Spatial variability of phenological phenotype of Picea crassifolia in Qilian Mountains and its internal mechanism
Jun DU1, Wen WANG1,2, Zhi-Bin HE,1,*, Long-Fei CHEN1, Peng-Fei LIN1, Xi ZHU1, Quan-Yan TIAN11Linze Inland River Basin Research Station, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Aims The aim of this study is to address how the spatial variability of phenological phenotype is characterized in natural populations, and what drives such spatial variability (genetic variation or phenotype plasticity). Methods The phenological process of 156 sample trees in 20 plots of five altitudes in Pailugou watershed was monitored for two consecutive years, and the saplings selected at different altitudes were transplanted in common garden. The investigation data were analyzed using ridge regression and lasso regression. Important findings All phenological stages of Picea crassifolia were delayed with altitude (0.57-1.36 d per 100 m). This spatial variability reflected a certain degree of provenance effect (i.e. genetic variation), with an average contribution rate of 20.8% that maintained at a relatively stable level between years. An interesting finding is that the spring phenology timing of high-altitude populations in the common garden was significantly earlier than low-altitude populations, which may be related to the lower threshold of pre-season heat accumulation required for bud differentiation of high-altitude populations. In addition, we found that pre-season heat accumulation, altitude, pre-season precipitation and freezing injury frequency played key roles in explaining phenological dynamics at the population level. Keywords:phenology;spatial variability;genetic variation;phenotype plasticity;Picea crassifolia
PDF (529KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 杜军, 王文, 何志斌, 陈龙飞, 蔺鹏飞, 朱喜, 田全彦. 祁连山青海云杉物候表型的空间分异及其内在机制. 植物生态学报, 2021, 45(8): 834-843. DOI: 10.17521/cjpe.2021.0100 DU Jun, WANG Wen, HE Zhi-Bin, CHEN Long-Fei, LIN Peng-Fei, ZHU Xi, TIAN Quan-Yan. Spatial variability of phenological phenotype of Picea crassifolia in Qilian Mountains and its internal mechanism. Chinese Journal of Plant Ecology, 2021, 45(8): 834-843. DOI: 10.17521/cjpe.2021.0100
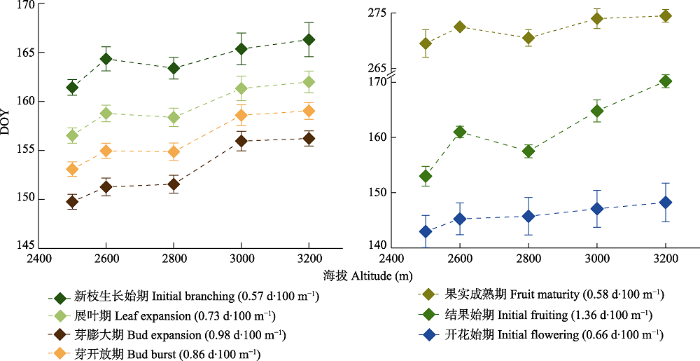
Fig. 1Variation of annual phenological stages of Picea crassifolia monitored in shady-slope plots (including small and large patches) in 2018 along the altitudinal gradient (mean ± SD). DOY, day of the year.
Table 1 表1 表1海拔、坡向及斑块大小对青海云杉种群物候空间分异的影响 Table 1Effects of altitude, aspect and patch size on the spatial differentiation of Picea crassifolia population phenology
影响因子 Impact factor
df
萌芽 Bud burst
展叶 Leaf expansion
抽枝 Branching
开花 Flowering
结果 Fruiting
MS
F
p
MS
F
p
MS
F
p
MS
F
p
MS
F
p
海拔 Altitude (AL)
4
155.23
38.70
0.00
216.80
82.50
0.00
239.76
72.91
0.00
63.27
4.44
0.00
849.72
744.96
0.00
坡向 Aspect (AS)
1
40.19
10.02
0.00
39.62
15.08
0.00
24.09
7.33
0.01
16.92
1.19
0.28
13.14
11.52
0.00
斑块大小 Patch size (PS)
1
0.95
0.24
0.63
1.83
0.70
0.41
1.65
0.50
0.48
0.07
0.01
0.94
0.02
0.01
0.91
AL × AS
4
1.58
0.39
0.71
2.09
0.80
0.53
1.42
0.43
0.79
0.12
0.01
1.00
8.14
7.14
0.00
AL × PS
4
0.39
0.10
0.98
0.72
0.28
0.89
0.61
0.19
0.95
0.40
0.03
1.00
2.35
2.06
0.12
AS × PS
1
40.76
10.16
0.00
22.16
8.43
0.01
5.89
1.79
0.19
59.16
4.15
0.05
1.27
1.11
0.30
AL × AS × PS
4
15.80
3.94
0.01
13.84
5.27
0.00
12.34
3.75
0.01
19.62
1.38
0.25
1.27
1.11
0.35
误差 Error
60
4.01
2.63
3.29
14.26
1.14
表中数字加粗部分表示差异显著(p < 0.05)。MS, 均方。 Bold font indicates significant difference (p < 0.05). MS, mean square.
Fig. 2Phenological pattern of Picea crassifolia populations in the common garden experiment versus elevation of provenance sites in 2018 (mean ± SD). DOY, day of the year.
Table 2 表2 表2青海云杉种群各物候阶段遗传分化所占比例 Table 2Genetic differentiation in phenological stages among populations of Picea crassifolia
阶段 Phase
2018
2019
D
F
p
D
F
p
萌芽期 Bud burst
13.4
2.669
0.043
19.9
3.636
0.011
展叶期 Leaf expansion
23.2
4.283
0.005
20.4
3.700
0.010
抽枝期 Branching
29.2
5.512
0.001
19.3
3.495
0.013
D, 方差分析估计的种源方差与总方差的比值(%); F, Fisher检验值; p, 种群效应的显著水平。加粗表示显著性差异(p < 0.05)。 D, the ratio (%) of the variance of provenance to total variance estimated by analysis of variance; F, Fisher’s test value; p, significance levels of the population effect. Bold indicates significant difference (p < 0.05).
Table 3 表3 表3青海云杉种群物候与环境要素的岭回归及Lasso回归分析结果 Table 3Ridge regression and Lasso regression analysis of the phenology of Picea crassifolia populations and environmental factors
变量 Variable
展叶 Leaf expansion (Scaled)
Lar序列 Lar sequence
开花 Flowering (Scaled)
Lar序列 Lar sequence
结果 Fruiting (Scaled)
Lar序列 Lar sequence
海拔 Altitude
4.80 (0.72)***
2
2.34 (0.69) ***
1
6.85 (0.23) ***
5
CH0(Oct.-Feb.)
-4.07 (0.38) ***
8
-1.42 (0.51) **
7
-7.00 (0.22) ***
10
CH5(Oct.-Feb.)
-2.52 (0.31) ***
-
-1.09 (0.43) *
-
-4.69 (0.18) ***
-
CDD0(Mar.-May)1
-5.93 (0.63) ***
5
-1.90 (0.57) ***
5
-9.49 (0.29) ***
-
CDD5(Mar.-May)1
-6.83 (0.79) ***
1
-2.13 (0.65) **
2
-10.65 (0.36) ***
1
CDD0(Mar.-May)2
0.74 (0.88)
6
-0.90 (0.94)
9
2.34 (0.45) ***
7
CDD5(Mar.-May)2
-
-
-
-
-
-
TR(Mar.-May)
1.01 (1.68)
9
1.27 (1.41)
6
4.27 (0.30) ***
9
RD(Mar.-May)
4.33 (1.27) ***
3
0.84 (1.11)
4
9.87 (0.80) ***
2
FD(May)
6.47 (0.83) ***
4
2.13 (0.67) **
3
10.11 (0.41) ***
3
HD(May)
3.60 (1.29) **
-
0.32 (0.97)
-
4.26 (0.62) ***
8
PSD(Mar.-May)
0.02 (1.15)
7
0.93 (1.21)
8
-1.68 (0.51) **
6
FSD(May)
-4.78 (0.60) ***
10
-1.98 (0.64) **
10
-6.50 (0.38) ***
4
CDD0(Mar.-May)1/CDD5(Mar.-May)1, 以0 ℃/5 ℃为基温计算3月至5月份的热量积累(气温); CDD0(Mar.-May)2/CDD5(Mar.-May)2, 以0 ℃/5 ℃为基温计算3月至5月份的热量积累(20 cm处土壤温度); CH0(Oct.-Feb.)/CH5(Oct.-Feb.), 以0 ℃/5 ℃为基温计算上年10月至来年2月的低温积累; FD(May), 5月份最低气温小于0 ℃的天数; FSD(May), 5月份冻土深度; HD(May), 5月份最高气温大于15 ℃的天数; PSD(Mar.-May), 3-5月份连续5天无降水的次数; RD(Mar.-May), 3-5月份降水天数; TR(Mar.-May), 3-5月份总降水量。偶数列为岭回归分析的标度系数(Scaled coefficient); 奇数列(除第一列)为Lasso回归分析的解释变量Lar序列(Lar sequence)。Lar序列加粗数字为筛选的主要解释变量。 CDD0(Mar.-May)1/CDD5(Mar.-May)1, calculation of heat accumulation (air temperature) from March to May with 0 °C/5 °C as the base temperature; CDD0(Mar.- May)2/ CDD5(Mar.- May)2, calculation of heat accumulation (soil temperature at 20 cm depth) from March to May with 0 °C/5 °C as the base temperature; CH0(Oct.-Feb.)/CH5(Oct.-Feb.), the chilling accumulation from October last year to February next year was calculated with 0 °C/5 °C as the base temperature; FD(May), days of minimum temperature less than 0 °C in May; FSD(May), permafrost depth in May; HD(May), days with maximum temperature higher than 15 °C in May; PSD(Mar.-May), frequency of no precipitation for five consecutive days from March to May; RD(Mar.-May), precipitation days from March to May; TR(Mar.-May), total rainfall from March to May. The even columns represent the scaled coefficient of ridge regression analysis; odd columns (except the first column) represent the explanatory variable Lar sequence of lasso regression analysis. The bold font in Lar sequence represents the main explanatory variables.
AndersonJT, GezonZJ (2015). Plasticity in functional traits in the context of climate change: a case study of the subalpine Boechera stricta (Brassicaceae) Global Change Biology, 21, 1689-1703. [本文引用: 1]
BressonCC, VitasseY, KremerA, DelzonS (2011). To what extent is altitudinal variation of functional traits driven by genetic adaptation in European oak and beech? Tree Physiology, 31, 1164-1174. [本文引用: 5]
ChenXQ, LiJ (2009). Relationships between Leymus chinensis phenology and meteorological factors in Inner Mongolia grasslands Acta Ecologica Sinica, 29, 5280-5290. [本文引用: 1]
CooperHF, GradyKC, CowanJA, BestRJ, AllanGJ, WhithamTG (2019). Genotypic variation in phenological plasticity: reciprocal common gardens reveal adaptive responses to warmer springs but not to fall frost Global Change Biology, 25, 187-200. [本文引用: 1]
DantecCF, DucasseH, CapdevielleX, FabreguettesO, DelzonS, Desprez-LoustauML (2015). Escape of spring frost and disease through phenological variations in oak populations along elevation gradients Journal of Ecology, 103, 1044-1056. [本文引用: 1]
DantecCF, VitasseY, BonhommeM, LouvetJM, KremerA, DelzonS (2014). Chilling and heat requirements for leaf unfolding in European beech and sessile oak populations at the southern limit of their distribution range International Journal of Biometeorology, 58, 1853-1864. [本文引用: 2]
DuJ, HeZB, YangJJ, ChenLF, ZhuX (2014). Detecting the effects of climate change on canopy phenology in coniferous forests in semi-arid mountain regions of China International Journal of Remote Sensing, 35, 6490-6507. [本文引用: 2]
DuJ, LiK, HeZB, ChenLF, LinPF, ZhuX (2020). Daily minimum temperature and precipitation control on spring phenology in arid-mountain ecosystems in China International Journal of Climatology, 40, 2568-2579. [本文引用: 1]
FreiER, GhazoulJ, MatterP, HeggliM, PluessAR (2014). Plant population differentiation and climate change: responses of grassland species along an elevational gradient Global Change Biology, 20, 441-455. [本文引用: 4]
GeQS, DaiJH, ZhengJY (2010). The progress of phenology studies and challenges to modern phenology research in China Bulletin of Chinese Academy of Sciences, 25, 310-316. [本文引用: 1]
GuoXL, KliszM, PuchałkaR, SilvestroR, FaubertP, BelienE, HuangJG, RossiS (2021). Common-garden experiment reveals clinal trends of bud phenology in black spruce populations from a latitudinal gradient in the boreal forest Journal of Ecology. DOI: 10.1111/1365-2745.13582. [本文引用: 1]
HwangT, SongC, VoseJM, BandLE (2011). Topography- mediated controls on local vegetation phenology estimated from MODIS vegetation index Landscape Ecology, 26, 541-556. [本文引用: 1]
JiRX, YuX, ChangY, ShenC, BaiXQ, XiaXL, YinWL, LiuC (2020). Geographical provenance variation of leaf anatomical structure of Caryopteris mongholica and its significance in response to environmental changes Chinese Journal of Plant Ecology, 44, 277-286. [本文引用: 1]
KosugiY, TakanashiS, UeyamaM, OhkuboS, TanakaH, MatsumotoK, YoshifujiN, AtakaM, SakabeA (2013). Determination of the gas exchange phenology in an evergreen coniferous forest from 7 years of eddy covariance flux data using an extended big-leaf analysis Ecological Research, 28, 373-385. [本文引用: 1]
KörnerC, BaslerD (2010). Phenology under global warming Science, 327, 1461-1462. [本文引用: 1]
KrepkowskiJ, BräuningA, GebrekirstosA, StroblS (2011). Cambial growth dynamics and climatic control of different tree life forms in tropical mountain forest in Ethiopia Trees, 25, 59-70. [本文引用: 1]
Leiblein-WildMC, TackenbergO (2014). Phenotypic variation of 38 European Ambrosia artemisiifolia populations measured in a common garden experiment Biological Invasions, 16, 2003-2015. [本文引用: 1]
LinY, WestG (2016). Reflecting conifer phenology using mobile terrestrial LiDAR: a case study of Pinus sylvestris growing under the Mediterranean climate in Perth, Australia Ecological Indicators, 70, 1-9. [本文引用: 1]
LiuXD, ZhaoWJ, ZhangXL, JingWM, FanLM (2013). Variation of soil nutrient content and pH value under Picea crassifolia forest in the Pailugou drainage basin in the Qilian mountains Arid Zone Research, 30, 1013-1020. [本文引用: 1]
PengXM, DuJ, YangB, XiaoSC, LiG (2019). Elevation- influenced variation in canopy and stem phenology of Qinghai spruce, central Qilian Mountains, northeastern Tibetan Plateau Trees, 33, 707-717. [本文引用: 1]
PeñuelasJ, FilellaI (2001). Responses to a warming world Science, 294, 793-795. [本文引用: 1]
RichardsonAD, HollingerDY, DailDB, LeeJT, MungerJW, O’keefeJ (2009). Influence of spring phenology on seasonal and annual carbon balance in two contrasting New England forests Tree physiology, 29, 321-331. [本文引用: 1]
RichardsonAD, KeenanTF, MigliavaccaM, RyuY, SonnentagO, ToomeyM (2013). Climate change, phenology, and phenological control of vegetation feedbacks to the climate system Agricultural and Forest Meteorology, 169, 156-173. [本文引用: 3]
ShenMG, PiaoSL, CongN, ZhangGX, JassensIA (2015). Precipitation impacts on vegetation spring phenology on the Tibetan Plateau Global Change Biology, 21, 3647-3656. [本文引用: 1]
SpringateDA, KoverPX (2014). Plant responses to elevated temperatures: a field study on phenological sensitivity and fitness responses to simulated climate warming Global Change Biology, 20, 456-465. [本文引用: 1]
TianQY, HeZB, XiaoSC, PengXM, DingAJ, LinPF (2017). Response of stem radial growth of Qinghai spruce (Picea crassifolia) to environmental factors in the Qilian Mountains of China Dendrochronologia, 44, 76-83. [本文引用: 1]
VitasseY, DelzonS, BressonCC, MichaletR, KremerA (2009). Altitudinal differentiation in growth and phenology among populations of temperate-zone tree species growing in a common garden Canadian Journal of Forest Research, 39, 1259-1269. [本文引用: 5]
VitasseY, HochG, RandinCF, LenzA, KollasC, ScheepensJF, KörnerC (2013). Elevational adaptation and plasticity in seedling phenology of temperate deciduous tree species Oecologia, 171, 663-678. [本文引用: 1]
VitasseY, SignarbieuxC, FuYH (2018). Global warming leads to more uniform spring phenology across elevations Proceedings of the National Academy of Sciences of the United States of America, 115, 1004-1008. [本文引用: 2]
VolisS, OrmanbekovaD, YermekbayevK (2015). Role of phenotypic plasticity and population differentiation in adaptation to novel environmental conditions Ecology and Evolution, 5, 3818-3829. [本文引用: 3]
WangSP, WangCS, DuanJC, ZhuXX, XuGP, LuoCY, ZhangZH, MengFD, LiYN, DuMY (2014). Timing and duration of phenological sequences of alpine plants along an elevation gradient on the Tibetan Plateau Agricultural and Forest Meteorology, 189- 190, 220-228. [本文引用: 1]
WareIM, van NulandME, SchweitzerJA, YangZ, SchadtCW, Sidak-LoftisLC, StoneNE, BuschJD, BaileyJK (2019). Climate-driven reduction of genetic variation in plant phenology alters soil communities and nutrient pools Global Change Biology, 25, 1514-1528. [本文引用: 1]
WilczekAM, BurghardtLT, CobbAR, CooperMD, WelchSM, SchmittJ (2010). Genetic and physiological bases for phenological responses to current and predicted climates Philosophical Transactions of the Royal Society of London B, Biological Sciences, 365, 3129-3147. [本文引用: 1]
YuZ, SunPS, LiuSR (2010). Phenological change of main vegetation types along a North-South Transect of Eastern China Chinese Journal of Plant Ecology, 34, 316-329. [本文引用: 1]
ZandlerH, BrenningA, SamimiC (2015). Quantifying dwarf shrub biomass in an arid environment: comparing empirical methods in a high dimensional setting Remote Sensing of Environment, 158, 140-155. [本文引用: 1]
ZhengJY, LiuY, GeQS, HaoZX (2015). Spring phenodate records derived from historical documents and reconstruction on temperature change in Central China during 1850-2008 Acta Geographica Sinica, 70, 696-704. [本文引用: 1]
... 近期的研究发现, 种群性状表达的空间异质性既体现了环境条件改变引起的表型可塑性, 同时也可能包含了种群在局部生境的自然选择作用下表现出的遗传分化现象(Bresson et al., 2011; Frei et al., 2014).量化植物性状空间分异的遗传表达与表型可塑性, 对于合理准确预测气候变化对植物动态的影响具有重要实践意义.本研究发现青海云杉营养生长的物候期具有显著的种源效应, 且其影响在年季之际相对稳定, 维持在20%左右的水平, 该结论与最近关于Picea mariana萌芽物候遗传分化的认识(Guo et al., 2021)基本一致.尽管由于移植样树尚未达到开花年龄, 未能监测到繁殖物候事件, 但前期的物候学研究表明开花时间同样具有很高的遗传力(Leiblein-Wild & Tackenberg, 2014), 因此, 异质生境中的群体分化可能是一个普遍存在的现象.另外, 我们发现同质园中移植种群所反映的海拔依赖性与原生境中自然种群的海拔梯度呈现相反的格局, 这是有别于其他多数物种研究(Bresson et al., 2011; Volis et al., 2015)的地方.事实上, Vitasse等(2009)在对Fagus sylvatica种群萌芽和开花物候研究时, 同样发现同质园内来自高海拔的种群物候期早于低海拔种群的现象.这可能与不同海拔种源的物候活动具有差异的热量需求有关(Dantec et al., 2014).相对于低海拔种群, 高海拔个体的芽分化往往需要较低的季前热量积累(图3); 当移植入低海拔的同质园后, 在冬季低温持续时间变化不大的情况下(图3), 较高的热量补偿可能加速了高海拔个体的物候响应.同时, 春季物候的提前还可能体现了具有短生长季的高海拔种群对生长周期延长的适应结果(Vitasse et al., 2009). ...
... ), 因此, 异质生境中的群体分化可能是一个普遍存在的现象.另外, 我们发现同质园中移植种群所反映的海拔依赖性与原生境中自然种群的海拔梯度呈现相反的格局, 这是有别于其他多数物种研究(Bresson et al., 2011; Volis et al., 2015)的地方.事实上, Vitasse等(2009)在对Fagus sylvatica种群萌芽和开花物候研究时, 同样发现同质园内来自高海拔的种群物候期早于低海拔种群的现象.这可能与不同海拔种源的物候活动具有差异的热量需求有关(Dantec et al., 2014).相对于低海拔种群, 高海拔个体的芽分化往往需要较低的季前热量积累(图3); 当移植入低海拔的同质园后, 在冬季低温持续时间变化不大的情况下(图3), 较高的热量补偿可能加速了高海拔个体的物候响应.同时, 春季物候的提前还可能体现了具有短生长季的高海拔种群对生长周期延长的适应结果(Vitasse et al., 2009). ...
内蒙古草原羊草物候与气象因子的关系 1 2009
... 植物物候作为生态环境变化的综合指示器, 一直被视为研究植物生长发育与气候关系的重要指标 (葛全胜等, 2010; Körner & Basler, 2010).在过去的30年内, 随着全球变化研究的不断深入, 人们普遍意识到植物物候在揭示气候变化对陆地生态系统长期影响中的重要地位(Richardson et al., 2013).国内外****就此针对各脆弱生态系统, 尤其是在干旱区山地, 围绕植物物候对气候变化的响应与反馈开展了大量研究工作, 试图了解气候变化对陆地生态系统的影响过程及程度(Dunne et al., 2003; 陈效逑和李倞, 2009; Wilczek et al., 2010; 郑景云等, 2015). ...
内蒙古草原羊草物候与气象因子的关系 1 2009
... 植物物候作为生态环境变化的综合指示器, 一直被视为研究植物生长发育与气候关系的重要指标 (葛全胜等, 2010; Körner & Basler, 2010).在过去的30年内, 随着全球变化研究的不断深入, 人们普遍意识到植物物候在揭示气候变化对陆地生态系统长期影响中的重要地位(Richardson et al., 2013).国内外****就此针对各脆弱生态系统, 尤其是在干旱区山地, 围绕植物物候对气候变化的响应与反馈开展了大量研究工作, 试图了解气候变化对陆地生态系统的影响过程及程度(Dunne et al., 2003; 陈效逑和李倞, 2009; Wilczek et al., 2010; 郑景云等, 2015). ...
Genotypic variation in phenological plasticity: reciprocal common gardens reveal adaptive responses to warmer springs but not to fall frost 1 2019
... 植物物候学研究存在明显的尺度效应, 包括对物候过程调控机理的认识, 例如区域尺度的物候模型往往不能准确预测单个物种的物候动态(Richardson et al, 2013).研究经验显示, 种群才是合理认知生物学规律的基本单元, 对于种群水平物候调控机理的解析能够有效提升物种环境响应的预测能力(Frei et al., 2014).本研究明确了不同海拔间青海云杉种群的物候过程具有一定的遗传分化现象, 但由于实验的局限性(仅在一个海拔设置了同质园实验), 无法定量表型可塑性的成分比例.尽管如此, 鉴于遗传分化和表型可塑性是植物在异质生境中的主要适应方式(Volis et al., 2015), 在遗传分化贡献率较低的情况下, 因环境改变而诱发的表型可塑性必然成为主导的环境适应机制.这种推论已在之前众多的相关研究(Vitasse et al., 2013; Frei et al., 2014; Cooper et al., 2019)中得以证实.因此, 本研究进一步对物候可塑性的环境调控作用进行了分析.研究发现, 对于多数表观物候阶段(包括营养生长与生殖生长)来说, 海拔起到了关键的指示作用, 主要与其影响局地尺度上的能量平衡和温度格局有关.与表观物候类似, 海拔同样能够引起径向生长周期的种群分异(Peng et al., 2019).作为反映温度调控的两个主要影响方式, 季前热量积累(CDD0/CDD5)与冬季低温的春化作用(CH0/CH5)均与青海云杉物候期呈现显著的负相关关系, 表明低温春化与热量积累的增加均有利于激发物候事件的提前发生.随着海拔的降低, 我们发现除了春季热量积累呈现显著的增加趋势外, 冬季有效低温的累积同样有所增加(图3), 与Vitasse等(2018)研究结果一致, 由此反映出温度的两种影响方式在塑造物候的海拔格局上发挥着协同的调控作用.在水分调节方面, 降水量及降水天数的增加并没有表现出对春季物候的促进作用, 而是在一定程度上推迟了展叶与开花的时间, 这一结果与区域尺度得到的认识(Shen et al., 2015; Du et al., 2020)相反, 原因可能是降水事件的发生往往伴随着气温的降低, 降水天数的增加提高了对季前热量需求的阈值, 由此导致种群物候期的延迟.除此之外, 晚春冻害与冻土融化深度也显著影响着初期阶段的物候进程: 气温升高后的倒春寒使得新萌发的枝叶易遭受冻害的风险, 由此可能导致新萌芽的死亡, 造成相应物候阶段的推迟; 冻土融化的深度意味着植株个体初生生长所能利用土壤水分的可得性, 冻土融化越深, 土壤水分的补给越多, 越有利于机体生理活动的有序进行. ...
Escape of spring frost and disease through phenological variations in oak populations along elevation gradients 1 2015
... 在山地系统中, 由于地形、气候、土壤等自然要素组合的特殊性, 在较小的空间尺度上可能会发生环境条件的剧烈改变(尤其气温的变化), 从而造成植物物候出现不同的表型特征(如早展叶与晚展叶、早开花与晚开花等, Dantec et al., 2015).目前关于物候表型分异的研究多集中在物候现象易于观测的落叶物种(包括乔木、灌木), 对常绿针叶树种没有足够的重视(Bresson et al., 2011; Springate & Kover, 2014; Anderson & Gezon, 2015).然而, 干旱区山地的一些常绿针叶树种, 如分布于我国祁连山地区的典型优势种——青海云杉(Picea crassifolia), 在维持区域碳的收支平衡、调节地表径流、涵养水源等方面发挥着重要的生态功能(刘贤德等, 2013).现阶段对常绿针叶树种的物候研究, 主要集中于区域尺度的遥感分析(余振等, 2010; Du et al., 2014)、群落尺度的通量反演(Richardson et al., 2009; Kosugi et al., 2013)、个体尺度的径向生长监测(Krepkowski et al., 2011; Tian et al., 2017)等方面, 尚缺乏对种群水平物候调控机理的认识.根据祁连山长期的物候监测数据, 青海云杉种群物候在海拔梯度上存在着较强的空间异质性, 这种空间分异是否存在及在多大程度上体现了种群间的遗传表达, 环境因子又如何调控非遗传分化部分(即表型可塑性)的物候响应, 都是种群水平物候研究亟待解决的科学问题. ...
Chilling and heat requirements for leaf unfolding in European beech and sessile oak populations at the southern limit of their distribution range 2 2014
Topography- mediated controls on local vegetation phenology estimated from MODIS vegetation index 1 2011
... 在山地生态系统中, 植物物候的空间分异与地形要素具有密切的联系, 其中, 海拔变量对物候格局的解释力已被众多典型案例所证实(Hwang et al., 2011).正是由于海拔效应的存在, Hopkins早在1918年就提出了适用于北美洲温带地区的“生物气候定律” (在其他条件不变的情况下, 海拔上升122 m, 植物春季物候期将推迟4天, 而秋季物候期则呈现相反的格局)(龚高法和简慰民, 1983).但对这一定律的认识正在被近年来的气候变暖效应所改变, 研究发现气候变暖导致不同海拔间春季物候期逐渐趋同(Vitasse et al., 2018).除了海拔影响外, 坡向也在一定程度上改变着物种的物候格局, 本研究中处于半阴坡的青海云杉林比阴坡具有更好的光照条件, 较多的季前热量积累有利于叶芽的萌发和花芽的开放.气象监测数据显示4-8月内半阴坡的平均气温比阴坡高0.22 ℃, 而同期相对湿度却无显著变化(-0.27%).相对坡向的影响, 具有不同分布面积的 林斑之间则没有表现出显著的差异.尽管林斑大小可能代表了不同的景观破碎化程度及生境质量(如土壤有机质含量), 但该结果表明由林斑大小引起的微环境变化不足以诱发青海云杉物候表型的明显改变. ...
蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义 1 2020
... 近年来, 同质园实验已被国内外****的研究证实为区分环境调控作用与遗传分化效应的一种有效手段(Vitasse et al., 2009; Ware et al., 2019; 纪若璇等, 2020).同质园的基本思想是将多个种源的同种个体在相同环境下栽培, 目的是探究其初生与次生代谢的响应.本研究中, 实验园位于海拔2 750 m处, 紧邻祁连山森林生态系统观测站, 周边建有综合环境监测系统, 可为实验园提供背景资料.选择一个5 m × 10 m的典型样地, 并在其周边架起围栏.按照遗传生态学原理, 通常情况下, 同质园实验的对象要求是植物的子代, 才有利于反映种群间的遗传分化现象; 但由于青海云杉种子的休眠时间较长, 并且幼苗生长缓慢, 因此, 本研究采用国外类似研究中对幼树移植的方式来代替种子移种(Frei et al., 2014), 不仅提高了幼树的成活率, 而且减少了生长过程所需的时间.实验最为关键的环节是保证同质园内土壤环境的一致性.本研究在实践中的做法如下: 在每个调查样地内随机选择幼树3-5株(基径1.5 cm)(5个海拔× 4个调查样地× 3-5株/调查样地), 将其连根带适量(少量)原土刨出(相对于净土移植, 这样做能提高移植过程中个体的存活率), 放入事先准备好的铁箱中运移至移栽地, 然后将其所携带的土壤去除, 种植于混合土壤(将所有海拔移植个体所携带的部分土壤充分混合在一起)中, 这样保证了所有海拔移植个体土壤环境的一致性.对所有个体进行编号标定.移植实验于2017年9月完成, 并于2018年开始进行物候监测. ...
蒙古莸叶片解剖结构的地理种源变异及其对环境变化响应的意义 1 2020
... 近年来, 同质园实验已被国内外****的研究证实为区分环境调控作用与遗传分化效应的一种有效手段(Vitasse et al., 2009; Ware et al., 2019; 纪若璇等, 2020).同质园的基本思想是将多个种源的同种个体在相同环境下栽培, 目的是探究其初生与次生代谢的响应.本研究中, 实验园位于海拔2 750 m处, 紧邻祁连山森林生态系统观测站, 周边建有综合环境监测系统, 可为实验园提供背景资料.选择一个5 m × 10 m的典型样地, 并在其周边架起围栏.按照遗传生态学原理, 通常情况下, 同质园实验的对象要求是植物的子代, 才有利于反映种群间的遗传分化现象; 但由于青海云杉种子的休眠时间较长, 并且幼苗生长缓慢, 因此, 本研究采用国外类似研究中对幼树移植的方式来代替种子移种(Frei et al., 2014), 不仅提高了幼树的成活率, 而且减少了生长过程所需的时间.实验最为关键的环节是保证同质园内土壤环境的一致性.本研究在实践中的做法如下: 在每个调查样地内随机选择幼树3-5株(基径1.5 cm)(5个海拔× 4个调查样地× 3-5株/调查样地), 将其连根带适量(少量)原土刨出(相对于净土移植, 这样做能提高移植过程中个体的存活率), 放入事先准备好的铁箱中运移至移栽地, 然后将其所携带的土壤去除, 种植于混合土壤(将所有海拔移植个体所携带的部分土壤充分混合在一起)中, 这样保证了所有海拔移植个体土壤环境的一致性.对所有个体进行编号标定.移植实验于2017年9月完成, 并于2018年开始进行物候监测. ...
Determination of the gas exchange phenology in an evergreen coniferous forest from 7 years of eddy covariance flux data using an extended big-leaf analysis 1 2013
... 在山地系统中, 由于地形、气候、土壤等自然要素组合的特殊性, 在较小的空间尺度上可能会发生环境条件的剧烈改变(尤其气温的变化), 从而造成植物物候出现不同的表型特征(如早展叶与晚展叶、早开花与晚开花等, Dantec et al., 2015).目前关于物候表型分异的研究多集中在物候现象易于观测的落叶物种(包括乔木、灌木), 对常绿针叶树种没有足够的重视(Bresson et al., 2011; Springate & Kover, 2014; Anderson & Gezon, 2015).然而, 干旱区山地的一些常绿针叶树种, 如分布于我国祁连山地区的典型优势种——青海云杉(Picea crassifolia), 在维持区域碳的收支平衡、调节地表径流、涵养水源等方面发挥着重要的生态功能(刘贤德等, 2013).现阶段对常绿针叶树种的物候研究, 主要集中于区域尺度的遥感分析(余振等, 2010; Du et al., 2014)、群落尺度的通量反演(Richardson et al., 2009; Kosugi et al., 2013)、个体尺度的径向生长监测(Krepkowski et al., 2011; Tian et al., 2017)等方面, 尚缺乏对种群水平物候调控机理的认识.根据祁连山长期的物候监测数据, 青海云杉种群物候在海拔梯度上存在着较强的空间异质性, 这种空间分异是否存在及在多大程度上体现了种群间的遗传表达, 环境因子又如何调控非遗传分化部分(即表型可塑性)的物候响应, 都是种群水平物候研究亟待解决的科学问题. ...
Phenology under global warming 1 2010
... 植物物候作为生态环境变化的综合指示器, 一直被视为研究植物生长发育与气候关系的重要指标 (葛全胜等, 2010; Körner & Basler, 2010).在过去的30年内, 随着全球变化研究的不断深入, 人们普遍意识到植物物候在揭示气候变化对陆地生态系统长期影响中的重要地位(Richardson et al., 2013).国内外****就此针对各脆弱生态系统, 尤其是在干旱区山地, 围绕植物物候对气候变化的响应与反馈开展了大量研究工作, 试图了解气候变化对陆地生态系统的影响过程及程度(Dunne et al., 2003; 陈效逑和李倞, 2009; Wilczek et al., 2010; 郑景云等, 2015). ...
Cambial growth dynamics and climatic control of different tree life forms in tropical mountain forest in Ethiopia 1 2011
... 在山地系统中, 由于地形、气候、土壤等自然要素组合的特殊性, 在较小的空间尺度上可能会发生环境条件的剧烈改变(尤其气温的变化), 从而造成植物物候出现不同的表型特征(如早展叶与晚展叶、早开花与晚开花等, Dantec et al., 2015).目前关于物候表型分异的研究多集中在物候现象易于观测的落叶物种(包括乔木、灌木), 对常绿针叶树种没有足够的重视(Bresson et al., 2011; Springate & Kover, 2014; Anderson & Gezon, 2015).然而, 干旱区山地的一些常绿针叶树种, 如分布于我国祁连山地区的典型优势种——青海云杉(Picea crassifolia), 在维持区域碳的收支平衡、调节地表径流、涵养水源等方面发挥着重要的生态功能(刘贤德等, 2013).现阶段对常绿针叶树种的物候研究, 主要集中于区域尺度的遥感分析(余振等, 2010; Du et al., 2014)、群落尺度的通量反演(Richardson et al., 2009; Kosugi et al., 2013)、个体尺度的径向生长监测(Krepkowski et al., 2011; Tian et al., 2017)等方面, 尚缺乏对种群水平物候调控机理的认识.根据祁连山长期的物候监测数据, 青海云杉种群物候在海拔梯度上存在着较强的空间异质性, 这种空间分异是否存在及在多大程度上体现了种群间的遗传表达, 环境因子又如何调控非遗传分化部分(即表型可塑性)的物候响应, 都是种群水平物候研究亟待解决的科学问题. ...
Phenotypic variation of 38 European Ambrosia artemisiifolia populations measured in a common garden experiment 1 2014
... 近期的研究发现, 种群性状表达的空间异质性既体现了环境条件改变引起的表型可塑性, 同时也可能包含了种群在局部生境的自然选择作用下表现出的遗传分化现象(Bresson et al., 2011; Frei et al., 2014).量化植物性状空间分异的遗传表达与表型可塑性, 对于合理准确预测气候变化对植物动态的影响具有重要实践意义.本研究发现青海云杉营养生长的物候期具有显著的种源效应, 且其影响在年季之际相对稳定, 维持在20%左右的水平, 该结论与最近关于Picea mariana萌芽物候遗传分化的认识(Guo et al., 2021)基本一致.尽管由于移植样树尚未达到开花年龄, 未能监测到繁殖物候事件, 但前期的物候学研究表明开花时间同样具有很高的遗传力(Leiblein-Wild & Tackenberg, 2014), 因此, 异质生境中的群体分化可能是一个普遍存在的现象.另外, 我们发现同质园中移植种群所反映的海拔依赖性与原生境中自然种群的海拔梯度呈现相反的格局, 这是有别于其他多数物种研究(Bresson et al., 2011; Volis et al., 2015)的地方.事实上, Vitasse等(2009)在对Fagus sylvatica种群萌芽和开花物候研究时, 同样发现同质园内来自高海拔的种群物候期早于低海拔种群的现象.这可能与不同海拔种源的物候活动具有差异的热量需求有关(Dantec et al., 2014).相对于低海拔种群, 高海拔个体的芽分化往往需要较低的季前热量积累(图3); 当移植入低海拔的同质园后, 在冬季低温持续时间变化不大的情况下(图3), 较高的热量补偿可能加速了高海拔个体的物候响应.同时, 春季物候的提前还可能体现了具有短生长季的高海拔种群对生长周期延长的适应结果(Vitasse et al., 2009). ...
Reflecting conifer phenology using mobile terrestrial LiDAR: a case study of Pinus sylvestris growing under the Mediterranean climate in Perth, Australia 1 2016
 ,1,*, 陈龙飞1, 蔺鹏飞1, 朱喜1, 田全彦1
,1,*, 陈龙飞1, 蔺鹏飞1, 朱喜1, 田全彦1

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT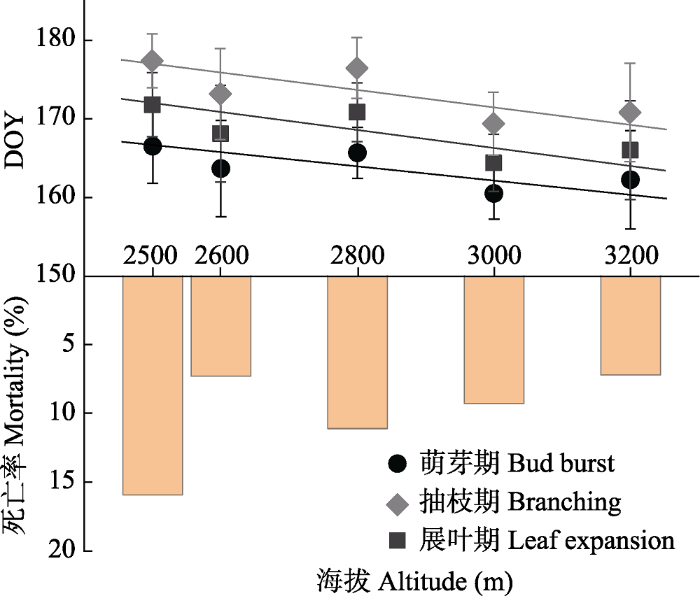 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT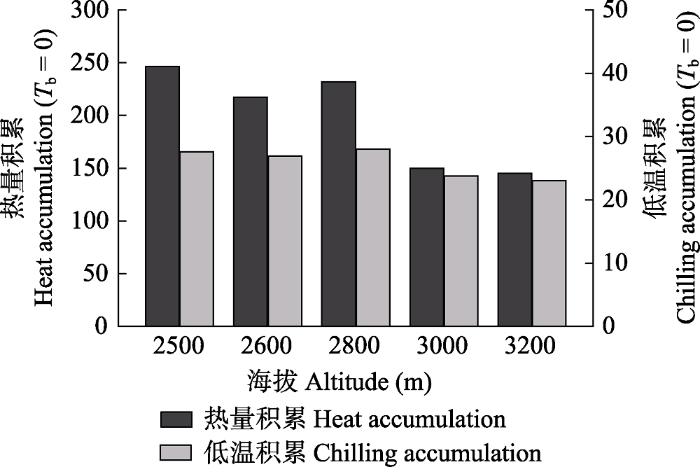 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}