 ,1,2,3,*
,1,2,3,*Relationships among xylem transport, anatomical structure and mechanical strength in stems and roots of three Podocarpaceae species
Shi-Tong LU1,2, Sen CHEN1,2, Yan LI1,2,3, Zhong-Yuan WANG1, Tian-Tian PAN1,2, Lin-Feng YE1,2, Jiang-Bo XIE,1,2,3,*通讯作者: *(0208xiejiangbo@163.com)
编委: 谭敦炎
责任编辑: 李敏
收稿日期:2020-12-4接受日期:2021-03-17
| 基金资助: |
Received:2020-12-4Accepted:2021-03-17
| Fund supported: |

摘要
水力失效是植物干旱死亡的主要机制。量化分析水力性状的种间和器官间差异是预测树木在气候变化下的响应甚至生存能力的基础。该研究对比分析了罗汉松科3种植物器官(茎和根)水平上水力功能性状的差异, 并探讨其与解剖结构和机械强度之间的关系。在湿生同质园内选择罗汉松科3种植物, 测定了茎和根木质部水力功能性状(最大比导率(Ks)和栓塞抗性(P50))、解剖结构性状(管胞直径(Dt)、水力直径(Dh)、管胞密度(Nt)、管胞壁厚(Tw)、纹孔膜直径(Dp)和纹孔密度(Np))和机械强度(木材密度(WD)和管胞厚度跨度比((t/b)2))。结果发现: (1)罗汉松科3种植物茎木质部不存在效率-安全权衡, 而根木质部存在权衡。(2)茎Ks与Dp显著正相关, 与(t/b)2和WD无关; 茎P50与Dp极显著负相关, 与(t/b)2和WD无关。(3)根Ks与Dh显著正相关, 与Tw和(t/b)2极显著负相关; 根P50与Tw、(t/b)2和WD均极显著正相关。在罗汉松科植物中, 根木质部性状与输水效率和栓塞抗性的密切关系是解释其存在效率-安全权衡的基础, 而茎木质部的过度建造是茎不存在效率-安全权衡的原因, 木质部的过度建造仍需要更多的实验证据。
关键词:
Abstract
Aims Hydraulic failure is one of the primary cause of plant mortality during drought. Thus, quantitative analysis on inter-specific and inter-organ variance in hydraulic traits can help us to predict the response and even survivability of species under climate change. Methods Here, three Podocarpaceae species (Podocarpus macrophyllus, P. macrophyllusvar. maki and Nageia nagi) grown in a mesic common garden were studied, with xylem hydraulic function (specific hydraulic conductivity (Ks); embolism resistance (P50)), anatomical structure (tracheid diameter (Dt); hydraulic diameter (Dh); tracheid wall thickness (Tw); tracheid density (Nt); pit membrane diameter (Dp); pit density (Np)) and mechanical strength (wood density (WD); tracheid thickness to span ratio ((t/b)2)) measured. Then, we analyzed hydraulic traits variance at the organ level (stem and root) in three Podocarpaceae species, and investigated the relationships among xylem hydraulic traits, anatomical structure and mechanical strength in stems and roots. Important findings We found that: 1) The stem xylem in three Podocarpaceae species exhibited no safety- efficiency trade-off. In contrast, the root xylem exhibited safety-efficiency trade-off. 2) For stems, Ks was positively correlated with Dp, but decoupled with stem WD and (t/b)2; Stem P50 was negatively correlated with Dp, but not correlated with WD and (t/b)2. 3) For roots, Ks was positively correlated with hydraulic diameter, but negatively correlated with root Tw and (t/b)2; Root P50 was positively correlated with Tw, (t/b)2 and WD. Root xylem traits exhibited strong relationships with both Ks and P50, demonstrating its cause-and-effect basis for the safety-efficiency tradeoff. On the other hand, the absence of safety-efficiency tradeoff in stems may be attributed to the overbuilt xylem of Podocarpaceae. More experimental evidence on the overbuilding of xylem is desired in the future study.
Keywords:
PDF (3051KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
陆世通, 陈森, 李彦, 王忠媛, 潘天天, 叶琳峰, 谢江波. 罗汉松科3种植物茎和根木质部水分运输、解剖结构与机械强度之间的关系. 植物生态学报, 2021, 45(6): 659-669. DOI: 10.17521/cjpe.2020.0402
LU Shi-Tong, CHEN Sen, LI Yan, WANG Zhong-Yuan, PAN Tian-Tian, YE Lin-Feng, XIE Jiang-Bo.

目前气候模型预测全球范围内干旱频率和强度都将增加(IPCC, 2013; Dietrich et al., 2019)。与干旱和高温有关的森林死亡事件在全球均有案例, 从热带雨林到北方寒带森林, 森林生长和死亡都对干旱高度敏感(Allen, 2009; Choat et al., 2012, 2018)。树木干旱死亡的机制主要有水力失效和碳饥饿(McDowell et al., 2008)。最近越来越多的研究支持水力失效是导致树木干旱死亡的原因(Rowland et al., 2015; Anderegg et al., 2016; Adams et al., 2017)。因此, 预测干旱对不同物种的影响就需要了解植物水力功能性状的种间差异(McElrone et al., 2004; Rosas et al., 2019)。
木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(Brodribb & Field, 2000; Zhu & Cao, 2009; Schumann et al., 2019)。P50代表了木质部抵抗栓塞的能力, 是确定木本物种耐旱极限的一个至关重要的性状(Choat et al., 2012)。此外, 栓塞抗性在不同分类群中差异很大, 即使是生长在相同条件下的近缘物种也存在显著差异, 栓塞抗性的变异是木质部输水效率-安全权衡存在的原因之一(van der Willigen et al., 2000; McCulloh et al., 2019)。
目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(Hacke et al., 2006, 2017)。管道直径决定了木质部输水效率, 影响了冻融胁迫引起的栓塞脆弱性, 并在较小程度上影响了干旱胁迫引起的栓塞脆弱性(Pittermann & Sperry, 2003, 2006; Choat et al., 2011; Hacke et al., 2017)。然而, 目前一些研究无法证实大直径管道对干旱引起的栓塞具有更高的脆弱性(Lens et al., 2011; Schuldt et al., 2016; Schumann et al., 2019)。因此, 木质部输水效率-安全权衡存在与否仍有争议。有研究表明: 对于水分胁迫引起的栓塞, 效率-安全权衡可以用纹孔来解释(Tyree & Zimmermann, 2002; Sperry et al., 2006; Schoonmaker et al., 2010)。在Hacke和Jansen (2009)的研究中, 裸子植物栓塞抗性与纹孔塞和纹孔口的相对大小有关。然而, 也有研究表明北方针叶树的纹孔输水效率与栓塞抗性之间只存在微弱的联系, 而南半球的针叶树并没有相关性(Hacke et al., 2004; Pittermann et al., 2010)。
水力功能性状与木质部的机械强度是相互依赖的。在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(Hacke et al., 2001; Pittermann et al., 2006a)。目前的研究表明了木材密度和管道厚度跨度比(管道厚度t与管腔跨度b的比值)与栓塞抗性有关(Hacke et al., 2001; Pittermann et al., 2006a; Lachenbruch & McCulloh, 2014)。因此, 机械强度也需要支持维管网络, 阻止因木质部负压引起的管道内爆(Pittermann et al., 2006a; Fan et al., 2017)。管道长度、直径和壁厚以及木材密度都会影响木质部输水效率和栓塞抗性以及管道强度(Sperry et al., 2006)。Baas等(2004)提出木质部“权衡”三角形, 权衡可能存在于木质部输水效率、安全性和机械强度之间。权衡可能会因木质部的过度建造而消失, 例如罗汉松科植物木质部相对于栓塞抗性有着更大的密度、厚度跨度之比, 且与栓塞抗性解耦联(Pittermann et al., 2006a; Brodribb, 2011)。
目前的研究集中于木本植物茎木质部性状的研究, 由于根深埋于土壤中, 比地上植物器官更难接近, 并且根的建筑复杂性为样品采样和标准水力测量方案带来了不确定性(Peters et al., 2020)。关于根木质部解剖和机械性状如何影响水分输送的研究仍较为欠缺(Pratt et al., 2007a; Rodriguez-Dominguez et al., 2018; Peters et al., 2020)。同时根是植物水力学途径的重要组成成分, 负责向所有地表植物组织输送水分(Rodriguez-Dominguez & Brodribb, 2020), 特别是在干旱时期, 针叶树维持生存依赖于已建立的根(Mackay et al., 2020)。
本研究对比了罗汉松科3种植物茎和根在水力功能和木质部解剖结构性状的差异, 并试图阐明3种植物茎和根水分运输与解剖结构和机械强度之间的关系。木质部输水效率量化为最大比导率, 木质部安全性量化为栓塞抗性(P50), 同时测量了相应的木质部解剖性状包括管胞直径、水力直径、管胞壁厚、管胞密度、纹孔膜直径、纹孔密度。用木材密度和管胞厚度跨度比来量化茎和根机械强度。根据木质部水分运输与解剖结构和机械强度的关系, 我们提出了两个问题: (1)在罗汉松科植物木质部是否存在效率-安全权衡, 这种权衡在茎和根之间是否有区别; (2)解剖结构和机械强度等性状如何影响木质部的输水效率和安全性之间的关系。
1 材料和方法
1.1 研究地概况
本研究在浙江省杭州市临安区湿生同质园内进行, 同质园(30.26° N, 119.73° E)位于浙江省西北部的天目山地区。天目山地区地处南方丘陵地带, 亚热带季风气候, 年降水量1 390-1 870 mm, 年平均气温14 ℃, 最冷月平均气温-2.6-3.4 ℃, 极端最低气温在-20.2- -13.1 ℃, 最热月平均气温19.9- 28.1 ℃, 极端最高气温29.1-38.2 ℃, 无霜期209- 235天。年日照时间为1 550-2 000 h, 年太阳辐射3 270-4 460 MJ·m-2。1.2 研究材料
本研究的目标树种为罗汉松科植物罗汉松(Podocarpus macrophyllus)、短叶罗汉松(P. macrophyllus var. maki)和竹柏(Nageia nagi)。在2018年11月间采样, 采样时间为8:00-11:00。所选植物立地条件基本一致, 生长状况良好(表1)。每株树至少采集3个树冠朝南方向1-2年生的枝条, 长度约1.5 m。在取枝条的正下方土壤中挖掘取根, 一般选择位于土层下10-20 cm深, 根直径在4-6 mm, 长度在40-60 cm的输水根, 小心地将根从土壤中取出并防止根木质部损伤。枝条和根在取样后迅速将其放入装有蒸馏水的黑色收纳箱中, 并用黑色塑料袋罩住并立即带回实验室。Table 1
表1
表1罗汉松科3种植物采样点及样树的基本特征(平均值±标准误)
Table 1
| 树种 Species | 海拔 Altitude (m) | 坡向 Slope direction | 坡度 Slope (°) | 土壤类型 Soil type | 土壤含水量 Soil water content (%) | 土壤容重 Soil bulk density (g·cm-3) | 树高 Tree height (m) | 胸径 DBH (cm) |
|---|---|---|---|---|---|---|---|---|
| 罗汉松 Podocarpus macrophyllus | 42.47 ± 0.23 | 西南 SW | 3 | 黄红壤亚类 Yellow red soil subclass | 36.87 ± 1.81 | 1.22 ± 0.15 | 5.93 ± 0.42 | 13.50 ± 1.55 |
| 短叶罗汉松 P. macrophyllus var. maki | 39.40 ± 0.44 | 西南 SW | 8 | 黄红壤亚类 Yellow red soil subclass | 35.24 ± 1.75 | 1.20 ± 0.10 | 2.89 ± 0.32 | 11.99 ± 1.54 |
| 竹柏 Nageia nagi | 41.48 ± 0.20 | 西南 SW | 1 | 黄红壤亚类 Yellow red soil subclass | 36.34 ± 1.74 | 1.23 ± 0.08 | 3.66 ± 0.18 | 8.52 ± 0.64 |
新窗口打开|下载CSV
1.3 研究方法
1.3.1 茎/根木质部输水效率和栓塞抗性所有茎段或根段均在水下剪取一段。茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28)。所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭。在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径。茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25)。然后小心地将茎段或根段基部上部2-3 cm的表皮剥离。该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率。测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(周洪华等, 2012; 刘存海等, 2014)。在高压(120 kPa)下反复冲刷10 min, 以确保潜在栓塞被清除, 直到达到最大导水率, 该过程反复进行直到最大导水率不再变化。在低压(6 kPa)下测量茎段或根段的最大导水率(Khmax, kg·m·MPa-1·s-1) (Pratt et al., 2007b; Schoonmaker et al., 2010; Johnson et al., 2016; Schumann et al., 2019)。采用Xylwin 3.2软件对其导水率和流量数据进行分析。根据所测的最大导水率和茎段或根段的长度, 最大比导率(Ks, kg·m-1·MPa-1·s-1)通过Khmax除以无髓和树皮的基部边材横截面积得到(Hajek et al., 2016; Schuldt et al., 2016)。
使用空气注入法(双端压力套技术)构建茎段或根段的脆弱性曲线。过去的研究表明了使用此法可以获得可靠的水力脆弱性曲线(Ennajeh et al., 2011)。在确定Khmax后, 将茎段或根段放入长度约为8 cm的双端压力套中, 与所加压力平衡。然后将茎段或根段从压力套上取下, 用测量Khmax的方法测量加压后的导水率(Kh, kg·m·MPa-1·s-1)。这个过程以0.2或0.3 MPa的增量重复进行(取决于植物或器官), 直到Kh降至最大值的10%以下(Johnson et al., 2016; Schumann et al., 2019)。导水率损失百分比计算如下: PLC = 100 × (1 - Kh/Khmax)。在R语言中使用“fitplc”软件包拟合PLC与水势之间的脆弱性曲线(Ogle et al., 2009; Duursma & Choat, 2017), 通过getPx函数从曲线中提取对应的P50。
1.3.2 木质部解剖结构
从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征。样品固定软化之后, 采用石蜡切片法将其制成永久切片(张军周等, 2013)。所有样品的横切面使用Leica DM3000显微镜(Leica Microsystems CMS GmbH, Wetzlar, Germany)在50倍镜和400倍镜下摄像。所有样品的纵切面在400倍镜放大下进行摄像。使用Image J图像处理软件分析照片。测量参数包括管胞直径(Dt, μm), 管胞密度(Nt, mm-2), 管胞壁厚(Tw, μm), 纹孔膜直径(Dp, μm), 纹孔密度(Np, mm-2)。由于管胞近似于椭圆, 根据Lewis等(1992)给出的公式D = [32 × (a × b)3/(a2 + b2)]1/4, 从椭圆的主轴(a)和次轴(b)计算出Dt。根据Tyree和Zimmermann (2002)给出的公式, 水力直径(Dh) = ∑D5/∑D4, 通过导管直径计算出Dh。对于管胞厚度跨度比((t/b)2), 随机选取4张400倍镜放大的横切面照片, 测量所有的管胞, 直到测量到50个管胞。其中t为相邻管胞的共同壁厚, 通过直接测量得到, b为管胞腔直径, 通过椭圆的主轴和次轴计算得到(Hacke et al., 2001; Pratt et al., 2007a)。选取400倍镜放大的茎和根纵切面图各4张, 测量纹孔面积, 将面积转化为Dp, 并估算每个管胞上的Np (Pittermann et al., 2006b)。
1.3.3 木材密度
根据Hacke等(2000)的方法测量茎和根的木材密度(WD), 对用于测定脆弱性曲线的茎和根木材密度进行测量。将木材样品树皮用剃刀除去后浸没在蒸馏水中。采用阿基米德原理确定木材样品的新鲜体积。然后将样品放置在75 ℃的烘箱烘48 h, 测量其干质量。木材密度即为干质量与新鲜体积之比。
1.3.4 统计分析
罗汉松科植物茎和根水力性状单因素方差分析(ANOVA)、木质部性状相关分析、图表制作等均在R 3.6.3中完成。
图1
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1三种罗汉松科植物木材解剖横切面和纵切面光学显微镜图像。A-D, 罗汉松茎横切图和纵切图、根横切图和纵切图。E-H, 短叶罗汉松茎横切图和纵切图、根横切图和纵切图。I-L, 竹柏茎横切图和纵切图、根横切图和纵切图。
Fig. 1Examples of light microscopy images of transverse and vertical sections of wood anatomy of three Podocarpaceae species. A-D, Transverse sections and vertical sections of stem, transverse sections and vertical sections of root in Podocarpus macrophyllus. E-H, Transverse sections and vertical sections of stem, transverse sections and vertical sections of root in P. macrophyllus var. maki. I-L, Transverse sections and vertical sections of stem, transverse sections and vertical sections of root in Nageia nagi.
2 结果和分析
2.1 输水效率与安全
罗汉松科3种植物茎Ks没有显著差异(罗汉松, (0.38 ± 0.02) kg·m-1·MPa-1·s-1; 短叶罗汉松, (0.30 ± 0.04) kg·m-1·MPa-1·s-1; 竹柏, (0.29 ± 0.01) kg·m-1·MPa-1·s-1)。茎P50在3种植物之间存在显著差异, 罗汉松((-4.75 ± 0.09) MPa)和短叶罗汉松茎P50 ((-5.06 ± 0.25) MPa)显著低于竹柏((-3.85 ± 0.26) MPa) (图2)。根Ks越大的物种, 其根P50越高(图2)。罗汉松根Ks ((3.56 ± 0.25) kg·m-1·MPa-1·s-1)最大, 其根P50 ((-0.72 ± 0.05) MPa)也显著高于短叶罗汉松((-1.32 ± 0.14) MPa)和竹柏((-2.60 ± 0.09) MPa) (图2)。图2
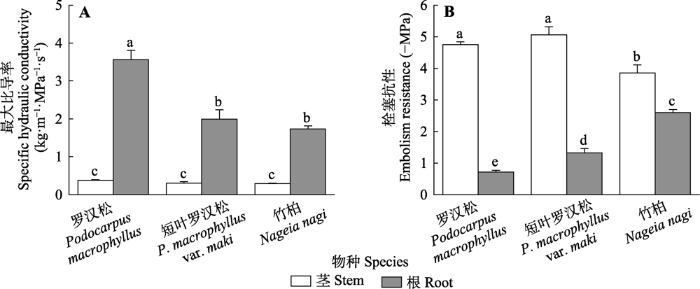 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2罗汉松科3种植物的水力功能性状图(平均值±标准误, n = 7)。不同小写字母表示在p ≤ 0.05水平上差异显著。
Fig. 2Hydraulic functional traits of three Podocarpaceae species (mean ± SE, n = 7). Different lowercase letters indicate significant differences at p ≤ 0.05 level.
3种植物茎Ks显著小于根Ks, 茎P50显著低于根P50 (图2)。
2.2 木质部解剖结构
竹柏茎Dt和Tw显著大于罗汉松(图3A、3D)。罗汉松根Dt最大, 根Tw最小(图3A、3D)。对比茎和根管胞结构, 根Dt和Tw显著大于茎(图3A、3D)。相反, 茎Nt显著大于根(图3C)。图3
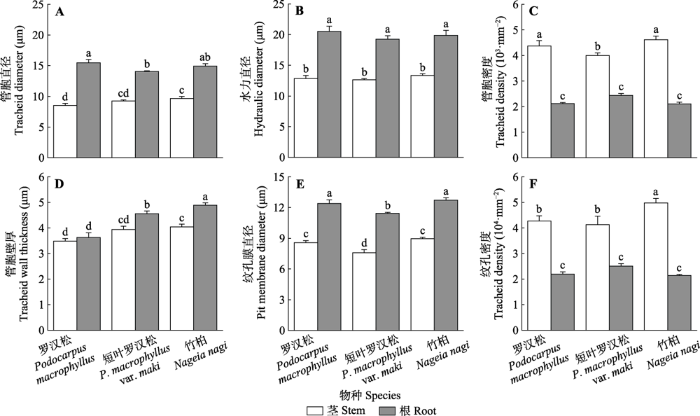 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3罗汉松科3种植物木质部解剖结构图(平均值±标准误, n = 7)。不同小写字母表示在p ≤ 0.05水平上差异显著。
Fig. 3Xylem anatomical structure of three Podocarpaceae species (mean ± SE, n = 7). Different lowercase letters indicate significant differences at p ≤ 0.05 level.
短叶罗汉松茎Dp显著小于罗汉松和竹柏, 竹柏茎Np显著大于罗汉松和短叶罗汉松(图3E、3F)。罗汉松和竹柏根Dp显著大于短叶罗汉松(图3E)。对比茎和根纹孔结构, 根Dp更大、Np更小(图3E、3F)。
2.3 木质部机械功能
3种植物的茎WD和(t/b)2没有显著差异(图4B)。竹柏根WD显著大于罗汉松和短叶罗汉松, 短叶罗汉松和竹柏根(t/b)2显著大于罗汉松(图4)。罗汉松和短叶罗汉松茎WD显著大于根(图4A)。3种植物茎(t/b)2均显著大于根(图4B)。图4
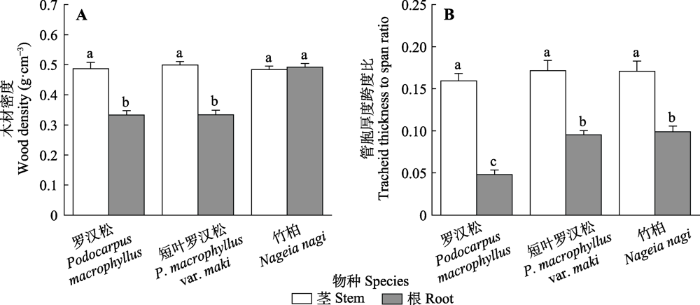 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4罗汉松科3种植物木质部机械强度图(平均值±标准误, n = 7)。不同小写字母表示在p ≤ 0.05水平上差异显著。
Fig. 4Xylem mechanical traits of three Podocarpaceae species (mean ± SE, n = 7). Different lowercase letters indicate significant differences at p ≤ 0.05 level.
2.4 木质部输水效率和安全性的关系
罗汉松科3种植物茎Ks和P50无关(p > 0.05, 图5A), 而根木质部存在效率-安全权衡(p < 0.05, 图5B)。图5
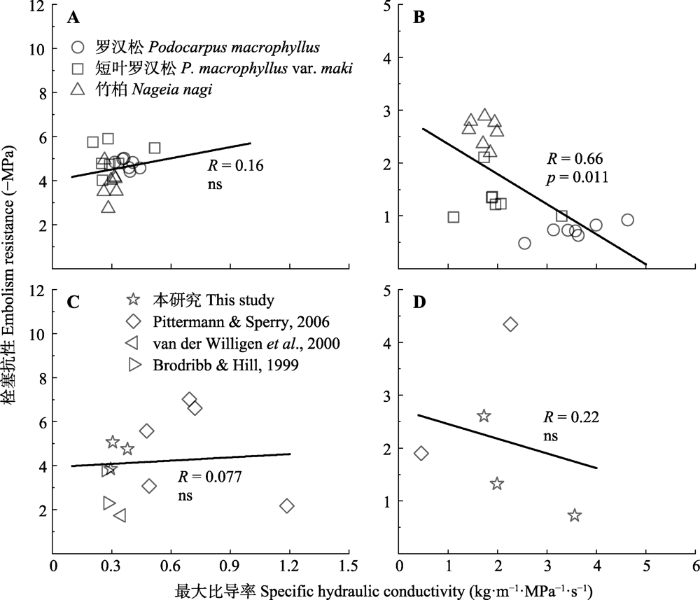 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5罗汉松科植物木质部输水效率与栓塞抗性的关系。A, 本研究罗汉松科3种植物茎。B, 本研究罗汉松科3种植物根。C, 罗汉松科12种植物茎(数据来源于Brodribb和Hill (1999), van der Willigen等(2000), Pittermann和Sperry (2006))。D, 罗汉松科5种植物根(数据来源于Pittermann和Sperry (2006))。ns, 没有显著关系。
Fig. 5Relationship between hydraulic conductivity and embolism resistance in the three Podocarpaceae species. A, The stems of three Podocarpaceae species in this paper. B, The roots of three Podocarpaceae species in this paper. C, The stems of twelve Podocarpaceae species (Data from Brodribb & Hill (1999), van der Willigen et al. (2000), Pittermann & Sperry (2006)). D, The roots of five Podocarpaceae species (Data from Pittermann & Sperry (2006)). ns, non-significant relationships.
基于先前发表的研究, 收集了12种罗汉松科植物的茎和5种罗汉松科植物根Ks和P50的数据。综合数据分析表明, 12种罗汉松科植物茎Ks和P50无关(p > 0.05, 图5C); 5种罗汉松科植物根Ks和P50无关(p > 0.05, 图5D)。
2.5 木质部性状之间的相关性
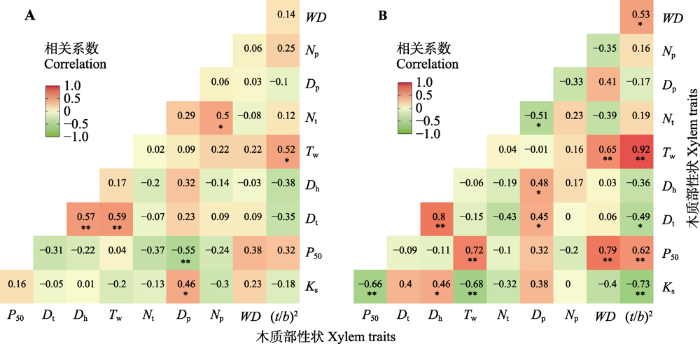
茎Ks与Dp显著正相关(p < 0.05), 与Dt和Tw无相关关系(p > 0.05, 图6A)。此外, Ks与茎的机械强度(WD和(t/b)2)相关性不显著(p > 0.05, 图6A)。茎P50与Dp显著负相关(p < 0.05), 与根Tw、(t/b)2和WD无相关关系(p > 0.05, 图6A)。图6
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6罗汉松科植物10个木质部性状之间的相关性分析。A, 茎木质部性状。B, 根木质部性状。*, p ≤ 0.05; **, p ≤ 0.01。Dh, 水力直径(μm); Dp, 纹孔膜直径(μm); Dt, 管胞直径(μm); Ks, 最大比导率(kg·m-1·MPa-1·s-1); Np, 纹孔密度(mm-2); Nt, 管胞密度(mm-2); P50, 导水率损失50%时的水势(-MPa); Tw, 管胞厚度(μm); WD, 木质部密度(g·cm-2); (t/b)2, 管胞厚度跨度比。
Fig. 6Correlation analysis of ten xylem traits in Podocarpaceae species. A, Xylem traits of stem. B, Xylem traits of root. *, p ≤ 0.05; **, p ≤ 0.01. Dh, hydraulic diameter (μm); Dp, pit membrane diameter (μm); Dt, tracheid diameter (μm); Ks, specific hydraulic conductivity (kg·m-1·MPa-1·s-1); Np, pit density (mm-2); Nt, tracheid density (mm2); P50, the xylem water potential causing 50% loss of hydraulic conductivity (-MPa); Tw, tracheid wall thickness (μm); WD, wood density (g·cm-2); (t/b)2, tracheid thickness to span ratio.
根Ks与Dh显著正相关(p < 0.05), 与Dp无相关关系(p > 0.05, 图6B)。根Ks与(t/b)2和Tw极显著负相关(p < 0.01, 图6B)。根P50与Dt和Dp均无相关关系(p > 0.05), 与根的机械强度(WD和(t/b)2)极显著正相关(p < 0.01), 与根Tw极显著正相关(p < 0.01, 图6B)。并且, 根Tw与(t/b)2和WD相关性极显著(p < 0.01), 即管胞壁越厚, 管胞(t/b)2和WD越大, 栓塞抗性也越强(p < 0.01, 图6B)。
3 讨论
3.1 茎和根的水力功能性状
罗汉松科3种植物茎中, 短叶罗汉松是茎栓塞抗性最强的物种, 与Brodribb和Hill (1999)所报道的P. lawrencei近似(P50, -5.6 MPa); 而竹柏茎栓塞抗性最弱(图2B)。根栓塞抗性有明显差异, 竹柏根栓塞抗性最强, 罗汉松最弱(图2B)。竹柏的管胞壁厚和木材密度最大, 其栓塞抗性也最高(图3D, 图4A), 与之前的研究结果相同, 即栓塞抗性的增加与木材密度和管胞壁厚的增加相关(Hacke et al., 2001; Fichot et al., 2010; Rosas et al., 2019)。罗汉松科3种植物根管胞和纹孔膜直径均比茎大(图3A、3E), 导致根的输水效率比茎更高, 但根高效的木质部更易引起栓塞发生和扩散(Loepfe et al., 2007; Schulte et al., 2015)。因此, 在水力功能方面表现为根栓塞抗性比茎弱(图2B), 导水率比茎高(图2A; Maherali et al., 2006; Pratt et al., 2007a; Hacke & Jansen, 2009; Johnson et al., 2016)。根更低的机械强度也意味着其管道强度更弱(图4)。上述结果代表了根可能是土壤到叶片水分运输通道的脆弱环节(McElrone et al., 2004; Maherali et al., 2006)。根比茎更脆弱可以允许植物在树干或枝条未发生空化的情况下先关闭气孔, 从而延迟不可逆的木质部损害(Martin-StPaul et al., 2017; Choat et al., 2018)。
3.2 茎和根木质部输水效率-安全权衡
本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡。对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(Brodribb & Hill, 1999; van der Willigen et al., 2000; Pittermann & Sperry, 2006; Gleason et al., 2016)。而本研究罗汉松科3种植物根木质部存在权衡。McElrone等(2004)对Juniperus ashei的研究也表明深根和浅根输水效率和安全性之间存在权衡。从效率和安全性的权衡来看, 根比茎更可能实现权衡(Hacke & Jansen, 2009)。首先, 茎木质部的结构异质性会改变输水效率和安全性之间的关系(Mayr & Cochard, 2003)。其次, 对于多数针叶树而言, 根的管胞较宽较长, 纹孔较大(Hacke & Jansen, 2009; Schulte et al., 2015), 上述结构提高了根的输水效率, 降低了栓塞抗性。本研究罗汉松科3种植物根管胞更宽、纹孔更大、木材密度和管胞厚跨比更小(图3, 图4)。此外, 其他木质部特有的权衡可能会混淆效率-安全权衡关系(Gleason et al., 2016)。3.3 木质部输水效率和安全性的决定因素
本研究表明茎管胞直径与输水效率和栓塞抗性没有联系, 而纹孔膜直径会影响茎输水效率和栓塞抗性(图6A), 茎纹孔膜直径越大, 输水效率就越大, 栓塞抗性就越弱。Hacke和Jansen (2009)研究表明在北方松科3种植物中, 茎纹孔膜直径与输水效率显著正相关, 本研究结果与之相似。考虑到复杂的纹孔结构包括纹孔膜孔隙分布、纹孔深度和纹孔塞-塞缘结构等, 都会影响输水效率和栓塞抗性(Domec et al., 2006; Pittermann et al., 2010; Schulte et al., 2015)。需要更多的工作来充分理解针叶树种纹孔结构和功能的差异及其与木质部输水效率和安全性的关系。罗汉松科植物茎机械性状(WD, (t/b)2)与茎输水效率和栓塞抗性无关, 无论是细胞水平((t/b)2), 还是组织水平(WD)(图6)。本研究结果与Pittermann等(2006a)对罗汉松科植物的研究结果相似, 其栓塞抗性与机械强度参数无关。Pittermann等(2006a)将这种现象归因于罗汉松科植物过度建造的木材: 由于自身栓塞保护作用, 木材抗旱成本高且输水效率低下, 在细胞水平上体现为厚度跨度之比(图4B), 组织水平上体现为更大的木材密度(图4A)。木材的过度建造在竹柏上尤为明显, 因为它具有远低于罗汉松和短叶罗汉松的栓塞抗性, 却拥有与罗汉松和短叶罗汉松相当的木材密度和管胞厚度跨度比, 导致罗汉松科植物茎机械强度与茎栓塞抗性解耦联(图4)。
根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(图6B), 与以往的研究结果(Sperry et al., 2006; Schuldt et al., 2016)一致, 即管胞直径在提高输水效率的同时并不会显著影响安全性(Tyree et al., 1994)。根栓塞抗性与管胞厚度跨度比显著正相关, 与Sperry等(2006)的研究结论相似。而根管胞厚度跨度比的增加显著降低了根输水效率(图6B), 意味着根输水效率和栓塞抗性与管胞厚度跨度比之间存在显著的权衡。根栓塞抗性也与组织水平上的机械强度(WD)密切正相关(图6B)。另外, 在根管胞壁厚与管胞厚度跨度比和木材密度之间存在密切的联系(图6B), 表明根管胞壁加厚是根机械强度增加的主因。与Fichot等(2010)的研究结果相似, 本研究也发现根管胞壁加厚与栓塞抗性的增强密切相关, 根管胞壁加厚可以加固根脆弱的管道, 增强根的机械强度, 从而增强了根栓塞抗性。因此, 对管胞壁较高的碳投入以弥补较大管道脆弱的栓塞抗性或许是植物应对干旱胁迫的策略(Hacke et al., 2001; Larter et al., 2017)。此外, 管胞壁加厚会影响纹孔膜性能包括纹孔膜孔隙和厚度, 而纹孔膜厚度的增加与木质部栓塞抗性有关(Jansen et al., 2009; Li et al., 2016)。
本研究的罗汉松科3种植物根木质部存在效率-安全权衡, 但茎木质部不存在(图5, 图6)。根木质部存在权衡的原因可能与管胞壁厚和根管胞厚度跨度比有关, 因为这两个性状都显示出与根输水效率和栓塞抗性的密切关系。Lens等(2011)对7种槭属(Acer)植物的研究表明木质部的关键性状与输水效率和栓塞抗性的密切关系是解释安全-效率权衡的基础。对木质部性状的相关性分析表明了根输水效率和栓塞抗性对结构需求是对立的, 例如根管胞厚度跨度比、管胞壁厚和木材密度是与更强栓塞抗性和更低输水效率相关的性状, 管胞直径和水力直径是与更高输水效率和更弱栓塞抗性相关的性状。本研究与Lens等(2011)的结论是相容的。茎输水效率和栓塞抗性与纹孔膜直径之间存在权衡, 但是因栓塞保护作用而过度建造的高密度木材可能会混淆效率-安全权衡(图6A)。因此, 效率-安全权衡依赖于木质部维管束网络性状之间的联系而不能归因于单一因素(Loepfe et al., 2007)。
4 结论
本研究证明在罗汉松科3种植物中: (1)根木质部存在效率-安全权衡。这种权衡与根管胞壁厚、管胞厚度跨度比和木材密度(根机械强度)密切相关, 并且根管胞壁加厚是根机械强度增强的主因。其次, 与茎相比, 根有更大的管胞和纹孔, 更小的管胞厚度跨度比和木材密度, 上述结构提高了根输水效率, 同时降低了栓塞抗性。(2)茎木质部不存在效率-安全权衡, 茎木材密度和管胞厚度跨度比均与其栓塞抗性无关, 暗示着罗汉松科茎木质部可能是过度建造的。上述结果表明效率和安全对多种结构需求的对立关系是权衡存在的结构基础, 并且在茎和根性状网络分析中得到了进一步证实。网络分析的结果强调了植物输水效率和栓塞抗性是由多种木质部性状共同决定的, 而不能归因于单个解剖结构。致谢
感谢荒漠与绿洲生态国家重点实验室开放基金资助。参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
[本文引用: 1]
[本文引用: 1]
DOI:10.1073/pnas.1525678113PMID:27091965 [本文引用: 1]

Drought-induced tree mortality has been observed globally and is expected to increase under climate change scenarios, with large potential consequences for the terrestrial carbon sink. Predicting mortality across species is crucial for assessing the effects of climate extremes on forest community biodiversity, composition, and carbon sequestration. However, the physiological traits associated with elevated risk of mortality in diverse ecosystems remain unknown, although these traits could greatly improve understanding and prediction of tree mortality in forests. We performed a meta-analysis on species' mortality rates across 475 species from 33 studies around the globe to assess which traits determine a species' mortality risk. We found that species-specific mortality anomalies from community mortality rate in a given drought were associated with plant hydraulic traits. Across all species, mortality was best predicted by a low hydraulic safety margin-the difference between typical minimum xylem water potential and that causing xylem dysfunction-and xylem vulnerability to embolism. Angiosperms and gymnosperms experienced roughly equal mortality risks. Our results provide broad support for the hypothesis that hydraulic traits capture key mechanisms determining tree death and highlight that physiological traits can improve vegetation model prediction of tree mortality during climate extremes.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1046/j.1469-8137.1999.00446.xURL [本文引用: 4]
DOI:10.1038/s41586-018-0240-xURL [本文引用: 2]
DOI:10.1038/nature11688URL [本文引用: 2]
DOI:10.1111/nph.2011.191.issue-4URL [本文引用: 1]
DOI:10.1111/1365-2745.13051 [本文引用: 1]
1. Temperate forests are predicted to experience an increased frequency and intensity of climate change-induced summer droughts and heat waves in the near future. Yet, while previous studies clearly showed a high drought sensitivity of different temperate tree species, the vulnerability of the physiological integrity of these trees remains unclear. 2. Here, we assessed the sensitivity of six temperate tree species to severe water limitation during three consecutive growing seasons, including the exceptional 2015 central European summer drought and heat wave. Specifically, we assessed stem increment growth, sap flow, water potentials, hydraulic vulnerability, and nonstructural carbohydrate contents in leaves and branches to determine how mature temperate trees responded to this exceptional weather event and how the observed responses relate to variation in xylem embolism and carbohydrate economy. 3. We found that the trees' predawn water potentials reached their minimum values during the 2015 summer drought and most species reduced their sap flow by up to 80%. Also, increment growth was strongly impaired with the onset of the drought in all species. Despite the strong responses in the trees' growth and water relations, all species exhibited minimum midday shoot water potentials well away from values associated with severe embolism (P-50). In addition, we detected no distinct decrease in nonstructural carbohydrate contents in leaves, bark, and stems throughout the drought event. 4. Synthesis. This study shows that mature individuals of six common central European forest tree species strongly reacted to a severe summer drought by reducing their water consumption and stopping growth. We found, however, no indications for xylem embolism or carbohydrate depletion in these trees. This suggests, that xylem embolism formation and carbohydrate reserve depletion are not routine in temperate trees during seasonal strong drought and reveals a low vulnerability of the physiological integrity of temperate trees during drought events as we describe here.
DOI:10.3732/ajb.93.11.1588URL [本文引用: 1]
URL [本文引用: 1]
DOI:10.1111/ppl.2011.142.issue-3URL [本文引用: 1]
URL [本文引用: 1]
[本文引用: 2]
[本文引用: 2]
DOI:10.1111/j.1469-8137.2009.02783.xPMID:19309447 [本文引用: 5]
While tracheid size of conifers is often a good proxy of water transport efficiency, correlations between conifer wood structure and transport safety remain poorly understood. It is hypothesized that at least some of the variation in bordered pit and tracheid structure is associated with both transport efficiency and embolism resistance. Stem and root samples from three boreal Pinaceae species were collected to test this hypothesis. Tracheid and pit anatomy were studied using light microscopy as well as scanning and transmission electron microscopy. While tracheid size explained at least 90% of the variation in specific conductivity for stem and root samples, the strongest correlations with embolism resistance occurred at the pit level. Both torus thickness and depth of the pit chamber showed a linear increase with greater vulnerability to cavitation. Greater embolism resistance was correlated with increasing wood density and tracheid wall reinforcement. A thinner torus may be more flexible and better able to seal the pit aperture. The pit chamber depth is proportional to the distance that the margo needs to deflect for pit aspiration.
DOI:10.1078/1439-1791-00006URL [本文引用: 1]
DOI:10.3732/ajb.91.3.386PMID:21653394 [本文引用: 1]
A model of xylem conduit function was applied to gymnosperm tracheids with torus-margo pit membranes for comparison with angiosperm vessels. Tracheids from 17 gymnosperm tree species with circular bordered pits and air-seed pressures from 0.8 to 11.8 MPa were analyzed. Tracheids were more reinforced against implosion than vessels, consistent with their double function in transport and support. Tracheid pits were 3.3 to 44 times higher in hydraulic conductivity than vessel pits because of greater membrane conductivity of the torus-margo configuration. Tight scaling between torus and pit size maximized pit conductivity. Higher pit conductivity allowed tracheids to be 1.7-3.4 times shorter than vessels and still achieve 95% of their lumen-limited maximum conductivity. Predicted tracheid lengths were consistent with measured lengths. The torus-margo structure is important for maximizing the conductivity of the inherently length-limited tracheid: replacing the torus-margo membrane with a vessel membrane caused stem tracheid conductivity to drop by 41%. Tracheids were no less hydraulically efficient than vessels if they were long enough to reach their lumen-limiting conductivity. However, this may only be possible for lumen diameters below approximately 60-70 μm.
DOI:10.1007/s004420100628PMID:28547229 [本文引用: 5]
Wood density (D ), an excellent predictor of mechanical properties, is typically viewed in relation to support against gravity, wind, snow, and other environmental forces. In contrast, we show the surprising extent to which variation in D and wood structure is linked to support against implosion by negative pressure in the xylem pipeline. The more drought-tolerant the plant, the more negative the xylem pressure can become without cavitation, and the greater the internal load on the xylem conduit walls. Accordingly, D was correlated with cavitation resistance. This trend was consistent with the maintenance of a safety factor from implosion by negative pressure: conduit wall span (b) and thickness (t) scaled so that (t/b) was proportional to cavitation resistance as required to avoid wall collapse. Unexpectedly, trends in D may be as much or more related to support of the xylem pipeline as to support of the plant.
PMID:16510385 [本文引用: 1]
We tested the hypothesis that greater cavitation resistance correlates with less total inter-vessel pit area per vessel (the pit area hypothesis) and evaluated a trade-off between cavitation safety and transport efficiency. Fourteen species of diverse growth form (vine, ring- and diffuse-porous tree, shrub) and family affinity were added to published data predominately from the Rosaceae (29 species total). Two types of vulnerability-to-cavitation curves were found. Ring-porous trees and vines showed an abrupt drop in hydraulic conductivity with increasing negative pressure, whereas hydraulic conductivity in diffuse-porous species generally decreased gradually. The ring-porous type curve was not an artifact of the centrifuge method because it was obtained also with the air-injection technique. A safety versus efficiency trade-off was evident when curves were compared across species: for a given pressure, there was a limited range of optimal vulnerability curves. The pit area hypothesis was supported by a strong relationship (r2 = 0.77) between increasing cavitation resistance and diminishing pit membrane area per vessel (A(P)). Small A(P) was associated with small vessel surface area and hence narrow vessel diameter (D) and short vessel length (L)--consistent with an increase in vessel flow resistance with cavitation resistance. This trade-off was amplified at the tissue level by an increase in xylem/vessel area ratio with cavitation resistance. Ring-porous species were more efficient than diffuse-porous species on a vessel basis but not on a xylem basis owing to higher xylem/vessel area ratios in ring-porous anatomy. Across four orders of magnitude, lumen and end-wall resistivities maintained a relatively tight proportionality with a near-optimal mean of 56% of the total vessel resistivity residing in the end-wall. This was consistent with an underlying scaling of L to D(3/2) across species. Pit flow resistance did not increase with cavitation safety, suggesting that cavitation pressure was not related to mean pit membrane porosity.
[本文引用: 2]
[本文引用: 1]
[本文引用: 1]
DOI:10.3732/ajb.0800248URL [本文引用: 1]
DOI:10.1093/treephys/tpw031PMID:27146334 [本文引用: 3]
Water transport from soils to the atmosphere is critical for plant growth and survival. However, we have a limited understanding about many portions of the whole-tree hydraulic pathway, because the vast majority of published information is on terminal branches. Our understanding of mature tree trunk hydraulic physiology, in particular, is limited. The hydraulic vulnerability segmentation hypothesis (HVSH) stipulates that distal portions of the plant (leaves, branches and roots) should be more vulnerable to embolism than trunks, which are nonredundant organs that require a massive carbon investment. In the current study, we compared vulnerability to loss of hydraulic function, leaf and xylem water potentials and the resulting hydraulic safety margins (in relation to the water potential causing 50% loss of hydraulic conductivity) in leaves, branches, trunks and roots of four angiosperms and four conifer tree species. Across all species, our results supported strongly the HVSH as leaves and roots were less resistant to embolism than branches or trunks. However, branches were consistently more resistant to embolism than any other portion of the plant, including trunks. Also, calculated whole-tree vulnerability to hydraulic dysfunction was much greater than vulnerability in branches. This was due to hydraulic dysfunction in roots and leaves at less negative water potentials than those causing branch or trunk dysfunction. Leaves and roots had narrow or negative hydraulic safety margins, but trunks and branches maintained positive safety margins. By using branch-based hydraulic information as a proxy for entire plants, much research has potentially overestimated embolism resistance, and possibly drought tolerance, for many species. This study highlights the necessity to reconsider past conclusions made about plant resistance to drought based on branch xylem only. This study also highlights the necessity for more research of whole-plant hydraulic physiology to better understand strategies of plant drought tolerance and the critical control points within the hydraulic pathway.© The Author 2016. Published by Oxford University Press. All rights reserved. For Permissions, please email: journals.permissions@oup.com.
DOI:10.1111/nph.13035PMID:25250668 [本文引用: 1]
This review presents a framework for evaluating how cells, tissues, organs, and whole plants perform both hydraulic and mechanical functions. The morphological alterations that affect dual functionality are varied: individual cells can have altered morphology; tissues can have altered partitioning to functions or altered cell alignment; and organs and whole plants can differ in their allocation to different tissues, or in the geometric distribution of the tissues they have. A hierarchical model emphasizes that morphological traits influence the hydraulic or mechanical properties; the properties, combined with the plant unit's environment, then influence the performance of that plant unit. As a special case, we discuss the mechanisms by which the proxy property wood density has strong correlations to performance but without direct causality. Traits and properties influence multiple aspects of performance, and there can be mutual compensations such that similar performance occurs. This compensation emphasizes that natural selection acts on, and a plant's viability is determined by, its performance, rather than its contributing traits and properties. Continued research on the relationships among traits, and on their effects on multiple aspects of performance, will help us better predict, manage, and select plant material for success under multiple stresses in the future.© 2014 The Authors. New Phytologist © 2014 New Phytologist Trust.
[本文引用: 1]
DOI:10.1111/nph.2011.190.issue-3URL [本文引用: 3]
DOI:10.1002/j.1537-2197.1992.tb13712.xPMID:30139134 [本文引用: 1]
A method for estimating the hydraulic diameter of a pore or conduit having a noncircular opening is presented with special reference to plant anatomy. An ellipse or a rectangle is inscribed within the opening, and the length of the short axis (a) is measured. The hydraulic diameter (D ) is estimated for the ellipse (D = 1.4a) or rectangle (D = 2a). Use of these equations often gives a more accurate estimate of the hydraulic diameter of a pore or conduit than does averaging the lengths of the short and long axes (D = [a + b]/2, b is the long axis) or assuming that the opening is circular (D = a). A table of the error in each method is included, and the errors inherent in the use of D are discussed. Because fewer measurements are required, estimation based on the measurement of one axis is much faster than calculation using both the short and long axes. The equations and table should permit anatomists and physiologists to rapidly determine the best method for estimating the hydraulic diameter of a pore or conduit, and to more accurately and quickly estimate the hydraulic diameters of large numbers of openings. However, because of potential pitfalls in applying theoretical fluid dynamics equations to real-world functional anatomy, botanists must ensure that their applications of hydraulic diameter are appropriate in each case.© 1992 Botanical Society of America.
DOI:10.1163/22941932-20160128URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
PMID:17509617 [本文引用: 2]
The xylem is one of the two long distance transport tissues in plants, providing a low resistance pathway for water movement from roots to leaves. Its properties determine how much water can be transported and transpired and, at the same time, the plant's vulnerability to transport dysfunctions (the formation and propagation of emboli) associated to important stress factors, such as droughts and frost. Both maximum transport efficiency and safety against embolism have classically been attributed to the properties of individual conduits or of the pit membrane connecting them. But this approach overlooks the fact that the conduits of the xylem constitute a network. The topology of this network is likely to affect its overall transport properties, as well as the propagation of embolism through the xylem, since, according to the air-seeding hypothesis, drought-induced embolism propagates as a contact process (i.e., between neighbouring conduits). Here we present a model of the xylem that takes into account its system-level properties, including the connectivity of the xylem network. With the tools of graph theory and assuming steady state and Darcy's flow we calculated the hydraulic conductivity of idealized wood segments at different water potentials. A Monte Carlo approach was adopted, varying the anatomical and topological properties of the segments within biologically reasonable ranges, based on data available from the literature. Our results showed that maximum hydraulic conductivity and vulnerability to embolism increase with the connectivity of the xylem network. This can be explained by the fact that connectivity determines the fraction of all the potential paths or conduits actually available for water transport and spread of embolism. It is concluded that the xylem can no longer be interpreted as the mere sum of its conduits, because the spatial arrangement of those conduits in the xylem network influences the main functional properties of this tissue. This brings new arguments into the long-standing discussion on the efficiency vs. safety trade-off in the plants' xylem.
[本文引用: 2]
DOI:10.1111/nph.16043PMID:31276231 [本文引用: 1]
Trees may survive prolonged droughts by shifting water uptake to reliable water sources, but it is unknown if the dominant mechanism involves activating existing roots or growing new roots during drought, or some combination of the two. To gain mechanistic insights on this unknown, a dynamic root-hydraulic modeling framework was developed that set up a feedback between hydraulic controls over carbon allocation and the role of root growth on soil-plant hydraulics. The new model was tested using a 5 yr drought/heat field experiment on an established piñon-juniper stand with root access to bedrock groundwater. Owing to the high carbon cost per unit root area, modeled trees initialized without adequate bedrock groundwater access experienced potentially lethal declines in water potential, while all of the experimental trees maintained nonlethal water potentials. Simulated trees were unable to grow roots rapidly enough to mediate the hydraulic stress, particularly during warm droughts. Alternatively, modeled trees initiated with root access to bedrock groundwater matched the hydraulics of the experimental trees by increasing their water uptake from bedrock groundwater when soil layers dried out. Therefore, the modeling framework identified a critical mechanism for drought response that required trees to shift water uptake among existing roots rather than growing new roots.© 2019 The Authors. New Phytologist © 2019 New Phytologist Trust.
DOI:10.1111/ele.12851PMID:28922708 [本文引用: 1]
Stomata play a significant role in the Earth's water and carbon cycles, by regulating gaseous exchanges between the plant and the atmosphere. Under drought conditions, stomatal control of transpiration has long been thought to be closely coordinated with the decrease in hydraulic capacity (hydraulic failure due to xylem embolism). We tested this hypothesis by coupling a meta-analysis of functional traits related to the stomatal response to drought and embolism resistance with simulations from a soil-plant hydraulic model. We report here a previously unreported phenomenon: the existence of an absolute limit by which stomata closure must occur to avoid rapid death in drought conditions. The water potential causing stomatal closure and the xylem pressure at the onset of embolism formation were equal for only a small number of species, and the difference between these two traits (i.e. safety margins) increased continuously with increasing embolism resistance. Our findings demonstrate the need to revise current views about the functional coordination between stomata and hydraulic traits and provide a mechanistic framework for modeling plant mortality under drought conditions.© 2017 John Wiley & Sons Ltd/CNRS.
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1469-8137.2008.02436.xPMID:18422905 [本文引用: 1]
Severe droughts have been associated with regional-scale forest mortality worldwide. Climate change is expected to exacerbate regional mortality events; however, prediction remains difficult because the physiological mechanisms underlying drought survival and mortality are poorly understood. We developed a hydraulically based theory considering carbon balance and insect resistance that allowed development and examination of hypotheses regarding survival and mortality. Multiple mechanisms may cause mortality during drought. A common mechanism for plants with isohydric regulation of water status results from avoidance of drought-induced hydraulic failure via stomatal closure, resulting in carbon starvation and a cascade of downstream effects such as reduced resistance to biotic agents. Mortality by hydraulic failure per se may occur for isohydric seedlings or trees near their maximum height. Although anisohydric plants are relatively drought-tolerant, they are predisposed to hydraulic failure because they operate with narrower hydraulic safety margins during drought. Elevated temperatures should exacerbate carbon starvation and hydraulic failure. Biotic agents may amplify and be amplified by drought-induced plant stress. Wet multidecadal climate oscillations may increase plant susceptibility to drought-induced mortality by stimulating shifts in hydraulic architecture, effectively predisposing plants to water stress. Climate warming and increased frequency of extreme events will probably cause increased regional mortality episodes. Isohydric and anisohydric water potential regulation may partition species between survival and mortality, and, as such, incorporating this hydraulic framework may be effective for modeling plant survival and mortality under future climate conditions.
DOI:10.1111/j.1469-8137.2004.01127.xPMID:33873731 [本文引用: 3]
•? To assess hydraulic architecture and limitations to water transport across whole trees, we compared xylem anatomy, vulnerability to cavitation (Ψ ) and specific hydraulic conductivity (K ) of stems, shallow roots and deep roots (from caves to 20 m depth) for four species: Juniperus ashei, Bumelia lanuginosa, Quercus fusiformis and Quercus sinuata. •? Mean, maximum and hydraulically weighted (D ) conduit diameters and K were largest in deep roots, intermediate in shallow roots, and smallest in stems (P < 0.05 for each). Mean vessel diameters of deep roots were 2.1-4.2-fold greater than in stems, and K was seven to 38 times larger in the deep roots. •? Ψ also increased from stems to roots with depth, as much as 24-fold from stems to deep roots in B. lanuginosa. For all species together, Ψ was positively correlated with both D and K, suggesting a potential trade-off exists between conducting efficiency and safety. •? The anatomical and hydraulic differences documented here suggest that the structure of deep roots minimizes flow resistance and maximizes deep water uptake.
DOI:10.1111/nph.2009.182.issue-2URL [本文引用: 1]
DOI:10.1093/jxb/eraa381URL [本文引用: 2]
DOI:10.1104/pp.110.158824PMID:20551212 [本文引用: 2]
Water transport in conifers occurs through single-celled tracheids that are connected to one another via intertracheid pit membranes. These membranes have two components: the porous margo, which allows water to pass through the membrane, and the impermeable torus, which functions to isolate gas-filled tracheids. During drought, tracheids can become air filled and thus hydraulically dysfunctional, a result of air entering through the pit membrane and nucleating cavitation in the water column. What are the hydraulic tradeoffs associated with cavitation resistance at the pit level, and how do they vary within the structural components of the intertracheid pit? To address these questions, we examined pit structure in 15 species of Cupressaceae exhibiting a broad range of cavitation resistances. Across species, cavitation resistance was most closely correlated to the ratio of the torus to pit aperture diameter but did not vary systematically with margo porosity. Furthermore, our data indicate that constraints on pit hydraulic efficiency are shared: the pit aperture limits pit conductivity in more drought-resistant taxa, while increased margo resistance is more likely to control pit conductivity in species that are more vulnerable to cavitation. These results are coupled with additional data concerning pit membrane structure and function and are discussed in the context of the evolutionary biogeography of the Cupressaceae.
PMID:16377751 [本文引用: 6]
Ice formation in the xylem sap produces air bubbles that under negative xylem pressures may expand and cause embolism in the xylem conduits. We used the centrifuge method to evaluate the relationship between freeze-thaw embolism and conduit diameter across a range of xylem pressures (Px) in the conifers Pinus contorta and Juniperus scopulorum. Vulnerability curves showing loss of conductivity (embolism) with Px down to -8 MPa were generated with versus without superimposing a freeze-thaw treatment. In both species, the freeze-thaw plus water-stress treatment caused more embolism than water stress alone. We estimated the critical conduit diameter (Df) above which a tracheid will embolize due to freezing and thawing and found that it decreased from 35 microm at a Px of -0.5 MPa to 6 microm at -8 MPa. Further analysis showed that the proportionality between diameter of the air bubble nucleating the cavitation and the diameter of the conduit (kL) declined with increasingly negative Px. This suggests that the bubbles causing cavitation are smaller in proportion to tracheid diameter in narrow tracheids than in wider ones. A possible reason for this is that the rate of dissolving increases with bubble pressure, which is inversely proportional to bubble diameter (La Place's law). Hence, smaller bubbles shrink faster than bigger ones. Last, we used the empirical relationship between Px and Df to model the freeze-thaw response in conifer species.
PMID:14532014 [本文引用: 1]
We tested the hypotheses that freezing-induced embolism is related to conduit diameter, and that conifers and angiosperms with conduits of equivalent diameter will exhibit similar losses of hydraulic conductivity in response to freezing. We surveyed the freeze-thaw response of conifers with a broad range of tracheid diameters by subjecting wood segments (root, stem and trunk wood) to a freeze-thaw cycle at -0.5 MPa in a centrifuge. Embolism increased as mean tracheid diameter exceeded 30 microm. Tracheids with a critical diameter greater than 43 microm were calculated to embolize in response to freezing and thawing at a xylem pressure of -0.5 MPa. To confirm that freezing-induced embolism is a function of conduit air content, we air-saturated stems of Abies lasiocarpa (Hook.) Nutt. (mean conduit diameter 13.7 +/- 0.7 microm) by pressurizing them 1 to 60 times above atmospheric pressure, prior to freezing and thawing. The air saturation method simulated the effect of increased tracheid size because the degree of super-saturation is proportional to a tracheid volume holding an equivalent amount of dissolved air at ambient pressure. Embolism increased when the dissolved air content was equivalent to a mean tracheid diameter of 30 microm at ambient air pressure. Our centrifuge and air-saturation data show that conifers are as vulnerable to freeze-thaw embolism as angiosperms with equal conduit diameter. We suggest that the hydraulic conductivity of conifer wood is maximized by increasing tracheid diameters in locations where freezing is rare. Conversely, the narrowing of tracheid diameters protects against freezing-induced embolism in cold climates.
[本文引用: 6]
DOI:10.3732/ajb.93.9.1265URL [本文引用: 1]
DOI:10.1111/nph.2007.174.issue-4URL [本文引用: 3]
DOI:10.1890/06-0780URL [本文引用: 1]
DOI:10.1038/nature15539URL [本文引用: 1]
DOI:10.1111/nph.15079PMID:29528498 [本文引用: 1]
The capacity of plant species to resist xylem cavitation is an important determinant of resistance to drought, mortality thresholds, geographic distribution and productivity. Unravelling the role of xylem cavitation vulnerability in plant evolution and adaptation requires a clear understanding of how this key trait varies between the tissues of individuals and between individuals of species. Here, we examine questions of variation within individuals by measuring how cavitation moves between organs of individual plants. Using multiple cameras placed simultaneously on roots, stems and leaves, we were able to record systemic xylem cavitation during drying of individual olive plants. Unlike previous studies, we found a consistent pattern of root > stem > leaf in terms of xylem resistance to cavitation. The substantial variation in vulnerability to cavitation, evident among individuals, within individuals and within tissues of olive seedlings, was coordinated such that plants with more resistant roots also had more resistant leaves. Preservation of root integrity means that roots can continue to supply water for the regeneration of drought-damaged aerial tissues after post-drought rain. Furthermore, coordinated variation in vulnerability between leaf, stem and root in olive plants suggests a strong selective pressure to maintain a fixed order of cavitation during drought.© 2018 The Authors. New Phytologist © 2018 New Phytologist Trust.
DOI:10.1111/nph.16177PMID:31498457 [本文引用: 1]
Efficient water transport from soil to leaves sustains stomatal opening and steady-state photosynthesis. The aboveground portion of this pathway is well-described, yet the roots and their connection with the soil are still poorly understood due to technical limitations. Here we used a novel rehydration technique to investigate changes in the hydraulic pathway between roots and soil and within the plant body as individual olive plants were subjected to a range of water stresses. Whole root hydraulic resistance (including the radial pathway from xylem to the soil-root interface) constituted 81% of the whole-plant resistance in unstressed plants, increasing to > 95% under a moderate level of water stress. The decline in this whole root hydraulic conductance occurred in parallel with stomatal closure and contributed significantly to the reduction in canopy conductance according to a hydraulic model. Our results demonstrate that losses in root hydraulic conductance, mainly due to a disconnection from the soil during moderate water stress in olive plants, are profound and sufficient to induce stomatal closure before cavitation occurs. Future studies will determine whether this core regulatory role of root hydraulics exists more generally among diverse plant species.© 2019 The Authors. New Phytologist © 2019 New Phytologist Trust.
[本文引用: 2]
[本文引用: 2]
DOI:10.1007/s00468-019-01874-xURL [本文引用: 4]
DOI:10.1111/nph.13437PMID:25944400 [本文引用: 3]
The flow of xylem sap in conifers is strongly dependent on the presence of a low resistance path through bordered pits, particularly through the pores present in the margo of the pit membrane. A computational fluid dynamics approach was taken, solving the Navier-Stokes equation for models based on the geometry of pits observed in tracheids from stems and roots of Picea mariana (black spruce) and Picea glauca (white spruce). Model solutions demonstrate a close, inverse relationship between the total resistance of bordered pits and the total area of margo pores. Flow through the margo was dominated by a small number of the widest pores. Particularly for pits where the margo component of flow resistance was low relative to that of the torus, pore location near the inner edge of the margo allowed for greater flow than that occurring through similar-sized pores near the outer edge of the margo. Results indicate a surprisingly large variation in pit structure and flow characteristics. Nonetheless, pits in roots have lower resistance to flow than those in stems because the pits were wider and consisted of a margo with a larger area in pores. © 2015 The Authors. New Phytologist © 2015 New Phytologist Trust.
DOI:10.1111/nph.2016.210.issue-2URL [本文引用: 5]
DOI:10.3732/ajb.93.10.1490PMID:21642096 [本文引用: 2]
The wide size range of conifer tracheids and angiosperm vessels has important consequences for function. In both conduit types, bigger is better for conducting efficiency. The gain in efficiency with size is maximized by the control of conduit shape, which balances end-wall and lumen resistances. Although vessels are an order of magnitude longer than tracheids of the same diameter, they are not necessarily more efficient because they lack the low end-wall resistance of tracheids with torus-margo pits. Instead, vessels gain conducting efficiency over tracheids by achieving wider maximum diameters. End-walls contributed 56-64% to total xylem resistance in both conduit types, indicating that length limits conducting efficiency. Tracheid dimensions may be more limited by unicellularity and the need to supply strength to homoxylous wood than by the need to protect against cavitation. In contrast, the greater size of the multicellular vessel is facilitated by fibers that strengthen heteroxylous wood. Vessel dimensions may be most limited by the need to restrict intervessel pitting and cavitation by air-seeding. Stressful habitats that promote narrow vessels should favor coexistence of conifers and angiosperms. The evolution of vessels in angiosperm wood may have required early angiosperms to survive a phase of mechanic and hydraulic instability.
DOI:10.1163/22941932-90001369URL [本文引用: 1]
[本文引用: 2]
DOI:10.1046/j.1469-8137.2000.00549.xURL [本文引用: 4]
DOI:10.3724/SP.J.1258.2013.00100URL [本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1258.2012.00019URL [本文引用: 1]
[本文引用: 1]
DOI:10.1007/s11258-009-9592-5URL [本文引用: 1]
A multi-species synthesis of physiological mechanisms in drought-induced tree mortality
1
2017
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Climate-induced forest dieback: an escalating global phenomenon
1
2009
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought- induced tree mortality across the globe
1
2016
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Evolution of xylem physiology//Hemsley AR, Poole I. The Evolution of Plant Physiology. Elsevier,
1
2004
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
A functional analysis of podocarp ecology
1
2011
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
Stem hydraulic supply is linked to leaf photosynthetic capacity: evidence from New Caledonian and Tasmanian rainforests
1
2000
... 木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(
The importance of xylem constraints in the distribution of conifer species
4
1999
... 罗汉松科3种植物茎Ks和P50无关(p > 0.05,
... Relationship between hydraulic conductivity and embolism resistance in the three Podocarpaceae species. A, The stems of three Podocarpaceae species in this paper. B, The roots of three Podocarpaceae species in this paper. C, The stems of twelve Podocarpaceae species (Data from
... 罗汉松科3种植物茎中, 短叶罗汉松是茎栓塞抗性最强的物种, 与
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
Triggers of tree mortality under drought
2
2018
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
Global convergence in the vulnerability of forests to drought
2
2012
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
... 木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(
Xylem traits mediate a trade-off between resistance to freeze-thaw-induced embolism and photosynthetic capacity in overwintering evergreens
1
2011
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
No role for xylem embolism or carbohydrate shortage in temperate trees during the severe 2015 drought
1
2019
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Bordered pit structure and function determine spatial patterns of air-seeding thresholds in xylem of Douglas-fir (Pseudotsuga menziesii; Pinaceae) trees
1
2006
... 本研究表明茎管胞直径与输水效率和栓塞抗性没有联系, 而纹孔膜直径会影响茎输水效率和栓塞抗性(
Fitplc—An R package to fit hydraulic vulnerability curves
1
2017
... 使用空气注入法(双端压力套技术)构建茎段或根段的脆弱性曲线.过去的研究表明了使用此法可以获得可靠的水力脆弱性曲线(
How reliable is the double-ended pressure sleeve technique for assessing xylem vulnerability to cavitation in woody angiosperms?
1
2011
... 使用空气注入法(双端压力套技术)构建茎段或根段的脆弱性曲线.过去的研究表明了使用此法可以获得可靠的水力脆弱性曲线(
Tradeoff between stem hydraulic efficiency and mechanical strength affects leaf-stem allometry in 28 Ficus tree species
1
2017
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
Common trade-offs between xylem resistance to cavitation and other physiological traits do not hold among unrelated Populus deltoides × Populus nigra hybrids
2
2010
... 罗汉松科3种植物茎中, 短叶罗汉松是茎栓塞抗性最强的物种, 与
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world’s woody plant species
2
2016
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
... ).此外, 其他木质部特有的权衡可能会混淆效率-安全权衡关系(
Embolism resistance of three boreal conifer species varies with pit structure
5
2009
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
... ).其次, 对于多数针叶树而言, 根的管胞较宽较长, 纹孔较大(
... 本研究表明茎管胞直径与输水效率和栓塞抗性没有联系, 而纹孔膜直径会影响茎输水效率和栓塞抗性(
Drought experience and cavitation resistance in six shrubs from the Great Basin, Utah
1
2000
... 根据
Analysis of circular bordered pit function II. Gymnosperm tracheids with torus- margo pit membranes
1
2004
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure
5
2001
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
... 的比值)与栓塞抗性有关(
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
... 罗汉松科3种植物茎中, 短叶罗汉松是茎栓塞抗性最强的物种, 与
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
Scaling of angiosperm xylem structure with safety and efficiency
1
2006
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
An ecophysiological and developmental perspective on variation in vessel diameter
2
2017
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... ;
Intraspecific variation in wood anatomical, hydraulic, and foliar traits in ten European beech provenances differing in growth yield
1
2016
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
1
2013
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Morphological variation of intervessel pit membranes and implications to xylem function in angiosperms
1
2009
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
A test of the hydraulic vulnerability segmentation hypothesis in angiosperm and conifer tree species
3
2016
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
... 使用空气注入法(双端压力套技术)构建茎段或根段的脆弱性曲线.过去的研究表明了使用此法可以获得可靠的水力脆弱性曲线(
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
Traits, properties, and performance: How woody plants combine hydraulic and mechanical functions in a cell, tissue, or whole plant
1
2014
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
Aridity drove the evolution of extreme embolism resistance and the radiation of conifer genus
1
2017
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer
3
2011
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 本研究的罗汉松科3种植物根木质部存在效率-安全权衡, 但茎木质部不存在(
... )植物的研究表明木质部的关键性状与输水效率和栓塞抗性的密切关系是解释安全-效率权衡的基础.对木质部性状的相关性分析表明了根输水效率和栓塞抗性对结构需求是对立的, 例如根管胞厚度跨度比、管胞壁厚和木材密度是与更强栓塞抗性和更低输水效率相关的性状, 管胞直径和水力直径是与更高输水效率和更弱栓塞抗性相关的性状.本研究与
Measuring the hydraulic diameter of a pore or conduit
1
1992
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem
1
2016
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
子午岭林区3种典型树木的水力结构特性比较
1
2014
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
子午岭林区3种典型树木的水力结构特性比较
1
2014
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
The relevance of xylem network structure for plant hydraulic efficiency and safety
2
2007
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
... 本研究的罗汉松科3种植物根木质部存在效率-安全权衡, 但茎木质部不存在(
Functional coordination between leaf gas exchange and vulnerability to xylem cavitation in temperate forest trees
2
2006
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
... ;
Conifers depend on established roots during drought: results from a coupled model of carbon allocation and hydraulics
1
2020
... 目前的研究集中于木本植物茎木质部性状的研究, 由于根深埋于土壤中, 比地上植物器官更难接近, 并且根的建筑复杂性为样品采样和标准水力测量方案带来了不确定性(
Plant resistance to drought depends on timely stomatal closure
1
2017
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
A new method for vulnerability analysis of small xylem areas reveals that compression wood of Norway spruce has lower hydraulic safety than opposite wood
1
2003
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
A dynamic yet vulnerable pipeline: integration and coordination of hydraulic traits across whole plants
1
2019
... 木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(
Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought?
1
2008
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Variation in xylem structure and function in stems and roots of trees to 20 m depth
3
2004
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
Hierarchical statistical modeling of xylem vulnerability to cavitation
1
2009
... 使用空气注入法(双端压力套技术)构建茎段或根段的脆弱性曲线.过去的研究表明了使用此法可以获得可靠的水力脆弱性曲线(
Non-invasive imaging reveals convergence in root and stem vulnerability to cavitation across five tree species
2
2020
... 目前的研究集中于木本植物茎木质部性状的研究, 由于根深埋于土壤中, 比地上植物器官更难接近, 并且根的建筑复杂性为样品采样和标准水力测量方案带来了不确定性(
... ;
The relationships between xylem safety and hydraulic efficiency in the Cupressaceae: the evolution of pit membrane form and function
2
2010
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 本研究表明茎管胞直径与输水效率和栓塞抗性没有联系, 而纹孔膜直径会影响茎输水效率和栓塞抗性(
Analysis of freeze-thaw embolism in conifers. The interaction between cavitation pressure and tracheid size
6
2006
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 罗汉松科3种植物茎Ks和P50无关(p > 0.05,
... ).D, 罗汉松科5种植物根(数据来源于
... ,
... ). D, The roots of five Podocarpaceae species (Data from
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
Tracheid diameter is the key trait determining the extent of freezing-induced embolism in conifers
1
2003
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem
6
2006a
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
... ;
... ).因此, 机械强度也需要支持维管网络, 阻止因木质部负压引起的管道内爆(
... 提出木质部“权衡”三角形, 权衡可能存在于木质部输水效率、安全性和机械强度之间.权衡可能会因木质部的过度建造而消失, 例如罗汉松科植物木质部相对于栓塞抗性有着更大的密度、厚度跨度之比, 且与栓塞抗性解耦联(
... 罗汉松科植物茎机械性状(WD, (t/b)2)与茎输水效率和栓塞抗性无关, 无论是细胞水平((t/b)2), 还是组织水平(WD)(
... 对罗汉松科植物的研究结果相似, 其栓塞抗性与机械强度参数无关.
Inter-tracheid pitting and the hydraulic efficiency of conifer wood: the role of tracheid allometry and cavitation protection
1
2006b
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
Relationships among xylem transport, biomechanics and storage in stems and roots of nine Rhamnaceae species of the California chaparral
3
2007a
... 目前的研究集中于木本植物茎木质部性状的研究, 由于根深埋于土壤中, 比地上植物器官更难接近, 并且根的建筑复杂性为样品采样和标准水力测量方案带来了不确定性(
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
Life history type and water stress tolerance in nine California chaparral species (Rhamnaceae)
1
2007b
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
Death from drought in tropical forests is triggered by hydraulics not carbon starvation
1
2015
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
Mapping xylem failure in disparate organs of whole plants reveals extreme resistance in olive roots
1
2018
... 目前的研究集中于木本植物茎木质部性状的研究, 由于根深埋于土壤中, 比地上植物器官更难接近, 并且根的建筑复杂性为样品采样和标准水力测量方案带来了不确定性(
Declining root water transport drives stomatal closure in olive under moderate water stress
1
2020
... 目前的研究集中于木本植物茎木质部性状的研究, 由于根深埋于土壤中, 比地上植物器官更难接近, 并且根的建筑复杂性为样品采样和标准水力测量方案带来了不确定性(
Adjustments and coordination of hydraulic, leaf and stem traits along a water availability gradient
2
2019
... 目前气候模型预测全球范围内干旱频率和强度都将增加(
... 罗汉松科3种植物茎中, 短叶罗汉松是茎栓塞抗性最强的物种, 与
Hydraulic acclimation to shading in boreal conifers of varying shade tolerance
2
2010
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
Xylem hydraulic safety and efficiency in relation to leaf and wood traits in three temperate Acer species differing in habitat preferences
4
2019
... 木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
... 使用空气注入法(双端压力套技术)构建茎段或根段的脆弱性曲线.过去的研究表明了使用此法可以获得可靠的水力脆弱性曲线(
Pit membrane structure is highly variable and accounts for a major resistance to water flow through tracheid pits in stems and roots of two boreal conifer species
3
2015
... 罗汉松科3种植物根管胞和纹孔膜直径均比茎大(
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
... 本研究表明茎管胞直径与输水效率和栓塞抗性没有联系, 而纹孔膜直径会影响茎输水效率和栓塞抗性(
How adaptable is the hydraulic system of European beech in the face of climate change-related precipitation reduction?
5
2016
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 水力功能性状与木质部的机械强度是相互依赖的.在针叶树中尤其明显, 因为针叶树的管胞不仅具有输水功能, 而且还具有对植物体的机械支持功能(
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
... ;
Size and function in conifer tracheids and angiosperm vessels
2
2006
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
Biophysical perspectives of xylem evolution: Is there a tradeoff of hydraulic efficiency for vulnerability to dysfunction?
1
1994
... 根管胞水力直径与根输水效率显著正相关但与栓塞抗性无关(
Xylem Structure and the Ascent of Sap.
2
2002
... 目前植物茎/根木质部相关的研究表明, 管道结构与其输水效率和安全性高度相关, 效率与安全在单个管道上的权衡导致在组织水平上的效率-安全权衡: 更高的栓塞抗性意味着更低的输水效率(
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
Xylem hydraulic characteristics of subtropical trees from contrasting habitats grown under identical environmental conditions
4
2000
... 木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(
... 罗汉松科3种植物茎Ks和P50无关(p > 0.05,
... ,
... 本研究中罗汉松科3种植物茎木质部不存在效率-安全权衡.对已发表的罗汉松科12种树种的数据进行分析, 发现它们的茎木质部也不存在权衡(
树轮生态学研究中微树芯石蜡切片制作的方法探讨
1
2013
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
树轮生态学研究中微树芯石蜡切片制作的方法探讨
1
2013
... 从测定导水率的茎段或根段上截取一段用于测定木质部解剖结构特征.样品固定软化之后, 采用石蜡切片法将其制成永久切片(
荒漠河岸林植物木质部导水与栓塞特征及其对干旱胁迫的响应
1
2012
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
荒漠河岸林植物木质部导水与栓塞特征及其对干旱胁迫的响应
1
2012
... 所有茎段或根段均在水下剪取一段.茎段平均长度为133.8 mm (SE = 1.78); 根段平均长度为140.4 mm (SE = 6.28).所有侧枝或侧根均被剪断, 并用螺纹密封胶带将产生的疤痕封闭.在进行水力测量之前, 用电子卡尺测定茎段或根段的长度和两端的直径.茎段的平均直径为4.01 mm (SE = 0.12), 根段的平均直径为4.02 mm (SE = 0.25).然后小心地将茎段或根段基部上部2-3 cm的表皮剥离.该部分连接到木质部导水率与栓塞测量系统XYL’EM-Plus (Bronkhorst, Montigny-les-Cormeilles, France), 用于测量导水率.测量溶液为20 mmol·L-1 KCl + 1 mmol·L-1 CaCl2溶液(
Hydraulic properties and photosynthetic rates in co-occurring lianas and trees in a seasonal tropical rainforest in southwestern China
1
2009
... 木质部水分运输效率(Ks)和导水率损失达到50%时的木质部水势(P50)是植物的两个重要水力功能性状(

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}