Research progress and prospect on the impacts of resource pulses on alien plant invasion
Wen-Chao QIN1,2,3,4, Zhi-Bin TAO3,5, Yong-Jian WANG6, Yan-Jie LIU7, Wei HUANG,,3,5,*1Research Center for Ecology and Environment of Qinghai-Tibetan Plateau, Tibet University, Lhasa 850000, China 2College of Science, Tibet University, Lhasa 850000, China 3Key Laboratory of Aquatic Botany and Watershed Ecology, Wuhan Botanical Garden, Chinese Academy of Sciences, Wuhan 430074, China 4University of Chinese Academy of Sciences, Beijing 100049, China 5Center of Conservation Biology, Core Botanical Gardens, Chinese Academy of Sciences, Wuhan 430074, China 6College of Horticulture and Forestry Sciences, Huazhong Agricultural University, Wuhan 430070, China 7Key Laboratory of Wetland Ecology and Environment, Northeast Institute of Geography and Agroecology, Chinese Academy Sciences, Changchun 130102, China
National Key R&D Program of China(2017YFC1200100) National Natural Science Foundation of China(31822007) National Natural Science Foundation of China(2071660) Natural Science Foundation of Hubei(2020CFA064)
Abstract Under global change, the variations of resources such as nutrients and water are not stabilized and often occur in the form of pulses with low frequency, large amplitude, and short duration. The existing evidence suggests that resource pulses have the potential to disrupt the inherent equilibrium of plant communities, and thereby play a critical role in another important component of global change, plant invasion. To date, studies on the impacts of global change on plant invasion have mainly focused on the constant changes of resources, but have ignored the resource fluctuation, in particular the role of resource pulses. Here, we present a literature review concerning the effects of resource pulses on plant invasion, including causes, types and impacts of resource pulses. Then we discuss the importance of resource pulses in plant invasion. Furthermore, our review puts forward some fields that are worthy to be paid more attention in the future, such as the importance of attributes and types of resource pulses, their interactive impacts on and potential mechanisms in plant invasion. Keywords:global change;biological invasion;invasive plant;resource availability;resource fluctuation;resource pulse
PDF (1062KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 秦文超, 陶至彬, 王永健, 刘艳杰, 黄伟. 资源脉冲对外来植物入侵影响的研究进展和展望. 植物生态学报, 2021, 45(6): 573-582. DOI: 10.17521/cjpe.2021.0068 QIN Wen-Chao, TAO Zhi-Bin, WANG Yong-Jian, LIU Yan-Jie, HUANG Wei. Research progress and prospect on the impacts of resource pulses on alien plant invasion. Chinese Journal of Plant Ecology, 2021, 45(6): 573-582. DOI: 10.17521/cjpe.2021.0068
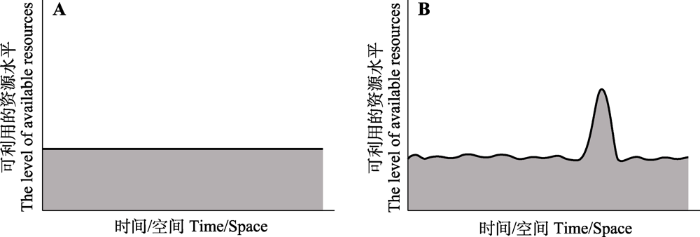
Fig. 1Two types of variations of the available resources in the ecosystem under global change. A, Stabilized variation—The variation of resources maintains a stabilized state. B, Resource pulse—The variation of resources with great fluctuation.
自Davis等于2000年提出资源波动假说之后, 相关研究不断增加。我们以2000-2021年为研究区间, 以TS = ((resources* or nutrient* or water* or drought* or temperature* or light*) and (resource pulses* or pulsed resources* or resources fluctuation* or spatial and temporal variation*) and (biological invasion* or invasive plant* or alien plant* or exotic plant*))为关键词, 在Web of Science进行了文献检索。初步检索出1 454篇文献。对这些文献我们通过以下规则进行了进一步的文献筛选: (1)研究范围属于生态学领域; (2)研究对象是外来入侵植物, 而非动物、微生物等; (3)研究包括资源的波动。经过筛选, 共得出有效文献83篇。具体研究的资源类型包括营养、水分、光3类, 相关研究分别有57、38、9篇(图2, 部分研究包含多种资源脉冲类型, 故数量有所重叠)。支持资源波动假说的文章有75篇, 持反对观点的文章有8篇。
AlpertP, BoneE, HolzapfelC (2000). Invasiveness, invasibility and the role of environmental stress in the spread of non-native plants Perspectives in Plant Ecology, Evolution and Systematics, 3, 52-66. [本文引用: 1]
BesawLM, ThelenGC, SutherlandS, MetlenK, CallawayRM (2011). Disturbance, resource pulses and invasion: short-term shifts in competitive effects, not growth responses, favour exotic annuals Journal of Applied Ecology, 48, 998-1006. DOI:10.1111/jpe.2011.48.issue-4URL [本文引用: 1]
BogdziewiczM, CroneEE, SteeleMA, ZwolakR (2017). Effects of nitrogen deposition on reproduction in a masting tree: benefits of higher seed production are trumped by negative biotic interactions Journal of Ecology, 105, 310-320. DOI:10.1111/jec.2017.105.issue-2URL [本文引用: 1]
BradleyBA, BlumenthalDM, WilcoveDS, ZiskaLH (2010a). Predicting plant invasions in an era of global change Trends in Ecology & Evolution, 25, 310-318. DOI:10.1016/j.tree.2009.12.003URL [本文引用: 2]
BradleyBA, WilcoveDS, OppenheimerM (2010b). Climate change increases risk of plant invasion in the Eastern United States Biological Invasions, 12, 1855-1872. DOI:10.1007/s10530-009-9597-yURL [本文引用: 1]
BrownKA, GurevitchJ (2004). Long-term impacts of logging on forest diversity in Madagascar Proceedings of the National Academy of Sciences of the United States of America, 101, 6045-6049. [本文引用: 1]
CampbellBD, GrimeJP (1989). A comparative study of plant responsiveness to the duration of episodes of mineral nutrient enrichment New Phytologist, 112, 261-267. DOI:10.1111/nph.1989.112.issue-2URL [本文引用: 1]
CharbonneauNC, FahrigL (2004). Influence of canopy cover and amount of open habitat in the surrounding landscape on proportion of alien plant species in forest sites Écoscience, 11, 278-281. DOI:10.1080/11956860.2004.11682833URL [本文引用: 1]
ChenD, XiongH, LinCG, HeW, ZhangZW, WangH, WangYJ (2019). Clonal integration benefits invasive alien plants under water variability in a native community Journal of Plant Ecology, 12, 574-582. DOI:10.1093/jpe/rty050 [本文引用: 2] Many invasive alien plant species are clonal and can greatly propagate and spread through clonal integration (sharing resources between connected ramets) in heterogeneous and variable environments. Here, we tested whether water variability influences clonal integration of invasive alien plant species and consequently facilitates their growth and dominance in a native community. We selected four typical invasive clonal plant species in China. Connected (with clonal integration) and disconnected (without clonal integration) clonal fragments were established either under constant watering or variable watering condition in an experimental native plant community consisting of three naturally co-occurring grassland species. Proximal part of the container received high nutrient and distal part received low nutrient. Clonal integration significantly increased biomass, aboveground mass and belowground mass of invasive alien plants in the proximal ramets, the distal ramets and the whole clone and decreased the growth of native community. Interestingly, clonal integration significantly increased the growth of invasive plants in variable watering. The positive effect of clonal integration was stronger in variable watering than in constant watering. Invasive plants with clonal integration had high biomass proportion (> 0.6) in the whole community. Our results suggest that invasive clonal plants benefit more from clonal integration in variable water environments when established in a native community, and to some extent, clonal integration potentially contribute greatly to the invasiveness of alien clonal plants when they enter a new community with resource variability.
ChenSP, BaiYF, ZhangLX, HanXG (2005). Comparing physiological responses of two dominant grass species to nitrogen addition in Xilin River Basin of China Environmental and Experimental Botany, 53, 65-75. DOI:10.1016/j.envexpbot.2004.03.002URL [本文引用: 2]
ChytrýM, WildJ, PyšekP, JarošíkV, DendonckerN, ReginsterI, PinoJ, MaskellLC, VilàM, PerglJ, KühnI, SpangenbergJH, SetteleJ (2012). Projecting trends in plant invasions in Europe under different scenarios of future land-use change Global Ecology and Biogeography, 21, 75-87. DOI:10.1111/j.1466-8238.2010.00573.xURL [本文引用: 1]
CraineJM, DybzinskiR (2013). Mechanisms of plant competition for nutrients, water and light Functional Ecology, 27, 833-840. DOI:10.1111/1365-2435.12081URL [本文引用: 1]
DavisMA, GrimeJP, ThompsonK (2000). Fluctuating resources in plant communities: a general theory of invasibility Journal of Ecology, 88, 528-534. DOI:10.1046/j.1365-2745.2000.00473.xURL [本文引用: 4]
EarlyR, BradleyBA, DukesJS, LawlerJJ, OldenJD, BlumenthalDM, GonzalezP, GrosholzED, IbañezI, MillerLP, SorteCJB, TatemAJ (2016). Global threats from invasive alien species in the twenty-first century and national response capacities Nature Communications, 7, 12485. DOI: 10.1038/ncomms12485. URL [本文引用: 1]
EasterlingDR, MeehlGA, ParmesanC, ChangnonSA, KarlTR, MearnsLO (2000). Climate extremes: observations, modeling, and impacts Science, 289, 2068-2074. PMID:11000103 [本文引用: 1] One of the major concerns with a potential change in climate is that an increase in extreme events will occur. Results of observational studies suggest that in many areas that have been analyzed, changes in total precipitation are amplified at the tails, and changes in some temperature extremes have been observed. Model output has been analyzed that shows changes in extreme events for future climates, such as increases in extreme high temperatures, decreases in extreme low temperatures, and increases in intense precipitation events. In addition, the societal infrastructure is becoming more sensitive to weather and climate extremes, which would be exacerbated by climate change. In wild plants and animals, climate-induced extinctions, distributional and phenological changes, and species' range shifts are being documented at an increasing rate. Several apparently gradual biological changes are linked to responses to extreme weather and climate events.
EnsE, HutleyLB, Rossiter-RachorNA, DouglasMM, SetterfieldSA (2015). Resource-use efficiency explains grassy weed invasion in a low-resource savanna in north Australia Frontiers in Plant Science, 6,560. DOI: 10.3389/fpls.?2015.00560. [本文引用: 1]
EzizA, YanZ, TianD, HanW, TangZ, FangJ (2017). Drought effect on plant biomass allocation: a meta-analysis Ecology and Evolution, 7, 11002-11010. DOI:10.1002/ece3.2017.7.issue-24URL [本文引用: 1]
FeltonAJ, SmithMD (2017). Integrating plant ecological responses to climate extremes from individual to ecosystem levels Philosophical Transactions of the Royal Society B: Biological Sciences, 372, 20160142. DOI: 10.1098/rstb.?2016.?0142. URL [本文引用: 1]
FrevolaDM, HovickSM (2019). The independent effects of nutrient enrichment and pulsed nutrient delivery on a common wetland invader and its native conspecific Oecologia, 191, 447-460. DOI:10.1007/s00442-019-04493-yURL [本文引用: 2]
FridleyJD (2012). Extended leaf phenology and the autumn niche in deciduous forest invasions Nature, 485, 359-362. DOI:10.1038/nature11056URL [本文引用: 1]
FunkJL (2008). Differences in plasticity between invasive and native plants from a low resource environment Journal of Ecology, 96, 1162-1173. DOI:10.1111/jec.2008.96.issue-6URL [本文引用: 2]
FunkJL (2013). The physiology of invasive plants in low- resource environments Conservation Physiology, 1. DOI: 10.1093/conphys/cot026. [本文引用: 3]
FunkJL, VitousekPM (2007). Resource-use efficiency and plant invasion in low-resource systems Nature, 446, 1079-1081. DOI:10.1038/nature05719URL [本文引用: 1]
GilliamFS (2006). Response of the herbaceous layer of forest ecosystems to excess nitrogen deposition Journal of Ecology, 94, 1176-1191. DOI:10.1111/jec.2006.94.issue-6URL [本文引用: 1]
GoldbergDE, RajaniemiT, GurevitchJ, Stewart-OatenA (1999). Empirical approaches to quantifying interaction intensity: competition and facilitation along productivity gradients Ecology, 80, 1118-1131. DOI:10.1890/0012-9658(1999)080[1118:EATQII]2.0.CO;2URL [本文引用: 1]
GrantPR, GrantBR, KellerLF, PetrenK (2000). Effects of El Niño events on Darwin’s finch productivity Ecology, 81, 2442-2457. [本文引用: 1]
GrattonC, DennoRF (2003). Inter-year carryover effects of a nutrient pulse on Spartina plants, herbivores, and natural enemies. Ecology, 84, 2692-2707. [本文引用: 1]
HarpoleWS, PottsDL, SudingKN (2007). Ecosystem responses to water and nitrogen amendment in a California grassland Global Change Biology, 13, 2341-2348. DOI:10.1111/gcb.2007.13.issue-11URL [本文引用: 1]
HobbsRJ, YatesS, MooneyHA (2007). Long-term data reveal complex dynamics in grassland in relation to climate and disturbance Ecological Monographs, 77, 545-568. DOI:10.1890/06-1530.1URL [本文引用: 1]
JabranK, DoğanMN (2020). Elevated CO2, temperature and nitrogen levels impact growth and development of invasive weeds in the Mediterranean region Journal of the Science of Food and Agriculture, 100, 4893-4900. DOI:10.1002/jsfa.v100.13URL [本文引用: 1]
JentschA, WhiteP (2019). A theory of pulse dynamics and disturbance in ecology Ecology, 100, e02734. DOI: 10.1002/ecy.2734. [本文引用: 1]
KnüselS, ConederaM, BugmannH, WunderJ (2019). Low litter cover, high light availability and rock cover favour the establishment of Ailanthus altissima in forests in southern Switzerland. NeoBiota, 46, 91-116. [本文引用: 1]
KreylingJ, BeierkuhnleinC, EllisL, JentschA (2008). Invasibility of grassland and heath communities exposed to extreme weather events-additive effects of diversity resistance and fluctuating physical environment Oikos, 117, 1542-1554. DOI:10.1111/j.0030-1299.2008.16653.xURL [本文引用: 1]
LaForgiaML, HarrisonSP, LatimerAM (2020). Invasive species interact with climatic variability to reduce success of natives Ecology, 101, e03022. DOI: 10.1002/ecy.3022. [本文引用: 1]
LarsonDL, AndersonPJ, NewtonW (2001). Alien plant invasion in mixed-grass prairie: effects of vegetation type and anthropogenic disturbance Ecological Applications, 11, 128-141. DOI:10.1890/1051-0761(2001)011[0128:APIIMG]2.0.CO;2URL [本文引用: 1]
LetnicM, TamayoB, DickmanCR (2005). The responses of mammals to La Niña (El Niño Southern Oscillation)- associated rainfall, predation, and wildfire in central Australia Journal of Mammalogy, 86, 689-703. DOI:10.1644/1545-1542(2005)086[0689:TROMTL]2.0.CO;2URL [本文引用: 1]
LiuS, StreichJ, BorevitzJO, RiceKJ, LiT, LiB, BradfordKJ (2019). Environmental resource deficit may drive the evolution of intraspecific trait variation in invasive plant populations Oikos, 128, 171-184. DOI:10.1111/oik.2019.v128.i2URL [本文引用: 1]
LiuY, OduorAMO, ZhangZ, ManeaA, ToothIM, LeishmanMR, XuX, van KleunenM (2017). Do invasive alien plants benefit more from global environmental change than native plants? Global Change Biology, 23, 3363-3370. DOI:10.1111/gcb.2017.23.issue-8URL [本文引用: 1]
LiuY, van KleunenM (2017). Responses of common and rare aliens and natives to nutrient availability and fluctuations Journal of Ecology, 105, 1111-1122. DOI:10.1111/jec.2017.105.issue-4URL [本文引用: 2]
LiuY, ZhangX, van KleunenM (2018). Increases and fluctuations in nutrient availability do not promote dominance of alien plants in synthetic communities of common natives Functional Ecology, 32, 2594-2604. DOI:10.1111/fec.2018.32.issue-11URL [本文引用: 2]
LüXT, HanXG (2010). Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia, China Plant and Soil, 327, 481-491. DOI:10.1007/s11104-009-0078-yURL [本文引用: 1]
MallakpourI, VillariniG (2017). Analysis of changes in the magnitude, frequency, and seasonality of heavy precipitation over the contiguous USA Theoretical and Applied Climatology, 130, 345-363. DOI:10.1007/s00704-016-1881-zURL [本文引用: 1]
MeehlGA, TebaldiC (2004). More intense, more frequent, and longer lasting heat waves in the 21st century Science, 305, 994-997. DOI:10.1126/science.1098704URL [本文引用: 1]
NiinemetsÜ, ValladaresF, CeulemansR (2003). Leaf-level phenotypic variability and plasticity of invasive Rhododendron ponticum and non-invasive Ilex aquifolium co-occurring at two contrasting European sites. Plant, Cell & Environment, 26, 941-956. [本文引用: 1]
NowlinWH, VanniMJ, YangLH (2008). Comparing resource pulses in aquatic and terrestrial ecosystems Ecology, 89, 647-659. DOI:10.1890/07-0303.1URL [本文引用: 1]
OshimaK, TakahashiK (2020). Forest disturbances promote invasion of alien herbaceous plants: a comparison of abundance and plant traits between alien and native species in thinned and unthinned stands Biological Invasions, 22, 2749-2762. DOI:10.1007/s10530-020-02283-9URL [本文引用: 2]
OsoneY, YazakiK, MasakiT, IshidaA (2014). Responses to nitrogen pulses and growth under low nitrogen availability in invasive and native tree species with differing successional status Journal of Plant Research, 127, 315-328. DOI:10.1007/s10265-013-0609-8PMID:24292716 [本文引用: 1] Invasive species are frequently found in recently disturbed sites. To examine how these disturbance-dependent invasive species exploit resource pulses resulting from disturbance, twelve physiological and morphological traits, including age-dependent responsiveness in leaf traits to nitrogen pulse, were compared between Bischofia javanica, an invasive tree species in Ogasawara islands, and three native Ogasawara species, each having a different successional status. When exposed to a nitrogen pulse, invasive B. javanica showed higher increases in photosynthetic capacity, leaf area, epidermal cell number and cell size in leaves of broad age classes, and root nitrogen absorption ability than two native mid-/late or late-successional species, but showed no particular superiority to a native pioneer species in these responses. Under low nitrogen, however, it showed the largest relative growth rate among the four species, while the native pioneer showed the lowest growth. From these results, we concluded that the combination of moderately high responsiveness to resource pulses and the ability to maintain steady growth under resource limitations may give B. javanica a competitive advantage over a series of native species with different successional status from early to late-successional stages.
OstfeldRS, KeesingF (2000). Pulsed resources and community dynamics of consumers in terrestrial ecosystems Trends in Ecology & Evolution, 15, 232-237. DOI:10.1016/S0169-5347(00)01862-0URL [本文引用: 2]
PainiDR, SheppardAW, CookDC, De BarroPJ, WornerSP, ThomasMB (2016). Global threat to agriculture from invasive species Proceedings of the National Academy of Sciences of the United States of America, 113, 7575-7579. DOI:10.1073/pnas.1602205113PMID:27325781 [本文引用: 1] Invasive species present significant threats to global agriculture, although how the magnitude and distribution of the threats vary between countries and regions remains unclear. Here, we present an analysis of almost 1,300 known invasive insect pests and pathogens, calculating the total potential cost of these species invading each of 124 countries of the world, as well as determining which countries present the greatest threat to the rest of the world given their trading partners and incumbent pool of invasive species. We find that countries vary in terms of potential threat from invasive species and also their role as potential sources, with apparently similar countries sometimes varying markedly depending on specifics of agricultural commodities and trade patterns. Overall, the biggest agricultural producers (China and the United States) could experience the greatest absolute cost from further species invasions. However, developing countries, in particular, Sub-Saharan African countries, appear most vulnerable in relative terms. Furthermore, China and the United States represent the greatest potential sources of invasive species for the rest of the world. The analysis reveals considerable scope for ongoing redistribution of known invasive pests and highlights the need for international cooperation to slow their spread.
Palacio-LópezK, GianoliE (2011). Invasive plants do not display greater phenotypic plasticity than their native or non-invasive counterparts: a meta-analysis Oikos, 120, 1393-1401. DOI:10.1111/more.2011.120.issue-9URL [本文引用: 1]
ParepaM, KahmenA, WernerRA, FischerM, BossdorfO (2019). Invasive knotweed has greater nitrogen-use efficiency than native plants: evidence from a 15N pulse-chasing experiment Oecologia, 191, 389-396. DOI:10.1007/s00442-019-04490-1URL [本文引用: 3]
PolisGA, HurdSD, JacksonCT, PiñeroFS (1997). El Niño effects on the dynamics and control of an island ecosystem in the Gulf of California Ecology, 78, 1884-1897. [本文引用: 1]
PostAK, KnappAK (2020). The importance of extreme rainfall events and their timing in a semi-arid grassland Journal of Ecology, 108, 2431-2443. DOI:10.1111/jec.v108.6URL [本文引用: 2]
PrevéyJS, SeastedtTR (2014). Seasonality of precipitation interacts with exotic species to alter composition and phenology of a semi-arid grassland Journal of Ecology, 102, 1549-1561. DOI:10.1111/jec.2014.102.issue-6URL [本文引用: 1]
RichardsCL, BossdorfO, MuthNZ, GurevitchJ, PigliucciM (2006). Jack of all trades, master of some? On the role of phenotypic plasticity in plant invasions Ecology Letters, 9, 981-993. DOI:10.1111/ele.2006.9.issue-8URL [本文引用: 1]
RosenzweigML (2001). The four questions: What does the introduction of exotic species do to diversity? Evolutionary Ecology Research, 3, 361-367. [本文引用: 1]
SchmidtKA (2003). Linking frequencies of acorn masting in temperate forests to long-term population growth rates in a songbird: the veery (Catharus fuscescens) Oikos, 103, 548-558. DOI:10.1034/j.1600-0706.2003.12462.xURL [本文引用: 2]
SeebensH, BacherS, BlackburnTM, CapinhaC, DawsonW, DullingerS, GenovesiP, HulmePE, van KleunenM, KühnI, JeschkeJM, LenznerB, LiebholdAM, PattisonZ, PerglJ, PyšekP, WinterM, EsslF (2021). Projecting the continental accumulation of alien species through to 2050 Global Change Biology, 27, 970-982. DOI:10.1111/gcb.v27.5URL [本文引用: 1]
SeebensH, BlackburnTM, DyerEE, GenovesiP, HulmePE, JeschkeJM, PagadS, PyšekP, WinterM, ArianoutsouM, BacherS, BlasiusB, BrunduG, CapinhaC, Celesti- GrapowL, et al. (2017). No saturation in the accumulation of alien species worldwide Nature Communications, 8, 14435. DOI: 10.1038/ncomms14435. PMID:28198420 [本文引用: 2] Although research on human-mediated exchanges of species has substantially intensified during the last centuries, we know surprisingly little about temporal dynamics of alien species accumulations across regions and taxa. Using a novel database of 45,813 first records of 16,926 established alien species, we show that the annual rate of first records worldwide has increased during the last 200 years, with 37% of all first records reported most recently (1970-2014). Inter-continental and inter-taxonomic variation can be largely attributed to the diaspora of European settlers in the nineteenth century and to the acceleration in trade in the twentieth century. For all taxonomic groups, the increase in numbers of alien species does not show any sign of saturation and most taxa even show increases in the rate of first records over time. This highlights that past efforts to mitigate invasions have not been effective enough to keep up with increasing globalization.
ShenXY, PengSL, ChenBM, PangJX, ChenLY, XuHM, HouYP (2011). Do higher resource capture ability and utilization efficiency facilitate the successful invasion of native plants? Biological Invasions, 13, 869-881. DOI:10.1007/s10530-010-9875-8URL [本文引用: 1]
SheppardCS, StanleyMC (2014). Does elevated temperature and doubled CO2 increase growth of three potentially invasive plants? Invasive Plant Science and Management, 7, 237-246. DOI:10.1614/IPSM-D-13-00038.1URL [本文引用: 2]
SinclairARE, KrebsCJ (2002). Complex numerical responses to top-down and bottom-up processes in vertebrate populations Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 357, 1221-1231. [本文引用: 1]
SmithLM (2013). Extended leaf phenology in deciduous forest invaders: mechanisms of impact on native communities Journal of Vegetation Science, 24, 979-987. DOI:10.1111/jvs.12087URL [本文引用: 1]
SunJ, JavedQ, AzeemA, UllahMS, RasoolG, DuD (2020). Addition of phosphorus and nitrogen support the invasiveness of Alternanthera Philoxeroides under water stress. CLEAN-Soil, Air, Water, 48, 2000059. DOI: 10.1002/clen.202000059. [本文引用: 1]
van KleunenM, BossdorfO, DawsonW (2018). The ecology and evolution of alien plants Annual Review of Ecology, Evolution, and Systematics, 49, 25-47. DOI:10.1146/annurev-ecolsys-110617-062654URL [本文引用: 1]
van KleunenM, DawsonW, EsslF, PerglJ, WinterM, WeberE, KreftH, WeigeltP, KarteszJ, NishinoM, AntonovaLA, BarcelonaJF, CabezasFJ, CárdenasD, Cárdenas-ToroJ, et al. (2015). Global exchange and accumulation of non-native plants Nature, 525, 100-103. DOI:10.1038/nature14910URL [本文引用: 1]
van KleunenM, WeberE, FischerM (2010). A meta-analysis of trait differences between invasive and non-invasive plant species Ecology Letters, 13, 235-245. DOI:10.1111/j.1461-0248.2009.01418.xPMID:20002494 [本文引用: 1] A major aim in ecology is identifying determinants of invasiveness. We performed a meta-analysis of 117 field or experimental-garden studies that measured pair-wise trait differences of a total of 125 invasive and 196 non-invasive plant species in the invasive range of the invasive species. We tested whether invasiveness is associated with performance-related traits (physiology, leaf-area allocation, shoot allocation, growth rate, size and fitness), and whether such associations depend on type of study and on biogeographical or biological factors. Overall, invasive species had significantly higher values than non-invasive species for all six trait categories. More trait differences were significant for invasive vs. native comparisons than for invasive vs. non-invasive alien comparisons. Moreover, for comparisons between invasive species and native species that themselves are invasive elsewhere, no trait differences were significant. Differences in physiology and growth rate were larger in tropical regions than in temperate regions. Trait differences did not depend on whether the invasive alien species originates from Europe, nor did they depend on the test environment. We conclude that invasive alien species had higher values for those traits related to performance than non-invasive species. This suggests that it might become possible to predict future plant invasions from species traits.
VetterVMS, KreylingJ, DenglerJ, ApostolovaI, Arfin-KhanMAS, BerauerBJ, BerwaersS, de BoeckHJ, NijsI, SchuchardtMA, SopotlievaD, von GillhausenP, WilfahrtPA, ZimmermannM, JentschA (2020). Invader presence disrupts the stabilizing effect of species richness in plant community recovery after drought Global Change Biology, 26, 3539-3551. DOI:10.1111/gcb.v26.6URL [本文引用: 2]
VitousekPM, D’AntonioCM, LoopeLL, WestbrooksR (1996). Biological invasions as global environmental change American Scientist, 84, 468-478. [本文引用: 2]
VitousekPM, MooneyHA, LubchencoJ, MelilloJM (1997). Human domination of Earthʼs ecosystems//Marzluff JM, Shulenberger E, Endlicher W, Alberti M, Bradley G, Ryan C, Simon U, ZumBrunnen C. Urban Ecology Springer, Boston. [本文引用: 2]
WalkerGA, RobertsonMP, GaertnerM, GallienL, RichardsonDM (2017). The potential range of Ailanthus altissima (tree of heaven) in South Africa: the roles of climate, land use and disturbance Biological Invasions, 19, 3675-3690. DOI:10.1007/s10530-017-1597-8URL [本文引用: 1]
WeberMJ, BrownML (2019). Recipient ecosystem productivity influences effects of resource pulses in mesocosms Hydrobiologia, 827, 183-199. DOI:10.1007/s10750-018-3764-zURL [本文引用: 1]
WeinerJ (2004). Allocation, plasticity and allometry in plants Perspectives in Plant Ecology, Evolution and Systematics, 6, 207-215. DOI:10.1078/1433-8319-00083URL [本文引用: 1]
WolkovichEM, ClelandEE (2011). The phenology of plant invasions: a community ecology perspective Frontiers in Ecology and the Environment, 9, 287-294. DOI:10.1890/100033URL [本文引用: 1]
WolkovichEM, DaviesTJ, SchaeferH, ClelandEE, CookBI, TraversSE, WillisCG, DavisCC (2013). Temperature- dependent shifts in phenology contribute to the success of exotic species with climate change American Journal of Botany, 100, 1407-1421. DOI:10.3732/ajb.1200478URL [本文引用: 1]
YangLH (2004). Periodical cicadas as resource pulses in North American forests Science, 306, 1565-1567. DOI:10.1126/science.1103114URL [本文引用: 1]
YangLH (2006). Interactions between a detrital resource pulse and a detritivore community Oecologia, 147, 522-532. DOI:10.1007/s00442-005-0276-0URL [本文引用: 1]
YangLH, BastowJL, SpenceKO, WrightAN (2008). What can we learn from resource pulses? Ecology, 89, 621-634. DOI:10.1890/07-0175.1URL [本文引用: 3]
... 1992年, 美国****Vitousek首次提出“全球环境变化(global environmental change)”的概念, 并指出全球变化终将对地球生态系统进程产生巨大影响.全球变化驱动因素一般包括大气CO2富集、气候变化、土地利用变化、氮沉降和生物入侵等(Vitousek et al., 1996; Bradley et al., 2010a; Sage, 2020).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
Cheatgrass is favored by warming but not CO2 enrichment in a semi-arid grassland 1 2016
... 1992年, 美国****Vitousek首次提出“全球环境变化(global environmental change)”的概念, 并指出全球变化终将对地球生态系统进程产生巨大影响.全球变化驱动因素一般包括大气CO2富集、气候变化、土地利用变化、氮沉降和生物入侵等(Vitousek et al., 1996; Bradley et al., 2010a; Sage, 2020).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
Effects of nitrogen deposition on reproduction in a masting tree: benefits of higher seed production are trumped by negative biotic interactions 1 2017
Invasibility of grassland and heath communities exposed to extreme weather events-additive effects of diversity resistance and fluctuating physical environment 1 2008
... 气候变化在全球范围内改变水文的循环, 同时也深刻影响着降雨格局的变化, 越来越多的证据表明全球范围内许多地方强降雨事件的频率和强度都在增强(Mallakpour & Villarini, 2017; Chen et al., 2019).强降雨事件幅度和频率的变化都会引起短暂的水资源脉冲, 对多数生态系统产生影响, 并可能改变群落的可入侵性(Kreyling et al., 2008).陆生生态系统中极端降雨引起的资源可用性变化往往通过促进外来植物入侵对群落结构和组成产生影响(Knapp et al., 2008).降雨事件会使北美干旱的生境中的外来植物大麦状雀麦(Bromus hordeaceus)的丰度大幅增加(Hobbs et al., 2007).在混合型草原中, 相比干旱的生境, 湿润环境中入侵植物的丰度更高, 其被入侵频率也相对更高(Larson et al., 2001).同时, 与有性繁殖的入侵植物相比, 水分脉冲对克隆性入侵植物的促进作用更强, 在一定程度上, 克隆整合对促进外来克隆植物入侵资源条件多变的群落具有潜在的促进作用(Chen et al., 2019). ...
Invasive species interact with climatic variability to reduce success of natives 1 2020
... 尽管从定义上来看, 资源脉冲是短暂性资源过剩的现象, 但资源脉冲可能对自然群落造成长期持续的影响.植物储备器官在资源丰富时期对过剩资源的储备(Noy-Meir, 1973), 可能在之后较长一段时间内作为植物的营养来源, 从而使资源脉冲的影响可以持续很长一段时间.生态系统对资源脉冲的响应幅度也是动态变化的, 其往往受到脉冲的幅度和时间及生态系统本身生产力的影响(Yang et al., 2010).研究表明生态系统的生产力是调节资源波动影响的重要因素, 对生态系统和食物网结构都能产生重要影响(Weber & Brown, 2019).不同的生态系统对资源脉冲的响应也有所不同, 一般水生生态系统对脉冲资源响应的持续时间和滞后时间都要短于陆地生态系统(Yang et al., 2010). ...
Forest disturbances promote invasion of alien herbaceous plants: a comparison of abundance and plant traits between alien and native species in thinned and unthinned stands 2 2020
Addition of phosphorus and nitrogen support the invasiveness of 1 2020
... 一种资源脉冲事件发生的同时, 往往伴随着另外一种或多种类型资源的脉冲事件, 如干旱环境中的降水事件可以同时引起生态系统中的养分和水分脉冲.研究表明土壤养分和水分的可利用性对群落的影响往往存在交互作用.如对内蒙古半干旱草原生态系统的研究证实了氮的有效性和水的有效性相互影响, 从而影响植物养分的吸收过程(Lü & Han, 2010).对加利福尼亚草地生态系统的研究指出草地生态系统对氮沉降的响应将在很大程度上取决于未来的降水模式(Harpole et al., 2007).这种资源动态变化的交互作用也会对生物入侵产生影响, 如营养和水分可利用性之间的相互作用会改变外来入侵种相对于本地植物的生长表现, 进而影响外来植物的成功入侵(Liu et al., 2018); 水分胁迫条件下, 氮和磷资源的添加会促进空心莲子草(Alternanthera philoxeroides)的入侵(Sun et al., 2020).但这些研究更多是关于资源平均水平变化对植物入侵的影响, 有关资源脉冲动态变化以及多种资源脉冲交互作用对外来植物入侵的影响还需进一步研究.加强多资源脉冲交互作用对生物入侵影响的研究, 可以丰富资源脉冲理论框架, 并且很可能会成为未来入侵生态学领域的一个重要方向. ...
High N, dry: experimental nitrogen deposition exacerbates native shrub loss and nonnative plant invasion during extreme drought 2 2017
... 1992年, 美国****Vitousek首次提出“全球环境变化(global environmental change)”的概念, 并指出全球变化终将对地球生态系统进程产生巨大影响.全球变化驱动因素一般包括大气CO2富集、气候变化、土地利用变化、氮沉降和生物入侵等(Vitousek et al., 1996; Bradley et al., 2010a; Sage, 2020).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
... 目前, 大部分关于全球变化背景下外来植物入侵的研究只关注环境因子的稳态变化, 即温度、氮沉降以及CO2浓度的稳定升高对外来植物入侵的影响(Valliere et al., 2017; Vetter et al., 2020).然而, 在自然界中这些环境因子并不是一成不变的, 其发生强度、时间和地点往往受人类活动的影响, 呈现出较大的波动性, 即非稳态变化(Easterling et al., 2000; Meehl & Tebaldi, 2004).在资源供给总量相同的前提下, 相对于环境因子稳态变化导致生态系统中可利用的资源维持在固定水平(图1A), 环境因子的非稳态变化往往导致生态系统中可利用的资源产生波动, 进而形成资源脉冲(resource pulse)(Yang et al., 2008, 图1B).已有研究表明: 生物从个体、种群、群落到生态系统水平, 对资源水平的稳态变化和资源脉冲的生态响应往往存在差别(Yang et al., 2008; Felton & Smith, 2017).因此, 研究资源脉冲对外来植物入侵的影响, 有助于准确评估全球变化对外来植物入侵的驱动作用, 阐明入侵植物在全球变化背景下的入侵机制. ...
The ecology and evolution of alien plants 1 2018
... 人类活动直接或间接地影响着生物的地理分布范围, 使许多生物跨越了地理隔阂, 进入到新的区域(Vitousek et al., 1997; Capinha et al., 2015).全球尺度的研究表明, 在人类活动的影响下, 已有超过1.3万种植物在新的区域建立起能够自我维持的种群, 成为归化种(van Kleunen et al., 2015; Seebens et al., 2017).此外, 最新研究表明外来生物增加的趋势尚未达到饱和, 还在不断增加(Seebens et al., 2017, 2021).部分归化物种迅速扩散, 改变了当地生态系统的结构和功能, 并可能对人类健康和经济发展造成巨大的不利影响, 最终成为入侵生物(invasive alien species)(Early et al., 2016; Paini et al., 2016).因此, 探究外来生物为什么能够成功归化, 以及在此基础上如何进一步形成入侵物种成为生态学研究的热点问题(van Kleunen et al., 2018). ...
Global exchange and accumulation of non-native plants 1 2015
... 人类活动直接或间接地影响着生物的地理分布范围, 使许多生物跨越了地理隔阂, 进入到新的区域(Vitousek et al., 1997; Capinha et al., 2015).全球尺度的研究表明, 在人类活动的影响下, 已有超过1.3万种植物在新的区域建立起能够自我维持的种群, 成为归化种(van Kleunen et al., 2015; Seebens et al., 2017).此外, 最新研究表明外来生物增加的趋势尚未达到饱和, 还在不断增加(Seebens et al., 2017, 2021).部分归化物种迅速扩散, 改变了当地生态系统的结构和功能, 并可能对人类健康和经济发展造成巨大的不利影响, 最终成为入侵生物(invasive alien species)(Early et al., 2016; Paini et al., 2016).因此, 探究外来生物为什么能够成功归化, 以及在此基础上如何进一步形成入侵物种成为生态学研究的热点问题(van Kleunen et al., 2018). ...
A meta-analysis of trait differences between invasive and non-invasive plant species 1 2010
... 群落中资源可用性的变化会影响植物对生物量的分配(Weiner, 2004; Eziz et al., 2017), 如对根茎比和植物组织中碳氮比的影响(Parepa et al., 2019).外来植物入侵成功往往与物种的生态特征和生活史特征密切相关(van Kleunen et al., 2010), 成功的入侵者通常具有快速获取有限资源以及将资源分配给不同的植物器官以提高其表现的能力(Goldberg et al., 1999).一般来说, 入侵植物会具有更高的表型可塑性(Richards et al., 2006; Funk, 2008), 如更高的水分利用效率、光合速率、生长速率以及更大的比叶面积等(Funk, 2013).当资源脉冲事件引起群落中临时性可用资源变化时, 这些机制提高了外来植物在群落中的表现, 进而促进其成功入侵(Harpole et al., 2016). ...
Invader presence disrupts the stabilizing effect of species richness in plant community recovery after drought 2 2020
... 1992年, 美国****Vitousek首次提出“全球环境变化(global environmental change)”的概念, 并指出全球变化终将对地球生态系统进程产生巨大影响.全球变化驱动因素一般包括大气CO2富集、气候变化、土地利用变化、氮沉降和生物入侵等(Vitousek et al., 1996; Bradley et al., 2010a; Sage, 2020).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
... 目前, 大部分关于全球变化背景下外来植物入侵的研究只关注环境因子的稳态变化, 即温度、氮沉降以及CO2浓度的稳定升高对外来植物入侵的影响(Valliere et al., 2017; Vetter et al., 2020).然而, 在自然界中这些环境因子并不是一成不变的, 其发生强度、时间和地点往往受人类活动的影响, 呈现出较大的波动性, 即非稳态变化(Easterling et al., 2000; Meehl & Tebaldi, 2004).在资源供给总量相同的前提下, 相对于环境因子稳态变化导致生态系统中可利用的资源维持在固定水平(图1A), 环境因子的非稳态变化往往导致生态系统中可利用的资源产生波动, 进而形成资源脉冲(resource pulse)(Yang et al., 2008, 图1B).已有研究表明: 生物从个体、种群、群落到生态系统水平, 对资源水平的稳态变化和资源脉冲的生态响应往往存在差别(Yang et al., 2008; Felton & Smith, 2017).因此, 研究资源脉冲对外来植物入侵的影响, 有助于准确评估全球变化对外来植物入侵的驱动作用, 阐明入侵植物在全球变化背景下的入侵机制. ...
Global environmental change: an introduction 1 1992
... 1992年, 美国****Vitousek首次提出“全球环境变化(global environmental change)”的概念, 并指出全球变化终将对地球生态系统进程产生巨大影响.全球变化驱动因素一般包括大气CO2富集、气候变化、土地利用变化、氮沉降和生物入侵等(Vitousek et al., 1996; Bradley et al., 2010a; Sage, 2020).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
Biological invasions as global environmental change 2 1996
... 1992年, 美国****Vitousek首次提出“全球环境变化(global environmental change)”的概念, 并指出全球变化终将对地球生态系统进程产生巨大影响.全球变化驱动因素一般包括大气CO2富集、气候变化、土地利用变化、氮沉降和生物入侵等(Vitousek et al., 1996; Bradley et al., 2010a; Sage, 2020).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
... ).生物入侵作为全球变化的一部分, 往往受到全球变化其他因素的影响(Vitousek et al., 1996).相关研究证实, 环境中上升的CO2浓度可以提高入侵植物相对于本地植物的竞争力(Bradley et al., 2010a, 2010b; Sheppard & Stanley, 2014; Liu et al., 2017; Jabran & Doğan, 2020), 当入侵植物为C3植物时这种效应尤其明显(Sheppard & Stanley, 2014).气候变暖往往通过提高外来植物存活率、加速植物生长以及提供空余生态位等方式促进外来植物入侵(Wolkovich et al., 2013; Zelikova et al., 2013; Blumenthal et al., 2016).此外, 由于入侵植物和本地植物间固有的物候差异(Wolkovich & Cleland, 2011), 降水格局的变化通常更利于入侵植物的生长(Blumenthal et al., 2008; Prevéy & Seastedt, 2014; Walker et al., 2017).农业生产导致的土地格局的变化会在一定程度上加速部分地区外来植物的扩散(Chytrý et al., 2012).氮沉降引起的土壤氮富集会对本地植物产生负面影响, 减少本地植物多样性, 从而促进外来植物入侵(Valliere et al., 2017; Vetter et al., 2020). ...
Human domination of Earthʼs ecosystems//Marzluff JM, Shulenberger E, Endlicher W, Alberti M, Bradley G, Ryan C, Simon U, ZumBrunnen C. Urban Ecology 2 1997
... 人类活动直接或间接地影响着生物的地理分布范围, 使许多生物跨越了地理隔阂, 进入到新的区域(Vitousek et al., 1997; Capinha et al., 2015).全球尺度的研究表明, 在人类活动的影响下, 已有超过1.3万种植物在新的区域建立起能够自我维持的种群, 成为归化种(van Kleunen et al., 2015; Seebens et al., 2017).此外, 最新研究表明外来生物增加的趋势尚未达到饱和, 还在不断增加(Seebens et al., 2017, 2021).部分归化物种迅速扩散, 改变了当地生态系统的结构和功能, 并可能对人类健康和经济发展造成巨大的不利影响, 最终成为入侵生物(invasive alien species)(Early et al., 2016; Paini et al., 2016).因此, 探究外来生物为什么能够成功归化, 以及在此基础上如何进一步形成入侵物种成为生态学研究的热点问题(van Kleunen et al., 2018). ...
 ,
, ,3,5,*
,3,5,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT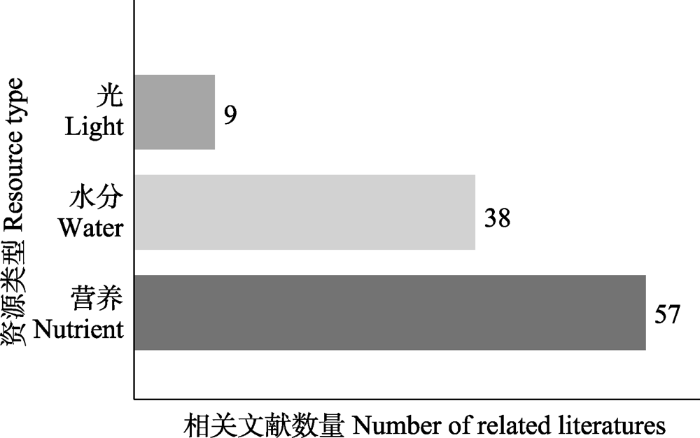 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}