Soil enzyme activities and their influencing factors among different alpine grasslands on the Qingzang Plateau
Zi-Wei WANG1,2,3, Song-Ze WAN3, Hong-Mao JIANG1,2, Yang HU1,2, Shu-Qin MA4, You-Chao CHEN5, Xu-Yang LU,1,*1Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China 2University of Chinese Academy of Science, Beijing 100049, China 3School of Forestry, Jiangxi Agricultural University, Nanchang 330045, China 4College of Tourism, Henan Normal University, Xinxiang, Henan 453007, China 5Wuhan Botanical Garden, Chinese Academy of Science, Wuhan 430074, China
Abstract Aims As a key factor of nutrient cycling in ecosystems, soil enzyme activity is an important indicator of soil quality and ecosystem function. However, there have been very few studies on the differences of soil enzyme activities among different types of alpine grassland ecosystems. Thus, the aims of this study were to compare the differences of soil enzyme activities among five different types of alpine grassland and to reveal their influencing environmental factors on the Qingzang Plateau. Methods Totally, 21 samples of five alpine grassland types, including alpine meadow, alpine steppe, alpine meadow steppe, alpine desert steppe and alpine desert on northern Qingzang Plateau, were selected for field in-situ investigation and sampling. The activities of 14 enzymes involved in the cycling of carbon (C), nitrogen (N) and phosphorus (P) were determined, and the relationships between enzymatic activities and environmental factors in alpine grassland were established. Important findings The activities of C-acquisition (invertase, cellulase, β-1,4-glucosidse, polyphenol oxidase and peroxidase), P-acquisition (alkaline phosphatase) enzymes and two N-acquisition (arylamidase and nitrite reductase) were significantly different among different alpine grassland types. Moreover, correlations were found among C-acquisition, N-acquisition and P-acquisition enzymes. A significant positive correlation was found between invertase and alkaline phosphatase, and between cellulase and N-acetyl-α-D-glucosaminidase. A significant negative correlation was found between polyphenol oxidase and nitrite reductase, N-acetyl-β-D- glucosaminidase. Soil organic matter (SOM) content, gram-negative bacteria content, ratio of nitrogen to phosphorus, gram-positive bacteria content, bacteria content, actinomycetes content, total nitrogen content and fungi content were the key factors influencing soil enzyme activity among the 19 environmental indicators, and SOM content had the greatest impact (explained 11.9%). The results demonstrated that the activities of C-acquisition, P-acquisition and two N-acquisition (arylamidase and nitrite reductase) enzymes were significantly different among different types of alpine grassland, and soil enzyme activities were mainly controlled by SOM content, microbes and N elements in alpine grassland ecosystems. Keywords:alpine meadow;alpine steppe;soil enzyme activity;soil microbes;environmental factor
PDF (2481KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 汪子微, 万松泽, 蒋洪毛, 胡扬, 马书琴, 陈有超, 鲁旭阳. 青藏高原不同高寒草地类型土壤酶活性及其影响因子. 植物生态学报, 2021, 45(5): 528-538. DOI: 10.17521/cjpe.2020.0139 WANG Zi-Wei, WAN Song-Ze, JIANG Hong-Mao, HU Yang, MA Shu-Qin, CHEN You-Chao, LU Xu-Yang. Soil enzyme activities and their influencing factors among different alpine grasslands on the Qingzang Plateau. Chinese Journal of Plant Ecology, 2021, 45(5): 528-538. DOI: 10.17521/cjpe.2020.0139
新窗口打开|下载原图ZIP|生成PPT 图2青藏高原高寒草地不同酶活性之间的相关分析。酶简写同表1。*, p < 0.05; **, p < 0.01。
Fig. 2Correlation analysis of different enzyme activities in alpine grasslands on the Qingzang Plateau. Abbreviations for enzymes are shown in Table 1. *, p < 0.05; **, p < 0.01.
Fig. 3Redundancy analysis (RDA) of the effects of environmental factors on soil enzyme activity in alpine grasslands on the Qingzang Plateau. AD1-AD3 represent three samples of alpine desert; ADS1-ADS5 represent five samples of alpine desert steppe; AM1- AM5 represent five samples of alpine meadow; AMS1-AMS3 represent three samples of alpine meadow steppe; AS1-AS5 represent five samples of alpine steppe. Act, actinomycetes; APB, above-ground plant biomass; APOM, above-ground plant organic matter; APTN, above-ground plant total nitrogen; C:N, C:P, N:P represent the ratio between carbon、nitrogen and phosphorus; G+, gram-positive bacteria; G-, gram-negative bacteria; ρb, bulk density; SOM, soil organic matter; TN, total nitrogen; TP, total phosphorus; UPB, below-ground plant biomass; UPOM, below-ground plant organic matter; UPTN, below-ground plant total nitrogen. Abbreviations for enzymes are shown in Table 1.
Table 2 表2 表2青藏高原高寒草地环境因子与酶活性之间的相关性 Table 2Correlations among environmental factors and enzyme activities in alpine grasslands on the Qingzang Plateau
环境因子 Environmental factor
INV
CEL
βG
PPO
POD
ARY
NAG
LAP
L-ASP
NAGLU
NR
NiRs
ASP
ALP
ρb
-0.482*
-0.346
0.272
0.211
0.193
0.013
-0.365
0.103
0.047
-0.137
-0.026
-0.020
0.086
-0.168
pH
-0.452*
0.234
0.169
0.049
0.238
0.074
-0.146
-0.033
-0.256
-0.155
-0.315
-0.016
-0.222
-0.245
SOM
0.601**
0.562**
-0.206
0.057
0.073
-0.441*
0.137
0.004
0.206
0.284
0.450*
-0.424
-0.054
0.437*
TN
0.552**
0.538*
-0.250
0.016
0.062
-0.337
0.156
-0.073
0.153
0.148
0.336
-0.244
-0.142
0.542*
TP
-0.016
-0.165
-0.057
0.048
-0.162
0.391
-0.174
0.101
-0.208
-0.214
-0.348
0.348
-0.103
-0.178
Bacteria
0.576**
0.582**
-0.177
0.033
0.145
-0.386
0.111
-0.064
0.218
0.249
0.366
-0.280
-0.230
0.364
Fungi
0.543*
0.510*
-0.152
0.056
0.214
-0.393
0.173
-0.057
0.327
0.135
0.320
-0.238
-0.229
0.339
Act
0.584**
0.545*
-0.136
0.039
0.024
-0.291
0.219
-0.117
0.175
0.312
0.332
-0.318
-0.288
0.386
G+
0.574**
0.571**
-0.187
0.030
0.174
-0.396
0.098
-0.069
0.200
0.246
0.353
-0.281
-0.272
0.357
G-
0.588**
0.595**
-0.186
0.037
0.155
-0.403
0.088
-0.051
0.203
0.270
0.382
-0.310
-0.229
0.374
APB
0.351
0.513*
0.008
-0.103
0.117
-0.283
0.164
-0.020
-0.088
0.068
0.097
-0.173
-0.257
0.102
UPB
0.470*
0.557**
-0.201
0.110
0.010
-0.426
-0.152
0.022
0.032
0.323
0.290
-0.335
-0.390
0.163
APOM
0.030
0.030
-0.176
0.291
0.019
-0.013
-0.421
-0.386
0.250
0.141
0.233
0.221
0.216
0.027
UPOM
0.198
0.061
-0.066
-0.050
-0.143
0.168
0.069
-0.411
-0.074
0.018
-0.109
0.318
0.060
-0.061
APTN
0.002
0.153
-0.007
-0.123
-0.213
-0.295
0.185
-0.220
-0.248
0.319
-0.391
-0.045
-0.191
-0.138
UPTN
-0.200
-0.323
-0.017
-0.101
-0.169
-0.216
0.062
-0.034
-0.034
-0.029
-0.356
0.062
-0.012
-0.288
C:N
0.070
0.075
-0.025
0.133
0.041
-0.161
-0.004
0.366
0.184
0.122
0.275
-0.277
0.014
-0.271
C:P
0.447*
0.324
-0.080
0.068
0.142
-0.462*
0.093
-0.015
0.037
0.193
0.334
-0.507*
-0.071
0.364
N:P
0.500*
0.484*
-0.081
0.012
0.155
-0.409
0.074
-0.203
-0.010
0.373
0.315
-0.408
-0.143
0.481*
环境因子与酶简写同图3和表1。*, p < 0.05; **, p < 0.01。 Abbreviations for environmental factors and enzymes are shown in Fig. 3 and Table 1. *, p < 0.05; **, p < 0.01.
AdamczykB,KilpeläinenP,KitunenV,SmolanderA(2014).Potential activities of enzymes involved in N, C, P and S cycling in boreal forest soil under different tree species .Pedobiologia,57, 97-102. DOI:10.1016/j.pedobi.2013.12.003URL [本文引用: 1]
BachHJ,MunchJC(2000).Identification of bacterial sources of soil peptidases .Biology and Fertility of Soils,31, 219-224. DOI:10.1007/s003740050648URL [本文引用: 1]
BaoSD(2000).Agrochemical Analysis of Soil.Agricultural Press,Beijing. 30-83. [本文引用: 1]
BurnsRG,DeforestJL,MarxsenJ,SinsabaughRL,StrombergerME,WallensteinMD,WeintraubMN,ZoppiniA(2013).Soil enzymes in a changing environment: current knowledge and future directions .Soil Biology & Biochemistry,58, 216-234. DOI:10.1016/j.soilbio.2012.11.009URL [本文引用: 1]
DjukicI,ZehetnerF,MentlerA,GerzabekMH(2010).Microbial community composition and activity in different Alpine vegetation zones .Soil Biology & Biochemistry,42, 155-161. DOI:10.1016/j.soilbio.2009.10.006URL [本文引用: 1]
DuanMJ,GaoQZ,GuoYQ,WanYF,LiYE,Ganzhuzhabu,Danjiuluobu,WeiLT,XiraozhuomaLT(2011).Species diversity distribution pattern of alpine grassland communities along an altitudinal gradient in the Northern Tibet .Pratacultural Science,28, 1845-1850. [本文引用: 1]
FuG,ShenZX(2017).Response of alpine soils to nitrogen addition on the Tibetan Plateau: a meta-analysis .Applied Soil Ecology,114, 99-104. DOI:10.1016/j.apsoil.2017.03.008URL [本文引用: 1]
GuanSY(1986).Journal of Natural Resources,Agriculture Press,Beijing. 294-297. [本文引用: 1]
HeQQ,WuYH,BingHJ,ZhouJ,WangJP(2020).Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau .Geoderma,374, 114424. DOI:10.1016/j.geoderma.2020.114424. DOI:10.1016/j.geoderma.2020.114424URL [本文引用: 1]
HenryHAL(2013).Reprint of “Soil extracellular enzyme dynamics in a changing climate” .Soil Biology & Biochemistry,56, 53-59. DOI:10.1016/j.soilbio.2012.10.022URL [本文引用: 1]
HernándezDL,HobbieSE(2010).The effects of substrate composition, quantity, and diversity on microbial activity .Plant and Soil,335, 397-411. DOI:10.1007/s11104-010-0428-9URL [本文引用: 1]
HongJT,WangXD,WuJB(2014).Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the Northern Tibetan Plateau .PLOS ONE,9, e109052. DOI:10.1371/journal.pone.0109052. DOI:10.1371/journal.pone.0109052URL [本文引用: 1]
JianSY,LiJW,ChenJ,WangGS,MayesMA,DzantorKE,HuiDF,LuoYQ(2016).Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: a meta-analysis .Soil Biology & Biochemistry,101, 32-43. DOI:10.1016/j.soilbio.2016.07.003URL [本文引用: 2]
JingX,ChenX,XiaoW,LinL,WangC,HeJS,ZhuB(2018).Soil enzymatic responses to multiple environmental drivers in the Tibetan grasslands: insights from two manipulative field experiments and a meta-analysis .Pedobiologia,71, 50-58. DOI:10.1016/j.pedobi.2018.10.001URL [本文引用: 2]
JingX,SandersNJ,ShiY,ChuHY,ClassenAT,ZhaoK,ChenLT,ShiY,JiangYX,HeJS(2015).The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate .Nature Communications,6, 8159. DOI:10.1038/ncomms9159. DOI:10.1038/ncomms9159PMID:26328906 [本文引用: 1] Jing, Xin; Zhao, Ke; Shi, Yue; He, Jin-Sheng Peking Univ, Dept Ecol, Coll Urban & Environm Sci, Beijing 100871, Peoples R China. Jing, Xin; Zhao, Ke; Shi, Yue; He, Jin-Sheng Peking Univ, Key Lab Earth Surface Proc, Minist Educ, Beijing 100871, Peoples R China. Sanders, Nathan J. Univ Copenhagen, Ctr Macroecol Evolut & Climate, Nat Hist Museum Denmark, DK-2100 Copenhagen, Denmark. Shi, Yu; Chu, Haiyan Chinese Acad Sci, Inst Soil Sci, State Key Lab Soil & Sustainable Agr, Nanjing 210008, Peoples R China. Classen, Aimee T. Univ Copenhagen, Nat Hist Museum Denmark, DK-2100 Copenhagen, Denmark. Chen, Litong; He, Jin-Sheng Chinese Acad Sci, Northwest Inst Plateau Biol, Key Lab Adaptat & Evolut Plateau Biota, Xining 810008, Peoples R China. Shi, Yue Chinese Acad Sci, Inst Bot, Beijing 100093, Peoples R China. Jiang, Youxu Chinese Acad Forestry, Inst Forest Ecol, Beijing 100091, Peoples R China.
KatsuyamaC,KondoN,SuwaY,YamagishiT,ItohM,OhteN,KimuraH,NagaosaK,KatoK(2008).Denitrification activity and relevant bacteria revealed by nitrite reductase gene fragments in soil of temperate mixed forest .Microbes and Environments,23, 337-345. PMID:21558728 [本文引用: 1] Denitrification activity and bacterial community constituents were investigated in both well-drained and poorly drained soils of a temperate forest in central Japan by (15)N tracer experiments and a cloning-sequencing approach. Denitrification activity was much higher in wet soil than in dry soil, based on (15)N(15)N ((30)N(2)) and (15)N(15)NO ((46)N(2)O) production. Labeled nitrate ((15)NO(3)(-)) was immediately reduced to (30)N(2) in wet soil, whereas it was only reduced to (46)N(2)O in dry soil. Thus, the wet soil at the lower end of the catchment is a functional site for the scavenging for NO(3)(-) and N(2)O. Nitrite reductase gene (nirK and nirS) fragments from these soils were PCR amplified, cloned, and sequenced. Both nirK and nirS fragments were detected in the wet soil, whereas only nirK fragments were detected in the dry soil. All the nirK and nirS clones showed less than 90% similarity to known clones. Numerous operational taxonomic units for nirK and nirS were found in the wet soil. Considerable diversification within the largest clade on the nirK phylogenetic tree, which contained no known sequence, was observed in wet soil. Thus, a wet soil environment can provide both the habitat and conditions for the expression of denitrification activity.
KeelerBL,HobbieSE,KelloggLE(2009).Effects of long- term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition .Ecosystems,12, 1-15. DOI:10.1007/s10021-008-9199-zURL [本文引用: 1]
KivlinSN,TresederKK(2014).Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition .Biogeochemistry,117, 23-37. DOI:10.1007/s10533-013-9852-2URL [本文引用: 2]
KuypersMMM,MarchantHK,KartalB(2018).The microbial nitrogen-cycling network .Nature Reviews Microbiology,16, 263-276. DOI:10.1038/nrmicro.2018.9PMID:29398704 [本文引用: 1] Nitrogen is an essential component of all living organisms and the main nutrient limiting life on our planet. By far, the largest inventory of freely accessible nitrogen is atmospheric dinitrogen, but most organisms rely on more bioavailable forms of nitrogen, such as ammonium and nitrate, for growth. The availability of these substrates depends on diverse nitrogen-transforming reactions that are carried out by complex networks of metabolically versatile microorganisms. In this Review, we summarize our current understanding of the microbial nitrogen-cycling network, including novel processes, their underlying biochemical pathways, the involved microorganisms, their environmental importance and industrial applications.
LiGL,KimS,HanSH,ChangHN,DuDL,SonYW(2018).Precipitation affects soil microbial and extracellular enzymatic responses to warming .Soil Biology & Biochemistry,120, 212-221. DOI:10.1016/j.soilbio.2018.02.014URL [本文引用: 1]
LiY,NieC,LiuYH,DuW,HeP(2019).Soil microbial community composition closely associates with specific enzyme activities and soil carbon chemistry in a long-term nitrogen fertilized grassland .Science of the Total Environment,654, 264-274. DOI:10.1016/j.scitotenv.2018.11.031URL [本文引用: 1]
LinB,ZhaoXR,ZhengY,QiS,LiuXZ(2017).Effect of grazing intensity on protozoan community, microbial biomass, and enzyme activity in an alpine meadow on the Tibetan Plateau .Journal of Soils and Sediments,17, 2752-2762. DOI:10.1007/s11368-017-1695-3URL [本文引用: 1]
LiuSB,RazaviBS,SuX,MaharjanM,ZarebanadkoukiM,BlagodatskayaE,KuzyakovY(2017).Spatio-temporal patterns of enzyme activities after manure application reflect mechanisms of niche differentiation between plants and microorganisms .Soil Biology & Biochemistry,112, 100-109. DOI:10.1016/j.soilbio.2017.05.006URL [本文引用: 1]
LiuXC,ZhangST(2019).Nitrogen addition shapes soil enzyme activity patterns by changing pH rather than the composition of the plant and microbial communities in an alpine meadow soil .Plant and Soil,440, 11-24. DOI:10.1007/s11104-019-04054-5URL [本文引用: 1]
LiuXJ,ZhangY,HanWX,TangAH,ShenJL,CuiZL,VitousekP,ErismanJW,GouldingK,ChristieP,FangmeierA,ZhangFS(2013).Enhanced nitrogen deposition over China .Nature,494, 459-462. DOI:10.1038/nature11917URL [本文引用: 1]
Lopez-AizpunM,Arango-MoraC,SantamariaC,LasherasE,SantamariaJM,CigandaVS,CardenasLM,ElustondoD(2018).Atmospheric ammonia concentration modulates soil enzyme and microbial activity in an oak forest affecting soil microbial biomass .Soil Biology & Biochemistry,116, 378-387. DOI:10.1016/j.soilbio.2017.10.020URL [本文引用: 1]
LuXY,YanY,SunJ,ZhangXK,ChenYC,WangXD,ChengGW(2015).Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: effects of grazing exclusion .Ecology and Evolution,5, 4492-4504. DOI:10.1002/ece3.2015.5.issue-19URL [本文引用: 1]
MaWJ,LiJ,GaoY,XingF,SunSN,ZhangT,ZhuXZ,ChenC,LiZ(2020).Responses of soil extracellular enzyme activities and microbial community properties to interaction between nitrogen addition and increased precipitation in a semi-arid grassland ecosystem .Science of the Total Environment,703, 134691. DOI:10.1016/S1002-0160(18) 60010-4. DOI:10.1016/S1002-0160(18) 60010-4URL [本文引用: 1]
ManzoniS,JacksonRB,TrofymowJA,PorporatoA(2008).The global stoichiometry of litter nitrogen mineralization .Science,321, 684-686. DOI:10.1126/science.1159792URL [本文引用: 1]
MazzonM,CavaniL,MargonA,SorrentiG,CiavattaC,MarzadoriC(2018).Changes in soil phenol oxidase activities due to long-term application of compost and mineral N in a walnut orchard .Geoderma,316, 70-77. DOI:10.1016/j.geoderma.2017.12.009URL [本文引用: 1]
MarxMC,WoodM,JarvisSC(2001).A microplate fluorimetric assay for the study of enzyme diversity in soils .Soil Biology & Biochemistry,33, 1633-1640. DOI:10.1016/S0038-0717(01)00079-7URL [本文引用: 1]
MoZH,LiYE,GaoQZ(2012).Simulation on productivity of main grassland ecosystems responding to climate change .Chinese Journal of Agrometeorology,33, 545-554. [本文引用: 1]
NannipieriP,Trasar-CepedaC,DickRP(2018).Soil enzyme activity: a brief history and biochemistry as a basis for appropriate interpretations and meta-analysis .Biology and Fertility of Soils,54, 11-19. DOI:10.1007/s00374-017-1245-6URL [本文引用: 1]
NiuL,LiuYH,LiY,OuyangSN(2015).Microbial community structure of the alpine meadow under different grazing styles in Naqu prefecture of Tibet .Chinese Journal of Applied Ecology,26, 2298-2306. [本文引用: 1]
QingQ,YangB,WymanCE(2010).Xylooligomers are strong inhibitors of cellulose hydrolysis by enzymes .Bioresource Technology,101, 9624-9630. DOI:10.1016/j.biortech.2010.06.137PMID:20708404 [本文引用: 1] Typically, the enzymatic hydrolysis rate of lignocellulosic biomass is fast initially but then slows down more rapidly than can be explained by just consumption of substrate. Although several factors including enzyme inhibition, enzyme deactivation, a drop in substrate reactivity, or nonproductive binding of enzyme to lignin could be responsible for this loss of effectiveness, we recently reported evidence that xylose, xylan, and xylooligomers dramatically decrease conversion rates and yields, but clarification was still needed for the magnitude of their effect. Therefore, in this study, xylan and various xylooligomers were added to Avicel hydrolysis at low enzyme loadings and found to have a greater effect than adding equal amounts of xylose derived from these materials or when added separately. Furthermore, xylooligomers were more inhibitory than xylan or xylose in terms of a decreased initial hydrolysis rate and a lower final glucose yield even for a low concentration of 1.67 mg/ml. At a higher concentration of 12.5mg/ml, xylooligomers lowered initial hydrolysis rates of Avicel by 82% and the final hydrolysis yield by 38%. Mixed DP xylooligomers showed strong inhibition on cellulase enzymes but not on beta-glucosidase enzymes. By tracking the profile change of xylooligomers, a large portion of the xylooligomers was found to be hydrolyzed by Spezyme CP enzyme preparations, indicating competitive inhibition by mixed xylooligomers. A comparison among glucose sugars and xylose sugars also showed that xylooligomers were more powerful inhibitors than well-established glucose and cellobiose.Copyright (c) 2010 Elsevier Ltd. All rights reserved.
RuiYC,WangYF,ChenCR,ZhouXQ,WangSP,XuZH,DuanJC,KangXM,LuSB,LuoCY(2012).Warming and grazing increase mineralization of organic P in an alpine meadow ecosystem of Qinghai-Tibet Plateau, China .Plant and Soil,357, 73-87. DOI:10.1007/s11104-012-1132-8URL [本文引用: 1]
SardansJ,PeñuelasJ,EstiarteM(2008).Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland .Applied Soil Ecology,39, 223-235. DOI:10.1016/j.apsoil.2007.12.011URL [本文引用: 2]
SinsabaughRL(2010).Phenol oxidase, peroxidase and organic matter dynamics of soil .Soil Biology & Biochemistry,42, 391-404. DOI:10.1016/j.soilbio.2009.10.014URL [本文引用: 2]
SinsabaughRL,AntibusRK,LinkinsAE,McclaughertyC,RayburnL,RepertD,WeilandT(1992).Wood decomposition over a first-order watershed: mass loss as a function of lignocellulase activity .Soil Biology & Biochemistry,24, 743-749. DOI:10.1016/0038-0717(92)90248-VURL [本文引用: 1]
SinsabaughRL,LauberCL,WeintraubMN,AhmedB,AllisonSD,CrenshawC,ContostaAR,CusackD,FreyS,GalloME,GartnerTB,HobbieSE,HollandK,KeelerBL,PowersJS,StursovaM,Takacs-VesbachC,WaldropMP,WallensteinMD,ZakDR,ZeglinLH(2008).Stoichiometry of soil enzyme activity at global scale .Ecology Letters,11, 1252-1264. DOI:10.1111/j.1461-0248.2008.01245.xPMID:18823393 [本文引用: 3] Extracellular enzymes are the proximate agents of organic matter decomposition and measures of these activities can be used as indicators of microbial nutrient demand. We conducted a global-scale meta-analysis of the seven-most widely measured soil enzyme activities, using data from 40 ecosystems. The activities of beta-1,4-glucosidase, cellobiohydrolase, beta-1,4-N-acetylglucosaminidase and phosphatase g(-1) soil increased with organic matter concentration; leucine aminopeptidase, phenol oxidase and peroxidase activities showed no relationship. All activities were significantly related to soil pH. Specific activities, i.e. activity g(-1) soil organic matter, also varied in relation to soil pH for all enzymes. Relationships with mean annual temperature (MAT) and precipitation (MAP) were generally weak. For hydrolases, ratios of specific C, N and P acquisition activities converged on 1 : 1 : 1 but across ecosystems, the ratio of C : P acquisition was inversely related to MAP and MAT while the ratio of C : N acquisition increased with MAP. Oxidative activities were more variable than hydrolytic activities and increased with soil pH. Our analyses indicate that the enzymatic potential for hydrolyzing the labile components of soil organic matter is tied to substrate availability, soil pH and the stoichiometry of microbial nutrient demand. The enzymatic potential for oxidizing the recalcitrant fractions of soil organic material, which is a proximate control on soil organic matter accumulation, is most strongly related to soil pH. These trends provide insight into the biogeochemical processes that create global patterns in ecological stoichiometry and organic matter storage.
SinsabaughRL,MoorheadDL(1994).Resource allocation to extracellular enzyme production: a model for nitrogen and phosphorus control of litter decomposition .Soil Biology & Biochemistry,26, 1305-1311. DOI:10.1016/0038-0717(94)90211-9URL [本文引用: 2]
ŠnajdrJ,CajthamlT,ValáškováV,MerhautováV,PetránkováM,SpetzP,LeppänenK,BaldrianP(2011).Transformation of Quercus petraea litter: successive changes in litter chemistry are reflected in differential enzyme activity and changes in the microbial community composition .Fems Microbiology Ecology,75, 291-303. DOI:10.1111/fem.2010.75.issue-2URL [本文引用: 1]
TorresIF,BastidaF,HernandezT,AlbaladejoJ,GarciaC(2015).Enzyme activity, microbial biomass and community structure in a long-term restored soil under semi-arid conditions .Soil Research,53, 553-560. DOI:10.1071/SR14297URL [本文引用: 1]
VeresZ,KotroczóZ,FeketeI,TóthJA,LajthaK,TownsendK,TóthmérészB(2015).Soil extracellular enzyme activities are sensitive indicators of detrital inputs and carbon availability .Applied Soil Ecology,92, 18-23. DOI:10.1016/j.apsoil.2015.03.006URL [本文引用: 1]
von SperberC,KriesH,TamburiniF,BernasconiSM,FrossardE(2014).The effect of phosphomonoesterases on the oxygen isotope composition of phosphate .Geochimica Et Cosmochimica Acta,125, 519-527. DOI:10.1016/j.gca.2013.10.010URL [本文引用: 1]
WalleniusK,RitaHN,MikkonenA,LappiK,LindstromK,HartikainenH,RaatelandA,NiemiR(2011).Effects of land use on the level, variation and spatial structure of soil enzyme activities and bacterial communities .Soil Biology & Biochemistry,43, 1464-1473. DOI:10.1016/j.soilbio.2011.03.018URL [本文引用: 1]
WallensteinMD,McMahonSK,SchimelJP(2009).Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils .Global Change Biology,15, 1631-1639. DOI:10.1111/gcb.2009.15.issue-7URL [本文引用: 1]
WangSG,HouYL(2004).Application of phospholipid fatty acid method in soil microbial analysis .Microbiology,31, 114-117. [本文引用: 1]
WangXX,DongSK,GaoQZ,ZhouHK,LiuSL,SuXK,LiYY(2014).Effects of short-term and long-term warming on soil nutrients, microbial biomass and enzyme activities in an alpine meadow on the Qinghai-Tibet Plateau of China .Soil Biology & Biochemistry,76, 140-142. DOI:10.1016/j.soilbio.2014.05.014URL [本文引用: 1]
WeintraubSR,WiederWR,ClevelandCC,TownsendAR(2013).Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest .Biogeochemistry,114, 313-326. DOI:10.1007/s10533-012-9812-2URL [本文引用: 1]
XiaoW,ChenX,JingX,ZhuB(2018).A meta-analysis of soil extracellular enzyme activities in response to global change .Soil Biology & Biochemistry,123, 21-32. DOI:10.1016/j.soilbio.2018.05.001URL [本文引用: 1]
XuG,LiuY,LongZJ,HuSL,ZhangYB,JiangH(2018).Effects of exotic plantation forests on soil edaphon and organic matter fractions .Science of the Total Environment,626, 59-68. DOI:10.1016/j.scitotenv.2018.01.088URL [本文引用: 1]
XuZF,HuR,XiongP,WanC,CaoG,LiuQ(2010).Initial soil responses to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China: nutrient availabilities, microbial properties and enzyme activities .Applied Soil Ecology,46, 291-299. DOI:10.1016/j.apsoil.2010.07.005URL [本文引用: 1]
YanZQ,QiYC,LiSJ,DongYS,PengQ,HeYL,LiZL(2017).Soil microorganisms and enzyme activity of grassland ecosystem affected by changes in precipitation pattern and increase in nitrogen deposition—A review .Microbiology,44, 1481-1490. [本文引用: 1]
YangYG,YangY,GengYQ,HuangGL,CuiXQ,HouM(2018).Effects of different land types on soil enzyme activity in the Qinghai lake region .Wetlands,38, 711-721. DOI:10.1007/s13157-018-1014-9URL [本文引用: 1]
YuCQ,ZhangYJ,ClausH,ZengR,ZhangXZ,WangJS(2012).Ecological and environmental issues faced by a developing Tibet .Environmental Science & Technology,46, 1979-1980. DOI:10.1021/es2047188URL [本文引用: 1]
ZhangC,WangJ,LiuGB,SongZL,FangLC(2019).Impact of soil leachate on microbial biomass and diversity affected by plant diversity .Plant and Soil,439, 505-523. DOI:10.1007/s11104-019-04032-x [本文引用: 1] AimsHigh plant diversity is usually linked with high soil microbial diversity, which is hypothesized to be attributed to a high diversity of components in the soil leachate, but experimental evidence is scarce. The aim of this study was to determine if the variation in soil leachate caused by plant diversity could affect the soil microbial community.MethodsA microcosm experiment was conducted to determine the effect of plant diversity on the soil microbial community by measuring soil leachate in a gradient of plant richness from levels 1 (one species) to 3 (three species).ResultsPlant richness significantly affected the diversity of soil leachate and microbial communities. The amount and diversity of soil leachate, microbial biomass carbon (C), basal respiration, beta-1,4-glucosidase activity, beta-1,4-N-acetylglucosaminidase activity, bacterial biomass, fungal biomass, total microorganism biomass, and microbial diversity (Shannon diversity index and evenness) were highest at richness level 3. Changes in the microbial community were best explained by variation in the amount and diversity of leachate. Linear regression and correlation analyses indicated that leachate diversity had a close association with microbial Shannon diversity and evenness, whereas leachate amount had a close association with microbial biomass C, total microbial biomass, bacterial biomass, enzyme activities, and abundance of microbial groups. An ordinary least squares multiple regression and the structural equation model demonstrated that leachate amount had a greater effect on microbial biomass than leachate diversity, which had a greater impact on microbial Shannon diversity and evenness.ConclusionsOur results indicate that plant diversity drives changes in soil microbial communities by altering the amount and diversity of leachate in the soil. The diversity of soil leachate determined the diversity of the microbial community to some extent.
ZhangXK,LuXY,WangXD(2014).The spatial and temporal variation of NDVI and its relationships to climatic factors in Northern Tibet over the period of 2000-2010—Take Shantsa for example .Journal of Mountain Research,32, 475-480. [本文引用: 1]
ZhangY,DongSK,GaoQZ,LiuSL,GanjurjavH,WangXX,SuXK,WuXY(2017).Soil bacterial and fungal diversity differently correlated with soil biochemistry in alpine grassland ecosystems in response to environmental changes .Scientific Reports,7, 43077. DOI:10.1038/srep43077. DOI:10.1038/srep43077PMID:28262753 [本文引用: 1] To understand effects of soil microbes on soil biochemistry in alpine grassland ecosystems under environmental changes, we explored relationships between soil microbial diversity and soil total nitrogen, organic carbon, available nitrogen and phosphorus, soil microbial biomass and soil enzyme activities in alpine meadow, alpine steppe and cultivated grassland on the Qinghai-Tibetan plateau under three-year warming, enhanced precipitation and yak overgrazing. Soil total nitrogen, organic carbon and NH-N were little affected by overgrazing, warming or enhanced precipitation in three types of alpine grasslands. Soil microbial biomass carbon and phosphorus along with the sucrase and phosphatase activities were generally stable under different treatments. Soil NO-N, available phosphorus, urease activity and microbial biomass nitrogen were increased by overgrazing in the cultivated grassland. Soil bacterial diversity was positively correlated with, while soil fungal diversity negatively with soil microbial biomass and enzyme activities. Soil bacterial diversity was negatively correlated with, while soil fungal diversity positively with soil available nutrients. Our findings indicated soil bacteria and fungi played different roles in affecting soil nutrients and microbiological activities that might provide an important implication to understand why soil biochemistry was generally stable under environmental changes in alpine grassland ecosystems.
ZhaoSC,LiKJ,ZhouW,QiuSJ,HuangSW,HeP(2016).Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China .Agriculture Ecosystems & Environment,216, 82-88. DOI:10.1016/j.agee.2015.09.028URL [本文引用: 1]
Potential activities of enzymes involved in N, C, P and S cycling in boreal forest soil under different tree species 1 2014
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
Identification of bacterial sources of soil peptidases 1 2000
... 本研究涉及的14种酶中, NAG活性较高, 一定程度上反映了N转化在高寒草地生态系统的重要性.因为NAG参与几丁质的分解, 参与了有机N的吸收(Sinsabaugh et al., 1992), 在其他β-1,4-连接的葡糖胺聚合物的降解中也发挥作用, 类似于βG在纤维素降解中的作用(Sinsabaugh et al., 2005).且由于酶是富含N的化合物, 其生产受到N可用性的严格控制(Keeler et al., 2009), 从而导致NAG的活性较高.因此, 参与N吸收的NAG间接反映了N转化过程在该区域的重要性.PPO可以降解木质素和可溶性酚类化合物(Mazzon et al., 2018), 是参与C循环过程的重要酶, NAG是N循环重要酶, 而相关分析表明NAG与PPO呈显著负相关关系, 这可能是因为PPO是分解复杂C的酶, 而NAG作为参与几丁质分解的N转化酶, 基于酶化学计量的meta分析发现, N的添加会加剧微生物量C限制, 从而造成两种酶之间的负相关(Chen et al., 2018).ALP活性直接影响有机磷在土壤中的分解和转化, 并加速有机P的脱磷率(von Sperber et al., 2014), 在AS中最高, AD中最低, 说明AS土壤中微生物养分需求受到P的限制最严重, 而AD土壤中的P限制较轻. ...
Microbial community composition and activity in different Alpine vegetation zones 1 2010
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局 1 2011
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
藏北高寒草地植物群落物种多样性沿海拔梯度的分布格局 1 2011
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
Response of alpine soils to nitrogen addition on the Tibetan Plateau: a meta-analysis 1 2017
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
Vegetation type rather than climate modulates the variation in soil enzyme activities and stoichiometry in subalpine forests in the eastern Tibetan Plateau 1 2020
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Reprint of “Soil extracellular enzyme dynamics in a changing climate” 1 2013
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
The effects of substrate composition, quantity, and diversity on microbial activity 1 2010
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the Northern Tibetan Plateau 1 2014
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: a meta-analysis 2 2016
Soil enzymatic responses to multiple environmental drivers in the Tibetan grasslands: insights from two manipulative field experiments and a meta-analysis 2 2018
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate 1 2015
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
Denitrification activity and relevant bacteria revealed by nitrite reductase gene fragments in soil of temperate mixed forest 1 2008
... 在测定的14种土壤酶活性中, C、N和P循环酶在5种高寒草地类型均存在显著差异的酶.其中参与C循环的5种酶均存在显著差异, 说明不同高寒草地类型是影响C循环酶活性的重要因子.而8种N循环酶中仅有2种酶活性在不同草地类型间存在显著差异, 这可能是由于土壤中N的累积比C的累积要慢很多(Veres et al., 2015; Kuypers et al., 2018), 为了平衡土壤中C含量高于N含量的现象, 微生物倾向于分泌更多的N 循环酶(Zhang et al., 2019), 并降低C循环酶的分泌, 且N获取策略具有多样性, N循环酶会与C的获取相耦合(Sinsabaugh et al., 2008), 同时, 分布在几类聚合物以及腐殖质分子中的N使得N的获取策略与特定类群的C底物偏好有关(Manzoni et al., 2008).因此, N循环酶不仅受环境因子的调控, 同时受C循环酶的底物特性及酶活性调控, 从而导致C循环酶和N循环酶在不同草地类型中差异不一致.另外, N转化过程中涉及N同化的两个酶NR与NiRs无显著相关关系, 可能是因为不同气候区域下土壤亚硝酸盐微生物群落分异造成的结果(Katsuyama et al., 2008). ...
Effects of long- term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition 1 2009
... 本研究涉及的14种酶中, NAG活性较高, 一定程度上反映了N转化在高寒草地生态系统的重要性.因为NAG参与几丁质的分解, 参与了有机N的吸收(Sinsabaugh et al., 1992), 在其他β-1,4-连接的葡糖胺聚合物的降解中也发挥作用, 类似于βG在纤维素降解中的作用(Sinsabaugh et al., 2005).且由于酶是富含N的化合物, 其生产受到N可用性的严格控制(Keeler et al., 2009), 从而导致NAG的活性较高.因此, 参与N吸收的NAG间接反映了N转化过程在该区域的重要性.PPO可以降解木质素和可溶性酚类化合物(Mazzon et al., 2018), 是参与C循环过程的重要酶, NAG是N循环重要酶, 而相关分析表明NAG与PPO呈显著负相关关系, 这可能是因为PPO是分解复杂C的酶, 而NAG作为参与几丁质分解的N转化酶, 基于酶化学计量的meta分析发现, N的添加会加剧微生物量C限制, 从而造成两种酶之间的负相关(Chen et al., 2018).ALP活性直接影响有机磷在土壤中的分解和转化, 并加速有机P的脱磷率(von Sperber et al., 2014), 在AS中最高, AD中最低, 说明AS土壤中微生物养分需求受到P的限制最严重, 而AD土壤中的P限制较轻. ...
Soil extracellular enzyme activities correspond with abiotic factors more than fungal community composition 2 2014
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
... ).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
The microbial nitrogen-cycling network 1 2018
... 在测定的14种土壤酶活性中, C、N和P循环酶在5种高寒草地类型均存在显著差异的酶.其中参与C循环的5种酶均存在显著差异, 说明不同高寒草地类型是影响C循环酶活性的重要因子.而8种N循环酶中仅有2种酶活性在不同草地类型间存在显著差异, 这可能是由于土壤中N的累积比C的累积要慢很多(Veres et al., 2015; Kuypers et al., 2018), 为了平衡土壤中C含量高于N含量的现象, 微生物倾向于分泌更多的N 循环酶(Zhang et al., 2019), 并降低C循环酶的分泌, 且N获取策略具有多样性, N循环酶会与C的获取相耦合(Sinsabaugh et al., 2008), 同时, 分布在几类聚合物以及腐殖质分子中的N使得N的获取策略与特定类群的C底物偏好有关(Manzoni et al., 2008).因此, N循环酶不仅受环境因子的调控, 同时受C循环酶的底物特性及酶活性调控, 从而导致C循环酶和N循环酶在不同草地类型中差异不一致.另外, N转化过程中涉及N同化的两个酶NR与NiRs无显著相关关系, 可能是因为不同气候区域下土壤亚硝酸盐微生物群落分异造成的结果(Katsuyama et al., 2008). ...
Precipitation affects soil microbial and extracellular enzymatic responses to warming 1 2018
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
Soil microbial community composition closely associates with specific enzyme activities and soil carbon chemistry in a long-term nitrogen fertilized grassland 1 2019
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Effect of grazing intensity on protozoan community, microbial biomass, and enzyme activity in an alpine meadow on the Tibetan Plateau 1 2017
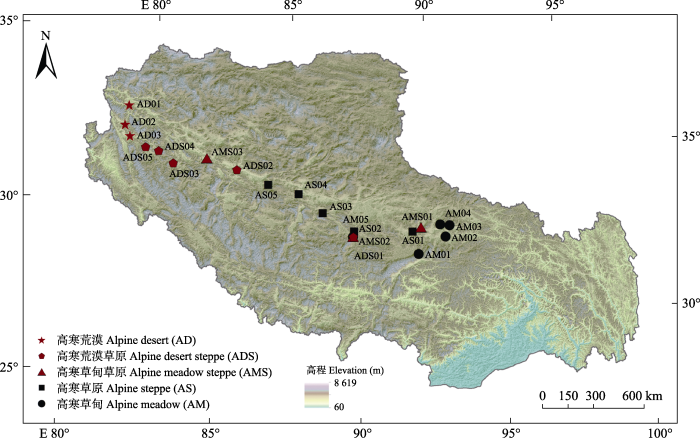
... 目前, 国内外在高寒草地土壤酶活性研究方面所开展的大量工作主要集中于其对模拟氮(N)沉降、施肥(N肥、磷(P)肥、粪肥等)(Liu et al., 2017; Liu & Zhang et al., 2019)、升温(Wang et al., 2014)、放牧强度(Lin et al., 2017)、土地利用方式(Yang et al., 2018)等的响应, 且仅在小区域或单一植被类型中进行.而高寒草地作为青藏高原主要的植被类型, 不同类型的高寒草地反映着青藏高原不同的生态环境, 有关不同高寒草地土壤酶活性的研究却严重缺少.在未来全球气候变化大背景下, 探讨不同高寒草地土壤酶活性及其对环境变化的响应机制, 对高寒草地生态系统的保护与分类管理具有重要意义, 同时对青藏高原养分循环的研究具有积极作用.本研究在藏北高原典型高寒草地分布区域选取高寒草甸(AM)、高寒草原(AS)、高寒草甸草原(AMS)、高寒荒漠草原(ADS)、高寒荒漠(AD) 5种植被类型, 测定了涉及碳(C)、N和P转化的14种土壤酶活性, 通过对青藏高原不同类型高寒草地土壤酶活性特征及其影响因子的研究, 丰富对土壤酶在高寒草地养分循环利用中作用规律的认识, 对预测青藏高原高寒草地生态系统土壤酶活性及其对全球变化的响应和机理提供科学参考, 同时为高寒草地生态系统初级生产力的维持提供理论依据. ...
Spatio-temporal patterns of enzyme activities after manure application reflect mechanisms of niche differentiation between plants and microorganisms 1 2017
... 目前, 国内外在高寒草地土壤酶活性研究方面所开展的大量工作主要集中于其对模拟氮(N)沉降、施肥(N肥、磷(P)肥、粪肥等)(Liu et al., 2017; Liu & Zhang et al., 2019)、升温(Wang et al., 2014)、放牧强度(Lin et al., 2017)、土地利用方式(Yang et al., 2018)等的响应, 且仅在小区域或单一植被类型中进行.而高寒草地作为青藏高原主要的植被类型, 不同类型的高寒草地反映着青藏高原不同的生态环境, 有关不同高寒草地土壤酶活性的研究却严重缺少.在未来全球气候变化大背景下, 探讨不同高寒草地土壤酶活性及其对环境变化的响应机制, 对高寒草地生态系统的保护与分类管理具有重要意义, 同时对青藏高原养分循环的研究具有积极作用.本研究在藏北高原典型高寒草地分布区域选取高寒草甸(AM)、高寒草原(AS)、高寒草甸草原(AMS)、高寒荒漠草原(ADS)、高寒荒漠(AD) 5种植被类型, 测定了涉及碳(C)、N和P转化的14种土壤酶活性, 通过对青藏高原不同类型高寒草地土壤酶活性特征及其影响因子的研究, 丰富对土壤酶在高寒草地养分循环利用中作用规律的认识, 对预测青藏高原高寒草地生态系统土壤酶活性及其对全球变化的响应和机理提供科学参考, 同时为高寒草地生态系统初级生产力的维持提供理论依据. ...
Nitrogen addition shapes soil enzyme activity patterns by changing pH rather than the composition of the plant and microbial communities in an alpine meadow soil 1 2019
... 目前, 国内外在高寒草地土壤酶活性研究方面所开展的大量工作主要集中于其对模拟氮(N)沉降、施肥(N肥、磷(P)肥、粪肥等)(Liu et al., 2017; Liu & Zhang et al., 2019)、升温(Wang et al., 2014)、放牧强度(Lin et al., 2017)、土地利用方式(Yang et al., 2018)等的响应, 且仅在小区域或单一植被类型中进行.而高寒草地作为青藏高原主要的植被类型, 不同类型的高寒草地反映着青藏高原不同的生态环境, 有关不同高寒草地土壤酶活性的研究却严重缺少.在未来全球气候变化大背景下, 探讨不同高寒草地土壤酶活性及其对环境变化的响应机制, 对高寒草地生态系统的保护与分类管理具有重要意义, 同时对青藏高原养分循环的研究具有积极作用.本研究在藏北高原典型高寒草地分布区域选取高寒草甸(AM)、高寒草原(AS)、高寒草甸草原(AMS)、高寒荒漠草原(ADS)、高寒荒漠(AD) 5种植被类型, 测定了涉及碳(C)、N和P转化的14种土壤酶活性, 通过对青藏高原不同类型高寒草地土壤酶活性特征及其影响因子的研究, 丰富对土壤酶在高寒草地养分循环利用中作用规律的认识, 对预测青藏高原高寒草地生态系统土壤酶活性及其对全球变化的响应和机理提供科学参考, 同时为高寒草地生态系统初级生产力的维持提供理论依据. ...
Enhanced nitrogen deposition over China 1 2013
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
Atmospheric ammonia concentration modulates soil enzyme and microbial activity in an oak forest affecting soil microbial biomass 1 2018
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Carbon, nitrogen, and phosphorus storage in alpine grassland ecosystems of Tibet: effects of grazing exclusion 1 2015
Responses of soil extracellular enzyme activities and microbial community properties to interaction between nitrogen addition and increased precipitation in a semi-arid grassland ecosystem 1 2020
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
The global stoichiometry of litter nitrogen mineralization 1 2008
... 在测定的14种土壤酶活性中, C、N和P循环酶在5种高寒草地类型均存在显著差异的酶.其中参与C循环的5种酶均存在显著差异, 说明不同高寒草地类型是影响C循环酶活性的重要因子.而8种N循环酶中仅有2种酶活性在不同草地类型间存在显著差异, 这可能是由于土壤中N的累积比C的累积要慢很多(Veres et al., 2015; Kuypers et al., 2018), 为了平衡土壤中C含量高于N含量的现象, 微生物倾向于分泌更多的N 循环酶(Zhang et al., 2019), 并降低C循环酶的分泌, 且N获取策略具有多样性, N循环酶会与C的获取相耦合(Sinsabaugh et al., 2008), 同时, 分布在几类聚合物以及腐殖质分子中的N使得N的获取策略与特定类群的C底物偏好有关(Manzoni et al., 2008).因此, N循环酶不仅受环境因子的调控, 同时受C循环酶的底物特性及酶活性调控, 从而导致C循环酶和N循环酶在不同草地类型中差异不一致.另外, N转化过程中涉及N同化的两个酶NR与NiRs无显著相关关系, 可能是因为不同气候区域下土壤亚硝酸盐微生物群落分异造成的结果(Katsuyama et al., 2008). ...
Changes in soil phenol oxidase activities due to long-term application of compost and mineral N in a walnut orchard 1 2018
... 本研究涉及的14种酶中, NAG活性较高, 一定程度上反映了N转化在高寒草地生态系统的重要性.因为NAG参与几丁质的分解, 参与了有机N的吸收(Sinsabaugh et al., 1992), 在其他β-1,4-连接的葡糖胺聚合物的降解中也发挥作用, 类似于βG在纤维素降解中的作用(Sinsabaugh et al., 2005).且由于酶是富含N的化合物, 其生产受到N可用性的严格控制(Keeler et al., 2009), 从而导致NAG的活性较高.因此, 参与N吸收的NAG间接反映了N转化过程在该区域的重要性.PPO可以降解木质素和可溶性酚类化合物(Mazzon et al., 2018), 是参与C循环过程的重要酶, NAG是N循环重要酶, 而相关分析表明NAG与PPO呈显著负相关关系, 这可能是因为PPO是分解复杂C的酶, 而NAG作为参与几丁质分解的N转化酶, 基于酶化学计量的meta分析发现, N的添加会加剧微生物量C限制, 从而造成两种酶之间的负相关(Chen et al., 2018).ALP活性直接影响有机磷在土壤中的分解和转化, 并加速有机P的脱磷率(von Sperber et al., 2014), 在AS中最高, AD中最低, 说明AS土壤中微生物养分需求受到P的限制最严重, 而AD土壤中的P限制较轻. ...
A microplate fluorimetric assay for the study of enzyme diversity in soils 1 2001
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
主要草原生态系统生产力对气候变化响应的模拟 1 2012
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
主要草原生态系统生产力对气候变化响应的模拟 1 2012
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
Soil enzyme activity: a brief history and biochemistry as a basis for appropriate interpretations and meta-analysis 1 2018
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Warming and grazing increase mineralization of organic P in an alpine meadow ecosystem of Qinghai-Tibet Plateau, China 1 2012
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland 2 2008
Transformation of Quercus petraea litter: successive changes in litter chemistry are reflected in differential enzyme activity and changes in the microbial community composition 1 2011
Effects of short-term and long-term warming on soil nutrients, microbial biomass and enzyme activities in an alpine meadow on the Qinghai-Tibet Plateau of China 1 2014
... 目前, 国内外在高寒草地土壤酶活性研究方面所开展的大量工作主要集中于其对模拟氮(N)沉降、施肥(N肥、磷(P)肥、粪肥等)(Liu et al., 2017; Liu & Zhang et al., 2019)、升温(Wang et al., 2014)、放牧强度(Lin et al., 2017)、土地利用方式(Yang et al., 2018)等的响应, 且仅在小区域或单一植被类型中进行.而高寒草地作为青藏高原主要的植被类型, 不同类型的高寒草地反映着青藏高原不同的生态环境, 有关不同高寒草地土壤酶活性的研究却严重缺少.在未来全球气候变化大背景下, 探讨不同高寒草地土壤酶活性及其对环境变化的响应机制, 对高寒草地生态系统的保护与分类管理具有重要意义, 同时对青藏高原养分循环的研究具有积极作用.本研究在藏北高原典型高寒草地分布区域选取高寒草甸(AM)、高寒草原(AS)、高寒草甸草原(AMS)、高寒荒漠草原(ADS)、高寒荒漠(AD) 5种植被类型, 测定了涉及碳(C)、N和P转化的14种土壤酶活性, 通过对青藏高原不同类型高寒草地土壤酶活性特征及其影响因子的研究, 丰富对土壤酶在高寒草地养分循环利用中作用规律的认识, 对预测青藏高原高寒草地生态系统土壤酶活性及其对全球变化的响应和机理提供科学参考, 同时为高寒草地生态系统初级生产力的维持提供理论依据. ...
Plant species- specific recognition of long and short β-1,3-linked glucans is mediated by different receptor systems 1 2020
Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest 1 2013
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
A meta-analysis of soil extracellular enzyme activities in response to global change 1 2018
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Effects of exotic plantation forests on soil edaphon and organic matter fractions 1 2018
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
Initial soil responses to experimental warming in two contrasting forest ecosystems, Eastern Tibetan Plateau, China: nutrient availabilities, microbial properties and enzyme activities 1 2010
... 土壤酶作为土壤组分中最活跃的一种有机成分 (Marx et al., 2001), 参与土壤中各种化学反应和生物化学过程, 在有机物质矿化分解、矿质营养元素循环迁移、能量转化、环境质量评价等过程中扮演重要角色(Sinsabaugh et al., 2008, 2014; Sinsabaugh, 2010).因其能够对外界因素引起的微生物群落活动、土壤有机质和其他土壤特性的微小变化提供快速而准确的信息, 经常被用作土壤微生物和生化过程的基本和敏感指标(Nannipieri et al., 2018; Li et al., 2019), 在生态系统中具有重要的作用.据报道, 有一系列生物和非生物因素会影响土壤酶的活动, 例如土壤的pH, 养分的可利用性, 微生物的生物量和群落组成, 温度和降水(Sinsabaugh & Follstad Shah, 2011), 这些因素大致可分为三类, 分别为资源可得性、环境条件和微生物群落(Xiao et al., 2018).Sinsabaugh和Moorhead (1994)提出“资源分配理论”, 指微生物会根据资源的可利用性调节酶的生产, 即微生物将改变其胞外酶作为一种生存策略——在复杂资源的情况下增加胞外酶, 而在易获得简单营养成分(如无机氮和可溶性碳)的情况下减少胞外酶(Wallenius et al., 2011).环境条件影响因素主要指由气候变化(Henry, 2013; Jing et al., 2018), 如温度(Wallenstein et al., 2009; Xu et al., 2010)和降水的变化直接影响土壤酶活性, 或者由其引起的土壤理化性质和养分有效性的改变, 进而间接影响土壤酶活性(Hernández & Hobbie, 2010).土壤微生物群落的组成和生物量大小决定了酶产生的潜力, 从而直接或间接影响土壤酶的活性(Torres et al., 2015; Lopez-Aizpun et al., 2018).植被类型的差异会通过以上3种因素及其综合作用影响土壤酶活性(Burns et al., 2013), 例如, 从天然次生林到人工林的转变可以调节土壤微生物酶的网络和功能(Xu et al., 2018), 而我国西南亚高山森林生态系统的土壤酶活性主要取决于植被类型的影响(He et al., 2020). ...
降水和氮沉降增加对草地土壤微生物与酶活性的影响研究进展 1 2017
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
降水和氮沉降增加对草地土壤微生物与酶活性的影响研究进展 1 2017
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
Effects of different land types on soil enzyme activity in the Qinghai lake region 1 2018
... 目前, 国内外在高寒草地土壤酶活性研究方面所开展的大量工作主要集中于其对模拟氮(N)沉降、施肥(N肥、磷(P)肥、粪肥等)(Liu et al., 2017; Liu & Zhang et al., 2019)、升温(Wang et al., 2014)、放牧强度(Lin et al., 2017)、土地利用方式(Yang et al., 2018)等的响应, 且仅在小区域或单一植被类型中进行.而高寒草地作为青藏高原主要的植被类型, 不同类型的高寒草地反映着青藏高原不同的生态环境, 有关不同高寒草地土壤酶活性的研究却严重缺少.在未来全球气候变化大背景下, 探讨不同高寒草地土壤酶活性及其对环境变化的响应机制, 对高寒草地生态系统的保护与分类管理具有重要意义, 同时对青藏高原养分循环的研究具有积极作用.本研究在藏北高原典型高寒草地分布区域选取高寒草甸(AM)、高寒草原(AS)、高寒草甸草原(AMS)、高寒荒漠草原(ADS)、高寒荒漠(AD) 5种植被类型, 测定了涉及碳(C)、N和P转化的14种土壤酶活性, 通过对青藏高原不同类型高寒草地土壤酶活性特征及其影响因子的研究, 丰富对土壤酶在高寒草地养分循环利用中作用规律的认识, 对预测青藏高原高寒草地生态系统土壤酶活性及其对全球变化的响应和机理提供科学参考, 同时为高寒草地生态系统初级生产力的维持提供理论依据. ...
Ecological and environmental issues faced by a developing Tibet 1 2012
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
Impact of soil leachate on microbial biomass and diversity affected by plant diversity 1 2019
... 在测定的14种土壤酶活性中, C、N和P循环酶在5种高寒草地类型均存在显著差异的酶.其中参与C循环的5种酶均存在显著差异, 说明不同高寒草地类型是影响C循环酶活性的重要因子.而8种N循环酶中仅有2种酶活性在不同草地类型间存在显著差异, 这可能是由于土壤中N的累积比C的累积要慢很多(Veres et al., 2015; Kuypers et al., 2018), 为了平衡土壤中C含量高于N含量的现象, 微生物倾向于分泌更多的N 循环酶(Zhang et al., 2019), 并降低C循环酶的分泌, 且N获取策略具有多样性, N循环酶会与C的获取相耦合(Sinsabaugh et al., 2008), 同时, 分布在几类聚合物以及腐殖质分子中的N使得N的获取策略与特定类群的C底物偏好有关(Manzoni et al., 2008).因此, N循环酶不仅受环境因子的调控, 同时受C循环酶的底物特性及酶活性调控, 从而导致C循环酶和N循环酶在不同草地类型中差异不一致.另外, N转化过程中涉及N同化的两个酶NR与NiRs无显著相关关系, 可能是因为不同气候区域下土壤亚硝酸盐微生物群落分异造成的结果(Katsuyama et al., 2008). ...
2000-2010年藏北申扎县植被NDVI时空变化与气候因子的关系 1 2014
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
2000-2010年藏北申扎县植被NDVI时空变化与气候因子的关系 1 2014
... 藏北高原主要位于念青唐古拉山和冈底斯山之间, 是青藏高原的核心地区, 平均海拔4 500 m以上(Hong et al., 2014), 南北纵跨约700 km (30.45°- 35.65° N), 东西横越约2 400 km (83.68°-95.17° E), 面积约44.6万km2, 大约占西藏自治区全部土地面积的37% (段敏杰等, 2011).该地区属高原亚寒带季风气候, 寒冷干燥, 多大风, 年平均气温低于0 ℃, 最冷月是1月, 最热月为7月, 年降水量低于500 mm, 全年降水量的80%集中于6-9月, 年蒸发量1 799.6 mm (张晓克等, 2014).年日照时间通常高于同纬度其他地区(年平均日照率约60%), 太阳年总辐射量约为6 000 MJ·m-2 (莫志鸿等, 2012). ...
Soil bacterial and fungal diversity differently correlated with soil biochemistry in alpine grassland ecosystems in response to environmental changes 1 2017
... 青藏高原作为世界上最大最高的单一地貌单元, 被称为世界“第三极”, 是高寒草地广泛分布的典型区域(Zhang et al., 2017), 其高寒生态系统是全球高寒生态系统的典型和重要组成部分, 对环境变化敏感(Fu & Shen, 2017).因此, 该区域对于研究亚洲和世界的气候变化和指示生态系统的变化具有关键作用(Djukic et al., 2010; Rui et al., 2012).青藏高原已经经历了过去几十年的环境急剧变化(如气候变暖、降水格局变化、氮沉积和冻土层解冻等)(Yu et al., 2012; Liu et al., 2013).这些环境变化将对地上和地下生物多样性及生态系统功能产生根本性影响(Jing et al., 2015).同时, 高寒草原的独特特征(例如低温、温度和降水的急剧变化, 生长季节短, 仅适应当地的植物物种, 相对年轻的土壤等)使得探究该区域土壤酶活性是否对环境变化敏感具有重要的科学意义(Jing et al., 2018). ...
Changes in soil microbial community, enzyme activities and organic matter fractions under long-term straw return in north-central China 1 2016
... 土壤酶活性与环境因子之间具有相互作用.非生物因素可以通过改变土壤微生物的活动或者群落结构从而间接影响土壤酶的活性(Kivlin et al., 2014), 而土壤养分循环和C周转又依赖于土壤酶活性(Weintraub et al., 2013).影响土壤酶活性的关键因子(SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量)中既包含生物因素又包含非生物因素.在本研究中, AM比其他4类高寒草地(AS、AMS、ADS和AD)具有更高的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:N、C:P、N:P及更低的容重、TP含量.而AD比其他4类高寒草地(AM、AS、AMS和ADS)具有更低的SOM含量、TN含量、微生物数量、地上和地下植物生物量、C:P、N:P及更高的TP含量、地下植物有机质含量.这些生物和非生物因子在不同生态系统的差异都可能导致了不同功能和类别的酶活性在5种高寒草地类型间的差异, 同时不同植被类型中凋落物类型的变化也会使酶活性产生差异(Adamczyk et al., 2014).本研究发现SOM含量、N:P、TN含量、细菌数量、真菌数量、放线菌数量、G+数量、G-数量关键因子均与INV和CEL存在显著正相关关系, 说明INV和CEL活性相对于其他酶更易受到环境变化的影响.在19个环境因子指标中, SOM含量对土壤酶的影响重要值最大, 这是因为SOM的分解能提高土壤酶活性和微生物数量, 同时微生物的代谢产物又可提高土壤酶活性(闫钟清等, 2017).Kivlin等(2014)对美国南加州17个站点的研究发现, 土壤酶最重要的调节剂是土壤中C和N的浓度, 本研究结果与之相近, SOM和TN含量是影响土壤酶活性的关键因子, 这可能是由于SOM和N是酶促底物的重要贡献源, 或其通过改变土壤物理性质从而影响酶活性, 而微生物在充足的物质条件和养分条件下可以对酶活性产生主要贡献(Zhao et al., 2016).Ma等(2020)对半干旱草原的研究发现土壤元素化学计量与土壤酶活性之间存在显著相关性, 例如, C:N和C:P分别与降解C和N的酶活性呈负相关关系.而本研究的结果是N:P显著影响C、P循环酶活性, 这可能说明N、P作为青藏高原高寒草地的限制元素, 其养分循环的耦合关系对C、P循环酶活性的调节作用在高寒草地相对重要.之前的研究表明土壤pH与土壤微生物和细胞外酶活性的变化密切相关(Li et al., 2018), 而本研究中pH对土壤酶活性并未产生显著影响. ...
 ,1,*
,1,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT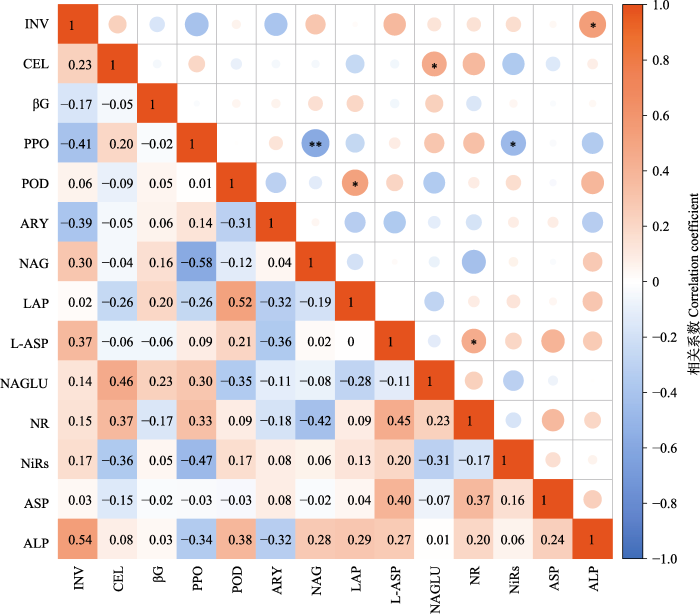 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT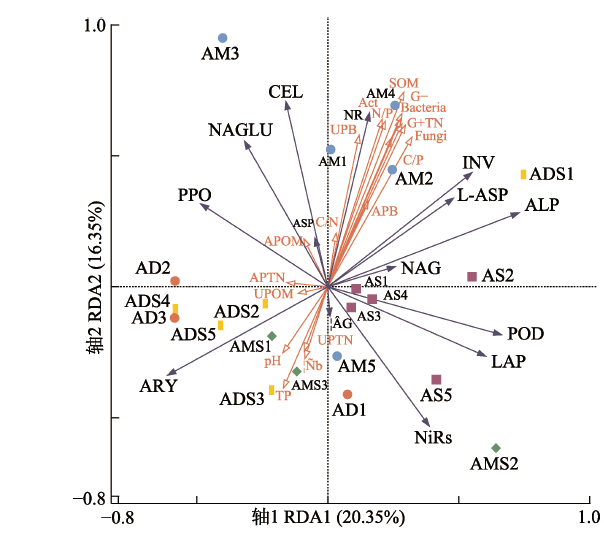 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}