董利军
1 , 李金花
, 1 , * , 陈珊
1 , 张瑞
1 , 孙建
2 , 马妙君
1 1 兰州大学生命科学学院, 草地农业生态系统国家重点实验室, 兰州 7300002 中国科学院地理科学与资源研究所生态网络观测与模拟重点实验室, 北京 100101Changes in soil organic carbon content and their causes during the degradation of alpine meadows in Zoigê Wetland Li-Jun DONG
1 , Jin-Hua LI
, 1 , * , Shan CHEN
1 , Rui ZHANG
1 , Jian SUN
2 , Miao-Jun MA
1 1 State Key Laboratory of Grassland Agro-ecosystem, School of Life Sciences, Lanzhou University, Lanzhou 730000, China2 Key Laboratory of Ecosystem Network Observation and Modelling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China通讯作者: *(
jinhuali@lzu.edu.cn )
编委: 王根绪
责任编辑: 李敏
收稿日期: 2020-07-13
接受日期: 2020-10-23
基金资助: 国家重点研发计划 (2017YFC0504802 )
Corresponding authors: *(
jinhuali@lzu.edu.cn )
Received: 2020-07-13
Accepted: 2020-10-23
摘要 土壤碳输入与输出之间的收支差决定土壤有机碳(SOC)含量。若尔盖湿地高寒草甸退化过程中, 土壤碳输入和输出哪个过程对SOC含量的影响占主导作用还不明确。该研究用空间序列代替时间序列的方法研究了若尔盖湿地高寒草甸不同退化阶段(高寒草甸(AM)、轻度退化高寒草甸(SD)和重度退化高寒草甸(HD)) SOC含量变化及原因。首先, 通过测定高寒草甸退化阶段上主要的土壤理化性状、微生物生物量、植物生物量和功能群组成的变化, 分析了退化阶段上土壤碳输入量的变化及原因; 其次, 结合室内土壤碳矿化培养实验结果和研究区的月平均气温以及土壤呼吸温度敏感性(
Q 10 )估算了该区域土壤碳输出, 并分析了其变化原因; 最后, 分析了造成SOC含量变化的主要原因和过程。结果表明: 在退化梯度上, 土壤含水量(
SWC )、SOC和全氮(TN)含量、微生物生物量碳氮含量降低; 植物群落组成逐渐从莎草科、禾本科占优势过渡到杂类草占优势, 且植物生物量降低; SOC矿化量降低; 有机碳潜在积累量降低(与AM阶段相比, SD和HD阶段有机碳潜在输入量、输出量和积累量分别降低了16%、18%、15%和59%、63%、41%)。
SWC 降低引起土壤容重、SOC含量、TN含量、全磷含量、C:N的改变, 进而导致植物功能群分布模式和土壤微生物的变化, 最终引起SOC输入和输出量的降低。
SWC 降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中SOC含量下降的主要原因。
关键词: 高寒草甸 ;
土壤有机碳 ;
碳输入 ;
土壤累积碳矿化 Abstract Aims The balance between soil organic carbon (SOC) input and output processes determines SOC content. However, it is not clear which of the two processes dominantly affect SOC content during the degradation of alpine meadows in Zoigê Wetland. In this study, the changes in SOC contents of alpine meadows and their causes at different degradation stages (alpine meadow (AM), slightly degraded alpine meadow (SD), and heavily degraded alpine meadow (HD)) in the Zoigê Wetland were investigated using the method of spatial sequence instead of temporal successional sequence.
Methods First, the changes in C input to soil and their causes along the degradation gradient were analyzed by investigating main soil physicochemical properties, microbial biomass, plant biomass and community composition of plant functional groups at different degradation stages. Secondly, the changes in the C output from soil were estimated based on lab incubation experiments of soil C mineralization and the temperature sensitivity of soil respiration (
Q 10 ) and monthly average air temperature of the Zoigê Wetland. Finally, the main causes and processes leading to changes in SOC content along the degradation gradient were analyzed.
Important findings The results showed that soil water content (
SWC ), SOC content, total nitrogen (TN) content, microbial biomass C and N content decreased with the increase of degradation. Plant community composition gradually changed from sedges and grasses dominated community to forbs dominated community. Plant biomass and SOC mineralization rate decreased during the degradation of alpine meadows. The potential accumulation of organic C reduced during the degradation (compared with AM, the potential input, output and accumulation of organic C in SD and HD decreased by 16%, 18%, 15% and 59%, 63%, 41%, respectively). The decrease in
SWC changed soil physical and chemical properties, including bulk density, SOC content, TN content, total phosphorus content, and C:N, which led to the shifts in the distribution pattern of plant functional groups and in soil microorganisms, consequently reducing the inputs and outputs of SOC. The decrease in potential plant-derived C input to soil caused by decreased
SWC was the main reason for the decline in SOC content along the degradation gradient of alpine meadows in Zoigê Wetland.
Keywords: alpine meadow ;
soil organic carbon ;
carbon input ;
soil accumulative carbon mineralization PDF (579KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 董利军, 李金花, 陈珊, 张瑞, 孙建, 马妙君. 若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析. 植物生态学报, 2021, 45(5): 507-515. DOI:
10.17521/cjpe.2020.0231 DONG Li-Jun, LI Jin-Hua, CHEN Shan, ZHANG Rui, SUN Jian, MA Miao-Jun.
Changes in soil organic carbon content and their causes during the degradation of alpine meadows in Zoigê Wetland .
Chinese Journal of Plant Ecology , 2021, 45(5): 507-515. DOI:
10.17521/cjpe.2020.0231 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物。SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 )。由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 )。因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 )。SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 )。土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 )。土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 )。
草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 )。随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 )。高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 )。此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 )。
若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 )。但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 )。若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 )。但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚。本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据。依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低。
1 材料和方法 1.1 研究区概况 研究区域位于四川省阿坝藏族羌族自治州若尔盖县花湖自然保护区内, 地理位置为33.92°-33.94° N, 102.82°-102.83° E; 海拔3 400 m左右, 年平均气温为1.1 ℃。年降水量600-800 mm, 雨季温暖湿润(5-9月), 降水量占全年的90%, 旱季寒冷干燥(10月至次年4月)。研究区域土壤类型主要为草甸土, 植被类型以高山草甸植被为主, 主要优势种有木里薹草(
Carex muliensis )、矮生嵩草(
Kobresia humilis )和二裂委陵菜(
Potentilla bifurca )(
Jin et al ., 2020 )。
1.2 实验设计与样品采集 研究区内放牧家畜主要为牦牛和绵羊。本研究根据调查区域内优势种和伴生种种类、植被盖度、表层水分概况及微地形条件(塔头高低、水位高低、有无鼠兔洞穴等), 采用空间序列代替时间序列的方法, 在若尔盖县花湖研究区划分了高寒草甸退化的3个阶段(高寒草甸(AM)、轻度退化高寒草甸(SD)、重度退化高寒草甸(HD)(
表1 )), 每个阶段选择5个25 m × 25 m的小区, 每个小区内部再随机选择3个5 m × 5 m的采样点, 共计45个采样点。通过调查和测定植物群落组成、生物量、土壤理化性质、微生物生物量、土壤碳矿化速率等估算有机碳潜在输入量和输出量, 探讨不同退化阶段草甸SOC含量的变化机制。
Table 1 表1 表1 若尔盖湿地高寒草甸退化阶段划分依据
Table 1
Classification basis of degradation stages of alpine meadows in Zoigê Wetland 草地类型 优势种 主要伴生种 塔头情况 鼠兔洞穴 总盖度 高寒草甸 木里薹草Carex muliensis 鹅绒委陵菜、矮生嵩草Potentilla anserine, Kobresia humilis 塔头高10-20 cm 无 92 轻度退化高寒草甸 木里薹草C. muliensis 鹅绒委陵菜、华扁穗草P . anserine, Blysmus 无塔头 有 97 重度退化高寒草甸 二裂委陵菜P . bifurca 鹅绒委陵菜、木里薹草P . anserine, C . muliensis 无塔头, 出现斑块化裸地, 部分土地沙化 有 64
新窗口打开 |
下载CSV 于2018年8月进行样品采集。植物样品采集时, 在每个退化阶段选取具有典型植被特征的15个采样点做植物样方(50 cm × 50 cm)调查, 记录主要物种名称、株高、盖度, 之后按莎草科、禾本科和杂类草分类齐地面剪下, 用烘干称质量法获得植物地上生物量。在剪过草的样方内用土钻(内径5 cm)取0-20 cm土在流水中冲洗土壤, 分离出根系, 然后在烘箱内65 ℃烘至恒质量, 称量获得根系生物量。土壤样品的采集采用五点取样法, 在5 m × 5 m的样地内, 沿对角线在每个取样点将植物地上部分贴地剪去, 用内径为3.5 cm的土钻取15钻0-20 cm土混合后去除根系、石块等杂物过2 mm筛, 一部分自然风干用于测定SOC含量、全氮(TN)含量等指标, 另一部分4 ℃冷藏, 用于微生物生物量测定和土壤碳矿化培养实验。
1.3 样品处理与测定 土壤基本理化性质采用常规分析法测定: 土壤含水量用烘干法; 容重用环刀法; pH用PHS-3酸度计; SOC含量用重铬酸钾外加热氧化法; TN和全磷(TP)消煮后用SmartChem200全自动化学分析仪(WestCo Scientific Instrument, Brookfielg, USA)测定。微生物生物量碳含量(
MBC )、氮含量(
MBN )、磷含量(
MBP )用氯仿熏蒸法测定。
土壤碳矿化速率的测定: 碱液吸收法是测定土壤呼吸的一种常用、简便的方法。青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化; 在室内20 ℃恒温培养条件下用碱液吸收法测得的CO
2 通量值与野外条件下用静态气室法测得的数值非常接近(
Song et al ., 2010 ), 说明该方法是一种简单易行且可靠的研究复杂环境条件下土壤呼吸的方法。因此, 本研究中采用通用的室内20 ℃恒温培养、碱液吸收法测定土壤碳矿化速率。简单过程如下: 相当于70 g干土的鲜土分别放入培养瓶中, 将土壤水分调节至田间持水量的60%, 置于培养箱中20 ℃无光条件下恒温培养, 在第1、2、3、5、7、9、12、15、19、23、27、31天分别测定土壤呼吸速率并计算碳累积矿化量(
李峰, 2019 )。
1.4 数据处理与计算 有机碳潜在输入量计算公式如下:
(1) PCI = 地上植物碳输入量+根系碳输入量+根系分泌物碳输入量其中, 地上植物碳输入量=
AGB ×植物碳含量/
SA; 根系碳输入量=
BGB ×根系碳含量× 20%/
SA; 根系分泌物碳输入量
= AGB ×植物碳含量× 10%/
SA (
Chapin III et al ., 2012 )。式中,
PCI 为有机碳潜在输入量(g·m
-2 ),
AGB 为样方地上生物量(g),
BGB 为根系生物量(g),
SA 为样方面积(m
2 )。
利用室内实验的土壤碳累积矿化量, 根据月平均气温和土壤呼吸温度敏感性(
Q 10 )估算出全年各月份的土壤碳累积矿化量, 公式如下:
(2) CCMn = CCM 0 × Q 10 × (Tn - T 0 )/10式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度。
由全年土壤碳累积矿化量估算出若尔盖全年的有机碳潜在输出量, 计算公式如下:
(3) PCO = CCM × BD × V /S 式中,
PCO 为有机碳潜在输出量(g·m
-2 );
CCM 为碳累积矿化量(g·kg
-1 );
BD 为容重(g·m
-3 );
V 为土壤体积(m
3 ),
V = 0.2 m
3 , 因为采样土层深度为0.2 m, 为与潜在输入量对应, 输出量对应的面积按1 m
2 算;
S = 1 m
2 , 表示单位面积。
有机碳潜在积累量用有机碳潜在输入量与潜在输出量的差值表示。
植物功能群的优势度用重要值表示, 计算如下: 重要值= (相对盖度+相对高度+相对生物量)/3 (
刘育红等, 2018 )。
采用Microsoft Excel 2013对实验数据进行计算和整理, SPSS 23.0分析软件对数据进行非参数检验, 用克鲁斯卡尔-沃利斯非参数检验进行多重比较; 对土壤及微生物碳氮磷含量、含水量、容重与植物类群及碳累积矿化量进行冗余分析(RDA); 对植物类群与退化梯度进行主成分分析(PCA)。用Origin 2018和Canoco 5软件作图。
2 结果和分析 2.1 退化梯度上土壤理化特征 从AM到SD、HD阶段, 土壤含水量的平均值分别为136%、74%、29%; SOC含量的平均值分别为210、149和46 g·kg
-1 ; TN含量的平均值分别为15、9和2 g·kg
-1 ; TP含量的平均值分别为1.2、1.8和1.4 g·kg
-1 ; C:N的平均值分别为15、18和22。
MBC 的平均值分别为1 256、587和440 mg·kg
-1 ;
MBN 的平均值分别为414、263和222 mg·kg
-1 ;
MBP 的平均值分别为58、69和42 mg·kg
-1 (
图1 )。土壤含水量、SOC含量、TN含量和C:N均表现为AM与SD之间差异不显著(
p > 0.05), AM、SD与HD之间存在显著差异(
p < 0.05); TP含量表现为AM与SD差异显著(
p < 0.05), HD与AM、SD均差异不显著(
p > 0.05);
MBC 和
MBN 均表现为AM与SD、HD差异显著(
p < 0.05), SD与HD差异不显著(
p > 0.05);
MBP 表现为AM与HD差异显著(
p < 0.05), HD与AM、SD均差异不显著(
p > 0.05)。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1若尔盖湿地高寒草甸退化梯度上土壤理化性质变化特征(平均值±标准误)。AM, 高寒草甸; HD, 重度退化高寒草甸; SD, 轻度退化高寒草甸。不同小写字母表示在退化梯度上差异显著(p < 0.05)。 Fig. 1Soil physicochemical characteristics along degradation gradients (mean ± SE ). AM, alpine meadow; HD, heavily degraded alpine meadow; SD, slightly degraded alpine meadow. Different lowercase letters indicate significant difference among the degradation stages (p < 0.05). 2.2 退化梯度上植物功能群PCA及RDA 2.2.1 退化梯度上植物功能群PCA
PCA结果显示: 莎草科、禾本科重要值沿第1排序轴自左向右减小, 反映出这两个功能群与高寒草甸退化程度间呈负相关关系; 杂类草重要值沿第1排序轴自左向右增大, 反映出该功能群与高寒草甸退化程度间呈正相关关系。AM阶段样地与莎草科和禾本科接近, SD阶段样地与杂类草、禾本科和莎草科均较接近, HD阶段与杂类草接近; 反映出随着退化程度的加重, 莎草科和禾本科优势度变弱, 而杂类草优势度增加(
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2植物功能群(禾本科、莎草科、杂类草)重要值与退化梯度主成分分析(PCA)。AM, 高寒草甸; HD, 重度退化高寒草甸; SD, 轻度退化高寒草甸。 Fig. 2Principal component analysis (PCA) of plant functional groups (grasses, sedges, and forbs) importance value and degradation gradient. AM, alpine meadow; HD, heavily degraded alpine meadow; SD, slightly degraded alpine meadow. 2.2.2 退化梯度上植物功能群重要值与土壤理化性质RDA
RDA二维排序图显示: 禾本科重要值与SOC含量、C:P、
SWC 、TN含量、
MBC 、
MBC :
MBN 、
MBC :
MBP 呈正相关关系, 与TP含量、容重呈负相关关系; 莎草科重要值与
MBP 呈正相关关系, 与C:N呈负相关关系; 杂类草重要值与容重呈正相关关系, 与SOC含量、TN含量和
SWC 呈负相关关系。该结果表明禾本科受SOC含量、C:P和容重的影响较大, 莎草科受C:N和
MBP 的影响较大, 杂类草受SOC含量和容重的影响较大(
图3A )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3植物功能群重要值(A)和土壤碳累积矿化量(B)与土壤因子的冗余分析(RDA)。C 1 、C 15 、C 31 分别表示第1、15、31天的土壤碳累积矿化量。BD, 容重; MBC, 微生物生物量碳含量; MBN, 微生物生物量氮含量; MBP, 微生物生物量磷含量; SOC, 土壤有机碳含量; SWC, 土壤含水量; TN, 全氮含量; TP, 全磷含量。 Fig. 3Redundancy analysis (RDA) of plant functional groups importance value (A), cumulative soil carbon mineralization (B) and soil physicochemical properties. C 1 , C 15, and C 31 represent the cumulative mineralization amount of soil carbon during 1day, 15 days, and 31 days of incubation, respectively. BD, bulk density; MBC, microbial biomass carbon content; MBN, microbial biomass nitrogen content; MBP, microbial biomass phosphorus content; SOC, soil organic carbon content; SWC, soil water content; TN, total nitrogen content; TP, total phosphorus content. 2.3 退化梯度上土壤碳累积矿化量与土壤因子RDA 土壤碳累积矿化量与土壤理化性质的RDA结果显示: 土壤碳累积矿化量与SOC含量、
SWC 、TN含量、C:P、N:P、
MBC :
MBN 正相关, 与容重和C:N负相关。该结果表明
SWC 、SOC含量和TN含量等的增加有利于土壤碳的矿化, 而土壤容重和C:N的增加不利于土壤碳的矿化(
图3B )。
2.4 退化梯度上土壤有机碳潜在积累量变化特征 在退化梯度上, 土壤有机碳潜在输入量、输出量和积累量均呈降低趋势, 但潜在输出量、积累量SD阶段与AM阶段之间差异不显著(
p > 0.05), 而各指标在HD阶段与AM、SD阶段相比显著降低(
p < 0.05)(
图4 )。与AM阶段相比, SD和HD阶段有机碳潜在输入量、输出量和积累量分别降低了16%、18%、15%和59%、63%、41%。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4退化梯度上土壤有机碳潜在输入量、输出量、积累量变化特征(平均值±标准误)。AM, 高寒草甸; HD, 重度退化高寒草甸; SD, 轻度退化高寒草甸。不同小写字母表示该指标在退化梯度上差异显著(p < 0.05)。 Fig. 4Characteristics of potential input, output, and accumulation of soil organic carbon along degradation gradients (mean ± SE ). AM, alpine meadow; HD, heavily degraded alpine meadow; SD, slightly degraded alpine meadow. Different lowercase letters indicate significant difference among the degradation stages (p < 0.05). 3 讨论 3.1 土壤主要理化特征的变化 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致。多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 )。与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 )。已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 )。
3.2 土壤碳输入和输出动态变化 SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 )。随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 )。这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 )。但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 )。
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 )。在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少。
SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 )。
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降。我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致。土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 )。在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 )。因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱。
3.3 土壤有机碳变化的主要原因 AM阶段微生物活性高, SOC分解量大, 但有机碳潜在输入量也高, 且输入量大于输出量, 使有机碳潜在积累量较高(
图4 ), 因此该阶段SOC总量最高; SD和HD阶段微生物活性减弱, SOC分解减少, 但是有机质输入量也减少, 导致有机碳潜在积累量降低(
图4 ), 造成SOC总量变小。从AM到HD阶段,
SWC 降低导致植物类群和生物量改变(
图2 ,
图3 )(
刘伟等, 2005 ;
唐明坤等, 2011 )、微生物生物量碳氮含量降低(
图1 ), 导致碳输入显著降低、碳输出显著减弱(与AM相比, SD和HD阶段有机碳潜在输入量分别降低了56和192 g·m
-2 , 潜在输出量分别降低了48和170 g·m
-2 ), 碳输入降低的量略大于碳输出降低的量, 但碳输入、输出的降幅基本相当(
图4 )。净碳积累量AM > SD >> HD, 该结果表明: 与未退化的高寒草甸相比, 重度退化高寒草甸土壤碳积累大大降低, 造成退化梯度上SOC含量降低。因此, 若尔盖高寒草甸退化过程中, 植物碳潜在输入量的减少和微生物分解作用的减弱共同影响SOC含量的变化, 但植物碳潜在输入量的减少是SOC含量降低的主导过程。
需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差。例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量。目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 )。在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 )。植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积。此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 )。另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出。不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 )。青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况。因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量。虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 )。该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势。上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量。因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理。
4 结论 随着草甸退化程度的加剧,
SWC 、SOC含量和TN含量降低; 植物群落从莎草科、禾本科为主的群落向以杂类草为主的群落转变。植物生物量降低导致碳潜在输入量严重降低; 微生物生物量碳氮含量和微生物活性降低导致碳潜在输出量显著降低; 碳输入降低的量略大于碳输出降低的量, 但碳输入、输出的降幅基本相当。因此, 土壤含水量的降低引起的植物碳潜在输入量的减少是若尔盖湿地高寒草甸退化过程中SOC含量降低的主导过程。
致谢 感谢若尔盖湿地国家级自然保护区提供野外工作条件; 感谢草地农业生态系统国家重点实验室提供实验设备条件; 感谢研究生李峰在实验中提供的支持与帮助。
[1] Chang ZQ Shi ZM Feng Q Su YH 2005 ).Temporal variation of soil respiration on sloping pasture of Heihe River basin and effects of temperature and soil moisture on itChinese Journal of Applied Ecology ,16, 1603 -1606 . [本文引用: 1] [常宗强 ,史作民 ,冯起 ,苏永红 (2005 ).黑河流域山区牧坡草地土壤呼吸的时间变化及水热因子影响应用生态学报 ,16, 1603 -1606 .] [本文引用: 1] [2] Chapin III FS Matson PA Vitousek PM 2012 ).Principles of Terrestrial Ecosystem Ecology [本文引用: 1] [3] Chen H 2018 ).Impact of Water Status on Soil Microbial Biomass and Community Structure in Typical Grasslands in Inner Mongolia China Agriculture University, Beijing . [本文引用: 1] [陈昊 (2018 ).水分状况对内蒙古典型草原土壤微生物量及群落结构的影响中国农业大学, 北京 .] [本文引用: 1] [4] Cong J Wang XL Liu X Zhang YG 2016 ).The distribution variation and key influencing factors of soil organic carbon of natural deciduous broadleaf forests along the latitudinal gradientActa Ecologica Sinica ,36, 333 -339 . DOI:
10.1016/j.chnaes.2016.06.002 URL [本文引用: 4] [5] Cox PM Betts RA Jones CD Spall SA Totterdell IJ 2000 ).Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate modelNature ,408, 184 -187 . DOI:
10.1038/35041539 URL [本文引用: 1] [6] Dlamini P Chivenge P Manson A Chaplot V 2014 ).Land degradation impact on soil organic carbon and nitrogen stocks of sub-tropical humid grasslands in South AfricaGeoderma 235-236 -372-381 . [本文引用: 4] [7] Dong SK Wen L Yi YY Wang XX Zhu L Li XY 2012 ).Soil-quality effects of grassland degradation and restoration on the Qinghai-Tibetan PlateauSoil Science Society of America Journal ,76, 2256 -2264 . DOI:
10.2136/sssaj2012.0092 URL [本文引用: 1] [8] Gao Y Zhou J Wang LM Guo JM Feng JX Wu H Lin GH 2019 ).Distribution patterns and controlling factors for the soil organic carbon in four mangrove forests of ChinaGlobal Ecology and Conservation ,17, e00575 . DOI:10.1016/j.gecco.2019.e00575 . DOI:
10.1016/j.gecco.2019.e00575 URL [本文引用: 1] [9] Guo XJ Du W Wang X Yang ZF 2013 ).Degradation and structure change of humic acids corresponding to water decline in Zoige peatland, Qinghai-Tibet PlateauScience of the Total Environment 445-446 -231-236 . [本文引用: 1] [10] Hu K Tao JP Huang K Hu J Wang W 2020 ).Effects of simulated root exudate carbon inputs on the dynamics of microbial communities during litter decompositionChinese Journal of Applied and Environmental Biology ,26, 417 -424 . [本文引用: 1] [胡凯 ,陶建平 ,黄科 ,胡靖 ,王微 (2020 ).模拟根系分泌物碳输入对凋落叶分解中微生物群落动态的影响应用与环境生物学报 ,26, 417 -424 .] [本文引用: 1] [11] Huang XY Chen KL Wu CY 2016 ).Diurnal variation and seasonal dynamics of soil respiration in the alpine meadow in the Qinghai Tibet PlateauYunnan Geographic Environment Research ,28, 66 -71 . [本文引用: 3] [黄晓宇 ,陈克龙 ,吴成永 (2016 ).青藏高原高寒草甸生长季土壤呼吸的昼夜变化及其季节动态云南地理环境研究 ,28, 66 -71 .] [本文引用: 3] [12] Jiang WG Lü J Wang CC Chen Z Liu YH 2017 ).Marsh wetland degradation risk assessment and change analysis: a case study in the Zoige Plateau, ChinaEcological Indicators ,82, 316 -326 . DOI:
10.1016/j.ecolind.2017.06.059 URL [本文引用: 1] [13] Jin XY Qiang HF Zhao L Jiang SZ Cui NB Cao Y Feng Y 2020 ).SPEI-based analysis of spatio-temporal variation characteristics for annual and seasonal drought in the Zoige Wetland, Southwest China from 1961 to 2016Theoretical and Applied Climatology ,139, 711 -725 . DOI:
10.1007/s00704-019-02981-y URL [本文引用: 1] [14] Lal R 2004 ).Soil carbon sequestration to mitigate climate changeGeoderma ,123, 1 -22 . DOI:
10.1016/j.geoderma.2004.01.032 URL [本文引用: 1] [15] Li F 2019 ).Effects of Fertilization on Organic Carbon Stimulation and Net Carbon Balance in Alpine Meadow Soil Lanzhou University, Lanzhou . [本文引用: 3] [李峰 (2019 ).施肥对高寒草甸土壤有机碳激发效应及净碳平衡的影响兰州大学, 兰州 .] [本文引用: 3] [16] Li JH Hou YL Zhang SX Li WJ Xu DH Knops JMH Shi XM 2018 ).Fertilization with nitrogen and/or phosphorus lowers soil organic carbon sequestration in alpine meadowsLand Degradation and Development ,29, 1634 -1641 . DOI:
10.1002/ldr.v29.6 URL [本文引用: 2] [17] Li JH Zhang R Cheng BH Ye LF Li WJ Shi XM 2021 ).Effect of nitrogen and phosphorus additions on decomposition and accumulation of soil organic carbon in alpine meadows on the Tibetan PlateauLand Degradation and Development ,32, 1467 -1477 . DOI:
10.1002/ldr.v32.3 URL [本文引用: 3] [18] Li ZW Gao P Hu XY Yi YJ Pan BZ You YC 2020 ).Coupled impact of decadal precipitation and evapotranspiration on peatland degradation in the Zoige basin, ChinaPhysical Geography ,41, 145 -168 . DOI:
10.1080/02723646.2019.1620579 URL [本文引用: 1] [19] Liu W Zhou HK Zhou L 2005 ).Biomass distribution pattern of degraded grassland in alpine meadowGrassland of China ,27, 9 -15 . [本文引用: 3] [刘伟 ,周华坤 ,周立 (2005 ).不同程度退化草地生物量的分布模式中国草地 ,27, 9 -15 .] [本文引用: 3] [20] Liu XM Luo JF Chen DC Zhu XW Zhou JX 2019 ).Floristic assembly and dominant population niches in degraded grasslands on the Zoigê PlateauJournal of Zhejiang A&F University ,36, 289 -297 . [本文引用: 3] [刘学敏 ,罗久富 ,陈德朝 ,朱欣伟 ,周金星 (2019 ).若尔盖高原不同退化程度草地植物种群生态位特征浙江农林大学学报 ,36, 289 -297 .] [本文引用: 3] [21] Liu YH Yang YW Zhang Y 2018 ).Redundancy analysis of the relationship between plant functional groups and soil factors in the degraded alpine meadowJournal of Ecology and Rural Environment ,34, 1112 -1121 . [本文引用: 5] [刘育红 ,杨元武 ,张英 (2018 ).退化高寒草甸植物功能群与土壤因子关系的冗余分析生态与农村环境学报 ,34, 1112 -1121 .] [本文引用: 5] [22] Liu YJ 2017 ).Responses of Soil Microbial Community Structure and Function to Water Condition Change in Wetland Nanchang University, Nanchang . [本文引用: 1] [刘亚军 (2017 ).湿地土壤微生物群落结构和功能对水分条件变化的响应南昌大学, 南昌 .] [本文引用: 1] [23] Luan JW Cui LJ Xiang CH Wu JH Song HT Ma QF 2014 ).Soil carbon stocks and quality across intact and degraded alpine wetlands in Zoige, east Qinghai-Tibet PlateauWetlands Ecology and Management ,22, 427 -438 . DOI:
10.1007/s11273-014-9344-8 URL [本文引用: 4] [24] Miller AJ Schuur EAG Chadwick OA 2001 ).Redox control of phosphorus pools in Hawaiian montane forest soilsGeoderma ,102, 219 -237 . DOI:
10.1016/S0016-7061(01)00016-7 URL [本文引用: 3] [25] Nie C Li Y Niu L Liu YH Shao R Xu X Tian YQ 2019 ).Soil respiration and its Q 10 response to various grazing systems of a typical steppe in Inner Mongolia, ChinaPeer J e7112 . DOI:10.7717/peerj.7112 . DOI:
10.7717/peerj.7112 URL [本文引用: 1] [26] Qin SJ Liu JS Wang GP 2006 ).Mechanism of phosphorus availability changing in soilChinese Journal of Soil Science ,37, 1012 -1016 . [本文引用: 2] [秦胜金 ,刘景双 ,王国平 (2006 ).影响土壤磷有效性变化作用机理土壤通报 ,37, 1012 -1016 .] [本文引用: 2] [27] Qiu PF Wu N Luo P Wang ZY Li MH 2009 ).Analysis of dynamics and driving factors of wetland landscape in Zoige, Eastern Qinghai-Tibetan PlateauJournal of Mountain Science ,6, 42 -55 . DOI:
10.1007/s11629-009-0230-4 URL [本文引用: 1] [28] Song MH Jiang J Cao GM Xu XL 2010 ).Effects of temperature, glucose and inorganic nitrogen inputs on carbon mineralization in a Tibetan alpine meadow soilEuropean Journal of Soil Biology ,46, 375 -380 . DOI:
10.1016/j.ejsobi.2010.09.003 URL [本文引用: 3] [29] Suh S Lee E Lee J 2009 ).Temperature and moisture sensitivities of CO2 efflux from lowland and alpine meadow soilsJournal of Plant Ecology ,2, 225 -231 . DOI:
10.1093/jpe/rtp021 URL [本文引用: 4] [30] Tang MK Li MF Zhao J Chen WW Suolang D Sun ZY 2011 ).Comparative study of the alpine grassland communities in varying degradation in Zoige County, Sichuan ProvinceGuihaia ,31, 775 -781 . [本文引用: 7] [唐明坤 ,李明富 ,赵杰 ,陈薇薇 ,索朗夺尔基 ,孙治宇 (2011 ).四川若尔盖县不同退化程度高寒草地群落比较研究广西植物 ,31, 775 -781 .] [本文引用: 7] [31] Wang D Lü YL Xu L Zhang HX Wang RM He NP 2013 ).The effect of moisture and temperature on soil C mineralization in wetland and steppe of the Zoige region, ChinaActa Ecologica Sinica ,33, 6436 -6443 . DOI:
10.5846/stxb URL [本文引用: 1] [王丹 ,吕瑜良 ,徐丽 ,张洪轩 ,王若梦 ,何念鹏 (2013 ).水分和温度对若尔盖湿地和草甸土壤碳矿化的影响生态学报 ,33, 6436 -6443 .] [本文引用: 1] [32] Wang JL Zhong ZM Wang ZH Chen BX Yu CQ Hu XX Shen ZX Dacizhuoga Zhang XZ 2014 ).Soil C/N distribution characteristics of alpine steppe ecosystem in Qinghai-Tibetan PlateauActa Ecologica Sinica ,34, 6678 -6691 . [本文引用: 1] [王建林 ,钟志明 ,王忠红 ,陈宝雄 ,余成群 ,胡兴祥 ,沈振西 ,大次卓嘎 ,张宪洲 (2014 ).青藏高原高寒草原生态系统土壤碳氮比的分布特征生态学报 ,34, 6678 -6691 .] [本文引用: 1] [33] Wu PF Hua C Wei X 2014 ).Response of plant functional groups to the de-gradational succession of alpine meadow in Zoigê CountyJournal of Southwest University for Nationalities (Natural Science Edition )321 -329 .[34] [吴鹏飞 ,华翠 ,魏雪 (2014 ).若尔盖高寒草甸不同功能群植物数量特征对退化演替的响应西南民族大学学报(自然科学版) ,40, 321 -329 .] [本文引用: 3] [35] Xu L Yu SX He NP Wen XF Shi PL Zhang YJ Dai JZ Wang RM 2013 ).Soil C mineralization and temperature sensitivity in alpine grasslands of the Qinghai-Xizang PlateauChinese Journal of Plant Ecology ,37, 988 -997 . DOI:
10.3724/SP.J.1258.2013.00102 URL [本文引用: 1] [徐丽 ,于书霞 ,何念鹏 ,温学发 ,石培礼 ,张扬建 ,代景忠 ,王若梦 (2013 ).青藏高原高寒草地土壤碳矿化及其温度敏感性植物生态学报 ,37, 988 -997 .] [本文引用: 1] [36] Yang G Peng C Chen H Dong F Wu N Yang Y Zhu Q 2017 ).Qinghai-Tibetan Plateau peatland sustainable. utilization under anthropogenic disturbances and climate changeEcosystem Health and Sustainability ,3, e01263 . DOI:10.1002/ehs2.1263 . DOI:
10.1002/ehs2.1263 URL [37] Yu HY Zha TG Zhang XX Ma LM 2019 ).Vertical distribution and influencing factors of soil organic carbon in the Loess Plateau, ChinaScience of the Total Environment ,693, 133632 . DOI:10.1016/j.scitotenv.2019.133632 . DOI:
10.1016/j.scitotenv.2019.133632 URL [本文引用: 3] [38] Zhang MM Chen W Lin L Zhang DG Wu YX Xiao HL 2019 ).A study of soil nutrient characteristics and soil soluble organic carbon levels in different types of alpine grassland in Qinghai ProvinceActa Prataculturae Sinica ,28, 20 -28 . [本文引用: 1] [张苗苗 ,陈伟 ,林丽 ,张德罡 ,吴玉鑫 ,肖海龙 (2019 ).青海省不同高寒草地土壤主要养分及可溶性有机碳特性研究草业学报 ,28, 20 -28 .] [本文引用: 1] [39] Zhao YF Hong MM Ou YS Huang Z Zhang YY Wang X 2018 ).The stoichiometric characteristics of soil C, N, P in mountain steppe of eastern Tibetan PlateauEcological Science ,37, 25 -32 . [本文引用: 2] [赵云飞 ,洪苗苗 ,欧延升 ,黄政 ,张洋洋 ,汪霞 (2018 ).青藏高原东部山地草地土壤碳、氮、磷元素计量特征生态科学 ,37, 25 -32 .] [本文引用: 2] [40] Zhu XC Shao MG 2018 ).Distribution, stock, and influencing factors of soil organic carbon in an alpine meadow in the hinterland of the Qinghai-Tibetan PlateauJournal of Earth System Science ,127, 71 . DOI:10.1007/S12040-018-0974-8 . DOI:
10.1007/S12040-018-0974-8 URL [本文引用: 1] 黑河流域山区牧坡草地土壤呼吸的时间变化及水热因子影响
1
2005
... 式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度. ...
黑河流域山区牧坡草地土壤呼吸的时间变化及水热因子影响
1
2005
... 式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度. ...
1
2012
... 其中, 地上植物碳输入量=
AGB ×植物碳含量/
SA; 根系碳输入量=
BGB ×根系碳含量× 20%/
SA; 根系分泌物碳输入量
= AGB ×植物碳含量× 10%/
SA (
Chapin III et al ., 2012 ).式中,
PCI 为有机碳潜在输入量(g·m
-2 ),
AGB 为样方地上生物量(g),
BGB 为根系生物量(g),
SA 为样方面积(m
2 ). ...
水分状况对内蒙古典型草原土壤微生物量及群落结构的影响
1
2018
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
水分状况对内蒙古典型草原土壤微生物量及群落结构的影响
1
2018
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
The distribution variation and key influencing factors of soil organic carbon of natural deciduous broadleaf forests along the latitudinal gradient
4
2016
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
Acceleration of global warming due to carbon-cycle feedbacks in a coupled climate model
1
2000
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
Land degradation impact on soil organic carbon and nitrogen stocks of sub-tropical humid grasslands in South Africa
4
2014
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
... ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
Soil-quality effects of grassland degradation and restoration on the Qinghai-Tibetan Plateau
1
2012
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
Distribution patterns and controlling factors for the soil organic carbon in four mangrove forests of China
1
2019
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
Degradation and structure change of humic acids corresponding to water decline in Zoige peatland, Qinghai-Tibet Plateau
1
2013
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
模拟根系分泌物碳输入对凋落叶分解中微生物群落动态的影响
1
2020
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
模拟根系分泌物碳输入对凋落叶分解中微生物群落动态的影响
1
2020
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
青藏高原高寒草甸生长季土壤呼吸的昼夜变化及其季节动态
3
2016
... 式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度. ...
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
青藏高原高寒草甸生长季土壤呼吸的昼夜变化及其季节动态
3
2016
... 式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度. ...
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
Marsh wetland degradation risk assessment and change analysis: a case study in the Zoige Plateau, China
1
2017
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
SPEI-based analysis of spatio-temporal variation characteristics for annual and seasonal drought in the Zoige Wetland, Southwest China from 1961 to 2016
1
2020
... 研究区域位于四川省阿坝藏族羌族自治州若尔盖县花湖自然保护区内, 地理位置为33.92°-33.94° N, 102.82°-102.83° E; 海拔3 400 m左右, 年平均气温为1.1 ℃.年降水量600-800 mm, 雨季温暖湿润(5-9月), 降水量占全年的90%, 旱季寒冷干燥(10月至次年4月).研究区域土壤类型主要为草甸土, 植被类型以高山草甸植被为主, 主要优势种有木里薹草(
Carex muliensis )、矮生嵩草(
Kobresia humilis )和二裂委陵菜(
Potentilla bifurca )(
Jin et al ., 2020 ). ...
Soil carbon sequestration to mitigate climate change
1
2004
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
施肥对高寒草甸土壤有机碳激发效应及净碳平衡的影响
3
2019
... 土壤碳矿化速率的测定: 碱液吸收法是测定土壤呼吸的一种常用、简便的方法.青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化; 在室内20 ℃恒温培养条件下用碱液吸收法测得的CO
2 通量值与野外条件下用静态气室法测得的数值非常接近(
Song et al ., 2010 ), 说明该方法是一种简单易行且可靠的研究复杂环境条件下土壤呼吸的方法.因此, 本研究中采用通用的室内20 ℃恒温培养、碱液吸收法测定土壤碳矿化速率.简单过程如下: 相当于70 g干土的鲜土分别放入培养瓶中, 将土壤水分调节至田间持水量的60%, 置于培养箱中20 ℃无光条件下恒温培养, 在第1、2、3、5、7、9、12、15、19、23、27、31天分别测定土壤呼吸速率并计算碳累积矿化量(
李峰, 2019 ). ...
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
... ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
施肥对高寒草甸土壤有机碳激发效应及净碳平衡的影响
3
2019
... 土壤碳矿化速率的测定: 碱液吸收法是测定土壤呼吸的一种常用、简便的方法.青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化; 在室内20 ℃恒温培养条件下用碱液吸收法测得的CO
2 通量值与野外条件下用静态气室法测得的数值非常接近(
Song et al ., 2010 ), 说明该方法是一种简单易行且可靠的研究复杂环境条件下土壤呼吸的方法.因此, 本研究中采用通用的室内20 ℃恒温培养、碱液吸收法测定土壤碳矿化速率.简单过程如下: 相当于70 g干土的鲜土分别放入培养瓶中, 将土壤水分调节至田间持水量的60%, 置于培养箱中20 ℃无光条件下恒温培养, 在第1、2、3、5、7、9、12、15、19、23、27、31天分别测定土壤呼吸速率并计算碳累积矿化量(
李峰, 2019 ). ...
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
... ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
Fertilization with nitrogen and/or phosphorus lowers soil organic carbon sequestration in alpine meadows
2
2018
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
... ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
Effect of nitrogen and phosphorus additions on decomposition and accumulation of soil organic carbon in alpine meadows on the Tibetan Plateau
3
2021
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
... ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
... ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
Coupled impact of decadal precipitation and evapotranspiration on peatland degradation in the Zoige basin, China
1
2020
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
不同程度退化草地生物量的分布模式
3
2005
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... AM阶段微生物活性高, SOC分解量大, 但有机碳潜在输入量也高, 且输入量大于输出量, 使有机碳潜在积累量较高(
图4 ), 因此该阶段SOC总量最高; SD和HD阶段微生物活性减弱, SOC分解减少, 但是有机质输入量也减少, 导致有机碳潜在积累量降低(
图4 ), 造成SOC总量变小.从AM到HD阶段,
SWC 降低导致植物类群和生物量改变(
图2 ,
图3 )(
刘伟等, 2005 ;
唐明坤等, 2011 )、微生物生物量碳氮含量降低(
图1 ), 导致碳输入显著降低、碳输出显著减弱(与AM相比, SD和HD阶段有机碳潜在输入量分别降低了56和192 g·m
-2 , 潜在输出量分别降低了48和170 g·m
-2 ), 碳输入降低的量略大于碳输出降低的量, 但碳输入、输出的降幅基本相当(
图4 ).净碳积累量AM > SD >> HD, 该结果表明: 与未退化的高寒草甸相比, 重度退化高寒草甸土壤碳积累大大降低, 造成退化梯度上SOC含量降低.因此, 若尔盖高寒草甸退化过程中, 植物碳潜在输入量的减少和微生物分解作用的减弱共同影响SOC含量的变化, 但植物碳潜在输入量的减少是SOC含量降低的主导过程. ...
不同程度退化草地生物量的分布模式
3
2005
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... AM阶段微生物活性高, SOC分解量大, 但有机碳潜在输入量也高, 且输入量大于输出量, 使有机碳潜在积累量较高(
图4 ), 因此该阶段SOC总量最高; SD和HD阶段微生物活性减弱, SOC分解减少, 但是有机质输入量也减少, 导致有机碳潜在积累量降低(
图4 ), 造成SOC总量变小.从AM到HD阶段,
SWC 降低导致植物类群和生物量改变(
图2 ,
图3 )(
刘伟等, 2005 ;
唐明坤等, 2011 )、微生物生物量碳氮含量降低(
图1 ), 导致碳输入显著降低、碳输出显著减弱(与AM相比, SD和HD阶段有机碳潜在输入量分别降低了56和192 g·m
-2 , 潜在输出量分别降低了48和170 g·m
-2 ), 碳输入降低的量略大于碳输出降低的量, 但碳输入、输出的降幅基本相当(
图4 ).净碳积累量AM > SD >> HD, 该结果表明: 与未退化的高寒草甸相比, 重度退化高寒草甸土壤碳积累大大降低, 造成退化梯度上SOC含量降低.因此, 若尔盖高寒草甸退化过程中, 植物碳潜在输入量的减少和微生物分解作用的减弱共同影响SOC含量的变化, 但植物碳潜在输入量的减少是SOC含量降低的主导过程. ...
若尔盖高原不同退化程度草地植物种群生态位特征
3
2019
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
若尔盖高原不同退化程度草地植物种群生态位特征
3
2019
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
退化高寒草甸植物功能群与土壤因子关系的冗余分析
5
2018
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... 植物功能群的优势度用重要值表示, 计算如下: 重要值= (相对盖度+相对高度+相对生物量)/3 (
刘育红等, 2018 ). ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
退化高寒草甸植物功能群与土壤因子关系的冗余分析
5
2018
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... 植物功能群的优势度用重要值表示, 计算如下: 重要值= (相对盖度+相对高度+相对生物量)/3 (
刘育红等, 2018 ). ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
湿地土壤微生物群落结构和功能对水分条件变化的响应
1
2017
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
湿地土壤微生物群落结构和功能对水分条件变化的响应
1
2017
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
Soil carbon stocks and quality across intact and degraded alpine wetlands in Zoige, east Qinghai-Tibet Plateau
4
2014
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
Redox control of phosphorus pools in Hawaiian montane forest soils
3
2001
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
Soil respiration and its
Q 10 response to various grazing systems of a typical steppe in Inner Mongolia, China
1
2019
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
影响土壤磷有效性变化作用机理
2
2006
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... ;
秦胜金等, 2006 ). ...
影响土壤磷有效性变化作用机理
2
2006
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... ;
秦胜金等, 2006 ). ...
Analysis of dynamics and driving factors of wetland landscape in Zoige, Eastern Qinghai-Tibetan Plateau
1
2009
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
Effects of temperature, glucose and inorganic nitrogen inputs on carbon mineralization in a Tibetan alpine meadow soil
3
2010
... 土壤碳矿化速率的测定: 碱液吸收法是测定土壤呼吸的一种常用、简便的方法.青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化; 在室内20 ℃恒温培养条件下用碱液吸收法测得的CO
2 通量值与野外条件下用静态气室法测得的数值非常接近(
Song et al ., 2010 ), 说明该方法是一种简单易行且可靠的研究复杂环境条件下土壤呼吸的方法.因此, 本研究中采用通用的室内20 ℃恒温培养、碱液吸收法测定土壤碳矿化速率.简单过程如下: 相当于70 g干土的鲜土分别放入培养瓶中, 将土壤水分调节至田间持水量的60%, 置于培养箱中20 ℃无光条件下恒温培养, 在第1、2、3、5、7、9、12、15、19、23、27、31天分别测定土壤呼吸速率并计算碳累积矿化量(
李峰, 2019 ). ...
... 需要指出的是: 本研究中通过估算植物碳输入和土壤碳矿化探讨了碳输入和输出过程对SOC含量的影响, 估算过程比较粗略且存在误差.例如, 在估算植物碳输入时, 我们忽略了凋落物自身的分解, 因为植物凋落物(特别是地上凋落物)有一部分会发生分解而没有输入到土壤中, 但分解的这部分目前在野外条件下很难估算, 只有通过同位素标记才可定量凋落物分解的量和截留在土壤中的凋落物量.目前, 我们已在青藏高原高寒草甸开展了一些相关研究: 有凋落物输入时土壤呼吸排放大于没有凋落物的土壤呼吸排放(
Li et al ., 2018 ); 进一步用标记葡萄糖和香草醛模拟植物凋落物碳时, 这些碳的输入会促进高寒草甸土壤有机碳的分解, 即产生正激发效应(
李峰, 2019 ;
Li et al ., 2021 ).在32天的培养时间里, 输入的葡萄糖-碳分解了3%, 大约97%截留在了土壤中; 香草醛-碳分解了14%, 大约86%截留在了土壤中; 凋落物碳的截留量大于土壤碳的分解量, 两种碳源的输入均产生净碳积累(
Li et al ., 2021 ).植物凋落物和根系分泌物的输入促进土壤有机碳的分解(
Li et al ., 2018 ,
2021 ;
李峰, 2019 ), 减少土壤碳的累积.此外, 根系分泌物碳输入量为0.6和1.2 mg·g
-1 时, 凋落物分解分别加速了26%和19% (
胡凯等, 2020 ).另外, 本研究中估算碳输出是基于20 ℃恒温培养条件下的矿化量, 这种培养方式和培养温度也可能高估土壤碳输出.不可否认, 不同温度下呼吸速率(例如本文的矿化速率)变化很大; 特别是在青藏高原, 已有的研究和我们自己的实验均已表明: 随着温度的升高, 微生物呼吸速率增加(
Song et al ., 2010 ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
... ).青藏高原气候变化莫测, 高寒草甸地温大致在-10-20 ℃变化(
Song et al ., 2010 ), 即便我们用变温培养, 也无法真正模拟青藏高原气温变化的实际情况.因此, 该研究中我们采用了通用的20 ℃恒温培养和最经典的碱液吸收法测定土壤碳矿化速率, 并基于该培养条件下的累计矿化量, 根据研究地区的月平均气温和
Q 10 估算全年各月份的土壤碳累积矿化量.虽然此方法有误差, 但该方法也是我们了解和模拟自然环境的方法之一, 并且被一些研究者所采纳并运用(
黄晓宇等, 2016 ).该方法产生的误差属于系统误差, 对试验结果的影响是同向的(同时偏大或偏小), 但不会改变各个退化阶段的变化趋势.上述研究表明, 本研究仅仅依据植物生物量和矿化量来估算植物碳输入量和土壤碳输出量, 可能高估碳输入量而低估碳输出量.因此, 今后的研究中, 需要更加精确的方法来定量草甸生态系统的碳输入和输出过程, 从而进一步揭示若尔盖高寒草甸退化过程中SOC含量降低的机理. ...
Temperature and moisture sensitivities of CO
2 efflux from lowland and alpine meadow soils
4
2009
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
... ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
... ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
四川若尔盖县不同退化程度高寒草地群落比较研究
7
2011
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... AM阶段微生物活性高, SOC分解量大, 但有机碳潜在输入量也高, 且输入量大于输出量, 使有机碳潜在积累量较高(
图4 ), 因此该阶段SOC总量最高; SD和HD阶段微生物活性减弱, SOC分解减少, 但是有机质输入量也减少, 导致有机碳潜在积累量降低(
图4 ), 造成SOC总量变小.从AM到HD阶段,
SWC 降低导致植物类群和生物量改变(
图2 ,
图3 )(
刘伟等, 2005 ;
唐明坤等, 2011 )、微生物生物量碳氮含量降低(
图1 ), 导致碳输入显著降低、碳输出显著减弱(与AM相比, SD和HD阶段有机碳潜在输入量分别降低了56和192 g·m
-2 , 潜在输出量分别降低了48和170 g·m
-2 ), 碳输入降低的量略大于碳输出降低的量, 但碳输入、输出的降幅基本相当(
图4 ).净碳积累量AM > SD >> HD, 该结果表明: 与未退化的高寒草甸相比, 重度退化高寒草甸土壤碳积累大大降低, 造成退化梯度上SOC含量降低.因此, 若尔盖高寒草甸退化过程中, 植物碳潜在输入量的减少和微生物分解作用的减弱共同影响SOC含量的变化, 但植物碳潜在输入量的减少是SOC含量降低的主导过程. ...
四川若尔盖县不同退化程度高寒草地群落比较研究
7
2011
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... AM阶段微生物活性高, SOC分解量大, 但有机碳潜在输入量也高, 且输入量大于输出量, 使有机碳潜在积累量较高(
图4 ), 因此该阶段SOC总量最高; SD和HD阶段微生物活性减弱, SOC分解减少, 但是有机质输入量也减少, 导致有机碳潜在积累量降低(
图4 ), 造成SOC总量变小.从AM到HD阶段,
SWC 降低导致植物类群和生物量改变(
图2 ,
图3 )(
刘伟等, 2005 ;
唐明坤等, 2011 )、微生物生物量碳氮含量降低(
图1 ), 导致碳输入显著降低、碳输出显著减弱(与AM相比, SD和HD阶段有机碳潜在输入量分别降低了56和192 g·m
-2 , 潜在输出量分别降低了48和170 g·m
-2 ), 碳输入降低的量略大于碳输出降低的量, 但碳输入、输出的降幅基本相当(
图4 ).净碳积累量AM > SD >> HD, 该结果表明: 与未退化的高寒草甸相比, 重度退化高寒草甸土壤碳积累大大降低, 造成退化梯度上SOC含量降低.因此, 若尔盖高寒草甸退化过程中, 植物碳潜在输入量的减少和微生物分解作用的减弱共同影响SOC含量的变化, 但植物碳潜在输入量的减少是SOC含量降低的主导过程. ...
水分和温度对若尔盖湿地和草甸土壤碳矿化的影响
1
2013
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
水分和温度对若尔盖湿地和草甸土壤碳矿化的影响
1
2013
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
青藏高原高寒草原生态系统土壤碳氮比的分布特征
1
2014
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
青藏高原高寒草原生态系统土壤碳氮比的分布特征
1
2014
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
Response of plant functional groups to the de-gradational succession of alpine meadow in Zoigê County
2014
若尔盖高寒草甸不同功能群植物数量特征对退化演替的响应
3
2014
... SOC的输入主要来自植物的茎、叶和根的碎屑、根系分泌物, 因此植物群落组成和生物量对SOC的输入有重要影响(
Cong et al ., 2016 ).随着退化程度的加深, 莎草科优势度减弱, 杂类草优势度逐渐变大, 到HD阶段杂类草已成为优势功能群(
图2 ).这与
唐明坤等(2011) 和
刘学敏等(2019) 的研究结果一致, 但与
吴鹏飞等(2014) 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... 的结果不一致; 不一致的原因是阶段划分依据与采样时间不同, 造成了植物生长情况不一样(
吴鹏飞等, 2014 ).但上述结果中重度退化阶段与之前的阶段相比, 均呈现莎草科和禾本科优势度变弱、杂类草优势度增强、地上生物量显著下降(
唐明坤等, 2011 ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
... ;
吴鹏飞等, 2014 ;
刘学敏等, 2019 ).
SWC 与禾本科、莎草科和SOC含量的相关性结果(
图3A )表明: 土壤含水量低不适合禾本科和莎草科的生长; 而杂类草更适宜在容重较大, 土壤含水量较低的环境生长(
刘学敏等, 2019 ).在退化梯度上杂类草优势度越来越大, 但其生物量不足以补偿莎草和禾草下降的部分(
刘伟等, 2005 ), 因此随着退化程度的加深, 植物生物量(包括地上生物量和根系生物量)逐渐降低, 造成了有机碳潜在输入量减少. ...
青藏高原高寒草地土壤碳矿化及其温度敏感性
1
2013
... 式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度. ...
青藏高原高寒草地土壤碳矿化及其温度敏感性
1
2013
... 式中,
CCMn 为各月份的土壤碳累积矿化量(g·kg
-1 );
CCM 0 为室内实验的土壤碳累积矿化量(g·kg
-1 );
Q 10 为呼吸温度敏感性(
黄晓宇等(2016) 的研究中
Q 10 为3.3, 低于青藏高原土壤呼吸的
Q 10 的研究(4.7)(
常宗强等, 2005 ;
徐丽等, 2013 ), 本研究中
Q 10 取二者平均值为4);
Tn 为各月平均气温(在
http://www.tianqihoubao.com 上查询了若尔盖2018年全年的气温, 计算出每个月份的平均气温);
T 0 为室内实验培养温度. ...
Qinghai-Tibetan Plateau peatland sustainable. utilization under anthropogenic disturbances and climate change
2017
Vertical distribution and influencing factors of soil organic carbon in the Loess Plateau, China
3
2019
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
... 本研究结果显示, 若尔盖湿地高寒草甸从AM到HD的退化梯度上,
SWC 、SOC含量和TN含量显著降低(
图1 ), 且SOC含量或TN含量与
SWC 显著正相关(
图3A ), 这与前人的研究结果(
Cong et al ., 2016 ;
刘育红等, 2018 )一致.多种原因造成了这种结果: 若尔盖湿地退化的主要原因是人为排水疏干(
唐明坤等, 2011 ), 其引起的水文条件的变化必然导致土壤理化性质的改变(
Guo et al ., 2013 ;
Yu et al ., 2019 ), 主要表现为土壤沙砾含量增加、土壤对有机质的物理保护作用减弱(
Dlamini et al ., 2014 ); 其次, 土壤养分有效性和阳离子交换能力降低, 造成不适合原有优势种生长的环境, 植物类群发生改变, 且植物生物量降低(
刘伟等, 2005 ), 导致土壤有机碳潜在输入量和积累量降低; 此外, 水分的丧失使土壤中的营养物质流失, 造成SOC和TN含量显著降低(
Dlamini et al ., 2014 ).与AM阶段相比, SD和HD阶段TP含量都增加, 水分条件的改变造成铁铝氧化物含量的变化可能是土壤全磷含量变化的重要原因(
Miller et al ., 2001 ;
秦胜金等, 2006 ).已有研究表明: 土壤水分的变化会改变土壤的氧化还原条件, 影响土壤中的铁铝氧化物含量, 而铁铝氧化物与土壤磷的吸附和释放过程直接相关(
Miller et al ., 2001 ), 但该过程的响应机制十分复杂, 目前研究结果不一致, 甚至出现了截然相反的结果(
Miller et al ., 2001 ;
秦胜金等, 2006 ). ...
... SOC的输出主要依赖于土壤的呼吸作用, SOC含量高并不一定导致土壤呼吸速率高; 高寒草甸土壤呼吸的
Q 10 受到放牧强度、SOC含量和质量、土壤湿度、温度等众多因素的影响(
Luan et al ., 2014 ;
Nie et al ., 2019 ;
Suh et al ., 2019 ).
Suh等(2009) 发现土壤呼吸速率在最适温度下受到土壤水分的强烈控制, 土壤湿度降低会导致土壤呼吸速率下降.我们的矿化结果表明: 随着退化程度的加深, SOC分解减弱(
图4 );
SWC 显著影响土壤碳累积矿化量(
图3B ), 这与其他****的研究结果(
Suh et al ., 2009 ;
王丹等, 2013 )一致.土壤含水量影响土壤微生物群落的活动, 改变底物向细胞表面流转的通量, 从而使碳周转速率发生变化, 影响SOC分解(
Cong et al ., 2016 ); 土壤含水量较低时虽然有利于气体的交换, 但是会减少土壤微生物生物量(
MBC 从AM阶段的1 256 mg·kg
-1 降低到HD阶段的440 mg·kg
-1 )、降低土壤微生物和酶的活性, 造成SOC分解减弱(
黄晓宇等, 2016 ;
Yu et al ., 2019 ).在退化梯度上, C:N增加, 碳累积矿化量降低(
图1 ,
图3B ), 表明随着草甸的退化, 微生物在分解SOC时存在的氮限制越来越严重(
王建林等, 2014 ).因此, 在退化梯度上
SWC 的降低和氮限制状况的加剧引起微生物量的减少和微生物活性的降低, 从而导致SOC分解减弱. ...
青海省不同高寒草地土壤主要养分及可溶性有机碳特性研究
1
2019
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
青海省不同高寒草地土壤主要养分及可溶性有机碳特性研究
1
2019
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
青藏高原东部山地草地土壤碳、氮、磷元素计量特征
2
2018
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
青藏高原东部山地草地土壤碳、氮、磷元素计量特征
2
2018
... 草地退化显著降低SOC含量(
Dong et al ., 2012 ;
Dlamini et al ., 2014 ;
赵云飞等, 2018 ).随着高寒草甸退化程度的加剧, SOC含量、氮磷含量、含水量等呈下降趋势; 植物功能群分布对土壤含水量、全氮含量等因子的变化更敏感(
刘育红等, 2018 ).高寒草甸退化后, 地上植物丰富度和植被盖度的变化导致土壤中凋落物和根系分泌物不同, 进而影响微生物对土壤碳氮的转化过程和转化速率(
张苗苗等, 2019 ).此外, 土壤团聚体通过包含和吸附等一系列相互作用对SOC起到了物理保护作用; 而草地退化引起的地上植被覆盖度的降低, 会增加降水造成的水土流失, 使土壤团聚体结构破坏, 导致SOC流失(
Dlamini et al ., 2014 ). ...
... 若尔盖湿地位于青藏高原东部边缘, 是世界上最大的高原泥炭沼泽型湿地(
Jiang et al ., 2017 ).但是, 从20世纪70年代开始, 由于气候变化、放牧活动和人为排水等因素的影响, 若尔盖湿地发生了严重的退化, 湿地面积减少, 草甸面积增加, 且已发生了不同程度的退化(
Qiu et al ., 2009 ;
唐明坤等2011 ;
Luan et al ., 2014 ;
Li et al ., 2020 ).若尔盖湿地高寒草甸随着退化的加剧, 植物群落盖度、物种丰富度、生物量降低(
唐明坤等, 2011 ), SOC含量显著降低(
刘育红等, 2018 ;
赵云飞等, 2018 ); 挖沟排水可能是该区高寒草甸退化的主要原因(
唐明坤等, 2011 ).但是, 若尔盖湿地高寒草甸退化过程中SOC含量降低的机理仍不明确, 特别是退化过程中碳输入和输出过程以及哪个过程对SOC含量降低的影响占主导作用还不清楚.本文分析了高寒草甸不同退化阶段SOC的输入和输出过程, 以期揭示草甸SOC降低的机制并为退化高寒草甸生态系统的生态恢复提供科学依据.依据前人研究(
Luan et al ., 2014 ;
Cong et al ., 2016 ;
刘育红等, 2018 ), 本文做出如下假设: 随着退化程度的加重, 若尔盖高寒草甸的碳输入显著降低, 碳输出减弱, 但碳输入对SOC的影响占主导作用, 最终导致SOC含量降低. ...
Distribution, stock, and influencing factors of soil organic carbon in an alpine meadow in the hinterland of the Qinghai-Tibetan Plateau
1
2018
... 土壤有机碳(SOC)是指以各种形态和状态存在于土壤中的含碳化合物, 它包括动物、植物和微生物残体以及分解、合成的产物.SOC通过影响土壤结构、肥力和持水能力影响自然生态系统的稳定性和生产力(
Zhu & Shao, 2018 ).由于SOC碳库容量巨大, 且与大气碳库之间有着直接的联系, 土壤碳库微小的改变即可引起大气CO
2 浓度的显著变化(
Lal, 2004 ).因此, SOC含量变化及其驱动力分析对于研究陆地生态系统的管理、区域或全球环境变化和碳循环有非常重要的意义(
Luan et al ., 2014 ;
Yu et al ., 2019 ).SOC含量取决于碳输入和输出之间的收支差; 凋落物的输入和土壤碳的矿化是SOC输入、输出的最主要过程(
Cox et al ., 2000 ).土壤含水量的变化会引起植物群落组成、植被覆盖度和初级生产力、根分泌过程的改变, 进而直接影响凋落物数量, 最终对SOC含量产生影响; 此外, 植物群落还可通过改变凋落物分解环境条件、凋落物基质质量、分解者群落结构与活性等非生物与生物因子而间接作用于SOC输出过程, 从而极大地影响土壤对碳的捕获能力(
Gao et al ., 2019 ).土壤含水量降低导致土壤微生物生物量和微生物呼吸速率降低(
Suh et al ., 2009 ); 而土壤水分增加则引起对水分响应敏感的革兰氏阴性细菌(主要为变形菌门和拟杆菌门)和真菌(主要为子囊菌门)类群的增加, 促进SOC的矿化, 降低SOC含量(
刘亚军, 2017 ;
陈昊, 2018 ). ...
 ,1,*, 陈珊1, 张瑞1, 孙建2, 马妙君1
,1,*, 陈珊1, 张瑞1, 孙建2, 马妙君1

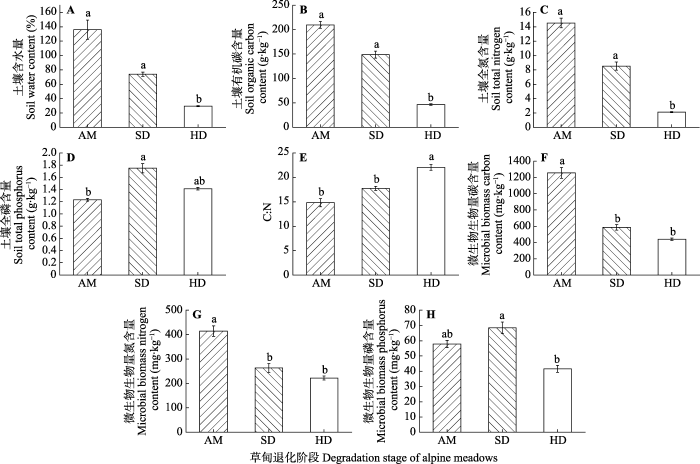 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT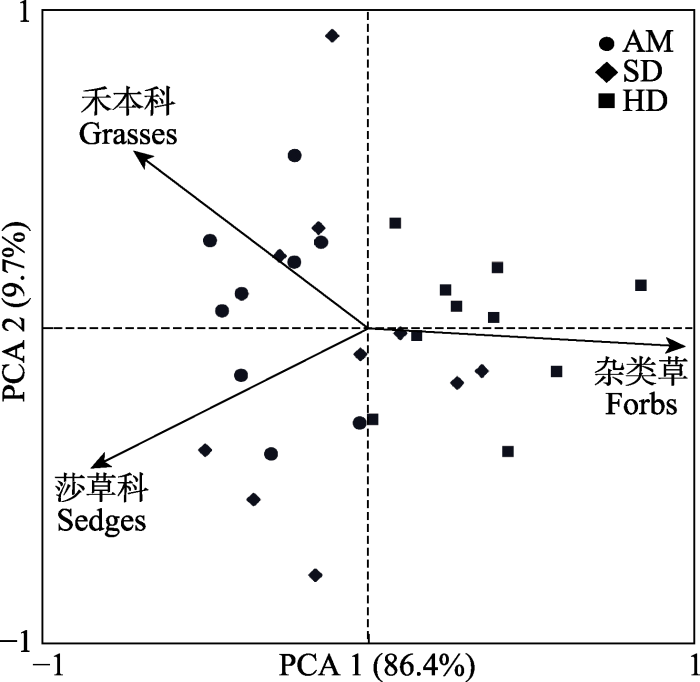 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT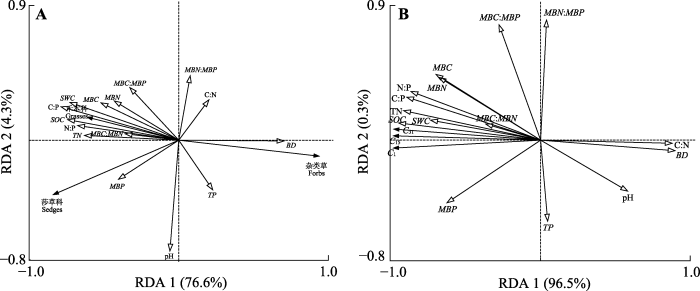 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT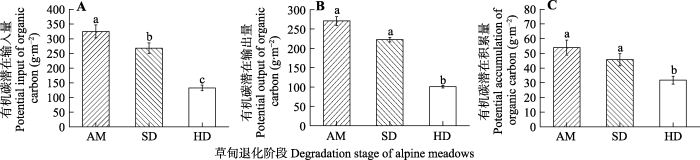 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}