Linkages of aboveground plant carbon accumulation rate with ecosystem multifunctionality in alpine grassland, Qingzang Plateau
Jian SUN,,1,*, Yi WANG1,2, Guo-Hua LIU31Key Laboratory of Observation and Simulation of Ecological Networks, Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 2College of Earth Sciences, Chengdu University of Technology, Chengdu 610059, China 3State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
Abstract Aims As one of the major terrestrial ecosystems of the world, a small fluctuation of grassland soil carbon (C) would affect the carbon cycle of the terrestrial ecosystem and ecosystem multifunctionlity (EMF). The carbon accumulation rate (CAR) of aboveground community well reflects the capacity and efficiency of carbon sequestration in a field from the start to the peak of a growing season. The changes in plant CAR could influence the ability of above- and below-ground community. Currently, the majority of studies have primarily focused on the relationship between community diversity and EMF, while the linkages of CAR with EMF were understudied. We aimed to explore the process and underlying mechanism of how CAR affecting EMF in alpine grassland community. Our results would improve the understanding of EMF maintenance mechanism and provide theoretical support for alpine ecosystem management. Methods We conducted a field transect survey which consists of a total of 115 sample sites of alpine grasslands on the Qingzang Plateau from July to August 2015. The ecosystem multifunctionality index (M) was calculated from 13 key ecosystem parameters including soil organic carbon content, total nitrogen content, total phosphorus content above- and belowground biomass etc. The normalized difference vegetation index (NDVI, 1982-2013) was adopted to obtain the phenology in 2015. We calculated the CAR value. To explore the underlying mechanism of how CAR affecting EMF, the annual total precipitation and temperature were extracted by the method of thin disk smooth spline interpolation based on observations of meteorological stations from 2011-2015. Important findings Belowground biomass, soil organic carbon content, total phosphorus content and microbial biomass carbon content had high weighting for CAR (0.58, 0.80, 0.83 and 0.79) and M (1.05, 0.98, 1.02 and 0.97). There was a significantly positive correlation between CAR and M (R2 = 0.45, p < 0.01). Our findings suggested that the synergism of plant community and soil elements affected CAR and further regulated EMF under the influences of precipitation and temperature. Keywords:ecosystem multifunctionlity;carbon accumulation rate;alpine grassland;soil organic carbon;Qingzang Plateau
PDF (1584KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 孙建, 王毅, 刘国华. 青藏高原高寒草地地上植物碳积累速率对生态系统多功能性的影响机制. 植物生态学报, 2021, 45(5): 496-506. DOI: 10.17521/cjpe.2020.0180 SUN Jian, WANG Yi, LIU Guo-Hua. Linkages of aboveground plant carbon accumulation rate with ecosystem multifunctionality in alpine grassland, Qingzang Plateau. Chinese Journal of Plant Ecology, 2021, 45(5): 496-506. DOI: 10.17521/cjpe.2020.0180
近年来, 在全球气候变化和人类活动的影响下, ****围绕草地生态系统功能以及物种多样性展开了一系列的研究, 但是多数研究主要集中于某种生态系统功能与驱动因子的关系(Wu et al., 2014)。随着研究的深入, 研究者逐渐在不同时间、空间、生境、土壤和气候条件下探讨生态系统多功能性(EMF)的变化(Zavaleta et al., 2010; Lefcheck et al., 2015)。如何量化人类活动、气候变化及其他环境因子对生态系统多功能性的影响, 以及这些驱动因子对多个生态系统功能的影响成为研究的热点(Byrnes et al., 2014)。特别是在全球气候变暖的背景下, 草地碳库作为陆地生态系统碳库的重要组成部分, 其较小幅度的波动, 将会影响整个陆地生态系统碳循环和EMF。因此, 深入研究草地植被固碳功能和固碳潜力对于适应和减缓气候变化、维持草地EMF具有重要意义。
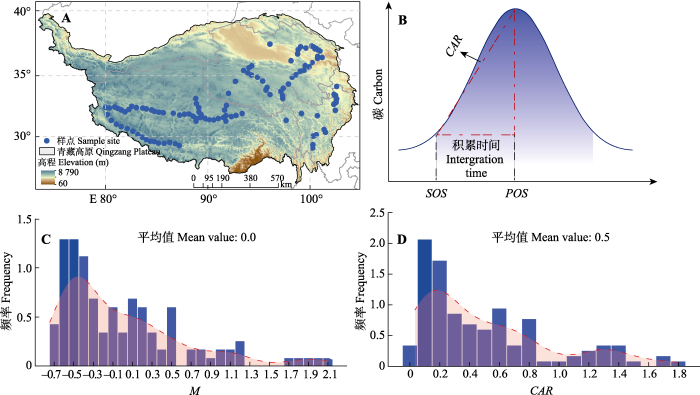
Fig. 1Sample sites in the study area (A), definition of carbon accumulation rate (CAR): the biomass production of plants from the timing of start of growing season (SOS) to the timing of the peak of growing season (POS)(B), frequency distribution of ecosystem multifunctionality index (M)(C), and frequency distribution of CAR (D).
物候数据来自Global Inventory Modelling and Mapping Studies (GIMMS)规范化差异植被指数第3版数据库(Gonsamo et al., 2018)。其中, 归一化植被指数(NDVI, 1982-2013年)是从Advanced Very High Resolution Radiometer (AVHRR)传感器获取的基于卫星的地表反射率数据中获得的。通过计算1982-2013年高原植物物候指标确定生长季初始时间(SOS)和生长季高峰时间(POS), 使用线性回归分析得到了2015年的物候指标。在本研究中, 首先采用傅立叶级数模型对NDVI数据集进行平滑(Wang et al., 2018):
Fig. 2Bivariate plots using linear mixed-effect models depicting the relationships of ecosystem multifunctionality index (M)(A) and carbon accumulation rate (CAR)(B) with ecosystem parameters of aboveground biomass (AGB), belowground biomass (BGB), leaf carbon (LC), leaf nitrogen (LN), leaf phosphorus (LP), microbial biomass carbon (MBC), microbial biomass nitrogen (MBN), soil available nitrogen (SAN), soil available phosphorus (SAP), soil organic carbon (SOC), soil total nitrogen (STN), soil total phosphorus (STP) content and soil water content (SWC)(mean ± SD). Except for EMF, other data were normalized.
Fig. 4Relationships between ecosystem multifunctionality index (M), carbon accumulation rate (CAR) and different ecosystem parameters. Belowground biomass (BGB), soil organic carbon (SOC), soil total phosphorus (STP) and microbial biomass carbon (MBC) content are ln-transformed data. The shade part indicates the 95% confidence interval.
Fig. 5Influence of annual total precipitation (ATP) and annual mean temperature (AMT) to ecosystem multifunctionality index (M) and carbon accumulation rate (CAR). ATP is ln-transformed data and AMT is (AMT + 12 °C) ln-transformed data. The shaded part indicates the 95% confidence interval.
Fig. 6Direct and indirect impacts of climatic and key factors on ecosystem multifunctionality index (M) and carbon accumulation rate (CAR). Path with significant effect is shown in the figure (p < 0.05), solid line represents direct effect and dotted line represents indirect effect; black arrow indicates positive effect and red arrow indicates negative effect. AMT, annual mean temperature; ATP, annual total precipitation; BGB, belowground biomass; MBC, microbial biomass carbon content; SOC, soil organic carbon content; STP, soil total nitrogen content.
3 讨论
3.1 生态系统参数对CAR和EMF的影响
本研究选择与生态系统碳、氮和磷循环相关的13种生态系统参数来表征生态系统多功能, 这些参数与植被生产力、养分循环、土壤有机碳蓄积等密切相关, 能够代表生态系统多功能性(Maestre et al., 2012)。研究发现, 植物叶片碳、氮和磷含量对EMF无显著效应, 对CAR有显著负效应(图2), 这与前人的研究结果(Jing et al., 2015)不一致。一般而言, 叶片的N和P特征是植物长期适应外界环境条件的结果, 这说明植物群落叶片的氮、磷含量可以适应环境因子的变动而迅速调节, 并没有形成较为稳定的元素特征关系, 这一点或许也可以解释为什么本研究植物叶片碳、氮和磷含量对EMF无显著影响(杨阔等, 2010)。而植物叶片C:N可指示植物对土壤因子等环境变化的适应性, 表征植物吸收营养物质同化碳的能力和反映植物对营养的利用效率(Thompson et al., 1997)。因此, 在低的养分条件下, 植物的生长虽然缓慢, 但养分利用效率较高, 具有高的C:N, 叶片碳含量高, 而叶片氮磷含量低(李丹等, 2016)。本研究发现植物叶片氮磷含量对CAR有显著负效应, 符合上述观点。
BaiJH,DengW,ZhangYX(2002).Spatial distribution of soil organic matter and nitrogen in soil of circular-zonary vegetation areas in Wulanpao Wetland, Inner Mongolia .Journal of Lake Science,14, 145-151. [本文引用: 1]
BaiY,WuJ,ClarkCM,NaeemS,PanQ,HuangJ,ZhangL,HanX(2010).Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from Inner Mongolia Grasslands .Global Change Biology,16, 358-372. DOI:10.1111/(ISSN)1365-2486URL [本文引用: 1]
BalachowskiJA,VolaireFA(2018).Implications of plant functional traits and drought survival strategies for ecological restoration .Journal of Applied Ecology,55, 631-640. DOI:10.1111/1365-2664.12979URL [本文引用: 1]
BaoSD(2000).Soil and Agricultural Chemistry Analysis.China Agriculture Press,Beijing. [本文引用: 1]
BaumannF,HeJS,SchmidtK,KühnP,ScholtenT(2009).Pedogenesis, permafrost, and soil moisture as controlling factors for soil nitrogen and carbon contents across the Tibetan Plateau .Global Change Biology,15, 3001-3017. DOI:10.1111/j.1365-2486.2009.01953.xURL [本文引用: 1]
BeheraN,SahaniU(2003).Soil microbial biomass and activity in response to Eucalyptus plantation and natural regeneration on tropical soil .Forest Ecology and Management,174, 1-11. DOI:10.1016/S0378-1127(02)00057-9URL [本文引用: 1]
ByrnesJEK,GamfeldtL,IsbellF,LefcheckJS,GriffinJN,HectorA,CardinaleBJ,HooperDU,DeeLE,Emmett DuffyJ(2014).Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions .Methods in Ecology and Evolution,5, 111-124. DOI:10.1111/2041-210X.12143URL [本文引用: 1]
Chapin IIIFS,MatsonPA,MooneyHA(2012).Principles of Terrestrial Ecosystem Ecology .Springer, San Francisco, USA. [本文引用: 1]
ChenZQ,ShaoQQ,LiuJY,WangJB(2012).Analysis of net primary productivity of terrestrial vegetation on the Qinghai- Tibetan Plateau based on MODIS remote sensing data .Science China: Earth Scienses,42, 402-410. [本文引用: 1]
FuYH,PiaoS,Op de BeeckM,CongN,ZhaoHF,ZhangY,MenzelA,JanssensIA(2014).Recent spring phenology shifts in western Central Europe based on multiscale observations .Global Ecology and Biogeography,23, 1255-1263. DOI:10.1111/geb.12210URL [本文引用: 1]
HuxmanTE,SmithMD,FayPA,KnappAK,ShawMR,LoikME,SmithSD,TissueDT,ZakJC,WeltzinJF,PockmanWT,SalaOE,HaddadBM,HarteJ,KochGW,SchwinningS,SmallEE,WilliamsDG(2004).Convergence across biomes to a common rain-use efficiency .Nature,429, 651-654. DOI:10.1038/nature02561URL [本文引用: 1]
JingX,SandersNJ,ShiY,ChuH,ClassenAT,ZhaoK,ChenL,ShiY,JiangY,HeJS(2015).The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate .Nature Communications,6, 8159. DOI:10.1038/ncomms9159. DOI:10.1038/ncomms9159PMID:26328906 [本文引用: 6] Jing, Xin; Zhao, Ke; Shi, Yue; He, Jin-Sheng Peking Univ, Dept Ecol, Coll Urban & Environm Sci, Beijing 100871, Peoples R China. Jing, Xin; Zhao, Ke; Shi, Yue; He, Jin-Sheng Peking Univ, Key Lab Earth Surface Proc, Minist Educ, Beijing 100871, Peoples R China. Sanders, Nathan J. Univ Copenhagen, Ctr Macroecol Evolut & Climate, Nat Hist Museum Denmark, DK-2100 Copenhagen, Denmark. Shi, Yu; Chu, Haiyan Chinese Acad Sci, Inst Soil Sci, State Key Lab Soil & Sustainable Agr, Nanjing 210008, Peoples R China. Classen, Aimee T. Univ Copenhagen, Nat Hist Museum Denmark, DK-2100 Copenhagen, Denmark. Chen, Litong; He, Jin-Sheng Chinese Acad Sci, Northwest Inst Plateau Biol, Key Lab Adaptat & Evolut Plateau Biota, Xining 810008, Peoples R China. Shi, Yue Chinese Acad Sci, Inst Bot, Beijing 100093, Peoples R China. Jiang, Youxu Chinese Acad Forestry, Inst Forest Ecol, Beijing 100091, Peoples R China.
KeelingHC,PhillipsOL(2007).The global relationship between forest productivity and biomass .Global Ecology and Biogeography,16, 618-631. DOI:10.1111/geb.2007.16.issue-5URL [本文引用: 1]
KleinJA,HarteJ,ZhaoXQ(2004).Experimental warming causes large and rapid species loss, dampened by simulated grazing, on the Tibetan Plateau .Ecology Letters,7, 1170-1179. DOI:10.1111/ele.2004.7.issue-12URL [本文引用: 1]
LambersH,RavenJA,ShaverGR,SmithSE(2008).Plant nutrient-acquisition strategies change with soil age .Trends in Ecology & Evolution,23, 95-103. DOI:10.1016/j.tree.2007.10.008URL [本文引用: 1]
LefcheckJS,ByrnesJEK,IsbellF,GamfeldtL,GriffinJN,EisenhauerN,HenselMJS,HectorA,CardinaleBJ,DuffyJE(2015).Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats .Nature Communications,6, 6936. DOI:10.1038/ncomms7936. DOI:10.1038/ncomms7936URL [本文引用: 1]
LiCB,PengYF,ZhaoDZ,NingY,ZhouGY(2016).Effects of precipitation change and nitrogen addition on community structure and plant diversity in an alpine steppe on the Qinghai-Tibetan Plateau .Research of Soil and Water Conservation,23, 185-191. [本文引用: 1]
LiD,KangS,ZhaoMY,ZhangQ,RenHJ,RenJ,ZhouJM,WangZ,WuRJ,NiuJM(2016).Relationships between soil nutrients and plant functional traits in different degradation stages of Leymus chinensis steppe in Nei Mongol, China .Chinese Journal of Plant Ecology,40, 991-1002. DOI:10.17521/cjpe.2015.0465URL [本文引用: 1]
LiL,GaoJQ,LeiGC,LüC,SuoL(2011).Distribution patterns of soil organic carbon and total nitrogen in Zoige peat land with different ground water table .Chinese Journal of Ecology,30, 2449-2455. [本文引用: 1]
LiXD,LiFX,ZhouBR,XiaoHB,YangXG,ZhouWF(2012).Study of the hydrothermal condition and aboveground biomass in typical alpine grassland in Tibetan Plateau .Plateau Meteorology,31, 1053-1058. [本文引用: 1]
LiuLL,JinZX,LiJH(2010).Plant species diversity in Sinocalycanthus chinensis community and its correlation with soil factors in Dalei Mountain of Zhejiang Province .Bulletin of Botanical Research,30, 57-64. [本文引用: 1]
MaestreFT,QueroJL,GotelliNJ,EscuderoA,OchoaV,Delgado-BaquerizoM,García-GómezM,BowkerMA,SoliveresS,EscolarC,García-PalaciosP,BerdugoM,ValenciaE,GozaloB,GallardoA,et al.(2012).Plant species richness and ecosystem multifunctionality in global drylands .Science,335, 214-218. DOI:10.1126/science.1215442URL [本文引用: 4]
PiaoS,CiaisP,FriedlingsteinP,PeylinP,ReichsteinM,LuyssaertS,MargolisH,FangJ,BarrA,ChenA,GrelleA,HollingerDY,LaurilaT,LindrothA,RichardsonAD,VesalaT(2008).Net carbon dioxide losses of northern ecosystems in response to autumn warming .Nature,451, 49-52. DOI:10.1038/nature06444URL [本文引用: 1]
SoliveresS,MaestreFT,EldridgeDJ,Delgado-BaquerizoM,QueroJL,BowkerMA,GallardoA(2014).Plant diversity and ecosystem multifunctionality peak at intermediate levels of woody cover in global drylands .Global Ecology and Biogeography,23, 1408-1416. PMID:25914607 [本文引用: 1] The global spread of woody plants into grasslands is predicted to increase over the coming century. While there is general agreement regarding the anthropogenic causes of this phenomenon, its ecological consequences are less certain. We analyzed how woody vegetation of differing cover affects plant diversity (richness and evenness) and multiple ecosystem functions (multifunctionality) in global drylands, and how this changes with aridity.224 dryland sites from all continents except Antarctica widely differing in their environmental conditions (from arid to dry-subhumid sites) and woody covers (from 0 to 100%).Using a standardized field survey, we measured the cover, richness and evenness of perennial vegetation. At each site, we measured 14 ecosystem functions related to soil fertility and the build-up of nutrient pools. These functions are critical for maintaining ecosystem function in drylands.Species richness and ecosystem multifunctionality were strongly influenced by woody vegetation, with both variables peaking at relative woody covers (RWC) of 41-60%. This relationship shifted with aridity. We observed linear positive effects of RWC in dry-subhumid sites. These positive trends shifted to hump-shaped RWC-diversity and multifunctionality relationships under semiarid environments. Finally, hump-shaped (richness, evenness) or linear negative (multifunctionality) effects of RWC were found under the most arid conditions.Plant diversity and multifunctionality peaked at intermediate levels of woody cover, although this relationship became increasingly positive under wetter environments. This comprehensive study accounts for multiple ecosystem attributes across a range of woody covers and environmental conditions. Our results help us to reconcile contrasting views of woody encroachment found in current literature and can be used to improve predictions of the likely effects of encroachment on biodiversity and ecosystem services.
SunHL,ZhengD,YaoTD,ZhangYL(2012).Protection and construction of the national ecological security shelter zone on Tibetan Plateau .Acta Geographica Sinica,67, 3-12. [本文引用: 1]
SunJ,ChengGW,LiWP,ShaYK,YangYC(2013).On the variation of NDVI with the principal climatic elements in the Tibetan Plateau .Remote Sensing,5, 1894-1911. DOI:10.3390/rs5041894URL [本文引用: 1]
SunJ,MaB,LuX(2018).Grazing enhances soil nutrient effects: trade-offs between aboveground and belowground biomass in alpine grasslands of the Tibetan Plateau .Land Degradation & Development,29, 337-348. DOI:10.1002/ldr.v29.2URL [本文引用: 1]
SunJ,ZhangZC,DongSK(2019).Adaptive management of alpine grassland ecosystems over Tibetan Plateau .Pratacultural Science,36, 933-938. [本文引用: 1]
SunJ,ZhouTC,LiuM,ChenYC,LiuGH,XuM,ShiPL,PengF,TsunekawaA,LiuY,WangXD,DongSK,ZhangYJ,LiYN(2020).Water and heat availability are drivers of the aboveground plant carbon accumulation rate in alpine grasslands on the Tibetan Plateau .Global Ecology and Biogeography,29, 50-64. DOI:10.1111/geb.v29.1URL [本文引用: 10]
ThompsonK,ParkinsonJA,BandSR,SpencerRE(1997).A comparative study of leaf nutrient concentrations in a regional herbaceous flora .New Phytologist,136, 679-689. DOI:10.1046/j.1469-8137.1997.00787.xURL [本文引用: 1]
van der HeijdenMGA,KlironomosJN,UrsicM,MoutoglisP,Streitwolf-EngelR,BollerT,WiemkenA,SandersIR(1998).Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity .Nature,396, 69-72. DOI:10.1038/23932URL [本文引用: 1]
WangJ,ZhouTC,PengPH(2018).Phenology response to climatic dynamic across China’s grasslands from 1985 to 2010 .ISPRS International Journal of Geo-Information,7, 290. DOI:10.3390/ijgi7080290. DOI:10.3390/ijgi7080290URL [本文引用: 1]
WangY,LiuBY,LiuM,SunJ,ZengT(2019).Synergistic and inhibitory effects of soil enzymes along desertified gradients of the Zoige alpine meadow .Pratacultural Science,36, 939-951. [本文引用: 2]
WangYH,SongXH,WangZW,KangJ,HanGD,WangZW(2018).Responses of plant above and underground productivity of Stipa breviflora desert steppe to stocking rates and precipitation .Acta Botanica Boreali-Occidentalia Sinica,38, 1526-1533. [本文引用: 1]
WuJ,ShenZ,ZhangX(2014).Precipitation and species composition primarily determine the diversity-productivity relationship of alpine grasslands on the Northern Tibetan Plateau .Alpine Botany,124, 13-25. DOI:10.1007/s00035-014-0125-zURL [本文引用: 1]
XiaJ,NiuS,CiaisP,JanssensIA,ChenJ,AmmannC,ArainA,BlankenPD,CescattiA,BonalD,BuchmannN,CurtisPS,ChenSP,DongJW,FlanaganLB,et al.(2015).Joint control of terrestrial gross primary productivity by plant phenology and physiology .Proceedings of the National Academy of Sciences of the United States of America,112, 2788-2793. [本文引用: 1]
XiongDP,ZhaoGS,WuJS,ShiPL,ZhangXZ(2016).The relationship between species diversity and ecosystem multifunctionality in alpine grasslands on the Tibetan Changtang Plateau .Acta Ecologica Sinica,36, 3362-3371. [本文引用: 2]
YanZQ,QiYC,PengQ,DongYS,HeYL,LiZL(2017).Advances in the effects of simulated precipitation and nitrogen deposition on grassland biomass .Acta Agrestia Sinica,25, 1165-1170. [本文引用: 1]
YangH,LIY,WUM,ZhangZ,LIL,WanS(2011).Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits .Global Change Biology,17, 2936-2944. DOI:10.1111/gcb.v17.9URL [本文引用: 1]
YangK,HuangJH,DongD,MaWH,HeJS(2010).Canopy leaf N and P stoichiometry in grassland communities of Qinghai-Tibetan Plateau, China .Chinese Journal of Plant Ecology,34, 17-22. [本文引用: 1]
YangYH,PiaoSL(2006).Variations in grassland vegetation cover in relation to climatic factors on the Tibetan Plateau .Acta Phytoecologica Sinica,30, 1-8. [本文引用: 1]
YuH,LuedelingE,XuJ(2010).Winter and spring warming result in delayed spring phenology on the Tibetan Plateau .Proceedings of the National Academy of Sciences of the United States of America,107, 22151-22156. [本文引用: 1]
YuanZY,ChenHYH(2015).Decoupling of nitrogen and phosphorus in terrestrial plants associated with global changes .Nature Climate Change,5, 465-469. DOI:10.1038/nclimate2549URL [本文引用: 1]
ZavaletaES,PasariJR,HulveyKB,TilmanGD(2010).Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity .Proceedings of the National Academy of Sciences of the United States of America,107, 1443-1446. [本文引用: 1]
ZhangBB,LiuF,DingJZ,FangK,YangGB,LiuL,ChenYL,LiF,YangYH(2016).Soil inorganic carbon stock in alpine grasslands on the Qinghai-Xizang Plateau: an updated evaluation using deep cores .Chinese Journal of Plant Ecology,40, 93-101. DOI:10.17521/cjpe.2015.0406URL [本文引用: 1]
ZhangG,ZhangY,DongJ,XiaoX(2013).Green-up dates in the Tibetan Plateau have continuously advanced from 1982 to 2011 .Proceedings of the National Academy of Sciences of the United States of America,110, 4309-4314. [本文引用: 1]
ZhangL,HaoBT,QiLX,LiYL,XuHM,YangLN,BaoY(2018).Dynamic responses of aboveground biomass and soil organic matter content to grassland restoration .Chinese Journal of Plant Ecology,42, 317-326. DOI:10.17521/cjpe.2017.0046URL [本文引用: 2]
ZhangX,MeiL,SongLH,LiuLC,ZhaoZY(2019).Effects of simulated nitrogen deposition on microbial community and greenhouse gases emission of Pinus massoniana soil .Acta Ecologica Sinica,39, 1917-1925. [本文引用: 2]
ZhaoDD,MaHY,LiY,WeiJP,WangZC(2019).Effects of water and nutrient additions on functional traits and aboveground biomass of Leymus chinensis .Chinese Journal of Plant Ecology,43, 501-511. DOI:10.17521/cjpe.2019.0041URL [本文引用: 2]
ZhaoXF,XuHL,ZhangP,TuWX,ZhangQQ(2014).Effects of nutrient and water additions on plant community structure and species diversity in desert grasslands .Chinese Journal of Plant Ecology,38, 167-177. DOI:10.3724/SP.J.1258.2014.00015URL [本文引用: 1]
Investigating the relationship between biodiversity and ecosystem multifunctionality: challenges and solutions 1 2014
... 近年来, 在全球气候变化和人类活动的影响下, ****围绕草地生态系统功能以及物种多样性展开了一系列的研究, 但是多数研究主要集中于某种生态系统功能与驱动因子的关系(Wu et al., 2014).随着研究的深入, 研究者逐渐在不同时间、空间、生境、土壤和气候条件下探讨生态系统多功能性(EMF)的变化(Zavaleta et al., 2010; Lefcheck et al., 2015).如何量化人类活动、气候变化及其他环境因子对生态系统多功能性的影响, 以及这些驱动因子对多个生态系统功能的影响成为研究的热点(Byrnes et al., 2014).特别是在全球气候变暖的背景下, 草地碳库作为陆地生态系统碳库的重要组成部分, 其较小幅度的波动, 将会影响整个陆地生态系统碳循环和EMF.因此, 深入研究草地植被固碳功能和固碳潜力对于适应和减缓气候变化、维持草地EMF具有重要意义. ...
Principles of Terrestrial Ecosystem Ecology 1 2012
... 目前, 相对于单个生态系统功能对EMF和CAR的影响而言, 气候要素的影响更加广泛, 且没有很好地将CAR和EMF联系起来, 关注点主要有两个方面(Jing et al., 2015; Sun et al., 2020).一方面体现在水热条件分别对CAR和EMF的显著影响, 另一方面主要表现在水热条件对生物多样性和EMF两者关系的调节作用上(Soliveres et al., 2014; Jing et al., 2015; Sun et al., 2020).本研究发现, 年降水量对CAR和EMF均有显著正效应, 而年平均气温对二者没有显著效应.实际上, 适宜的水热条件和土壤资源(如土壤水和养分)的变化, 能够解释CAR的差异.具体而言, 水资源不足、相对高温、土壤养分匮乏和资源胁迫的环境下, 会严重阻碍CAR, 进一步能影响EMF (Sun et al., 2020).而且, 水分限制会降低光合作用, 进而降低光照条件下叶片吸收CO2的能力(Chapin III et al., 2012), 因此, 我们的研究结果出现年降水量与CAR和M的显著线性正相关关系(图5A、5C).我们的研究发现, 年平均气温与CAR和M的相关关系较弱, 这和前人的研究(Jing et al., 2015)有很多一致性.当然, 区域性因素是影响本试验结果的重要方面, 例如: 针对青海海北高寒草甸, 有研究表明, 温度变化能显著影响植物群落物种组成和多样性, 从而有可能进一步影响群落生产力、CAR和EMF (Klein et al., 2004); 在青藏高原高寒草原, 降水显著影响高寒草原群落盖度和物种组成(李长斌等, 2016), 从而增加群落CAR和EMF; 此外, 有研究指出, 降水量<450 mm的区域内, 影响青藏高原植被生产力变化的主导因子为降水; 降水量>450 mm的区域, 植被生产力变化的主导因子为气温(陈卓奇等, 2012).而且, 经过长时间序列的观察发现, 在青藏高原地区, 气温对植被的影响具有季节性, AGB和夏季平均气温的相关性较高, 与其他季节的平均气温相关性均不显著(李晓东等, 2012); 对干旱生态系统的研究也发现温度升高不会影响EMF (Maestre et al., 2012).在高寒草地生态系统中, 季节性变化明显, 温差很大, 群落对温度的适应性很强, 因此EMF不会对气温的变化有较大响应(Baumann et al., 2009). ...
基于MODIS的青藏高原植被净初级生产力研究 1 2012
... 目前, 相对于单个生态系统功能对EMF和CAR的影响而言, 气候要素的影响更加广泛, 且没有很好地将CAR和EMF联系起来, 关注点主要有两个方面(Jing et al., 2015; Sun et al., 2020).一方面体现在水热条件分别对CAR和EMF的显著影响, 另一方面主要表现在水热条件对生物多样性和EMF两者关系的调节作用上(Soliveres et al., 2014; Jing et al., 2015; Sun et al., 2020).本研究发现, 年降水量对CAR和EMF均有显著正效应, 而年平均气温对二者没有显著效应.实际上, 适宜的水热条件和土壤资源(如土壤水和养分)的变化, 能够解释CAR的差异.具体而言, 水资源不足、相对高温、土壤养分匮乏和资源胁迫的环境下, 会严重阻碍CAR, 进一步能影响EMF (Sun et al., 2020).而且, 水分限制会降低光合作用, 进而降低光照条件下叶片吸收CO2的能力(Chapin III et al., 2012), 因此, 我们的研究结果出现年降水量与CAR和M的显著线性正相关关系(图5A、5C).我们的研究发现, 年平均气温与CAR和M的相关关系较弱, 这和前人的研究(Jing et al., 2015)有很多一致性.当然, 区域性因素是影响本试验结果的重要方面, 例如: 针对青海海北高寒草甸, 有研究表明, 温度变化能显著影响植物群落物种组成和多样性, 从而有可能进一步影响群落生产力、CAR和EMF (Klein et al., 2004); 在青藏高原高寒草原, 降水显著影响高寒草原群落盖度和物种组成(李长斌等, 2016), 从而增加群落CAR和EMF; 此外, 有研究指出, 降水量<450 mm的区域内, 影响青藏高原植被生产力变化的主导因子为降水; 降水量>450 mm的区域, 植被生产力变化的主导因子为气温(陈卓奇等, 2012).而且, 经过长时间序列的观察发现, 在青藏高原地区, 气温对植被的影响具有季节性, AGB和夏季平均气温的相关性较高, 与其他季节的平均气温相关性均不显著(李晓东等, 2012); 对干旱生态系统的研究也发现温度升高不会影响EMF (Maestre et al., 2012).在高寒草地生态系统中, 季节性变化明显, 温差很大, 群落对温度的适应性很强, 因此EMF不会对气温的变化有较大响应(Baumann et al., 2009). ...
基于MODIS的青藏高原植被净初级生产力研究 1 2012
... 目前, 相对于单个生态系统功能对EMF和CAR的影响而言, 气候要素的影响更加广泛, 且没有很好地将CAR和EMF联系起来, 关注点主要有两个方面(Jing et al., 2015; Sun et al., 2020).一方面体现在水热条件分别对CAR和EMF的显著影响, 另一方面主要表现在水热条件对生物多样性和EMF两者关系的调节作用上(Soliveres et al., 2014; Jing et al., 2015; Sun et al., 2020).本研究发现, 年降水量对CAR和EMF均有显著正效应, 而年平均气温对二者没有显著效应.实际上, 适宜的水热条件和土壤资源(如土壤水和养分)的变化, 能够解释CAR的差异.具体而言, 水资源不足、相对高温、土壤养分匮乏和资源胁迫的环境下, 会严重阻碍CAR, 进一步能影响EMF (Sun et al., 2020).而且, 水分限制会降低光合作用, 进而降低光照条件下叶片吸收CO2的能力(Chapin III et al., 2012), 因此, 我们的研究结果出现年降水量与CAR和M的显著线性正相关关系(图5A、5C).我们的研究发现, 年平均气温与CAR和M的相关关系较弱, 这和前人的研究(Jing et al., 2015)有很多一致性.当然, 区域性因素是影响本试验结果的重要方面, 例如: 针对青海海北高寒草甸, 有研究表明, 温度变化能显著影响植物群落物种组成和多样性, 从而有可能进一步影响群落生产力、CAR和EMF (Klein et al., 2004); 在青藏高原高寒草原, 降水显著影响高寒草原群落盖度和物种组成(李长斌等, 2016), 从而增加群落CAR和EMF; 此外, 有研究指出, 降水量<450 mm的区域内, 影响青藏高原植被生产力变化的主导因子为降水; 降水量>450 mm的区域, 植被生产力变化的主导因子为气温(陈卓奇等, 2012).而且, 经过长时间序列的观察发现, 在青藏高原地区, 气温对植被的影响具有季节性, AGB和夏季平均气温的相关性较高, 与其他季节的平均气温相关性均不显著(李晓东等, 2012); 对干旱生态系统的研究也发现温度升高不会影响EMF (Maestre et al., 2012).在高寒草地生态系统中, 季节性变化明显, 温差很大, 群落对温度的适应性很强, 因此EMF不会对气温的变化有较大响应(Baumann et al., 2009). ...
Recent spring phenology shifts in western Central Europe based on multiscale observations 1 2014
... 全球气候变暖对草地生态系统的植物物候和植物碳(C)积累有重要影响, 而且未来还会继续发生变化, 甚至进一步影响到EMF (Fu et al., 2014).植物中碳的含量会影响和控制生产力、呼吸和分解的生化反应, 对气候变化相当敏感(Yu et al., 2010).目前关于植物碳的研究多关注生长季高峰期植物的生产力或碳含量, 发现水热条件的有效性是影响植物光合作用、呼吸作用、生物化学和物候条件的重要因素(Keeling & Phillips, 2007; Lambers et al., 2008).而且, 植物可以根据气候变化调整光合作用和呼吸作用之间的平衡, 进而调整营养积累(Huxman et al., 2004; Zhang et al., 2013).此外, 由于营养元素控制植物光合作用新细胞的产生, 土壤中的养分(如氮(N)和磷(P))含量是影响植物C含量的另一个决定因素(Heimann & Reichstein, 2008).前人对植物群落影响的研究大多关注碳储存和物候条件以及这些过程的驱动因素(Piao et al., 2008), 对于植物物候和植物碳储量动态之间相互作用的关注有限, 重要的是, 不同环境下植物物候和生存策略的变化可能导致时间尺度上植物碳积累的变化(Xia et al., 2015; Yuan & Chen, 2015).因此, 从生长季开始到生长季生物量峰值的地上部分碳累积速率(CAR)可能对于理解植物适应性策略和EMF更为关键(Sun et al., 2020).之前研究发现, 青藏高原水热状况通过调节土壤营养的动态, 进而调节植被碳积累速率, 例如在高寒草原, 干旱气候和土壤养分的资源匮乏共同制约着植被群落碳积累速率(Sun et al., 2020), 可能进一步调控生态系统功能, 在高寒草甸则反之.另外, 有研究指出, 不同草地利用方式也会影响草地生态系统, 而且在未来降水空间格局变化的背景下, 它们共同决定了生态系统植被类型、净初级生产力和生态系统碳积累, 进而影响水源涵养和碳氮固定、积累等方面的生态功能(杨元合和朴世龙, 2006; 孙鸿烈等, 2012; Sun et al., 2020). ...
Peak season plant activity shift towards spring is reflected by increasing carbon uptake by extratropical ecosystems 1 2018
... 物候数据来自Global Inventory Modelling and Mapping Studies (GIMMS)规范化差异植被指数第3版数据库(Gonsamo et al., 2018).其中, 归一化植被指数(NDVI, 1982-2013年)是从Advanced Very High Resolution Radiometer (AVHRR)传感器获取的基于卫星的地表反射率数据中获得的.通过计算1982-2013年高原植物物候指标确定生长季初始时间(SOS)和生长季高峰时间(POS), 使用线性回归分析得到了2015年的物候指标.在本研究中, 首先采用傅立叶级数模型对NDVI数据集进行平滑(Wang et al., 2018): ...
Terrestrial ecosystem carbon dynamics and climate feedbacks 1 2008
... 全球气候变暖对草地生态系统的植物物候和植物碳(C)积累有重要影响, 而且未来还会继续发生变化, 甚至进一步影响到EMF (Fu et al., 2014).植物中碳的含量会影响和控制生产力、呼吸和分解的生化反应, 对气候变化相当敏感(Yu et al., 2010).目前关于植物碳的研究多关注生长季高峰期植物的生产力或碳含量, 发现水热条件的有效性是影响植物光合作用、呼吸作用、生物化学和物候条件的重要因素(Keeling & Phillips, 2007; Lambers et al., 2008).而且, 植物可以根据气候变化调整光合作用和呼吸作用之间的平衡, 进而调整营养积累(Huxman et al., 2004; Zhang et al., 2013).此外, 由于营养元素控制植物光合作用新细胞的产生, 土壤中的养分(如氮(N)和磷(P))含量是影响植物C含量的另一个决定因素(Heimann & Reichstein, 2008).前人对植物群落影响的研究大多关注碳储存和物候条件以及这些过程的驱动因素(Piao et al., 2008), 对于植物物候和植物碳储量动态之间相互作用的关注有限, 重要的是, 不同环境下植物物候和生存策略的变化可能导致时间尺度上植物碳积累的变化(Xia et al., 2015; Yuan & Chen, 2015).因此, 从生长季开始到生长季生物量峰值的地上部分碳累积速率(CAR)可能对于理解植物适应性策略和EMF更为关键(Sun et al., 2020).之前研究发现, 青藏高原水热状况通过调节土壤营养的动态, 进而调节植被碳积累速率, 例如在高寒草原, 干旱气候和土壤养分的资源匮乏共同制约着植被群落碳积累速率(Sun et al., 2020), 可能进一步调控生态系统功能, 在高寒草甸则反之.另外, 有研究指出, 不同草地利用方式也会影响草地生态系统, 而且在未来降水空间格局变化的背景下, 它们共同决定了生态系统植被类型、净初级生产力和生态系统碳积累, 进而影响水源涵养和碳氮固定、积累等方面的生态功能(杨元合和朴世龙, 2006; 孙鸿烈等, 2012; Sun et al., 2020). ...
Phenology response to climatic dynamic across China’s grasslands from 1985 to 2010 1 2018
... 物候数据来自Global Inventory Modelling and Mapping Studies (GIMMS)规范化差异植被指数第3版数据库(Gonsamo et al., 2018).其中, 归一化植被指数(NDVI, 1982-2013年)是从Advanced Very High Resolution Radiometer (AVHRR)传感器获取的基于卫星的地表反射率数据中获得的.通过计算1982-2013年高原植物物候指标确定生长季初始时间(SOS)和生长季高峰时间(POS), 使用线性回归分析得到了2015年的物候指标.在本研究中, 首先采用傅立叶级数模型对NDVI数据集进行平滑(Wang et al., 2018): ...
Precipitation and species composition primarily determine the diversity-productivity relationship of alpine grasslands on the Northern Tibetan Plateau 1 2014
... 近年来, 在全球气候变化和人类活动的影响下, ****围绕草地生态系统功能以及物种多样性展开了一系列的研究, 但是多数研究主要集中于某种生态系统功能与驱动因子的关系(Wu et al., 2014).随着研究的深入, 研究者逐渐在不同时间、空间、生境、土壤和气候条件下探讨生态系统多功能性(EMF)的变化(Zavaleta et al., 2010; Lefcheck et al., 2015).如何量化人类活动、气候变化及其他环境因子对生态系统多功能性的影响, 以及这些驱动因子对多个生态系统功能的影响成为研究的热点(Byrnes et al., 2014).特别是在全球气候变暖的背景下, 草地碳库作为陆地生态系统碳库的重要组成部分, 其较小幅度的波动, 将会影响整个陆地生态系统碳循环和EMF.因此, 深入研究草地植被固碳功能和固碳潜力对于适应和减缓气候变化、维持草地EMF具有重要意义. ...
Joint control of terrestrial gross primary productivity by plant phenology and physiology 1 2015
... 全球气候变暖对草地生态系统的植物物候和植物碳(C)积累有重要影响, 而且未来还会继续发生变化, 甚至进一步影响到EMF (Fu et al., 2014).植物中碳的含量会影响和控制生产力、呼吸和分解的生化反应, 对气候变化相当敏感(Yu et al., 2010).目前关于植物碳的研究多关注生长季高峰期植物的生产力或碳含量, 发现水热条件的有效性是影响植物光合作用、呼吸作用、生物化学和物候条件的重要因素(Keeling & Phillips, 2007; Lambers et al., 2008).而且, 植物可以根据气候变化调整光合作用和呼吸作用之间的平衡, 进而调整营养积累(Huxman et al., 2004; Zhang et al., 2013).此外, 由于营养元素控制植物光合作用新细胞的产生, 土壤中的养分(如氮(N)和磷(P))含量是影响植物C含量的另一个决定因素(Heimann & Reichstein, 2008).前人对植物群落影响的研究大多关注碳储存和物候条件以及这些过程的驱动因素(Piao et al., 2008), 对于植物物候和植物碳储量动态之间相互作用的关注有限, 重要的是, 不同环境下植物物候和生存策略的变化可能导致时间尺度上植物碳积累的变化(Xia et al., 2015; Yuan & Chen, 2015).因此, 从生长季开始到生长季生物量峰值的地上部分碳累积速率(CAR)可能对于理解植物适应性策略和EMF更为关键(Sun et al., 2020).之前研究发现, 青藏高原水热状况通过调节土壤营养的动态, 进而调节植被碳积累速率, 例如在高寒草原, 干旱气候和土壤养分的资源匮乏共同制约着植被群落碳积累速率(Sun et al., 2020), 可能进一步调控生态系统功能, 在高寒草甸则反之.另外, 有研究指出, 不同草地利用方式也会影响草地生态系统, 而且在未来降水空间格局变化的背景下, 它们共同决定了生态系统植被类型、净初级生产力和生态系统碳积累, 进而影响水源涵养和碳氮固定、积累等方面的生态功能(杨元合和朴世龙, 2006; 孙鸿烈等, 2012; Sun et al., 2020). ...
 ,
, ,1,*, 王毅1,2, 刘国华3
,1,*, 王毅1,2, 刘国华3

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT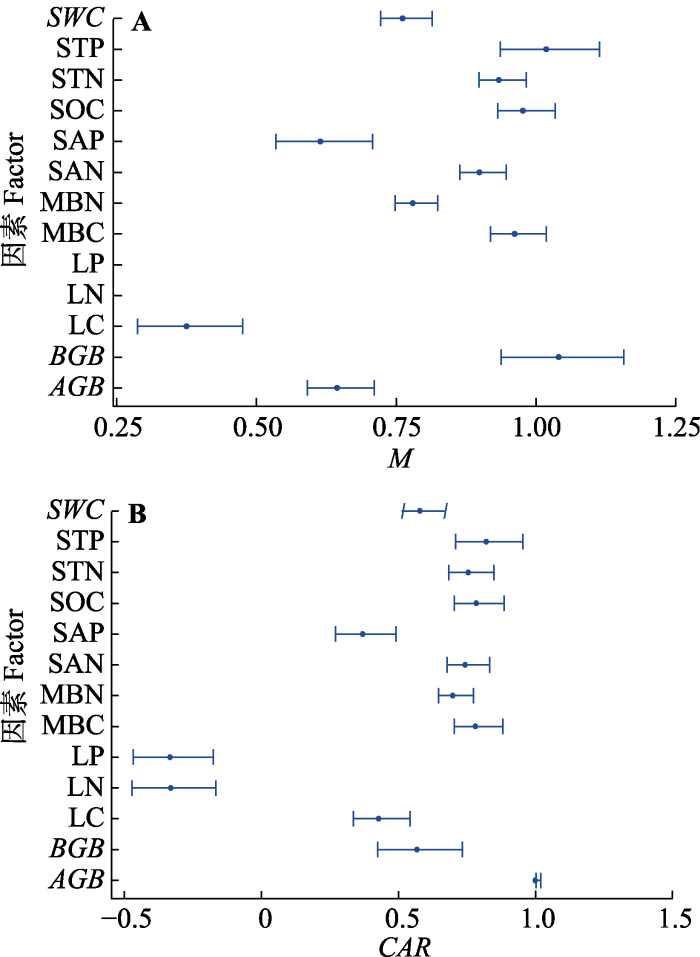 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT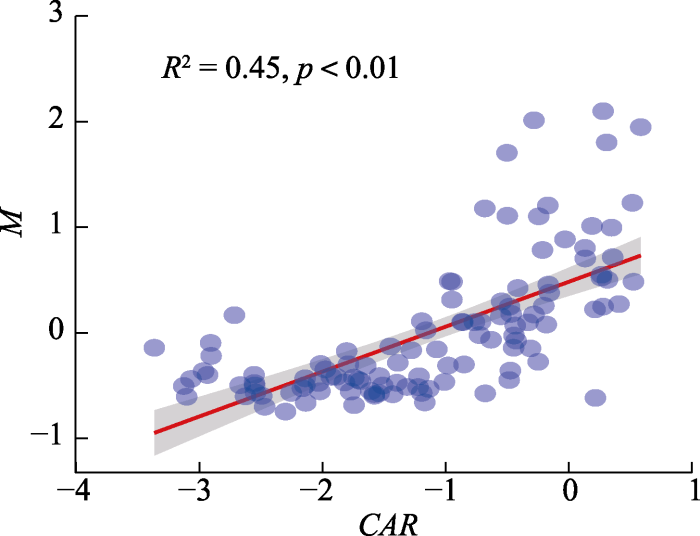 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT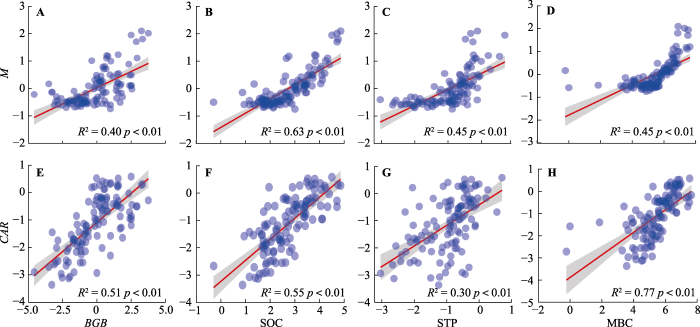 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT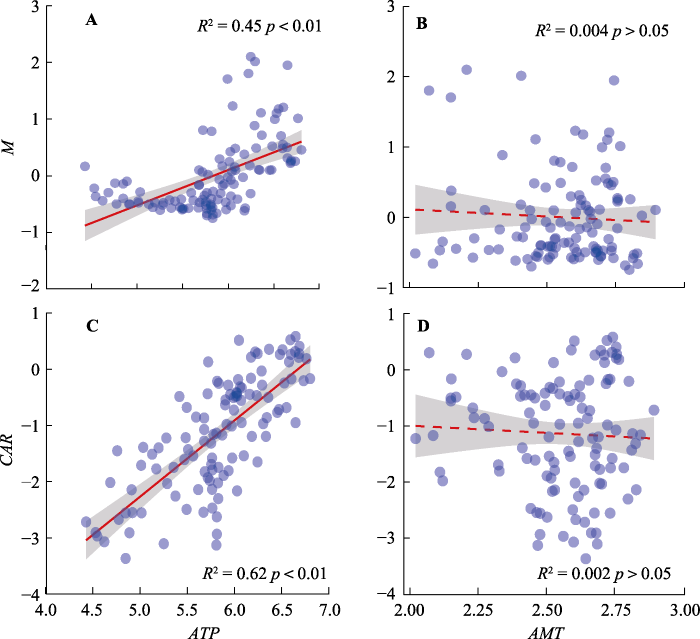 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}