李孝龙1,4, 周俊,,1,*, 彭飞2, 钟宏韬3, Hans LAMBERS31中国科学院、水利部成都山地灾害与环境研究所, 成都 610041 2中国科学院西北生态环境资源研究院沙漠与沙漠化重点实验室, 兰州 730000 3School of Biological Sciences and Institute of Agriculture, University of Western Australia, 35 Stirling Highway, Perth, WA 6009, Australia 4中国科学院大学, 北京 100049
Temporal trends of plant nutrient-acquisition strategies with soil age and their ecological significance
Xiao-Long LI1,4, Jun ZHOU,,1,*, Fei PENG2, Hong-Tao ZHONG3, LAMBERS Hans31Institute of Mountain Hazards and Environment, Chinese Academy of Sciences, Chengdu 610041, China 2Key Laboratory of Desert and Desertification, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou 730000, China 3School of Biological Sciences and Institute of Agriculture, University of Western Australia, 35 Stirling Highway, Perth, WA 6009, Australia 4University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Changes in soil nutrient availability and primary succession of vegetation often co-occur during the processes of natural soil development. A low availability of nitrogen (N) and phosphorus (P) resources is common in the very early and late stage of soil development, respectively. Plants have evolved different nutrient-acquisition strategies (NASs) in response to low nutrient availability. Although the changes and responses of plant NASs to soil nutrients may affect primary succession and species diversity, the temporal trends and underlying mechanisms of plant NASs with soil development remain unknown. We reviewed 104 studies mainly carried out on soil chronosequences to clarify changes in plant NASs with soil age and its ecological significance. We classify plant NASs into Fine root, Microbial, Specialized root, Carnivorous and Parasitic strategies. We argue that the diversity of plant NASs changes with increasing soil age following a dumbbell-pattern, while reaching the maximum in the late stage of soil development. The role of Microbial and Fine root strategies in plants acquiring nutrients gradually decreases with increasing soil age, while the minimum and maximum role of Specialized root strategies in plants acquiring P is in the intermediate and late stages of soil development, respectively. The effects of NASs on interspecific relationships of plants vary with soil age. Specifically, pioneer plants with biological N fixation and specialized root strategies usually increase available soil N and regolith-derived nutrients to facilitate the colonization of subsequent plants in the early stage of soil development. During the early-intermediate stage, NASs mainly affect plant competitiveness in acquiring relatively abundantly available nutrients from soil. The facilitation and competition affected by NASs contribute to plant species turnover in the first two stages. In the late stage, diverse NASs enable plants to acquire distinct forms of nutrients from different soil spaces and complementary NASs enable plants to take up soil nutrients mobilized by their neighbors. Together with the interactions between NASs and soil pathogens, these processes contribute the coexistence and diversity of plant species in this stage when most soil nutrients have a very low availability. We propose that it is necessary to quantify the relationships between changes in soil nutrient availability (including concentrations and fractions) and plant NASs with soil age. More studies are also needed to quantify contributions of NASs to primary succession, diversity of plant species and soil development. Keywords:root traits;mycorrhizal symbiosis;cluster root;rhizosphere exudates;biodiversity;primary succession;pedogenesis;chronosequence
PDF (4320KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 李孝龙, 周俊, 彭飞, 钟宏韬, Hans LAMBERS. 植物养分捕获策略随成土年龄的变化及生态学意义. 植物生态学报, 2021, 45(7): 714-727. DOI: 10.17521/cjpe.2021.0160 LI Xiao-Long, ZHOU Jun, PENG Fei, ZHONG Hong-Tao, Hans LAMBERS. Temporal trends of plant nutrient-acquisition strategies with soil age and their ecological significance. Chinese Journal of Plant Ecology, 2021, 45(7): 714-727. DOI: 10.17521/cjpe.2021.0160
植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008)。根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1)。微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为。特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015)。细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019)。植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002)。寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005)。
Fig. 2Morphology of plant nutrient-acquisition strategies. A, Hyphae of the arbuscular mycorrhizal fungus Glomus caledonium growing into soil from a host root of Trifolium repens (Fabaceae). Photo by Iver Jakobsen; reprinted with permission of Springer-Verlag (Olsson et al., 2002). B, A seedling of Pinus sylvestris (Pinaceae) growing in a microcosm in association with the ectomycorrhizal fungus Suillus bovinus. The fungal mycelium can be seen spreading in the soil in the microcosm (arrowhead) and proliferating locally to form well-defined dense patches (arrows). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). C, Ericoid mycorrhizal root of Woollsia pungens, showing epidermal cells colonized by coils of an ericoid mycorrhizal fungus (stained blue, arrowed). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., 2015). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., 2020). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.
细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020)。细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015)。草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019)。农作物也存在细根策略。与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016)。
特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H)。虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008)。土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011)。缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020)。有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006)。
食虫策略植物捕获养分通常包括3个阶段: 产生诱饵、捕获猎物和消化猎物体内的有机物(Król et al., 2012)。盆栽实验表明, 植物能够消化吸收猎物体内的几丁质和蛋白质, 并且对蛋白质的利用效率更高(Pavlovič et al., 2016); 高山捕虫堇(Pinguicula alpina)能够捕获昆虫并利用其体内的N和P (Aldenius et al., 1983)。目前已发现约有4 000种植物通过寄生策略获取养分, 其受土壤养分的影响相对较小(Press & Phoenix, 2005)。
2 植物NASs随成土年龄的变化
2.1 成土早期的植物NASs
成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3)。多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径。夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002)。新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003)。在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强。例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011)。美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013)。尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3)。例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015)。
Fig. 3Temporal changes in plant nutrient-acquisition strategies (NASs) with increasing soil age. AM, arbuscular mycorrhiza; CR, cluster roots; DR, dauciform roots; ECM, ectomycorrhiza; FR, fine roots; N fixation, nitrogen fixation; OM, orchid mycorrhiza; Ptase, phosphatases; SR, sand-banding roots; VR, vellozioid roots. The change in intensity of the background color represents abundance in diversity of NASs. The width of irregular polygons on the top of the figure represents the general trend of three classes of NASs with soil age. The size of cycles represents the relative importance of a particular NAS in plant acquiring nutrients in different stages of pedogenesis. The dotted circle of FR indicates that more evidence is needed to support the role of the fine root strategy in plant acquiring nutrients. The change in intensity of color in soil nitrogen (N) and phosphorus (P) pool rectangles represents variation in total N and P availability, respectively, with soil age.
2.2 成土中期的植物NASs
成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3)。澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015)。相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002)。新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011)。亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020)。目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015)。由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限。同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3)。
2.3 成土后期的植物NASs
长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016)。由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3)。由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3)。例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015)。在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019)。在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001)。新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011)。
成土后期植物NASs的多样性显著升高。澳大利亚西南部Jurien Bay序列后期, 土壤总P含量降至6-10 mg·kg-1, 用稀有物种评估的植物NASs多样性增加了一倍, 与NASs相关的性状分离度增加了46% (Zemunik et al., 2015)。加拿大养分贫瘠的沿海沙丘序列, 随着土壤年龄增加, 植物NASs的Simpson指数增加超过1.5倍(Roy-Bolduc et al., 2016)。成土过程中总P的减少是导致植物NASs多样性增加的原因之一。澳大利亚西南部Jurien Bay土壤序列的总P含量与植物NASs多样性呈负相关关系, 总P的减少使植物产生多种NASs以获取土壤中的P (Zemunik et al., 2015)。在澳大利亚西南部总P为50 mg·kg-1左右的土壤中, Xylomelum occidentale发育了特别的NASs, 既不释放大量有机酸也不形成菌根, 而是释放磷酸酶水解土壤中的RNA和磷脂(Zhong et al., 2021), 增加了成土后期NASs的丰富度。
自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015)。NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化。互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019)。
3.1 成土早期的NASs与先锋群落建立
成土早期第一阶段, 植物NASs主要通过改善土壤养分条件和启动养分循环促进先锋群落形成(图4)。这一阶段中有效N、P的含量均很低, 具有固N能力的植物最先定居, 增加土壤N库和其他矿质养分的有效性, 改善土壤养分状况, 为后续植物的定居和繁殖提供条件。例如, 在贡嘎山海螺沟冰川退缩后仅几年的冰碛物上, 先锋固N植物黄芪(Astragalus mahoshanicus)可为邻近的非固N植物川滇柳(Salix rehderiana)提供N源, 促进川滇柳的生长, 川滇柳取代黄芪成为该演替序列下一阶段的优势物种(Wang et al., 2021)。新西兰Kanakana Dome高原发生的原生演替中, 固N灌木Coriaria arborea使有机层土壤的N库增加了10倍, 促进了后续物种Griselinia littoralis的定居(Walker et al., 2003)。美国俄勒冈州温带雨林的红桤木(Alnus rubra)的根瘤固N策略不仅增加了土壤N库, 而且大量被固定的N经硝化作用后溶解土壤矿物, 提高了土壤中P和钙(Ca)等其他矿质养分的有效性(Perakis & Pett-Ridge, 2019)。一些具有特殊根系策略的植物也能够通过风化矿物来提升土壤矿质养分水平。巴西东南部生长于岩石上的翡若翠科植物可发育毛状根(VR), 并释放大量羧化物溶解硅酸盐矿物独居石中的P等矿质养分, 改善土壤养分条件(Augusto et al., 2019; Teodoro et al., 2019)。日本Honshu岛废弃采石场形成的5-15 cm厚的薄层土壤上, 菌根和根瘤固N策略促进了先锋植物豆科短梗胡枝子(Lespedeza cyrtobotrya)的生长和定居(Maki et al., 2008)。Muñoz等(2021)总结火山沉积物上发生的原生演替, 发现先锋植物形成根瘤吸收更多的N, 形成菌根或释放有机酸吸收更多的P, 所以能够在贫瘠土壤上最先定居。因此, 在成土早期第一阶段, 植物NASs主要通过微生物和特殊根系等策略提升植物的养分水平和土壤的有效养分库, 并通过促进作用影响先锋植物群落的形成(图4)。
Fig. 4Conceptual scheme showing effects of plant nutrient-acquisition strategies (NASs) on vegetation primary succession and coexistence and diversity of species. The change in intensity of color in soil nitrogen (N) and phosphorus (P) pool rectangle represents variations in total N and P pool with soil age, respectively. In the first stage of pedogenesis, plant NASs contribute to the establishment of pioneer community through facilitation. For example, N fixation and carboxylate-releasing strategies improve soil available nutrient pools and thus facilitate the settlement, growth and reproduction of subsequent species. In the early-intermediate stage, there are adequate soil available nutrients, NASs contribute to the competitiveness of plant species. Species with more effective NASs may have advantages in competition. And thus this will result in species turnover and promote succession. In the late stage, soil available nutrients are low or impoverished, especially for P, plant have to develop diverse NASs to acquire limited nutrients. The relationships between these NASs are mainly complementary. Diverse and complementary NASs have important impacts on the coexistence and diversity in the late stage of pedogenesis, although we still do not know the exact contribution to them. In addition, as proposed by Lambers et al. (2018), the different abilities against pathogens of plant species with different NASs also contribute the coexistence of plant species at this stage.
AbrahãoA, deBritto Costa P, LambersH, AndradeSAL, SawayaACHF, RyanMH, OliveiraRS (2019). Soil types select for plants with matching nutrient-acquisition and -use traits in hyperdiverse and severely nutrient-impoverished campos rupestres and cerrado in Central Brazil Journal of Ecology, 107, 1302-1316. DOI:10.1111/jec.2019.107.issue-3URL [本文引用: 1]
TeodoroGS, LambersH, NascimentoDL, AdriánLópez de Andrade S, RyanMH, SilvaOliveira R (2020). Vellozioid roots allow for habitat specialization among rock- and soil-dwelling Velloziaceae in campos rupestres Functional Ecology, 34, 442-457. DOI:10.1111/fec.v34.2URL [本文引用: 3]
AbrahãoA, LambersH, SawayaACHF, MazzaferaP, OliveiraRS (2014). Convergence of a specialized root trait in plants from nutrient-impoverished soils: phosphorus-acquisition strategy in a nonmycorrhizal cactus Oecologia, 176, 345-355. DOI:10.1007/s00442-014-3033-4PMID:25135179 [本文引用: 1] In old, phosphorus (P)-impoverished habitats, root specializations such as cluster roots efficiently mobilize and acquire P by releasing large amounts of carboxylates in the rhizosphere. These specialized roots are rarely mycorrhizal. We investigated whether Discocactus placentiformis (Cactaceae), a common species in nutrient-poor campos rupestres over white sands, operates in the same way as other root specializations. Discocactus placentiformis showed no mycorrhizal colonization, but exhibited a sand-binding root specialization with rhizosheath formation. We first provide circumstantial evidence for carboxylate exudation in field material, based on its very high shoot manganese (Mn) concentrations, and then firm evidence, based on exudate analysis. We identified predominantly oxalic acid, but also malic, citric, lactic, succinic, fumaric, and malonic acids. When grown in nutrient solution with P concentrations ranging from 0 to 100 μM, we observed an increase in total carboxylate exudation with decreasing P supply, showing that P deficiency stimulated carboxylate release. Additionally, we tested P solubilization by citric, malic and oxalic acids, and found that they solubilized P from the strongly P-sorbing soil in its native habitat, when the acids were added in combination and in relatively low concentrations. We conclude that the sand-binding root specialization in this nonmycorrhizal cactus functions similar to that of cluster roots, which efficiently enhance P acquisition in other habitats with very low P availability.
AdamczykB, SietiöOM, BiasiC, HeinonsaloJ (2019). Interaction between tannins and fungal necromass stabilizes fungal residues in boreal forest soils New Phytologist, 223, 16-21. DOI:10.1111/nph.15729PMID:30721536 [本文引用: 1]
AdamecL (2002). Leaf absorption of mineral nutrients in carnivorous plants stimulates root nutrient uptake New Phytologist, 155, 89-100. DOI:10.1046/j.1469-8137.2002.00441.xURL [本文引用: 1]
Ahmad-RamliMF, CornulierT, JohnsonD (2013). Partitioning of soil phosphorus regulates competition between Vaccinium vitis-idaea and Deschampsia cespitosa Ecology and Evolution, 3, 4243-4252. DOI:10.1002/ece3.2013.3.issue-12URL [本文引用: 1]
AlbornozFE, BurgessTI, LambersH, EtchellsH, LalibertéE (2017). Native soilborne pathogens equalize differences in competitive ability between plants of contrasting nutrient- acquisition strategies Journal of Ecology, 105, 549-557. DOI:10.1111/jec.2017.105.issue-2URL [本文引用: 1]
AlbornozFE, LambersH, TurnerBL, TesteFP, LalibertéE (2016). Shifts in symbiotic associations in plants capable of forming multiple root symbioses across a long-term soil chronosequence Ecology and Evolution, 6, 2368-2377. DOI:10.1002/ece3.2016.6.issue-8URL [本文引用: 2]
AldeniusJ, CarlssonB, KarlssonS (1983). Effects of insect trapping on growth and nutrient content of Pinguicula vulgaris L. in relation to the nutrient content of the substrate New Phytologist, 93, 53-59. DOI:10.1111/nph.1983.93.issue-1URL [本文引用: 1]
AugustoL, FaninN, BakkerMR (2019). When plants eat rocks: functional adaptation of roots on rock outcrops Functional Ecology, 33, 760-761. DOI:10.1111/fec.2019.33.issue-5URL [本文引用: 2]
BahadurA, JinZC, LongXL, JiangSJ, ZhangQ, PanJB, LiuYJ, FengHY (2019). Arbuscular mycorrhizal fungi alter plant interspecific interaction under nitrogen fertilization European Journal of Soil Biology, 93, 103094. DOI: 10.1016/j.ejsobi.2019.103094. URL [本文引用: 1]
CabalC, Martínez-GarcíaR, deCastro Aguilar A, ValladaresF, PacalaSW (2020). The exploitative segregation of plant roots Science, 370, 1197-1199. DOI:10.1126/science.aba9877URL [本文引用: 1]
CaplanJS, MeinersSJ, Flores-MorenoH, McCormackML (2019). Fine-root traits are linked to species dynamics in a successional plant community Ecology, 100, e02588. DOI: 10.1002/ecy.2588. [本文引用: 1]
CeulemansT, BodéS, BollynJ, HarpoleS, CoorevitsK, PeetersG, vanAcker K, SmoldersE, BoeckxP, HonnayO (2017). Phosphorus resource partitioning shapes phosphorus acquisition and plant species abundance in grasslands Nature Plants, 3, 16224. DOI: 10.1038/nplants.2016.224. PMID:28134925 [本文引用: 1] Species diversity is commonly hypothesized to result from trade-offs for different limiting resources, providing separate niches for coexisting species. As soil nutrients occur in multiple chemical forms, plant differences in acquisition of the same element derived from different compounds may represent unique niche dimensions. Because plant productivity of ecosystems is often limited by phosphorus, and because plants have evolved diverse adaptations to acquire soil phosphorus, a promising yet untested hypothesis is phosphorus resource partitioning. Here, we provided two different chemical forms of phosphorus to sown grassland mesocosms to investigate phosphorus acquisition of eight plant species that are common in European grasslands, and to identify subsequent patterns of plant abundance. For the first time, we show that the relative abundance of grassland plant species can be influenced by soil phosphorus forms, as higher abundance was linked to higher acquisition of a specific form of phosphorus. These results were supported by a subsequent isotope dilution experiment using intact grassland sods that were treated with different inorganic or organic phosphorus forms. Here, 5 out of 14 species showed greater phosphorus acquisition in the inorganic phosphorus treatment, and 4 in the organic phosphorus treatments. Furthermore, for the species used in both experiments we found similar acquisition patterns. Our results support the hypothesis of phosphorus resource partitioning and may provide a new mechanistic framework to explain high plant diversity in phosphorus-poor ecosystems. As world biodiversity hotspots are almost invariably related to phosphorus limitation, our results may thus also be key to understanding biodiversity loss in an era of ever-increasing nutrient enrichment.
ChadwickOA, DerryLA, VitousekPM, HuebertBJ, HedinLO (1999). Changing sources of nutrients during four million years of ecosystem development Nature, 397, 491-497. DOI:10.1038/17276URL [本文引用: 1]
ChenJH, ZhouDS, NiuM, BiePF, XieTZ, ZhaoR, MuCL (2020). Comparative analysis on the fine root traits of the four native broad-leaved trees in the hilly region of central Sichuan Province Journal of Nanjing Forestry University (Natural Sciences Edition), 44, 31-38. [本文引用: 1]
ChenMM, ChenBD, WangXJ, ZhuYG, WangYS (2009). Influences of arbuscular mycorrhizal fungi (AMF) on the growth and ecological stoichiometry of clover and ryegrass grown in monoculture or in mixture at different phosphorus (P) levels Acta Ecologica Sinica, 29, 1980-1986. [本文引用: 1]
CrewsTE, KitayamaK, FownesJH, RileyRH, HerbertDA, Mueller-DomboisD, VitousekPM (1995). Changes in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii Ecology, 76, 1407-1424. DOI:10.2307/1938144URL [本文引用: 2]
EissenstatDM (1992). Costs and benefits of constructing roots of small diameter Journal of Plant Nutrition, 15, 763-782. DOI:10.1080/01904169209364361URL [本文引用: 1]
HayesP, TurnerBL, LambersH, LalibertéE (2014). Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence Journal of Ecology, 102, 396-410. DOI:10.1111/1365-2745.12196URL [本文引用: 1]
HoldawayRJ, RichardsonSJ, DickieIA, PeltzerDA, CoomesDA (2011). Species- and community-level patterns in fine root traits along a 120000-year soil chronosequence in temperate rain forest Journal of Ecology, 99, 954-963. DOI:10.1111/jec.2011.99.issue-4URL [本文引用: 3]
JiWL, LiWZ, ZhuKQ, MartinJL (2008). Review of the biological characteristics of dauciform roots Acta Prataculturae Sinica, (2), 118-125. [本文引用: 1]
JiangJ, SongMH (2010). Review of the roles of plants and soil microorganisms in regulating ecosystem nutrient cycling Chinese Journal of Plant Ecology, 34, 979-988. [本文引用: 1]
KellerAB, BrzostekER, CraigME, FisherJB, PhillipsRP (2021). Root-derived inputs are major contributors to soil carbon in temperate forests, but vary by mycorrhizal type Ecology Letters, 24, 626-635. DOI:10.1111/ele.v24.4URL [本文引用: 1]
KnoblochováT, KohoutP, PüschelD, DoubkováP, FrouzJ, CajthamlT, KuklaJ, VosátkaM, RydlováJ (2017). Asymmetric response of root-associated fungal communities of an arbuscular mycorrhizal grass and an ectomycorrhizal tree to their coexistence in primary succession Mycorrhiza, 27, 775-789. DOI:10.1007/s00572-017-0792-xPMID:28752181 [本文引用: 2] The arbuscular mycorrhizal (AM) grass Calamagrostis epigejos and predominantly ectomycorrhizal (EcM) tree Salix caprea co-occur at post-mining sites spontaneously colonized by vegetation. During succession, AM herbaceous vegetation is replaced by predominantly EcM woody species. To better understand the interaction of AM and EcM plants during vegetation transition, we studied the reciprocal effects of these species' coexistence on their root-associated fungi (RAF). We collected root and soil samples from three different microenvironments: stand of C. epigejos, under S. caprea canopy, and contact zone where roots of the two species interacted. RAF communities and mycorrhizal colonization were determined in sampled roots, and the soil was tested for EcM and AM inoculation potentials. Although the microenvironment significantly affected composition of the RAF communities in both plant species, the effect was greater in the case of C. epigejos RAF communities than in that of S. caprea RAF communities. The presence of S. caprea also significantly decreased AM fungal abundance in soil as well as AM colonization and richness of AM fungi in C. epigejos roots. Changes observed in the abundance and community composition of AM fungi might constitute an important factor in transition from AM-dominated to EcM-dominated vegetation during succession.
KoideRT, KabirZ (2000). Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate New Phytologist, 148, 511-517. DOI:10.1046/j.1469-8137.2000.00776.xPMID:33863024 [本文引用: 1] Organic phosphorus sources make up a large fraction of the total P in some soils. Vesicular-arbuscular mycorrhizal fungi provide a large surface area for the absorption of inorganic P. The question of whether or not they have direct access to organic P by producing extracellular phosphatases has hitherto been controversial because experiments had not been performed in the absence of other soil microorganisms. We used a split-dish in vitro carrot mycorrhiza system free from contaminating microorganisms. The extraradical hyphae of Glomus intraradices hydrolysed both 5-bromo-4-chloro-3-indolyl phosphate and phenolphthalein diphosphate. Moreover, they transferred significantly more P to roots when they had access to inositol hexaphosphoric acid (phytate) than when they did not. Thus we show unequivocally that extraradical hyphae of G. intraradices can hydrolyse organic P, and, further, that the resultant inorganic P can be taken up and transported to host roots.
KrólE, PłachnoBJ, AdamecL, StolarzM, DziubińskaH, TrębaczK (2012). Quite a few reasons for calling carnivores “the most wonderful plants in the world” Annals of Botany, 109, 47-64. DOI:10.1093/aob/mcr249URL [本文引用: 1]
LambersH, AlbornozF, KotulaL, LalibertéE, RanathungeK, TesteFP, ZemunikG (2018). How belowground interactions contribute to the coexistence of mycorrhizal and non- mycorrhizal species in severely phosphorus-impoverished hyperdiverse ecosystems Plant and Soil, 424, 11-33. DOI:10.1007/s11104-017-3427-2URL [本文引用: 4]
LambersH, BrundrettMC, RavenJA, HopperSD (2011). Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies Plant and Soil, 348, 7-27. DOI:10.1007/s11104-011-0977-6URL [本文引用: 3]
LambersH, ShaneMW, CramerMD, PearseSJ, VeneklaasEJ (2006). Root structure and functioning for efficient acquisition of phosphorus: matching morphological and physiological traits Annals of Botany, 98, 693-713. PMID:16769731 [本文引用: 1] Global phosphorus (P) reserves are being depleted, with half-depletion predicted to occur between 2040 and 2060. Most of the P applied in fertilizers may be sorbed by soil, and not be available for plants lacking specific adaptations. On the severely P-impoverished soils of south-western Australia and the Cape region in South Africa, non-mycorrhizal species exhibit highly effective adaptations to acquire P. A wide range of these non-mycorrhizal species, belonging to two monocotyledonous and eight dicotyledonous families, produce root clusters. Non-mycorrhizal species with root clusters appear to be particularly effective at accessing P when its availability is extremely low.There is a need to develop crops that are highly effective at acquiring inorganic P (Pi) from P-sorbing soils. Traits such as those found in non-mycorrhizal root-cluster-bearing species in Australia, South Africa and other P-impoverished environments are highly desirable for future crops. Root clusters combine a specialized structure with a specialized metabolism. Native species with such traits could be domesticated or crossed with existing crop species. An alternative approach would be to develop future crops with root clusters based on knowledge of the genes involved in development and functioning of root clusters.Root clusters offer enormous potential for future research of both a fundamental and a strategic nature. New discoveries of the development and functioning of root clusters in both monocotyledonous and dicotyledonous families are essential to produce new crops with superior P-acquisition traits.
LiHB, LiuBT, McCormackML, MaZQ, GuoDL (2017). Diverse belowground resource strategies underlie plant species coexistence and spatial distribution in three grasslands along a precipitation gradient New Phytologist, 216, 1140-1150. DOI:10.1111/nph.2017.216.issue-4URL [本文引用: 1]
LiuBT, LiHB, ZhuB, KoideRT, EissenstatDM, GuoDL (2015). Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species New Phytologist, 208, 125-136. DOI:10.1111/nph.2015.208.issue-1URL [本文引用: 3]
LyuY, TangHL, LiHG, ZhangFS, RengelZ, WhalleyWR, ShenJB (2016). Major crop species show differential balance between root morphological and physiological responses to variable phosphorus supply Frontiers in Plant Science, 7, 1939. DOI: 10.3389/fpls.2016.01939. [本文引用: 2]
MaZQ, GuoDL, XuXL, LuMZ, BardgettRD, EissenstatDM, McCormackML, HedinLO (2018). Evolutionary history resolves global organization of root functional traits Nature, 555, 94-97. DOI:10.1038/nature25783URL [本文引用: 1]
MakiT, NomachiM, YoshidaS, EzawaT (2008). Plant symbiotic microorganisms in acid sulfate soil: significance in the growth of pioneer plants Plant and Soil, 310, 55-65. DOI:10.1007/s11104-008-9628-yURL [本文引用: 1]
MasudaG, MaruyamaH, LambersH, WasakiJ (2021). Formation of dauciform roots by Japanese native Cyperaceae and their contribution to phosphorus dynamics in soils Plant and Soil, 461, 107-118. DOI:10.1007/s11104-020-04565-6URL [本文引用: 2]
MayfieldMM, LevineJM (2010). Opposing effects of competitive exclusion on the phylogenetic structure of communities Ecology Letters, 13, 1085-1093. DOI:10.1111/j.1461-0248.2010.01509.xURL [本文引用: 1]
MiaoY, WuHF, MaCD, KongDL (2013). Relationship between mycorrhizal fungi and functional traits in absorption roots: researchprogress and synthesis Chinese Journal of Plant Ecology, 37, 1035-1042. DOI:10.3724/SP.J.1258.2013.00107URL [本文引用: 1]
MulerAL, OliveiraRS, LambersH, VeneklaasEJ (2014). Does cluster-root activity benefit nutrient uptake and growth of co-existing species Oecologia, 174, 23-31. DOI:10.1007/s00442-013-2747-zPMID:23934064 [本文引用: 1] Species that inhabit phosphorus- (P) and micronutrient-impoverished soils typically have adaptations to enhance the acquisition of these nutrients, for example cluster roots in Proteaceae. However, there are several species co-occurring in the same environment that do not produce similar specialised roots. This study aims to investigate whether one of these species (Scholtzia involucrata) can benefit from the mobilisation of P or micronutrients by the cluster roots of co-occurring Banksia attenuata, and also to examine the response of B. attenuata to the presence of S. involucrata. We conducted a greenhouse experiment, using a replacement series design, where B. attenuata and S. involucrata shared a pot at proportions of 2:0, 1:2 and 0:4. S. involucrata plants grew more in length, were heavier and had higher manganese (Mn) concentrations in their young leaves when grown next to one individual of B. attenuata and one individual of S. involucrata than when grown with three conspecifics. All S. involucrata individuals were colonised by arbuscular mycorrhizal fungi, and possibly Rhizoctonia. Additionally, P concentration was higher in the young leaves of B. attenuata when grown with another B. attenuata than when grown with two individuals of S. involucrata, despite the smaller size of the S. involucrata individuals. Our results demonstrate that intraspecific competition was stronger than interspecific competition for S. involucrata, but not for B. attenuata. We conclude that cluster roots of B. attenuata facilitate the acquisition of nutrients by neighbouring shrubs by making P and Mn more available for their neighbours.
MuñozG, OrlandoJ, Zuñiga-FeestA (2021). Plants colonizing volcanic deposits: root adaptations and effects on rhizosphere microorganisms Plant and Soil, 461, 265-279. DOI:10.1007/s11104-020-04783-yURL [本文引用: 1]
NastoMK, Alvarez-ClareS, LekbergY, SullivanBW, TownsendAR, ClevelandCC (2014). Interactions among nitrogen fixation and soil phosphorus acquisition strategies in lowland tropical rain forests Ecology Letters, 17, 1282-1289. DOI:10.1111/ele.2014.17.issue-10URL [本文引用: 1]
NaveLE, NadelhofferKJ, MoineJM, DiepenLTA, CoochJK, DykeNJ (2013). Nitrogen uptake by trees and mycorrhizal fungi in a successional northern temperate forest: insights from multiple isotopic methods Ecosystems, 16, 590-603. DOI:10.1007/s10021-012-9632-1URL [本文引用: 1]
NewmanGS, CobleAA, HaskinsKE, KowlerAL, HartSC (2020). The expanding role of deep roots during long-term terrestrial ecosystem development Journal of Ecology, 108, 2256-2269. DOI:10.1111/jec.v108.6URL [本文引用: 1]
NurfadilahS, SwartsND, DixonKW, LambersH, MerrittDJ (2013). Variation in nutrient-acquisition patterns by mycorrhizal fungi of rare and common orchids explains diversification in a global biodiversity hotspot Annals of Botany, 111, 1233-1241. DOI:10.1093/aob/mct064PMID:23532043 [本文引用: 3] Many terrestrial orchids have an obligate requirement for mycorrhizal associations to provide nutritional support from germination to establishment. This study will investigate the ability of orchid mycorrhizal fungi (OMF) to utilize a variety of nutrient sources in the nutrient-impoverished (low organic) soils of the Southwest Australian Floristic Region (SWAFR) in order to effectively compete, survive and sustain the orchid host.Mycorrhizal fungi representing key OMF genera were isolated from three common and widespread species: Pterostylis recurva, Caladenia flava and Diuris corymbosa, and one rare and restricted species: Drakaea elastica. The accessibility of specific nutrients was assessed by comparing growth including dry biomass of OMF in vitro on basal CN MMN liquid media.Each of the OMF accessed and effectively utilized a wide variety of nutrient compounds, including carbon (C) sources, inorganic and organic nitrogen (N) and inorganic and organic phosphorus (P). The nutrient compounds utilized varied between the genera of OMF, most notably sources of N.These results suggest that OMF can differentiate between niches (micro-niche specialization) in a constrained, highly resource-limited environment such as the SWAFR. Phosphorus is the most limited macronutrient in SWAFR soils and the ability to access phytate by OMF indicates a characterizing functional capacity of OMF from the SWAFR. Furthermore, compared with OMF isolated from the rare D. elastica, OMF associating with the common P. recurva produced far greater biomass over a wider variety of nutritional sources. This suggests a broader tolerance for habitat variation providing more opportunities for the common orchid for recruitment and establishment at a site.
PanFJ, LiangYM, WangKL, ZhangW (2018). Responses of fine root functional traits to soil nutrient limitations in a karst ecosystem of southwest China Forests, 9, 743. DOI: 10.3390/f9120743. URL [本文引用: 1]
PearsonHL, VitousekPM (2002). Soil phosphorus fractions and symbiotic nitrogen fixation across a substrate-age gradient in Hawaii Ecosystems, 5, 587-596. DOI:10.1007/s10021-002-0172-yURL [本文引用: 5]
PerakisSS, Pett-RidgeJC (2019). Nitrogen-fixing red alder trees tap rock-derived nutrients Proceedings of the National Academy of Sciences of the United States of America, 116, 5009-5014. DOI:10.1073/pnas.1814782116PMID:30804181 [本文引用: 1] Symbiotic nitrogen (N)-fixing trees supply significant N inputs to forest ecosystems, leading to increased soil fertility, forest growth, and carbon storage. Rapid growth and stoichiometric constraints of N fixers also create high demands for rock-derived nutrients such as phosphorus (P), while excess fixed N can generate acidity and accelerate leaching of rock-derived nutrients such as calcium (Ca). This ability of N-fixing trees to accelerate cycles of Ca, P, and other rock-derived nutrients has fostered speculation of a direct link between N fixation and mineral weathering in terrestrial ecosystems. However, field evidence that N-fixing trees have enhanced access to rock-derived nutrients is lacking. Here we use strontium (Sr) isotopes as a tracer of nutrient sources in a mixed-species temperate rainforest to show that N-fixing trees access more rock-derived nutrients than nonfixing trees. The N-fixing tree red alder (), on average, took up 8 to 18% more rock-derived Sr than five co-occurring nonfixing tree species, including two with high requirements for rock-derived nutrients. The increased access to rock-derived nutrients occurred despite spatial variation in community-wide Sr sources across the forest, and only N fixers had foliar Sr isotopes that differed significantly from soil exchangeable pools. We calculate that increased uptake of rock-derived nutrients by N-fixing alder requires a 64% increase in weathering supply of nutrients over nonfixing trees. These findings provide direct evidence that an N-fixing tree species can also accelerate nutrient inputs from rock weathering, thus increasing supplies of multiple nutrients that limit carbon uptake and storage in forest ecosystems.
PhoenixGK, JohnsonDA, MuddimerSP, LeakeJR, CameronDD (2020). Niche differentiation and plasticity in soil phosphorus acquisition among co-occurring plants Nature Plants, 6, 349-354. DOI:10.1038/s41477-020-0624-4PMID:32203292 [本文引用: 2] How species coexist despite competing for the same resources that are in limited supply is central to our understanding of the controls on biodiversity. Resource partitioning may facilitate coexistence, as co-occurring species use different sources of the same limiting resource. In plant communities, however, direct evidence for partitioning of the commonly limiting nutrient, phosphorus (P), has remained scarce due to the challenges of quantifying P?acquisition from its different chemical forms present in soil. To address this, we used P to directly trace P?uptake from DNA, orthophosphate and calcium phosphate into monocultures and mixed communities of plants growing in grassland soil. We show that co-occurring plants acquire P from these important organic and mineral sources in different proportions, and that differences in P source use are consistent with the species' root adaptations for P?acquisition. Furthermore, the net benefit arising from niche plasticity (the gain in P?uptake for a species in a mixed community compared to monoculture) correlates with species abundance in the wild, suggesting that niche plasticity for P is a driver of community structure. This evidence for P?resource partitioning and niche plasticity may explain the high levels of biodiversity frequently found in P-limited ecosystems worldwide.
PlaystedCWS, JohnstonME, RamageCM, EdwardsDG, CawthrayGR, LambersH (2006). Functional significance of dauciform roots: exudation of carboxylates and acid phosphatase under phosphorus deficiency in Caustis blakei (Cyperaceae ). New Phytologist, 170, 491-500. PMID:16626471 [本文引用: 2] Caustis blakei produces an intriguing morphological adaptation by inducing dauciform roots in response to phosphorus (P) deficiency. We tested the hypothesis that these hairy, swollen lateral roots play a similar role to cluster roots in the exudation of organic chelators and ectoenzymes known to aid the chemical mobilization of sparingly available soil nutrients, such as P. Dauciform-root development and exudate composition (carboxylates and acid phosphatase activity) were analysed in C. blakei plants grown in nutrient solution under P-starved conditions. The distribution of dauciform roots in the field was determined in relation to soil profile depth and matrix. The percentage of dauciform roots of the entire root mass was greatest at the lowest P concentration ([P]) in solution, and was suppressed with increasing solution [P], while in the field dauciform roots were predominantely located in the upper soil horizons, and decreased with increasing soil depth. Citrate was the major carboxylate released in an exudative burst from mature dauciform roots, which also produced elevated levels of acid phosphatase activity. Malonate was the dominant internal carboxylate present, with the highest concentration in young dauciform roots. The high concentration of carboxylates and phosphatases released from dauciform roots, combined with their prolific distribution in the organic surface layer of nutrient-impoverished soils, provides an ecophysiological advantage for enhancing nutrient acquisition.
PressMC, PhoenixGK (2005). Impacts of parasitic plants on natural communities New Phytologist, 166, 737-751. [本文引用: 2]
QuMH, YuYC, LiS, ZhangJC (2019). Advances in research on activation of mineral nutrients by arbuscular mycorrhizal fungi Journal of Zhejiang A&F University, 36, 394-405. [本文引用: 1]
Roy-BolducA, LalibertéE, BoudreauS, HijriM (2016). Strong linkage between plant and soil fungal communities along a successional coastal dune system FEMS Microbiology Ecology, 92, fiw156. DOI: 10.1093/femsec/fiw156. [本文引用: 2]
RyanMH, TibbettM, Edmonds-TibbettT, SuriyagodaLDB, LambersH, CawthrayGR, PangJ (2012). Carbon trading for phosphorus gain: the balance between rhizosphere carboxylates and arbuscular mycorrhizal symbiosis in plant phosphorus acquisition Plant, Cell & Environment, 35, 2170-2180. [本文引用: 1]
SeeCR, LukeMcCormack M, HobbieSE, Flores-MorenoH, SilverWL, KennedyPG (2019). Global patterns in fine root decomposition: climate, chemistry, mycorrhizal association and woodiness Ecology Letters, 22, 946-953. DOI:10.1111/ele.2019.22.issue-6URL [本文引用: 1]
SemchenkoM, LepikA, AbakumovaM, ZobelK (2018). Different sets of belowground traits predict the ability of plant species to suppress and tolerate their competitors Plant and Soil, 424, 157-169. DOI:10.1007/s11104-017-3282-1URL [本文引用: 1]
SmithSE, AndersonIC, SmithFA (2015). Mycorrhizal associations and phosphorus acquisition: from cells to ecosystems //Plaxton WC, Lambers H. Annual Plant Reviews Volume 48: Phosphorus Metabolism in Plants. John Wiley & Sons, Hoboken, USA. 409-439. [本文引用: 2]
SmithSE, SmithFA, JakobsenI (2004). Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake New Phytologist, 162, 511-524. DOI:10.1111/nph.2004.162.issue-2URL [本文引用: 1]
SongYC, LiXL, FengG (2001). Effect of phosphatase activity on soil organic phosphorus loss in the environment of clover growth Acta Ecologica Sinica, 21, 1130-1135. [本文引用: 1]
SongMY, YuL, FuSL, KorpelainenH, LiCY (2020). Stoichiometric flexibility and soil bacterial communities respond to nitrogen fertilization and neighbor competition at the early stage of primary succession Biology and Fertility of Soils, 56, 1121-1135. DOI:10.1007/s00374-020-01495-4URL [本文引用: 1]
TedersooL, BahramM, ZobelM (2020). How mycorrhizal associations drive plant population and community biology Science, 367, eaba1223. DOI: 10.1126/science.aba1223. URL [本文引用: 2]
TeodoroGS, LambersH, NascimentoDL, de Britto CostaP, Flores-BorgesDNA, AbrahãoA, MayerJLS, SawayaACHF, LadeiraFSB, AbdalaDB, PérezCA, OliveiraRS (2019). Specialized roots of Velloziaceae weather quartzite rock while mobilizing phosphorus using carboxylates Functional Ecology, 33, 762-773. [本文引用: 4]
TesteFP, KardolP, TurnerBL, WardleDA, ZemunikG, RentonM, LalibertéE (2017). Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands Science, 355, 173-176. DOI:10.1126/science.aai8291URL [本文引用: 2]
TesteFP, LalibertéE, LambersH, AuerY, KramerS, KandelerE (2016). Mycorrhizal fungal biomass and scavenging declines in phosphorus-impoverished soils during ecosystem retrogression Soil Biology & Biochemistry, 92, 119-132. DOI:10.1016/j.soilbio.2015.09.021URL [本文引用: 2]
TesteFP, DixonKW, LambersH, ZhouJ, VeneklaasEJ (2020). The potential for phosphorus benefits through root placement in the rhizosphere of phosphorus-mobilising neighbours Oecologia, 193, 843-855. DOI:10.1007/s00442-020-04733-6URL [本文引用: 1]
TianZM, LiCJ, WangC, ZhaoZJ (2000). Comparative studies on organic acid exudation by tips of proteoid and non proteoid roots of phosphorus deficient white lupin Plant Physiology Journal, 36, 317-322. [本文引用: 1]
TurnerBL, CondronLM (2013). Pedogenesis, nutrient dynamics, and ecosystem development: the legacy of T. W. Walker and J. K. Syers Plant and Soil, 367, 1-10. DOI:10.1007/s11104-013-1750-9URL [本文引用: 1]
VitousekPM, ShearerG, KohlDH (1989). Foliar 15N natural abundance in Hawaiian rainforest: patterns and possible mechanisms Oecologia, 78, 383-388. DOI:10.1007/BF00379113PMID:28312585 [本文引用: 1] Foliar samples were obtained from symbiotic nitrogen-fixers and control plants (non-fixers) along elevational and primary successional gradients in volcanic sites in Hawai'i. Most control plants had negative δN values (range-10.1 to +0.7‰), while most nitrogen-fixers were near 0‰. Foliar N in the native tree Metrosideros polymorpha did not vary with elevation (from sea level to tree-line), but it did increase substantially towards 0‰ on older soils. The soil in an 197-yr-old site had a δN value of approximately-2‰, while in a ~67000-yr-old site it was +3.6‰. We suggest that inputs of N-depleted nitrogen from precipitation coupled with very low nitrogen outputs cause the strongly negative δN values in non-nitrogen-fixing plants on early successional sites.
WaddellHA, SimpsonRJ, RyanMH, LambersH, GardenDL, RichardsonAE (2017). Root morphology and its contribution to a large root system for phosphorus uptake by Rytidosperma species (wallaby grass ). Plant and Soil, 412, 7-19. DOI:10.1007/s11104-016-2933-yURL [本文引用: 1]
WalkerLR, ClarksonBD, SilvesterWB, ClarksonBR (2003). Colonization dynamics and facilitative impacts of a nitrogen- fixing shrub in primary succession Journal of Vegetation Science, 14, 277-290. DOI:10.1111/jvs.2003.14.issue-2URL [本文引用: 4]
WangCG, HanSJ, ZhouYM, ZhangJH, ZhengXB, DaiGH, LiMH (2016). Fine root growth and contribution to soil carbon in a mixed mature Pinus koraiensis forest Plant and Soil, 400, 275-284. DOI:10.1007/s11104-015-2724-xURL [本文引用: 1]
WangJP, HeQQ, WuYH, ZhuH, SunHY, ZhouJ, WangDP, LiJJ, BingHJ (2021). Effects of pioneer N2-fixing plants on the resource status and establishment of neighboring non-N2-fixing plants in a newly formed glacier floodplain, eastern Tibetan Plateau Plant and Soil, 458, 261-276. DOI:10.1007/s11104-020-04462-yURL [本文引用: 1]
WangP, MouP, LiYB (2012). Review of root nutrient foraging plasticity and root competition of plants Chinese Journal of Plant Ecology, 36, 1184-1196. DOI:10.3724/SP.J.1258.2012.01184URL [本文引用: 1]
WangYL, GengQQ, HuangJH, WangCH, LiL, HasiM, NiuGX (2021). Effects of nitrogen addition and planting density on the growth and biological nitrogen fixation of Lespedeza davurica Chinese Journal of Plant Ecology, 45, 13-22. DOI:10.17521/cjpe.2020.0185URL [本文引用: 1]
WangX, PearseSJ, LambersH (2013). Cluster-root formation and carboxylate release in three Lupinus species as dependent on phosphorus supply, internal phosphorus concentration and relative growth rate Annals of Botany, 112, 1449-1459. DOI:10.1093/aob/mct210URL [本文引用: 1]
WardleDA (2004). Ecosystem properties and forest decline in contrasting long-term chronosequences Science, 305, 509-513. PMID:15205475 [本文引用: 1] During succession, ecosystem development occurs; but in the long-term absence of catastrophic disturbance, a decline phase eventually follows. We studied six long-term chronosequences, in Australia, Sweden, Alaska, Hawaii, and New Zealand; for each, the decline phase was associated with a reduction in tree basal area and an increase in the substrate nitrogen-to-phosphorus ratio, indicating increasing phosphorus limitation over time. These changes were often associated with reductions in litter decomposition rates, phosphorus release from litter, and biomass and activity of decomposer microbes. Our findings suggest that the maximal biomass phase reached during succession cannot be maintained in the long-term absence of major disturbance, and that similar patterns of decline occur in forested ecosystems spanning the tropical, temperate, and boreal zones.
WeiSP, JiaLM (2014). Research progress of mineral weathering by ectomycorrhizal fungi Chinese Journal of Ecology, 33, 3447-3454. [本文引用: 1]
WenZH, LiHB, ShenQ, TangXM, XiongCY, LiHG, PangJY, RyanMH, LambersH, ShenJB (2019). Tradeoffs among root morphology, exudation and mycorrhizal symbioses for phosphorus-acquisition strategies of 16 crop species New Phytologist, 223, 882-895. DOI:10.1111/nph.2019.223.issue-2URL [本文引用: 2]
YangY, JiangCH, HeYJ, OuJ, WangPP, SiJP, HeMH, LinY (2017). Effects of arbuscular mycorrhizal networks on the N and P contents and stoichiometry of three plants species from karst area Plant Physiology Journal, 53, 2078-2090. [本文引用: 1]
YaoQ, LiXL, FengG, ChristieP (2001). Mobilization of sparingly soluble inorganic phosphates by the external mycelium of an abuscular mycorrhizal fungus Plant and Soil, 230, 279-285. DOI:10.1023/A:1010367501363URL [本文引用: 1]
YuRP, LiXX, XiaoZH, LambersH, LiL (2020). Phosphorus facilitation and covariation of root traits in steppe species New Phytologist, 226, 1285-1298. DOI:10.1111/nph.v226.5URL [本文引用: 1]
ZemunikG, LambersH, TurnerBL, LalibertéE, OliveiraRS (2018). High abundance of non-mycorrhizal plant species in severely phosphorus-impoverished Brazilian campos rupestres Plant and Soil, 424, 255-271. DOI:10.1007/s11104-017-3503-7URL [本文引用: 1]
ZemunikG, TurnerBL, LambersH, LalibertéE (2015). Diversity of plant nutrient-acquisition strategies increases during long-term ecosystem development Nature Plants, 1, 15050. DOI: 10.1038/nplants.2015.50. URL [本文引用: 8]
ZhangW, YangL, WangZJ (2015). Advance and development trend of biological nitrogen fixation research Journal of Yunnan Agricultural University (Natural Science), 30, 810-821. [本文引用: 1]
ZhangYT, ZhuM, XianY, ShenH, ZhaoJ, GuoT (2012). Influence of mycorrhizal inoculation on competition between plant species and inorganic phosphate forms Acta Ecologica Sinica, 32, 7091-7101. DOI:10.5846/stxbURL [本文引用: 1]
ZhongHT, ZhouJ, AzmiA, ArrudaAJ, DooletteAL, SmernikRJ, LambersH (2021). Xylomelum occidentale (Proteaceae) accesses relatively mobile soil organic phosphorus without releasing carboxylates Journal of Ecology, 109, 246-259. DOI:10.1111/jec.v109.1URL [本文引用: 1]
ZhouJ, BingH, WuY, SunH, WangJ (2018). Weathering of primary mineral phosphate in the early stages of ecosystem development in the Hailuogou Glacier foreland chronosequence European Journal of Soil Science, 69, 450-461. DOI:10.1111/ejss.2018.69.issue-3URL [本文引用: 1]
ZhouJ, BingHJ, WuYH, YangZJ, WangJP, SunHY, LuoJ, LiangJH (2016). Rapid weathering processes of a 120-year-old chronosequence in the Hailuogou Glacier foreland, Mt. Gongga, SW China Geoderma, 267, 78-91. DOI:10.1016/j.geoderma.2015.12.024URL [本文引用: 1]
ZhouJ, WuYH, PrietzelJ, BingHJ, YuD, SunSQ, LuoJ, SunHY (2013). Changes of soil phosphorus speciation along a 120-year soil chronosequence in the Hailuogou Glacier retreat area (Gongga Mountain, SW China). Geoderma, 195-196, 251-259. DOI:10.1016/j.geoderma.2012.12.010URL [本文引用: 2]
ZhouJ, WuYH, TurnerBL, SunHY, WangJP, BingHJ, LuoJ, HeXL, ZhuH, HeQQ (2019). Transformation of soil organic phosphorus along the Hailuogou post-glacial chronosequence, southeastern edge of the Tibetan Plateau Geoderma, 352, 414-421. DOI:10.1016/j.geoderma.2019.05.038 [本文引用: 1] Organic phosphorus (P) accumulates in soil during pedogenesis, yet information on the composition and transformation of organic P during the early stages of soil development remains scarce. We studied the top 5 cm of mineral soil immediately beneath the organic horizon from six sites (0, 35, 45, 57, 85 and 125 years) along the Hailuogou glacier foreland chronosequence, on the southeastern edge of the Tibetan Plateau. Phosphorus compounds in the soils were determined by NaOH-EDTA extraction and solution phosphorus-31 nuclear magnetic resonance (P-31 NMR) spectroscopy. Extractable P was dominated by phosphomonoesters (up to 51.5%) and orthophosphate (37.9-44.6%) throughout the chronosequence. The phosphomonoesters were mainly hydrolysis products of RNA and phospholipids and followed a unimodal pattern with soil age, with maximum concentration at the 57-year-old site. myo-Inositol hexakisphosphate was not detected, although scyllo-inositol hexakisphosphate accounted for 4.7-9.3% of the extracted P and D-chiro- and neo-inositol hexakisphosphates occurred small amounts in a few soils (1.1-3.3% of the extracted P). DNA accounted for 4.0-8.3% of extracted P and increased continuously along the chronosequence, associated with increased inputs of plant and microbial residues and stronger sorption to soil surfaces during the rapid decline in soil pH. Pyrophosphate, an inorganic polyphosphate, occurred in small concentrations (up to 5.0%) that fluctuated with soil age. We conclude that organic P compounds accumulate rapidly in the top mineral soil during the early stages of pedogenesis, predominantly as relatively labile compounds from plant and microbial residues.
ZhouJ, Zúñiga-FeestA, LambersH (2020). In the beginning, there was only bare regolith-Then some plants arrived and changed the regolith Journal of Plant Ecology, 13, 511-516. DOI:10.1093/jpe/rtaa030URL [本文引用: 2]
Soil types select for plants with matching nutrient-acquisition and -use traits in hyperdiverse and severely nutrient-impoverished campos rupestres and cerrado in Central Brazil 1 2019
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Vellozioid roots allow for habitat specialization among rock- and soil-dwelling Velloziaceae in campos rupestres 3 2020
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. 10.17521/cjpe.2021.0160.F0002
Morphology of plant nutrient-acquisition strategies. A, Hyphae of the arbuscular mycorrhizal fungus Glomus caledonium growing into soil from a host root of Trifolium repens (Fabaceae). Photo by Iver Jakobsen; reprinted with permission of Springer-Verlag (Olsson et al., 2002). B, A seedling of Pinus sylvestris (Pinaceae) growing in a microcosm in association with the ectomycorrhizal fungus Suillus bovinus. The fungal mycelium can be seen spreading in the soil in the microcosm (arrowhead) and proliferating locally to form well-defined dense patches (arrows). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). C, Ericoid mycorrhizal root of Woollsia pungens, showing epidermal cells colonized by coils of an ericoid mycorrhizal fungus (stained blue, arrowed). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., <xref ref-type="bibr" rid="b49">2015</xref>). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., <xref ref-type="bibr" rid="b2">2020</xref>). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... ). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., 2020). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
Convergence of a specialized root trait in plants from nutrient-impoverished soils: phosphorus-acquisition strategy in a nonmycorrhizal cactus 1 2014
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
Interaction between tannins and fungal necromass stabilizes fungal residues in boreal forest soils 1 2019
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Leaf absorption of mineral nutrients in carnivorous plants stimulates root nutrient uptake 1 2002
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
Partitioning of soil phosphorus regulates competition between Vaccinium vitis-idaea and Deschampsia cespitosa 1 2013
... 多样的NASs还可影响植物生态位, 使其利用不同形态的养分, 促进植物共存.在澳大利亚西南部养分极其缺乏的成熟土壤中, 同一群落中的各种兰科植物发育不同的兰花菌根, 使不同种的兰花可吸收利用不同形态的N, 从而实现共存(Nurfadilah et al., 2013).P缺乏的欧洲草地中, 部分植物偏向于释放有机酸活化吸收无机P, 而另一部分植物偏向释放磷酸酶活化吸收有机P, 由此促进具有不同NASs的植物数量的增加和共存(Ceulemans et al., 2017).Ahmad-Ramli等(2013)发现Vaccinium通过释放磷酸单酯酶水解葡萄糖六磷酸盐获取P, 与其共存的Deschampsia则通过释放植酸酶水解植酸盐吸收P.英国古老石灰土上7种共存的植物, 与单独培育时相比, 混合培育时具有特殊根系的植物更多地活化和利用有机P, 具有菌根策略的禾本科植物以及具有固N策略的豆科植物更多地利用土壤中的Ca和P, 说明NASs可通过调节植物生态位导致物种的稳定共存(Phoenix et al., 2020). ...
Native soilborne pathogens equalize differences in competitive ability between plants of contrasting nutrient- acquisition strategies 1 2017
... 植物NASs与土壤病原体等的相互作用也可影响植物的共存和多样性.养分贫乏的成熟土壤上, 尽管具有特殊根系策略的植物可有效利用难溶态P, 提高竞争能力, 但因其根系缺乏外皮而容易受病原体的侵害, 因此竞争能力受到一定程度的抑制; 菌根植物捕获难溶态P的能力较弱, 但菌根真菌可帮助植物抵御病原体, 使具有不同NASs的植物的竞争能力达到一定程度的平衡, 从而对共存和多样性产生影响(Albornoz et al., 2017; Lambers et al., 2018; Tedersoo et al., 2020).澳大利亚西南部灌木林的土壤生物因素有利于ECM植物的生长, 但抑制了排根和固N策略植物的生长; 模型模拟发现, 这些抑制和促进效应的同时发生是此处成为全球生物多样性“热点区”的一个关键机制(Teste et al., 2017). ...
Shifts in symbiotic associations in plants capable of forming multiple root symbioses across a long-term soil chronosequence 2 2016
... 成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3).澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015).相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002).新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011).亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020).目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015).由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限.同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Effects of insect trapping on growth and nutrient content of Pinguicula vulgaris L. in relation to the nutrient content of the substrate 1 1983
... 食虫策略植物捕获养分通常包括3个阶段: 产生诱饵、捕获猎物和消化猎物体内的有机物(Król et al., 2012).盆栽实验表明, 植物能够消化吸收猎物体内的几丁质和蛋白质, 并且对蛋白质的利用效率更高(Pavlovič et al., 2016); 高山捕虫堇(Pinguicula alpina)能够捕获昆虫并利用其体内的N和P (Aldenius et al., 1983).目前已发现约有4 000种植物通过寄生策略获取养分, 其受土壤养分的影响相对较小(Press & Phoenix, 2005). ...
When plants eat rocks: functional adaptation of roots on rock outcrops 2 2019
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Arbuscular mycorrhizal fungi alter plant interspecific interaction under nitrogen fertilization 1 2019
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
The exploitative segregation of plant roots 1 2020
... 成土后期, 土壤中的有效养分(尤其是P)逐渐耗竭(Walker & Syers, 1976), 原生演替通常已经完成并形成顶极群落, 甚至可能由于养分的耗竭导致开始出现逆向演替(Wardle et al., 2004), 多样且互补的NASs使群落中的植物可获取不同空间中的养分, 有助于各种植物达到稳定共存的状态(图4).例如, 最近的研究发现, 相邻植物根系存在空间重叠时, 植物会减少远距离的养分吸收, 转而在邻近茎干的空间产生大量细根, 以增强植物对近距离资源的吸收能力, 使得同为细根策略的植物从不同土壤空间中吸收养分, 减少对养分的竞争以达到共存(Cabal et al., 2020).江西有效养分极低的砖红壤上生长的14种植物中, 浅根植物主要通过细根策略获取N、P和钾(K), 而深根植物则更多地依靠AM策略捕获养分, 是养分缺乏地区植物共存的关键机制(Liu et al., 2015). ...
Fine-root traits are linked to species dynamics in a successional plant community 1 2019
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Phosphorus resource partitioning shapes phosphorus acquisition and plant species abundance in grasslands 1 2017
... 多样的NASs还可影响植物生态位, 使其利用不同形态的养分, 促进植物共存.在澳大利亚西南部养分极其缺乏的成熟土壤中, 同一群落中的各种兰科植物发育不同的兰花菌根, 使不同种的兰花可吸收利用不同形态的N, 从而实现共存(Nurfadilah et al., 2013).P缺乏的欧洲草地中, 部分植物偏向于释放有机酸活化吸收无机P, 而另一部分植物偏向释放磷酸酶活化吸收有机P, 由此促进具有不同NASs的植物数量的增加和共存(Ceulemans et al., 2017).Ahmad-Ramli等(2013)发现Vaccinium通过释放磷酸单酯酶水解葡萄糖六磷酸盐获取P, 与其共存的Deschampsia则通过释放植酸酶水解植酸盐吸收P.英国古老石灰土上7种共存的植物, 与单独培育时相比, 混合培育时具有特殊根系的植物更多地活化和利用有机P, 具有菌根策略的禾本科植物以及具有固N策略的豆科植物更多地利用土壤中的Ca和P, 说明NASs可通过调节植物生态位导致物种的稳定共存(Phoenix et al., 2020). ...
Changing sources of nutrients during four million years of ecosystem development 1 1999
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
川中丘陵区4种乡土阔叶树细根性状对比研究 1 2020
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
川中丘陵区4种乡土阔叶树细根性状对比研究 1 2020
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
不同磷水平土壤接种丛枝菌根真菌对植物生长和养分吸收的影响 1 2009
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
不同磷水平土壤接种丛枝菌根真菌对植物生长和养分吸收的影响 1 2009
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
丛枝菌根真菌在土壤氮素循环中的作用 1 2014
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
丛枝菌根真菌在土壤氮素循环中的作用 1 2014
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Changes in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii 2 1995
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Costs and benefits of constructing roots of small diameter 1 1992
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
Linking root traits to nutrient foraging in arbuscular mycorrhizal trees in a temperate forest 1 2015
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
1 2010
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Foliar nutrient concentrations and resorption efficiency in plants of contrasting nutrient-acquisition strategies along a 2-million-year dune chronosequence 1 2014
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
Species- and community-level patterns in fine root traits along a 120000-year soil chronosequence in temperate rain forest 3 2011
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... 成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3).澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015).相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002).新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011).亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020).目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015).由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限.同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
胡萝卜状根的生物学特性研究综述 1 2008
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
胡萝卜状根的生物学特性研究综述 1 2008
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
植物与土壤微生物在调控生态系统养分循环中的作用 1 2010
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
植物与土壤微生物在调控生态系统养分循环中的作用 1 2010
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Root-derived inputs are major contributors to soil carbon in temperate forests, but vary by mycorrhizal type 1 2021
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Asymmetric response of root-associated fungal communities of an arbuscular mycorrhizal grass and an ectomycorrhizal tree to their coexistence in primary succession 2 2017
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Extraradical hyphae of the mycorrhizal fungus Glomus intraradices can hydrolyse organic phosphate 1 2000
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Quite a few reasons for calling carnivores “the most wonderful plants in the world” 1 2012
... 食虫策略植物捕获养分通常包括3个阶段: 产生诱饵、捕获猎物和消化猎物体内的有机物(Król et al., 2012).盆栽实验表明, 植物能够消化吸收猎物体内的几丁质和蛋白质, 并且对蛋白质的利用效率更高(Pavlovič et al., 2016); 高山捕虫堇(Pinguicula alpina)能够捕获昆虫并利用其体内的N和P (Aldenius et al., 1983).目前已发现约有4 000种植物通过寄生策略获取养分, 其受土壤养分的影响相对较小(Press & Phoenix, 2005). ...
How belowground interactions contribute to the coexistence of mycorrhizal and non- mycorrhizal species in severely phosphorus-impoverished hyperdiverse ecosystems 4 2018
Conceptual scheme showing effects of plant nutrient-acquisition strategies (NASs) on vegetation primary succession and coexistence and diversity of species. The change in intensity of color in soil nitrogen (N) and phosphorus (P) pool rectangle represents variations in total N and P pool with soil age, respectively. In the first stage of pedogenesis, plant NASs contribute to the establishment of pioneer community through facilitation. For example, N fixation and carboxylate-releasing strategies improve soil available nutrient pools and thus facilitate the settlement, growth and reproduction of subsequent species. In the early-intermediate stage, there are adequate soil available nutrients, NASs contribute to the competitiveness of plant species. Species with more effective NASs may have advantages in competition. And thus this will result in species turnover and promote succession. In the late stage, soil available nutrients are low or impoverished, especially for P, plant have to develop diverse NASs to acquire limited nutrients. The relationships between these NASs are mainly complementary. Diverse and complementary NASs have important impacts on the coexistence and diversity in the late stage of pedogenesis, although we still do not know the exact contribution to them. In addition, as proposed by Lambers et al. (<xref ref-type="bibr" rid="b31">2018</xref>), the different abilities against pathogens of plant species with different NASs also contribute the coexistence of plant species at this stage.Fig. 43.2 成土早-中期的NASs与物种更替成土早期第二阶段及成土中期, 随着土壤有效养分状况的改善, 群落密度和物种多样性有所增加, NASs影响不同植物对有效养分的竞争能力, 养分捕获能力强的植物在演替过程中逐渐占据优势, 促进物种更替和群落演替的进行(图4).火山爆发后进行的原生演替中, 后续植物Griselinia littoralis吸收先锋固N植物Coriaria arborea固定的N和活化的P, 地下和地上生物量都增加了3-6倍, 逐渐替代C. arborea (Walker et al., 2003).捷克一个矿山废弃地的原生演替中, 具有ECM的演替后续植物黄花柳(Salix caprea)比具有AM的先锋植物拂子茅(Calamagrostis epigejos)有更强的N吸收能力, 从而逐渐替代拂子茅成为优势种(Knoblochová et al., 2017).贡嘎山冰川退缩区土壤发育30多年后, 土壤有效P和N显著增加, 植被演替进入灌丛-阔叶林阶段, 冬瓜杨(Populus purdomii)和川滇柳开始竞争对N的吸收; 由于冬瓜杨的细根具有更强的N吸收能力(Song et al., 2020), 促进冬瓜杨取代川滇柳成为该演替序列下一阶段优势物种.土壤发育80多年后, 土壤中有机P的比例显著增加(Zhou et al., 2013, 2019), 具有ECM策略的峨眉冷杉(Abies fabri)可有效矿化并利用有机P, 而冬瓜杨的细根策略无法高效利用有机P, 因此, NASs对P捕获能力的差异可能是峨眉冷杉取代冬瓜杨成为这一时期优势物种的重要原因.不过, 成土早-中期NASs对物种更替的影响机制, 仍缺乏大量坚实的证据. ...
... Conceptual scheme showing effects of plant nutrient-acquisition strategies (NASs) on vegetation primary succession and coexistence and diversity of species. The change in intensity of color in soil nitrogen (N) and phosphorus (P) pool rectangle represents variations in total N and P pool with soil age, respectively. In the first stage of pedogenesis, plant NASs contribute to the establishment of pioneer community through facilitation. For example, N fixation and carboxylate-releasing strategies improve soil available nutrient pools and thus facilitate the settlement, growth and reproduction of subsequent species. In the early-intermediate stage, there are adequate soil available nutrients, NASs contribute to the competitiveness of plant species. Species with more effective NASs may have advantages in competition. And thus this will result in species turnover and promote succession. In the late stage, soil available nutrients are low or impoverished, especially for P, plant have to develop diverse NASs to acquire limited nutrients. The relationships between these NASs are mainly complementary. Diverse and complementary NASs have important impacts on the coexistence and diversity in the late stage of pedogenesis, although we still do not know the exact contribution to them. In addition, as proposed by Lambers et al. (2018), the different abilities against pathogens of plant species with different NASs also contribute the coexistence of plant species at this stage.Fig. 43.2 成土早-中期的NASs与物种更替成土早期第二阶段及成土中期, 随着土壤有效养分状况的改善, 群落密度和物种多样性有所增加, NASs影响不同植物对有效养分的竞争能力, 养分捕获能力强的植物在演替过程中逐渐占据优势, 促进物种更替和群落演替的进行(图4).火山爆发后进行的原生演替中, 后续植物Griselinia littoralis吸收先锋固N植物Coriaria arborea固定的N和活化的P, 地下和地上生物量都增加了3-6倍, 逐渐替代C. arborea (Walker et al., 2003).捷克一个矿山废弃地的原生演替中, 具有ECM的演替后续植物黄花柳(Salix caprea)比具有AM的先锋植物拂子茅(Calamagrostis epigejos)有更强的N吸收能力, 从而逐渐替代拂子茅成为优势种(Knoblochová et al., 2017).贡嘎山冰川退缩区土壤发育30多年后, 土壤有效P和N显著增加, 植被演替进入灌丛-阔叶林阶段, 冬瓜杨(Populus purdomii)和川滇柳开始竞争对N的吸收; 由于冬瓜杨的细根具有更强的N吸收能力(Song et al., 2020), 促进冬瓜杨取代川滇柳成为该演替序列下一阶段优势物种.土壤发育80多年后, 土壤中有机P的比例显著增加(Zhou et al., 2013, 2019), 具有ECM策略的峨眉冷杉(Abies fabri)可有效矿化并利用有机P, 而冬瓜杨的细根策略无法高效利用有机P, 因此, NASs对P捕获能力的差异可能是峨眉冷杉取代冬瓜杨成为这一时期优势物种的重要原因.不过, 成土早-中期NASs对物种更替的影响机制, 仍缺乏大量坚实的证据. ...
... 植物NASs与土壤病原体等的相互作用也可影响植物的共存和多样性.养分贫乏的成熟土壤上, 尽管具有特殊根系策略的植物可有效利用难溶态P, 提高竞争能力, 但因其根系缺乏外皮而容易受病原体的侵害, 因此竞争能力受到一定程度的抑制; 菌根植物捕获难溶态P的能力较弱, 但菌根真菌可帮助植物抵御病原体, 使具有不同NASs的植物的竞争能力达到一定程度的平衡, 从而对共存和多样性产生影响(Albornoz et al., 2017; Lambers et al., 2018; Tedersoo et al., 2020).澳大利亚西南部灌木林的土壤生物因素有利于ECM植物的生长, 但抑制了排根和固N策略植物的生长; 模型模拟发现, 这些抑制和促进效应的同时发生是此处成为全球生物多样性“热点区”的一个关键机制(Teste et al., 2017). ...
Plant mineral nutrition in ancient landscapes: high plant species diversity on infertile soils is linked to functional diversity for nutritional strategies 3 2011
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Plant nutrient-acquisition strategies change with soil age. 10 2008
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
Morphology of plant nutrient-acquisition strategies. A, Hyphae of the arbuscular mycorrhizal fungus Glomus caledonium growing into soil from a host root of Trifolium repens (Fabaceae). Photo by Iver Jakobsen; reprinted with permission of Springer-Verlag (Olsson et al., 2002). B, A seedling of Pinus sylvestris (Pinaceae) growing in a microcosm in association with the ectomycorrhizal fungus Suillus bovinus. The fungal mycelium can be seen spreading in the soil in the microcosm (arrowhead) and proliferating locally to form well-defined dense patches (arrows). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). C, Ericoid mycorrhizal root of Woollsia pungens, showing epidermal cells colonized by coils of an ericoid mycorrhizal fungus (stained blue, arrowed). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., <xref ref-type="bibr" rid="b49">2015</xref>). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., <xref ref-type="bibr" rid="b2">2020</xref>). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... Morphology of plant nutrient-acquisition strategies. A, Hyphae of the arbuscular mycorrhizal fungus Glomus caledonium growing into soil from a host root of Trifolium repens (Fabaceae). Photo by Iver Jakobsen; reprinted with permission of Springer-Verlag (Olsson et al., 2002). B, A seedling of Pinus sylvestris (Pinaceae) growing in a microcosm in association with the ectomycorrhizal fungus Suillus bovinus. The fungal mycelium can be seen spreading in the soil in the microcosm (arrowhead) and proliferating locally to form well-defined dense patches (arrows). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). C, Ericoid mycorrhizal root of Woollsia pungens, showing epidermal cells colonized by coils of an ericoid mycorrhizal fungus (stained blue, arrowed). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., 2015). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., 2020). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... ). C, Ericoid mycorrhizal root of Woollsia pungens, showing epidermal cells colonized by coils of an ericoid mycorrhizal fungus (stained blue, arrowed). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., 2015). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., 2020). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... ). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., 2020). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
... ., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Root structure and functioning for efficient acquisition of phosphorus: matching morphological and physiological traits 1 2006
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
Diverse belowground resource strategies underlie plant species coexistence and spatial distribution in three grasslands along a precipitation gradient 1 2017
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species 3 2015
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
... 成土后期, 土壤中的有效养分(尤其是P)逐渐耗竭(Walker & Syers, 1976), 原生演替通常已经完成并形成顶极群落, 甚至可能由于养分的耗竭导致开始出现逆向演替(Wardle et al., 2004), 多样且互补的NASs使群落中的植物可获取不同空间中的养分, 有助于各种植物达到稳定共存的状态(图4).例如, 最近的研究发现, 相邻植物根系存在空间重叠时, 植物会减少远距离的养分吸收, 转而在邻近茎干的空间产生大量细根, 以增强植物对近距离资源的吸收能力, 使得同为细根策略的植物从不同土壤空间中吸收养分, 减少对养分的竞争以达到共存(Cabal et al., 2020).江西有效养分极低的砖红壤上生长的14种植物中, 浅根植物主要通过细根策略获取N、P和钾(K), 而深根植物则更多地依靠AM策略捕获养分, 是养分缺乏地区植物共存的关键机制(Liu et al., 2015). ...
Major crop species show differential balance between root morphological and physiological responses to variable phosphorus supply 2 2016
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
Evolutionary history resolves global organization of root functional traits 1 2018
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
Plant symbiotic microorganisms in acid sulfate soil: significance in the growth of pioneer plants 1 2008
Formation of dauciform roots by Japanese native Cyperaceae and their contribution to phosphorus dynamics in soils 2 2021
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Opposing effects of competitive exclusion on the phylogenetic structure of communities 1 2010
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
菌根真菌与吸收根功能性状的关系: 研究进展与评述 1 2013
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
菌根真菌与吸收根功能性状的关系: 研究进展与评述 1 2013
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
Does cluster-root activity benefit nutrient uptake and growth of co-existing species 1 2014
... 互补性的植物NASs可为邻近植物提供其自身无法活化的有效养分, 从而促进群落中植物的稳定共存.例如, 澳大利亚西南部的Banksia menziesii发育排根活化大量的P和Ca, 而与其相邻的无法直接活化P的菌根植物则通过菌丝获取这些由B. menziesii活化的养分, 成为该地区植物共存的一个重要机制((Muler et al., 2014; Teste et al., 2014).Teste等(2020)进一步研究发现B. menziesii活化的P可达土壤有效P库的80%, 是相邻植物获取P的主要来源.内蒙古草原上, 植物根系分泌的羧化物既可活化难溶的无机磷, 后被邻近的植物通过细根吸收, 也可将土壤矿物固定的有机P释放到土壤溶液中, 再被邻近的菌根植物通过磷酸酶矿化后吸收, 展现了互补性的NASs对植物共存的影响(Yu et al., 2020). ...
Plants colonizing volcanic deposits: root adaptations and effects on rhizosphere microorganisms 1 2021
Interactions among nitrogen fixation and soil phosphorus acquisition strategies in lowland tropical rain forests 1 2014
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Nitrogen uptake by trees and mycorrhizal fungi in a successional northern temperate forest: insights from multiple isotopic methods 1 2013
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
The expanding role of deep roots during long-term terrestrial ecosystem development 1 2020
... 成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3).澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015).相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002).新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011).亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020).目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015).由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限.同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3). ...
Variation in nutrient-acquisition patterns by mycorrhizal fungi of rare and common orchids explains diversification in a global biodiversity hotspot 3 2013
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
... 多样的NASs还可影响植物生态位, 使其利用不同形态的养分, 促进植物共存.在澳大利亚西南部养分极其缺乏的成熟土壤中, 同一群落中的各种兰科植物发育不同的兰花菌根, 使不同种的兰花可吸收利用不同形态的N, 从而实现共存(Nurfadilah et al., 2013).P缺乏的欧洲草地中, 部分植物偏向于释放有机酸活化吸收无机P, 而另一部分植物偏向释放磷酸酶活化吸收有机P, 由此促进具有不同NASs的植物数量的增加和共存(Ceulemans et al., 2017).Ahmad-Ramli等(2013)发现Vaccinium通过释放磷酸单酯酶水解葡萄糖六磷酸盐获取P, 与其共存的Deschampsia则通过释放植酸酶水解植酸盐吸收P.英国古老石灰土上7种共存的植物, 与单独培育时相比, 混合培育时具有特殊根系的植物更多地活化和利用有机P, 具有菌根策略的禾本科植物以及具有固N策略的豆科植物更多地利用土壤中的Ca和P, 说明NASs可通过调节植物生态位导致物种的稳定共存(Phoenix et al., 2020). ...
Mineral nutrition of Campos rupestres plant species on contrasting nutrient-impoverished soil types 4 2015
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. 10.17521/cjpe.2021.0160.F0002
Morphology of plant nutrient-acquisition strategies. A, Hyphae of the arbuscular mycorrhizal fungus Glomus caledonium growing into soil from a host root of Trifolium repens (Fabaceae). Photo by Iver Jakobsen; reprinted with permission of Springer-Verlag (Olsson et al., 2002). B, A seedling of Pinus sylvestris (Pinaceae) growing in a microcosm in association with the ectomycorrhizal fungus Suillus bovinus. The fungal mycelium can be seen spreading in the soil in the microcosm (arrowhead) and proliferating locally to form well-defined dense patches (arrows). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). C, Ericoid mycorrhizal root of Woollsia pungens, showing epidermal cells colonized by coils of an ericoid mycorrhizal fungus (stained blue, arrowed). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., <xref ref-type="bibr" rid="b49">2015</xref>). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., <xref ref-type="bibr" rid="b33">2008</xref>). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., <xref ref-type="bibr" rid="b2">2020</xref>). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... ). D, Nodules of Astragalus mahoshanicus (Fabaceae). E, Cluster roots of Banksia grandis (Proteaceae). Photo by ZHONG Hong-Tao. F, Sand-binding roots of Actinocephalus cabralensis (Ericaulaceae)(Oliveira et al., 2015). G, Dauciform roots of Tetraria species (Cyperaceae). Reprinted with permission of Elsevier B.V. (Lambers et al., 2008). H, Vellozioid roots of Barbacenia tomentosa (Velloziaceae) collected in field (Abrahão et al., 2020). EMF, ericoid mycorrhizal fungus; RE, root epidermis; RVS, root vascular system.Fig. 2 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
Effects of nitrogen and phosphorus availability on fine-root dynamics in Hawaiian montane forests 1 2001
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Responses of fine root functional traits to soil nutrient limitations in a karst ecosystem of southwest China 1 2018
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
A carnivorous sundew plant prefers protein over chitin as a source of nitrogen from its traps 1 2016
... 食虫策略植物捕获养分通常包括3个阶段: 产生诱饵、捕获猎物和消化猎物体内的有机物(Król et al., 2012).盆栽实验表明, 植物能够消化吸收猎物体内的几丁质和蛋白质, 并且对蛋白质的利用效率更高(Pavlovič et al., 2016); 高山捕虫堇(Pinguicula alpina)能够捕获昆虫并利用其体内的N和P (Aldenius et al., 1983).目前已发现约有4 000种植物通过寄生策略获取养分, 其受土壤养分的影响相对较小(Press & Phoenix, 2005). ...
Soil phosphorus fractions and symbiotic nitrogen fixation across a substrate-age gradient in Hawaii 5 2002
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... 植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... 成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3).澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015).相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002).新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011).亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020).目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015).由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限.同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Nitrogen-fixing red alder trees tap rock-derived nutrients 1 2019
Niche differentiation and plasticity in soil phosphorus acquisition among co-occurring plants 2 2020
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
... 多样的NASs还可影响植物生态位, 使其利用不同形态的养分, 促进植物共存.在澳大利亚西南部养分极其缺乏的成熟土壤中, 同一群落中的各种兰科植物发育不同的兰花菌根, 使不同种的兰花可吸收利用不同形态的N, 从而实现共存(Nurfadilah et al., 2013).P缺乏的欧洲草地中, 部分植物偏向于释放有机酸活化吸收无机P, 而另一部分植物偏向释放磷酸酶活化吸收有机P, 由此促进具有不同NASs的植物数量的增加和共存(Ceulemans et al., 2017).Ahmad-Ramli等(2013)发现Vaccinium通过释放磷酸单酯酶水解葡萄糖六磷酸盐获取P, 与其共存的Deschampsia则通过释放植酸酶水解植酸盐吸收P.英国古老石灰土上7种共存的植物, 与单独培育时相比, 混合培育时具有特殊根系的植物更多地活化和利用有机P, 具有菌根策略的禾本科植物以及具有固N策略的豆科植物更多地利用土壤中的Ca和P, 说明NASs可通过调节植物生态位导致物种的稳定共存(Phoenix et al., 2020). ...
Functional significance of dauciform roots: exudation of carboxylates and acid phosphatase under phosphorus deficiency in Caustis blakei (Cyperaceae 2 2006
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Impacts of parasitic plants on natural communities 2 2005
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
... 食虫策略植物捕获养分通常包括3个阶段: 产生诱饵、捕获猎物和消化猎物体内的有机物(Król et al., 2012).盆栽实验表明, 植物能够消化吸收猎物体内的几丁质和蛋白质, 并且对蛋白质的利用效率更高(Pavlovič et al., 2016); 高山捕虫堇(Pinguicula alpina)能够捕获昆虫并利用其体内的N和P (Aldenius et al., 1983).目前已发现约有4 000种植物通过寄生策略获取养分, 其受土壤养分的影响相对较小(Press & Phoenix, 2005). ...
丛枝菌根真菌对矿质养分活化作用研究进展 1 2019
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
丛枝菌根真菌对矿质养分活化作用研究进展 1 2019
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Strong linkage between plant and soil fungal communities along a successional coastal dune system 2 2016
... 成土后期植物NASs的多样性显著升高.澳大利亚西南部Jurien Bay序列后期, 土壤总P含量降至6-10 mg·kg-1, 用稀有物种评估的植物NASs多样性增加了一倍, 与NASs相关的性状分离度增加了46% (Zemunik et al., 2015).加拿大养分贫瘠的沿海沙丘序列, 随着土壤年龄增加, 植物NASs的Simpson指数增加超过1.5倍(Roy-Bolduc et al., 2016).成土过程中总P的减少是导致植物NASs多样性增加的原因之一.澳大利亚西南部Jurien Bay土壤序列的总P含量与植物NASs多样性呈负相关关系, 总P的减少使植物产生多种NASs以获取土壤中的P (Zemunik et al., 2015).在澳大利亚西南部总P为50 mg·kg-1左右的土壤中, Xylomelum occidentale发育了特别的NASs, 既不释放大量有机酸也不形成菌根, 而是释放磷酸酶水解土壤中的RNA和磷脂(Zhong et al., 2021), 增加了成土后期NASs的丰富度. ...
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Carbon trading for phosphorus gain: the balance between rhizosphere carboxylates and arbuscular mycorrhizal symbiosis in plant phosphorus acquisition 1 2012
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Global patterns in fine root decomposition: climate, chemistry, mycorrhizal association and woodiness 1 2019
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Different sets of belowground traits predict the ability of plant species to suppress and tolerate their competitors 1 2018
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Cluster roots: a curiosity in context 1 2005
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
Mycorrhizal associations and phosphorus acquisition: from cells to ecosystems 2 2015
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Functional diversity in arbuscular mycorrhizal (AM) symbioses: the contribution of the mycorrhizal P uptake pathway is not correlated with mycorrhizal responses in growth or total P uptake 1 2004
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
菌根真菌磷酸酶活性对红三叶草生境中土壤有机磷亏缺的影响 1 2001
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
菌根真菌磷酸酶活性对红三叶草生境中土壤有机磷亏缺的影响 1 2001
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Stoichiometric flexibility and soil bacterial communities respond to nitrogen fertilization and neighbor competition at the early stage of primary succession 1 2020
How mycorrhizal associations drive plant population and community biology 2 2020
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
... 植物NASs与土壤病原体等的相互作用也可影响植物的共存和多样性.养分贫乏的成熟土壤上, 尽管具有特殊根系策略的植物可有效利用难溶态P, 提高竞争能力, 但因其根系缺乏外皮而容易受病原体的侵害, 因此竞争能力受到一定程度的抑制; 菌根植物捕获难溶态P的能力较弱, 但菌根真菌可帮助植物抵御病原体, 使具有不同NASs的植物的竞争能力达到一定程度的平衡, 从而对共存和多样性产生影响(Albornoz et al., 2017; Lambers et al., 2018; Tedersoo et al., 2020).澳大利亚西南部灌木林的土壤生物因素有利于ECM植物的生长, 但抑制了排根和固N策略植物的生长; 模型模拟发现, 这些抑制和促进效应的同时发生是此处成为全球生物多样性“热点区”的一个关键机制(Teste et al., 2017). ...
Specialized roots of Velloziaceae weather quartzite rock while mobilizing phosphorus using carboxylates 4 2019
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
Plant-soil feedback and the maintenance of diversity in Mediterranean-climate shrublands 2 2017
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
... 植物NASs与土壤病原体等的相互作用也可影响植物的共存和多样性.养分贫乏的成熟土壤上, 尽管具有特殊根系策略的植物可有效利用难溶态P, 提高竞争能力, 但因其根系缺乏外皮而容易受病原体的侵害, 因此竞争能力受到一定程度的抑制; 菌根植物捕获难溶态P的能力较弱, 但菌根真菌可帮助植物抵御病原体, 使具有不同NASs的植物的竞争能力达到一定程度的平衡, 从而对共存和多样性产生影响(Albornoz et al., 2017; Lambers et al., 2018; Tedersoo et al., 2020).澳大利亚西南部灌木林的土壤生物因素有利于ECM植物的生长, 但抑制了排根和固N策略植物的生长; 模型模拟发现, 这些抑制和促进效应的同时发生是此处成为全球生物多样性“热点区”的一个关键机制(Teste et al., 2017). ...
Mycorrhizal fungal biomass and scavenging declines in phosphorus-impoverished soils during ecosystem retrogression 2 2016
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
... ., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Complementary plant nutrient-acquisition strategies promote growth of neighbour species 2 2014
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
... 互补性的植物NASs可为邻近植物提供其自身无法活化的有效养分, 从而促进群落中植物的稳定共存.例如, 澳大利亚西南部的Banksia menziesii发育排根活化大量的P和Ca, 而与其相邻的无法直接活化P的菌根植物则通过菌丝获取这些由B. menziesii活化的养分, 成为该地区植物共存的一个重要机制((Muler et al., 2014; Teste et al., 2014).Teste等(2020)进一步研究发现B. menziesii活化的P可达土壤有效P库的80%, 是相邻植物获取P的主要来源.内蒙古草原上, 植物根系分泌的羧化物既可活化难溶的无机磷, 后被邻近的植物通过细根吸收, 也可将土壤矿物固定的有机P释放到土壤溶液中, 再被邻近的菌根植物通过磷酸酶矿化后吸收, 展现了互补性的NASs对植物共存的影响(Yu et al., 2020). ...
The potential for phosphorus benefits through root placement in the rhizosphere of phosphorus-mobilising neighbours 1 2020
... 互补性的植物NASs可为邻近植物提供其自身无法活化的有效养分, 从而促进群落中植物的稳定共存.例如, 澳大利亚西南部的Banksia menziesii发育排根活化大量的P和Ca, 而与其相邻的无法直接活化P的菌根植物则通过菌丝获取这些由B. menziesii活化的养分, 成为该地区植物共存的一个重要机制((Muler et al., 2014; Teste et al., 2014).Teste等(2020)进一步研究发现B. menziesii活化的P可达土壤有效P库的80%, 是相邻植物获取P的主要来源.内蒙古草原上, 植物根系分泌的羧化物既可活化难溶的无机磷, 后被邻近的植物通过细根吸收, 也可将土壤矿物固定的有机P释放到土壤溶液中, 再被邻近的菌根植物通过磷酸酶矿化后吸收, 展现了互补性的NASs对植物共存的影响(Yu et al., 2020). ...
缺磷白羽扇豆排根与非排根区根尖分泌有机酸的比较 1 2000
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
缺磷白羽扇豆排根与非排根区根尖分泌有机酸的比较 1 2000
... 特殊根系策略包括形成排根(CR)、固沙根(SR)、胡萝卜状根(DR)和毛状根(VR)等(图2E-2H).虽然这些特殊根系的形态和结构各不相同, 但都可在较短时间内(通常为几天)释放比菌根多几个数量级的羧化物等物质来活化土壤中被吸附固定的养分(主要是P), 增加土壤有效P的含量(田中民等, 2000; Shane & Lambers, 2005; Lambers et al., 2006; 吉文丽等, 2008).土壤有效P极低的环境中, 山龙眼科植物利用排根策略活化难利用态P已被广泛报道(Lambers et al., 2008, 2011).缺P的巴西中部地区, 约44.4%的Eriocaulaceae植物发育固沙根活化土壤P (Oliveira et al., 2015), 而生长于岩石上的Velloziaceae植物则发育毛状根, 促进岩石释放无机养分(Teodoro et al., 2019; Abrahão et al., 2020).有机质较高但有效P极低的浅层土壤中, Caustis blakei可发育胡萝卜状根, 释放大量的有机酸和磷酸酶矿化有机磷(Playsted et al., 2006). ...
Pedogenesis, nutrient dynamics, and ecosystem development: the legacy of T. W. Walker and J. K. Syers 1 2013
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
Foliar 15N natural abundance in Hawaiian rainforest: patterns and possible mechanisms 1 1989
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
Root morphology and its contribution to a large root system for phosphorus uptake by Rytidosperma species (wallaby grass 1 2017
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
Colonization dynamics and facilitative impacts of a nitrogen- fixing shrub in primary succession 4 2003
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... ., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... 成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3).澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015).相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002).新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011).亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020).目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015).由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限.同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
... 成土后期, 土壤中的有效养分(尤其是P)逐渐耗竭(Walker & Syers, 1976), 原生演替通常已经完成并形成顶极群落, 甚至可能由于养分的耗竭导致开始出现逆向演替(Wardle et al., 2004), 多样且互补的NASs使群落中的植物可获取不同空间中的养分, 有助于各种植物达到稳定共存的状态(图4).例如, 最近的研究发现, 相邻植物根系存在空间重叠时, 植物会减少远距离的养分吸收, 转而在邻近茎干的空间产生大量细根, 以增强植物对近距离资源的吸收能力, 使得同为细根策略的植物从不同土壤空间中吸收养分, 减少对养分的竞争以达到共存(Cabal et al., 2020).江西有效养分极低的砖红壤上生长的14种植物中, 浅根植物主要通过细根策略获取N、P和钾(K), 而深根植物则更多地依靠AM策略捕获养分, 是养分缺乏地区植物共存的关键机制(Liu et al., 2015). ...
Fine root growth and contribution to soil carbon in a mixed mature Pinus koraiensis forest 1 2016
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Effects of pioneer N2-fixing plants on the resource status and establishment of neighboring non-N2-fixing plants in a newly formed glacier floodplain, eastern Tibetan Plateau 1 2021
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
土壤养分空间异质性与植物根系的觅食反应 1 2004
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
氮肥和种植密度对达乌里胡枝子的生长与生物固氮的影响 1 2021
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
氮肥和种植密度对达乌里胡枝子的生长与生物固氮的影响 1 2021
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Cluster-root formation and carboxylate release in three Lupinus species as dependent on phosphorus supply, internal phosphorus concentration and relative growth rate 1 2013
Ecosystem properties and forest decline in contrasting long-term chronosequences 1 2004
... 成土后期, 土壤中的有效养分(尤其是P)逐渐耗竭(Walker & Syers, 1976), 原生演替通常已经完成并形成顶极群落, 甚至可能由于养分的耗竭导致开始出现逆向演替(Wardle et al., 2004), 多样且互补的NASs使群落中的植物可获取不同空间中的养分, 有助于各种植物达到稳定共存的状态(图4).例如, 最近的研究发现, 相邻植物根系存在空间重叠时, 植物会减少远距离的养分吸收, 转而在邻近茎干的空间产生大量细根, 以增强植物对近距离资源的吸收能力, 使得同为细根策略的植物从不同土壤空间中吸收养分, 减少对养分的竞争以达到共存(Cabal et al., 2020).江西有效养分极低的砖红壤上生长的14种植物中, 浅根植物主要通过细根策略获取N、P和钾(K), 而深根植物则更多地依靠AM策略捕获养分, 是养分缺乏地区植物共存的关键机制(Liu et al., 2015). ...
外生菌根真菌对矿物风化作用研究进展 1 2014
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
外生菌根真菌对矿物风化作用研究进展 1 2014
... 植物是影响成土过程的重要因子.首先, 植物为获取生长发育所需的矿质养分, 通过各种NASs客观上导致母岩的风化、促进成土过程的进行(魏松坡和贾黎明, 2014; Augusto et al., 2019; Zhou et al., 2020).然而, 目前的研究多关注成土早期有效养分不足如何导致植物NASs发生变化, 少有研究定量解析植物NASs对成土过程的贡献.其次, 植物NASs对成土过程中有机C的积累具有重要贡献(Keller et al., 2021).在成土和演替早期, 土壤有效养分含量较低, 先锋植物会大量增加地下生物量、尤其是细根的生产力以获取足够的养分(George et al., 2010); 由于细根的生物量和周转率大于叶片, 因此, 植物根系成为土壤有机C的一个重要来源.如东北红松林的原位研究发现, 细根对土壤C的贡献是叶片凋落物贡献量的1.2倍(Wang et al., 2016).此外, 被菌根真菌侵染的植物根系比非菌根更难分解(See et al., 2019), 尤其是与根际有机酸发生络合反应后更不易被分解(Adamczyk et al., 2019), 从而更易形成可长期存在的有机C库.不过, 目前仍不清楚各成土阶段中不同植物NASs对矿物风化和有机C积累的贡献, 也不清楚这些贡献随成土年龄的变化, 值得未来开展更多的研究工作.这些研究成果不仅有助于阐明成土作用的生物学机制, 而且有益于揭示土壤发育与植物群落演替之间的协同关系, 具有重要的生态学意义. ...
Tradeoffs among root morphology, exudation and mycorrhizal symbioses for phosphorus-acquisition strategies of 16 crop species 2 2019
... 植物养分捕获策略(nutrient-acquisition strategies, NASs)是指植物为克服土壤有效养分的不足, 通过改变根系形态、分泌特定化学物质或利用微生物的作用等增强自身养分获取能力的行为(Lambers et al., 2008).根据捕获养分方式的不同, 植物NASs可分为微生物策略(microbial strategy)、特殊根系策略(specialized root strategy)、细根策略(fine root strategy)、食虫策略(carnivorous strategy)和寄生策略(parasitic strategy)等(图1).微生物策略主要是指植物通过非共生微生物或形成根系-微生物互惠共生体(如菌根或根瘤)等方式增强养分获取能力的行为.特殊根系策略主要是指植物形成与普通根系不同且可释放大量羧化物(carboxylates)的形态特殊的根系(如排根cluster roots), 以活化土壤中难以被植物直接利用的养分(Abrahão et al., 2014; Hayes et al., 2014; Oliveira et al., 2015).细根策略主要是指植物通过产生更多细根、增加比根长和分支比等, 增加根与土壤接触的面积, 从而增加根系直接吸收养分的比表面积, 使植物根系在相同的碳(C)支出下, 更有效地探索和吸收土壤中的养分(Liu et al., 2015; Ma et al., 2018; Wen et al., 2019).植物除了通过根系捕获养分以外, 还能通过制造陷阱捕猎小型虫类, 消化吸收猎物体内的养分, 这种策略被称为食虫策略(Adamec, 2002).寄生策略则是指植物通过寄生或半寄生的方式从宿主植物根系或地上部分的维管组织中获取生长所需养分的行为(Press & Phoenix, 2005). ...
... 细根策略的主要特征是植物通过生长更多的侧根、形成分支比和比根长更大的细根以获得更强的养分捕获能力(Eissenstat, 1992; 王庆成和程云环, 2004; 苗原等, 2013; Waddell et al., 2017; 陈俊华等, 2020).细根策略常与菌根策略同时发生, 但随着植物吸收根根径减小, 植物主要通过细根直接吸收养分, 对菌根吸收养分的依赖度降低(Eissenstat et al., 2015).草地生态系统中, 吸收根更细、分支更密集的植物不易被菌根侵染, 植物分配给根系的C主要用于细根的发育, 并主要通过细根直接吸收养分(Li et al., 2017; Wen et al., 2019).农作物也存在细根策略.与豆科植物相比, 玉米(Zea mays)、小麦(Triticum aestivum)和油菜(Brassica napus)的吸收根更细、比根长相对更大, 以通过细根吸收更多养分(Lyu et al., 2016). ...
丛枝菌根网对喀斯特适生植物氮、磷化学计量特征的影响 1 2017
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
丛枝菌根网对喀斯特适生植物氮、磷化学计量特征的影响 1 2017
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
Mobilization of sparingly soluble inorganic phosphates by the external mycelium of an abuscular mycorrhizal fungus 1 2001
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
Phosphorus facilitation and covariation of root traits in steppe species 1 2020
... 互补性的植物NASs可为邻近植物提供其自身无法活化的有效养分, 从而促进群落中植物的稳定共存.例如, 澳大利亚西南部的Banksia menziesii发育排根活化大量的P和Ca, 而与其相邻的无法直接活化P的菌根植物则通过菌丝获取这些由B. menziesii活化的养分, 成为该地区植物共存的一个重要机制((Muler et al., 2014; Teste et al., 2014).Teste等(2020)进一步研究发现B. menziesii活化的P可达土壤有效P库的80%, 是相邻植物获取P的主要来源.内蒙古草原上, 植物根系分泌的羧化物既可活化难溶的无机磷, 后被邻近的植物通过细根吸收, 也可将土壤矿物固定的有机P释放到土壤溶液中, 再被邻近的菌根植物通过磷酸酶矿化后吸收, 展现了互补性的NASs对植物共存的影响(Yu et al., 2020). ...
High abundance of non-mycorrhizal plant species in severely phosphorus-impoverished Brazilian campos rupestres 1 2018
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
Diversity of plant nutrient-acquisition strategies increases during long-term ecosystem development 8 2015
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
... 成土中期, 土壤N、P总量处于中等水平, 养分主要以有机态或次生矿物结合态存在, 有效养分相对较多(Walker & Syers, 1976), 固N、菌根和细根策略在养分捕获上的作用有所减弱; 可有效活化P的特殊根系策略降至最低(图3).澳大利亚西南部的Jurien Bay序列中期与早期相比, Acacia rostellifera的根瘤生物量占总生物量中的比值降低了约40% (Albornoz et al., 2016), AM和ECM植物的盖度均显著降低(Zemunik et al., 2015).相对于早期, 夏威夷土壤序列中期固N树种的根际固N速率最多减少到1/8 (Pearson & Vitousek, 2002).新西兰的Franz Josef序列中期, 细根吸收的N和P相对早期减少了50% (Holdaway et al., 2011).亚利桑那州北部的土壤序列中期, 有机层土壤的细根数量相对早期减少了近一半(Newman et al., 2020).目前仅有研究报道澳大利亚西南部的Jurien Bay序列中期发现植物发育了排根策略(Zemunik et al., 2015).由于成土中期养分较为充足, 因此, 针对这一时期NASs的研究成果较为有限.同时, 我们推测, 这一时期植物获取养分相对较为容易, 从植物养分获取权衡的角度考虑, 这一时期的植物倾向于将更少的C投入NASs的发育(图3). ...
... 长时间的淋溶作用使成土后期土壤的P库接近耗竭, 有限的P主要以有机态或闭蓄态存在, N的形态以有机态为主, 植物生长通常受到P或N、P的共同限制(Walker & Syers, 1976; Chadwick et al., 1999; Lambers et al., 2008; Teste et al., 2016).由于土壤中的闭蓄态P难以被菌根和细根直接吸收(Ryan et al., 2012; Pan et al., 2018), 需要被释放到土壤溶液中才能被磷酸酶高效水解, 而菌根释放的羧化物和磷酸酶相对较少, 只能活化数量有限的有机P和闭蓄态P (Nurfadilah et al., 2013; Smith et al., 2015; Albornoz et al., 2016), 因此, 成土后期微生物和细根策略对植物捕获P的贡献相对较弱(图3).由于植物的特殊根系可释放大量有机酸活化闭蓄态P, 因此, 这一时期特殊根系及磷酸酶成为植物获取P的主要策略(Playsted et al., 2006; Lambers et al., 2008; Masuda et al., 2021; 图3).例如, 相对前中期, 澳大利亚西南部的Jurien Bay土壤序列后期AM植物的相对盖度与早期相比减少了42%以上(Teste et al., 2016), 而特殊根系植物的相对盖度增大, 特殊根系策略成为植物获取土壤有效P最主要的途径(Zemunik et al., 2015).在土壤养分贫瘠的巴西Minas Gerais地区, AM植物根系被侵染部分占总根系长度的比例从成土早期的71%下降到后期的小于1%, 特殊根系植物的相对盖度与有效P呈负线性相关关系(Zemunik et al., 2018; Abrahão et al., 2019).在N、P共同限制的夏威夷土壤序列后期, 固N树种的固N速率远低于成土早期(Pearson & Vitousek, 2002); 尽管植物细根的质量和密度增加, 但细根P浓度变化不大, 说明后期植物细根吸收养分的能力有所减弱(Ostertag, 2001).新西兰的Franz Josef土壤年代序列后期, 同样发现细根中N、P浓度比早期更低, 也可表明成土后期植物细根养分吸收能力的减弱(Holdaway et al., 2011). ...
... 成土后期植物NASs的多样性显著升高.澳大利亚西南部Jurien Bay序列后期, 土壤总P含量降至6-10 mg·kg-1, 用稀有物种评估的植物NASs多样性增加了一倍, 与NASs相关的性状分离度增加了46% (Zemunik et al., 2015).加拿大养分贫瘠的沿海沙丘序列, 随着土壤年龄增加, 植物NASs的Simpson指数增加超过1.5倍(Roy-Bolduc et al., 2016).成土过程中总P的减少是导致植物NASs多样性增加的原因之一.澳大利亚西南部Jurien Bay土壤序列的总P含量与植物NASs多样性呈负相关关系, 总P的减少使植物产生多种NASs以获取土壤中的P (Zemunik et al., 2015).在澳大利亚西南部总P为50 mg·kg-1左右的土壤中, Xylomelum occidentale发育了特别的NASs, 既不释放大量有机酸也不形成菌根, 而是释放磷酸酶水解土壤中的RNA和磷脂(Zhong et al., 2021), 增加了成土后期NASs的丰富度. ...
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
生物固氮的研究进展及发展趋势 1 2015
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
生物固氮的研究进展及发展趋势 1 2015
... 微生物共生策略包括丛枝菌根(AM)、外生菌根(ECM)、兰花菌根(OM)、欧石南类菌根(ERM)和根瘤固N等(图2A-2D).根瘤共生体促进植物获取N的作用已被广泛报道(Pearson & Vitousek, 2002; 张武等, 2015; 王银柳等, 2021).菌根增强植物获取养分的能力主要包括通过外延菌丝拓展根系获取有效养分的空间和释放有机酸与酶等物质活化非有效态养分(蒋婧和宋明华, 2010; 杨应等, 2017; 屈明华等, 2019).研究发现, 发育了菌根策略的植物不再通过根毛或者表皮, 而是通过真菌吸收有效养分(Smith et al., 2004).多个实验室和原位研究均发现, AM和ECM共生体不仅有助于植物从受N限制的土壤中获取有效N (陈永亮等, 2014; Knoblochová et al., 2017; Bahadur et al., 2019), 而且可释放一定量的有机酸和磷酸酶活化土壤中的难溶态有机P, 增加土壤P的有效性, 再通过菌根将有效P传输给植物(Koide & Kabir, 2000; 宋勇春等, 2001; Nurfadilah et al., 2013; Smith et al., 2015).微生物非共生策略是指微生物不与根系形成共生体、但可固N或溶解矿物以提升植物获取有效养分的行为.目前较为明确的微生物非共生策略主要包括联合固N和自由微生物溶P等两类. ...
接种AM真菌对玉米和油菜种间竞争及土壤无机磷组分的影响 1 2012
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
接种AM真菌对玉米和油菜种间竞争及土壤无机磷组分的影响 1 2012
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
Xylomelum occidentale (Proteaceae) accesses relatively mobile soil organic phosphorus without releasing carboxylates 1 2021
... 成土后期植物NASs的多样性显著升高.澳大利亚西南部Jurien Bay序列后期, 土壤总P含量降至6-10 mg·kg-1, 用稀有物种评估的植物NASs多样性增加了一倍, 与NASs相关的性状分离度增加了46% (Zemunik et al., 2015).加拿大养分贫瘠的沿海沙丘序列, 随着土壤年龄增加, 植物NASs的Simpson指数增加超过1.5倍(Roy-Bolduc et al., 2016).成土过程中总P的减少是导致植物NASs多样性增加的原因之一.澳大利亚西南部Jurien Bay土壤序列的总P含量与植物NASs多样性呈负相关关系, 总P的减少使植物产生多种NASs以获取土壤中的P (Zemunik et al., 2015).在澳大利亚西南部总P为50 mg·kg-1左右的土壤中, Xylomelum occidentale发育了特别的NASs, 既不释放大量有机酸也不形成菌根, 而是释放磷酸酶水解土壤中的RNA和磷脂(Zhong et al., 2021), 增加了成土后期NASs的丰富度. ...
Weathering of primary mineral phosphate in the early stages of ecosystem development in the Hailuogou Glacier foreland chronosequence 1 2018
... 成土早期第一阶段(通常在几十年以内), 土壤N库由0开始逐渐积累, 绝大部分P以原生矿物的形式存在, 有效性较低(Walker & Syers, 1976; Vitousek et al., 1989; Turner & Condron, 2013; Zhou et al., 2018), 植物NASs以微生物策略和细根策略为主, 仅存在少量的特殊根系策略(图3).多种气候和地质条件下的研究均发现成土早期根瘤固N作用普遍存在(Pearson & Vitousek, 2002; Walker et al., 2003), 是成土早期植物获取N和土壤中N积累的主要途径.夏威夷土壤序列早期的Acacia koa植物主要通过根瘤固N作用获取N来满足生长所需(Pearson & Vitousek, 2002).新西兰Kanakana Dome高原的成土早期, Coriaria arborea的根瘤固N作用使得土壤有机层的N不断积累(Walker et al., 2003).在成土早期第二阶段, 土壤中已积累了一定浓度的有效N和P (图3), 由于菌根和细根策略可高效吸收低浓度的可溶性养分, 因此这两种策略的重要性在这一阶段有所增强.例如, 新西兰的Franz Josef土壤序列的早期, 植物主要通过细根吸收N和P来满足生长需求(Holdaway et al., 2011).美国五大湖地区冰川沉积物上发育的有效N不足的早期土壤中, 植物形成大量ECM菌丝体活化并吸收土壤中难溶性的N (Nave et al., 2013).尽管成土早期较低的土壤有效P含量可导致特殊根系策略发生(Zhou et al., 2020), 如南美的Velloziaceae植物发育毛状根(VR)溶解包裹于原生矿物中的P (Teodoro et al., 2019), 但由于细根和菌根也可溶解暴露于土壤溶液中的无机矿物P (Yao et al., 2001; Lyu et al., 2016), 因此, 这一时期的P捕获策略通常以菌根和细根为主(图3).例如, 澳大利亚西南部的Jurien Bay序列上, 成土早期AM植物的盖度超过了75%, 而排根和固沙根植物的盖度都未达到10% (Zemunik et al., 2015). ...
Rapid weathering processes of a 120-year-old chronosequence in the Hailuogou Glacier foreland, Mt. Gongga, SW China 1 2016
Changes of soil phosphorus speciation along a 120-year soil chronosequence in the Hailuogou Glacier retreat area (Gongga Mountain, SW China). 2 2013
... 自然条件下植被原生演替与成土作用通常同时进行, 成土过程中N和P等养分的含量和形态不断变化(Walker & Syers, 1976; Crew et al., 1995; Zhou et al., 2013), 导致植物NASs的组成和多样性随之改变(Zemunik et al., 2015).NASs的变化影响植物竞争养分的能力, 可引起植物的种间关系(陈梅梅等, 2009; Mayfield & Levine, 2010; 张宇亭等, 2012)、植物-土壤反馈(PSFs)(Teste et al., 2017)和生态位(Tedersoo et al., 2020)等发生变化.互补的NASs使共存植物共同利用附近的养分(Teste et al., 2014; Liu et al., 2015; Phoenix et al., 2020), 而竞争性的NASs会抑制邻近植物获取养分(Semchenko et al., 2018), 从而影响物种更替、群落结构(Nasto et al., 2014)、物种多样性(Lambers et al., 2011)和植被原生演替过程(Roy-Bolduc et al., 2016; Caplan et al., 2019). ...
 ,
, ,1,*, 彭飞2, 钟宏韬3, Hans LAMBERS3
,1,*, 彭飞2, 钟宏韬3, Hans LAMBERS3

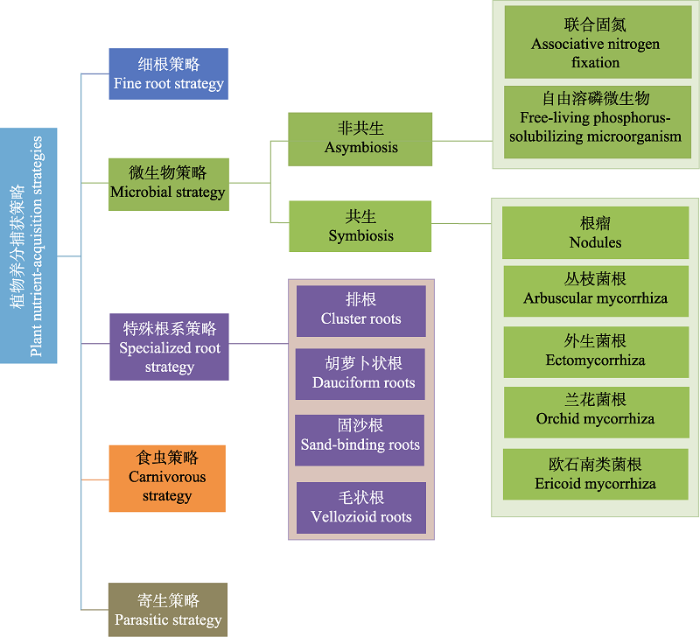 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT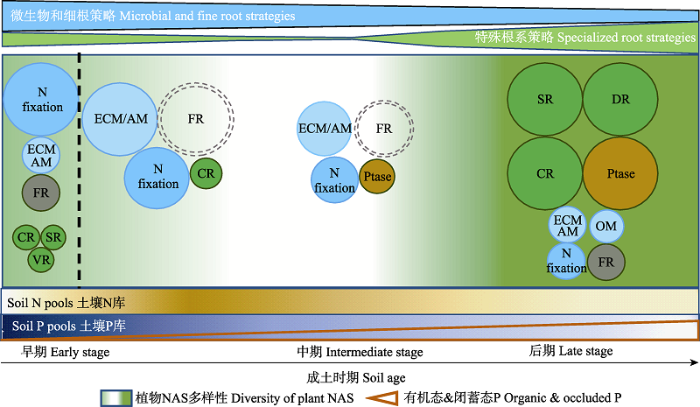 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT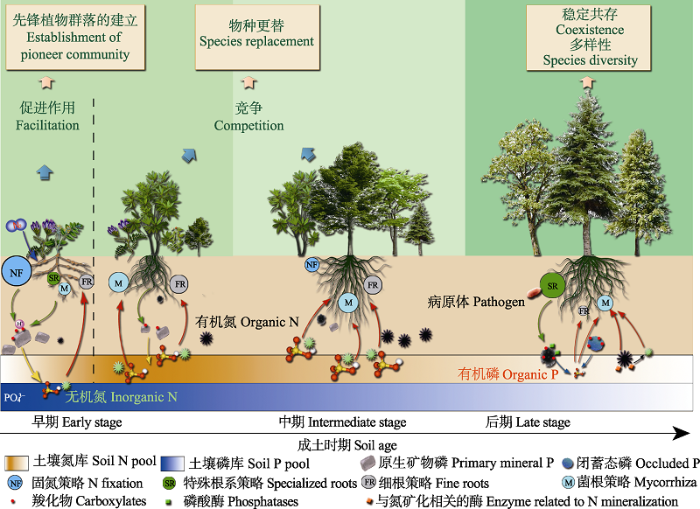 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}