孙建
, , 1 , 2 , 3 , * , 刘国华
2 , 3 1 中国科学院青藏高原研究所青藏高原地球系统与资源环境国家重点实验室, 北京 1001012 中国科学院生态环境研究中心城市与区域生态学国家重点实验室, 北京 1000853 中国科学院大学资源与环境学院, 北京 100190Alpine grassland on the Qingzang Plateau: pattern and process Jian SUN
, , 1 , 2 , 3 , * , Guo-Hua LIU
2 , 3 1 State Key Laboratory of Tibetan Plateau Earth System, Resources and Environment (TPESRE), Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing 100101, China2 State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China3 College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100190, China通讯作者: *孙建:ORCID: 0000-0001-8765-5015(E-mail:
jshe@pku.edu.cn )
编委: 唐志尧
责任编辑: 李敏
收稿日期: 2009-01-12
接受日期: 2009-06-3
Corresponding authors: *(E-mail:
jshe@pku.edu.cn )
Received: 2009-01-12
Accepted: 2009-06-3
PDF (5220KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 孙建, 刘国华. 青藏高原高寒草地: 格局与过程. 植物生态学报, 2021, 45(5): 429-433. DOI:
10.17521/cjpe.2021.0099 SUN Jian, LIU Guo-Hua.
Alpine grassland on the Qingzang Plateau: pattern and process .
Chinese Journal of Plant Ecology , 2021, 45(5): 429-433. DOI:
10.17521/cjpe.2021.0099 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 )。青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
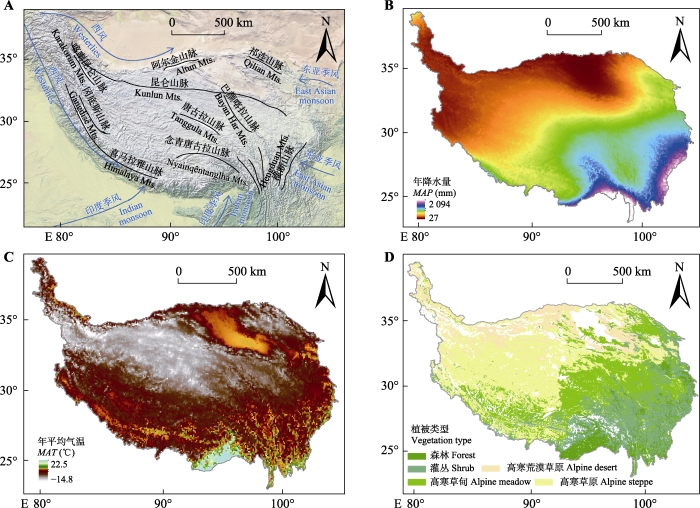
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性。喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 )。受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 )。高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 )。由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1青藏高原季风与主要山脉格局(A)、年降水量(B)、年平均气温(C)和植被类型地理格局(D)。 Fig. 1Pattern of monsoon and main mountains (A), mean annual precipitation (MAP )(B), mean annual air temperature (MAT )(C), geographical pattern of vegetation types (D) on the Qingzang Plateau. 目前, 青藏高原整体呈变暖趋势, 导致青藏高原冰川融化, 湖泊增加, 湖泊总数由1990年的868个增加到2015年的1 207个, 湖泊总水面面积从38 823.3 km
2 增加到48 793.0 km
2 (
Sun et al ., 2018 )。此外, 持续增加的冻土活动层厚度和大气氮沉降速率(
Wu & Zhang, 2010 ), 以及不断增加的人类活动, 如围栏和放牧等生态系统管理措施也在改变和修饰着高寒生态系统(
Sun et al ., 2020 )。这些全球变化的主要表现形式必将对高寒植被地理格局和过程产生深刻的影响, 理解其中的功能变化和过程机制, 有助于我们深入理解高寒草地的分布变化和发展趋势, 以推进青藏高原生态安全屏障建设, 支撑高原可持续发展, 这对国家生态文明建设等方面具有重要的参考价值。值得深度关注的是, 大量的研究都考虑自然过程, 忽略了无处不在的放牧利用方式和基本覆盖全域的围栏管理措施带来的影响, 导致真实的格局、过程和机制很难被深刻地理解或阐明。因此建议未来高寒草地格局与过程研究, 应该综合考虑人类活动要素。
近年来, 国内外****围绕全球变化背景下的高寒草地生态系统格局和过程等主题开展了大量工作, 并已经取得了重要进展(
图2 )。技术层面上, 随着测试手段的提升, 研究呈现技术多样性, 如卫星遥感、模型模拟、大尺度样带调查、无人机航拍和定位观测等; 研究尺度从区域、景观、生态系统和群落尺度发展到微观分子和基因组层面的研究等; 观测指标更为完整, 从基础指标观测完善为涵盖水分、土壤、生物、水域和气候五大观测指标体系; 驱动因素考虑逐渐全面, 涵盖干旱、冻融、鼠害、杂草入侵、放牧和围栏管理等驱动过程, 从环境因素发展到自然因素和人类活动以及整个生境对高寒生态系统影响的考虑; 关注点从营养级、食物网、地上和地下独立过程发展到整个生态系统各个过程地上和地下的链接; 研究机理更为深入, 从单纯的植物和植被分类, 植被地理格局与环境要素的关联, 发展到植被结构、功能、特征和过程的地理格局与环境要素的关联和响应与适应机制方面。新的时期、新的技术和理论, 为青藏高原植被地理格局和过程研究提供了新的研究方向。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2青藏高原高寒草地生态系统格局与过程研究概念图。 Fig. 2Mind map of alpine grassland ecosystem pattern and process on the Qingzang Plateau. 基于以上论述的意义和目的, 我们组织了“青藏高原高寒草地: 格局与过程”专辑, 以期推进后续的相关研究。本专辑收录了13篇文章, 内容涉及基于样带尺度上的地上生态系统多功能性, 土壤酶活性和有机质化学组成变化格局与成因, 以及在降水梯度上对高寒草地养分限制的机制探讨, 同时对海拔梯度上的植物功能性状也进行了梳理。基于站点的观测研究探讨了土壤营养、土壤微生物、植物多样性等动态变化与成因, 涉及人工草地、天然草地、鼠兔干扰草地和退化草地等不同草地类型和草地利用方式。
在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用。此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制。颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去。此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子。该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平。海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考。
针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因。
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识。
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位。针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性。而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本。此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段。
致谢 感谢中国科学院青藏高原研究所高原植被地理与可持续管理研究组的王毅、叶冲冲、贺文、米琦、李邵宇、张锦涛、刘天源、李若玮、李洋、何豫川、兰翔宇和孙乐等的长期科考实践和讨论, 这对本文的形成至关重要。
[1] Chen Z Wang H Wang JZ Shi HJ Liu HY He JS 2021 ).Estimation on seasonal dynamics of alpine grassland aboveground biomass using phenology camera-derivedNDVI. Chinese Journal of Plant Ecolog y487 -495 . [本文引用: 1] [陈哲 ,汪浩 ,王金洲 ,石慧瑾 ,刘慧颖 ,贺金生 (2021 ).基于物候相机归一化植被指数估算高寒草地植物地上生物量的季节动态植物生态学报 ,45, 487 -495 .] [本文引用: 1] [2] Dong LJ Li JH Chen S Zhang R Sun J Ma MJ 2021 ).Changes in soil organic carbon content and their causes during the degradation of alpine meadows in Zoigê WetlandChinese Journal of Plant Ecology ,45, 507 -515 . [本文引用: 1] [董利军 ,李金花 ,陈姗 ,张瑞 ,孙建 ,马妙君 (2021 ).若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析植物生态学报 ,45, 507 -515 .] [本文引用: 1] [3] Goldblum D Carrier C 2017 ).Climate Change and Biogeography John Wiley & Sons, San Francisco, USA . [本文引用: 1] [4] Guo K Zheng D 2002 ).Regional differentiation of vegetation on the West Kunlun, the West Karakorum, and the North-west Himalaya and the implication for the ecological environmentActa Phytoecologica Sinica ,26, 17 -22 . [本文引用: 1] [郭柯 ,郑度 (2002 ).西昆仑、西喀喇昆仑和西北喜马拉雅地区植被的地域分异及其指示意义植物生态学报 ,26, 17 -22 .] [本文引用: 1] [5] Jiang X Niu KC 2021 ).Effects of grass mixed-sowing on soil microbial diversity on the Qingzang (Tibetan) PlateauChinese Journal of Plant Ecology ,45, 539 -551 . [本文引用: 1] [姜鑫 ,牛克昌 (2021 ).青藏高原禾草混播对土壤微生物多样性的影响植物生态学报 ,45, 539 -551 .] [本文引用: 1] [6] Li J Chen YY Qiao FY Zhi DG Guo ZG 2021 ).Effect of disturbance by plateau pika on the β diversity of an alpine meadowChinese Journal of Plant Ecolog y476 -486 . [本文引用: 1] [李捷 ,陈莹莹 ,乔福云 ,郅堤港 ,郭正刚 (2021 ).高原鼠兔干扰对高寒草甸β多样性的影响植物生态学报 ,45, 476 -486 .] [本文引用: 1] [7] Liu P Wang WY Zhou HK Mao XF Liu YF 2021 ).Dynamics of soil soluble nitrogen and plant productivity in artificial pastures on the Qingzang PlateauChinese Journal of Plant Ecology ,45, 562 -572 . [本文引用: 1] [刘攀 ,王文颖 ,周华坤 ,毛旭峰 ,刘艳方 (2021 ).青藏高原人工草地土壤可溶性氮组分与植被生产力动态变化过程植物生态学报 ,45, 562 -572 .] [本文引用: 1] [8] Luo MM Chen Y Yang G Hu B Li W Chen H 2021 ).Short-term response of soil prokaryotic community structure to water level restoration in degraded peatland of the Zoigê PlateauChinese Journal of Plant Ecology ,45, 552 -561 . [本文引用: 1] [罗明没 ,陈悦 ,杨刚 ,胡斌 ,李玮 ,陈槐 (2021 ).若尔盖退化泥炭地土壤原核微生物群落结构对水位恢复的短期响应植物生态学报 ,45, 552 -561 .] [本文引用: 1] [9] Ma SQ Wang ZW Chen YC Lu XY 2021 ).Effect of soil organic matter chemical compositions on soil protease and urease activity in alpine grassland soils in Northern Xizang, ChinaChinese Journal of Plant Ecology ,45, 516 -527 . [本文引用: 1] [马书琴 ,汪子微 ,陈有超 ,鲁旭阳 (2021 ).藏北高寒草地土壤有机质化学组成对土壤蛋白酶和脲酶活性的影响,植物生态学报 ,45, 516 -527 .] [本文引用: 1] [10] Sun J Fu BJ Zhao WW Liu SL Liu GH Zhou HK Shao XQ Chen YC Zhang Y Deng YF 2021 ).Optimizing grazing exclusion practices to achieve Goal 15 of the sustainable development goals in the Tibetan PlateauScience Bulletin 10.1016/j.scib.2021.03.014 .DOI:
10.1016/j.scib.2021.03.014 [本文引用: 1] [11] Sun J Liu M Fu BJ Kemp D Zhao WW Liu GH Han GD Wilkes A Lu XY Chen YC Cheng GW Zhou TC Hou G Zhan TY Peng F et al. 2020 ).Reconsidering the efficiency of grazing exclusion using fences on the Tibetan PlateauScience Bulletin ,65, 1405 -1414 . DOI:
10.1016/j.scib.2020.04.035 URL [本文引用: 1] [12] Sun J Wang Y Liu GH 2021 ).Linkages of aboveground plant carbon accumulation rate with ecosystem multifunctionality in alpine grassland, Qingzang PlateauChinese Journal of Plant Ecology ,45, 496 -506 . [本文引用: 1] [孙建 ,王毅 ,刘国华 (2021 ).青藏高原高寒草地地上植物碳积累速率对生态系统多功能性的影响机制植物生态学报 ,45, 496 -506 .] [本文引用: 1] [13] Sun J Zhang ZC Dong SK 2019 ).Adaptive management of alpine grassland ecosytems over Tibetan PlateauPratacultural Science ,36, 933 -938 . [本文引用: 1] [孙建 ,张振超 ,董世魁 (2019 ).青藏高原高寒草地生态系统的适应性管理草业科学 ,36, 933 -938 .] [本文引用: 1] [14] Sun J Zhou TC Liu M Chen YC Shang H Zhu LP Shedayi AA Yu H Cheng GW Liu GH Xu M Deng W Fan JH Lu XY Sha YK 2018 ).Linkages of the dynamics of glaciers and lakes with the climate elements over the Tibetan PlateauEarth-Science Reviews ,185, 308 -324 . DOI:
10.1016/j.earscirev.2018.06.012 URL [本文引用: 1] [15] Wang Y Sun J Ye CC Zeng T 2021 ).Climatic factors drive the aboveground ecosystem functions of alpine grassland via soil microbial biomass nitrogen on the Qingzang PlateauChinese Journal of Plant Ecology ,45, 434 -443 . [本文引用: 1] [王毅 ,孙建 ,叶冲冲 ,曾涛 (2021 ).气候因子通过土壤微生物生物量氮促进青藏高原高寒草地地上生态系统功能植物生态学报 ,45, 434 -443 .] [本文引用: 1] [16] Wang ZW Wan SZ Jiang HM Hu Y Ma SQ Chen YC Lu XY 2021 ).Soil enzyme activities and their influencing factors among different alpine grasslands on the Qingzang PlateauChinese Journal of Plant Ecology ,45, 528 -538 . [本文引用: 1] [汪子微 ,万松泽 ,蒋洪毛 ,胡扬 ,马书琴 ,陈有超 ,鲁旭阳 (2021 ).青藏高原不同高寒草地类型土壤酶活性及其影响因子植物生态学报 ,45, 528 -538 .] [本文引用: 1] [17] Wu QB Zhang TJ 2010 ).Changes in active layer thickness over the Qinghai-Tibetan Plateau from 1995 to 2007Journal of Geophysical Research ,115, D09107 . DOI:10.1029/2009JD012974 .DOI:
10.1029/2009JD012974 [本文引用: 1] [18] Xiang X Huang YM Yang CY Li ZQ Chen HY Pan YP Huo JX Ren L 2021 ).Effect of altitude on community-level plant functional traits in the Qinghai Lake Basin, ChinaChinese Journal of Plant Ecolog y456 -466 . [本文引用: 1] [向响 ,黄永梅 ,杨崇曜 ,李泽卿 ,陈慧颖 ,潘莹萍 ,霍佳璇 ,任梁 (2021 ).海拔对青海湖流域群落水平植物功能性状的影响植物生态学报 ,45, 456 -466 .] [本文引用: 1] [19] Xue PF Li WL Zhu GF Zhou HK Liu CL Yan HP 2021 ).Changes in the pattern of an alpine wetland landscape in Maqu County in the first meander of the Yellow RiverChinese Journal of Plant Ecolog y467 -475 . [本文引用: 1] [薛鹏飞 ,李文龙 ,朱高峰 ,周华坤 ,刘陈立 ,晏和飘 (2021 ).黄河首曲玛曲县高寒湿地景观格局演变植物生态学报 ,45, 467 -475 .] [本文引用: 1] [20] Yao TD Chen FH Cui P Ma YM Xu BQ Zhu LP Zhang F Wang WC Ai LK Yang XX 2017 ).From Tibetan Plateau to Third Pole and Pan-Third PoleBulletin of Chinese Academy of Sciences ,32, 924 -931 . [本文引用: 1] [姚檀栋 ,陈发虎 ,崔鹏 ,马耀明 ,徐柏青 ,朱立平 ,张凡 ,王伟财 ,艾丽坤 ,杨晓新 (2017 ).从青藏高原到第三极和泛第三极中国科学院院刊 ,32, 924 -931 .] [本文引用: 1] [21] Zong N Shi PL Zhao GS Zheng LL Niu B Zhou TC Hou G 2021 ).Variations of nitrogen and phosphorus limitation along the environmental gradient in alpine grasslands on the Northern Xizang PlateauChinese Journal of Plant Ecolog y444 -455 . [本文引用: 1] [宗宁 ,石培礼 ,赵广帅 ,郑莉莉 ,牛犇 ,周天财 ,侯阁 (2021 ).降水量变化对藏北高寒草地养分限制的影响植物生态学报 ,45, 444 -455 .] [本文引用: 1] 基于物候相机归一化植被指数估算高寒草地植物地上生物量的季节动态
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
基于物候相机归一化植被指数估算高寒草地植物地上生物量的季节动态
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
Climate Change and Biogeography 1
2017
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
西昆仑、西喀喇昆仑和西北喜马拉雅地区植被的地域分异及其指示意义
1
2002
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
西昆仑、西喀喇昆仑和西北喜马拉雅地区植被的地域分异及其指示意义
1
2002
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
青藏高原禾草混播对土壤微生物多样性的影响
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
青藏高原禾草混播对土壤微生物多样性的影响
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
高原鼠兔干扰对高寒草甸β多样性的影响
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
高原鼠兔干扰对高寒草甸β多样性的影响
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
青藏高原人工草地土壤可溶性氮组分与植被生产力动态变化过程
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
青藏高原人工草地土壤可溶性氮组分与植被生产力动态变化过程
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
若尔盖退化泥炭地土壤原核微生物群落结构对水位恢复的短期响应
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
若尔盖退化泥炭地土壤原核微生物群落结构对水位恢复的短期响应
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
藏北高寒草地土壤有机质化学组成对土壤蛋白酶和脲酶活性的影响,
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
藏北高寒草地土壤有机质化学组成对土壤蛋白酶和脲酶活性的影响,
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
Optimizing grazing exclusion practices to achieve Goal 15 of the sustainable development goals in the Tibetan Plateau
1
2021
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
Reconsidering the efficiency of grazing exclusion using fences on the Tibetan Plateau
1
2020
... 目前, 青藏高原整体呈变暖趋势, 导致青藏高原冰川融化, 湖泊增加, 湖泊总数由1990年的868个增加到2015年的1 207个, 湖泊总水面面积从38 823.3 km
2 增加到48 793.0 km
2 (
Sun et al ., 2018 ).此外, 持续增加的冻土活动层厚度和大气氮沉降速率(
Wu & Zhang, 2010 ), 以及不断增加的人类活动, 如围栏和放牧等生态系统管理措施也在改变和修饰着高寒生态系统(
Sun et al ., 2020 ).这些全球变化的主要表现形式必将对高寒植被地理格局和过程产生深刻的影响, 理解其中的功能变化和过程机制, 有助于我们深入理解高寒草地的分布变化和发展趋势, 以推进青藏高原生态安全屏障建设, 支撑高原可持续发展, 这对国家生态文明建设等方面具有重要的参考价值.值得深度关注的是, 大量的研究都考虑自然过程, 忽略了无处不在的放牧利用方式和基本覆盖全域的围栏管理措施带来的影响, 导致真实的格局、过程和机制很难被深刻地理解或阐明.因此建议未来高寒草地格局与过程研究, 应该综合考虑人类活动要素. ...
青藏高原高寒草地地上植物碳积累速率对生态系统多功能性的影响机制
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
青藏高原高寒草地地上植物碳积累速率对生态系统多功能性的影响机制
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
青藏高原高寒草地生态系统的适应性管理
1
2019
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
青藏高原高寒草地生态系统的适应性管理
1
2019
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
Linkages of the dynamics of glaciers and lakes with the climate elements over the Tibetan Plateau
1
2018
... 目前, 青藏高原整体呈变暖趋势, 导致青藏高原冰川融化, 湖泊增加, 湖泊总数由1990年的868个增加到2015年的1 207个, 湖泊总水面面积从38 823.3 km
2 增加到48 793.0 km
2 (
Sun et al ., 2018 ).此外, 持续增加的冻土活动层厚度和大气氮沉降速率(
Wu & Zhang, 2010 ), 以及不断增加的人类活动, 如围栏和放牧等生态系统管理措施也在改变和修饰着高寒生态系统(
Sun et al ., 2020 ).这些全球变化的主要表现形式必将对高寒植被地理格局和过程产生深刻的影响, 理解其中的功能变化和过程机制, 有助于我们深入理解高寒草地的分布变化和发展趋势, 以推进青藏高原生态安全屏障建设, 支撑高原可持续发展, 这对国家生态文明建设等方面具有重要的参考价值.值得深度关注的是, 大量的研究都考虑自然过程, 忽略了无处不在的放牧利用方式和基本覆盖全域的围栏管理措施带来的影响, 导致真实的格局、过程和机制很难被深刻地理解或阐明.因此建议未来高寒草地格局与过程研究, 应该综合考虑人类活动要素. ...
气候因子通过土壤微生物生物量氮促进青藏高原高寒草地地上生态系统功能
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
气候因子通过土壤微生物生物量氮促进青藏高原高寒草地地上生态系统功能
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
青藏高原不同高寒草地类型土壤酶活性及其影响因子
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
青藏高原不同高寒草地类型土壤酶活性及其影响因子
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
Changes in active layer thickness over the Qinghai-Tibetan Plateau from 1995 to 2007
1
2010
... 目前, 青藏高原整体呈变暖趋势, 导致青藏高原冰川融化, 湖泊增加, 湖泊总数由1990年的868个增加到2015年的1 207个, 湖泊总水面面积从38 823.3 km
2 增加到48 793.0 km
2 (
Sun et al ., 2018 ).此外, 持续增加的冻土活动层厚度和大气氮沉降速率(
Wu & Zhang, 2010 ), 以及不断增加的人类活动, 如围栏和放牧等生态系统管理措施也在改变和修饰着高寒生态系统(
Sun et al ., 2020 ).这些全球变化的主要表现形式必将对高寒植被地理格局和过程产生深刻的影响, 理解其中的功能变化和过程机制, 有助于我们深入理解高寒草地的分布变化和发展趋势, 以推进青藏高原生态安全屏障建设, 支撑高原可持续发展, 这对国家生态文明建设等方面具有重要的参考价值.值得深度关注的是, 大量的研究都考虑自然过程, 忽略了无处不在的放牧利用方式和基本覆盖全域的围栏管理措施带来的影响, 导致真实的格局、过程和机制很难被深刻地理解或阐明.因此建议未来高寒草地格局与过程研究, 应该综合考虑人类活动要素. ...
海拔对青海湖流域群落水平植物功能性状的影响
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
海拔对青海湖流域群落水平植物功能性状的影响
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
黄河首曲玛曲县高寒湿地景观格局演变
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
黄河首曲玛曲县高寒湿地景观格局演变
1
2021
... 针对退化草地,
董利军等(2021) 开展了若尔盖湿地高寒草甸退化过程中土壤有机碳含量变化及成因分析, 认为土壤含水量的降低导致的植物碳潜在输入量的降低是若尔盖湿地高寒草甸退化过程中土壤有机碳含量下降的主要原因.
罗明没等(2021) 通过野外原位试验和水位恢复梯度的设置, 揭示了短期水位恢复并没有改变原核微生物的α多样性, 土壤pH、有机碳、全氮等控制了退化泥炭地原核微生物群落结构随短期水位恢复过程中的变异, 该研究在一定程度上丰富了原核微生物群落结构对短期水位响应的认识.
李捷等(2021) 尝试采用β多样性揭示植物群落随小型啮齿草食动物干扰梯度变化的生态过程, 确立了冰草(
Agropyron cristatum )、臭蒿(
Artemisia hedinii )和小花草玉梅(
Anemone rivularis var.
flore-minore )对保护高原鼠兔干扰下高寒草甸植物群落多样性的关键地位.针对人工草地,
姜鑫和牛克昌(2021) 研究了青藏高原禾草混播对土壤微生物多样性的影响, 认为在野外条件下高寒草地禾草混播并不增加土壤微生物多样性.而
刘攀等(2021) 筛选出适应高寒生境的草种, 认为土壤营养是维持高寒人工草地生产力和稳定性的根本.此外, 本专辑还收录了景观格局演变(
薛鹏飞等, 2021 )和基于物候相机归一化植被指数(
NDVI )估算高寒草地植物地上生物量的季节动态(
陈哲等, 2021 )的研究文章, 旨在展示青藏高原高寒草地格局和过程研究的新的方法和技术手段. ...
从青藏高原到第三极和泛第三极
1
2017
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
从青藏高原到第三极和泛第三极
1
2017
... 随着全球变化问题的凸显, 如全球温度升高、人类活动加剧、极端气候频发、大气成分改变以及海洋水质恶化等, 越来越多的研究者开始关注气候变化对全球生物地理格局和过程的影响(
Goldblum & Carrier, 2017 ).青藏高原是世界上海拔最高、面积最大的高原, 约占中国陆地国土总面积的26%, 平均海拔4 000 m以上, 被称为“世界屋脊” (
孙建等, 2019 ), 是高寒草地格局与过程研究的天然实验室, 具有典型的水平地带性.喜马拉雅山、喀喇昆仑山和昆仑山, 阻挡了西风对青藏高原的水汽输送, 在高原“抽气系统”的影响下, 印度季风和东亚季风带来的水汽被源源不断地抽提到高海拔的青藏高原, 从第一阶梯开始, 水汽不断地消耗, 因此在高原面上, 形成了从东南到西北递减的降水梯度(
郭柯和郑度, 2002 ).受降水梯度的长期影响, 植被类型分异明显, 形成了森林、灌丛、高寒草甸、高寒草原和高寒荒漠草原的植被地理格局(
图1 ).高寒草地作为青藏高原主要的生态系统, 对畜牧业发展起到重要的支撑作用(
Sun et al ., 2021 ), 发挥着极其重要的气候调节和水源涵养等生态系统功能, 是我国重要的战略资源储备要地和生态安全屏障(
姚檀栋等, 2017 ).由于青藏高原高寒、生长周期短和土壤贫瘠的特点, 高寒草地对环境的变化极为敏感. ...
降水量变化对藏北高寒草地养分限制的影响
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
降水量变化对藏北高寒草地养分限制的影响
1
2021
... 在多功能性方面, 基于青藏高原地区115个样点进行了草地群落结构和土壤属性等要素的样带调查, 综合植物地上生物量, 叶片碳、氮和磷含量等参数计算多功能性, 讨论了青藏高原高寒草地地上生态系统功能与环境要素的关系(
王毅等, 2021 ), 并针对碳积累速率对生态系统多功能性开展了进一步剖析(
孙建等, 2021 ), 发现土壤微生物生物量氮和群落地上碳积累速率在调控高寒草地生态系统多功能方面发挥重要作用.此外,
宗宁等(2021) 通过在藏北降水梯度下高寒草甸、高寒草甸草原、高寒草原和高寒荒漠草原的氮磷养分添加试验, 发现随着降水量降低, 高寒草地养分限制模式从氮限制逐渐过渡到氮磷共同限制.颇具实际生产指导的意义在于, 利用养分添加恢复不同类型退化高寒草地时, 应将氮磷限制模式的差异考虑进去.此外, 也开展了不同草地类型土壤酶活性格局与过程等相关研究(
汪子微等, 2021 ),
马书琴等(2021) 研究发现不同高寒草地类型间碳循环酶、磷循环酶和两种氮循环酶(芳香氨基酶和亚硝酸盐还原酶)活性差异显著, 且土壤有机质、微生物数量和氮含量等是影响高寒草地生态系统土壤酶活性的关键因子.该研究在国内较早地开展热裂解气质联用技术(Py-GC/MS)分析, 并比较不同类型草地土壤有机质化学组成, 发现高寒草地类型和土壤有机质化学是影响高寒草地土壤蛋白酶活性的重要因素, 而对土壤脲酶活性的影响均未达到显著水平.海拔梯度上,
向响等(2021) 研究了青海湖流域群落水平植物功能性状的影响, 认为随海拔变化的热量和深层土壤养分含量是群落水平植物功能性状变化的主要影响因子, 特别是针对热量的理解, 可为今后相关研究提供理论参考. ...
 ,
, ,1,2,3,*, 刘国华2,3
,1,2,3,*, 刘国华2,3
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT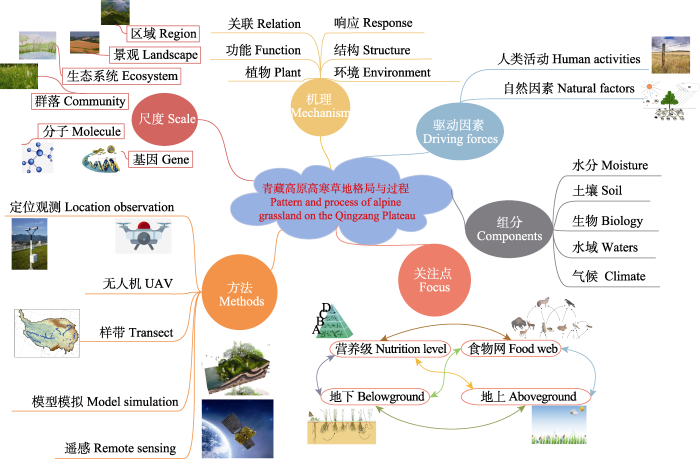 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}