下降, 该结果与上述的结果不一致, 可能是因为本研究采用的是全日照下的全天高光连续处理3天, 而Huang等(2018b)研究的高光处理时间较短(处理时间仅为30和60 min)从而尚未影响到PSI的活性.另有研究表明, Y(NA)增加也可作为PSI发生光抑制的指标(Kim et al., 2005 ; Yamori et al., 2016 ; Yang et al., 2019a ).本研究中, 正午强光下三七具有较高的Y(NA)(图10A), 该结果与拟南芥(Arabidopsis thaliana)和白及(Bletilla striata)从低到高光转变后Y(NA)升高的结果(Huang et al., 2019 ; Yang et al., 2019a )一致, 由此推测, 环境光强增加后, 因无法形成足够的pH, 类囊体管腔酸化水平低, 从PSII到PSI的电子流引起受体侧羟基自由基的积累, 最终导致PSI发生光抑制(黄伟, 2012; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
Steady-state and dynamic photosynthetic performance and nitrogen partitioning in the shade-demanding plant Panax notoginseng under different levels of growth irradiance 2 2014
Photosynthesis, light energy partitioning, and photoprotection in the shade-demanding species Panax notoginseng under high and low level of growth irradiance 1 2016
Photosynthetic acclimation to light in woody and herbaceous species: a comparison of leaf structure, pigment content and chlorophyll fluorescence characteristics measured in the field 2 2012
... 自然环境中生长的植物, 其PSI通常对高光环境不敏感(Barth et al., 2001 ).一般只有在低温弱光、常温频闪光或是饱和脉冲处理植物时, PSI才会发生光抑制(Zhang & Scheller, 2004; 梁英等, 2018; Yamamoto & Shikanai, 2019).而滇南九节(Psychotria henryi)和镰叶肾蕨(Nephrolepis falciformis)等喜阴植物叶片的PSI却在恒定高光处理下发生光抑制(Huang et al., 2017 , 2018a).PSI发生光抑制的显著特征是PSI活性下降(Huang et al., 2018b ; 梁英等, 2018).Huang等(2018b)研究发现, 三七在高光下处理30和60 min时, Pm不受影响, 说明三七的PSI对高光胁迫不敏感.但本研究中, 短时环境强光导致三七的Pm显著下降, 该结果与上述的结果不一致, 可能是因为本研究采用的是全日照下的全天高光连续处理3天, 而Huang等(2018b)研究的高光处理时间较短(处理时间仅为30和60 min)从而尚未影响到PSI的活性.另有研究表明, Y(NA)增加也可作为PSI发生光抑制的指标(Kim et al., 2005 ; Yamori et al., 2016 ; Yang et al., 2019a ).本研究中, 正午强光下三七具有较高的Y(NA)(图10A), 该结果与拟南芥(Arabidopsis thaliana)和白及(Bletilla striata)从低到高光转变后Y(NA)升高的结果(Huang et al., 2019 ; Yang et al., 2019a )一致, 由此推测, 环境光强增加后, 因无法形成足够的pH, 类囊体管腔酸化水平低, 从PSII到PSI的电子流引起受体侧羟基自由基的积累, 最终导致PSI发生光抑制(黄伟, 2012; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
Photosystem I acceptor side limitation is a prerequisite for the reversible decrease in the maximum extent of P700 oxidation after short-term chilling in the light in four plant species with different chilling sensitivities 1 2005
... 自然环境中生长的植物, 其PSI通常对高光环境不敏感(Barth et al., 2001 ).一般只有在低温弱光、常温频闪光或是饱和脉冲处理植物时, PSI才会发生光抑制(Zhang & Scheller, 2004; 梁英等, 2018; Yamamoto & Shikanai, 2019).而滇南九节(Psychotria henryi)和镰叶肾蕨(Nephrolepis falciformis)等喜阴植物叶片的PSI却在恒定高光处理下发生光抑制(Huang et al., 2017 , 2018a).PSI发生光抑制的显著特征是PSI活性下降(Huang et al., 2018b ; 梁英等, 2018).Huang等(2018b)研究发现, 三七在高光下处理30和60 min时, Pm不受影响, 说明三七的PSI对高光胁迫不敏感.但本研究中, 短时环境强光导致三七的Pm显著下降, 该结果与上述的结果不一致, 可能是因为本研究采用的是全日照下的全天高光连续处理3天, 而Huang等(2018b)研究的高光处理时间较短(处理时间仅为30和60 min)从而尚未影响到PSI的活性.另有研究表明, Y(NA)增加也可作为PSI发生光抑制的指标(Kim et al., 2005 ; Yamori et al., 2016 ; Yang et al., 2019a ).本研究中, 正午强光下三七具有较高的Y(NA)(图10A), 该结果与拟南芥(Arabidopsis thaliana)和白及(Bletilla striata)从低到高光转变后Y(NA)升高的结果(Huang et al., 2019 ; Yang et al., 2019a )一致, 由此推测, 环境光强增加后, 因无法形成足够的pH, 类囊体管腔酸化水平低, 从PSII到PSI的电子流引起受体侧羟基自由基的积累, 最终导致PSI发生光抑制(黄伟, 2012; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
Relative importance of photosynthetic traits and allocation patterns as correlates of seedling shade tolerance of 13 tropical trees 1 1994
CO2 response of cyclic electron flow around PSI (CEF-PSI) in tobacco leaves—Relative electron fluxes through PSI and PSII determine the magnitude of non-photochemical quenching (NPQ) of chl fluorescence 1 2005
Overexpressed superoxide dismutase and catalase act synergistically to protect the repair of PSII during photoinhibition in Synechococcus elongatus PCC 7942 1 2016
Regulation of antenna structure and electron transport in Photosystem II of Pisum sativum under elevated temperature probed by the fast polyphasic chlorophyll a fluorescence transient: OKJIP 1 1997
Stimulation of cyclic electron flow around photosystem I upon a sudden transition from low to high light in two angiosperms Arabidopsis thaliana and Bletilla striata 4 2019a
... 自然环境中生长的植物, 其PSI通常对高光环境不敏感(Barth et al., 2001 ).一般只有在低温弱光、常温频闪光或是饱和脉冲处理植物时, PSI才会发生光抑制(Zhang & Scheller, 2004; 梁英等, 2018; Yamamoto & Shikanai, 2019).而滇南九节(Psychotria henryi)和镰叶肾蕨(Nephrolepis falciformis)等喜阴植物叶片的PSI却在恒定高光处理下发生光抑制(Huang et al., 2017 , 2018a).PSI发生光抑制的显著特征是PSI活性下降(Huang et al., 2018b ; 梁英等, 2018).Huang等(2018b)研究发现, 三七在高光下处理30和60 min时, Pm不受影响, 说明三七的PSI对高光胁迫不敏感.但本研究中, 短时环境强光导致三七的Pm显著下降, 该结果与上述的结果不一致, 可能是因为本研究采用的是全日照下的全天高光连续处理3天, 而Huang等(2018b)研究的高光处理时间较短(处理时间仅为30和60 min)从而尚未影响到PSI的活性.另有研究表明, Y(NA)增加也可作为PSI发生光抑制的指标(Kim et al., 2005 ; Yamori et al., 2016 ; Yang et al., 2019a ).本研究中, 正午强光下三七具有较高的Y(NA)(图10A), 该结果与拟南芥(Arabidopsis thaliana)和白及(Bletilla striata)从低到高光转变后Y(NA)升高的结果(Huang et al., 2019 ; Yang et al., 2019a )一致, 由此推测, 环境光强增加后, 因无法形成足够的pH, 类囊体管腔酸化水平低, 从PSII到PSI的电子流引起受体侧羟基自由基的积累, 最终导致PSI发生光抑制(黄伟, 2012; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
... ; Yang et al., 2019a )一致, 由此推测, 环境光强增加后, 因无法形成足够的pH, 类囊体管腔酸化水平低, 从PSII到PSI的电子流引起受体侧羟基自由基的积累, 最终导致PSI发生光抑制(黄伟, 2012; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
... ; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
... ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
Photosynthetic regulation under fluctuating light in field-grownCerasus cerasoides: a comparison of young and mature leaves 1 2019b
... 自然环境中生长的植物, 其PSI通常对高光环境不敏感(Barth et al., 2001 ).一般只有在低温弱光、常温频闪光或是饱和脉冲处理植物时, PSI才会发生光抑制(Zhang & Scheller, 2004; 梁英等, 2018; Yamamoto & Shikanai, 2019).而滇南九节(Psychotria henryi)和镰叶肾蕨(Nephrolepis falciformis)等喜阴植物叶片的PSI却在恒定高光处理下发生光抑制(Huang et al., 2017 , 2018a).PSI发生光抑制的显著特征是PSI活性下降(Huang et al., 2018b ; 梁英等, 2018).Huang等(2018b)研究发现, 三七在高光下处理30和60 min时, Pm不受影响, 说明三七的PSI对高光胁迫不敏感.但本研究中, 短时环境强光导致三七的Pm显著下降, 该结果与上述的结果不一致, 可能是因为本研究采用的是全日照下的全天高光连续处理3天, 而Huang等(2018b)研究的高光处理时间较短(处理时间仅为30和60 min)从而尚未影响到PSI的活性.另有研究表明, Y(NA)增加也可作为PSI发生光抑制的指标(Kim et al., 2005 ; Yamori et al., 2016 ; Yang et al., 2019a ).本研究中, 正午强光下三七具有较高的Y(NA)(图10A), 该结果与拟南芥(Arabidopsis thaliana)和白及(Bletilla striata)从低到高光转变后Y(NA)升高的结果(Huang et al., 2019 ; Yang et al., 2019a )一致, 由此推测, 环境光强增加后, 因无法形成足够的pH, 类囊体管腔酸化水平低, 从PSII到PSI的电子流引起受体侧羟基自由基的积累, 最终导致PSI发生光抑制(黄伟, 2012; Yang et al., 2019a ).这说明, 短时环境光强的骤增导致典型阴生植物三七的PSI发生光抑制.与PSII不同, PSI发生光抑制通常需要一周或更长的时间才能得以恢复, 若是造成光损伤则会直接导致植株的死亡(Zivcaket al., 2015 ; Yang et al., 2019a ).因此, 植物必须具有可行的机制来保护PSI在波动光下不受损.围绕PSI的循环电子流对PSI起到重要的光保护作用, 这在从弱光转到强光环境下的九节、三七和高盆樱桃(Cerasus cerasoides)中均得到了证实(Huang et al., 2015b , 2018b; Yang et al., 2019b ).本研究中, 短期环境光照骤增导致围绕PSI的循环电子流被大量激活(图10, 图11), 不仅接收了来自PSI的电子, 还能使P700保持高氧化态以保护PSI不受损(黄伟等, 2012; Huang et al., 2018c ).但围绕PSI的循环电子流的激发也不能完全保护强光下的PSI免受光抑制. ...
Photosynthetic characteristics and light energy conversions under different light environments in five tree species occupying dominant status at different stages of subtropical forest succession 1 2015
 ,1,2, 双升普1,2, 张金燕1,2, 寸竹1,2, 孟珍贵1,2, 李龙根1,2, 沙本才1, 陈军文
,1,2, 双升普1,2, 张金燕1,2, 寸竹1,2, 孟珍贵1,2, 李龙根1,2, 沙本才1, 陈军文 ,1,2,*
,1,2,*

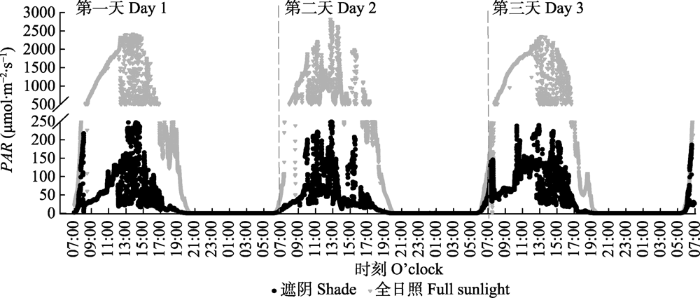 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT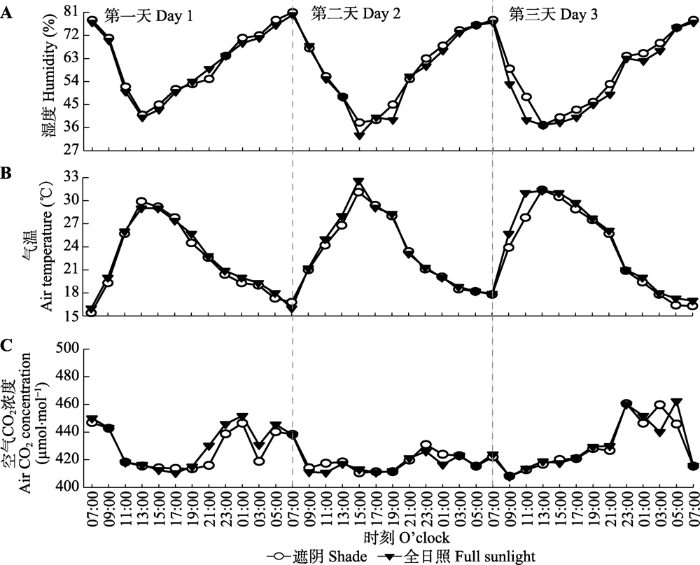 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT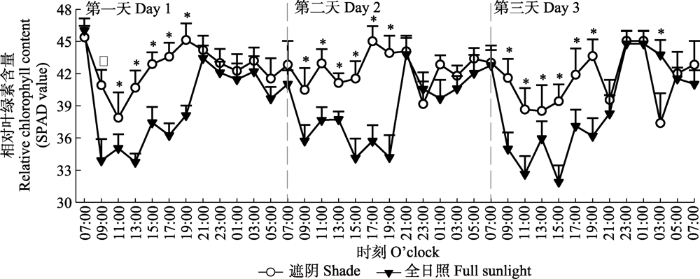 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT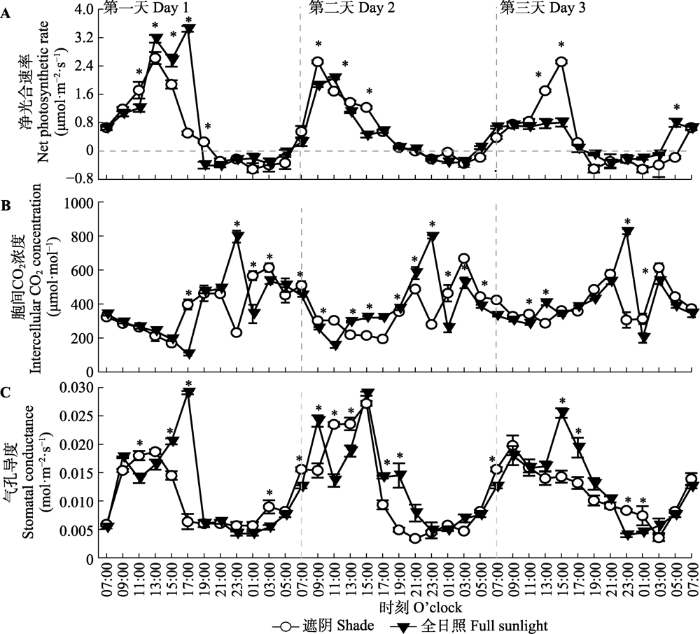 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT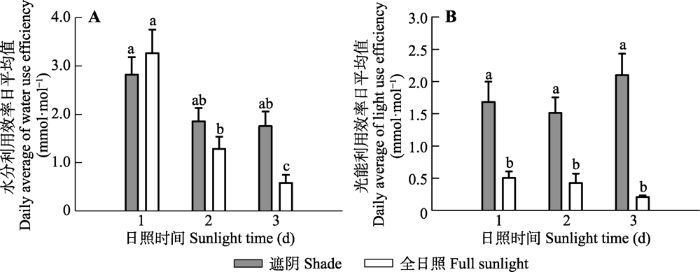 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT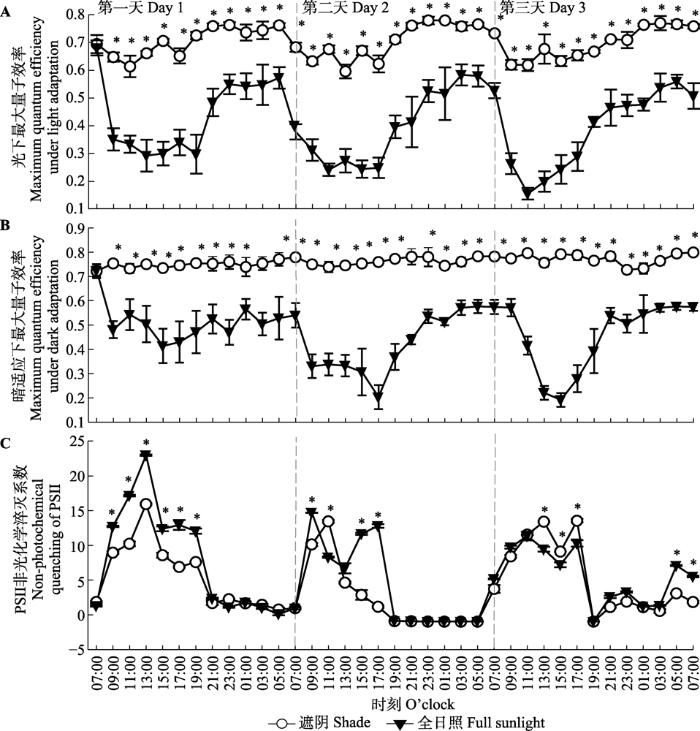 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT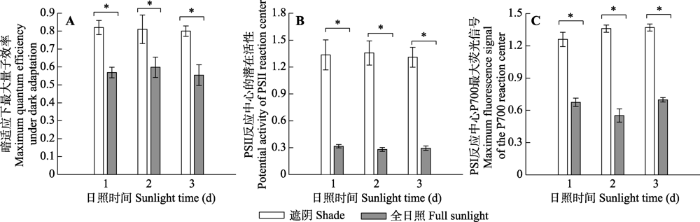 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT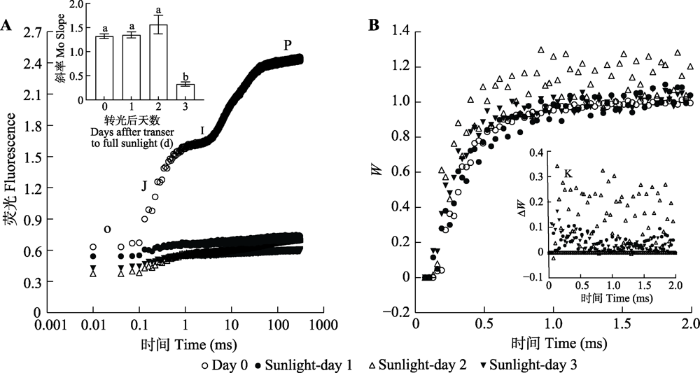 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT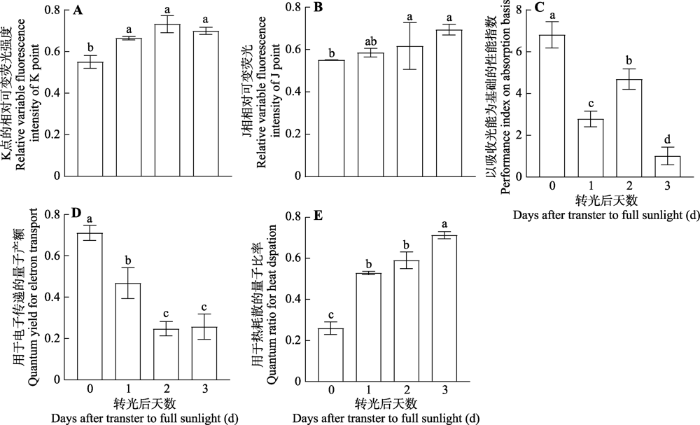 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT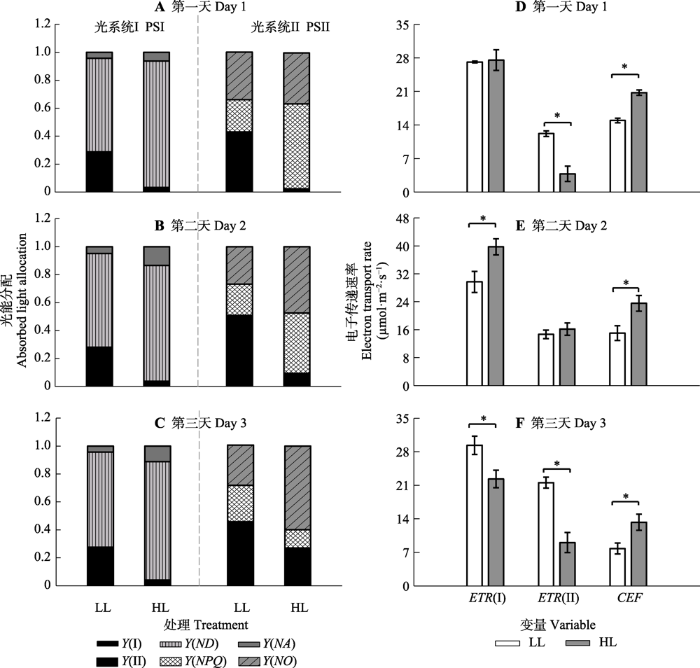 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT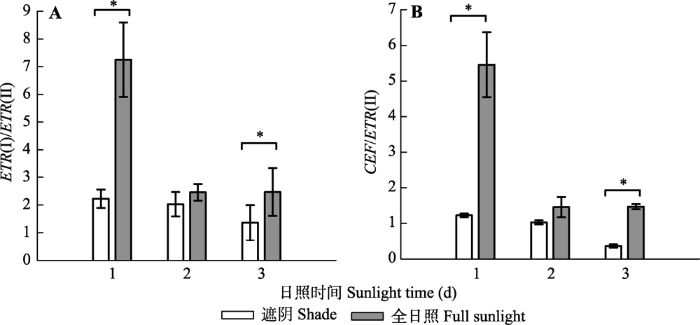 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}