Temporal variation and resorption of nutrients in plant culms and leaves in Hulun Buir grassland
Xiao-Jing ZHANG1,2, Xiao-Sa LIANG1,2, Wang MA1, Zheng-Wen WANG,,1,*1Erguna Forest-Steppe Ecotone Research Station, Institute of Applied Ecology, Chinese Academy of Sciences, Shenyang 110016, China 2University of Chinese Academy of Sciences, Beijing 100049, China
Abstract Aims Nutrient resorption in different organs of plants is important for plant nutrient use strategies and elemental biogeochemical cycles. Previous researches on nutrient resorption have focused on leaves, but neglected culms. Additionally, leaves of different species have been collected at the same time during the peak growth stage in previous studies, ignoring the different peak times of nutrient content of different species, which leads to the underestimation of nutrient resorption efficiency. Methods In order to explore the seasonal variation of nutrients and nutrient resorption efficiency in the culms and leaves of herbaceous plants, 22 common plants in Hulun Buir grassland were chosen as the research objects to determine the temporal dynamics of nutrient content during the growing season, and the resorption efficiency of nitrogen (N) and phosphorus (P) in plant culms and leaves. Important findings The content of N, P in plants had obvious temporal dynamics during the growing season, showing an increasing trend first and then a decreasing trend. For most of the 22 species, the maximum content occurred in the middle and late August, but the peak time differed among different species. The resorption efficiency of N in leaves was higher than in culms, but that of P did not differ between leaves and culms. Nutrient resorption efficiency of plants was closely related to the nutrient content at the senescence stage, but not to that at the growth stage. In previous studies, different plants were sampled at the same time during the growth stage, which led to underestimation of N and P resorption efficiency of culms and leaves. This study re-examined the sampling strategy in nutrient resorption studies, and showed that the sampling time of mature tissues in the growth stage could be determined according to the peak nutrient content time of different species. Keywords:herbaceous;element content;temporal dynamics;plant organ;nutrient resorption
PDF (1732KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 张效境, 梁潇洒, 马望, 王正文. 呼伦贝尔草地植物茎秆和叶片中养分的时间动态与回收. 植物生态学报, 2021, 45(7): 738-748. DOI: 10.17521/cjpe.2021.0125 ZHANG Xiao-Jing, LIANG Xiao-Sa, MA Wang, WANG Zheng-Wen. Temporal variation and resorption of nutrients in plant culms and leaves in Hulun Buir grassland. Chinese Journal of Plant Ecology, 2021, 45(7): 738-748. DOI: 10.17521/cjpe.2021.0125
氮(N)、磷(P)在植物生长发育过程中起关键作用(Han et al., 2005), 是陆地生态系统主要的限制性营养元素。植物中N、P养分主要源于两部分, 其一是直接吸收土壤中的有效矿质养分, 其二是养分回收。养分回收指养分从衰老的植物组织转移到新生组织或贮藏组织中再利用的过程(Killingbeck, 1986)。研究表明陆地植物叶片平均约62.1%的N和64.9%的P被回收至多年生组织或者器官中(Vergutz et al., 2012), 因此养分回收不仅能降低植物对土壤养分供给的依赖(Aerts & Chapin III, 1999), 还能影响凋落物分解、养分循环等生态过程。衡量植物养分回收能力的指标有养分回收效率(叶片脱落前回收养分的比例)和养分回收能力(衰老过程中养分降低的水平)(Killingbeck, 1996)。在一个完整的生长周期内, 植物的N、P含量并不是一成不变的(Chapin III & Kedrowski, 1983; Estiarte & Peñuelas, 2015), 随着生育阶段的变化, 植物会调整养分的吸收及不同器官内养分的分配, 表现出明显的时间动态。由于不同植物养分含量达到峰值的时间不同, 传统的养分回收的测定方式可能导致对植物养分回收效率的估测存在偏差。
温带草地构成了我国草地的主体, 不仅发挥着重要的经济效益和社会效益(石岳等, 2013), 还是我国北方重要的生态屏障(沈海花等, 2016; 潘庆民等, 2018), 在生物多样性维持、碳固持等方面发挥着不可或缺的作用(方精云等, 2010)。养分是温带草地生态系统生产力重要的限制因子, 目前已有一些研究对温带草地植物的养分回收进行了探究(Huang et al., 2012; Lü et al., 2012), 然而这些研究只关注少数几个优势物种, 且研究对象集中于植物叶片。次有种及从属种在维持植物群落稳定的过程中发挥着重要作用, 尤其是当群落遭受环境胁迫时(Mariotte et al., 2015), 而草地植物茎秆是植物体的重要组成部分(任海彦等, 2009), 在结构与功能上与叶片存在显著差异, 因此分析不同植物茎秆和叶片的养分回收模式有助于我们进一步理解草地植物的生存与竞争策略(Eckstein et al., 1999; Yuan & Chen, 2015), 同时也有助于生态系统养分循环过程与机制理论体系的优化和完善(McGroddy et al., 2004; Vergutz et al., 2012)。
样品采集方法: 设置6块10 m × 10 m固定样方, 每个物种在固定样方内选取10株以上长势良好植株, 采集地上部分, 形成混合样。本研究共采集22种呼伦贝尔草原常见种, 具体采样物种名称详见表1。
Table 1 表1 表1呼伦贝尔地区取样物种名、功能类群与茎秆和叶片的区分 Table 1Sampling species name, functional groups of plants in Hulun Buir area and whether culm and leaf were distinguished or not
物种 Species
生活型 Life form
区分茎秆和叶片 Distinguishing culm and leaf
羊草 Leymus chinensis
多年生禾草 Perennial grasses
√
狼针草 Stipa baicalensis
多年生禾草 Perennial grasses
×
糙隐子草 Cleistogenes squarrosa
多年生禾草 Perennial grasses
√
草 Koeleria macrantha
多年生禾草 Perennial grasses
×
柄状薹草 Carex pediformis
多年生莎草 Perennial sedges
×
寸草 Carex duriuscula
多年生莎草 Perennial sedges
×
广布野豌豆 Vicia cracca
多年生豆科 Perennial legumes
√
披针叶野决明 Thermopsis lanceolata
多年生豆科 Perennial legumes
√
狭叶沙参 Adenophora gmelinii
多年生杂草 Perennial forbs
√
龙蒿 Artemisia dracunculus
多年生杂草 Perennial forbs
√
阿尔泰狗娃花 Aster altaicus
多年生杂草 Perennial forbs
√
麻花头 Klasea centauroides
多年生杂草 Perennial forbs
√
裂叶蒿 Artemisia tanacetifolia
多年生杂草 Perennial forbs
×
冷蒿 Artemisia frigida
多年生杂草 Perennial forbs
√
达乌里芯芭 Cymbaria daurica
多年生杂草 Perennial forbs
√
瓣蕊唐松草 Thalictrum petaloideum
多年生杂草 Perennial forbs
√
白头翁 Pulsatilla chinensis
多年生杂草 Perennial forbs
×
二裂委陵菜 Potentilla bifurca
多年生杂草 Perennial forbs
×
星毛委陵菜 Potentilla acaulis
多年生杂草 Perennial forbs
×
菊叶委陵菜 Potentilla tanacetifolia
多年生杂草 Perennial forbs
×
野鸢尾 Iris dichotoma
多年生杂草 Perennial forbs
×
囊花鸢尾 Iris ventricosa
多年生杂草 Perennial forbs
×
√, 区分茎秆和叶片; ×, 不区分茎秆和叶片。 √, distinguishing culm and leaf; ×, not distinguishing culm and leaf.
植物养分回收效率为考虑质量损失校正因子(MLCF)的衰老组织和成熟组织相比养分含量下降的百分比(Kazakou et al., 2007; Vergutz et al., 2012; Yuan & Chen, 2015), 具体公式如下:
$N u R E=\left(1-\frac{N u t_{\text {senesced }}}{N u t_{\text {green }}} M L C F\right) \times 100 \%$ 式中, Nutgreen指生长盛期叶片最高养分含量, Nutsenesced衰老期叶片养分含量, MLCF是质量损失校正因子, 指衰老叶片干质量与成熟叶片干质量的比值(Vergutz et al., 2012)。本研究通过对植物生长盛期多次取样, 获取养分含量最大值作为成熟叶片的养分含量计算养分回收效率, 并使用禾草和非禾本科草本植物MLCF对不同类群植物的回收效率进行校正(Vergutz et al., 2012)。植物茎秆的养分回收效率计算与叶片相同。
Fig. 1Temporal variation of nitrogen (N) content in leaves and culms. A, Artemisia dracunculus. B, Cleistogenes squarrosa. C, Adenophora gmelinii. D, Vicia cracca. E, Cymbaria daurica. F, Aster altaicus. G, Klasea centauroides. H, Leymus chinensis. I, Thermopsis lanceolata. J, Artemisia frigida. K, Thalictrum petaloideum. Circle, leaves; triangle, culms. The solid line indicates that leaf N content is related to sampling time (p < 0.05); the dotted line indicates that culm N content is related to sampling time (p < 0.05).
Fig. 2Temporal variation of phosphorus (P) content in leaves and culms. A, Artemisia dracunculus. B, Cleistogenes squarrosa. C, Adenophora gmelinii. D, Vicia cracca. E, Cymbaria daurica. F, Aster altaicus. G, Klasea centauroides. H, Leymus chinensis. I, Thermopsis lanceolata. J, Artemisia frigida. K, Thalictrum petaloideum. Circle, leaves; triangle, culms. The solid line indicates that leaf P content is related to sampling time (p < 0.05); The dotted line indicates that culm P content is related to sampling time (p < 0.05).
Fig. 4Correlations between nitrogen (N) and phosphorus (P) resorption efficiency. Circle, leaves; triangle, culms. The solid line indicates that leaf N resorption efficiency is related to P resorption efficiency (p < 0.05).
Fig. 5Correlations between nitrogen (N) and phosphorus (P) contents at the growth and senescence stages and nutrient resorption efficiency. A, N content and N resorption efficiency at the growth stage. B, P content and P resorption efficiency at the growth stage. C, N content and N resorption efficiency at the senescence stage. D, P content and P resorption efficiency at the senescence stage. Circle, leaves; triangle, culms. The solid line indicates that leaf resorption efficiency is related to nutrient content (p < 0.05); the dotted line indicates that culm resorption efficiency is related to nutrient content (p < 0.05).
Table 4 表4 表4不同计算方法获得的呼伦贝尔草地植物茎秆和叶片氮(N)、磷(P)回收效率的差异状况(平均值±标准差) Table 4Difference in nitrogen (N) and phosphorus (P) resorption efficiency of Hulun Buirr grassland plant culms and leaves obtained by different calculation methods (mean ± SD)
器官 Organ
养分 Nutrient
样本量 n
回收效率 Resorption efficiency (%)
低估值 Underestimation value (%)
基于养分最大含量 Based on maximum nutrient content
基于8月20日养分含量 Based on nutrient content of August 20
叶片 Leaf
N
22
66.75 ± 7.70
64.69 ± 8.80
2.06 ± 3.31
P
22
77.26 ± 7.97
71.32 ± 14.32
5.94 ± 9.35
茎秆 Culm
N
11
61.65 ± 4.24
58.72 ± 6.55
2.93 ± 4.85
P
11
78.33 ± 9.90
70.93 ± 15.33
7.39 ± 11.81
低估值中数值加粗表示配对t检验差异显著(p < 0.05)。 Values in bold fonts mean significant underestimation values based on paired t-tests (p < 0.05).
AertsR (1996). Nutrient resorption from senescing leaves of perennials: Are there general patterns Journal of Ecology, 84, 597-608. DOI:10.2307/2261481URL [本文引用: 1]
AertsR, ChapinIII FS (1999). The mineral nutrition of wild plants revisited: are-evaluation of processes and patterns Advances in Ecological Research, 30, 1-67. [本文引用: 2]
BrantAN, ChenHYH (2015). Patterns and mechanisms of nutrient resorption in plants Critical Reviews in Plant Sciences, 34, 471-486. DOI:10.1080/07352689.2015.1078611URL [本文引用: 1]
ChapinIII FS, KedrowskiRA (1983). Seasonal-changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees Ecology, 64, 376-391. DOI:10.2307/1937083URL [本文引用: 4]
ChapinIII FS, MoilanenL (1991). Nutritional controls over nitrogen and phosphorus resorption from Alaskan birch leaves Ecology, 72, 709-715. DOI:10.2307/2937210URL [本文引用: 1]
CharlesedwardsDA, StutzelH, FerrarisR, BeechDF (1987). An analysis of spatial variation in the nitrogen-content of leaves from different horizons within a canopy Annals of Botany, 60, 421-426. DOI:10.1093/oxfordjournals.aob.a087463URL [本文引用: 1]
DuEZ, TerrerC, PellegriniAFA, AhlströmA, vanLissa CJ, ZhaoX, XiaN, WuXH, JacksonRB (2020). Global patterns of terrestrial nitrogen and phosphorus limitation Nature Geoscience, 13, 221-226. DOI:10.1038/s41561-019-0530-4URL [本文引用: 2]
EcksteinRL, KarlssonPS, WeihM (1999). Leaf life span and nutrient resorption as determinants of plant nutrient conservation in temperate-arctic regions New Phytologist, 143, 177-189. DOI:10.1046/j.1469-8137.1999.00429.xURL [本文引用: 1]
FangJY, YangYH, MaWH, MohammatA, ShenHH (2010). Ecosystem carbon stocks and their changes in China’s grasslands Science China Life Sciences, 40, 566-576. [本文引用: 1]
FreschetGT, CornelissenJHC, vanLogtestijn RSP, AertsR (2010). Substantial nutrient resorption from leaves, stems and roots in a subarctic flora: What is the link with other resource economics traits New Phytologist, 186, 879-889. DOI:10.1111/nph.2010.186.issue-4URL [本文引用: 2]
HanWX, FangJY, GuoDL, ZhangY (2005). Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China New Phytologist, 168, 377-385. DOI:10.1111/nph.2005.168.issue-2URL [本文引用: 2]
HättenschwilerS, AeschlimannB, CoûteauxMM, RoyJ, BonalD (2008). High variation in foliage and leaf litter chemistry among 45 tree species of a neotropical rainforest community New Phytologist, 179, 165-175. DOI:10.1111/j.1469-8137.2008.02438.xPMID:18422903 [本文引用: 2] Distinct ecosystem level carbon : nitrogen : phosphorus (C : N : P) stoichiometries in forest foliage have been suggested to reflect ecosystem-scale selection for physiological strategies in plant nutrient use. Here, this hypothesis was explored in a nutrient-poor lowland rainforest in French Guiana. Variation in C, N and P concentrations was evaluated in leaf litter and foliage from neighbour trees of 45 different species, and the litter concentrations of major C fractions were also measured. Litter C ranged from 45.3 to 52.4%, litter N varied threefold (0.68-2.01%), and litter P varied seven-fold (0.009-0.062%) among species. Compared with foliage, mean litter N and P concentrations decreased by 30% and 65%, respectively. Accordingly, the range in mass-based N : P shifted from 14 to 55 in foliage to 26 to 105 in litter. Resorption proficiencies indicated maximum P withdrawal in most species, but with a substantial increase in variation in litter P compared with foliage. These data suggest that constrained ecosystem-level C : N : P ratios do not preclude the evolution of highly diversified strategies of nutrient use and conservation among tropical rainforest tree species. The resulting large variation in litter quality will influence stoichiometric constraints within the decomposer food web, with potentially far-ranging consequences on nutrient dynamics and plant-soil feedbacks.
HeJS, WangL, FlynnDFB, WangXP, MaWH, FangJY (2008). Leaf nitrogen:phosphorus stoichiometry across Chinese grassland biomes Oecologia, 155, 301-310. DOI:10.1007/s00442-007-0912-yURL [本文引用: 1]
HuangJY, YuHL, WangB, LiLH, XiaoGJ, YuanZY (2012). Nutrient resorption based on different estimations of five perennial herbaceous species from the grassland in Inner Mongolia, China Journal of Arid Environments, 76, 1-8. DOI:10.1016/j.jaridenv.2011.08.018URL [本文引用: 1]
KazakouE, GarnierE, NavasML, RoumetC, CollinC, LaurentG (2007). Components of nutrient residence time and the leaf economics spectrum in species from Mediterranean old-fields differing in successional status Functional Ecology, 21, 235-245. DOI:10.1111/fec.2007.21.issue-2URL [本文引用: 2]
KillingbeckKT (1984). Nitrogen and phosphorus resorption dynamics of five tree species in a Kansas gallery forest American Midland Naturalist, 111, 155-164. DOI:10.2307/2425554URL [本文引用: 1]
KillingbeckKT (1986). Litterfall dynamics and element use efficiency in a Kansas gallery forest American Midland Naturalist, 116, 180-189. DOI:10.2307/2425950URL [本文引用: 1]
KillingbeckKT (1996). Nutrients in senesced leaves: keys to the search for potential resorption and resorption proficiency Ecology, 77, 1716-1727. DOI:10.2307/2265777URL [本文引用: 1]
KobeRK, LepczykCA, IyerM (2005). Resorption efficiency decreases with increasing green leaf nutrients in a global data set. Ecology, 86, 2780-2792. DOI:10.1890/04-1830URL [本文引用: 2]
KoerselmanW, MeulemanAFM (1996). The vegetation N:P ratio: a new tool to detect the nature of nutrient limitation Journal of Applied Ecology, 33, 1441-1450. DOI:10.2307/2404783URL [本文引用: 1]
LiuJQ, WangXY, GuoY, WangSL, ZhouL, DaiLM, YuDP (2015). Seasonal dynamics and resorption efficiencies of foliar nutrients in three dominant woody plants that grow at the treeline on Changbai Mountain Acta Ecologica Sinica, 35, 165-171. [本文引用: 3]
McGroddyME, DaufresneT, HedinLO (2004). Scaling of C:N stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios Ecology, 85, 2390-2401. DOI:10.1890/03-0351URL [本文引用: 1]
PanQM, XueJG, TaoJ, XuMY, ZhangWH (2018). Current status of grassland degradation and measures for grassland restoration in Northern China Chinese Science Bulletin, 63, 1642-1650. [本文引用: 1]
ReedSC, TownsendAR, DavidsonEA, ClevelandCC (2012). Stoichiometric patterns in foliar nutrient resorption across multiple scales New Phytologist, 196, 173-180. DOI:10.1111/nph.2012.196.issue-1URL [本文引用: 1]
RenHY, ZhengSX, BaiYF (2009). Effects of grazing on foliage biomass allocation of grassland communities in Xilin River basin, Inner Mongolia Chinese Journal of Plant Ecology, 33, 1065-1074. [本文引用: 1]
ShiY, MaYL, MaWH, LiangCZ, ZhaoXQ, FangJY, HeJS (2013). Large scale patterns of forage yield and quality across Chinese grasslands Chinese Science Bulletin, 58, 226-239. [本文引用: 1]
SunSC, ChenLZ (2001). Leaf nutrient dynamics and resorption efficiency of Quercus liaotungensis in the Dongling Mountain region Acta Phytoecologica Sinica, 25, 76-82. [本文引用: 2]
TsujiiY, OnodaY, KitayamaK (2017). Phosphorus and nitrogen resorption from different chemical fractions in senescing leaves of tropical tree species on Mount Kinabalu, Borneo Oecologia, 185, 171-180. DOI:10.1007/s00442-017-3938-9PMID:28871400 [本文引用: 1] Nutrient resorption, a process by which plants degrade organic compounds and resorb their nutrients from senescing tissues, is a crucial plant function to increase growth and fitness in nutrient-poor environments. Tropical trees on phosphorus (P)-poor soils are particularly known to have high P-resorption efficiency (PRE, the percentage of P resorbed from senescing leaves before abscission per total P in green leaves). However, the biochemical mechanisms underlying this greater PRE remain unclear. In this study, we determined the P concentration in easily soluble, nucleic acid, lipid and residual fractions for green and senescent leaves of 22 tree species from three sites, which differed in P availability, on the lower flanks of Mt. Kinabalu, Borneo. PRE varied from 24 to 93% and was higher in species from the P-poor site. P-resorption rate was greatest from the lipid fraction, the nucleic acid fraction, and lowest in the easily soluble fraction and the residual fraction when all the species were pooled. For species with higher PRE, P-resorption rate of the residual fraction was relatively high and was comparable in magnitude to that of the other labile fractions. This suggests that tree species inhabiting P-poor environments increased PRE by improving the degradation of recalcitrant compounds. This study suggests that plants selectively degrade organic compounds depending on environmental conditions, which is a key mechanism underlying the variation of PRE.
van HeerwaardenLM, ToetS, AertsR (2003). Current measures of nutrient resorption efficiency lead to a substantial underestimation of real resorption efficiency: facts and solutions Oikos, 101, 664-669. DOI:10.1034/j.1600-0706.2003.12351.xURL [本文引用: 1]
VergutzL, ManzoniS, PorporatoA, NovaisRF, JacksonRB (2012). Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants Ecological Monographs, 82, 205-220. DOI:10.1890/11-0416.1URL [本文引用: 7]
WangHY, WangZW, DingR, HouSL, YangGJ, LüXT, HanXG (2018). The impacts of nitrogen deposition on community N:P stoichiometry do not depend on phosphorus availability in a temperate meadow steppe Environmental Pollution, 242, 82-89. DOI:10.1016/j.envpol.2018.06.088URL [本文引用: 1]
XingXR, HanXG, ChenLZ (2000). A review on research of plant nutrient use efficiency Chinese Journal of Applied Ecology, 11, 785-790. PMID:11767544 [本文引用: 1] The concept of nutrient use efficiency is the central to the understanding of ecosystem function. We reviewed the concept of nutrient use efficiency and resorption, its expression and calculation, affecting factors and biochemical basis, we also analyzed the current problems in the studies of nutrient use efficiency, and pointed out the directions for future research work of this field. [ 邢雪荣, 韩兴国, 陈灵芝 (2000). 植物养分利用效率研究综述 应用生态学报, 11, 785-790.] PMID:11767544 [本文引用: 1]
YasumuraY, HikosakaK, HiroseT (2006). Seasonal changes in photosynthesis, nitrogen content and nitrogen partitioning in Lindera umbellata leaves grown in high or low irradiance Tree Physiology, 26, 1315-1323. PMID:16815833 [本文引用: 1] Seasonal changes in photosynthetic capacity, leaf nitrogen (N) content and N partitioning were studied from before leaf maturation (spring) until death (autumn) in high- and low-light-exposed leaves of a deciduous shrub, Lindera umbellata var. membranacea (Maxim.) Momiyama growing in a natural forest in northeast Japan. In spring, light-saturated photosynthetic rate (Pmax) was low despite high leaf N and Rubisco contents, indicating that the photosynthetic apparatus was not yet functionally developed. Rubisco seemed to be only partially active. In summer and autumn, Pmax per unit leaf N increased and changes in Pmax were correlated with changes in leaf N and two photosynthetic components, Rubisco and chlorophyll. Changes in these components paralleled the changes in leaf N. During leaf senescence, about 70% of leaf N was resorbed. Metabolic proteins that accounted for the majority of leaf N in summer were highly degradable and more than sufficient to explain the high N-resorption efficiency. Structural proteins represented only a small part of leaf N and were relatively resistant to degradation and thus contributed little to N resorption. Leaf N partitioning between metabolic and structural proteins determined the amount of retranslocatable N, but did not strictly determine the N content of a dead leaf or N-resorption efficiency.
YuanZY, ChenHYH (2009). Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation Global Ecology and Biogeography, 18, 11-18. DOI:10.1111/geb.2009.18.issue-1URL [本文引用: 1]
YuanZY, ChenHYH (2015). Negative effects of fertilization on plant nutrient resorption Ecology, 96, 373-380. PMID:26240859 [本文引用: 3] Plants in infertile habitats are thought to have a high rate of nutrient resorption to enable them reuse nutrients more efficiently than those in fertile habitats. However, there is still much debate on how plant nutrient resorption responds to nutrient availability. Here we used a meta-analysis from a global data set of 9703 observations at 306 sites from 508 published articles to examine the effects of nitrogen (N) and phosphorus (P) fertilization on plant foliar N and P concentrations and resorption efficiency. We found that N fertilization enhanced N concentration in green leaves by 27% and P fertilization enhanced green-leaf P by 73% on average. The N and P concentrations in senesced leaves also increased with respective nutrient fertilization. Resorption efficiencies (percentage of nutrient recovered from senescing leaves) of both N and P declined in response to respective nutrient fertilization. Combined N and P fertilization also had negative effects on both N and P resorption efficiencies. Whether nutrient resorption efficiency differs among plant growth types and among ecosystems, however, remains uncertain due to the limited sample sizes when analyzed by plant growth types or ecosystem types. Our analysis indicates that fertilization decreases plant nutrient resorption and the view that nutrient resorption is a critical nutrient conservation strategy for plants in nutrient-poor environments cannot be abandoned. The response values to fertilization presented in our analysis can help improve biogeochemical models.
ZongN, ShiPL, GengSB, MaWL (2017). Nitrogen and phosphorus resorption efficiency of forests in North China Chinese Journal of Eco-Agriculture, 25, 520-529. [本文引用: 1]
Plasticity in leaf and stem nutrient resorption proficiency potentially reinforces plant-soil feedbacks and microscale heterogeneity in a semi-arid grassland 1 2012
... 温带草地构成了我国草地的主体, 不仅发挥着重要的经济效益和社会效益(石岳等, 2013), 还是我国北方重要的生态屏障(沈海花等, 2016; 潘庆民等, 2018), 在生物多样性维持、碳固持等方面发挥着不可或缺的作用(方精云等, 2010).养分是温带草地生态系统生产力重要的限制因子, 目前已有一些研究对温带草地植物的养分回收进行了探究(Huang et al., 2012; Lü et al., 2012), 然而这些研究只关注少数几个优势物种, 且研究对象集中于植物叶片.次有种及从属种在维持植物群落稳定的过程中发挥着重要作用, 尤其是当群落遭受环境胁迫时(Mariotte et al., 2015), 而草地植物茎秆是植物体的重要组成部分(任海彦等, 2009), 在结构与功能上与叶片存在显著差异, 因此分析不同植物茎秆和叶片的养分回收模式有助于我们进一步理解草地植物的生存与竞争策略(Eckstein et al., 1999; Yuan & Chen, 2015), 同时也有助于生态系统养分循环过程与机制理论体系的优化和完善(McGroddy et al., 2004; Vergutz et al., 2012). ...
Subordinate plants mitigate drought effects on soil ecosystem processes by stimulating fungi 1 2015
... 温带草地构成了我国草地的主体, 不仅发挥着重要的经济效益和社会效益(石岳等, 2013), 还是我国北方重要的生态屏障(沈海花等, 2016; 潘庆民等, 2018), 在生物多样性维持、碳固持等方面发挥着不可或缺的作用(方精云等, 2010).养分是温带草地生态系统生产力重要的限制因子, 目前已有一些研究对温带草地植物的养分回收进行了探究(Huang et al., 2012; Lü et al., 2012), 然而这些研究只关注少数几个优势物种, 且研究对象集中于植物叶片.次有种及从属种在维持植物群落稳定的过程中发挥着重要作用, 尤其是当群落遭受环境胁迫时(Mariotte et al., 2015), 而草地植物茎秆是植物体的重要组成部分(任海彦等, 2009), 在结构与功能上与叶片存在显著差异, 因此分析不同植物茎秆和叶片的养分回收模式有助于我们进一步理解草地植物的生存与竞争策略(Eckstein et al., 1999; Yuan & Chen, 2015), 同时也有助于生态系统养分循环过程与机制理论体系的优化和完善(McGroddy et al., 2004; Vergutz et al., 2012). ...
Scaling of C:N stoichiometry in forests worldwide: implications of terrestrial redfield-type ratios 1 2004
... 温带草地构成了我国草地的主体, 不仅发挥着重要的经济效益和社会效益(石岳等, 2013), 还是我国北方重要的生态屏障(沈海花等, 2016; 潘庆民等, 2018), 在生物多样性维持、碳固持等方面发挥着不可或缺的作用(方精云等, 2010).养分是温带草地生态系统生产力重要的限制因子, 目前已有一些研究对温带草地植物的养分回收进行了探究(Huang et al., 2012; Lü et al., 2012), 然而这些研究只关注少数几个优势物种, 且研究对象集中于植物叶片.次有种及从属种在维持植物群落稳定的过程中发挥着重要作用, 尤其是当群落遭受环境胁迫时(Mariotte et al., 2015), 而草地植物茎秆是植物体的重要组成部分(任海彦等, 2009), 在结构与功能上与叶片存在显著差异, 因此分析不同植物茎秆和叶片的养分回收模式有助于我们进一步理解草地植物的生存与竞争策略(Eckstein et al., 1999; Yuan & Chen, 2015), 同时也有助于生态系统养分循环过程与机制理论体系的优化和完善(McGroddy et al., 2004; Vergutz et al., 2012). ...
中国北方草原退化现状与恢复技术 1 2018
... 温带草地构成了我国草地的主体, 不仅发挥着重要的经济效益和社会效益(石岳等, 2013), 还是我国北方重要的生态屏障(沈海花等, 2016; 潘庆民等, 2018), 在生物多样性维持、碳固持等方面发挥着不可或缺的作用(方精云等, 2010).养分是温带草地生态系统生产力重要的限制因子, 目前已有一些研究对温带草地植物的养分回收进行了探究(Huang et al., 2012; Lü et al., 2012), 然而这些研究只关注少数几个优势物种, 且研究对象集中于植物叶片.次有种及从属种在维持植物群落稳定的过程中发挥着重要作用, 尤其是当群落遭受环境胁迫时(Mariotte et al., 2015), 而草地植物茎秆是植物体的重要组成部分(任海彦等, 2009), 在结构与功能上与叶片存在显著差异, 因此分析不同植物茎秆和叶片的养分回收模式有助于我们进一步理解草地植物的生存与竞争策略(Eckstein et al., 1999; Yuan & Chen, 2015), 同时也有助于生态系统养分循环过程与机制理论体系的优化和完善(McGroddy et al., 2004; Vergutz et al., 2012). ...
中国北方草原退化现状与恢复技术 1 2018
... 温带草地构成了我国草地的主体, 不仅发挥着重要的经济效益和社会效益(石岳等, 2013), 还是我国北方重要的生态屏障(沈海花等, 2016; 潘庆民等, 2018), 在生物多样性维持、碳固持等方面发挥着不可或缺的作用(方精云等, 2010).养分是温带草地生态系统生产力重要的限制因子, 目前已有一些研究对温带草地植物的养分回收进行了探究(Huang et al., 2012; Lü et al., 2012), 然而这些研究只关注少数几个优势物种, 且研究对象集中于植物叶片.次有种及从属种在维持植物群落稳定的过程中发挥着重要作用, 尤其是当群落遭受环境胁迫时(Mariotte et al., 2015), 而草地植物茎秆是植物体的重要组成部分(任海彦等, 2009), 在结构与功能上与叶片存在显著差异, 因此分析不同植物茎秆和叶片的养分回收模式有助于我们进一步理解草地植物的生存与竞争策略(Eckstein et al., 1999; Yuan & Chen, 2015), 同时也有助于生态系统养分循环过程与机制理论体系的优化和完善(McGroddy et al., 2004; Vergutz et al., 2012). ...
Stoichiometric patterns in foliar nutrient resorption across multiple scales 1 2012
 ,
, ,1,*
,1,*

 草 Koeleria macrantha
草 Koeleria macrantha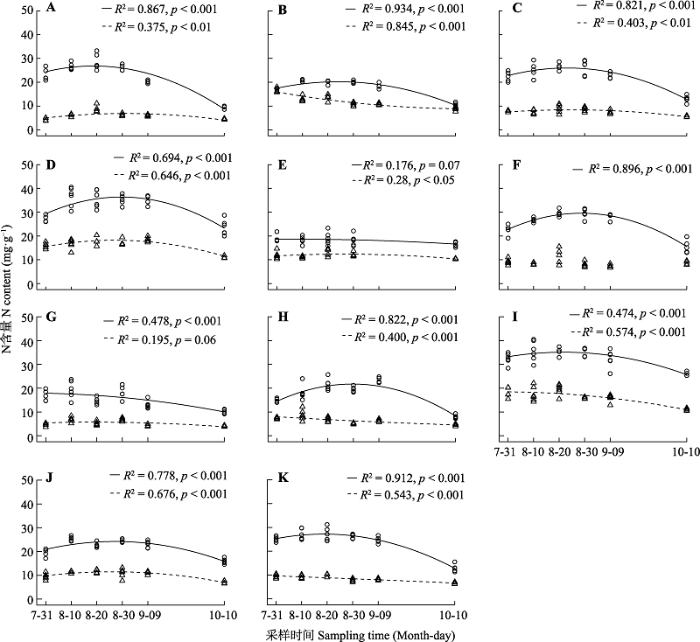 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT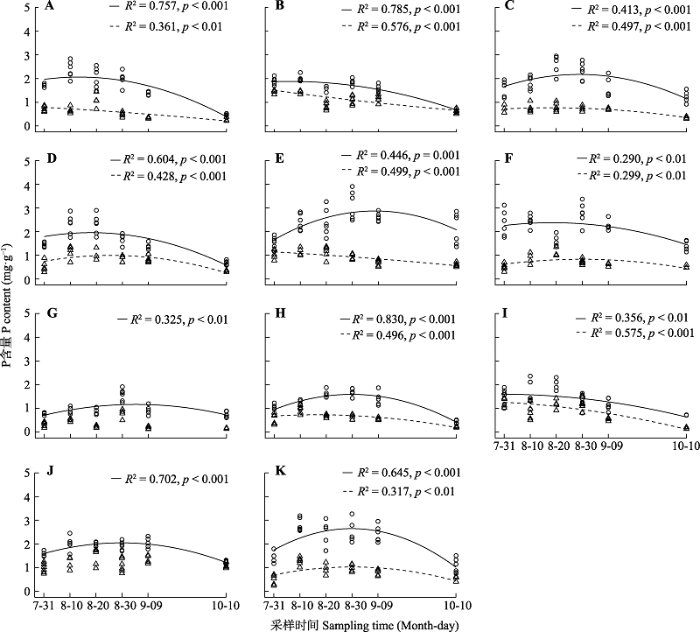 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT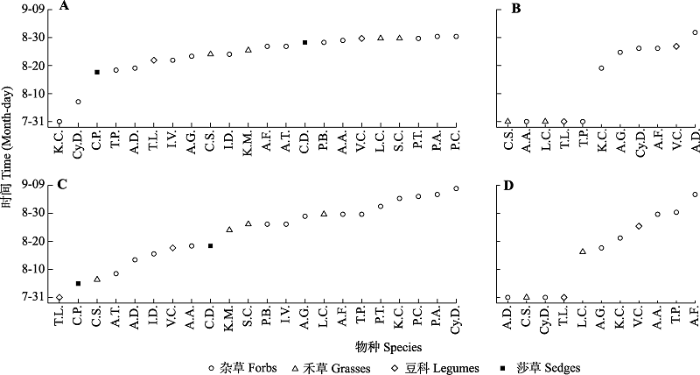 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT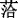 草; L.C., 羊草; P.A., 星毛委陵菜; P.B., 二裂委陵菜; P.C., 白头翁; P.T., 菊叶委陵菜; S.B., 狼针草; T.L., 披针叶野决明; T.P., 瓣蕊唐松草; V.C., 广布野豌豆。
草; L.C., 羊草; P.A., 星毛委陵菜; P.B., 二裂委陵菜; P.C., 白头翁; P.T., 菊叶委陵菜; S.B., 狼针草; T.L., 披针叶野决明; T.P., 瓣蕊唐松草; V.C., 广布野豌豆。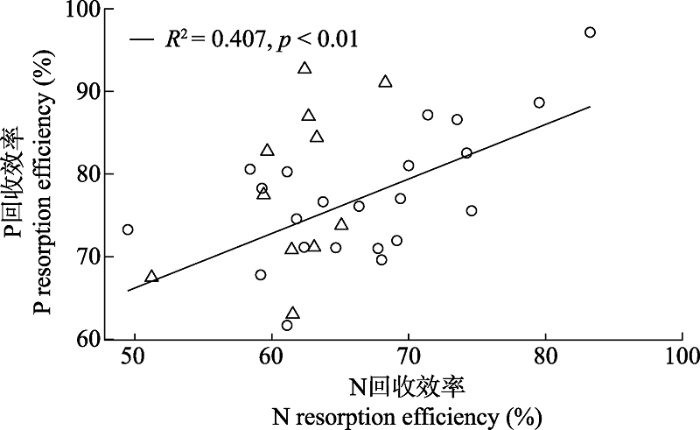 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}