Effects of nitrogen input on carbon cycle and carbon budget in a coastal salt marsh
Guang-Xuan HAN,,*, Juan-Yong LI, Wen-Di QUCAS Key Laboratory of Coastal Environmental Processes and Ecological Remediation, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai, Shandong 264003, China; and University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Coastal salt marshes are an effective blue carbon sink to mitigate climate warming, but their ecosystem stability and carbon sink function are threatened by the large amount of nitrogen input caused by coastal eutrophication. Under the action of regular tides, the high nitrogen content in the coastal waters will have a profound effect on the key processes of carbon cycle such as plant photosynthetic carbon fixation, carbon allocation in plant-soil system, and soil carbon release in the salt marsh. This study reviewed the effects of nitrogen input on plant photosynthetic carbon fixation, carbon allocation in plant-soil system, decomposition of soil organic carbon, formation and release of soil dissolved organic carbon (DOC), and carbon sequestration in the salt marsh. Based on the shortcomings of current research, this review proposed the directions of future research, including the effects of nitrogen input on plant photosynthetic carbon fixation and carbon allocation in plant-soil system, the microbial mechanism of soil organic carbon decomposition, production and lateral exchange of soil DOC, and the potential impact of different forms of nitrogen input on soil carbon sequestration in the salt marsh. Overall, this study aims to improve the understanding of impacts of nitrogen input on the key carbon processes and the mechanisms of carbon sequestration in a salt marsh, and to provide new ideas for assessing the potential changes of carbon pools under the influence of eutrophication of coastal waters in the salt marsh wetlands. Keywords:nitrogen input;carbon cycle;carbon allocation;carbon sink;coastal salt marsh;tidal action
PDF (1423KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 韩广轩, 李隽永, 屈文笛. 氮输入对滨海盐沼湿地碳循环关键过程的影响及机制. 植物生态学报, 2021, 45(4): 321-333. DOI: 10.17521/cjpe.2020.0353 HAN Guang-Xuan, LI Juan-Yong, QU Wen-Di. Effects of nitrogen input on carbon cycle and carbon budget in a coastal salt marsh. Chinese Journal of Plant Ecology, 2021, 45(4): 321-333. DOI: 10.17521/cjpe.2020.0353
Fig. 1Effect of nitrogen (N) input under tidal action on key processes of carbon (C) cycle in a salt marsh. Periodic tides bring N from nearshore eutrophic water into coastal salt marsh ecosystems, changing the stoichiometric relationship of soil nutrient elements in salt marsh wetland. The exogenous N input could have important impacts on the key processes of carbon cycle, such as photosynthesis and respiration of plants, distribution of photosynthetic products, decomposition of soil organic carbon and loss of dissolved organic carbon (DOC). DIC, dissolved inorganic carbon; POC, particulate organic carbon.
Fig. 2Effect of nitrogen (N) input on decomposition of soil organic carbon in a salt marsh (adapted from Hester et al., 2018 ). Microbial processes are directly or indirectly affected by N input. Plants mainly absorb NH4+-N, which stimulates plant productivity and the deposition of organic matter from the rhizosphere, thus stimulating the material circulation of rhizosphere and the metabolism of rhizosphere microorganisms. Plant derived carbon input and higher N utilization jointly stimulate microbial heterotrophic respiration, and then stimulate the production and emission of CO2 in aerobic condition. In anaerobic condition, excessive NO3--N is converted into NH4+-N, which will inhibit CH4 oxidation.
Fig. 3Effect of nitrogen (N) input on production and release of dissolved organic carbon (DOC) in a salt marsh. N input is generally accompanied by DOC input, which affects vegetation growth. It alters DOC content in soil and water through plant litter and microbial biomass, thus also interfering with CO2 and CH4 emissions and DOC output.
5 氮输入对盐沼湿地土壤碳库的影响
滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 )。结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 )。据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统。与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性。研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 )。氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011)。为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1)。评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的。已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 )。同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入。如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 )。准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义。
Table 1 表1 表1氮(N)输入对盐沼湿地及其他湿地类型土壤碳库的影响 Table 1Effect of nitrogen (N) input on carbon sink function of wetland ecosystems
AngelR, ClausP, ConradR (2012). Methanogenic archaea are globally ubiquitous in aerated soils and become active under wet anoxic conditions The ISME Journal, 6, 847-862. DOI:10.1038/ismej.2011.141URL [本文引用: 1]
AnisfeldSC, HillTD (2012). Fertilization effects on elevation change and belowground carbon balance in a long island sound tidal marsh Estuaries and Coasts, 35, 201-211. DOI:10.1007/s12237-011-9440-4URL
ArmitageAR, FourqureanJW (2016). Carbon storage in seagrass soils: long-term nutrient history exceeds the effects of near-term nutrient enrichment Biogeosciences, 13, 313-321. DOI:10.5194/bg-13-313-2016URL [本文引用: 1]
BauerJE, CaiWJ, RaymondPA, BianchiTS, HopkinsonCS, RegnierPAG (2013). The changing carbon cycle of the coastal ocean Nature, 504, 61-70. DOI:10.1038/nature12857URL [本文引用: 1]
BolinderMA, KättererT, AndrénO, ParentLE (2012). Estimating carbon inputs to soil in forage-based crop rotations and modeling the effects on soil carbon dynamics in a Swedish long-term field experiment Canadian Journal of Soil Science, 92, 821-833. DOI:10.4141/cjss2012-036URL [本文引用: 1]
BragazzaL, FreemanC, JonesT, RydinH, LimpensJ, FennerN, EllisT, GerdolR, HájekM, HájekT, IacuminP, KutnarL, TahvanainenT, TobermanH (2006). Atmospheric nitrogen deposition promotes carbon loss from peat bogs Proceedings of the National Academy of Sciences of the United States of America, 103, 19386-19389. PMID:17151199 [本文引用: 2] Peat bogs have historically represented exceptional carbon (C) sinks because of their extremely low decomposition rates and consequent accumulation of plant remnants as peat. Among the factors favoring that peat accumulation, a major role is played by the chemical quality of plant litter itself, which is poor in nutrients and characterized by polyphenols with a strong inhibitory effect on microbial breakdown. Because bogs receive their nutrient supply solely from atmospheric deposition, the global increase of atmospheric nitrogen (N) inputs as a consequence of human activities could potentially alter the litter chemistry with important, but still unknown, effects on their C balance. Here we present data showing the decomposition rates of recently formed litter peat samples collected in nine European countries under a natural gradient of atmospheric N deposition from approximately 0.2 to 2 g.m(-2).yr(-1). We found that enhanced decomposition rates for material accumulated under higher atmospheric N supplies resulted in higher carbon dioxide (CO2) emissions and dissolved organic carbon release. The increased N availability favored microbial decomposition (i) by removing N constraints on microbial metabolism and (ii) through a chemical amelioration of litter peat quality with a positive feedback on microbial enzymatic activity. Although some uncertainty remains about whether decay-resistant Sphagnum will continue to dominate litter peat, our data indicate that, even without such changes, increased N deposition poses a serious risk to our valuable peatland C sinks.
BubierJL, MooreTR, BledzkiLA (2007). Effects of nutrient addition on vegetation and carbon cycling in an ombrotrophic bog Global Change Biology, 13, 1168-1186. DOI:10.1111/gcb.2007.13.issue-6URL [本文引用: 2]
Chapin IIIFS, MatsonPA, MooneyHA (2011). Carbon input to terrestrial ecosystems//Chapin III FS, Matson PA, Mooney HA. Principles of Terrestrial Ecosystem Ecology. Springer New York. [本文引用: 1]
ChenDM, LanZC, HuSJ, BaiYF (2015a). Effects of nitrogen enrichment on belowground communities in grassland: relative role of soil nitrogen availability vs. soil acidification Soil Biology & Biochemistry, 89, 99-108. DOI:10.1016/j.soilbio.2015.06.028URL [本文引用: 1]
ChenH, LiD, GurmesaGA, YuG, LiL, ZhangW, FangH, MoJ (2015b). Effects of nitrogen deposition on carbon cycle in terrestrial ecosystems of China: a meta-analysis Environmental Pollution, 206, 352-360. DOI:10.1016/j.envpol.2015.07.033URL [本文引用: 1]
ChenXP, WangGX, ZhangT, MaoTX, WeiD, SongCL, HuZY, HuangKW (2017). Effects of warming and nitrogen fertilization on GHG flux in an alpine swamp meadow of a permafrost region Science of the Total Environment, 601- 602, 1389-1399. [本文引用: 5]
ChenZM, XuYL, HeYJ, ZhouXH, FanJL, YuHY, DingWX (2018). Nitrogen fertilization stimulated soil heterotrophic but not autotrophic respiration in cropland soils: a greater role of organic over inorganic fertilizer Soil Biology & Biochemistry, 116, 253-264. DOI:10.1016/j.soilbio.2017.10.029URL [本文引用: 1]
CloughBF (1992). Primary productivity and growth of mangrove forests//Robertson AI, Alongi DM Tropical Mangrove Ecosystems. The American Geophysical Union, Washington D. C. 225-249. [本文引用: 1]
CotrufoMF, WallensteinMD, BootCM, DenefK, PaulE (2013). The Microbial Efficiency-Matrix Stabilization (MEMS) framework integrates plant litter decomposition with soil organic matter stabilization: Do labile plant inputs form stable soil organic matter? Global Change Biology, 19, 988-995. DOI:10.1111/gcb.12113URL [本文引用: 1]
DeeganLA, JohnsonDS, WarrenRS, PetersonBJ, FleegerJW, FagherazziS, WollheimWM (2012). Coastal eutrophication as a driver of salt marsh loss Nature, 490, 388-392. DOI:10.1038/nature11533URL [本文引用: 7]
DuE, TerrerC, PellegriniAFA, AhlströmA, van LissaCJ, ZhaoX, XiaN, WuX, JacksonRB (2020). Global patterns of terrestrial nitrogen and phosphorus limitation Nature Geoscience, 13, 221-226. DOI:10.1038/s41561-019-0530-4URL [本文引用: 1]
DuarteCM, MiddelburgJJ, CaracoN (2005). Major role of marine vegetation on the oceanic carbon cycle Biogeosciences, 2, 1-8. DOI:10.5194/bg-2-1-2005URL [本文引用: 2]
FangC, YeJS, GongY, PeiJ, YuanZ, XieC, ZhuY, YuY (2017). Seasonal responses of soil respiration to warming and nitrogen addition in a semi-arid alfalfa-pasture of the Loess Plateau, China Science of the Total Environment, 590-591, 729-738. [本文引用: 2]
FellmanJB, D’AmoreDV, HoodE, CunninghamP (2017). Vulnerability of wetland soil carbon stocks to climate warming in the perhumid coastal temperate rainforest Biogeochemistry, 133, 165-179. DOI:10.1007/s10533-017-0324-yURL [本文引用: 1]
Fernández-MartínezM, ViccaS, JanssensIA, SardansJ, LuyssaertS, CampioliM, Chapin IIIFS, CiaisP, MalhiY, ObersteinerM, PapaleD, PiaoSL, ReichsteinM, RodàF, PeñuelasJ (2014). Nutrient availability as the key regulator of global forest carbon balance Nature Climate Change, 4, 471-476. DOI:10.1038/nclimate2177URL [本文引用: 1]
GaciaE, DuarteCM, MiddelburgJJ (2002). Carbon and nutrient deposition in a Mediterranean seagrass (Posidonia oceanica) meadow Limnology and Oceanography, 47, 23-32. DOI:10.4319/lo.2002.47.1.0023URL [本文引用: 1]
GrahamSA, MendelssohnIA (2016). Contrasting effects of nutrient enrichment on below-ground biomass in coastal wetlands Journal of Ecology, 104, 249-260. DOI:10.1111/1365-2745.12498URL [本文引用: 1]
GuerrieriR, MencucciniM, SheppardLJ, SaurerM, PerksMP, LevyP, SuttonMA, BorghettiM, GraceJ (2011). The legacy of enhanced N and S deposition as revealed by the combined analysis of δ 13C, δ 18O and δ 15N in tree rings Global Change Biology, 17, 1946-1962. DOI:10.1111/j.1365-2486.2010.02362.xURL [本文引用: 1]
HanG, XingQ, YuJ, LuoY, LiD, YangL, WangG, MaoP, XieB, MikleN (2014). Agricultural reclamation effects on ecosystem CO2 exchange of a coastal wetland in the Yellow River Delta Agriculture Ecosystems & Environment, 196, 187-198. DOI:10.1016/j.agee.2013.09.012URL [本文引用: 1]
HanGX ( 2017). Effect of tidal action and drying-wetting cycles on carbon exchange in a salt marsh: progress and prospects Acta Ecologica Sinica, 37, 8170-8178. [本文引用: 6]
HayesMA, JesseA, TabetB, ReefR, KeuskampJA, LovelockCE (2017). The contrasting effects of nutrient enrichment on growth, biomass allocation and decomposition of plant tissue in coastal wetlands Plant and Soil, 416,193-204. DOI:10.1007/s11104-017-3206-0URL [本文引用: 2]
HerbertER, Schubauer-BeriganJP, CraftCB (2020). Effects of 10 yr of nitrogen and phosphorus fertilization on carbon and nutrient cycling in a tidal freshwater marsh Limnology and Oceanography, 65, 1669-1687. DOI:10.1002/lno.v65.8URL [本文引用: 4]
HögbergMN, BrionesMJI, KeelSG, MetcalfeDB, CampbellC, MidwoodAJ, ThorntonB, HurryV, LinderS, NäsholmT, HögbergP (2010). Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest New Phytologist, 187, 485-493. DOI:10.1111/nph.2010.187.issue-2URL [本文引用: 1]
HuY, WangL, FuX, YanJ, WuJ, TsangY, LeY, SunY (2016). Salinity and nutrient contents of tidal water affects soil respiration and carbon sequestration of high and low tidal flats of Jiuduansha wetlands in different ways Science of the Total Environment, 565, 637-648. DOI:10.1016/j.scitotenv.2016.05.004URL [本文引用: 1]
HuxhamM, LangatJ, TamoohF, KennedyH, MencucciniM, SkovMW, KairoJ (2010). Decomposition of mangrove roots: effects of location, nutrients, species identity and mix in a Kenyan forest Estuarine Coastal and Shelf Science, 88, 135-142. DOI:10.1016/j.ecss.2010.03.021URL [本文引用: 1]
IversenCM, BridghamSD, KelloggLE (2010). Scaling plant nitrogen use and uptake efficiencies in response to nutrient addition in peatlands Ecology, 91, 693-707. DOI:10.1890/09-0064.1URL [本文引用: 1]
JonesDL, KiellandK (2012). Amino acid, peptide and protein mineralization dynamics in a taiga forest soil Soil Biology & Biochemistry, 55, 60-69. DOI:10.1016/j.soilbio.2012.06.005URL [本文引用: 1]
JuutinenS, BubierJL, MooreTR (2010). Responses of vegetation and ecosystem CO2 exchange to 9 years of nutrient addition at Mer Bleue bog Ecosystems, 13, 874-887. DOI:10.1007/s10021-010-9361-2URL [本文引用: 1]
KeuskampJA, FellerIC, LaanbroekHJ, VerhoevenJTA, HeftingMM (2015). Short- and long-term effects of nutrient enrichment on microbial exoenzyme activity in mangrove peat Soil Biology & Biochemistry, 81, 38-47. DOI:10.1016/j.soilbio.2014.11.003URL [本文引用: 1]
KibaT, KrappA (2016). Plant nitrogen acquisition under low availability: regulation of uptake and root architecture Plant and Cell Physiology, 57, 707-714. DOI:10.1093/pcp/pcw052URL [本文引用: 1]
KirwanML, Patrick MegonigalJ (2013). Tidal wetland stability in the face of human impacts and sea-level rise Nature, 504, 53-60. DOI:10.1038/nature12856URL [本文引用: 1]
KivimäkiSK, SheppardLJ, LeithID, GraceJ (2013). Long-term enhanced nitrogen deposition increases ecosystem respiration and carbon loss from a Sphagnum bog in the Scottish Borders Environmental and Experimental Botany, 90, 53-61. DOI:10.1016/j.envexpbot.2012.09.003URL
LiW, ZhangH, HuangG, LiuR, ZhaoC, McDowellNG (2020). Effects of nitrogen enrichment on tree carbon allocation: a global synthesis Global Ecology and Biogeography, 29, 573-589. DOI:10.1111/geb.v29.3URL [本文引用: 1]
LiuJ, WuN, WangH, SunJ, PengB, JiangP, BaiE (2016). Nitrogen addition affects chemical compositions of plant tissues, litter and soil organic matter Ecology, 97, 1796-1806. DOI:10.1890/15-1683.1URL [本文引用: 1]
MaTT, LiXW, BaiJH, DingSY, ZhouFW, CuiBS (2019). Four decadesʼ dynamics of coastal blue carbon storage driven by land use/land cover transformation under natural and anthropogenic processes in the Yellow River Delta, China Science of the Total Environment, 655, 741-750. DOI:10.1016/j.scitotenv.2018.11.287URL [本文引用: 1]
MacreadiePI, AntonA, RavenJA, BeaumontN, ConnollyRM, FriessDA, KellewayJJ, KennedyH, KuwaeT, LaveryPS, LovelockCE, SmaleDA, ApostolakiET, AtwoodTB, BaldockJ, et al. (2019). The future of Blue Carbon science Nature Communications, 10, 3998. DOI: https://www.plant-ecology.com/article/2021/1005-264X/10.1038/%20s41467-019-11693-w. PMID:31488846 [本文引用: 2] The term Blue Carbon (BC) was first coined a decade ago to describe the disproportionately large contribution of coastal vegetated ecosystems to global carbon sequestration. The role of BC in climate change mitigation and adaptation has now reached international prominence. To help prioritise future research, we assembled leading experts in the field to agree upon the top-ten pending questions in BC science. Understanding how climate change affects carbon accumulation in mature BC ecosystems and during their restoration was a high priority. Controversial questions included the role of carbonate and macroalgae in BC cycling, and the degree to which greenhouse gases are released following disturbance of BC ecosystems. Scientists seek improved precision of the extent of BC ecosystems; techniques to determine BC provenance; understanding of the factors that influence sequestration in BC ecosystems, with the corresponding value of BC; and the management actions that are effective in enhancing this value. Overall this overview provides a comprehensive road map for the coming decades on future research in BC science.
MajdiH, ÖhrvikJ (2004). Interactive effects of soil warming and fertilization on root production, mortality, and longevity in a Norway spruce stand in Northern Sweden Global Change Biology, 10, 182-188. DOI:10.1111/gcb.2004.10.issue-2URL [本文引用: 1]
ManzoniS, TaylorP, RichterA, PorporatoA, ÅgrenGI (2012). Environmental and stoichiometric controls on microbial carbon-use efficiency in soils New Phytologist, 196, 79-91. DOI:10.1111/nph.2012.196.issue-1URL [本文引用: 1]
MaoQ, LuX, MoH, GundersenP, MoJ (2018). Effects of simulated N deposition on foliar nutrient status, N metabolism and photosynthetic capacity of three dominant understory plant species in a mature tropical forest Science of the Total Environment, 610-611, 555-562. [本文引用: 3]
McLeodE, ChmuraGL, BouillonS, SalmR, BjörkM, DuarteCM, LovelockCE, SchlesingerWH, SillimanBR (2011). A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2 Frontiers in Ecology and the Environment, 9, 552-560. DOI:10.1890/110004URL [本文引用: 3]
MinK, KangH, LeeD (2011). Effects of ammonium and nitrate additions on carbon mineralization in wetland soils Soil Biology & Biochemistry, 43, 2461-2469. DOI:10.1016/j.soilbio.2011.08.019URL [本文引用: 2]
MitschWJ, GosselinkJG (2011). Wetlands.4th ed. John Wiley & Sons, Hobgen USA. [本文引用: 1]
MouXJ, SunZG, WangLL, WangCY (2011). Nitrogen cycle of a typical Suaeda salsa marsh ecosystem in the Yellow River estuary Journal of Environmental Sciences, 23, 958-967. DOI:10.1016/S1001-0742(10)60530-XURL [本文引用: 1]
PardoLH, FennME, GoodaleCL, GeiserLH, DriscollCT, AllenEB, BaronJS, BobbinkR, BowmanWD, ClarkCM, EmmettB, GilliamFS, GreaverTL, HallSJ, LilleskovEA, et al. (2011). Effects of nitrogen deposition and empirical nitrogen critical loads for ecoregions of the United States Ecological Applications, 21, 3049-3082. DOI:10.1890/10-2341.1URL [本文引用: 1]
PastoreMA, MegonigalJP, LangleyJA (2017). Elevated CO2 and nitrogen addition accelerate net carbon gain in a brackish marsh Biogeochemistry, 133, 73-87. DOI:10.1007/s10533-017-0312-2URL [本文引用: 1]
PauschJ, KuzyakovY (2018). Carbon input by roots into the soil: quantification of rhizodeposition from root to ecosystem scale Global Change Biology, 24, 1-12. DOI:10.1111/gcb.2018.24.issue-1URL [本文引用: 1]
PengY, PengZ, ZengX, Houx IIIJH (2019). Effects of nitrogen- phosphorus imbalance on plant biomass production: a global perspective Plant and Soil, 436, 245-252. DOI:10.1007/s11104-018-03927-5URL [本文引用: 5]
PenningsSC (2012). Ecology: the big picture of marsh loss Nature, 490, 352-353. DOI:10.1038/490352aURL [本文引用: 3]
RadabaughKR, MoyerRP, ChappelAR, PowellCE, BociuI, ClarkBC, SmoakJM (2018). Coastal blue carbon assessment of mangroves, salt marshes, and salt barrens in Tampa Bay, Florida, USA Estuaries and Coasts, 41, 1496-1510. DOI:10.1007/s12237-017-0362-7URL [本文引用: 1]
RomeroLM, Smith IIITJ, FourqureanJW (2005). Changes in mass and nutrient content of wood during decomposition in a South Florida mangrove forest Journal of Ecology, 93, 618-631. DOI:10.1111/jec.2005.93.issue-3URL [本文引用: 2]
RouskJ, BrookesPC, BååthE (2010). Investigating the mechanisms for the opposing pH relationships of fungal and bacterial growth in soil Soil Biology & Biochemistry, 42, 926-934. DOI:10.1016/j.soilbio.2010.02.009URL [本文引用: 1]
ShenH, DongS, LiS, XiaoJ, HanY, YangM, ZhangJ, GaoX, XuY, LiY, ZhiY, LiuS, DongQ, ZhouH, YeomansJC (2019). Effects of simulated N deposition on photosynthesis and productivity of key plants from different functional groups of alpine meadow on Qinghai-Tibetan Plateau Environmental Pollution, 251, 731-737. DOI:10.1016/j.envpol.2019.05.045 [本文引用: 1] Nitrogen (N) deposition may alter physiological process of plants in grassland ecosystem. However, little is known about the response mechanism of individual plants in alpine regions to N deposition. We conducted a field experiment, and three treatments including 0 kg Nha(-l) year(-1) (CK), 8 kgNha(-l) year(-1) (Low N), and 72 kg N ha(-1) year(-1) (High N) were established to simulate N deposition in alpine meadow of Qinghai-Tibetan plateau. Our objectives were to determine the influence of N deposition on photosynthesis of different functional types of herbage species in alpine meadow, and finally characterize the links of plant productivity and photosynthesis with soil nutrients. The results showed that responses of alpine plants were species-specific under N deposition. Compared with grass species Agropyron cristatum and forb species Thalictrum aquilegifolium, the sedge species Carex melanantha was much more sensitive to N deposition; a lower N load (8 kgNha(-1)year(-1)) can cause a negative effect on its photosynthesis and productivity. Additionally, N deposition can promote plant N uptake and significantly decreased the C (carbon)/N (nitrogen) ratio. Compared with CK and low N deposition, high N deposition inhibited the photosynthesis and growth of the forb species Thalictrum aquilegifolium and sedge species Carex melanantha. In all three functional types of herbage species, the grass species A. cristatum tended to show a much higher photosynthetic capacity and better growth potential; thus, suggesting that grass species A. cristatum will be a more adaptative alpine plants under N deposition. Our findings suggested that plant photosynthetic responses to N deposition were species-specific, low N deposition was not beneficial for all the herbage species, and N deposition may change plant composition by the differential photosynthetic responses among species in alpine grassland. Plant composition shift to grass-dorminant in alpine regions might be attributed to a much higher photosynthetic potential and N use efficiency of grass species. (C) 2019 Elsevier Ltd.
ShermanRE, FaheyTJ, MartinezP (2003). Spatial patterns of biomass and aboveground net primary productivity in a mangrove ecosystem in the Dominican Republic Ecosystems, 6, 384-398. DOI:10.1007/s10021-002-0191-8URL [本文引用: 1]
ShipleyB, MezianeD (2002). The balanced-growth hypothesis and the allometry of leaf and root biomass allocation Functional Ecology, 16, 326-331. DOI:10.1046/j.1365-2435.2002.00626.xURL [本文引用: 1]
SinsabaughRL, BelnapJ, RudgersJ, KuskeCR, MartinezN, SandquistD (2015). Soil microbial responses to nitrogen addition in arid ecosystems Frontiers in Microbiology, 6, 819. DOI: https://www.plant-ecology.com/article/2021/1005-264X/10.3389/fmicb.2015.00819. PMID:26322030 [本文引用: 1] The N cycle of arid ecosystems is influenced by low soil organic matter, high soil pH, and extremes in water potential and temperature that lead to open canopies and development of biological soil crusts (biocrusts). We investigated the effects of N amendment on soil microbial dynamics in a Larrea tridentata-Ambrosia dumosa shrubland site in southern Nevada USA. Sites were fertilized with a NO3-NH4 mix at 0, 7, and 15 kg N ha(-1) y(-1) from March 2012 to March 2013. In March 2013, biocrust (0-0.5 cm) and bulk soils (0-10 cm) were collected beneath Ambrosia canopies and in the interspaces between plants. Biomass responses were assessed as bacterial and fungal SSU rRNA gene copy number and chlorophyll a concentration. Metabolic responses were measured by five ecoenzyme activities and rates of N transformation. By most measures, nutrient availability, microbial biomass, and process rates were greater in soils beneath the shrub canopy compared to the interspace between plants, and greater in the surface biocrust horizon compared to the deeper 10 cm soil profile. Most measures responded positively to experimental N addition. Effect sizes were generally greater for bulk soil than biocrust. Results were incorporated into a meta-analysis of arid ecosystem responses to N amendment that included data from 14 other studies. Effect sizes were calculated for biomass and metabolic responses. Regressions of effect sizes, calculated for biomass, and metabolic responses, showed similar trends in relation to N application rate and N load (rate x duration). The critical points separating positive from negative treatment effects were 88 kg ha(-1) y(-1) and 159 kg ha(-1), respectively, for biomass, and 70 kg ha(-1) y(-1) and 114 kg ha(-1), respectively, for metabolism. These critical values are comparable to those for microbial biomass, decomposition rates and respiration reported in broader meta-analyses of N amendment effects in mesic ecosystems. However, large effect sizes at low N addition rates indicate that arid ecosystems are sensitive to modest increments in anthropogenic N deposition.
SinsabaughRL, MoorheadDL (1994). Resource allocation to extracellular enzyme production: a model for nitrogen and phosphorus control of litter decomposition Soil Biology & Biochemistry, 26, 1305-1311. DOI:10.1016/0038-0717(94)90211-9URL [本文引用: 1]
SmithMD, La PierreKJ, CollinsSL, KnappAK, GrossKL, BarrettJE, FreySD, GoughL, MillerRJ, MorrisJT, RustadLE, YarieJ (2015). Global environmental change and the nature of aboveground net primary productivity responses: insights from long-term experiments Oecologia, 177, 935-947. DOI:10.1007/s00442-015-3230-9URL [本文引用: 1]
SongCC, LiuDY, YangGS, SongYY, MaoR (2011). Effect of nitrogen addition on decomposition of Calamagrostis angustifolia litters from freshwater marshes of Northeast China Ecological Engineering, 37, 1578-1582. DOI:10.1016/j.ecoleng.2011.03.036URL
SongCC, WangLL, TianHQ, LiuDY, LuCQ, XuXF, ZhangLH, YangGS, WanZM (2013). Effect of continued nitrogen enrichment on greenhouse gas emissions from a wetland ecosystem in the Sanjiang Plain, Northeast China: a 5 year nitrogen addition experiment Journal of Geophysical Research, 118, 741-751. [本文引用: 1]
SongMH, JiangJ, CaoGM, XuXL (2010). Effects of temperature, glucose and inorganic nitrogen inputs on carbon mineralization in a Tibetan alpine meadow soil European Journal of Soil Biology, 46, 375-380. DOI:10.1016/j.ejsobi.2010.09.003URL [本文引用: 1]
TaoBX, LiuCY, ZhangBH, DongJ (2018). Effects of inorganic and organic nitrogen additions on CO2 emissions in the coastal wetlands of the Yellow River Delta, China Atmospheric Environment, 185, 159-167. DOI:10.1016/j.atmosenv.2018.05.009URL [本文引用: 1]
TippingE, RoweEC, EvansCD, MillsRTE, EmmettBA, ChaplowJS, HallJR (2012). N14C: a plant-soil nitrogen and carbon cycling model to simulate terrestrial ecosystem responses to atmospheric nitrogen deposition Ecological Modelling, 247, 11-26. DOI:10.1016/j.ecolmodel.2012.08.002URL [本文引用: 1]
van WijnenHJ, BakkerJP (1999). Nitrogen and phosphorus limitation in a coastal barrier salt marsh: the implications for vegetation succession Journal of Ecology, 87, 265-272. DOI:10.1046/j.1365-2745.1999.00349.xURL
VitousekPM, HättenschwilerS, OlanderL, AllisonS (2002). Nitrogen and nature AMBIO: A Journal of the Human Environment, 31, 97-101. PMID:12078015 [本文引用: 1] Anthropogenic changes to the global N cycle are important in part because added N alters the composition, productivity, and other properties of many natural ecosystems substantially. Why does added N have such a large impact? Why is N in short supply in so many natural ecosystems? Processes that slow the cycling of N relative to other elements and processes that control ecosystem-level inputs and outputs of N could cause N supply to limit the dynamics of ecosystems. We discuss stoichiometric differences between terrestrial plants and other organisms, the abundance of protein-precipitating plant defenses, and the nature of the C-N bond in soil organic matter as factors that can slow N cycling. For inputs, the energetic costs of N fixation and their consequences, the supply of nutrients other than N, and preferential grazing on N-fixers all could constrain the abundance and/or activity of biological N-fixers. Together these processes drive and sustain N limitation in many natural terrestrial ecosystems.
VivancoL, IrvineIC, MartinyJBH (2015). Nonlinear responses in salt marsh functioning to increased nitrogen addition Ecology, 96, 936-947. PMID:26230015 [本文引用: 3] Salt marshes provide storm protection to shorelines, sequester carbon (C), and mitigate coastal eutrophication. These valuable coastal ecosystems are confronted with increasing nitrogen (N) inputs from anthropogenic sources, such as agricultural runoff, wastewater, and atmospheric deposition. To inform predictions of salt marsh functioning and sustainability in the future, we characterized the response of a variety of plant, microbial, and sediment responses to a seven-level gradient of N addition in three Californian salt marshes after 7 and 14 months of N addition. The marshes showed variable responses to the experimental N gradient that can be grouped as neutral (root biomass, sediment respiration, potential carbon mineralization, and potential net nitrification), linear (increasing methane flux, decreasing potential net N mineralization, and increasing sediment inorganic N), and nonlinear (saturating aboveground plant biomass and leaf N content, and exponentially increasing sediment inorganic and organic N). The three salt marshes showed quantitative differences in most ecosystem properties and processes rates; however, the form of the response curves to N addition were generally consistent across the three marshes, indicating that the responses observed may be applicable to other marshes in the region. Only for sediment properties (inorganic and organic N pool) did the shape of the response differ significantly between marshes. Overall, the study suggests salt marshes are limited in their ability to sequester C and N with future increases in N, even without further losses in marsh area.
WangB, XinM, WeiQ, XieL (2018). A historical overview of coastal eutrophication in the China Seas Marine Pollution Bulletin, 136, 394-400. DOI:10.1016/j.marpolbul.2018.09.044URL [本文引用: 2]
WangJ, GaoY, ZhangY, YangJ, SmithMD, KnappAK, EissenstatDM, HanX (2019). Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe Global Change Biology, 25, 2958-2969. DOI:10.1111/gcb.v25.9URL [本文引用: 6]
WangQ, WangS, HeT, LiuL, WuJ (2014). Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils Soil Biology & Biochemistry, 71, 13-20. DOI:10.1016/j.soilbio.2014.01.004URL [本文引用: 1]
WangTT, ZhuZK, ZhuHH, TangZZ, PangJ, LiBZ, SuYR, GeTD, WuJS ( 2017). Input and distribution of photosynthesized carbon in soil-rice system affected by water management and nitrogen fertilization Environmental Science, 38, 1227-1234. [本文引用: 2]
WendelS, MooreT, BubierJ, BlodauC (2011). Experimental nitrogen, phosphorus, and potassium deposition decreases summer soil temperatures, water contents, and soil CO2 concentrations in a northern bog Biogeosciences, 8, 585-595. DOI:10.5194/bg-8-585-2011URL [本文引用: 1]
WuY, BlodauC, MooreTR, BubierJ, JuutinenS, LarmolaT (2015). Effects of experimental nitrogen deposition on peatland carbon pools and fluxes: a modelling analysis Biogeosciences, 12, 79-101. DOI:10.5194/bg-12-79-2015URL [本文引用: 1]
XiaoLL, XieBH, LiuJC, ZhangHX, HanGX, WangOM, LiuFH (2017). Stimulation of long-term ammonium nitrogen deposition on methanogenesis by Methanocellaceae in a coastal wetland Science of the Total Environment, 595, 337-343. DOI:10.1016/j.scitotenv.2017.03.279URL [本文引用: 2]
XiaoML, ZangHD, LiuSL, YeRZ, ZhuZK, SuYR, WuJS, GeTD (2019). Nitrogen fertilization alters the distribution and fates of photosynthesized carbon in rice-soil systems: a13C-CO2 pulse labeling study Plant and Soil, 445, 101-112. DOI:10.1007/s11104-019-04030-zURL [本文引用: 5]
XuX, SchimelJP, ThorntonPE, SongX, YuanF, GoswamiS (2014). Substrate and environmental controls on microbial assimilation of soil organic carbon: a framework for Earth system models Ecology Letters, 17, 547-555. DOI:10.1111/ele.2014.17.issue-5URL [本文引用: 1]
YangK, ZhuJJ, GuJC, XuS, YuLZ, WangZQ (2018). Effects of continuous nitrogen addition on microbial properties and soil organic matter in a Larix gmeliniiplantation in China Journal of Forestry Research, 29, 85-92. DOI:10.1007/s11676-017-0430-7URL [本文引用: 1]
YaoRJ, YangJS, WangXP, XieWP, ZhengFL, LiHQ, TangC, ZhuH (2021). Response of soil characteristics and bacterial communities to nitrogen fertilization gradients in a coastal salt-affected agroecosystem Land Degradation & Development, 32, 338-353. DOI:10.1002/ldr.v32.1URL [本文引用: 1]
ZhouZH, WangCK, ZhengMH, JiangLF, LuoYQ (2017). Patterns and mechanisms of responses by soil microbial communities to nitrogen addition Soil Biology & Biochemistry, 115, 433-441. DOI:10.1016/j.soilbio.2017.09.015URL [本文引用: 3]
Nitrogen saturation in temperate forest ecosystems: hypotheses revisited 1 1998
... 在氮有效性较高、分解率低的潮汐盐沼湿地, 氮输入一方面可能会促进植被根系生长, 提高土壤有机质输入, 加快有机质积累, 从而短期内大量氮输入可能会提高湿地的碳汇量(Hayes et al., 2017 ).对于森林生态系统而言, 新碳的加入能激发促进土壤老碳的更新循环.氮输入驱动下增加的碳大部分都是新的植物生物量, 而非土壤碳.因此, 氮输入对这类生态系统长期土壤碳汇反而具有消极的影响(Tipping et al., 2012 ; Mills et al., 2014 ).因此, 相比之下, 由于沼泽、泥炭地等生态系统土壤有机质分解受到高湿度和富含难分解化合物的限制, 新输入的植物凋落物等有机质被封存和隔离, 甚至能够在土壤中埋藏成百上千年(Dise, 2009).有研究表明, 在滨海湿地生态系统中, 氮输入促进芦苇(Phragmites australis)生长、提高芦苇生物量, 增加了土壤中的碳含量(Qu et al., 2020 ).另一方面, 氮输入通过影响植被的地下生物量分配进而影响土壤碳库.已有研究表明, 盐沼湿地在氮有效性较高时, 总生物量会增加, 但当氮有效性较低时, 地下部分的生物量分配会相对提高(Clough, 1992; Sherman et al., 2003 ).生态系统氮饱和假说认为, 向氮饱和的生态系统中继续加氮会导致土壤功能的丧失, 例如养分循环和氮固持功能等减弱, 进而对植物生长产生影响(Aber et al., 1998 ). ...
Carbon cycling and storage in mangrove forests 1 2014
The changing carbon cycle of the coastal ocean 1 2013
... 外源氮输入影响盐沼湿地土壤DOC的产生和累积, 进而影响盐沼湿地横向碳流失.盐沼湿地土壤中的碳流失主要有2种方式, 一种是以CO2和CH4等气体形式排放, 另一种是以DOC形式流失.尽管DOC仅占土壤总有机碳的0.04%-0.22%, 但它却是有机碳库中最活跃和不容忽视的组成部分(Bauer et al., 2013 ).例如, 由于受到充沛降雨和周期性的潮汐作用, 横向碳输出量大约占温带滨海湿地总碳输出量的40% (Majidzadeh et al., 2017 ).毫无疑问, 外源氮输入对盐沼湿地土壤DOC的产生和释放具有显著影响(图3).首先, 外源氮输入能促进盐沼植被生长和生物量增加, 而植物凋落物和根系分泌物增加能促进土壤DOC的形成和释放.但是长期氮富集可能导致盐沼湿地退化, 根际激发效应促进了分解, 从而降低了DOC含量(Deegan et al., 2012 ).其次, 氮输入通过影响土壤pH间接影响土壤有机质中DOC释放, 同时氮类型也会改变DOC对氮输入的响应(Chang et al., 2018 ).铵态氮增加通常能降低土壤pH, 抑制土壤中DOC淋溶, 从而减少土壤有机碳的横向流失; 但硝态氮增加则通过提高土壤pH, 从而促进DOC释放(Chang et al., 2018 ; Preston et al., 2020 ).再次, 氮输入增加了土壤微生物生物量, 加快了微生物新陈代谢活动, 从而加速了土壤DOC分解(Fellman et al., 2017 ).然而目前研究多集中在氮输入对盐沼湿地垂直方向碳流失(CO2和CH4)的影响, 忽视了近岸水体富营养化背景下以DOC为主要形式的陆海横向碳交换, 制约着对滨海盐沼湿地碳循环过程的整体理解以及对其碳收支的准确评估. ...
Estimating carbon inputs to soil in forage-based crop rotations and modeling the effects on soil carbon dynamics in a Swedish long-term field experiment 1 2012
... 氮输入不仅从整体上影响植物光合固碳能力, 还能影响植物光合产物在植物-土壤系统中的分配比例.植物地上、地下生物量都是盐沼湿地碳汇的重要组成部分(韩广轩, 2017; Hayes et al., 2017 ).首先, 植物地上部分能够拦截海水中的泥沙及颗粒有机质, 帮助盐沼湿地快速抬升高程, 增大其有机碳的储存空间; 同时, 植物地上部分是植物作为初级生产者固定光合碳的器官, 其生物量大小对滨海盐沼湿地的初级生产力具有直接影响.植物地下部分一方面能够分泌根际沉积物, 是土壤可溶性有机碳等易分解碳的重要来源; 另一方面, 植物地下根系能够锚定沉积物, 防止潮汐作用对有机质的侵蚀等(Kirwan & Patrick Megonigal, 2013; Mao et al., 2018 ).因此, 植物生物量的空间分配能够反映近期光合同化的新碳向根系和土壤碳库的转移规律, 是土壤碳输入和碳埋藏的重要参考指标, 深刻影响着土壤碳库的变化(Bolinderet al., 2012 ).通常情况下, 植物光合固定的碳即时分配大小为茎叶>根>土壤, 大部分光合固定碳都留在地上部分, 氮增加则会更加提高植物同化碳向植物地上部分的流动, 同时显著增加根际土壤中碳的累积与回收率, 而降低植物根系中的光合碳分配(王婷婷等, 2017).随着同位素技术的发展和应用, 通过测定土壤或植物中的碳同位素自然丰度值可以量化光合碳在植物-土壤系统中的动态变化、周转规律及其对环境变化的响应(Wang et al., 2019 ; Xiaoet al., 2019 ).例如, 13C脉冲标记实验表明, 适量氮输入显著提高即时光合碳分配给根系的比例(Wang et al., 2019 ).氮输入通常刺激植物生长早期同化碳向土壤方向迁移, 但生长后期可能降低, 这种由生长阶段造成的差异使得光合碳在根系中的分配模式对氮输入的响应具有一定的不确定性(王婷婷等, 2017; Xiao et al., 2019 ). ...
Atmospheric nitrogen deposition promotes carbon loss from peat bogs 2 2006
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... 排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Declining oxygen in the global ocean and coastal waters 1 2018
... 陆源氮通过地表径流进入近海, 导致近岸海域富营养化日益加剧(Deegan et al., 2012 ; Xu et al., 2020 ), 是目前全球海岸带面临的最为严重的一个环境问题.在潮汐作用下, 大量氮必然会进入盐沼等滨海湿地生态系统, 并被盐沼植被截留, 进而改变其光合碳输入、植物-土壤碳分配、土壤有机碳分解和土壤可溶性有机碳释放等碳循环关键过程(Hu et al., 2016 ; 韩广轩, 2017; Herbert et al., 2020 ), 并对其碳汇功能产生广泛且深刻的影响(图1).相比工业化之前, 近海水体氮负荷持续加重, 全球范围内由陆地向海洋输出的氮至少增加了10倍, 且主要以无机氮(DIN)为主要氮类型(Deegan et al., 2012 ; Breitburg et al., 2018 ).监测及模拟结果表明, 2000-2050年间, 中国沿海总氮输入量将增加30%-200% (Strokal et al., 2014 ; Wang et al., 2018 ).因此, 阐明氮输入对盐沼湿地碳循环关键过程的影响机制, 将有助于揭示氮输入对盐沼湿地蓝色碳汇形成过程与机制的影响, 并为预测近岸海域富营养化背景下盐沼湿地碳库的潜在变化趋势提供科学依据. ...
Effects of nutrient addition on vegetation and carbon cycling in an ombrotrophic bog 2 2007
... 氮输入对盐沼湿地植物光合固碳影响并非简单的线性变化, 存在阈值效应, 主要受时间尺度、氮输入类型和水平的影响(Vivanco et al., 2015 ; Peng et al., 2019 ; Xiao et al., 2019 ; Herbert et al., 2020 ).大部分生态系统都会受到氮的限制, 因此外源氮输入能通过提高植被的氮利用效率显著提高植物光合固碳能力, 刺激植物地上或者地下部分生长, 进而增加土壤碳输入(Fernández-Martínezet al., 2014 ; Herbert et al., 2020 ).但是, 达到一定阈值后, 继续加氮可能抑制这种正效应, 甚至产生金属毒害作用从而抑制植物正常生长(Bubier et al., 2007 ; Peng et al., 2019 ).就盐沼湿地而言, 特别是新生湿地, 通常是氮限制, 因此短期内大量氮输入可能会提高其植物光合固碳能力.例如, 加氮能通过改变植株密度、促进土壤磷流动以及增大微域环境CO2浓度等途径增加植物光合固碳能力(Wu et al., 2015 ).但是长期大量氮输入可能导致生态系统逐渐走向氮饱和, 甚至导致生态系统从氮限制转变为磷限制, 从而降低植物光合固碳能力对氮输入的响应(Chen et al., 2017 ; Peng et al., 2019 ). ...
Nitrogen addition reduces dissolved organic carbon leaching in a montane forest 3 2018
... 外源氮输入影响盐沼湿地土壤DOC的产生和累积, 进而影响盐沼湿地横向碳流失.盐沼湿地土壤中的碳流失主要有2种方式, 一种是以CO2和CH4等气体形式排放, 另一种是以DOC形式流失.尽管DOC仅占土壤总有机碳的0.04%-0.22%, 但它却是有机碳库中最活跃和不容忽视的组成部分(Bauer et al., 2013 ).例如, 由于受到充沛降雨和周期性的潮汐作用, 横向碳输出量大约占温带滨海湿地总碳输出量的40% (Majidzadeh et al., 2017 ).毫无疑问, 外源氮输入对盐沼湿地土壤DOC的产生和释放具有显著影响(图3).首先, 外源氮输入能促进盐沼植被生长和生物量增加, 而植物凋落物和根系分泌物增加能促进土壤DOC的形成和释放.但是长期氮富集可能导致盐沼湿地退化, 根际激发效应促进了分解, 从而降低了DOC含量(Deegan et al., 2012 ).其次, 氮输入通过影响土壤pH间接影响土壤有机质中DOC释放, 同时氮类型也会改变DOC对氮输入的响应(Chang et al., 2018 ).铵态氮增加通常能降低土壤pH, 抑制土壤中DOC淋溶, 从而减少土壤有机碳的横向流失; 但硝态氮增加则通过提高土壤pH, 从而促进DOC释放(Chang et al., 2018 ; Preston et al., 2020 ).再次, 氮输入增加了土壤微生物生物量, 加快了微生物新陈代谢活动, 从而加速了土壤DOC分解(Fellman et al., 2017 ).然而目前研究多集中在氮输入对盐沼湿地垂直方向碳流失(CO2和CH4)的影响, 忽视了近岸水体富营养化背景下以DOC为主要形式的陆海横向碳交换, 制约着对滨海盐沼湿地碳循环过程的整体理解以及对其碳收支的准确评估. ...
... ).铵态氮增加通常能降低土壤pH, 抑制土壤中DOC淋溶, 从而减少土壤有机碳的横向流失; 但硝态氮增加则通过提高土壤pH, 从而促进DOC释放(Chang et al., 2018 ; Preston et al., 2020 ).再次, 氮输入增加了土壤微生物生物量, 加快了微生物新陈代谢活动, 从而加速了土壤DOC分解(Fellman et al., 2017 ).然而目前研究多集中在氮输入对盐沼湿地垂直方向碳流失(CO2和CH4)的影响, 忽视了近岸水体富营养化背景下以DOC为主要形式的陆海横向碳交换, 制约着对滨海盐沼湿地碳循环过程的整体理解以及对其碳收支的准确评估. ...
... 综上所述, 近岸水体富营养化背景下, 大量氮输入对盐沼湿地植物光合碳输入、植物-土壤碳分配和土壤碳输出等碳循环关键过程产生深刻影响, 进而影响其碳汇功能的准确评估.首先, 氮输入对湿地植物光合固碳的影响存在阈值效应(Vivanco et al., 2015 ; Peng et al., 2019 ).低水平氮输入可以增强植物光合固碳能力, 但是达到一定阈值后, 继续加氮可能抑制这种正效应(Peng et al., 2019 ).其次, 氮输入影响植物光合产物在植物-土壤系统中的分配比例.光合固定碳向根系和土壤碳库的分配和转移规律, 深刻影响着土壤碳库的走向(Xu et al., 2020 ).再次, 氮输入会影响土壤有机碳的分解, 但是影响结果与氮输入类型和水平、土壤初始碳氮含量、施氮时间尺度等有很大的关系, 研究结论甚至截然相反(Chen et al., 2017 ; Zhou et al., 2017 ).另外, 氮输入对盐沼湿地土壤DOC的产生和累积有显著影响(Deegan et al., 2012 ; Chang et al., 2018 ), 进而影响盐沼湿地横向碳流失(韩广轩, 2017).最后, 氮输入在提高植被固碳潜力的同时, 也会促进土壤, 尤其是深层土壤中有机碳分解(Macket al., 2004 ).因此, 氮输入对盐沼湿地碳循环过程的影响存在不确定性, 无疑将在很大程度上制约着对盐沼湿地碳循环和碳汇功能的整体理解以及水体富营养化背景下盐沼湿地“蓝碳”吸存能力的评估和预测. ...
Carbon input to terrestrial ecosystems//Chapin III FS, Matson PA, Mooney HA. Principles of Terrestrial Ecosystem Ecology. Springer 1 2011
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
Effects of nitrogen enrichment on belowground communities in grassland: relative role of soil nitrogen availability vs. soil acidification 1 2015a
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
Effects of nitrogen deposition on carbon cycle in terrestrial ecosystems of China: a meta-analysis 1 2015b
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
Effects of warming and nitrogen fertilization on GHG flux in an alpine swamp meadow of a permafrost region 5 2017
... 氮输入对盐沼湿地植物光合固碳影响并非简单的线性变化, 存在阈值效应, 主要受时间尺度、氮输入类型和水平的影响(Vivanco et al., 2015 ; Peng et al., 2019 ; Xiao et al., 2019 ; Herbert et al., 2020 ).大部分生态系统都会受到氮的限制, 因此外源氮输入能通过提高植被的氮利用效率显著提高植物光合固碳能力, 刺激植物地上或者地下部分生长, 进而增加土壤碳输入(Fernández-Martínezet al., 2014 ; Herbert et al., 2020 ).但是, 达到一定阈值后, 继续加氮可能抑制这种正效应, 甚至产生金属毒害作用从而抑制植物正常生长(Bubier et al., 2007 ; Peng et al., 2019 ).就盐沼湿地而言, 特别是新生湿地, 通常是氮限制, 因此短期内大量氮输入可能会提高其植物光合固碳能力.例如, 加氮能通过改变植株密度、促进土壤磷流动以及增大微域环境CO2浓度等途径增加植物光合固碳能力(Wu et al., 2015 ).但是长期大量氮输入可能导致生态系统逐渐走向氮饱和, 甚至导致生态系统从氮限制转变为磷限制, 从而降低植物光合固碳能力对氮输入的响应(Chen et al., 2017 ; Peng et al., 2019 ). ...
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... 滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 ).结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 ).据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
... 综上所述, 近岸水体富营养化背景下, 大量氮输入对盐沼湿地植物光合碳输入、植物-土壤碳分配和土壤碳输出等碳循环关键过程产生深刻影响, 进而影响其碳汇功能的准确评估.首先, 氮输入对湿地植物光合固碳的影响存在阈值效应(Vivanco et al., 2015 ; Peng et al., 2019 ).低水平氮输入可以增强植物光合固碳能力, 但是达到一定阈值后, 继续加氮可能抑制这种正效应(Peng et al., 2019 ).其次, 氮输入影响植物光合产物在植物-土壤系统中的分配比例.光合固定碳向根系和土壤碳库的分配和转移规律, 深刻影响着土壤碳库的走向(Xu et al., 2020 ).再次, 氮输入会影响土壤有机碳的分解, 但是影响结果与氮输入类型和水平、土壤初始碳氮含量、施氮时间尺度等有很大的关系, 研究结论甚至截然相反(Chen et al., 2017 ; Zhou et al., 2017 ).另外, 氮输入对盐沼湿地土壤DOC的产生和累积有显著影响(Deegan et al., 2012 ; Chang et al., 2018 ), 进而影响盐沼湿地横向碳流失(韩广轩, 2017).最后, 氮输入在提高植被固碳潜力的同时, 也会促进土壤, 尤其是深层土壤中有机碳分解(Macket al., 2004 ).因此, 氮输入对盐沼湿地碳循环过程的影响存在不确定性, 无疑将在很大程度上制约着对盐沼湿地碳循环和碳汇功能的整体理解以及水体富营养化背景下盐沼湿地“蓝碳”吸存能力的评估和预测. ...
Nitrogen fertilization stimulated soil heterotrophic but not autotrophic respiration in cropland soils: a greater role of organic over inorganic fertilizer 1 2018
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Global carbon sequestration in tidal, saline wetland soils 2 2003
... 滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 ).结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 ).据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
... (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
Dynamics of carbon sequestration in a coastal wetland using radiocarbon measurements 1 2004
Effect of nitrogen (N) input on decomposition of soil organic carbon in a salt marsh (adapted from <xref ref-type="bibr" rid="b36">Hester <i>et al</i>., 2018 </xref>). Microbial processes are directly or indirectly affected by N input. Plants mainly absorb NH<sub>4</sub><sup>+</sup>-N, which stimulates plant productivity and the deposition of organic matter from the rhizosphere, thus stimulating the material circulation of rhizosphere and the metabolism of rhizosphere microorganisms. Plant derived carbon input and higher N utilization jointly stimulate microbial heterotrophic respiration, and then stimulate the production and emission of CO<sub>2</sub> in aerobic condition. In anaerobic condition, excessive NO<sub>3</sub><sup>-</sup>-N is converted into NH<sub>4</sub><sup>+</sup>-N, which will inhibit CH<sub>4</sub> oxidation. Fig. 2 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... Effect of nitrogen (N) input on decomposition of soil organic carbon in a salt marsh (adapted from Hester et al., 2018 ). Microbial processes are directly or indirectly affected by N input. Plants mainly absorb NH4+-N, which stimulates plant productivity and the deposition of organic matter from the rhizosphere, thus stimulating the material circulation of rhizosphere and the metabolism of rhizosphere microorganisms. Plant derived carbon input and higher N utilization jointly stimulate microbial heterotrophic respiration, and then stimulate the production and emission of CO2 in aerobic condition. In anaerobic condition, excessive NO3--N is converted into NH4+-N, which will inhibit CH4 oxidation. Fig. 2 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... 排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Quantification of effects of season and nitrogen supply on tree below-ground carbon transfer to ectomycorrhizal fungi and other soil organisms in a boreal pine forest 1 2010
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Salinity and nutrient contents of tidal water affects soil respiration and carbon sequestration of high and low tidal flats of Jiuduansha wetlands in different ways 1 2016
... 陆源氮通过地表径流进入近海, 导致近岸海域富营养化日益加剧(Deegan et al., 2012 ; Xu et al., 2020 ), 是目前全球海岸带面临的最为严重的一个环境问题.在潮汐作用下, 大量氮必然会进入盐沼等滨海湿地生态系统, 并被盐沼植被截留, 进而改变其光合碳输入、植物-土壤碳分配、土壤有机碳分解和土壤可溶性有机碳释放等碳循环关键过程(Hu et al., 2016 ; 韩广轩, 2017; Herbert et al., 2020 ), 并对其碳汇功能产生广泛且深刻的影响(图1).相比工业化之前, 近海水体氮负荷持续加重, 全球范围内由陆地向海洋输出的氮至少增加了10倍, 且主要以无机氮(DIN)为主要氮类型(Deegan et al., 2012 ; Breitburg et al., 2018 ).监测及模拟结果表明, 2000-2050年间, 中国沿海总氮输入量将增加30%-200% (Strokal et al., 2014 ; Wang et al., 2018 ).因此, 阐明氮输入对盐沼湿地碳循环关键过程的影响机制, 将有助于揭示氮输入对盐沼湿地蓝色碳汇形成过程与机制的影响, 并为预测近岸海域富营养化背景下盐沼湿地碳库的潜在变化趋势提供科学依据. ...
Decomposition of mangrove roots: effects of location, nutrients, species identity and mix in a Kenyan forest 1 2010
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
Scaling plant nitrogen use and uptake efficiencies in response to nutrient addition in peatlands 1 2010
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
Reduction of forest soil respiration in response to nitrogen deposition 1 2010
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
Amino acid, peptide and protein mineralization dynamics in a taiga forest soil 1 2012
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
Responses of vegetation and ecosystem CO2 exchange to 9 years of nutrient addition at Mer Bleue bog 1 2010
... 滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 ).结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 ).据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
Short- and long-term effects of nutrient enrichment on microbial exoenzyme activity in mangrove peat 1 2015
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
Plant nitrogen acquisition under low availability: regulation of uptake and root architecture 1 2016
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
Nitrogen addition differently alters GHGs production and soil microbial community of tidal salt marsh soil depending on the types of halophyte 0 2020
Tidal wetland stability in the face of human impacts and sea-level rise 1 2013
... 氮输入不仅从整体上影响植物光合固碳能力, 还能影响植物光合产物在植物-土壤系统中的分配比例.植物地上、地下生物量都是盐沼湿地碳汇的重要组成部分(韩广轩, 2017; Hayes et al., 2017 ).首先, 植物地上部分能够拦截海水中的泥沙及颗粒有机质, 帮助盐沼湿地快速抬升高程, 增大其有机碳的储存空间; 同时, 植物地上部分是植物作为初级生产者固定光合碳的器官, 其生物量大小对滨海盐沼湿地的初级生产力具有直接影响.植物地下部分一方面能够分泌根际沉积物, 是土壤可溶性有机碳等易分解碳的重要来源; 另一方面, 植物地下根系能够锚定沉积物, 防止潮汐作用对有机质的侵蚀等(Kirwan & Patrick Megonigal, 2013; Mao et al., 2018 ).因此, 植物生物量的空间分配能够反映近期光合同化的新碳向根系和土壤碳库的转移规律, 是土壤碳输入和碳埋藏的重要参考指标, 深刻影响着土壤碳库的变化(Bolinderet al., 2012 ).通常情况下, 植物光合固定的碳即时分配大小为茎叶>根>土壤, 大部分光合固定碳都留在地上部分, 氮增加则会更加提高植物同化碳向植物地上部分的流动, 同时显著增加根际土壤中碳的累积与回收率, 而降低植物根系中的光合碳分配(王婷婷等, 2017).随着同位素技术的发展和应用, 通过测定土壤或植物中的碳同位素自然丰度值可以量化光合碳在植物-土壤系统中的动态变化、周转规律及其对环境变化的响应(Wang et al., 2019 ; Xiaoet al., 2019 ).例如, 13C脉冲标记实验表明, 适量氮输入显著提高即时光合碳分配给根系的比例(Wang et al., 2019 ).氮输入通常刺激植物生长早期同化碳向土壤方向迁移, 但生长后期可能降低, 这种由生长阶段造成的差异使得光合碳在根系中的分配模式对氮输入的响应具有一定的不确定性(王婷婷等, 2017; Xiao et al., 2019 ). ...
Long-term enhanced nitrogen deposition increases ecosystem respiration and carbon loss from a Sphagnum bog in the Scottish Borders 0 2013
Vegetation feedbacks of nutrient addition lead to a weaker carbon sink in an ombrotrophic bog 0 2013
Effects of nitrogen enrichment on tree carbon allocation: a global synthesis 1 2020
Four decadesʼ dynamics of coastal blue carbon storage driven by land use/land cover transformation under natural and anthropogenic processes in the Yellow River Delta, China 1 2019
... 滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 ).结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 ).据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
Ecosystem carbon storage in arctic tundra reduced by long-term nutrient fertilization 1 2004
... 综上所述, 近岸水体富营养化背景下, 大量氮输入对盐沼湿地植物光合碳输入、植物-土壤碳分配和土壤碳输出等碳循环关键过程产生深刻影响, 进而影响其碳汇功能的准确评估.首先, 氮输入对湿地植物光合固碳的影响存在阈值效应(Vivanco et al., 2015 ; Peng et al., 2019 ).低水平氮输入可以增强植物光合固碳能力, 但是达到一定阈值后, 继续加氮可能抑制这种正效应(Peng et al., 2019 ).其次, 氮输入影响植物光合产物在植物-土壤系统中的分配比例.光合固定碳向根系和土壤碳库的分配和转移规律, 深刻影响着土壤碳库的走向(Xu et al., 2020 ).再次, 氮输入会影响土壤有机碳的分解, 但是影响结果与氮输入类型和水平、土壤初始碳氮含量、施氮时间尺度等有很大的关系, 研究结论甚至截然相反(Chen et al., 2017 ; Zhou et al., 2017 ).另外, 氮输入对盐沼湿地土壤DOC的产生和累积有显著影响(Deegan et al., 2012 ; Chang et al., 2018 ), 进而影响盐沼湿地横向碳流失(韩广轩, 2017).最后, 氮输入在提高植被固碳潜力的同时, 也会促进土壤, 尤其是深层土壤中有机碳分解(Macket al., 2004 ).因此, 氮输入对盐沼湿地碳循环过程的影响存在不确定性, 无疑将在很大程度上制约着对盐沼湿地碳循环和碳汇功能的整体理解以及水体富营养化背景下盐沼湿地“蓝碳”吸存能力的评估和预测. ...
... 外源氮输入影响盐沼湿地土壤DOC的产生和累积, 进而影响盐沼湿地横向碳流失.盐沼湿地土壤中的碳流失主要有2种方式, 一种是以CO2和CH4等气体形式排放, 另一种是以DOC形式流失.尽管DOC仅占土壤总有机碳的0.04%-0.22%, 但它却是有机碳库中最活跃和不容忽视的组成部分(Bauer et al., 2013 ).例如, 由于受到充沛降雨和周期性的潮汐作用, 横向碳输出量大约占温带滨海湿地总碳输出量的40% (Majidzadeh et al., 2017 ).毫无疑问, 外源氮输入对盐沼湿地土壤DOC的产生和释放具有显著影响(图3).首先, 外源氮输入能促进盐沼植被生长和生物量增加, 而植物凋落物和根系分泌物增加能促进土壤DOC的形成和释放.但是长期氮富集可能导致盐沼湿地退化, 根际激发效应促进了分解, 从而降低了DOC含量(Deegan et al., 2012 ).其次, 氮输入通过影响土壤pH间接影响土壤有机质中DOC释放, 同时氮类型也会改变DOC对氮输入的响应(Chang et al., 2018 ).铵态氮增加通常能降低土壤pH, 抑制土壤中DOC淋溶, 从而减少土壤有机碳的横向流失; 但硝态氮增加则通过提高土壤pH, 从而促进DOC释放(Chang et al., 2018 ; Preston et al., 2020 ).再次, 氮输入增加了土壤微生物生物量, 加快了微生物新陈代谢活动, 从而加速了土壤DOC分解(Fellman et al., 2017 ).然而目前研究多集中在氮输入对盐沼湿地垂直方向碳流失(CO2和CH4)的影响, 忽视了近岸水体富营养化背景下以DOC为主要形式的陆海横向碳交换, 制约着对滨海盐沼湿地碳循环过程的整体理解以及对其碳收支的准确评估. ...
Environmental and stoichiometric controls on microbial carbon-use efficiency in soils 1 2012
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
Effects of simulated N deposition on foliar nutrient status, N metabolism and photosynthetic capacity of three dominant understory plant species in a mature tropical forest 3 2018
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
... 了植株的生长(Harpole et al., 2016 ; Wang et al., 2018 ); (2)高浓度NH4+的潜在阳离子毒性可能限制植物的生长(Wei et al., 2013 ); (3)氮添加引起群落内部的物种更替.此外, 植物光合作用对氮的响应还取决于植物本身的性能.例如, 某些莎草属(Cyperus)的植物对氮较为敏感, 即使较低的氮负荷也会对其光合能力产生负面影响; 但一些氮利用效率较高、生存能力较强的物种则更能适应氮负荷较高的生境, 并提高自身的光合能力和生产力(Mao et al., 2018 ; Shen et al., 2019 ). ...
... 氮输入不仅从整体上影响植物光合固碳能力, 还能影响植物光合产物在植物-土壤系统中的分配比例.植物地上、地下生物量都是盐沼湿地碳汇的重要组成部分(韩广轩, 2017; Hayes et al., 2017 ).首先, 植物地上部分能够拦截海水中的泥沙及颗粒有机质, 帮助盐沼湿地快速抬升高程, 增大其有机碳的储存空间; 同时, 植物地上部分是植物作为初级生产者固定光合碳的器官, 其生物量大小对滨海盐沼湿地的初级生产力具有直接影响.植物地下部分一方面能够分泌根际沉积物, 是土壤可溶性有机碳等易分解碳的重要来源; 另一方面, 植物地下根系能够锚定沉积物, 防止潮汐作用对有机质的侵蚀等(Kirwan & Patrick Megonigal, 2013; Mao et al., 2018 ).因此, 植物生物量的空间分配能够反映近期光合同化的新碳向根系和土壤碳库的转移规律, 是土壤碳输入和碳埋藏的重要参考指标, 深刻影响着土壤碳库的变化(Bolinderet al., 2012 ).通常情况下, 植物光合固定的碳即时分配大小为茎叶>根>土壤, 大部分光合固定碳都留在地上部分, 氮增加则会更加提高植物同化碳向植物地上部分的流动, 同时显著增加根际土壤中碳的累积与回收率, 而降低植物根系中的光合碳分配(王婷婷等, 2017).随着同位素技术的发展和应用, 通过测定土壤或植物中的碳同位素自然丰度值可以量化光合碳在植物-土壤系统中的动态变化、周转规律及其对环境变化的响应(Wang et al., 2019 ; Xiaoet al., 2019 ).例如, 13C脉冲标记实验表明, 适量氮输入显著提高即时光合碳分配给根系的比例(Wang et al., 2019 ).氮输入通常刺激植物生长早期同化碳向土壤方向迁移, 但生长后期可能降低, 这种由生长阶段造成的差异使得光合碳在根系中的分配模式对氮输入的响应具有一定的不确定性(王婷婷等, 2017; Xiao et al., 2019 ). ...
A blueprint for blue carbon: toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2 3 2011
Changes in mass and nutrient content of wood during decomposition in a South Florida mangrove forest 2 2005
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
... ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
Investigating the mechanisms for the opposing pH relationships of fungal and bacterial growth in soil 1 2010
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
Effects of simulated N deposition on photosynthesis and productivity of key plants from different functional groups of alpine meadow on Qinghai-Tibetan Plateau 1 2019
... 了植株的生长(Harpole et al., 2016 ; Wang et al., 2018 ); (2)高浓度NH4+的潜在阳离子毒性可能限制植物的生长(Wei et al., 2013 ); (3)氮添加引起群落内部的物种更替.此外, 植物光合作用对氮的响应还取决于植物本身的性能.例如, 某些莎草属(Cyperus)的植物对氮较为敏感, 即使较低的氮负荷也会对其光合能力产生负面影响; 但一些氮利用效率较高、生存能力较强的物种则更能适应氮负荷较高的生境, 并提高自身的光合能力和生产力(Mao et al., 2018 ; Shen et al., 2019 ). ...
Spatial patterns of biomass and aboveground net primary productivity in a mangrove ecosystem in the Dominican Republic 1 2003
... 在氮有效性较高、分解率低的潮汐盐沼湿地, 氮输入一方面可能会促进植被根系生长, 提高土壤有机质输入, 加快有机质积累, 从而短期内大量氮输入可能会提高湿地的碳汇量(Hayes et al., 2017 ).对于森林生态系统而言, 新碳的加入能激发促进土壤老碳的更新循环.氮输入驱动下增加的碳大部分都是新的植物生物量, 而非土壤碳.因此, 氮输入对这类生态系统长期土壤碳汇反而具有消极的影响(Tipping et al., 2012 ; Mills et al., 2014 ).因此, 相比之下, 由于沼泽、泥炭地等生态系统土壤有机质分解受到高湿度和富含难分解化合物的限制, 新输入的植物凋落物等有机质被封存和隔离, 甚至能够在土壤中埋藏成百上千年(Dise, 2009).有研究表明, 在滨海湿地生态系统中, 氮输入促进芦苇(Phragmites australis)生长、提高芦苇生物量, 增加了土壤中的碳含量(Qu et al., 2020 ).另一方面, 氮输入通过影响植被的地下生物量分配进而影响土壤碳库.已有研究表明, 盐沼湿地在氮有效性较高时, 总生物量会增加, 但当氮有效性较低时, 地下部分的生物量分配会相对提高(Clough, 1992; Sherman et al., 2003 ).生态系统氮饱和假说认为, 向氮饱和的生态系统中继续加氮会导致土壤功能的丧失, 例如养分循环和氮固持功能等减弱, 进而对植物生长产生影响(Aber et al., 1998 ). ...
The balanced-growth hypothesis and the allometry of leaf and root biomass allocation 1 2002
Effect of nitrogen addition on decomposition of Calamagrostis angustifolia litters from freshwater marshes of Northeast China 0 2011
Effect of continued nitrogen enrichment on greenhouse gas emissions from a wetland ecosystem in the Sanjiang Plain, Northeast China: a 5 year nitrogen addition experiment 1 2013
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Effects of temperature, glucose and inorganic nitrogen inputs on carbon mineralization in a Tibetan alpine meadow soil 1 2010
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Increasing eutrophication in the coastal seas of China from 1970 to 2050 1 2014
... 陆源氮通过地表径流进入近海, 导致近岸海域富营养化日益加剧(Deegan et al., 2012 ; Xu et al., 2020 ), 是目前全球海岸带面临的最为严重的一个环境问题.在潮汐作用下, 大量氮必然会进入盐沼等滨海湿地生态系统, 并被盐沼植被截留, 进而改变其光合碳输入、植物-土壤碳分配、土壤有机碳分解和土壤可溶性有机碳释放等碳循环关键过程(Hu et al., 2016 ; 韩广轩, 2017; Herbert et al., 2020 ), 并对其碳汇功能产生广泛且深刻的影响(图1).相比工业化之前, 近海水体氮负荷持续加重, 全球范围内由陆地向海洋输出的氮至少增加了10倍, 且主要以无机氮(DIN)为主要氮类型(Deegan et al., 2012 ; Breitburg et al., 2018 ).监测及模拟结果表明, 2000-2050年间, 中国沿海总氮输入量将增加30%-200% (Strokal et al., 2014 ; Wang et al., 2018 ).因此, 阐明氮输入对盐沼湿地碳循环关键过程的影响机制, 将有助于揭示氮输入对盐沼湿地蓝色碳汇形成过程与机制的影响, 并为预测近岸海域富营养化背景下盐沼湿地碳库的潜在变化趋势提供科学依据. ...
Effects of inorganic and organic nitrogen additions on CO2 emissions in the coastal wetlands of the Yellow River Delta, China 1 2018
... 滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 ).结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 ).据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
N14C: a plant-soil nitrogen and carbon cycling model to simulate terrestrial ecosystem responses to atmospheric nitrogen deposition 1 2012
... 在氮有效性较高、分解率低的潮汐盐沼湿地, 氮输入一方面可能会促进植被根系生长, 提高土壤有机质输入, 加快有机质积累, 从而短期内大量氮输入可能会提高湿地的碳汇量(Hayes et al., 2017 ).对于森林生态系统而言, 新碳的加入能激发促进土壤老碳的更新循环.氮输入驱动下增加的碳大部分都是新的植物生物量, 而非土壤碳.因此, 氮输入对这类生态系统长期土壤碳汇反而具有消极的影响(Tipping et al., 2012 ; Mills et al., 2014 ).因此, 相比之下, 由于沼泽、泥炭地等生态系统土壤有机质分解受到高湿度和富含难分解化合物的限制, 新输入的植物凋落物等有机质被封存和隔离, 甚至能够在土壤中埋藏成百上千年(Dise, 2009).有研究表明, 在滨海湿地生态系统中, 氮输入促进芦苇(Phragmites australis)生长、提高芦苇生物量, 增加了土壤中的碳含量(Qu et al., 2020 ).另一方面, 氮输入通过影响植被的地下生物量分配进而影响土壤碳库.已有研究表明, 盐沼湿地在氮有效性较高时, 总生物量会增加, 但当氮有效性较低时, 地下部分的生物量分配会相对提高(Clough, 1992; Sherman et al., 2003 ).生态系统氮饱和假说认为, 向氮饱和的生态系统中继续加氮会导致土壤功能的丧失, 例如养分循环和氮固持功能等减弱, 进而对植物生长产生影响(Aber et al., 1998 ). ...
Nitrogen and phosphorus limitation in a coastal barrier salt marsh: the implications for vegetation succession 0 1999
Nitrogen and nature 1 2002
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
Nonlinear responses in salt marsh functioning to increased nitrogen addition 3 2015
... 氮输入对盐沼湿地植物光合固碳影响并非简单的线性变化, 存在阈值效应, 主要受时间尺度、氮输入类型和水平的影响(Vivanco et al., 2015 ; Peng et al., 2019 ; Xiao et al., 2019 ; Herbert et al., 2020 ).大部分生态系统都会受到氮的限制, 因此外源氮输入能通过提高植被的氮利用效率显著提高植物光合固碳能力, 刺激植物地上或者地下部分生长, 进而增加土壤碳输入(Fernández-Martínezet al., 2014 ; Herbert et al., 2020 ).但是, 达到一定阈值后, 继续加氮可能抑制这种正效应, 甚至产生金属毒害作用从而抑制植物正常生长(Bubier et al., 2007 ; Peng et al., 2019 ).就盐沼湿地而言, 特别是新生湿地, 通常是氮限制, 因此短期内大量氮输入可能会提高其植物光合固碳能力.例如, 加氮能通过改变植株密度、促进土壤磷流动以及增大微域环境CO2浓度等途径增加植物光合固碳能力(Wu et al., 2015 ).但是长期大量氮输入可能导致生态系统逐渐走向氮饱和, 甚至导致生态系统从氮限制转变为磷限制, 从而降低植物光合固碳能力对氮输入的响应(Chen et al., 2017 ; Peng et al., 2019 ). ...
... 综上所述, 近岸水体富营养化背景下, 大量氮输入对盐沼湿地植物光合碳输入、植物-土壤碳分配和土壤碳输出等碳循环关键过程产生深刻影响, 进而影响其碳汇功能的准确评估.首先, 氮输入对湿地植物光合固碳的影响存在阈值效应(Vivanco et al., 2015 ; Peng et al., 2019 ).低水平氮输入可以增强植物光合固碳能力, 但是达到一定阈值后, 继续加氮可能抑制这种正效应(Peng et al., 2019 ).其次, 氮输入影响植物光合产物在植物-土壤系统中的分配比例.光合固定碳向根系和土壤碳库的分配和转移规律, 深刻影响着土壤碳库的走向(Xu et al., 2020 ).再次, 氮输入会影响土壤有机碳的分解, 但是影响结果与氮输入类型和水平、土壤初始碳氮含量、施氮时间尺度等有很大的关系, 研究结论甚至截然相反(Chen et al., 2017 ; Zhou et al., 2017 ).另外, 氮输入对盐沼湿地土壤DOC的产生和累积有显著影响(Deegan et al., 2012 ; Chang et al., 2018 ), 进而影响盐沼湿地横向碳流失(韩广轩, 2017).最后, 氮输入在提高植被固碳潜力的同时, 也会促进土壤, 尤其是深层土壤中有机碳分解(Macket al., 2004 ).因此, 氮输入对盐沼湿地碳循环过程的影响存在不确定性, 无疑将在很大程度上制约着对盐沼湿地碳循环和碳汇功能的整体理解以及水体富营养化背景下盐沼湿地“蓝碳”吸存能力的评估和预测. ...
A historical overview of coastal eutrophication in the China Seas 2 2018
... 陆源氮通过地表径流进入近海, 导致近岸海域富营养化日益加剧(Deegan et al., 2012 ; Xu et al., 2020 ), 是目前全球海岸带面临的最为严重的一个环境问题.在潮汐作用下, 大量氮必然会进入盐沼等滨海湿地生态系统, 并被盐沼植被截留, 进而改变其光合碳输入、植物-土壤碳分配、土壤有机碳分解和土壤可溶性有机碳释放等碳循环关键过程(Hu et al., 2016 ; 韩广轩, 2017; Herbert et al., 2020 ), 并对其碳汇功能产生广泛且深刻的影响(图1).相比工业化之前, 近海水体氮负荷持续加重, 全球范围内由陆地向海洋输出的氮至少增加了10倍, 且主要以无机氮(DIN)为主要氮类型(Deegan et al., 2012 ; Breitburg et al., 2018 ).监测及模拟结果表明, 2000-2050年间, 中国沿海总氮输入量将增加30%-200% (Strokal et al., 2014 ; Wang et al., 2018 ).因此, 阐明氮输入对盐沼湿地碳循环关键过程的影响机制, 将有助于揭示氮输入对盐沼湿地蓝色碳汇形成过程与机制的影响, 并为预测近岸海域富营养化背景下盐沼湿地碳库的潜在变化趋势提供科学依据. ...
... 了植株的生长(Harpole et al., 2016 ; Wang et al., 2018 ); (2)高浓度NH4+的潜在阳离子毒性可能限制植物的生长(Wei et al., 2013 ); (3)氮添加引起群落内部的物种更替.此外, 植物光合作用对氮的响应还取决于植物本身的性能.例如, 某些莎草属(Cyperus)的植物对氮较为敏感, 即使较低的氮负荷也会对其光合能力产生负面影响; 但一些氮利用效率较高、生存能力较强的物种则更能适应氮负荷较高的生境, 并提高自身的光合能力和生产力(Mao et al., 2018 ; Shen et al., 2019 ). ...
Molecular mechanisms of water table lowering and nitrogen deposition in affecting greenhouse gas emissions from a Tibetan alpine wetland 0 2017
Asymmetry in above- and belowground productivity responses to N addition in a semi-arid temperate steppe 6 2019
... 氮被认为是限制陆地生态系统净初级生产力的最重要的一种营养元素, 植物可以从土壤中以无机物(如硝酸盐和铵盐)或有机物(如尿素、氨基酸和肽)的形式通过根部获得氮(Jones & Kielland, 2012; Kiba & Krapp, 2016).最新的研究表明, 全球超过50%的天然陆地生态系统受到氮限制(Du et al., 2020 ).氮被生物固定到土壤之后, 非常容易通过淋溶或挥发从生态系统中移除, 例如以溶解性有机氮(DON)和DIN形式的氮流失(Vitousek et al., 2002 ).从植物组织到植物个体, 再到整个群落的组成部分, 氮利用效率均存在较大的差异(Wang et al., 2019 ).氮输入一方面通过改变植株整体生物量以及植株地上和地下间、叶片和木质组织间生物量的分配等, 进而影响植株的氮利用效率(Iversen et al., 2010 ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
... ; Wang et al., 2019 ).例如, 氮是光捕获组织及植物生物量的重要组分, 氮在叶片光合酶中占很大比例, 因此叶片氮含量与光合能力之间普遍存在正相关关系(Chapin III et al., 2011 ; Mao et al., 2018 ).适量增加土壤氮有效性能提高植物叶片生物量及叶片氮含量, 进而通过3种途径提高植物生物量: 增加CO2的吸收、通过改变CO2同化作用及气孔导度提高叶片的水分利用效率、减少光消耗(Guerrieri et al., 2011 ).另一方面, 长期氮输入可能对植物群落演替产生影响, 从而影响植被群落的氮吸收利用效率(Wang et al., 2019 ).通常提高氮有效性会显著降低植物和群落的氮吸收利用效率, 这种现象主要有三方面的原因: (1)解除氮限制后, 其他环境因子(例如土壤水分、温度、光照强度等)变得更为关键, 限制 ...
Nitrogen deposition weakens plant-microbe interactions in grassland ecosystems 1 2013
... 了植株的生长(Harpole et al., 2016 ; Wang et al., 2018 ); (2)高浓度NH4+的潜在阳离子毒性可能限制植物的生长(Wei et al., 2013 ); (3)氮添加引起群落内部的物种更替.此外, 植物光合作用对氮的响应还取决于植物本身的性能.例如, 某些莎草属(Cyperus)的植物对氮较为敏感, 即使较低的氮负荷也会对其光合能力产生负面影响; 但一些氮利用效率较高、生存能力较强的物种则更能适应氮负荷较高的生境, 并提高自身的光合能力和生产力(Mao et al., 2018 ; Shen et al., 2019 ). ...
Experimental nitrogen, phosphorus, and potassium deposition decreases summer soil temperatures, water contents, and soil CO2 concentrations in a northern bog 1 2011
... 滨海湿地土壤碳库功能是滨海湿地一种重要的生态系统服务功能, 并在全球陆地生态系统碳循环中起着关键作用, 滨海湿地碳储量是其他陆地生态系统碳储量的50多倍(Ma et al., 2019 ).结合实际观测和生态模型预测发现, 盐沼湿地中的土壤碳累积速率为(218 ± 24) g·m -2·a-1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 相比之下, 温带、热带森林和北方森林的土壤碳累积速率仅为0.7-13.1 g·m-2·a-1 (McLeod et al., 2011 ).据估算, 全球盐沼湿地的年际碳储量约为(87.2 ± 9.6) Tg·a -1 (Chmura et al., 2003 ; Duarte et al., 2005 ), 其碳存储密度远高于具有高植被覆盖度和地上生物量的森林生态系统.与森林生态系统的植被光合固碳不同, 滨海湿地的固碳主要是由于沉积物的累积和土壤有机质的难分解性.研究发现, 滨海湿地常年有大量沉积物的累积, 且沉积物中有机碳的固存率高, 植物产生的大量有机物质难以分解, 并以土壤有机质的形式存在于地下, 因而滨海湿地生态系统具有很高的土壤固碳潜力(McLeod et al., 2011 ).氮作为陆地生态系统最重要的养分, 氮的有效性对滨海湿地土壤碳收支也有着重要的影响(Mitsch & Gosselink, 2011).为明确外源氮输入对盐沼湿地及其他湿地类型土壤碳库的影响, 许多研究通过原位控制实验、模型模拟以及meta分析等手段对氮输入背景下的湿地土壤碳储量进行了整体评估(表1).评估结果不尽相同, 究其原因, 土壤碳库存是由碳输入和碳输出之间的平衡所决定的.已有的研究表明, 土壤呼吸作为湿地主要的碳输出途径, 对氮输入的响应也是不同的, 如中国高寒湿地中, NO3--N的添加降低了土壤的呼吸速率(Chen et al., 2017 ), 然而在加拿大高寒湿地以及中国滨海湿地中, 氮添加反而促进了土壤呼吸, 从而加快了土壤的碳输出(Juutinen et al., 2010 , Tao et al., 2018 ).同样, 氮添加也可以促进植被的生长并吸收更多CO2, 提高湿地土壤地上和地下生物量, 从而提高土壤的碳输入.如在美国和加拿大的湿地碳循环研究中发现, 氮添加会通过提高生物量进而提高土壤的碳储存(Wendel et al., 2011 , Pastore et al., 2017 ).准确测量或估算湿地生态系统碳储存变化对外源氮输入的响应对研究湿地对全球生态系统碳平衡的贡献具有重要意义. ...
Effects of experimental nitrogen deposition on peatland carbon pools and fluxes: a modelling analysis 1 2015
... 氮输入对盐沼湿地植物光合固碳影响并非简单的线性变化, 存在阈值效应, 主要受时间尺度、氮输入类型和水平的影响(Vivanco et al., 2015 ; Peng et al., 2019 ; Xiao et al., 2019 ; Herbert et al., 2020 ).大部分生态系统都会受到氮的限制, 因此外源氮输入能通过提高植被的氮利用效率显著提高植物光合固碳能力, 刺激植物地上或者地下部分生长, 进而增加土壤碳输入(Fernández-Martínezet al., 2014 ; Herbert et al., 2020 ).但是, 达到一定阈值后, 继续加氮可能抑制这种正效应, 甚至产生金属毒害作用从而抑制植物正常生长(Bubier et al., 2007 ; Peng et al., 2019 ).就盐沼湿地而言, 特别是新生湿地, 通常是氮限制, 因此短期内大量氮输入可能会提高其植物光合固碳能力.例如, 加氮能通过改变植株密度、促进土壤磷流动以及增大微域环境CO2浓度等途径增加植物光合固碳能力(Wu et al., 2015 ).但是长期大量氮输入可能导致生态系统逐渐走向氮饱和, 甚至导致生态系统从氮限制转变为磷限制, 从而降低植物光合固碳能力对氮输入的响应(Chen et al., 2017 ; Peng et al., 2019 ). ...
Stimulation of long-term ammonium nitrogen deposition on methanogenesis by Methanocellaceae in a coastal wetland 2 2017
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
... 排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Nitrogen fertilization alters the distribution and fates of photosynthesized carbon in rice-soil systems: a13C-CO2 pulse labeling study 5 2019
... 氮输入对盐沼湿地植物光合固碳影响并非简单的线性变化, 存在阈值效应, 主要受时间尺度、氮输入类型和水平的影响(Vivanco et al., 2015 ; Peng et al., 2019 ; Xiao et al., 2019 ; Herbert et al., 2020 ).大部分生态系统都会受到氮的限制, 因此外源氮输入能通过提高植被的氮利用效率显著提高植物光合固碳能力, 刺激植物地上或者地下部分生长, 进而增加土壤碳输入(Fernández-Martínezet al., 2014 ; Herbert et al., 2020 ).但是, 达到一定阈值后, 继续加氮可能抑制这种正效应, 甚至产生金属毒害作用从而抑制植物正常生长(Bubier et al., 2007 ; Peng et al., 2019 ).就盐沼湿地而言, 特别是新生湿地, 通常是氮限制, 因此短期内大量氮输入可能会提高其植物光合固碳能力.例如, 加氮能通过改变植株密度、促进土壤磷流动以及增大微域环境CO2浓度等途径增加植物光合固碳能力(Wu et al., 2015 ).但是长期大量氮输入可能导致生态系统逐渐走向氮饱和, 甚至导致生态系统从氮限制转变为磷限制, 从而降低植物光合固碳能力对氮输入的响应(Chen et al., 2017 ; Peng et al., 2019 ). ...
... 氮输入不仅从整体上影响植物光合固碳能力, 还能影响植物光合产物在植物-土壤系统中的分配比例.植物地上、地下生物量都是盐沼湿地碳汇的重要组成部分(韩广轩, 2017; Hayes et al., 2017 ).首先, 植物地上部分能够拦截海水中的泥沙及颗粒有机质, 帮助盐沼湿地快速抬升高程, 增大其有机碳的储存空间; 同时, 植物地上部分是植物作为初级生产者固定光合碳的器官, 其生物量大小对滨海盐沼湿地的初级生产力具有直接影响.植物地下部分一方面能够分泌根际沉积物, 是土壤可溶性有机碳等易分解碳的重要来源; 另一方面, 植物地下根系能够锚定沉积物, 防止潮汐作用对有机质的侵蚀等(Kirwan & Patrick Megonigal, 2013; Mao et al., 2018 ).因此, 植物生物量的空间分配能够反映近期光合同化的新碳向根系和土壤碳库的转移规律, 是土壤碳输入和碳埋藏的重要参考指标, 深刻影响着土壤碳库的变化(Bolinderet al., 2012 ).通常情况下, 植物光合固定的碳即时分配大小为茎叶>根>土壤, 大部分光合固定碳都留在地上部分, 氮增加则会更加提高植物同化碳向植物地上部分的流动, 同时显著增加根际土壤中碳的累积与回收率, 而降低植物根系中的光合碳分配(王婷婷等, 2017).随着同位素技术的发展和应用, 通过测定土壤或植物中的碳同位素自然丰度值可以量化光合碳在植物-土壤系统中的动态变化、周转规律及其对环境变化的响应(Wang et al., 2019 ; Xiaoet al., 2019 ).例如, 13C脉冲标记实验表明, 适量氮输入显著提高即时光合碳分配给根系的比例(Wang et al., 2019 ).氮输入通常刺激植物生长早期同化碳向土壤方向迁移, 但生长后期可能降低, 这种由生长阶段造成的差异使得光合碳在根系中的分配模式对氮输入的响应具有一定的不确定性(王婷婷等, 2017; Xiao et al., 2019 ). ...
... ; Xiao et al., 2019 ). ...
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
... 氮输入对盐沼湿地土壤有机碳分解存在不确定性, 或促进、或抑制或不显著.研究发现, 氮输入可以促进湿地土壤CO2和CH4排放(Fang et al., 2017 ; Herbert et al., 2020 ), 但是在氮输入量很高的情况下, 土壤CO2排放量会逐渐趋向饱和, 甚至受到抑制(Xiao et al., 2019 ).氮输入可以促进土壤CO2排放, 这是由于氮可能促进土壤酶活性, 降低氮对生物代谢的胁迫和限制, 同时改善凋落物质量(Bragazza et al., 2006 ; Song et al., 2013 ; Fang et al., 2017 ).但是, 土壤微生物也会受到氮、磷等变化范围的限制.在施氮量较高的情况下, 土壤CO2排放量会呈现饱和趋势(Bragazza et al., 2006 ).也有研究发现, 氮输入对土壤有机碳分解和土壤CO2排放的影响可能是中性的(Chen et al., 2017 ), 这可能是由于碳对土壤微生物的限制作用(Song et al., 2010 ), 也可能是因为氮输入后会刺激植被的自养呼吸, 但却通过抑制有机质分解降低其异养呼吸(Högberget al., 2010 ; Wang et al., 2014 ).值得注意的是, 由于土壤肥力的差异, 光合碳输入对根际土壤的“激发效应”产生或正或负的影响, 从而调节土壤有机碳分解和碳埋藏过程(Pausch & Kuzyakov, 2018).另外, 氮输入通过影响微生物群落结构和功能进而影响CH4产生(Sinsabaugh et al., 2015 ).CH4由产甲烷古菌生成(Angel et al., 2012 ), 探究产甲烷古菌的反应可能是了解土壤CH4排放对氮输入响应的关键(Xiao et al., 2017 ).铵态氮和硝态氮由于生物化学性质的差异, 它们对有机碳矿化的影响也不同(Chen et al., 2017 , 2018).目前大量研究揭示了氮输入量和土壤有机碳分解的关系, 但是有关氮类型对土壤有机碳稳定性的影响研究较少.例如, 相对NO3-, Juncus acutiflorus湿地优先选择NH4+作为氮源, 导致根际中NO3-过剩, 改变了根际氮循环动态, 从而有利于提高微生物的种类和数量(Hester et al., 2018 ).当氮输入量较高时, 土壤微生物可能会优先选择能耗更低的铵态氮; 添加NH4+和硝酸铵会减少CO2排放, 但也有研究发现添加NO3-对土壤CO2排放影响不大(Min et al., 2011 ).过量的NO3-会促进厌氧呼吸, 过量的NH4+会抑制CH4氧化, 可能会导致CH4排放不稳定(Hester et al., 2018 ).我们前期在黄河三角洲4年的野外控制实验发现, 铵态氮在全年均提高了CH4排放; 硝态氮虽然在淹水期对CH4排放具有促进作用, 但影响较小(Xiao et al., 2017 ).另外, 我们还发现, 铵态氮对土壤呼吸有显著促进作用, 但是硝态氮施加对年平均土壤呼吸速率没有显著影响(Qu et al., 2020 ).铵态氮和硝态氮对土壤有机碳分解的影响不同, 这种差异也可能是土壤pH和酶活性的响应不同导致的(Min et al., 2011 ). ...
Human eutrophication drives biogeographic salt marsh productivity patterns in China 2 2020
... 陆源氮通过地表径流进入近海, 导致近岸海域富营养化日益加剧(Deegan et al., 2012 ; Xu et al., 2020 ), 是目前全球海岸带面临的最为严重的一个环境问题.在潮汐作用下, 大量氮必然会进入盐沼等滨海湿地生态系统, 并被盐沼植被截留, 进而改变其光合碳输入、植物-土壤碳分配、土壤有机碳分解和土壤可溶性有机碳释放等碳循环关键过程(Hu et al., 2016 ; 韩广轩, 2017; Herbert et al., 2020 ), 并对其碳汇功能产生广泛且深刻的影响(图1).相比工业化之前, 近海水体氮负荷持续加重, 全球范围内由陆地向海洋输出的氮至少增加了10倍, 且主要以无机氮(DIN)为主要氮类型(Deegan et al., 2012 ; Breitburg et al., 2018 ).监测及模拟结果表明, 2000-2050年间, 中国沿海总氮输入量将增加30%-200% (Strokal et al., 2014 ; Wang et al., 2018 ).因此, 阐明氮输入对盐沼湿地碳循环关键过程的影响机制, 将有助于揭示氮输入对盐沼湿地蓝色碳汇形成过程与机制的影响, 并为预测近岸海域富营养化背景下盐沼湿地碳库的潜在变化趋势提供科学依据. ...
... 综上所述, 近岸水体富营养化背景下, 大量氮输入对盐沼湿地植物光合碳输入、植物-土壤碳分配和土壤碳输出等碳循环关键过程产生深刻影响, 进而影响其碳汇功能的准确评估.首先, 氮输入对湿地植物光合固碳的影响存在阈值效应(Vivanco et al., 2015 ; Peng et al., 2019 ).低水平氮输入可以增强植物光合固碳能力, 但是达到一定阈值后, 继续加氮可能抑制这种正效应(Peng et al., 2019 ).其次, 氮输入影响植物光合产物在植物-土壤系统中的分配比例.光合固定碳向根系和土壤碳库的分配和转移规律, 深刻影响着土壤碳库的走向(Xu et al., 2020 ).再次, 氮输入会影响土壤有机碳的分解, 但是影响结果与氮输入类型和水平、土壤初始碳氮含量、施氮时间尺度等有很大的关系, 研究结论甚至截然相反(Chen et al., 2017 ; Zhou et al., 2017 ).另外, 氮输入对盐沼湿地土壤DOC的产生和累积有显著影响(Deegan et al., 2012 ; Chang et al., 2018 ), 进而影响盐沼湿地横向碳流失(韩广轩, 2017).最后, 氮输入在提高植被固碳潜力的同时, 也会促进土壤, 尤其是深层土壤中有机碳分解(Macket al., 2004 ).因此, 氮输入对盐沼湿地碳循环过程的影响存在不确定性, 无疑将在很大程度上制约着对盐沼湿地碳循环和碳汇功能的整体理解以及水体富营养化背景下盐沼湿地“蓝碳”吸存能力的评估和预测. ...
Substrate and environmental controls on microbial assimilation of soil organic carbon: a framework for Earth system models 1 2014
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
Effects of continuous nitrogen addition on microbial properties and soil organic matter in a Larix gmeliniiplantation in China 1 2018
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
Response of soil characteristics and bacterial communities to nitrogen fertilization gradients in a coastal salt-affected agroecosystem 1 2021
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
Patterns and mechanisms of responses by soil microbial communities to nitrogen addition 3 2017
... 氮输入可通过影响植物生长、根系活动、凋落物分解、微生物特性等, 进而影响土壤有机碳分解.盐沼湿地作为一个巨大的蓝色碳汇, 其碳存储主要以土壤有机碳的形式存在(Macreadie et al., 2019 ).氮输入影响植物生长和碳分配, 光合作用产物由叶片输送到细根, 影响细根生长及土壤呼吸.我们前期的研究发现, 盐沼湿地植物冠层光合作用在日尺度上对土壤呼吸动态变化具有明显的调节作用(Han et al., 2014 ).同时, 土壤微生物过程直接或间接受到氮输入的影响(图2).氮输入通过改变土壤氮有效性, 调节土壤微生物生长、活性、群落组成/多样性及酶活性, 进而对土壤有机碳分解产生影响(Zhou et al., 2017 ; Yang et al., 2018 ; Xiao et al., 2019 ; Qu et al., 2020 ).一方面, 氮输入可以通过提高土壤中碳、氮的含量从而改变土壤微生物生物量和微生物活性, 导致土壤有机质组分分解的变化.当底物中的氮含量较高时, 施氮更容易使微生物大量聚集, 形成较为稳定的土壤有机质; 当底物中氮含量较低时, 施氮通常促进土壤呼吸作用, 土壤中的碳不容易留存下来(Manzoni et al., 2012 ; Cotrufo et al., 2013 ; Xu et al., 2014 ).另一方面, 氮输入可能会降低土壤pH, 调动土壤中的铝, 从而抑制微生物活性.氮输入可以降低真菌和细菌的比例(F:B), 同时也会引起土壤酸化(提高F:B)(Rousk et al., 2010 ; Chen et al., 2015a ).另外, 土壤酶的功能通常分为氧化或者水解, 氧化酶降解木质素等难降解的化合物, 而水解酶降解纤维素等简单的化合物(Sinsabaugh & Moorhead, 1994).较高的矿质氮输入可以促进纤维素的分解, 但也会抑制难降解的木质素有机物的分解, 从而延缓惰性有机质分解. ...
... 同时, 潮汐盐沼湿地的氮有效性也会影响土壤有机质分解(Romero et al., 2005 ; Huxham et al., 2010 ), 并对促进微生物分解植物碎屑很重要(Romero et al., 2005 ), 但过高的氮输入也会抑制植物根系及土壤的碳分解.因此, 氮添加充足时, 氮输入抑制了有机质分解, 同时考虑到植被生产力的提高, 从而可能会提高潮汐盐沼湿地土壤碳汇能力(Janssens et al., 2010 ; Keuskamp et al., 2015 ).因而, 评估氮输入对盐沼湿地碳汇功能时应当区分氮限制生态系统和富氮生态系统(Chen et al., 2015b ), 目前大多数潮汐盐沼湿地都是贫氮生态系统(Mou et al., 2011 ).对于贫氮生态系统, 氮输入能刺激植被生长固碳, 改善土壤养分、生化特性和代谢活性, 使其更适合于微生物生境(Zhou et al., 2017 ; Yaoet al., 2021 ), 增加土壤有机碳含量, 提高有机碳分解速率.例如, 位于美国马萨诸塞州东北部的一个长达9年的研究通过野外控制实验发现近岸水体富营养化导致的长期大量氮输入可能造成生态系统氮饱和, 从而制约潮汐盐沼湿地土壤的“蓝碳”功能(Deegan et al., 2012 ). ...
... 综上所述, 近岸水体富营养化背景下, 大量氮输入对盐沼湿地植物光合碳输入、植物-土壤碳分配和土壤碳输出等碳循环关键过程产生深刻影响, 进而影响其碳汇功能的准确评估.首先, 氮输入对湿地植物光合固碳的影响存在阈值效应(Vivanco et al., 2015 ; Peng et al., 2019 ).低水平氮输入可以增强植物光合固碳能力, 但是达到一定阈值后, 继续加氮可能抑制这种正效应(Peng et al., 2019 ).其次, 氮输入影响植物光合产物在植物-土壤系统中的分配比例.光合固定碳向根系和土壤碳库的分配和转移规律, 深刻影响着土壤碳库的走向(Xu et al., 2020 ).再次, 氮输入会影响土壤有机碳的分解, 但是影响结果与氮输入类型和水平、土壤初始碳氮含量、施氮时间尺度等有很大的关系, 研究结论甚至截然相反(Chen et al., 2017 ; Zhou et al., 2017 ).另外, 氮输入对盐沼湿地土壤DOC的产生和累积有显著影响(Deegan et al., 2012 ; Chang et al., 2018 ), 进而影响盐沼湿地横向碳流失(韩广轩, 2017).最后, 氮输入在提高植被固碳潜力的同时, 也会促进土壤, 尤其是深层土壤中有机碳分解(Macket al., 2004 ).因此, 氮输入对盐沼湿地碳循环过程的影响存在不确定性, 无疑将在很大程度上制约着对盐沼湿地碳循环和碳汇功能的整体理解以及水体富营养化背景下盐沼湿地“蓝碳”吸存能力的评估和预测. ...
 ,
, ,*, 李隽永, 屈文笛中国科学院烟台海岸带研究所, 中国科学院海岸带环境过程与生态修复重点实验室, 山东烟台 264003; 中国科学院大学, 北京 100049
,*, 李隽永, 屈文笛中国科学院烟台海岸带研究所, 中国科学院海岸带环境过程与生态修复重点实验室, 山东烟台 264003; 中国科学院大学, 北京 100049

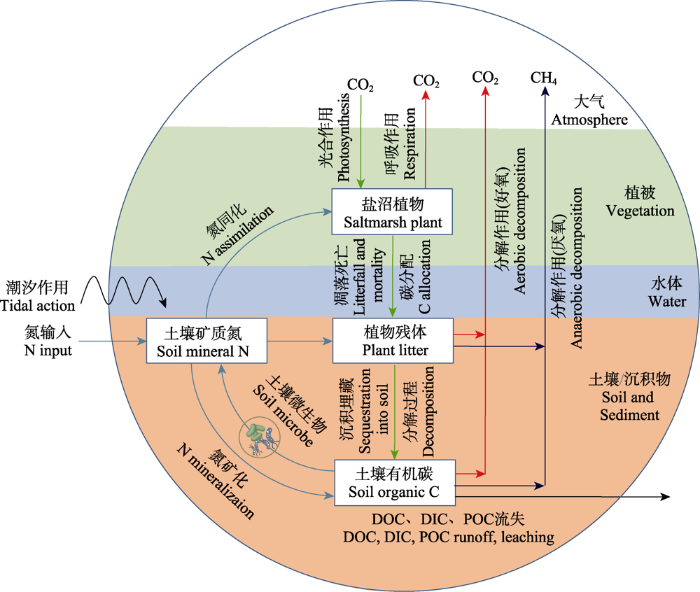 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT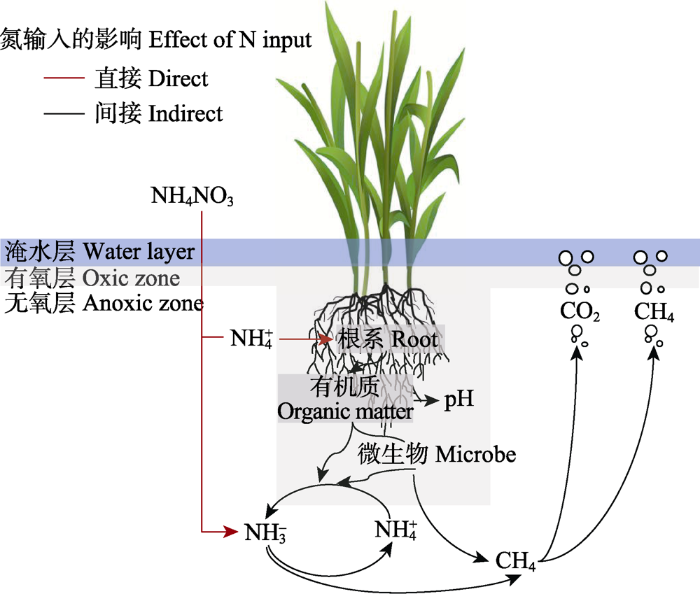 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT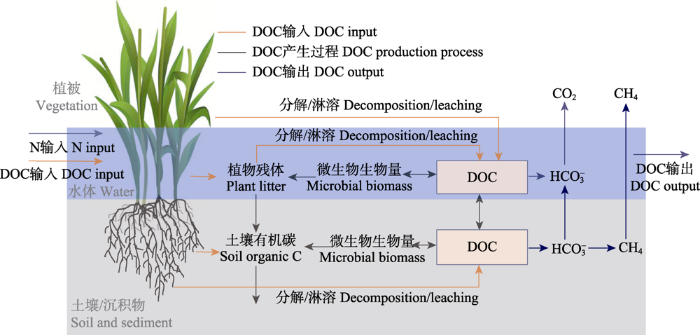 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT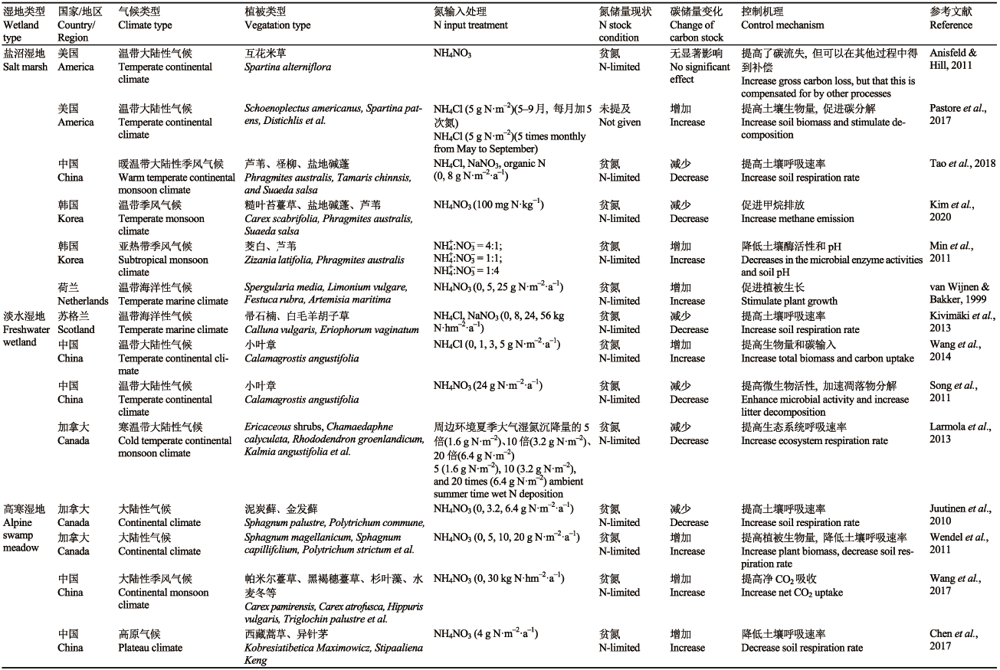


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}