Decomposition characteristics of leaf litters and roots of six main plant species and their relationships with functional traits in Stipa grandis steppe
Wei-Na ZHU1,3, Guo-Long ZHANG1,3, Pu-Jin ZHANG2, Qian-Qian ZHANG1,3, Jin-Tao REN1,3, Bu-Yun XU1,3, Hua QING,1,3,*1Inner Mongolia Key Laboratory of Grassland Ecology, School of Ecology and Environment, Inner Mongolia University, Hohhot 010020, China 2Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Hohhot 010031, China 3Ministry of Education Key Laboratory of Ecology and Resource Use of the Mongolian Plateau, Inner Mongolia University, Hohhot 010020, China
Natural Science Foundation of Nei Mongol of China(2018MS03072) National Natural Science Foundation of China(31960246) National Natural Science Foundation of China(31560174) Innovative Foundation of Nei Mongol Agricultural and Animal Husbandry Sciences(2020CXJJM11)
Abstract Aims It is very important to investigate the relationships between litter decomposition characteristics and plant functional traits in understanding the maintenance mechanism of ecosystem functions. Methods In order to study the main driving factors that affect the leaf litters and root decomposition of different species, this study took the leaf litters and roots of six main plant species Stipa grandis, Cleistogenes squarrosa, Anemarrhena asphodeloides, Leymus chinensis, Convolvulus ammannii and Carex korshinskyi in S. grandis steppe. The litter bag method was used to study the decomposition rate constant of both leaf litters and root through 501 days of field incubation. Plant functional traits including leaf dry matter content, root specific surface area, root tissue density, contents of C, N and different cellulose components of the leaf and root litters were determined and the relationships between decomposition characteristics and functional traits of leaf litters and root across six plant species were examined. Important findingsThe results showed that there were significant interspecific differences in leaf and root traits of six plant species. The ratios of maximum to minimum values for most traits were between 1 and 2, while the difference in some traits, such as C:N and specific surface area of roots between species was nearly 4 times. For the six plant species, the overall trend of the mass residue and decomposition rate constant of the leaf litter and root during 501 days of decomposition all showed the rapid decomposition in the early stage, relatively slow decomposition in the middle stage and the slowest decomposition in the later stage. During the decomposition process of leaf litters and roots, Cleistogenes squarrosa showed the slowest one, while the leaf litter decomposition of Anemarrhena asphodeloides was the fastest, and the root decomposition of Convolvulus ammannii was the fastest. Through the correlation analysis and stepwise regression analysis, it was found that the decomposition process of leaf litters and roots was affected by different traits in different decomposition periods. The structural carbohydrate content was the main factor affecting the early and late decomposition of litters and the early decomposition of roots, while the non-structural carbohydrate content was the main factor affecting the middle and late decomposition of roots. In addition, the decomposition rate of leaf litters in the middle stage of decomposition was mainly affected by leaf dry matter content, while the decomposition rates of roots in the middle and late stages of decomposition were also significantly affected by C:N and N content, respectively. Our results present the important guide for the prediction of carbon and nutrient cycling process in the S. grandis steppe. Keywords:Stipa grandis steppe;litter;plant functional trait;decomposition rate
PDF (1772KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 朱蔚娜, 张国龙, 张璞进, 张迁迁, 任瑾涛, 徐步云, 清华. 大针茅草原6种主要植物叶凋落物和根系分解特征与功能性状的关系. 植物生态学报, 2021, 45(6): 606-616. DOI: 10.17521/cjpe.2020.0268 ZHU Wei-Na, ZHANG Guo-Long, ZHANG Pu-Jin, ZHANG Qian-Qian, REN Jin-Tao, XU Bu-Yun, QING Hua. Decomposition characteristics of leaf litters and roots of six main plant species and their relationships with functional traits in Stipa grandis steppe. Chinese Journal of Plant Ecology, 2021, 45(6): 606-616. DOI: 10.17521/cjpe.2020.0268
6种植物的叶凋落物与根系经研磨过40目筛后, 分别称取0.50 g放入滤袋封口, 采用van Soest (1963)的提取方法, 使用ANKOM纤维分析仪(A2000I, ANKOM, New York, USA)进行非结构性碳水化合物含量和结构性碳水化合物(半纤维素、纤维素及木质素)含量的测定。凋落物的C、N含量采用CHNS元素分析仪(Elementar Vario EL III, Elementar, Hanau, Germany)进行测定。
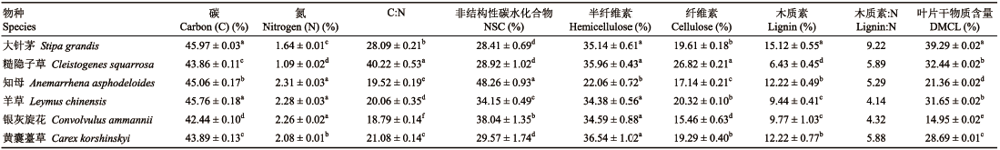
Table 1 表1 表1内蒙古大针茅草原6种植物叶凋落物的功能性状及叶片干物质含量(平均值2标准误) Table 1Functional traits and dry matter content of leaves of leaf ltter of six plant species in Stipa grandis steppe in Nei Mongol (mean2SE)
不同小写字母代表同列中差异显著(p < 0.05)。 Different lowercase ltters represent signifcant difrences in the same column (p < 0.05). DMCL, dry matter content of lcaves; NSC, non-stuctural carbobydrate.
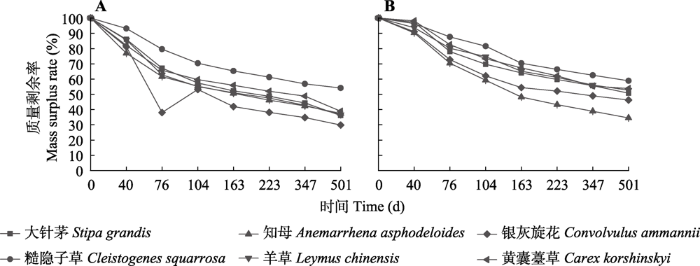
新窗口打开|下载原图ZIP|生成PPT 图1内蒙古大针茅草原6种植物叶凋落物和根分解过程中的质量剩余率变化(平均值±标准误, n = 3)。A, 叶凋落物分解。B, 根系分解。由于银灰旋花叶片较小且易碎, 第76天处理时出现较大异常。
Fig. 1Variation of mass residual rate during the decomposition process of leaf litters and roots of six plant species in Stipa grandis steppe in Nei Mongol (mean ± SE, n = 3). A, Decomposition of leaf litters. B, Decomposition of roots. Because Convolvulus ammannii leaves were small and fragile, and a big abnormality appeared at 76 daysʼ treatment.
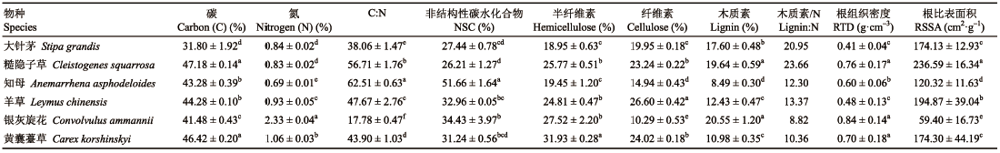
Table 3 表3 表3内蒙古大针茅草 原6种植物叶凋落物与根在不同分解时期的拟合分解速率常数(平均值±标准误) Table 3Functional traits of roots of six plant species in Stipa grandis steppe in Nei Mongol (mean±SE)
k,分解速率常数;R2,相关系数。 k decomposition rate constant; R2, correlation cofficient.
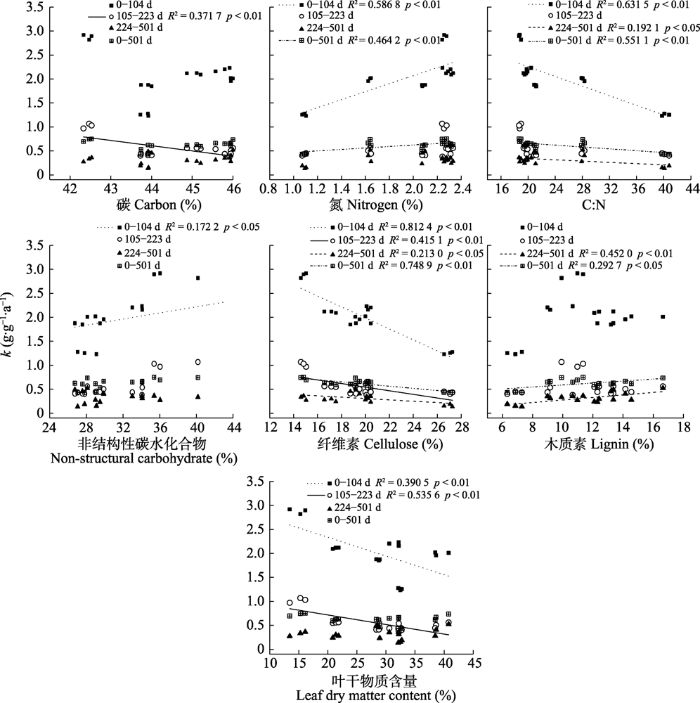
Fig. 2Relationship between functional traits of leaf litter and decomposition rate constant (k) of six plant species in Stipa grandis steppe in Nei Mongol. Functional traits that were not correlated with k for any of the four periods were not plotted (p > 0.05).
Fig. 3Relationship between functional traits of litters of plant roots and decomposition rate constant (k) of six plant species in Stipa grandis steppe in Nei Mongol. Functional traits that were not correlated with k of the four periods were not plotted (p > 0.05).
Table 4 表4 表4内蒙古大针茅草原6种植物叶凋落物和根系多元逐步回归模型系数及显著性 Table 4Multiple stepwise regression model coefficients and significance of leaf and root litters of six plant species in Stipa grandis steppe in Nei Mongol
分解时间段 Decomposition period (d)
凋落物类型 Litter type
模型 Model
r
调整后的R2 Adjusted R2
F
显著性 Significance
赤池信息 准则值 AICc
0-104
叶 Leaf
kL = 4.336 - 0.116X6
0.907
0.812
74.639
<0.001
1.213 0
kL = 5.448 - 0.140X6 - 0.057X7
0.945
0.878
62.069
<0.001
-4.300 6
kL = 4.968 - 0.149X6 - 0.055X7 + 0.19X5
0.962
0.910
58.202
<0.001
-7.094 9
根 Root
kR = 2.411 - 0.055X6
0.826
0.663
34.410
<0.001
4.521 8
kR = 3.118 - 0.06X6 - 0.041X7
0.946
0.881
64.217
<0.001
6.992 0
kR = 3.937 - 0.056X6 - 0.046X7 - 0.019X1
0.976
0.942
92.731
<0.001
-22.226 0
105-223
叶 Leaf
kL = 1.131 - 0.020X9
0.750
0.536
20.607
<0.001
-11.592 8
根 Root
kR = 0.131 + 0.014X4
0.767
0.563
22.863
<0.001
-19.749 6
kR = -0.069 + 0.012X4 + 0.006X3
0.905
0.795
34.016
<0.001
-31.214 4
224-501
叶 Leaf
kL = 0.15 + 0.027X7
0.695
0.451
14.985
0.001
-34.327 3
kL = -0.272 + 0.03X7 + 0.008X6
0.777
0.551
11.440
0.001
-31.549 6
根 Root
kR = 0.071 + 0.04X4
0.679
0.427
13.685
0.002
-58.761 2
kR = 0.120 + 0.004X4 - 0.047X2
0.846
0.678
18.867
<0.001
-23.870 1
kR = 0.406 + 0.001X4 - 0.106X2 - 0.001X10
0.899
0.768
19.751
<0.001
-70.138 3
0-501
叶 Leaf
kL = -1.059 - 0.023X6
0.874
0.749
51.631
<0.001
-51.201 6
kL = -1.050 - 0.029X6 + 0.005X9
0.924
0.835
43.920
<0.001
-56.544 5
kL = -1.407 - 0.044X6 + 0.01X9 - 0.019X7
0.960
0.904
54.266
<0.001
-63.616 2
根 Root
kR = 0.119 + 0.012X4
0.944
0.885
131.926
<0.001
-58.386 3
kR = 0.212 + 0.012X4 - 0.137X11
0.960
0.911
88.248
<0.001
-60.832 1
kL, 叶凋落物分解速率常数; kR, 根系分解速率常数。X1, 碳(C)含量(%); X2, 氮(N)含量(%); X3, C:N; X4, 非结构性碳水化合物含量(%); X5, 半纤维素含量(%); X6, 纤维素含量(%); X7, 木质素含量(%); X8, 木质素:N; X9, 叶干物质含量(%); X10, 根比表面积(cm2·g-1); X11, 根组织密度(g·cm-3)。 kL, decomposition rate constant of leaf litter; kR, decomposition rate constant of roots. X1, the content of carbon (C)(%); X2, the content of nitrogen (N)(%); X3, C:N; X4, the content of non-structural carbohydrate (%); X5, the content of hemicellulose (%); X6, the content of cellulose (%); X7, the content of lignin (%); X8, lignin:N; X9, the content of dry matter in leaves (%); X10, the specific surface area of roots (cm2·g-1); X11, the density of root tissues (g·cm-3). AICc, corrected Akaike’s Information Criterion.
BalasubramanianD, ArunachalamK, DasAK, ArunachalamA (2012). Decomposition and nutrient release of Eichhornia crassipes (Mart.) Solms. under different trophic conditions in wetlands of eastern Himalayan foothills Ecological Engineering, 44, 111-122. DOI:10.1016/j.ecoleng.2012.03.002URL [本文引用: 1]
BardgettRD, MommerL, de VriesFT (2014). Going underground: root traits as drivers of ecosystem processes Trends in Ecology & Evolution, 29, 692-699. DOI:10.1016/j.tree.2014.10.006URL [本文引用: 1]
BardgettRD, ShineA (1999). Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands Soil Biology & Biochemistry, 31, 317-321. DOI:10.1016/S0038-0717(98)00121-7URL [本文引用: 1]
CebrianJ (1999). Patterns in the fate of production in plant communities The American Naturalist, 154, 449-468. DOI:10.1086/303244PMID:10523491 [本文引用: 1] I examine, through an extensive compilation of published reports, the nature and variability of carbon flow (i.e., primary production, herbivory, detrital production, decomposition, export, and biomass and detrital storage) in a range of aquatic and terrestrial plant communities. Communities composed of more nutritional plants (i.e., higher nutrient concentrations) lose higher percentages of production to herbivores, channel lower percentages as detritus, experience faster decomposition rates, and, as a result, store smaller carbon pools. These results suggest plant palatability as a main limiting factor of consumer metabolical and feeding rates across communities. Hence, across communities, plant nutritional quality may be regarded as a descriptor of the importance of herbivore control on plant biomass ("top-down" control), the rapidity of nutrient and energy recycling, and the magnitude of carbon storage. These results contribute to an understanding of how much and why the trophic routes of carbon flow, and their ecological implications, vary across plant communities. They also offer a basis to predict the effects of widespread enhancement of plant nutritional quality due to large-scale anthropogenic eutrophication on carbon balances in ecosystems.
ClemmensenKE, BahrA, OvaskainenO, DahlbergA, EkbladA, WallanderH, StenlidJ, FinlayRD, WardleDA, LindahlBD (2013). Roots and associated fungi drive long-term carbon sequestration in boreal forest Science, 339, 1615-1618. DOI:10.1126/science.1231923PMID:23539604 [本文引用: 1] Boreal forest soils function as a terrestrial net sink in the global carbon cycle. The prevailing dogma has focused on aboveground plant litter as a principal source of soil organic matter. Using (14)C bomb-carbon modeling, we show that 50 to 70% of stored carbon in a chronosequence of boreal forested islands derives from roots and root-associated microorganisms. Fungal biomarkers indicate impaired degradation and preservation of fungal residues in late successional forests. Furthermore, 454 pyrosequencing of molecular barcodes, in conjunction with stable isotope analyses, highlights root-associated fungi as important regulators of ecosystem carbon dynamics. Our results suggest an alternative mechanism for the accumulation of organic matter in boreal forests during succession in the long-term absence of disturbance.
CornelissenJHC, Castro-DíezP, CarnelliAL (1998). Variation in relative growth rate among woody species//Lambers H, Poorter H, van Vuuren MMI. Inherent Variation in Plant Growth: Physiological Mechanisms and Ecological Consequences Backhuys Publishers, Leiden, the Netherlands.363-392. [本文引用: 1]
CornelissenJHC, ThompsonK (1997). Functional leaf attributes predict litter decomposition rate in herbaceous plants New Phytologist, 135, 109-114. DOI:10.1046/j.1469-8137.1997.00628.xPMID:33863150 [本文引用: 1] We tested the hypothesis that functional attributes of living leaves provide a basis for predicting the decomposition rate of leaf litter. The data were obtained from standardized screening tests on 38 British herbaceous species. Graminoid monocots had physically tougher leaves with higher silicon contents than did herbaceous dicots, and this corresponded with the lesser decomposibility of the former. Total base content of living leaves was a good predictor of litter decomposition rate, but the evolutionary and ecological basis for this relationship appeared to differ between graminoid monocots and herbaceous dicots. In the monocots, litter decomposition rate was strongly predicted by leaf potassium content, which appeared to reflect other growth- related plant attributes such as seedling maximum relative growth rate, foliar nitrogen and phosphorus content, specific leaf area and short leaf lifespan. In the dicots the relationship between total leaf base content and litter decomposition rate was not unambiguously explained by growth-related leaf attributes, possibly because of the considerable calcium uptake by dicots, which varies according to calcium availability in the soil.
CornwellWK, CornelissenJHC, AmatangeloK, DorrepaalE, EvinerVT, GodoyO, HobbieSE, HoorensB, KurokawaH, Pérez-HarguindeguyN, QuestedHM, SantiagoLS, WardleDA, WrightIJ, AertsR, et al. (2008). Plant species traits are the predominant control on litter decomposition rates within biomes worldwide Ecology Letters, 11, 1065-1071. DOI:10.1111/j.1461-0248.2008.01219.xPMID:18627410 [本文引用: 4] Worldwide decomposition rates depend both on climate and the legacy of plant functional traits as litter quality. To quantify the degree to which functional differentiation among species affects their litter decomposition rates, we brought together leaf trait and litter mass loss data for 818 species from 66 decomposition experiments on six continents. We show that: (i) the magnitude of species-driven differences is much larger than previously thought and greater than climate-driven variation; (ii) the decomposability of a species' litter is consistently correlated with that species' ecological strategy within different ecosystems globally, representing a new connection between whole plant carbon strategy and biogeochemical cycling. This connection between plant strategies and decomposability is crucial for both understanding vegetation-soil feedbacks, and for improving forecasts of the global carbon cycle.
CusackDF, ChouWW, YangWH, HarmonME, SilverWL, TeamTL (2009). Controls on long-term root and leaf litter decomposition in neotropical forests Global Change Biology, 15, 1339-1355. DOI:10.1111/gcb.2009.15.issue-5URL [本文引用: 3]
DíazS, CabidoM (2001). Vive la différence: plant functional diversity matters to ecosystem processes Trends in Ecology & Evolution, 16, 646-655. DOI:10.1016/S0169-5347(01)02283-2URL [本文引用: 1]
FreschetGT, CornwellWK, WardleDA, ElumeevaTG, LiuW, JacksonBG, OnipchenkoVG, SoudzilovskaiaNA, TaoJ, CornelissenJHC (2013). Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide Journal of Ecology, 101, 943-952. DOI:10.1111/1365-2745.12092URL [本文引用: 1]
García-PalaciosP, PrietoI, OurcivalJM, HättenschwilerS (2016). Disentangling the litter quality and soil microbial contribution to leaf and fine root litter decomposition responses to reduced rainfall Ecosystems, 19, 490-503. DOI:10.1007/s10021-015-9946-xURL [本文引用: 1]
HuangfuCH, WeiZS (2018). Nitrogen addition drives convergence of leaf litter decomposition rates between Flaveria bidentis and native plant Plant Ecology, 219, 1355-1368. DOI:10.1007/s11258-018-0884-5URL [本文引用: 2]
JiaBR(2019). Litter decomposition and its underlying mechanisms Chinese Journal of Plant Ecology, 43, 648-657. DOI:10.17521/cjpe.2019.0097URL [本文引用: 2]
JiangL, KouL, LiSG (2018). Alterations of early-stage decomposition of leaves and absorptive roots by deposition of nitrogen and phosphorus have contrasting mechanisms Soil Biology & Biochemistry, 127, 213-222. DOI:10.1016/j.soilbio.2018.09.037URL [本文引用: 1]
KazakouE, VileD, ShipleyB, GalletC, GarnierE (2006). Blackwell Publishing Ltd Co-variations in litter decomposition, leaf traits and plant growth in species from a Mediterranean old-field succession Functional Ecology, 20, 21-30. [本文引用: 1]
LanY, CuiBS, YouZY, LiX, HanZ, ZhangYT, ZhangY (2012). Litter decomposition of six macrophytes in a eutrophic shallow lake (Baiyangdian Lake, China) Clean—Soil, Air, Water, 40, 1159-1166. DOI:10.1002/clen.201200056URL [本文引用: 1]
LiGH(2010). Effect of organic amendments and chemical fertilizer on soil microbial activity, biomass and community structure Chinese Agricultural Science Bulletin, 26(14), 204-208. [本文引用: 1]
LiuP, HuangJH, HanXG, SunOJ (2009). Litter decomposition in semiarid grassland of Inner Mongolia, China Rangeland Ecology & Management, 62, 305-313. DOI:10.2111/08-040.1URL [本文引用: 1]
LiuX, XiongYM, LiaoBW (2017). Relative contributions of leaf litter and fine roots to soil organic matter accumulation in mangrove forests Plant and Soil, 421, 493-503. DOI:10.1007/s11104-017-3477-5URL [本文引用: 1]
MaCE, XiongYM, LiL, GuoDL (2016). Root and leaf decomposition become decoupled over time: implications for below- and above-ground relationships Functional Ecology, 30, 1239-1246. DOI:10.1111/fec.2016.30.issue-7URL [本文引用: 1]
MeierCL, BowmanWD (2008a). Links between plant litter chemistry, species diversity, and below-ground ecosystem function Proceedings of the National Academy of Sciences of the United States of America, 105,19780-19785. [本文引用: 1]
MengYY, ZhangLM, YuanYS, JiaX, ChengH, HuangfuCH(2021). Effects of soil moisture content and litter quality on decomposition of Carex thunbergii fine roots and leaf litter Research of Environmental Sciences, 34, 707-714. [本文引用: 2]
OʼLearHA, SeastedtTR (1994). Landscape patterns of litter decomposition in alpine tundra Oecologia, 99, 95-101. DOI:10.1007/BF00317088URL [本文引用: 1]
OlsonJS (1963). Energy storage and the balance of producers and decomposers in ecological systems Ecology, 44, 322-331. DOI:10.2307/1932179URL [本文引用: 1]
PetragliaA, CacciatoriC, ChelliS, FenuG, CalderisiG, GarganoD, AbeliT, OrsenigoS, CarbognaniM (2019). Litter decomposition: effects of temperature driven by soil moisture and vegetation type Plant and Soil, 435, 187-200. DOI:10.1007/s11104-018-3889-xURL [本文引用: 1]
ShiY, WenZM, GongSH(2011). Comparisons of relationships between leaf and fine root traits in hilly area of the Loess Plateau, Yanhe River basin, Shaanxi Province, China Acta Ecologica Sinica, 31, 6805-6814. [本文引用: 1]
SunT, DongLL, WangZW, LüX, MaoZJ (2016). Effects of long-term nitrogen deposition on fine root decomposition and its extracellular enzyme activities in temperate forests Soil Biology & Biochemistry, 93, 50-59. DOI:10.1016/j.soilbio.2015.10.023URL [本文引用: 1]
SunT, HobbieSE, BergB, ZhangH, WangQ, WangZ, HättenschwilerS (2018). Contrasting dynamics and trait controls in first-order root compared with leaf litter decomposition Proceedings of the National Academy of Sciences of the United States of America, 115, 10392-10397. [本文引用: 1]
SunT, MaoZJ, HanYY (2013). Slow decomposition of very fine roots and some factors controlling the process: a 4-year experiment in four temperate tree species Plant and Soil, 372, 445-458. DOI:10.1007/s11104-013-1755-4URL [本文引用: 1]
van SoestPJ (1963). Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and lignin Journal of Association of Official Agricultural Chemists, 46, 829-835. [本文引用: 1]
ViolleC, NavasML, VileD, KazakouE, FortunelC, HummelI, GarnierE (2007). Let the concept of trait be functional! Oikos, 116, 882-892. DOI:10.1111/oik.2007.116.issue-5URL [本文引用: 2]
WangJ, LiuLL, WangX, YangS, ZhangBB, LiP, QiaoCL, DengMF, LiuWX (2017a). High night-time humidity and dissolved organic carbon content support rapid decomposition of standing litter in a semi-arid landscape Functional Ecology, 31, 1659-1668. DOI:10.1111/fec.2017.31.issue-8URL [本文引用: 2]
WangJ, YangS, ZhangBB, LiuWX, DengMF, ChenSP, LiuLL (2017b). Temporal dynamics of ultraviolet radiation impacts on litter decomposition in a semi-arid ecosystem Plant and Soil, 419, 71-81. DOI:10.1007/s11104-017-3290-1URL [本文引用: 2]
WangX, XuZW, LüXT, WangRZ, CaiJP, YangS, LiMH, JiangY (2017c). Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland Plant and Soil, 418, 241-253. DOI:10.1007/s11104-017-3288-8URL [本文引用: 1]
WeedonJT, CornwellWK, CornelissenJHC, ZanneAE, WirthC, CoomesDA (2009). Global meta-analysis of wood decomposition rates: a role for trait variation among tree species? Ecology Letters, 12, 45-56. DOI:10.1111/j.1461-0248.2008.01259.xPMID:19016827 [本文引用: 1] The carbon flux from woody debris, a crucial uncertainty within global carbon-climate models, is simultaneously affected by climate, site environment and species-based variation in wood quality. In the first global analysis attempting to explicitly tease out the wood quality contribution to decomposition, we found support for our hypothesis that, under a common climate, interspecific differences in wood traits affect woody debris decomposition patterns. A meta-analysis of 36 studies from all forested continents revealed that nitrogen, phosphorus, and C : N ratio correlate with decomposition rates of angiosperms. In addition, gymnosperm wood consistently decomposes slower than angiosperm wood within common sites, a pattern that correlates with clear divergence in wood traits between the two groups. New empirical studies are needed to test whether this difference is due to a direct effect of wood trait variation on decomposer activity or an indirect effect of wood traits on decomposition microsite environment. The wood trait-decomposition results point to an important role for changes in the wood traits of dominant tree species as a driver of carbon cycling, with likely feedback to atmospheric CO(2) particularly where angiosperm species replace gymnosperms regionally. Truly worldwide upscaling of our results will require further site-based multi-species wood trait and decomposition data, particularly from low-latitude ecosystems.
WirthC, LichsteinJW (2009). The imprint of species turnover on old-growth forest carbon balances—Insights from a trait-based model of forest dynamics//Wirth C, Gleixner G, Heimann M. Old-Growth Forests Springer, Berlin. 81-113. [本文引用: 1]
XiongYM, FanPP, FuSL, ZengH, GuoDL (2013). Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees Plant and Soil, 363, 19-31. DOI:10.1007/s11104-012-1290-8URL [本文引用: 1]
YangZM, Hasitamier, LiuXM(2016). Effects of grazing on the composition of soil animals and their decomposition function to Stipa grandis litter in Inner Mongolia typical steppe, China Chinese Journal of Applied Ecology, 27, 2864-2874. DOI:10.13287/j.1001-9332.201609.033PMID:29732849 [本文引用: 2] Grazing has been considered to be an important factor determining the composition of soil animals and decomposition of leaf litter in grassland ecosystem. Sampling plots were selected in ungrazed grassland, grazed grassland and sandy land. Litter bags were used to compare the changes of physicochemical properties of Stipa grandis litter and the composition of soil fauna in the process of the litter decomposition in Baiyinxile, Inner Mongolia, since 2010 to 2012. A total number of 67056 soil animals were captured, belonging to five phyla and eight classes, including 23 families of mites and 19 families of insects. After 780 days' decomposition, the loss of the organic matter of S. grandis litter was from 92.5% to 40.0% in the ungrazed grassland, and to 41.3% in the grazed grassland, with no significant difference observed. However, there was a significant difference (P<0.05) between the ratio of litter residues of the ungrazed grassland (50.0%) and that of the grazed grassland (23.0%). The abundance of soil animals in the residual litters was significantly decreased in the grazed grassland compared to the ungrazed grassland. When the litter was moved into the sandy land, the decomposition rate of organic matter in the residual litter was not significant changed but the ratio of litter residue declined significantly, and the composition of mite community in the resi-dual litter changed significantly. Our results illustrated that grazing activity could affect the composition and abundance of soil fauna in temperate grassland, but slightly influenced the decomposition of organic matter. Therefore, soil animals had relatively weak direct effects on the decomposition of litter in this semi-arid region. [ 杨志敏, 哈斯塔米尔, 刘新民(2016). 放牧对内蒙古典型草原大针茅凋落物中土壤动物组成及其分解功能的影响 应用生态学报, 27, 2864-2874.] PMID:29732849 [本文引用: 2]
ZanneAE, OberleB, DunhamKM, MiloAM, WaltonML, YoungDF (2015). A deteriorating state of affairs: How endogenous and exogenous factors determine plant decay rates Journal of Ecology, 103, 1421-1431. DOI:10.1111/jec.2015.103.issue-6URL [本文引用: 2]
ZhaoHM, HuangG, MaJ, LiY, TangLS (2014). Decomposition of aboveground and root litter for three desert herbs: mass loss and dynamics of mineral nutrients Biology and Fertility of Soils, 50, 745-753. DOI:10.1007/s00374-013-0892-5URL [本文引用: 3]
ZhouSX, HuangCD, XiangYB, HanBH, XiaoYX, TangJD(2016). Effects of simulated nitrogen deposition on lignin and cellulose degradation of foliar litter in natural evergreen broad-leaved forest in Rainy Area of Western China Chinese Journal of Applied Ecology, 27, 1368-1374. [本文引用: 3]
ZhouX, HeZ, DingF, LiL, StoffellaPJ (2018). Biomass decaying and elemental release of aquatic macrophyte detritus in waterways of the Indian River Lagoon basin, South Florida, USA Science of the Total Environment, 635, 878-891. DOI:10.1016/j.scitotenv.2018.04.047URL [本文引用: 1]
Decomposition and nutrient release of Eichhornia crassipes (Mart.) Solms. under different trophic conditions in wetlands of eastern Himalayan foothills 1 2012
... 很多研究结果表明, 凋落物初始的N含量、C:N与凋落物分解显著相关, N含量越高, C:N越低, 凋落物分解越快(Balasubramanian et al., 2012; Lan et al., 2012; Li et al., 2012).本研究中, 6种植物的叶凋落物N含量远高于其根系的N含量, 这也可能是叶凋落物分解较快的原因.同时通过多元逐步回归发现N含量和C:N也是本研究中影响根系分解的主要影响因素, 但从相关性分析中发现, 叶凋落物的N含量与其分解前期和整个分解过程的k呈正相关关系, 而根系的N含量仅与其分解后期的k呈负相关关系, 这可能与凋落物分解过程中自身木质素的含量有关.另外, 研究发现微生物的生长和活性与N含量相关, 凋落物的N含量高可以间接增加微生物活性(李桂花, 2010), 而微生物生长代谢繁殖较快会加速凋落物的分解.同理, C:N的高低对微生物生长和凋落物的分解具有相同的影响, 所以相关分析发现本研究中6种植物的叶凋落物在分解前期和后期C:N越低, k越高, 分解速率越快.但是本研究还发现, 根系在分解的中期和后期表现为C:N越低, k越低, 分解速率越慢, 这可能是因为根系在分解的中期和后期受木质素含量影响较大, 因此木质素:N也常作为凋落物分解速率的影响因素(孟盈盈等, 2021).通过上述分析可以看出, 在不同分解阶段, 影响6种植物叶凋落物和根系分解的因素不同, 正或负的效应可能也受性状之间相互关系的影响, 因而叶凋落物和根系分解速率的调控因子也有所不同. ...
Going underground: root traits as drivers of ecosystem processes 1 2014
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Linkages between plant litter diversity, soil microbial biomass and ecosystem function in temperate grasslands 1 1999
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Patterns in the fate of production in plant communities 1 1999
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Roots and associated fungi drive long-term carbon sequestration in boreal forest 1 2013
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Variation in relative growth rate among woody species//Lambers H, Poorter H, van Vuuren MMI. Inherent Variation in Plant Growth: Physiological Mechanisms and Ecological Consequences 1 1998
... 另外, 本研究中叶片干物质含量、根比表面积和根组织密度分别与6种植物叶凋落物和根系的k表现出一定的相关性, 尤其是叶片干物质含量与叶凋落物在分解中期的k呈显著负相关关系.早有研究表明, 植物的一些物理性状对其凋落物分解过程具有显著影响(Cornelissen & Thompson, 1997; Cornelissen et al., 1998), 尤其是叶片干物质含量(Kazakou et al., 2006), 高的叶片干物质含量代表慢速生长的植物策略, 对应的植物凋落物分解速率也慢, 我们的研究结果与之一致. ...
... 另外, 本研究中叶片干物质含量、根比表面积和根组织密度分别与6种植物叶凋落物和根系的k表现出一定的相关性, 尤其是叶片干物质含量与叶凋落物在分解中期的k呈显著负相关关系.早有研究表明, 植物的一些物理性状对其凋落物分解过程具有显著影响(Cornelissen & Thompson, 1997; Cornelissen et al., 1998), 尤其是叶片干物质含量(Kazakou et al., 2006), 高的叶片干物质含量代表慢速生长的植物策略, 对应的植物凋落物分解速率也慢, 我们的研究结果与之一致. ...
Plant species traits are the predominant control on litter decomposition rates within biomes worldwide 4 2008
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
... ; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
... ; Cornwell et al., 2008; Weedon et al., 2009). ...
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Controls on long-term root and leaf litter decomposition in neotropical forests 3 2009
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
Vive la différence: plant functional diversity matters to ecosystem processes 1 2001
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Linking litter decomposition of above- and below-ground organs to plant-soil feedbacks worldwide 1 2013
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Disentangling the litter quality and soil microbial contribution to leaf and fine root litter decomposition responses to reduced rainfall 1 2016
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Diversity meets decomposition 1 2010
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Nitrogen addition drives convergence of leaf litter decomposition rates between Flaveria bidentis and native plant 2 2018
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
凋落物分解及其影响机制 2 2019
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... ; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
凋落物分解及其影响机制 2 2019
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... ; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Alterations of early-stage decomposition of leaves and absorptive roots by deposition of nitrogen and phosphorus have contrasting mechanisms 1 2018
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Blackwell Publishing Ltd Co-variations in litter decomposition, leaf traits and plant growth in species from a Mediterranean old-field succession 1 2006
... 另外, 本研究中叶片干物质含量、根比表面积和根组织密度分别与6种植物叶凋落物和根系的k表现出一定的相关性, 尤其是叶片干物质含量与叶凋落物在分解中期的k呈显著负相关关系.早有研究表明, 植物的一些物理性状对其凋落物分解过程具有显著影响(Cornelissen & Thompson, 1997; Cornelissen et al., 1998), 尤其是叶片干物质含量(Kazakou et al., 2006), 高的叶片干物质含量代表慢速生长的植物策略, 对应的植物凋落物分解速率也慢, 我们的研究结果与之一致. ...
Litter decomposition of six macrophytes in a eutrophic shallow lake (Baiyangdian Lake, China) 1 2012
... 很多研究结果表明, 凋落物初始的N含量、C:N与凋落物分解显著相关, N含量越高, C:N越低, 凋落物分解越快(Balasubramanian et al., 2012; Lan et al., 2012; Li et al., 2012).本研究中, 6种植物的叶凋落物N含量远高于其根系的N含量, 这也可能是叶凋落物分解较快的原因.同时通过多元逐步回归发现N含量和C:N也是本研究中影响根系分解的主要影响因素, 但从相关性分析中发现, 叶凋落物的N含量与其分解前期和整个分解过程的k呈正相关关系, 而根系的N含量仅与其分解后期的k呈负相关关系, 这可能与凋落物分解过程中自身木质素的含量有关.另外, 研究发现微生物的生长和活性与N含量相关, 凋落物的N含量高可以间接增加微生物活性(李桂花, 2010), 而微生物生长代谢繁殖较快会加速凋落物的分解.同理, C:N的高低对微生物生长和凋落物的分解具有相同的影响, 所以相关分析发现本研究中6种植物的叶凋落物在分解前期和后期C:N越低, k越高, 分解速率越快.但是本研究还发现, 根系在分解的中期和后期表现为C:N越低, k越低, 分解速率越慢, 这可能是因为根系在分解的中期和后期受木质素含量影响较大, 因此木质素:N也常作为凋落物分解速率的影响因素(孟盈盈等, 2021).通过上述分析可以看出, 在不同分解阶段, 影响6种植物叶凋落物和根系分解的因素不同, 正或负的效应可能也受性状之间相互关系的影响, 因而叶凋落物和根系分解速率的调控因子也有所不同. ...
Mycorrhizal controls on belowground litter quality 1 2003
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
不同施肥对土壤微生物活性、群落结构和生物量的影响 1 2010
... 很多研究结果表明, 凋落物初始的N含量、C:N与凋落物分解显著相关, N含量越高, C:N越低, 凋落物分解越快(Balasubramanian et al., 2012; Lan et al., 2012; Li et al., 2012).本研究中, 6种植物的叶凋落物N含量远高于其根系的N含量, 这也可能是叶凋落物分解较快的原因.同时通过多元逐步回归发现N含量和C:N也是本研究中影响根系分解的主要影响因素, 但从相关性分析中发现, 叶凋落物的N含量与其分解前期和整个分解过程的k呈正相关关系, 而根系的N含量仅与其分解后期的k呈负相关关系, 这可能与凋落物分解过程中自身木质素的含量有关.另外, 研究发现微生物的生长和活性与N含量相关, 凋落物的N含量高可以间接增加微生物活性(李桂花, 2010), 而微生物生长代谢繁殖较快会加速凋落物的分解.同理, C:N的高低对微生物生长和凋落物的分解具有相同的影响, 所以相关分析发现本研究中6种植物的叶凋落物在分解前期和后期C:N越低, k越高, 分解速率越快.但是本研究还发现, 根系在分解的中期和后期表现为C:N越低, k越低, 分解速率越慢, 这可能是因为根系在分解的中期和后期受木质素含量影响较大, 因此木质素:N也常作为凋落物分解速率的影响因素(孟盈盈等, 2021).通过上述分析可以看出, 在不同分解阶段, 影响6种植物叶凋落物和根系分解的因素不同, 正或负的效应可能也受性状之间相互关系的影响, 因而叶凋落物和根系分解速率的调控因子也有所不同. ...
不同施肥对土壤微生物活性、群落结构和生物量的影响 1 2010
... 很多研究结果表明, 凋落物初始的N含量、C:N与凋落物分解显著相关, N含量越高, C:N越低, 凋落物分解越快(Balasubramanian et al., 2012; Lan et al., 2012; Li et al., 2012).本研究中, 6种植物的叶凋落物N含量远高于其根系的N含量, 这也可能是叶凋落物分解较快的原因.同时通过多元逐步回归发现N含量和C:N也是本研究中影响根系分解的主要影响因素, 但从相关性分析中发现, 叶凋落物的N含量与其分解前期和整个分解过程的k呈正相关关系, 而根系的N含量仅与其分解后期的k呈负相关关系, 这可能与凋落物分解过程中自身木质素的含量有关.另外, 研究发现微生物的生长和活性与N含量相关, 凋落物的N含量高可以间接增加微生物活性(李桂花, 2010), 而微生物生长代谢繁殖较快会加速凋落物的分解.同理, C:N的高低对微生物生长和凋落物的分解具有相同的影响, 所以相关分析发现本研究中6种植物的叶凋落物在分解前期和后期C:N越低, k越高, 分解速率越快.但是本研究还发现, 根系在分解的中期和后期表现为C:N越低, k越低, 分解速率越慢, 这可能是因为根系在分解的中期和后期受木质素含量影响较大, 因此木质素:N也常作为凋落物分解速率的影响因素(孟盈盈等, 2021).通过上述分析可以看出, 在不同分解阶段, 影响6种植物叶凋落物和根系分解的因素不同, 正或负的效应可能也受性状之间相互关系的影响, 因而叶凋落物和根系分解速率的调控因子也有所不同. ...
Litter quality and interactions of macrophytes tissues decomposition in a eutrophic shallow lake 2 2012
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
... 很多研究结果表明, 凋落物初始的N含量、C:N与凋落物分解显著相关, N含量越高, C:N越低, 凋落物分解越快(Balasubramanian et al., 2012; Lan et al., 2012; Li et al., 2012).本研究中, 6种植物的叶凋落物N含量远高于其根系的N含量, 这也可能是叶凋落物分解较快的原因.同时通过多元逐步回归发现N含量和C:N也是本研究中影响根系分解的主要影响因素, 但从相关性分析中发现, 叶凋落物的N含量与其分解前期和整个分解过程的k呈正相关关系, 而根系的N含量仅与其分解后期的k呈负相关关系, 这可能与凋落物分解过程中自身木质素的含量有关.另外, 研究发现微生物的生长和活性与N含量相关, 凋落物的N含量高可以间接增加微生物活性(李桂花, 2010), 而微生物生长代谢繁殖较快会加速凋落物的分解.同理, C:N的高低对微生物生长和凋落物的分解具有相同的影响, 所以相关分析发现本研究中6种植物的叶凋落物在分解前期和后期C:N越低, k越高, 分解速率越快.但是本研究还发现, 根系在分解的中期和后期表现为C:N越低, k越低, 分解速率越慢, 这可能是因为根系在分解的中期和后期受木质素含量影响较大, 因此木质素:N也常作为凋落物分解速率的影响因素(孟盈盈等, 2021).通过上述分析可以看出, 在不同分解阶段, 影响6种植物叶凋落物和根系分解的因素不同, 正或负的效应可能也受性状之间相互关系的影响, 因而叶凋落物和根系分解速率的调控因子也有所不同. ...
科尔沁沙地植物成熟叶片性状与叶凋落物分解的关系 2 2008
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... ).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
科尔沁沙地植物成熟叶片性状与叶凋落物分解的关系 2 2008
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... ).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Litter decomposition in semiarid grassland of Inner Mongolia, China 1 2009
... 在内蒙古典型草原, 关于凋落物分解影响因素的研究主要集中于湿度、紫外辐射、水分和养分以及利用方式变化对凋落物分解的影响(Liu et al., 2009; 杨志敏等, 2016; Wang et al., 2017a, 2017b, 2017c).例如, 杨志敏等(2016)发现放牧并未对大针茅(Stipa grandis)叶凋落物分解速率产生显著影响.Wang等(2017b)发现紫外辐射可以通过改变糙隐子草(Cleistogenes squarrosa)和克氏针茅(S. krylovii)叶凋落物不同分解时期的生物降解性以及微生物活性促进其分解.尽管这些研究从不同方面深入探讨了草原凋落物的分解过程和影响因素, 但仍存在一些不足.首先大部分分解实验主要以草原群落的优势植物为研究对象, 缺乏对其他植物凋落物的分解研究, 尤其对于根系分解过程的研究仍不足; 其次, 植物功能性状在草原凋落物分解过程中的作用并不清楚, 尤其对于叶凋落物和根系分解的影响效应有待研究.因此, 本研究以大针茅草原6种植物的叶凋落物和根系为研究对象, 在分析叶凋落物和根系的分解特征与功能性状的基础上, 探讨两者之间的关系, 阐明大针茅草原叶凋落物和根系分解的主要影响因素, 以期为预测大针茅典型草原的碳和养分循环过程提供参考数据. ...
Relative contributions of leaf litter and fine roots to soil organic matter accumulation in mangrove forests 1 2017
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Root and leaf decomposition become decoupled over time: implications for below- and above-ground relationships 1 2016
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Links between plant litter chemistry, species diversity, and below-ground ecosystem function 1 2008a
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Global patterns in root decomposition: comparisons of climate and litter quality effects 2 2001
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Litter chemistry and chemical diversity drive ecosystem processes in forest ponds 1 2016
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Effects of long-term nitrogen deposition on fine root decomposition and its extracellular enzyme activities in temperate forests 1 2016
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Contrasting dynamics and trait controls in first-order root compared with leaf litter decomposition 1 2018
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
Slow decomposition of very fine roots and some factors controlling the process: a 4-year experiment in four temperate tree species 1 2013
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and lignin 1 1963
... 6种植物的叶凋落物与根系经研磨过40目筛后, 分别称取0.50 g放入滤袋封口, 采用van Soest (1963)的提取方法, 使用ANKOM纤维分析仪(A2000I, ANKOM, New York, USA)进行非结构性碳水化合物含量和结构性碳水化合物(半纤维素、纤维素及木质素)含量的测定.凋落物的C、N含量采用CHNS元素分析仪(Elementar Vario EL III, Elementar, Hanau, Germany)进行测定. ...
Let the concept of trait be functional! 2 2007
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
... ), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
High night-time humidity and dissolved organic carbon content support rapid decomposition of standing litter in a semi-arid landscape 2 2017a
... 在内蒙古典型草原, 关于凋落物分解影响因素的研究主要集中于湿度、紫外辐射、水分和养分以及利用方式变化对凋落物分解的影响(Liu et al., 2009; 杨志敏等, 2016; Wang et al., 2017a, 2017b, 2017c).例如, 杨志敏等(2016)发现放牧并未对大针茅(Stipa grandis)叶凋落物分解速率产生显著影响.Wang等(2017b)发现紫外辐射可以通过改变糙隐子草(Cleistogenes squarrosa)和克氏针茅(S. krylovii)叶凋落物不同分解时期的生物降解性以及微生物活性促进其分解.尽管这些研究从不同方面深入探讨了草原凋落物的分解过程和影响因素, 但仍存在一些不足.首先大部分分解实验主要以草原群落的优势植物为研究对象, 缺乏对其他植物凋落物的分解研究, 尤其对于根系分解过程的研究仍不足; 其次, 植物功能性状在草原凋落物分解过程中的作用并不清楚, 尤其对于叶凋落物和根系分解的影响效应有待研究.因此, 本研究以大针茅草原6种植物的叶凋落物和根系为研究对象, 在分析叶凋落物和根系的分解特征与功能性状的基础上, 探讨两者之间的关系, 阐明大针茅草原叶凋落物和根系分解的主要影响因素, 以期为预测大针茅典型草原的碳和养分循环过程提供参考数据. ...
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
Temporal dynamics of ultraviolet radiation impacts on litter decomposition in a semi-arid ecosystem 2 2017b
... 在内蒙古典型草原, 关于凋落物分解影响因素的研究主要集中于湿度、紫外辐射、水分和养分以及利用方式变化对凋落物分解的影响(Liu et al., 2009; 杨志敏等, 2016; Wang et al., 2017a, 2017b, 2017c).例如, 杨志敏等(2016)发现放牧并未对大针茅(Stipa grandis)叶凋落物分解速率产生显著影响.Wang等(2017b)发现紫外辐射可以通过改变糙隐子草(Cleistogenes squarrosa)和克氏针茅(S. krylovii)叶凋落物不同分解时期的生物降解性以及微生物活性促进其分解.尽管这些研究从不同方面深入探讨了草原凋落物的分解过程和影响因素, 但仍存在一些不足.首先大部分分解实验主要以草原群落的优势植物为研究对象, 缺乏对其他植物凋落物的分解研究, 尤其对于根系分解过程的研究仍不足; 其次, 植物功能性状在草原凋落物分解过程中的作用并不清楚, 尤其对于叶凋落物和根系分解的影响效应有待研究.因此, 本研究以大针茅草原6种植物的叶凋落物和根系为研究对象, 在分析叶凋落物和根系的分解特征与功能性状的基础上, 探讨两者之间的关系, 阐明大针茅草原叶凋落物和根系分解的主要影响因素, 以期为预测大针茅典型草原的碳和养分循环过程提供参考数据. ...
Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland 1 2017c
... 在内蒙古典型草原, 关于凋落物分解影响因素的研究主要集中于湿度、紫外辐射、水分和养分以及利用方式变化对凋落物分解的影响(Liu et al., 2009; 杨志敏等, 2016; Wang et al., 2017a, 2017b, 2017c).例如, 杨志敏等(2016)发现放牧并未对大针茅(Stipa grandis)叶凋落物分解速率产生显著影响.Wang等(2017b)发现紫外辐射可以通过改变糙隐子草(Cleistogenes squarrosa)和克氏针茅(S. krylovii)叶凋落物不同分解时期的生物降解性以及微生物活性促进其分解.尽管这些研究从不同方面深入探讨了草原凋落物的分解过程和影响因素, 但仍存在一些不足.首先大部分分解实验主要以草原群落的优势植物为研究对象, 缺乏对其他植物凋落物的分解研究, 尤其对于根系分解过程的研究仍不足; 其次, 植物功能性状在草原凋落物分解过程中的作用并不清楚, 尤其对于叶凋落物和根系分解的影响效应有待研究.因此, 本研究以大针茅草原6种植物的叶凋落物和根系为研究对象, 在分析叶凋落物和根系的分解特征与功能性状的基础上, 探讨两者之间的关系, 阐明大针茅草原叶凋落物和根系分解的主要影响因素, 以期为预测大针茅典型草原的碳和养分循环过程提供参考数据. ...
Global meta-analysis of wood decomposition rates: a role for trait variation among tree species? 1 2009
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
The imprint of species turnover on old-growth forest carbon balances—Insights from a trait-based model of forest dynamics//Wirth C, Gleixner G, Heimann M. Old-Growth Forests 1 2009
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Slow decomposition and limited nitrogen release by lower order roots in eight Chinese temperate and subtropical trees 1 2013
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
放牧对内蒙古典型草原大针茅凋落物中土壤动物组成及其分解功能的影响 2 2016
... 在内蒙古典型草原, 关于凋落物分解影响因素的研究主要集中于湿度、紫外辐射、水分和养分以及利用方式变化对凋落物分解的影响(Liu et al., 2009; 杨志敏等, 2016; Wang et al., 2017a, 2017b, 2017c).例如, 杨志敏等(2016)发现放牧并未对大针茅(Stipa grandis)叶凋落物分解速率产生显著影响.Wang等(2017b)发现紫外辐射可以通过改变糙隐子草(Cleistogenes squarrosa)和克氏针茅(S. krylovii)叶凋落物不同分解时期的生物降解性以及微生物活性促进其分解.尽管这些研究从不同方面深入探讨了草原凋落物的分解过程和影响因素, 但仍存在一些不足.首先大部分分解实验主要以草原群落的优势植物为研究对象, 缺乏对其他植物凋落物的分解研究, 尤其对于根系分解过程的研究仍不足; 其次, 植物功能性状在草原凋落物分解过程中的作用并不清楚, 尤其对于叶凋落物和根系分解的影响效应有待研究.因此, 本研究以大针茅草原6种植物的叶凋落物和根系为研究对象, 在分析叶凋落物和根系的分解特征与功能性状的基础上, 探讨两者之间的关系, 阐明大针茅草原叶凋落物和根系分解的主要影响因素, 以期为预测大针茅典型草原的碳和养分循环过程提供参考数据. ...
A deteriorating state of affairs: How endogenous and exogenous factors determine plant decay rates 2 2015
... 凋落物分解是一个关键的生态系统功能, 影响养分循环和土壤有机质的形成(Meier & Bowman, 2008b; Wirth & Lichstein, 2009; Petraglia et al., 2019).据统计, 陆地生态系统中90%的生物量以凋落物分解的形式回归到生态系统中, 以维持其正常运转(Cebrian, 1999).已有研究表明, 凋落物分解过程受内源(凋落物自身化学组成和形态特征)(Cornwell et al., 2008; Meier & Bowman, 2008a; Stoler et al., 2016)和外源(气候、地形、环境背景)(OʼLear & Seastedt, 1994; Zanne et al., 2015)等因素及其相互作用的影响(Bardgett & Shine, 1999; Cornwell et al., 2008; Gessner et al., 2010; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
... ; Zanne et al., 2015).其中, 凋落物分解受到内源因素的影响可总结为是受到植物功能性状的影响.功能性状是植物在进化过程中适应外界环境所表现的特征, 能够响应外部环境变化, 影响生态系统功能(Díaz & Cabido, 2001; Violle et al., 2007), 对植物个体的生长、生存有重要意义(Violle et al., 2007).这些与植物获取资源或者防御有关的特征, 解释了大部分凋落物分解过程(分解速率)所产生的变化(Silver & Miya, 2001; Cornwell et al., 2008; Weedon et al., 2009). ...
Decomposition of aboveground and root litter for three desert herbs: mass loss and dynamics of mineral nutrients 3 2014
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... )或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
模拟氮沉降对华西雨屏区天然常绿阔叶林凋落物木质素和纤维素降解的影响 3 2016
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
... 目前很多研究将凋落物分解速率与植物的功能性状相联系(Silver & Miya, 2001; Cornwell et al., 2008; 李玉霖等, 2008; Zhao et al., 2014).例如李玉霖等(2008)对科尔沁沙地20种植物的叶凋落物分解及其与11个叶性状的关系进行研究, 发现叶片的一些性状可以预测叶凋落物分解速率.因为叶片的性状易于观测, 所以目前大部分地上凋落物的分解研究仍以叶片为主(贾丙瑞, 2019).但是越来越多的研究表明, 土壤有机碳可能大部分源于细根的死亡和分解(Clemmensen et al., 2013; Bardgett et al., 2014).与叶凋落物相比, 虽然细根在植物体生物量中占比较小, 但由于其较低的分解速率(Freschet et al., 2013; Ma et al., 2016; Sun et al., 2016; Liu et al., 2017), 对土壤碳(C)和氮(N)的贡献可能要大于地上凋落物(Langley & Hungate, 2003; Xiong et al., 2013; 贾丙瑞, 2019), 这使得根系分解研究同样成为目前C、N等养分循环研究的热点.已有研究表明叶凋落物和根系的初始化学成分与分解速率之间的影响效应并不一致(García-Palacios et al., 2016; Jiang et al., 2018), 例如有研究认为, 与叶凋落物相比, 根系的结构性碳水化合物含量较多, 导致其分解速率和养分释放较慢(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018), 但也有研究发现如果根系的非结构性碳水化合物含量较高, 可能会导致其分解速率变快(Sun et al., 2013)或高于叶凋落物(Zhao et al., 2014).但是同种植物叶和根系的功能性状本身存在一定关联性, 这些性状和凋落物分解速率之间的关系有待于深入研究. ...
... 本研究中, 在整个分解过程中根系的分解慢于叶凋落物(图1; 表3), 这与不少研究结果(Zhao et al.,2014; 孟盈盈等, 2021)相同, 这主要因为6种植物根系的木质素含量普遍高于叶凋落物, 而木质素是微生物较难分解的物质, 其含量较高就会对凋落物的分解产生一定的抑制作用(Cusack et al., 2009; 周世兴等, 2016; Huangfu & Wei, 2018).很多研究认为非结构性碳水化合物作为易降解的成分, 其含量越高对凋落物分解越有利(Wang et al., 2017a; Zhou et al., 2018).Li等(2012)研究发现, 分解早期凋落物分解较快与非结构性碳水化合物和半纤维的快速释放有关.本研究虽然通过相关分析发现叶凋落物和根系在分解前期的k与非结构性碳水化合物含量呈显著正相关关系, 但多元逐步回归分析发现在前期的分解过程中, 叶凋落物的木质素、纤维素、半纤维含量以及根系的纤维素、木质素含量与其k呈显著负相关关系, 说明在分解前期, 叶凋落物和根系的分解速率主要受结构性碳水化合物含量的制约.而在分解后期, 6种植物叶凋落物分解速率的主要影响因素并未发生变化, 但根系分解速率的主要影响因素则由结构性碳水化合物转变为非结构性碳水化合物及C:N和N含量, 这一结果与人们对于根系中结构性碳水化合物和难降解的物质是控制根系分解速率的主要因素(Cusack et al., 2009; 周世兴等, 2016)的认识不同.虽然通过相关分析发现根系木质素含量对其中期和后期的分解速率有较强的抑制性, 但通过多元逐步回归分析我们发现它并不是影响根系中后期分解速率的主要影响因素, 并且在根系分解的整个过程中, 非结构性碳水化合物含量是影响其分解速率的主要因素.最近Sun等(2018)在对阔叶红松林35种木本植物的凋落物进行6年的分解实验后发现, 与叶凋落物分解的影响因素不同, 非结构性碳水化合物及次生代谢产物对细根的分解起着主导作用, 这与本研究中根系分解速率的主要影响因素相符. ...
 ,1,3,*
,1,3,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT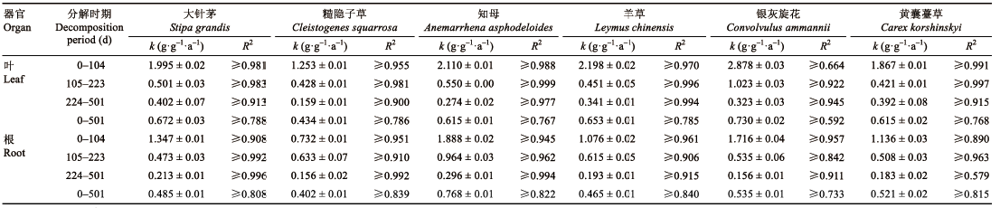
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT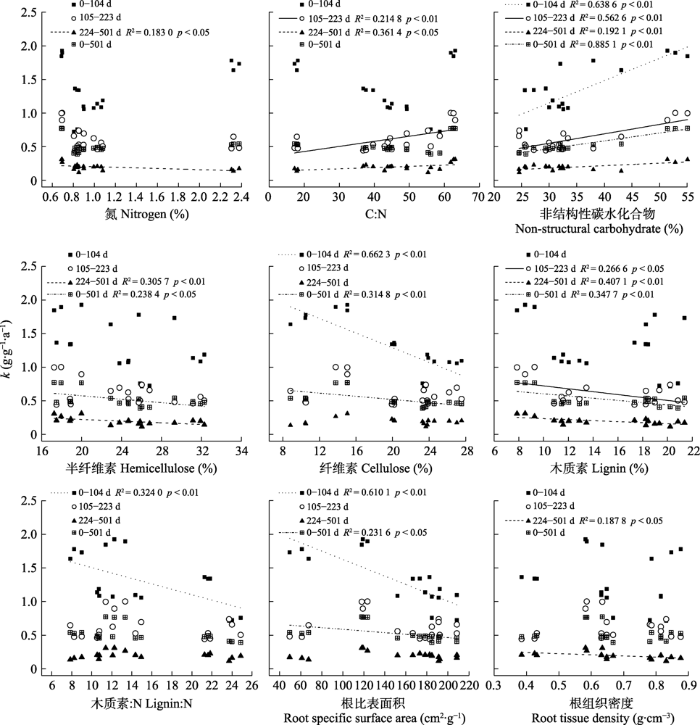 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}