Effects of nitrogen addition at different levels on soil microorganisms in saline-alkaline grassland of northern China
Jian-Qiang YANG1, Hua-Jie DIAO2,3,4, Shu-Ya HU4, Chang-Hui WANG,2,3,4,*1College of Life Science, Shanxi Agricultural University, Taigu, Shanxi 030801, China 2College of Grassland Science, Shanxi Agricultural University, Taigu, Shanxi 030801, China 3Youyu Loess Plateau Grassland Ecosystem Research Station, Youyu, Shanxi 037200, China 4State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Bejing 100093, China
National Natural Science Foundation of China(31770526) National Key R&D Program of China(2017YFA0604802) Research Projects of Shanxi Province’s Doctoral Graduates and Postdoctoral Researchers Working in Shanxi Province(SXBYKY2021052)
Abstract Aims Nitrogen (N) availability is an important limiting factor for grassland ecosystem productivity, and soil microorganisms are the main driving factor on soil N transformation. With the increase of atmospheric N deposition, the response of soil microbial characteristics to different nitrogen input levels is still unclear especially in saline-alkaline grassland. Methods The experiment was conducted in Youyu Loess Plateau Grassland Ecosystem Research Station, Shanxi Province. Eight different nitrogen addition levels were set, which were 0, 1, 2, 4, 8, 16, 24 and 32 g·m-2·a-1, respectively. The Ammonia-oxidizing microorganisms (i.e. ammonia-oxidizing bacteria (AOB) and ammonia- oxidizing archaea (AOA)) abundance, soil bacterial and fungal abundance, as well as soil microbial biomass carbon (MBC) and nitrogen (MBN) content were measured in the growing season (May to September) in 2020 to explore the effects of different levels of N addition on soil microbial characteristics. Important findings Our results showed that: (1) Sampling month had a significant effect on soil AOB, bacteria, fungal abundance and MBC, MBN content due to the variation in soil temperature and soil water content in the growing season. (2) N addition had a significant effect on soil AOB abundance, while had no effects on soil MBC, MBN content, and bacterial and fungal composition. (3) Higher N addition (24 and 32 g·m-2·a-1) significantly increased the abundance of ammonia-oxidizing bacteria (AOB) on the early growth stage (May to August), while having no effect on late growth period (September). (4) Soil microorganisms were mainly regulated by soil cations concentrations and soil pH values, which explained the variation of soil microorganisms by 21.8% and 17.2%, respectively. We found that soil microorganisms were not sensitive to N addition in saline-alkaline grassland, while AOB showed a significant increase under higher N addition, indicating that higher N addition might promote soil N transformation. Keywords:saline alkaline grassland;nitrogen addition;Ammonia-oxidizing microorganisms;bacteria;fungi
PDF (1344KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 杨建强, 刁华杰, 胡姝娅, 王常慧. 不同水平氮添加对盐渍化草地土壤微生物特征的影响. 植物生态学报, 2021, 45(7): 780-789. DOI: 10.17521/cjpe.2021.0072 YANG Jian-Qiang, DIAO Hua-Jie, HU Shu-Ya, WANG Chang-Hui. Effects of nitrogen addition at different levels on soil microorganisms in saline-alkaline grassland of northern China. Chinese Journal of Plant Ecology, 2021, 45(7): 780-789. DOI: 10.17521/cjpe.2021.0072
土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力。氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019)。氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012)。研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程。综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少。并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008)。因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究。
Table 1 表1 表1氮(N)水平、月份及其交互作用对盐渍化草地土壤微生物特征(氨氧化古菌和氨氧化细菌、细菌、真菌丰度和微生物生物量碳、微生物生物量氮含量)的影响 Table 1Effects of nitrogen (N) levels, month and their interactions on soil microorganism (AOA, AOB, bacteria, fungi abundance and MBC, MBN content) in saline-alkaline grassland
CaiJP, LuoWT, LiuHY, FengX, ZhangYY, WangRZ, XuZW, ZhangYG, JiangY (2017). Precipitation-mediated responses of soil acid buffering capacity to long-term nitrogen addition in a semi-arid grassland Atmospheric Environment, 170, 312-318. DOI:10.1016/j.atmosenv.2017.09.054URL [本文引用: 2]
ChengY, WangJ, MaryB, ZhangJB, CaiZC, ChangSX (2013). Soil pH has contrasting effects on gross and net nitrogen mineralizations in adjacent forest and grassland soils in central Alberta, Canada Soil Biology & Biochemistry, 57, 848-857. DOI:10.1016/j.soilbio.2012.08.021URL [本文引用: 1]
FiererN, JacksonRB (2006). The diversity and biogeography of soil bacterial communities Proceedings of the National Academy of Sciences of the United States of America, 103, 626-631. PMID:16407148 [本文引用: 1] For centuries, biologists have studied patterns of plant and animal diversity at continental scales. Until recently, similar studies were impossible for microorganisms, arguably the most diverse and abundant group of organisms on Earth. Here, we present a continental-scale description of soil bacterial communities and the environmental factors influencing their biodiversity. We collected 98 soil samples from across North and South America and used a ribosomal DNA-fingerprinting method to compare bacterial community composition and diversity quantitatively across sites. Bacterial diversity was unrelated to site temperature, latitude, and other variables that typically predict plant and animal diversity, and community composition was largely independent of geographic distance. The diversity and richness of soil bacterial communities differed by ecosystem type, and these differences could largely be explained by soil pH (r(2) = 0.70 and r(2) = 0.58, respectively; P < 0.0001 in both cases). Bacterial diversity was highest in neutral soils and lower in acidic soils, with soils from the Peruvian Amazon the most acidic and least diverse in our study. Our results suggest that microbial biogeography is controlled primarily by edaphic variables and differs fundamentally from the biogeography of "macro" organisms.
GruberN, GallowayJN (2008). An Earth-system perspective of the global nitrogen cycle Nature, 451, 293-296. DOI:10.1038/nature06592URL [本文引用: 1]
HuygheC (2010). New utilizations for the grassland areas and the forage plants: What matters Fourrages, 203, 213-219. [本文引用: 1]
KanakidouM, MyriokefalitakisS, DaskalakisN, FanourgakisG, NenesA, BakerAR, TsigaridisK, MihalopoulosN (2016). Past, present and future atmospheric nitrogen deposition Journal of the Atmospheric Sciences, 73, 2039-2047. DOI:10.1175/JAS-D-15-0278.1URL [本文引用: 1]
KuypersMMM, MarchantHK, KartalB (2018). The microbial nitrogen-cycling network Nature Reviews Microbiology, 16, 263-276. DOI:10.1038/nrmicro.2018.9PMID:29398704 [本文引用: 2] Nitrogen is an essential component of all living organisms and the main nutrient limiting life on our planet. By far, the largest inventory of freely accessible nitrogen is atmospheric dinitrogen, but most organisms rely on more bioavailable forms of nitrogen, such as ammonium and nitrate, for growth. The availability of these substrates depends on diverse nitrogen-transforming reactions that are carried out by complex networks of metabolically versatile microorganisms. In this Review, we summarize our current understanding of the microbial nitrogen-cycling network, including novel processes, their underlying biochemical pathways, the involved microorganisms, their environmental importance and industrial applications.
LauraRD (1976). Effects of alkali salts on carbon and nitrogen mineralization of organic matter in soil Plant and Soil, 44, 587-596. DOI:10.1007/BF00011378URL [本文引用: 1]
LiJG, PuLJ, HanMF, ZhuM, ZhangRS, XiangYZ (2014). Soil salinization research in China: advances and prospects Journal of Geographical Sciences, 24, 943-960. DOI:10.1007/s11442-014-1130-2URL [本文引用: 1]
LiY, XuXH, SunW, ShenY, RenTT, HuangJH, WangCH (2019). Effects of different forms and levels of N additions on soil potential net N mineralization rate in meadow steppe, Nei Mongol, China Chinese Journal of Plant Ecology, 43, 174-184. DOI:10.17521/cjpe.2018.0245URL [本文引用: 2]
LuM, YangYH, LuoYQ, FangCM, ZhouXH, ChenJK, YangX, LiB (2011). Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis New Phytologist, 189, 1040-1050. DOI:10.1111/nph.2011.189.issue-4URL [本文引用: 1]
LucasRW, KlaminderJ, FutterMN, BishopKH, EgnellG, LaudonH, HögbergP (2011). A meta-analysis of the effects of nitrogen additions on base cations: implications for plants, soils, and streams Forest Ecology and Management, 262, 95-104. DOI:10.1016/j.foreco.2011.03.018URL [本文引用: 2]
Martinez-BeltranJ, Licona-ManzurC (2005). Overview of salinity problems in the world and FAO strategies to address the problem//Proceedings of the International Salinity Forum. Riverside, California, USA.311-313. [本文引用: 1]
NicolGW, LeiningerS, SchleperC, ProsserJI (2008). The influence of soil pH on the diversity, abundance and transcriptional activity of ammonia oxidizing archaea and bacteria Environmental Microbiology, 10, 2966-2978. DOI:10.1111/emi.2008.10.issue-11URL [本文引用: 1]
NormanJS, BarrettJE (2014). Substrate and nutrient limitation of ammonia-oxidizing bacteria and archaea in temperate forest soil Soil Biology & Biochemistry, 69, 141-146. DOI:10.1016/j.soilbio.2013.11.003URL [本文引用: 1]
RathKM, FiererN, MurphyDV, RouskJ (2019). Linking bacterial community composition to soil salinity along environmental gradients The ISME Journal, 13, 836-846. DOI:10.1038/s41396-018-0313-8URL [本文引用: 3]
RathKM, RouskJ (2015). Salt effects on the soil microbial decomposer community and their role in organic carbon cycling: a review Soil Biology & Biochemistry, 81, 108-123. DOI:10.1016/j.soilbio.2014.11.001URL [本文引用: 4]
RouskJ, BååthE, BrookesPC, LauberCL, LozuponeC, CaporasoJG, KnightR, FiererN (2010). Soil bacterial and fungal communities across a pH gradient in an arable soil The ISME Journal, 4, 1340-1351. DOI:10.1038/ismej.2010.58URL [本文引用: 1]
TianDS, NiuSL (2015). A global analysis of soil acidification caused by nitrogen addition Environmental Research Letters, 10, 024019. DOI: 10.1088/1748-9326/10/2/024019. URL [本文引用: 5]
WangC, Butterbach-BahlK, HeN, WangQ, XingX, HanX (2015). Nitrogen addition and mowing affect microbial nitrogen transformations in a C4 grassland in Northern China European Journal of Soil Science, 66, 485-495. DOI:10.1111/ejss.2015.66.issue-3URL [本文引用: 1]
WeiL, SuJS, JingGH, ZhaoJ, LiuJ, ChengJM, JinJW (2018). Nitrogen addition decreased soil respiration and its components in a long-term fenced grassland on the Loess Plateau Journal of Arid Environments, 152, 37-44. DOI:10.1016/j.jaridenv.2018.01.017URL [本文引用: 1]
XuXH, DiaoHJ, TanCY, HaoJ, ShenY, DongKH, WangCH (2021). Response of soil net nitrogen mineralization to different levels of nitrogen addition in a saline-alkaline grassland of northern China Chinese Journal of Plant Ecology, 45, 85-95. DOI:10.17521/cjpe.2020.0153URL [本文引用: 2]
YangC, WangXZ, MiaoFH, LiZY, TangW, SunJ (2020). Assessing the effect of soil salinization on soil microbial respiration and diversities under incubation conditions Applied Soil Ecology, 155, 103671. DOI: 10.1016/j.apsoil.2020.103671. URL [本文引用: 3]
YangJS, ZhanC, LiYZ, ZhouD, YuY, YuJB (2018). Effect of salinity on soil respiration in relation to dissolved organic carbon and microbial characteristics of a wetland in the Liaohe River estuary, Northeast China Science of the Total Environment, 642, 946-953. DOI:10.1016/j.scitotenv.2018.06.121URL [本文引用: 1]
YingJY, LiXX, WangNN, LanZC, HeJZ, BaiYF (2017). Contrasting effects of nitrogen forms and soil pH on ammonia oxidizing microorganisms and their responses to long-term nitrogen fertilization in a typical steppe ecosystem Soil Biology & Biochemistry, 107, 10-18. DOI:10.1016/j.soilbio.2016.12.023URL [本文引用: 6]
YuanBC, LiZZ, LiuH, GaoM, ZhangYY (2007). Microbial biomass and activity in salt affected soils under arid conditions Applied Soil Ecology, 35, 319-328. DOI:10.1016/j.apsoil.2006.07.004URL [本文引用: 3]
ZhangL, HuangJH, BaiYF, HanXG (2009). Effect of nitrogen addition on net nitrogen mineralization in Leymus chinensis grassland, Inner Mongolia, China Chinese Journal of Plant Ecology, 33, 563-569. [本文引用: 1]
Chronic nitrogen addition differentially affects gross nitrogen transformations in alpine and temperate grassland soils 1 2020
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Current insights into the autotrophic thaumarchaeal ammonia oxidation in acidic soils 1 2012
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
氨氧化微生物生态学与氮循环研究进展 1 2009
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
氨氧化微生物生态学与氮循环研究进展 1 2009
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
New utilizations for the grassland areas and the forage plants: What matters 1 2010
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Effects of alkali salts on carbon and nitrogen mineralization of organic matter in soil 1 1976
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Soil salinization research in China: advances and prospects 1 2014
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis 1 2011
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
A meta-analysis of the effects of nitrogen additions on base cations: implications for plants, soils, and streams 2 2011
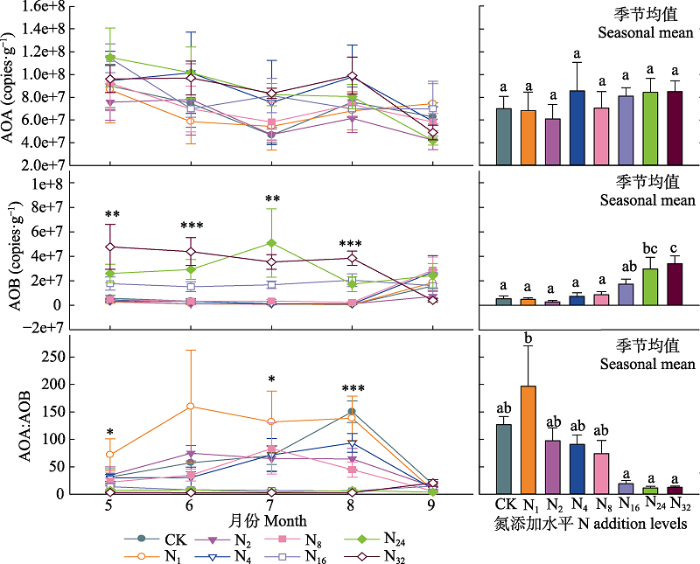
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
Nutrient addition reduces carbon sequestration in a Tibetan grassland soil: disentangling microbial and physical controls 2 2020
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
... ; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
... ).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
Soil bacterial and fungal communities across a pH gradient in an arable soil 1 2010
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
A global analysis of soil acidification caused by nitrogen addition 5 2015
Nitrogen addition and mowing affect microbial nitrogen transformations in a C4 grassland in Northern China 1 2015
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Nitrogen addition decreased soil respiration and its components in a long-term fenced grassland on the Loess Plateau 1 2018
Assessing the effect of soil salinization on soil microbial respiration and diversities under incubation conditions 3 2020
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
... ., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
... ., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
Effect of salinity on soil respiration in relation to dissolved organic carbon and microbial characteristics of a wetland in the Liaohe River estuary, Northeast China 1 2018
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
Contrasting effects of nitrogen forms and soil pH on ammonia oxidizing microorganisms and their responses to long-term nitrogen fertilization in a typical steppe ecosystem 6 2017
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
... ., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
... ., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
氮素添加对内蒙古羊草草原净氮矿化的影响 1 2009
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
氮素添加对内蒙古羊草草原净氮矿化的影响 1 2009
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Soil pH is equally important as salinity in shaping bacterial communities in saline soils under halophytic vegetation 1 2018
... 本研究结果表明盐渍化草地土壤微生物对氮输入的响应不敏感(细菌和真菌丰度及微生物生物量碳、氮含量变化不显著), 这与草地较高的盐渍化程度有关.首先, 较高的盐渍化会抑制土壤微生物活性, 大量的研究表明随着盐渍化程度的升高, 土壤微生物生物量降低(Yuan et al., 2007; Rath & Rousk, 2015), 土壤微生物活性降低(Yuan et al., 2007; Rath & Rousk, 2015; Yang et al., 2018, 2020).盐渍化程度不同的土壤中土壤微生物群落结构与组成存在较大差异(Yang et al., 2020).在我们的实验地点发现土壤阳离子浓度和土壤pH是调控土壤微生物的关键因子, 这与已有的研究结果(Rath & Rousk, 2015; Yang et al., 2020)一致.氮添加会引起土壤的酸化(Cai et al., 2017), 这是由于氮的输入增加了土壤的硝化作用, 使得氮转化过程中H+释放增加, 同时硝酸盐淋溶损失增加, 引起土壤酸性增加(Lucas et al., 2011; Cai et al., 2017), 影响土壤微生物的群落和组成(Rousk et al., 2010; Zhao et al., 2018).土壤具有一定的酸缓冲能力, 一方面与土壤自身的pH高低有关, 研究表明, 较高的土壤pH往往具有相对较强的抵抗力(Bowman et al., 2008; Rath et al., 2019).因此在本试验中, 尽管随着氮输入水平的提高土壤pH不断降低, 但不同施氮水平下土壤pH差异并不显著, 氮添加诱导的酸化作用较弱.另一方面土壤交换性阳离子对土壤的酸化起到主要的酸缓冲作用(Bowman et al., 2008; Tian & Niu, 2015), 盐渍化草地较高的土壤交换性阳离子能够阻止氮输入引起的pH降低, 减弱氮输入诱导的pH改变对土壤微生物的影响, 从而调控氮添加对微生物的影响(Bowmanet al., 2008; Lucas et al., 2011; Tian & Niu, 2015).因此, 盐渍化草地土壤pH和土壤阳离子浓度共同调控着土壤微生物特征, 而相对较低的土壤pH和土壤阳离子浓度的改变使得土壤微生物特征对氮输入的响应不敏感. ...
Modeling patterns of nonlinearity in ecosystem responses to temperature, CO2, and precipitation changes 1 2008
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
Patterns and mechanisms of responses by soil microbial communities to nitrogen addition 1 2017
... 土壤微生物是调控土壤氮转化的主要生物因子(Kuypers et al., 2018), 影响氮有效性的同时改变了生态系统生产力.氮沉降增加会改变土壤微生物特征, 影响土壤的氮转化过程(Laura, 1976; Lu et al., 2011; Ying et al., 2017), 但前人的研究结果(张璐等, 2009; Wang et al., 2015; Hao et al., 2020)并不一致, 可能与氮的添加量和形态有关(李阳等, 2019).氨氧化过程是硝化作用的第一步, 在自然生态系统中, 这是由氨氧化微生物(氨氧化细菌和氨氧化古菌)驱动的(贺纪正和张丽梅, 2009; He et al., 2012).研究表明氮的输入会影响到氨氧化微生物数量(Ying et al., 2017), 土壤细菌和真菌的组成(Luo et al., 2020)以及土壤微生物生物量(Zhou et al., 2017), 进而影响到土壤氮的转化过程.综合考虑氨氧化微生物(氨氧化古菌和氨氧化细菌), 土壤细菌和真菌的丰度, 以及土壤微生物生物量对不同氮水平添加的响应的研究较少.并且随着氮输入水平的提高, 生态系统功能和过程往往会呈现出非线性的响应(Zhou et al., 2008).因此不同氮添加水平是如何影响土壤微生物特征的仍需要深入研究. ...
 ,2,3,4,*
,2,3,4,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT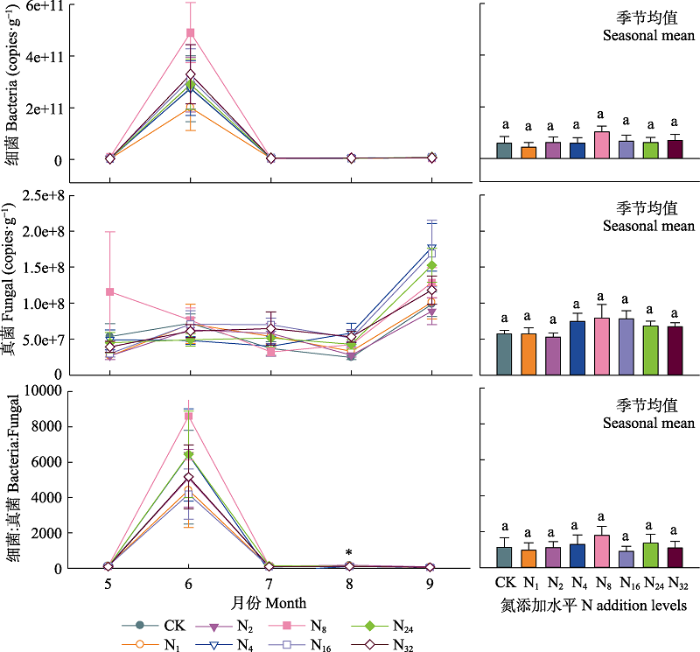 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT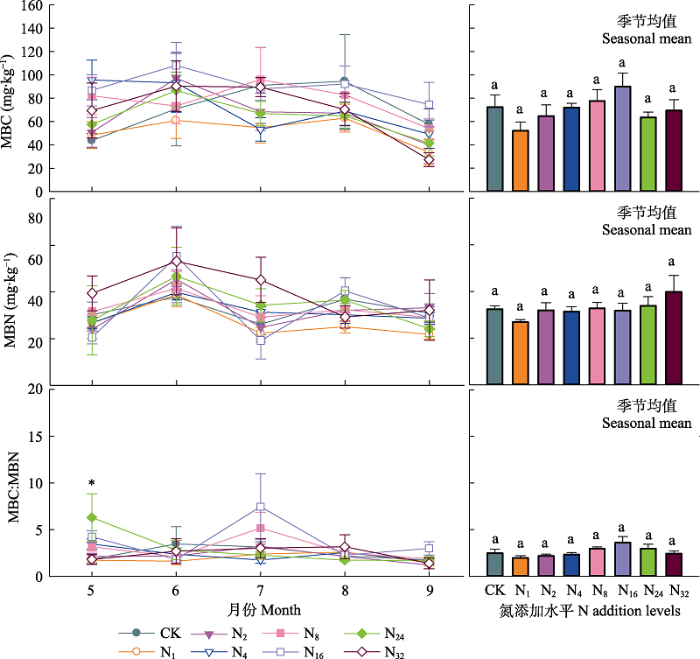 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT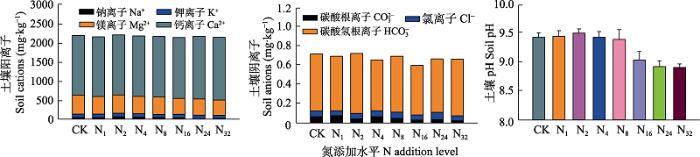 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT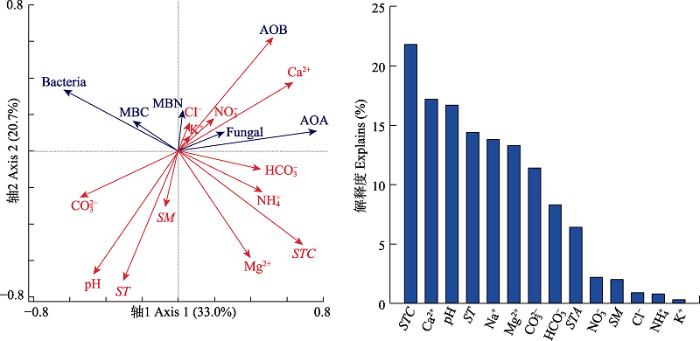 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}