Climatic factors drive the aboveground ecosystem functions of alpine grassland via soil microbial biomass nitrogen on the Qingzang Plateau
Yi WANG1,2, Jian SUN,,2,*, Chong-Chong YE2,3, Tao ZENG,1,*1College of Earth Sciences, Chengdu University of Technology, Chengdu 610059, China 2Key Laboratory of Observation and Simulation of Ecological Networks, Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 3School of Civil Engineering and Architecture, Southwest Petroleum University, Chengdu 610500, China
Abstract Aims In recent years, under the background of climate change and human activities, the trend of biodiversity loss is increasing. Such accelerated loss in biodiversity could bring serious consequences to ecosystem functions. At present, the research on ecosystem function ignores the important driving role of carbon and nitrogen cycling in soil and microorganism on the above ground ecosystem functions. Any changes of soil carbon, nitrogen and microorganism may affect the ability of belowground community, which can have substantial effects on the aboveground ecosystem functions. Our aim was to explore the driving factors and key mechanism of abovegroud ecosystem functions (AEF) in alpine grassland. Methods From July to August 2015, we conducted a transect survey in alpine grasslands to measure plant community and soil properties across Qingzang Plateau. There were in total 115 sample sites. The aboveground ecosystem function was calculated based on the aboveground biomass, leaf carbon, leaf nitrogen and leaf phosphorus. The effects of key elements such as soil organic carbon, total nitrogen and biomass on the aboveground ecosystem function were analyzed. Combined with mean annual precipitation and air temperature, we explored important drivers of AEF and related mechanisms. Important findings Precipitation has a greater impact on aboveground ecosystem functions, while air temperature has a minor impact. Mean annual precipitation, soil microbial nitrogen content and aridity index had relative higher importance to aboveground ecosystem functions. Specificially, mean annual precipitation, soil microbial nitrogen content and aridity index accounted for the variations of 21.1%, 10.9% and 10.1%, respectively. The findings indicated that soil properties might play more important roles than plant community and productivity to aboveground ecosystem functions. Considering the cascading impacts of climate factors on soil nutrients cycling and microorganisms, soil microbial biomass nitrogen content plays an important role in regulating AEF of alpine grassland, Qingzang Plateau. Keywords:aboveground ecosystem function;alpine grassland;soil total nitrogen;soil microbial biomass nitrogen;Qingzang Plateau
PDF (5659KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 王毅, 孙建, 叶冲冲, 曾涛. 气候因子通过土壤微生物生物量氮促进青藏高原高寒草地地上生态系统功能. 植物生态学报, 2021, 45(5): 434-443. DOI: 10.17521/cjpe.2020.0204 WANG Yi, SUN Jian, YE Chong-Chong, ZENG Tao. Climatic factors drive the aboveground ecosystem functions of alpine grassland via soil microbial biomass nitrogen on the Qingzang Plateau. Chinese Journal of Plant Ecology, 2021, 45(5): 434-443. DOI: 10.17521/cjpe.2020.0204
高寒草地是青藏高原最主要的生态系统, 在气候调节、水源涵养和碳氮固定等方面发挥着重要的生态功能(杨元合和朴世龙, 2006; 孙鸿烈等, 2012; Sun et al., 2020)。青藏高原独特的高海拔特征、寒冷的气候条件和严苛的植物生长环境导致高寒草地生态系统非常脆弱, 生态系统功能对环境变化非常敏感(Jing et al., 2015; 熊定鹏等, 2016)。在地上群落生物多样性和EMF的关系格局方面, 研究发现, 降水量显著影响物种丰富度和EMF, 但年降水量并未显著影响物种多样性指数, 强调了群落物种丰富度对维持EMF重要意义, 这意味着物种的丧失可能会给高寒草地EMF和生态服务带来更为严重的后果(熊定鹏等, 2016)。EMF与地上和地下生物多样性关系方面, 研究发现二者的联系是由气候条件决定的(Jing et al., 2015)。作为对气候条件十分敏感的青藏高原, 目前尚缺乏降水梯度下地下土壤关键因子对高寒草地生态系统地上功能影响的主要控制因子和机理的研究。因此, 本研究选择青藏高原地区, 基于样带上115个样点调查分析, 综合运用13种生态系统功能参数和3个气候因子, 试图回答以下两个科学问题: (1)驱动高寒草地AEF的主要因子是什么? (2)这种因子是如何作用于AEF的? 以期为高寒草地生态系统多样性保护、多功能性维持和可持续利用管理提供科学依据。
1 材料和方法
1.1 试验设计
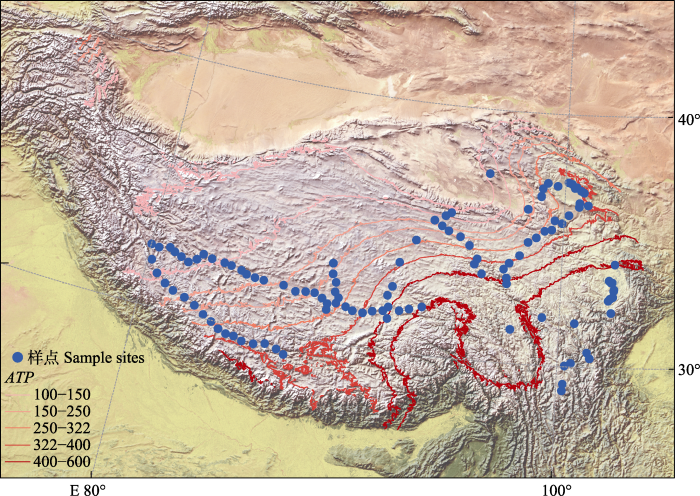
2015年7-8月, 在青藏高原进行试验取样, 覆盖了四川、青海、甘肃和西藏地区(80°-105° E, 27°-37° N, 海拔3 030-5 000 m), 共计115个研究样点(图1)。该地区主要植被类型是高寒草甸和高寒草原, 主要土壤类型有毛毡土、冷钙质土和棕色钙质土(Sun et al., 2020)。每个样地大小10 m × 10 m, 在每个样地随机放置3个样方框(50 cm × 50 cm)取样调查。分种记录每个样方的植物种类、高度和盖度, 然后与地表平齐采集样方内植物地上部分, 用信封装好, 以获得地上生物量数据。用直径4 cm的土钻在各个小样方内采集土壤(0-10 cm土层)样品, 用于测定地下生物量和土壤养分。以上样品采集后, 均严格按照标准处理方式保存并带回实验室进行测定。
2015年平均气温(AMT)和年降水量(ATP)数据来源于中国气象数据网(http://data.cma.cn/), 依据1 km分辨率的栅格数据, 使用Anusplin 4.2 (Centre for Resource and 220 Environmental Studies, Australian National University, Canberra, Australia)进行空间插值。利用ArcGIS 10.2软件从数据库中提取研究区115个样点的气候数据。干旱指数(AI)(Sun et al., 2020)计算如下:
$AI = \frac{{ATP}}{{AMT + 10}}$
1.4 生态系统多功能性
选择与植物生长(AGB)(熊定鹏等, 2016)和植物营养(LC、LN和LP)(Jing et al., 2015)密切相关的重要生态系统参数来表征AEF (表1)。为量化AEF, 采用生态系统多功能性(EMF)的量化方式(Maestre et al., 2012)。首先计算115个调查样地4种生态系统参数的Z分数, 单样本Kolmogorov-Smirnov检验结果表明LN和AEF值服从正态分布(p > 0.05), 其余参数均不服从正态分布。随后, 在计算Z分数之前先进行归一化处理。Z分数计算公式如下:
${Z_{ij}} = ({x_{ij}} - {\lambda _j})/{\delta _j}$ Table 1 表1 表1青藏高原样带调查各指标描述性统计 Table 1Descriptive statistics of each index in the transect survey on the Qingzang Plateau
调查指标 Survey indicator
p
平均值 Mean
标准误 SE
最大值 Max
最小值 Min
地上生态系统功能 Aboveground ecosystem functions
0.200
0.05
0.54
1.77
-1.70
地上生物量 Aboveground biomass (g·m-2)
0.000
91.15
77.22
318.48
7.56
地下生物量 Belowground biomass (g·m-2)
0.000
3.20
6.16
42.53
0.01
叶片碳含量 Leaf carbon content (g·kg-1)
0.000
40.59
3.75
45.12
27.42
叶片磷含量 Leaf phosphorus content (g·kg-1)
0.001
1.67
0.79
5.32
0.34
叶片氮含量 Leaf nitrogen content (g·kg-1)
0.098
17.98
4.34
34.44
5.90
土壤含水量 Soil water content (%)
0.000
0.15
0.13
0.53
0.01
土壤速效磷含量 Soil available phosphorus content (mg·kg-1)
Fig. 2Relationships between climate factors, belowground biomass, soil factors and aboveground ecosystem functions (AEF). A, The correlation between AEF value and each factor; black box indicates positive correlation, and red box indicates negative correlation, * and ** represented significantly correlated with AEF value (p < 0.05 and p < 0.01). B, Principal component analysis of AEF value and each factor. AI, aridity index; AMT, mean annual air temperature; ATP, mean annual precipitation; BGB, belowground biomass; EMF, ecosystem multi-function; MBC, microbial biomass carbon content; MBN, microbial biomass nitrogen content; SAN, soil available nitrogen content; SAP, soil available phosphorus content; SOC, soil organic carbon content; STN, soil total nitrogen content; STP, soil total phosphorus content.
2.2 气候和土壤因子对AEF的作用
ATP、MBC和AI对AEF值的相对重要性贡献较高(重要性分别为21.1%、10.9%和10.1%)(图3), 综合相关性结果, 对气候因子和土壤碳氮要素进行分析。ATP、AI、STN和MBN与AEF值均呈现显著线性正相关关系(p < 0.05)。而且, AEF值对ATP (R2 = 0.09, k = 0.74)和AI (R2 = 0.05, k = 0.51)有较高的敏感性, 对MBN (R2 = 0.05, k = 0.27)敏感性相对较低(图4)。ATP和SOC、STN、MBC和MBN均呈显著线性正相关关系(p < 0.01), 尤其是ATP与MBC和MBN拟合优度和斜率较高(R2 = 0.54, k = 1.87; R2 = 0.53, k = 1.53)。同样, AI和SOC、STN、MBC和MBN呈显著线性正相关关系(p < 0.01), 尤其是AI与MBC和MBN拟合优度和斜率较高(R2 = 0.52, k = 1.76; R2 = 0.54, k = 1.46)(图5)。
Fig. 6Effects of climatic and soil factors on aboveground ecosystem functions value (AEF). The path with significant effect is shown in the figure (p < 0.05), the solid lines indicate a positive effect and the dotted line indicates a positive effect. AI, aridity index; ATP, mean annual precipitation; MBC, microbial biomass carbon content; MBN, microbial biomass nitrogen content; SOC, soil organic carbon content; STN, soil total nitrogen content.
3 讨论
3.1 气候因子对AEF的影响
研究发现, 年平均气温与AEF值之间没有显著关系(图2), 这与Jing等(2015)关于年平均气温对生态系统功能的影响不显著的研究结果一致。这可能是因为在高寒草地生态系统中昼夜温差很大, 类似于季节性变化(Baumann et al., 2009)。实际上, 气候条件对青藏高原高寒草地生态系统植被群落结构和物种丰富度的空间分布格局的形成至关重要(Sun et al., 2020)。研究区2015年年降水量从东南至西北由860 mm递减至143 mm, 水分变化是本研究中影响AEF值的重要因素, 二者之间具有正相关关系(图2, 图4)。这一结果与羌塘高寒草地和对整个青藏高原高寒草地样带物种多样性和降水量的研究结果(杨元合等, 2004; 熊定鹏等, 2016)相符。一方面, 降水量的增加能够促进生态系统对碳的积累, 而且间接地促进土壤养分的矿化和积累, 从而在本研究结果中观察到土壤碳氮含量随降水量的增加呈现显著增加的趋势(图5)(Niu et al., 2007)。另一方面, 本研究中降水量和干旱指数与AEF值、土壤有机碳、土壤全氮、微生物生物量碳和氮含量等养分循环相关的参数呈极显著的正相关关系(图5), 这些结果表明气候因子, 特别是水分条件是决定青藏高原高寒草地AEF的重要限制因子。此外, 也有研究指出, 在高寒草地生态系统, 气候调节土壤肥力和微生物的分解功能, 连接着物种多样性和EMF之间的关系(Jing et al., 2015)。有研究证明, 气候调节和土壤肥力对陆地生态系统微生物的分解功能产生重要影响, 微生物多样性的任何损失都可能降低多功能性(Delgado-Baquerizo et al., 2016), 这些都有助于解释我们的结果。
3.2 土壤因子对AEF的影响
本研究发现年降水量、干旱指数和土壤微生物生物量氮含量对AEF值有着较大的贡献(图3)。有研究指出物种丰富度对EMF的影响极为关键, 是干旱生态系统多功能性的主要驱动力(Maestre et al., 2012)。综合92个试验研究发现, 生物多样性能够增强生态系统在营养和生境水平中的多功能性(Lefcheck et al., 2015)。我们的结果与前人的研究并不矛盾, 因为气候和物种丰富度维持碳氮循环, 而碳氮循环维持着碳氮的固定和土壤肥力相关的生态系统功能(王淑平等, 2002; Whitford, 2002)。一般来说, 物种丰富度的增加通常通过提高地上、地下生产力和根系凋落物碳库来实现土壤碳累积, 而且, 物种多样性能够通过改变凋落物质量以及土壤的生物和非生物属性来影响土壤有机质分解, 从而影响EMF, 这是一个协同促进的过程(Cong et al., 2015)。本研究结果表明, 土壤养分和微生物对地上生态系统功能有促进作用, 这与Jing等(2015)在青藏高原的研究结果一致, 这对草地生态系统物种多样性保护的认识具有重要意义。
BaiJB,XuXL,FuG,SongMH,HeYT,JiangJ(2011).Effects of temperature and nitrogen input on nitrogen mineralization in alpine soils on Tibetan Plateau .Agricultural Science & Technology,12, 1909-1912. [本文引用: 1]
BaoSD(2000).Soil and Agricultural Chemistry Analysis.China Agriculture Press,Beijing. [本文引用: 1]
BardgettRD,van der PuttenWH(2014).Belowground biodiversity and ecosystem functioning .Nature,515, 505-511. DOI:10.1038/nature13855URL [本文引用: 1]
BaumannF,HeJ,SchmidtK,KühnP,ScholtenT(2009).Pedogenesis, permafrost, and soil moisture as controlling factors for soil nitrogen and carbon contents across the Tibetan Plateau .Global Change Biology,15, 3001-3017. DOI:10.1111/j.1365-2486.2009.01953.xURL [本文引用: 1]
BradfordMA,WoodSA,BardgettRD,BlackHIJ,BonkowskiM,EggersT,GraystonSJ,KandelerE,ManningP,SetäläH,JonesTH(2014).Discontinuity in the responses of ecosystem processes and multifunctionality to altered soil community composition .Proceedings of the National Academy of Sciences of the United States of America,111, 14478-14483. [本文引用: 1]
ChenQL,DingJ,ZhuD,HuHW,Delgado-BaquerizoM,MaYB,HeJZ,ZhuYG(2019).Rare microbial taxa as the major drivers of ecosystem multifunctionality in long-term fertilized soils .Soil Biology & Biochemistry,141, 107686. DOI:10.1016/j.soilbio.2019.107686. DOI:10.1016/j.soilbio.2019.107686URL [本文引用: 1]
CongWF,van RuijvenJ,van der WerfW,de DeynGB,MommerL,BerendseF,HofflandE(2015).Plant species richness leaves a legacy of enhanced root litter-induced decomposition in soil .Soil Biology & Biochemistry,80, 341-348. DOI:10.1016/j.soilbio.2014.10.017URL [本文引用: 1]
Delgado-BaquerizoM,MaestreFT,ReichPB,JeffriesTC,GaitanJJ,EncinarD,BerdugoM,CampbellCD,SinghBK(2016).Microbial diversity drives multifunctionality in terrestrial ecosystems .Nature Communications,7, 10541. DOI:10.1038/ncomms10541. DOI:10.1038/ncomms10541PMID:26817514 [本文引用: 2] Despite the importance of microbial communities for ecosystem services and human welfare, the relationship between microbial diversity and multiple ecosystem functions and services (that is, multifunctionality) at the global scale has yet to be evaluated. Here we use two independent, large-scale databases with contrasting geographic coverage (from 78 global drylands and from 179 locations across Scotland, respectively), and report that soil microbial diversity positively relates to multifunctionality in terrestrial ecosystems. The direct positive effects of microbial diversity were maintained even when accounting simultaneously for multiple multifunctionality drivers (climate, soil abiotic factors and spatial predictors). Our findings provide empirical evidence that any loss in microbial diversity will likely reduce multifunctionality, negatively impacting the provision of services such as climate regulation, soil fertility and food and fibre production by terrestrial ecosystems.
FriedmanJH,PopescuBE(2008).Predictive learning via rule ensembles .The Annals of Applied Statistics,2, 916-954. [本文引用: 1]
FuYW,TianDS,WangJS,NiuSL,ZhaoKT(2019).Patterns and affecting factors of nitrogen use efficiency of plant leaves and roots in Nei Mongol and Qinghai-Xizang Plateau grasslands .Chinese Journal of Plant Ecology,43, 566-575. DOI:10.17521/cjpe.2019.0044URL [本文引用: 1]
GansJD,WolinskyM,DunbarJ(2005).Computational improvements reveal great bacterial diversity and high metal toxicity in soil .Science,309, 1387-1390. DOI:10.1126/science.1112665URL [本文引用: 1]
HooperDU,BignellDE,BrownVK,BrussardL,Mark DangerfieldJ,WallDH,WardleDA,ColemanDC,GillerKE,LavelleP,van der PuttenWH,de RuiterPC,RusekJ,SilverWL,TiedjeJM,WoltersV(2000).Interactions between aboveground and belowground biodiversity in terrestrial ecosystems: patterns, mechanisms, and feedbacks .BioScience,50, 1049-1061. DOI:10.1641/0006-3568(2000)050[1049:IBAABB]2.0.CO;2URL [本文引用: 1]
HooperDU,ChapinFS,EwelJJ,HectorA,InchaustiP,LavorelS,LawtonJH,LodgeDM,LoreauM,NaeemS,SchmidB,SetäläH,SymstadAJ,VandermeerJ,WardleDA(2005).Effects of biodiversity on ecosystem functioning: a consensus of current knowledge .Ecological Monographs,75, 3-35. DOI:10.1890/04-0922URL [本文引用: 1]
JingX,SandersNJ,ShiY,ChuHY,ClassenAT,ZhaoK,ChenLT,ShiY,JiangYX,HeJS(2015).The links between ecosystem multifunctionality and above- and belowground biodiversity are mediated by climate .Nature Communications,6, 8159. DOI:10.1038/ncomms9159. DOI:10.1038/ncomms9159PMID:26328906 [本文引用: 9] Jing, Xin; Zhao, Ke; Shi, Yue; He, Jin-Sheng Peking Univ, Dept Ecol, Coll Urban & Environm Sci, Beijing 100871, Peoples R China. Jing, Xin; Zhao, Ke; Shi, Yue; He, Jin-Sheng Peking Univ, Key Lab Earth Surface Proc, Minist Educ, Beijing 100871, Peoples R China. Sanders, Nathan J. Univ Copenhagen, Ctr Macroecol Evolut & Climate, Nat Hist Museum Denmark, DK-2100 Copenhagen, Denmark. Shi, Yu; Chu, Haiyan Chinese Acad Sci, Inst Soil Sci, State Key Lab Soil & Sustainable Agr, Nanjing 210008, Peoples R China. Classen, Aimee T. Univ Copenhagen, Nat Hist Museum Denmark, DK-2100 Copenhagen, Denmark. Chen, Litong; He, Jin-Sheng Chinese Acad Sci, Northwest Inst Plateau Biol, Key Lab Adaptat & Evolut Plateau Biota, Xining 810008, Peoples R China. Shi, Yue Chinese Acad Sci, Inst Bot, Beijing 100093, Peoples R China. Jiang, Youxu Chinese Acad Forestry, Inst Forest Ecol, Beijing 100091, Peoples R China.
LefcheckJS,ByrnesJEK,IsbellF,GamfeldtL,GriffinJN,EisenhauerN,HenselMJS,HectorA,CardinaleBJ,DuffyJE(2015).Biodiversity enhances ecosystem multifunctionality across trophic levels and habitats .Nature Communications,6, 6936. DOI:10.1038/ncomms7936. DOI:10.1038/ncomms7936URL [本文引用: 1]
LiG,WangLJ,LiYJ,QiaoJ,ZhangHF,SongXL,YangDL(2013).Effects of different vegetation restoration patterns on the diversity of soil nitrogen-fixing microbes in Hulunbeier sandy land, Inner Mongolia of North China .Chinese Journal of Applied Ecology,24, 1639-1646. [本文引用: 1]
LiL,GaoJQ,LeiGC,LüC,SuoL(2011).Distribution patterns of soil organic carbon and total nitrogen in Zoige peat land with different ground water table .Chinese Journal of Ecology,30, 2449-2455. [本文引用: 2]
LuoYQ,SuB,CurrieWS,DukesJS,FinziA,HartwigU,HungateB,Mc MurtrieRE,OrenR,PartonWJ,PatakiDE,ShawMR,ZakDR,FieldCB(2004).Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide .BioScience,54, 731-739. DOI:10.1641/0006-3568(2004)054[0731:PNLOER]2.0.CO;2URL [本文引用: 1]
MaestreFT,QueroJL,GotelliNJ,EscuderoA,OchoaV,Delgado-BaquerizoM,Garcia-GomezM,BowkerMA,SoliveresS,EscolarC,Garcia-PalaciosP,BerdugoM,ValenciaE,GozaloB,GallardoA,et al.(2012).Plant species richness and ecosystem multifunctionality in global drylands .Science,335, 214-218. DOI:10.1126/science.1215442URL [本文引用: 3]
MeyerST,PtacnikR,HillebrandH,BesslerH,BuchmannN,EbelingA,EisenhauerN,EngelsC,FischerM,HalleS,KleinAM,OelmannY,RoscherC,RottstockT,ScherberC,et al.(2018).Biodiversity-multifunctionality relationships depend on identity and number of measured functions .Nature Ecology & Evolution,2, 44-49. [本文引用: 1]
NiuSL,WuMY,HanY,XiaJY,LiLH,WanSQ(2007).Water-mediated responses of ecosystem carbon fluxes to climatic change in a temperate steppe .New Phytologist,177, 209-219. DOI:10.1111/nph.2008.177.issue-1URL [本文引用: 1]
ShipleyB,MezianeD(2002).The balanced-growth hypothesis and the allometry of leaf and root biomass allocation .Functional Ecology,16, 326-331. DOI:10.1046/j.1365-2435.2002.00626.xURL [本文引用: 1]
SunHL,ZhengD,YaoTD,ZhangYL(2012).Protection and construction of the national ecological security shelter zone on Tibetan Plateau .Acta Geographica Sinica,67, 3-12. [本文引用: 1]
SunJ,ZhouTC,LiuM,ChenYC,LiuGH,XuM,ShiPL,PengF,TsunekawaA,LiuY,WangXD,DongSK,ZhangYJ,LiYN(2020).Water and heat availability are drivers of the aboveground plant carbon accumulation rate in alpine grasslands on the Tibetan Plateau .Global Ecology and Biogeography,29, 50-64. DOI:10.1111/geb.v29.1URL [本文引用: 5]
van der HeijdenMGA,KlironomosJN,UrsicM,MoutoglisP,Streitwolf-EngelR,BollerT,WiemkenA,SandersIR(1998).Mycorrhizal fungal diversity determines plant biodiversity, ecosystem variability and productivity .Nature,396, 69-72. DOI:10.1038/23932URL [本文引用: 2]
WaggC,BenderSF,WidmerF,van der HeijdenMGA(2014).Soil biodiversity and soil community composition determine ecosystem multifunctionality .Proceedings of the National Academy of Sciences of the United States of America,111, 5266-5270. [本文引用: 2]
WangDL,WangL(2019).A new perspective on the concept of grassland management .Chinese Science Bulletin,64, 1106-1113. [本文引用: 1]
WangSP,ZhouGS,LüYC,ZouJJ(2002).Distribution of soil carbon, nitrogen and phosphorus along Northeast China Transect (NECT) and their relationships with climatic factors .Acta Phytoecologica Sinica,26, 513-517. [本文引用: 1]
WangX,XuZW,LüX,WangRZ,CaiJP,YangS,LiMH,JiangY(2017).Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland .Plant and Soil,418, 241-253. DOI:10.1007/s11104-017-3288-8URL [本文引用: 1]
WangY,LiuBY,LiuM,SunJ,ZengT(2019).Synergistic and inhibitory effects of soil enzymes along desertified gradients of the Zoige alpine meadow .Pratacultural Science,36, 939-951. [本文引用: 1]
WardleDA,BardgettRD,KlironomosJN,SetäläH,van der PuttenWH,WallDH(2004).Ecological linkages between aboveground and belowground biota .Science,304, 1629-1633. PMID:15192218 [本文引用: 1] All terrestrial ecosystems consist of aboveground and belowground components that interact to influence community- and ecosystem-level processes and properties. Here we show how these components are closely interlinked at the community level, reinforced by a greater degree of specificity between plants and soil organisms than has been previously supposed. As such, aboveground and belowground communities can be powerful mutual drivers, with both positive and negative feedbacks. A combined aboveground-belowground approach to community and ecosystem ecology is enhancing our understanding of the regulation and functional significance of biodiversity and of the environmental impacts of human-induced global change phenomena.
WhitfordWG(2002).Ecology of Desert Systems.Academic Press, San Diego,USA. [本文引用: 1]
XiongDP,ZhaoGS,WuJS,ShiPL,ZhangXZ(2016).The relationship between species diversity and ecosystem multifunctionality in alpine grasslands on the Tibetan Changtang Plateau .Acta Ecologica Sinica,36, 3362-3371. [本文引用: 4]
XuZW,LiMH,ZimmermannNE,LiSP,LiH,RenHY,SunH,HanXG,JiangY,JiangL(2018).Plant functional diversity modulates global environmental change effects on grassland productivity .Journal of Ecology,106, 1941-1951. DOI:10.1111/jec.2018.106.issue-5URL [本文引用: 1]
YanZQ,QiYC,PengQ,DongYS,HeYL,LiZL(2017).Advances in the effects of simulated precipitation and nitrogen deposition on grassland biomass .Acta Agrestia Sinica,25, 1165-1170. [本文引用: 2]
YangXX,RenF,ZhouHK,HeJS(2014).Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau .Chinese Journal of Plant Ecology,38, 159-166. DOI:10.3724/SP.J.1258.2014.00014URL [本文引用: 1]
YangYH,PiaoSL(2006).Variations in grassland vegetation cover in relation to climatic factors on the Tibetan Plateau .Journal of Plant Ecology (Chinese Version),30, 1-8. [本文引用: 1]
YangYH,RaoS,HuHF,ChenAP,JiCJ,ZhuB,ZuoWY,LiXR,ShenHH,WangZH,TangYH,FangJY(2004).Plant species richness of alpine grasslands in relation to environmental factors and biomass on the Tibetan Plateau .Chinese Biodiversity,12, 200-205. [本文引用: 1]
ZavaletaES,PasariJR,HulveyKB,TilmanGD(2010).Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity .Proceedings of the National Academy of Sciences of the United States of America,107, 1443-1446. DOI:10.1073/pnas.0906829107PMID:20080690 [本文引用: 1] Society places value on the multiple functions of ecosystems from soil fertility to erosion control to wildlife-carrying capacity, and these functions are potentially threatened by ongoing biodiversity losses. Recent empirically based models using individual species' traits suggest that higher species richness is required to provide multiple ecosystem functions. However, no study to date has analyzed the observed functionality of communities of interacting species over multiple temporal scales to assess the relationship between biodiversity and multifunctionality. We use data from the longest-running biodiversity-functioning field experiment to date to test how species diversity affects the ability of grassland ecosystems to provide threshold levels of up to eight ecosystem functions simultaneously. Across years and every combination of ecosystem functions, minimum-required species richness consistently increases with the number of functions considered. Moreover, tradeoffs between functions and variability among years prevent any one community type from providing high levels of multiple functions, regardless of its diversity. Sustained multifunctionality, therefore, likely requires both higher species richness than single ecosystem functionality and a diversity of species assemblages across the landscape. Effects of temperature and nitrogen input on nitrogen mineralization in alpine soils on Tibetan Plateau 1 2011
... 随着研究的深入, 研究者逐渐认识到地下群落对AEF的驱动作用, 地下部分是影响草地生态系统结构、功能和规律中最不确定的因素(王毅等, 2019).土壤生物群落也被视为调节生态系统功能的关键因子(van der Heijden et al., 1998), 因为土壤生物群落结构的简化会降低EMF (van der Heijden et al., 1998; Wagg et al., 2014; Jing et al., 2015).不同的生态类型, 影响其生态系统功能的主导因素可能不同, 例如: 在森林生态系统中, 物种丰富度和功能多样性对土壤养分有显著正效应, 功能多样性能很好地解释生态系统功能(Delgado-Baquerizo et al., 2017).在全球干旱生态系统中, 土壤微生物群落驱动旱地EMF对全球变化的抵抗力(Delgado-Baquerizo et al., 2017).由于草地生态系统普遍受到氮限制, 在全球氮富集的背景下, 土壤氮可能成为影响和驱动EMF的关键(符义稳等, 2019).有研究发现, 随着全球气候变暖, CO2浓度增高, 土壤中不稳定碳的含量增加, 这些不稳定碳通过根系分泌物以及根的转化来增加土壤微生物对氮的需求, 从而增加植物和土壤微生物对可利用氮的竞争, 导致生态系统过程的氮限制, 降低生态系统功能(Luo et al., 2004).在草地生态系统中, 氮对植物生产力、碳储量和养分周转有正效应, 土壤水分和氮有效性增加可以提高植物群落的生产力, 进而提高地下功能复杂性(Bradford et al., 2014; 杨晓霞等, 2014; Wang et al., 2017).此外, 对青藏高原高寒草地的研究发现, 输入的无机氮被微生物固定, 能够显著促进土壤氮矿化速率, 从而促使原来有机氮的矿化和释放, 对生态系统功能产生非常积极的作用(Bai et al., 2011).土壤微生物的关键作用在于其能促进凋落物分解和有机物矿化等, 这些过程促进物质和能量在地上和地下群落之间转移, 在维持生态系统功能方面发挥着关键作用(Hooper et al., 2000; Wardle et al., 2004).一部分土壤微生物类群具有固定大气中氮的能力, 能将N2转变为植物可利用的形态, 这一类群的数量变化, 对土壤氮补充、平衡和调节, 以及对土壤结构、肥力以及生态系统功能至关重要(李刚等, 2013).此外, 研究发现, 在长期肥沃土壤环境下, 稀有微生物是生态系统多功能性的主要驱动力(Chen et al., 2019).因此, 全球气候变化导致土壤碳氮含量以及微生物任何变化都有可能改变地上和地下群落维持AEF的能力(Gans et al., 2005; Bardgett & van der Putten, 2014; Tedersoo et al., 2014). ...
... 在全球变化影响下, 生物多样性正在加速丧失, 生态系统结构不断简化, 生态系统服务和功能受到严重破坏(Hooper et al., 2005; Wagg et al., 2014; 孙建等, 2019).生物多样性与生态系统功能成为生态学领域研究的热点, 并且在全球范围内开展了广泛的研究(Jing et al., 2015; López-Rojo et al., 2019).其中, 影响生态系统功能的驱动因子成为关注的重点, 并且从不同研究层面都获得了一定的成果.物种层面, 通过分析不同时空尺度和环境条件, 在综合考虑了地上、地下生产力、群落抗入侵能力、植物氮、土壤碳氮要素等多种生态系统功能后, 发现维持生态系统多个功能比单个功能需要更多的物种, 这是因为生态系统多功能间的互补性, 整体功能比单个功能更易受物种丧失的影响(Zavaleta et al., 2010; Perkins et al., 2015; Meyer et al., 2018).群落层面, 随物种丰富度的增加, 维持生态系统多功能的概率也会增加, 然而物种丰富度并不是驱动生态系统多功能性(EMF)的唯一生物因素.非生物因子层面, 气候因子调节EMF与地上和地下生物多样性之间的联系, 区域尺度的气候变化可以改变生物多样性对EMF的影响(Jing et al., 2015).目前, 生态系统地下部分提供诸如碳氮养分循环等基础服务功能和碳氮养分对地上生态系统功能(AEF)潜在的作用尚需进一步明确. ...
青藏高原高寒草地生态系统的适应性管理 1 2019
... 在全球变化影响下, 生物多样性正在加速丧失, 生态系统结构不断简化, 生态系统服务和功能受到严重破坏(Hooper et al., 2005; Wagg et al., 2014; 孙建等, 2019).生物多样性与生态系统功能成为生态学领域研究的热点, 并且在全球范围内开展了广泛的研究(Jing et al., 2015; López-Rojo et al., 2019).其中, 影响生态系统功能的驱动因子成为关注的重点, 并且从不同研究层面都获得了一定的成果.物种层面, 通过分析不同时空尺度和环境条件, 在综合考虑了地上、地下生产力、群落抗入侵能力、植物氮、土壤碳氮要素等多种生态系统功能后, 发现维持生态系统多个功能比单个功能需要更多的物种, 这是因为生态系统多功能间的互补性, 整体功能比单个功能更易受物种丧失的影响(Zavaleta et al., 2010; Perkins et al., 2015; Meyer et al., 2018).群落层面, 随物种丰富度的增加, 维持生态系统多功能的概率也会增加, 然而物种丰富度并不是驱动生态系统多功能性(EMF)的唯一生物因素.非生物因子层面, 气候因子调节EMF与地上和地下生物多样性之间的联系, 区域尺度的气候变化可以改变生物多样性对EMF的影响(Jing et al., 2015).目前, 生态系统地下部分提供诸如碳氮养分循环等基础服务功能和碳氮养分对地上生态系统功能(AEF)潜在的作用尚需进一步明确. ...
Water and heat availability are drivers of the aboveground plant carbon accumulation rate in alpine grasslands on the Tibetan Plateau 5 2020
... 高寒草地是青藏高原最主要的生态系统, 在气候调节、水源涵养和碳氮固定等方面发挥着重要的生态功能(杨元合和朴世龙, 2006; 孙鸿烈等, 2012; Sun et al., 2020).青藏高原独特的高海拔特征、寒冷的气候条件和严苛的植物生长环境导致高寒草地生态系统非常脆弱, 生态系统功能对环境变化非常敏感(Jing et al., 2015; 熊定鹏等, 2016).在地上群落生物多样性和EMF的关系格局方面, 研究发现, 降水量显著影响物种丰富度和EMF, 但年降水量并未显著影响物种多样性指数, 强调了群落物种丰富度对维持EMF重要意义, 这意味着物种的丧失可能会给高寒草地EMF和生态服务带来更为严重的后果(熊定鹏等, 2016).EMF与地上和地下生物多样性关系方面, 研究发现二者的联系是由气候条件决定的(Jing et al., 2015).作为对气候条件十分敏感的青藏高原, 目前尚缺乏降水梯度下地下土壤关键因子对高寒草地生态系统地上功能影响的主要控制因子和机理的研究.因此, 本研究选择青藏高原地区, 基于样带上115个样点调查分析, 综合运用13种生态系统功能参数和3个气候因子, 试图回答以下两个科学问题: (1)驱动高寒草地AEF的主要因子是什么? (2)这种因子是如何作用于AEF的? 以期为高寒草地生态系统多样性保护、多功能性维持和可持续利用管理提供科学依据. ...
... 2015年7-8月, 在青藏高原进行试验取样, 覆盖了四川、青海、甘肃和西藏地区(80°-105° E, 27°-37° N, 海拔3 030-5 000 m), 共计115个研究样点(图1).该地区主要植被类型是高寒草甸和高寒草原, 主要土壤类型有毛毡土、冷钙质土和棕色钙质土(Sun et al., 2020).每个样地大小10 m × 10 m, 在每个样地随机放置3个样方框(50 cm × 50 cm)取样调查.分种记录每个样方的植物种类、高度和盖度, 然后与地表平齐采集样方内植物地上部分, 用信封装好, 以获得地上生物量数据.用直径4 cm的土钻在各个小样方内采集土壤(0-10 cm土层)样品, 用于测定地下生物量和土壤养分.以上样品采集后, 均严格按照标准处理方式保存并带回实验室进行测定. ...
... 2015年平均气温(AMT)和年降水量(ATP)数据来源于中国气象数据网(http://data.cma.cn/), 依据1 km分辨率的栅格数据, 使用Anusplin 4.2 (Centre for Resource and 220 Environmental Studies, Australian National University, Canberra, Australia)进行空间插值.利用ArcGIS 10.2软件从数据库中提取研究区115个样点的气候数据.干旱指数(AI)(Sun et al., 2020)计算如下: ...
... 研究发现, 年平均气温与AEF值之间没有显著关系(图2), 这与Jing等(2015)关于年平均气温对生态系统功能的影响不显著的研究结果一致.这可能是因为在高寒草地生态系统中昼夜温差很大, 类似于季节性变化(Baumann et al., 2009).实际上, 气候条件对青藏高原高寒草地生态系统植被群落结构和物种丰富度的空间分布格局的形成至关重要(Sun et al., 2020).研究区2015年年降水量从东南至西北由860 mm递减至143 mm, 水分变化是本研究中影响AEF值的重要因素, 二者之间具有正相关关系(图2, 图4).这一结果与羌塘高寒草地和对整个青藏高原高寒草地样带物种多样性和降水量的研究结果(杨元合等, 2004; 熊定鹏等, 2016)相符.一方面, 降水量的增加能够促进生态系统对碳的积累, 而且间接地促进土壤养分的矿化和积累, 从而在本研究结果中观察到土壤碳氮含量随降水量的增加呈现显著增加的趋势(图5)(Niu et al., 2007).另一方面, 本研究中降水量和干旱指数与AEF值、土壤有机碳、土壤全氮、微生物生物量碳和氮含量等养分循环相关的参数呈极显著的正相关关系(图5), 这些结果表明气候因子, 特别是水分条件是决定青藏高原高寒草地AEF的重要限制因子.此外, 也有研究指出, 在高寒草地生态系统, 气候调节土壤肥力和微生物的分解功能, 连接着物种多样性和EMF之间的关系(Jing et al., 2015).有研究证明, 气候调节和土壤肥力对陆地生态系统微生物的分解功能产生重要影响, 微生物多样性的任何损失都可能降低多功能性(Delgado-Baquerizo et al., 2016), 这些都有助于解释我们的结果. ...
... 本研究发现年降水量、干旱指数和土壤微生物生物量氮含量对AEF值有着较大的贡献(图3).有研究指出物种丰富度对EMF的影响极为关键, 是干旱生态系统多功能性的主要驱动力(Maestre et al., 2012).综合92个试验研究发现, 生物多样性能够增强生态系统在营养和生境水平中的多功能性(Lefcheck et al., 2015).我们的结果与前人的研究并不矛盾, 因为气候和物种丰富度维持碳氮循环, 而碳氮循环维持着碳氮的固定和土壤肥力相关的生态系统功能(王淑平等, 2002; Whitford, 2002).一般来说, 物种丰富度的增加通常通过提高地上、地下生产力和根系凋落物碳库来实现土壤碳累积, 而且, 物种多样性能够通过改变凋落物质量以及土壤的生物和非生物属性来影响土壤有机质分解, 从而影响EMF, 这是一个协同促进的过程(Cong et al., 2015).本研究结果表明, 土壤养分和微生物对地上生态系统功能有促进作用, 这与Jing等(2015)在青藏高原的研究结果一致, 这对草地生态系统物种多样性保护的认识具有重要意义. ...
中国东北样带(NECT)土壤碳、氮、磷的梯度分布及其与气候因子的关系 1 2002
... 本研究发现年降水量、干旱指数和土壤微生物生物量氮含量对AEF值有着较大的贡献(图3).有研究指出物种丰富度对EMF的影响极为关键, 是干旱生态系统多功能性的主要驱动力(Maestre et al., 2012).综合92个试验研究发现, 生物多样性能够增强生态系统在营养和生境水平中的多功能性(Lefcheck et al., 2015).我们的结果与前人的研究并不矛盾, 因为气候和物种丰富度维持碳氮循环, 而碳氮循环维持着碳氮的固定和土壤肥力相关的生态系统功能(王淑平等, 2002; Whitford, 2002).一般来说, 物种丰富度的增加通常通过提高地上、地下生产力和根系凋落物碳库来实现土壤碳累积, 而且, 物种多样性能够通过改变凋落物质量以及土壤的生物和非生物属性来影响土壤有机质分解, 从而影响EMF, 这是一个协同促进的过程(Cong et al., 2015).本研究结果表明, 土壤养分和微生物对地上生态系统功能有促进作用, 这与Jing等(2015)在青藏高原的研究结果一致, 这对草地生态系统物种多样性保护的认识具有重要意义. ...
Responses of litter decomposition and nutrient release rate to water and nitrogen addition differed among three plant species dominated in a semi-arid grassland 1 2017
... 随着研究的深入, 研究者逐渐认识到地下群落对AEF的驱动作用, 地下部分是影响草地生态系统结构、功能和规律中最不确定的因素(王毅等, 2019).土壤生物群落也被视为调节生态系统功能的关键因子(van der Heijden et al., 1998), 因为土壤生物群落结构的简化会降低EMF (van der Heijden et al., 1998; Wagg et al., 2014; Jing et al., 2015).不同的生态类型, 影响其生态系统功能的主导因素可能不同, 例如: 在森林生态系统中, 物种丰富度和功能多样性对土壤养分有显著正效应, 功能多样性能很好地解释生态系统功能(Delgado-Baquerizo et al., 2017).在全球干旱生态系统中, 土壤微生物群落驱动旱地EMF对全球变化的抵抗力(Delgado-Baquerizo et al., 2017).由于草地生态系统普遍受到氮限制, 在全球氮富集的背景下, 土壤氮可能成为影响和驱动EMF的关键(符义稳等, 2019).有研究发现, 随着全球气候变暖, CO2浓度增高, 土壤中不稳定碳的含量增加, 这些不稳定碳通过根系分泌物以及根的转化来增加土壤微生物对氮的需求, 从而增加植物和土壤微生物对可利用氮的竞争, 导致生态系统过程的氮限制, 降低生态系统功能(Luo et al., 2004).在草地生态系统中, 氮对植物生产力、碳储量和养分周转有正效应, 土壤水分和氮有效性增加可以提高植物群落的生产力, 进而提高地下功能复杂性(Bradford et al., 2014; 杨晓霞等, 2014; Wang et al., 2017).此外, 对青藏高原高寒草地的研究发现, 输入的无机氮被微生物固定, 能够显著促进土壤氮矿化速率, 从而促使原来有机氮的矿化和释放, 对生态系统功能产生非常积极的作用(Bai et al., 2011).土壤微生物的关键作用在于其能促进凋落物分解和有机物矿化等, 这些过程促进物质和能量在地上和地下群落之间转移, 在维持生态系统功能方面发挥着关键作用(Hooper et al., 2000; Wardle et al., 2004).一部分土壤微生物类群具有固定大气中氮的能力, 能将N2转变为植物可利用的形态, 这一类群的数量变化, 对土壤氮补充、平衡和调节, 以及对土壤结构、肥力以及生态系统功能至关重要(李刚等, 2013).此外, 研究发现, 在长期肥沃土壤环境下, 稀有微生物是生态系统多功能性的主要驱动力(Chen et al., 2019).因此, 全球气候变化导致土壤碳氮含量以及微生物任何变化都有可能改变地上和地下群落维持AEF的能力(Gans et al., 2005; Bardgett & van der Putten, 2014; Tedersoo et al., 2014). ...
若尔盖地区沙化草地土壤酶协同和抑制效应 1 2019
... 随着研究的深入, 研究者逐渐认识到地下群落对AEF的驱动作用, 地下部分是影响草地生态系统结构、功能和规律中最不确定的因素(王毅等, 2019).土壤生物群落也被视为调节生态系统功能的关键因子(van der Heijden et al., 1998), 因为土壤生物群落结构的简化会降低EMF (van der Heijden et al., 1998; Wagg et al., 2014; Jing et al., 2015).不同的生态类型, 影响其生态系统功能的主导因素可能不同, 例如: 在森林生态系统中, 物种丰富度和功能多样性对土壤养分有显著正效应, 功能多样性能很好地解释生态系统功能(Delgado-Baquerizo et al., 2017).在全球干旱生态系统中, 土壤微生物群落驱动旱地EMF对全球变化的抵抗力(Delgado-Baquerizo et al., 2017).由于草地生态系统普遍受到氮限制, 在全球氮富集的背景下, 土壤氮可能成为影响和驱动EMF的关键(符义稳等, 2019).有研究发现, 随着全球气候变暖, CO2浓度增高, 土壤中不稳定碳的含量增加, 这些不稳定碳通过根系分泌物以及根的转化来增加土壤微生物对氮的需求, 从而增加植物和土壤微生物对可利用氮的竞争, 导致生态系统过程的氮限制, 降低生态系统功能(Luo et al., 2004).在草地生态系统中, 氮对植物生产力、碳储量和养分周转有正效应, 土壤水分和氮有效性增加可以提高植物群落的生产力, 进而提高地下功能复杂性(Bradford et al., 2014; 杨晓霞等, 2014; Wang et al., 2017).此外, 对青藏高原高寒草地的研究发现, 输入的无机氮被微生物固定, 能够显著促进土壤氮矿化速率, 从而促使原来有机氮的矿化和释放, 对生态系统功能产生非常积极的作用(Bai et al., 2011).土壤微生物的关键作用在于其能促进凋落物分解和有机物矿化等, 这些过程促进物质和能量在地上和地下群落之间转移, 在维持生态系统功能方面发挥着关键作用(Hooper et al., 2000; Wardle et al., 2004).一部分土壤微生物类群具有固定大气中氮的能力, 能将N2转变为植物可利用的形态, 这一类群的数量变化, 对土壤氮补充、平衡和调节, 以及对土壤结构、肥力以及生态系统功能至关重要(李刚等, 2013).此外, 研究发现, 在长期肥沃土壤环境下, 稀有微生物是生态系统多功能性的主要驱动力(Chen et al., 2019).因此, 全球气候变化导致土壤碳氮含量以及微生物任何变化都有可能改变地上和地下群落维持AEF的能力(Gans et al., 2005; Bardgett & van der Putten, 2014; Tedersoo et al., 2014). ...
若尔盖地区沙化草地土壤酶协同和抑制效应 1 2019
... 随着研究的深入, 研究者逐渐认识到地下群落对AEF的驱动作用, 地下部分是影响草地生态系统结构、功能和规律中最不确定的因素(王毅等, 2019).土壤生物群落也被视为调节生态系统功能的关键因子(van der Heijden et al., 1998), 因为土壤生物群落结构的简化会降低EMF (van der Heijden et al., 1998; Wagg et al., 2014; Jing et al., 2015).不同的生态类型, 影响其生态系统功能的主导因素可能不同, 例如: 在森林生态系统中, 物种丰富度和功能多样性对土壤养分有显著正效应, 功能多样性能很好地解释生态系统功能(Delgado-Baquerizo et al., 2017).在全球干旱生态系统中, 土壤微生物群落驱动旱地EMF对全球变化的抵抗力(Delgado-Baquerizo et al., 2017).由于草地生态系统普遍受到氮限制, 在全球氮富集的背景下, 土壤氮可能成为影响和驱动EMF的关键(符义稳等, 2019).有研究发现, 随着全球气候变暖, CO2浓度增高, 土壤中不稳定碳的含量增加, 这些不稳定碳通过根系分泌物以及根的转化来增加土壤微生物对氮的需求, 从而增加植物和土壤微生物对可利用氮的竞争, 导致生态系统过程的氮限制, 降低生态系统功能(Luo et al., 2004).在草地生态系统中, 氮对植物生产力、碳储量和养分周转有正效应, 土壤水分和氮有效性增加可以提高植物群落的生产力, 进而提高地下功能复杂性(Bradford et al., 2014; 杨晓霞等, 2014; Wang et al., 2017).此外, 对青藏高原高寒草地的研究发现, 输入的无机氮被微生物固定, 能够显著促进土壤氮矿化速率, 从而促使原来有机氮的矿化和释放, 对生态系统功能产生非常积极的作用(Bai et al., 2011).土壤微生物的关键作用在于其能促进凋落物分解和有机物矿化等, 这些过程促进物质和能量在地上和地下群落之间转移, 在维持生态系统功能方面发挥着关键作用(Hooper et al., 2000; Wardle et al., 2004).一部分土壤微生物类群具有固定大气中氮的能力, 能将N2转变为植物可利用的形态, 这一类群的数量变化, 对土壤氮补充、平衡和调节, 以及对土壤结构、肥力以及生态系统功能至关重要(李刚等, 2013).此外, 研究发现, 在长期肥沃土壤环境下, 稀有微生物是生态系统多功能性的主要驱动力(Chen et al., 2019).因此, 全球气候变化导致土壤碳氮含量以及微生物任何变化都有可能改变地上和地下群落维持AEF的能力(Gans et al., 2005; Bardgett & van der Putten, 2014; Tedersoo et al., 2014). ...
Ecological linkages between aboveground and belowground biota 1 2004
... 随着研究的深入, 研究者逐渐认识到地下群落对AEF的驱动作用, 地下部分是影响草地生态系统结构、功能和规律中最不确定的因素(王毅等, 2019).土壤生物群落也被视为调节生态系统功能的关键因子(van der Heijden et al., 1998), 因为土壤生物群落结构的简化会降低EMF (van der Heijden et al., 1998; Wagg et al., 2014; Jing et al., 2015).不同的生态类型, 影响其生态系统功能的主导因素可能不同, 例如: 在森林生态系统中, 物种丰富度和功能多样性对土壤养分有显著正效应, 功能多样性能很好地解释生态系统功能(Delgado-Baquerizo et al., 2017).在全球干旱生态系统中, 土壤微生物群落驱动旱地EMF对全球变化的抵抗力(Delgado-Baquerizo et al., 2017).由于草地生态系统普遍受到氮限制, 在全球氮富集的背景下, 土壤氮可能成为影响和驱动EMF的关键(符义稳等, 2019).有研究发现, 随着全球气候变暖, CO2浓度增高, 土壤中不稳定碳的含量增加, 这些不稳定碳通过根系分泌物以及根的转化来增加土壤微生物对氮的需求, 从而增加植物和土壤微生物对可利用氮的竞争, 导致生态系统过程的氮限制, 降低生态系统功能(Luo et al., 2004).在草地生态系统中, 氮对植物生产力、碳储量和养分周转有正效应, 土壤水分和氮有效性增加可以提高植物群落的生产力, 进而提高地下功能复杂性(Bradford et al., 2014; 杨晓霞等, 2014; Wang et al., 2017).此外, 对青藏高原高寒草地的研究发现, 输入的无机氮被微生物固定, 能够显著促进土壤氮矿化速率, 从而促使原来有机氮的矿化和释放, 对生态系统功能产生非常积极的作用(Bai et al., 2011).土壤微生物的关键作用在于其能促进凋落物分解和有机物矿化等, 这些过程促进物质和能量在地上和地下群落之间转移, 在维持生态系统功能方面发挥着关键作用(Hooper et al., 2000; Wardle et al., 2004).一部分土壤微生物类群具有固定大气中氮的能力, 能将N2转变为植物可利用的形态, 这一类群的数量变化, 对土壤氮补充、平衡和调节, 以及对土壤结构、肥力以及生态系统功能至关重要(李刚等, 2013).此外, 研究发现, 在长期肥沃土壤环境下, 稀有微生物是生态系统多功能性的主要驱动力(Chen et al., 2019).因此, 全球气候变化导致土壤碳氮含量以及微生物任何变化都有可能改变地上和地下群落维持AEF的能力(Gans et al., 2005; Bardgett & van der Putten, 2014; Tedersoo et al., 2014). ...
1 2002
... 本研究发现年降水量、干旱指数和土壤微生物生物量氮含量对AEF值有着较大的贡献(图3).有研究指出物种丰富度对EMF的影响极为关键, 是干旱生态系统多功能性的主要驱动力(Maestre et al., 2012).综合92个试验研究发现, 生物多样性能够增强生态系统在营养和生境水平中的多功能性(Lefcheck et al., 2015).我们的结果与前人的研究并不矛盾, 因为气候和物种丰富度维持碳氮循环, 而碳氮循环维持着碳氮的固定和土壤肥力相关的生态系统功能(王淑平等, 2002; Whitford, 2002).一般来说, 物种丰富度的增加通常通过提高地上、地下生产力和根系凋落物碳库来实现土壤碳累积, 而且, 物种多样性能够通过改变凋落物质量以及土壤的生物和非生物属性来影响土壤有机质分解, 从而影响EMF, 这是一个协同促进的过程(Cong et al., 2015).本研究结果表明, 土壤养分和微生物对地上生态系统功能有促进作用, 这与Jing等(2015)在青藏高原的研究结果一致, 这对草地生态系统物种多样性保护的认识具有重要意义. ...
羌塘高寒草地物种多样性与生态系统多功能关系格局 4 2016
... 高寒草地是青藏高原最主要的生态系统, 在气候调节、水源涵养和碳氮固定等方面发挥着重要的生态功能(杨元合和朴世龙, 2006; 孙鸿烈等, 2012; Sun et al., 2020).青藏高原独特的高海拔特征、寒冷的气候条件和严苛的植物生长环境导致高寒草地生态系统非常脆弱, 生态系统功能对环境变化非常敏感(Jing et al., 2015; 熊定鹏等, 2016).在地上群落生物多样性和EMF的关系格局方面, 研究发现, 降水量显著影响物种丰富度和EMF, 但年降水量并未显著影响物种多样性指数, 强调了群落物种丰富度对维持EMF重要意义, 这意味着物种的丧失可能会给高寒草地EMF和生态服务带来更为严重的后果(熊定鹏等, 2016).EMF与地上和地下生物多样性关系方面, 研究发现二者的联系是由气候条件决定的(Jing et al., 2015).作为对气候条件十分敏感的青藏高原, 目前尚缺乏降水梯度下地下土壤关键因子对高寒草地生态系统地上功能影响的主要控制因子和机理的研究.因此, 本研究选择青藏高原地区, 基于样带上115个样点调查分析, 综合运用13种生态系统功能参数和3个气候因子, 试图回答以下两个科学问题: (1)驱动高寒草地AEF的主要因子是什么? (2)这种因子是如何作用于AEF的? 以期为高寒草地生态系统多样性保护、多功能性维持和可持续利用管理提供科学依据. ...
 ,
, ,2,*, 叶冲冲2,3, 曾涛
,2,*, 叶冲冲2,3, 曾涛

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT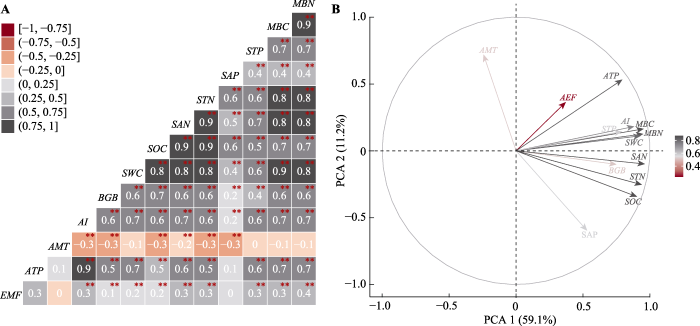 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT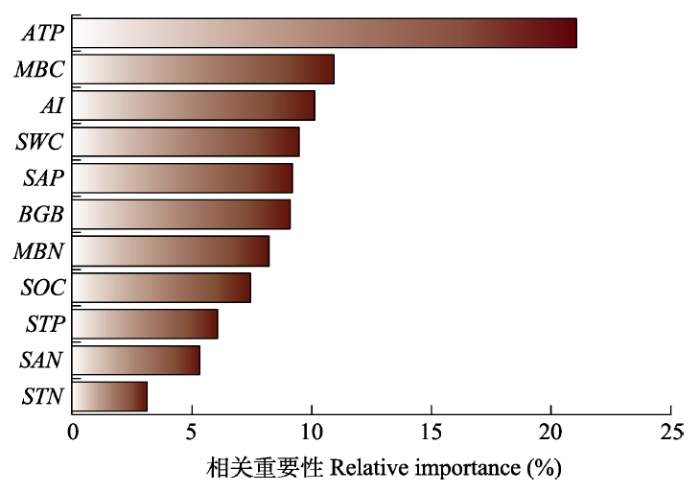 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT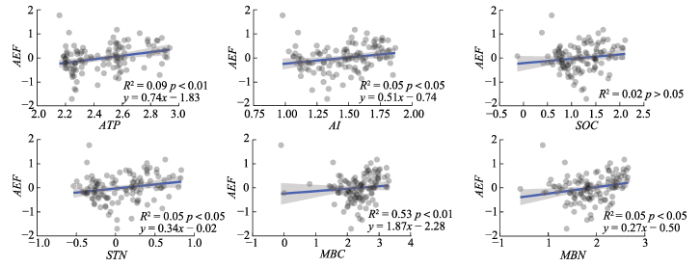 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT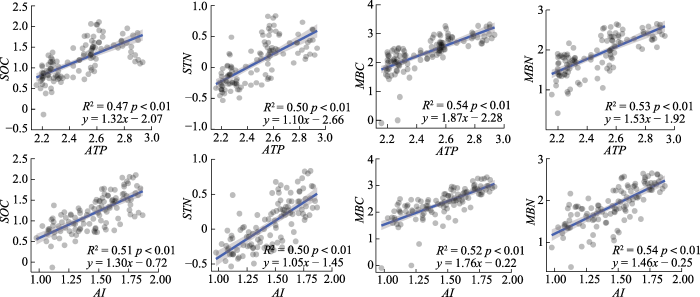 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT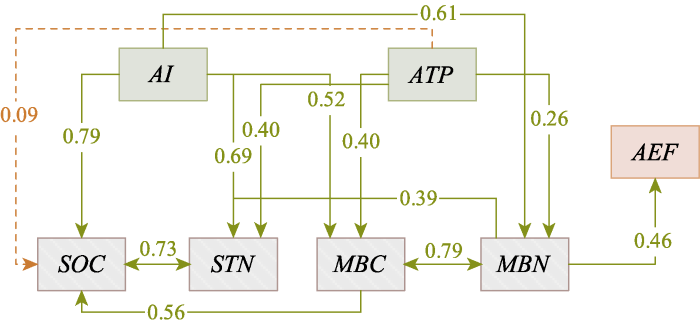 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}