 ,
, ,1,2,*
,1,2,*Responses of soil nitrogen in different soil organic matter fractions to long-term nitrogen addition in a semi-arid grassland
Yun-Tao WU1,2, Sen YANG1,2, Xin WANG1, Jun-Sheng HUANG1, Bin WANG1,2, Wei-Xing LIU1, Ling-Li LIU,,1,2,*通讯作者: * 刘玲莉: ORCID: 0000-0002-5696-3151,lingli.liu@ibcas.ac.cn
编委: 白娥
责任编辑: 赵航
收稿日期:2021-02-3接受日期:2021-03-30
| 基金资助: |
Corresponding authors: *lingli.liu@ibcas.ac.cn
Received:2021-02-3Accepted:2021-03-30
| Fund supported: |

摘要
土壤氮库对生态系统的养分循环至关重要。目前多数研究主要关注氮沉降对土壤总氮的影响, 而对土壤不同有机质组分的氮库对氮沉降响应的研究较为缺乏。该研究基于内蒙古典型草地的长期多水平施氮(0、8、32、64 g·m-2·a-1)实验平台, 利用土壤密度分级方法, 探究氮添加处理13年后典型草地中两种土壤有机质组分(颗粒态有机质(POM), 矿质结合态有机质(MAOM))氮含量的变化及调控机制。结果显示: 土壤总碳含量、POM和MAOM的碳含量在施氮处理间均没有显著差异。土壤总氮含量则随着施氮水平增加呈显著增加的趋势, 同时施氮处理下POM的氮含量显著上升, 而MAOM的氮含量没有变化。进一步分析发现, 施氮促进植物地上生物量积累, 增加了凋落物量及其氮含量, 从而导致POM的氮含量增加。由于MAOM主要通过黏土矿物等吸附土壤中小分子有机质形成, 其氮含量受土壤中黏粒与粉粒含量影响, 而与氮添加水平无显著相关关系。该研究结果表明长期氮添加促进土壤氮库积累, 但增加的氮主要分布在稳定性较低的POM中, 受干扰后容易从生态系统中流失。为了更准确地评估和预测氮沉降对陆地生态系统的氮循环过程的影响, 应考虑土壤中不同有机质组分的差异响应。
关键词:
Abstract
Aims Soil nitrogen (N) plays a vital role in regulating the structure and function of ecosystems and is affected by N deposition. Most previous studies focus on the responses of the N content in bulk soil to N deposition, but the responses of the N content in different soil organic matter (SOM) fractions remain unclear. We aimed to investigate how long-term N addition influenced soil N of different SOM fractions in a semi-arid grassland. Methods A manipulated N addition experiment with 4 levels of N addition (0, 8, 32 and 64 g·m-2·a-1) has been conducted for 13 years in Duolun country, Nei Mongol. SOM was separated to particulate organic matter (POM) and mineral associated organic matter (MAOM) by density fractionation. The plant and soil properties were also measured. Important findings The results showed that N addition had no significant effect on the carbon (C) content in bulk soil, POM, or MAOM. With increasing levels of N addition, the N content in bulk soil and in POM increased significantly. Furthermore, we found that the increased N content of POM was mainly associated with greater aboveground biomass following N addition. The N content of MAOM is mainly correlated with soil texture, but was not affected by N addition. These results suggest that continuous N addition can increase the soil N in bulk soil, but the increased N is mostly distributed in labile POM pools, which can be vulnerable to land use and climate change.
Keywords:
PDF (1302KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
武运涛, 杨森, 王欣, 黄俊胜, 王斌, 刘卫星, 刘玲莉. 草地土壤有机质不同组分氮库对长期氮添加的响应. 植物生态学报, 2021, 45(7): 790-798. DOI: 10.17521/cjpe.2021.0044
WU Yun-Tao, YANG Sen, WANG Xin, HUANG Jun-Sheng, WANG Bin, LIU Wei-Xing, LIU Ling-Li.

氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al., 2008)。环境保护政策和经济结构调整使得中国氮沉降速率在2000年后趋于稳定, 但中国依然是全球大气氮沉降量最高的一个区域(Yu et al., 2019)。当大气氮沉降进入到生态系统后, 约25%的氮被储存在土壤有机质中(Niu et al., 2016)。土壤氮含量的增加一方面缓解了生态系统氮限制, 促进植物生长, 另一方面也导致了土壤酸化, 抑制微生物分解活动(Wei et al., 2013; Yang et al., 2019)。土壤有机质是由化学组成、形成机制与稳定性皆存在差异的多个组分构成(Christensen, 2001; Schmidt et al., 2011)。土壤有机质不同组分的氮含量及氮矿化速率存在较大差异, 对外源氮的固持能力亦不同(Six et al., 2002)。目前多数研究主要关注氮沉降对土壤总氮的影响(Lu et al., 2011), 而对土壤有机质不同组分的氮含量对氮沉降响应的研究仍非常缺乏。定量评估大气沉降的氮在不同土壤有机质组分中的分配, 对准确判断陆地生态系统对大气氮沉降的固持能力有重要意义。
土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al., 2007)。大多数研究认为POM是密度小于1.6-1.85 g·cm-3和(或)粒径大于50-63 μm的有机质组分, MAOM是密度大于1.6-1.85 g·cm-3和(或)粒径小于50-63 μm的有机质组分(Lavallee et al., 2020)。POM主要来源于半分解的植物残体, 因其在土壤中通常以游离态形式存在, 使POM的氮容易被微生物代谢利用(Meyer et al., 2012; Francesca Cotrufo et al., 2015)。而MAOM主要来源于微生物残体与代谢产物等(Mikutta & Kaiser, 2011)。虽然MAOM的碳氮比低于POM, 但是由于土壤矿物的物理和化学保护, MAOM中的氮不容易被植物和异养微生物利用(刘满强等, 2007; Dungait et al., 2012; Lavallee et al., 2020)。
大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al., 2015)。氮沉降模拟实验表明, 氮添加通过提高草地生态系统生产力和改变群落的物种组成, 增加了凋落物的输入量及其氮含量(Liu & Greaver, 2010; Yang et al., 2019)。凋落物输入量的上升增加了分解过程中的底物数量, 促进土壤中POM形成(Lavallee et al., 2020), 所以长期氮添加可能会增加POM的氮含量。氮添加导致的土壤酸化会抑制土壤微生物代谢作用(Liu et al., 2018), 减缓凋落物等向MAOM转化。从而, 氮添加可能降低MAOM的含量。但是氮添加显著降低了土壤中真菌与细菌的比例(Liu et al., 2018), 由于真菌的碳氮比高于细菌, 所以氮添加促进了土壤中低碳氮比的微生物残体等数量的积累, 可能增加土壤中MAOM的氮含量。此外, 土壤中MAOM的含量与土壤黏土矿物含量(主要是黏粒和粉粒含量)呈显著正相关关系(Hassink, 1997)。一般认为在未受严重干扰的自然生态系统中, 土壤质地变化不显著, 因此通过矿物等吸附形成的有机质(MAOM)含量是相对稳定的(Castellano et al., 2015)。所以, 长期氮添加导致的微生物过程改变是否会影响土壤中MAOM的氮含量尚不清楚。
为探讨氮沉降增加对土壤中POM和MAOM氮含量的影响, 本研究基于内蒙古多伦县典型草地生态系统的长期多水平氮添加实验平台, 通过土壤密度分级的方法, 研究长期氮添加对典型草地中不同土壤有机质组分氮含量的影响。同时结合对植物、土壤和微生物过程的监测, 解析POM和MAOM氮含量响应氮沉降的机制。本研究的主要假设为: (1)氮添加促进了地上生物量的积累, 可能通过增加凋落物的输入量, 提高土壤中POM的氮含量; (2)氮添加导致了土壤酸化, 可能通过抑制微生物的代谢作用, 减少土壤中MAOM的氮含量。
1 材料和方法
1.1 研究区域与实验设计
本研究位于内蒙古多伦县典型草地生态系统(42.03° N, 116.28° E; 海拔1 324 m)。该区域年平均气温为2.1 ℃, 年降水量为382.2 mm, 降水集中在植被生长季(5-9月)。自然条件下大气氮沉降速率约为1.6 g·m-2·a-1 (Zhang et al., 2008)。根据联合国粮农组织(FAO)分类标准, 该区域的土壤类型为栗钙土, 土壤的平均密度为1.3 g·cm-3, 平均pH为6.8。该区域的植被以多年生物种为主, 优势种包括西北针茅(Stipa Sareptana var. krylovii)、冰草(Agropyron cristatum)、冷蒿(Artemisia frigida)、糙隐子草(Cleistogenes squarrosa)等。2003年开始氮添加实验。实验样地包含64个样方(面积为10 m × 15 m, 缓冲区距离为5 m), 按照拉丁方实验设计划分为8行8列。每年的7月对实验样方进行8个水平的施氮处理(0、1、2、4、8、16、32、64 g·m-2·a-1)。从2005年开始, 每年的8月末对奇数行样方通过刈割来模拟放牧。本研究只考虑氮添加的效应, 因此生物量测定及土壤取样分析等在偶数行(非刈割处理)样方进行。综合考虑分析成本、工作量和氮添加处理梯度范围, 本研究选取了其中4个氮添加处理(0、8、32、64 g·m-2·a-1)进行实验和分析。
1.2 取样方法
植物地上生物量(AGB)在2016年8月中旬测定。在每个样方中随机放置一个1 m × 1 m的样方框, 按物种刈割样方框内的所有地上植被, 并在烘箱内65 ℃条件下烘至恒质量。将样方框内的所有植物样品称质量并加和来表征该实验样方区的AGB。土壤样品在2016年8月中旬收集。在每个样方内用直径5 cm的土钻随机取3钻表层土壤(0-15 cm), 混匀为一个土壤样品。利用2 mm的网筛去除土壤样品中较大的根和石头, 冷藏保存并带回实验室。一部分样品被冷冻保存, 用于测定微生物生物量碳氮含量和土壤溶解性无机氮(DIN)含量; 一部分样品自然风干后用于测定土壤的理化性质, 如碳氮含量、机械组成、土壤 pH、有机质组分等。微生物生物量碳、氮含量采用氯仿熏蒸法测定(Beck et al., 1997), 即在25 ℃下将经过氯仿熏蒸24 h的土壤和对照处理的土壤, 用50 mL 0.5 mol·L-1 K2SO4浸提, 利用总有机碳分析仪(TOC-3000, Analytik JenaAG, Jena, Germany)测定两份浸提液中溶解性碳、氮的浓度, 熏蒸和未熏蒸样品的浓度差除以转化系数(KC = 0.38, KN = 0.45)后, 得到土壤中微生物生物量碳、氮含量(Brookes et al., 1985; Vance et al., 1987)。土壤DIN (铵态氮和硝态氮)含量利用50 mL 2 mol·L-1 KCl溶液浸提, 通过全自动流动分析仪(SAN-System, Breda, Skalar, the Netherlands)测定土壤浸提液和空白标准液中铵态氮和硝态氮浓度, 通过标准曲线计算土壤中DIN含量。一部分土壤风干样品, 利用激光粒度分析仪(Malvern Mastersizer 2000, Malvern, Worcestershire, UK)测定土壤质地(土壤质地按照国际制(ISSS Working Group RB)土壤质地分类依据可以分为黏粒(<0.002 mm), 粉粒(0.002-0.02 mm)以及砂粒(0.02-2 mm)); 利用pH电位计按照土壤和水的重量体积比(W/V) = 1:2.5的比例测定土壤pH。另一部分风干样品经过粉碎后, 称取20 mg粉碎后样品, 利用元素分析仪(CNS Element Analysensysteme, GmbH, Hanau, German)测定样品的碳氮含量。
为了探究不同有机质组分的质量及其氮含量的变化, 本研究通过土壤密度分级的方法将土壤有机质分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)。即: 称取30 g自然风干的土壤样品于250 mL的离心瓶中, 加入150 mL密度为1.8 g·cm-3的NaI溶液, 利用220 r·min-1的摇床振荡3 h, 水平静置24 h后将上层悬浮的自由态POM转移至0.22 μm孔径的滤纸上。剩余沉淀液用150 J·mL-1的能量超声处理, 释放被土壤团聚体包裹的POM, 沉淀液经过高速离心(4 250 r·min-1) 10 min后, 将上层悬浮的包裹态POM转移至0.22 μm孔径的滤纸上。两部分的POM用蒸馏水冲洗至溶液的电导率<2 μs, 重复上述所有操作3次。将剩余的沉淀液高速离心(4 250 r·min-1) 30 min后, 转移至0.22 μm孔径的滤纸上, 用蒸馏水冲洗至溶液的电导率<50 μs, 得到的组分即为MAOM。将POM和MAOM置于50 ℃烘箱中烘至恒质量, 粉碎后利用元素分析仪测定每个组分的碳氮含量。相关方法参见Wiesmeier等(2015)。
1.3 统计分析
研究利用单因素方差分析和最小显著性差异(LSD)法检验氮添加对土壤碳氮含量、不同有机质组分的碳氮含量、土壤质地、土壤 pH、植物AGB和微生物生物量的影响。在统计分析中, 我们参考经典的统计方法, 仅在方差分析显著的情况下对不同处理水平间进行了多重比较, 以减小统计结果中一类错误(即原假设H0符合实际情况, 检验结果却将其否定)发生的概率(Forthofer & Lee, 1996)。同时利用简单线性回归来估计两种组分的碳、氮含量对两个变量(氮添加水平、黏粒和粉粒的含量)的变化响应。采用基于step-AIC的模型筛选、广义线性混合模型(GLMM)和结构方程模型(SEM)的方法来探讨氮添加处理和土壤质地对两种有机质组分氮含量影响的可能机制。其中在模型筛选中, 响应变量为POM和MAOM的氮含量, 预测变量为AGB、植物碳氮比、微生物生物量碳含量、微生物异养呼吸速率、土壤质地相关参数。GLMM中固定效应为氮添加水平和土壤质地, 随机效应为样品的重复数。所有的数据利用Shapiro-Wilk test和Leveneʼs test进行了正态性和方差齐性的检验。数据分析在R 3.5.4软件中进行。统计结果仅在p < 0.05时认为存在统计显著性。2 结果和分析
2.1 长期氮添加对植物、土壤和微生物的影响
植物、土壤和微生物对长期氮添加的响应不同(图1)。随施氮水平的增加, 植物AGB显著增加。施氮处理显著增加了土壤中的DIN含量, 但显著降低了土壤pH及微生物生物量碳(MBC)含量。此外, 长期氮添加背景下, 土壤质地在不同施氮水平样地之间没有显著的差异(图2)。图1
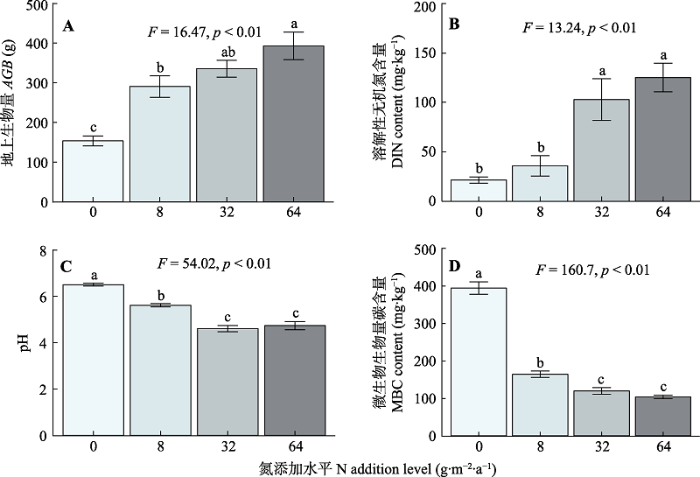 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1氮添加对地上生物量(A), 溶解性无机氮含量(B), 土壤pH (C)和微生物生物量碳含量(D)的影响(平均值±标准误)。F和p为方差分析的结果, 不同小写字母表示不同氮添加水平间存在显著差异(p < 0.05)。
Fig. 1Effects of nitrogen (N) addition on aboveground biomass (AGB)(A), dissolved inorganic nitrogen (DIN) content (B), soil pH (C) and microbial biomass carbon (MBC) content (D)(mean ± SE). F and p are the results of one-way ANOVA. Different lowercase letters indicate significant difference among N addition levels (p < 0.05).
图2
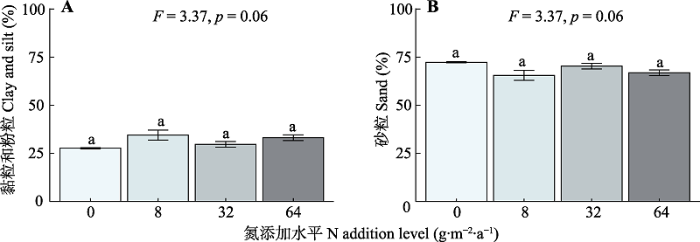 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2氮添加对土壤黏粒与粉粒含量(A)和砂粒含量(B)的影响(平均值±标准误)。F和p为方差分析的结果, 不同小写字母表示不同氮添加水平间存在显著差异(p < 0.05)。
Fig. 2Effects of nitrogen (N) addition on soil clay and silt (A) and sand (B) content (mean ± SE). F and p are the results of one-way ANOVA. Different lowercase letters indicate significant difference (p < 0.05) among different N addition levels.
2.2 长期氮添加对土壤有机质组分的影响
与对照相比, 氮添加不改变土壤总碳含量, 但显著提高土壤总氮含量; 同时土壤碳氮比沿施氮梯度无显著变化(图3)。土壤有机质中POM的含量约为6.2%, MAOM含量为93.8%, 施氮处理并未显著改变土壤中POM和MAOM的相对含量(图4A)。此外, 随着氮添加量的增加, 土壤中POM和MAOM的碳含量均无显著变化(图4B)。POM的氮含量沿施氮梯度显著增加, 但MAOM的氮含量在施氮处理间无显著变化(图4C)。图3
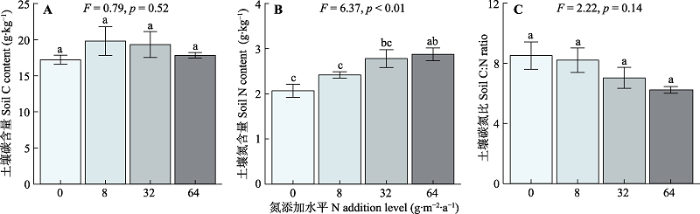 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3氮添加对土壤碳含量(A)、氮含量(B)和碳氮比(C)的影响(平均值±标准误)。F和p为方差分析的结果, 不同小写字母表示不同氮添加水平间存在显著差异(p < 0.05)。
Fig. 3Effects of nitrogen (N) addition on soil carbon (C) content (A), soil N content (B) and soil C:N (C)(mean ± SE). F and p are the results of one-way ANOVA. Different lowercase letters indicate significant difference (p < 0.05) among different N addition levels.
图4
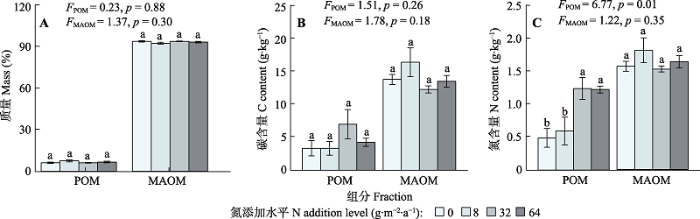 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4氮添加对土壤中颗粒态有机质(POM)和矿质结合态有机质(MAOM)的相对含量(A)、碳含量(B)和氮含量(C)的影响(平均值±标准误)。F和p为方差分析的结果, 不同字母表示不同氮添加水平间存在显著差异(p < 0.05)。
Fig. 4Effect of nitrogen (N) addition on the relative mass (A), carbon (C) content (B) and N content (C) of particulate organic matter (POM) and mineral associated organic matter (MAOM)(mean ± SE). F and p are the results of one-way ANOVA. Different lowercase letters indicate significant difference (p < 0.05) among different N addition levels.
2.3 土壤有机质组分变化的影响因素
POM和MAOM的碳含量与氮添加量无显著线性相关关系(图5A)。然而, POM的氮含量与氮添加量之间呈显著正相关关系(R2 = 0.49, p < 0.01, 图5C), 而MAOM的氮含量与氮添加量无显著线性相关关系(p = 0.75, 图5C)。此外, POM的碳、氮含量与土壤黏粒和粉粒含量均无明显相关关系(碳: p = 0.69, 图5B; 氮: p = 0.75, 图5D)。同时, MAOM的碳含量与土壤黏粒和粉粒含量也无显著相关关系(p = 0.06, 图5B); 但是, MAOM的氮含量与土壤黏粒和粉粒含量呈显著线性正相关关系(R2 = 0.23, p = 0.03, 图5D)。GLMM的结果表明: 氮添加和土壤质地(黏粒和粉粒)对POM的碳含量均无显著作用(表1), 而MAOM的碳含量的变化与氮添加和土壤质地显著相关(氮添加: p < 0.05; 土壤质地: p = 0.01)。POM的氮含量主要受到氮添加处理的影响(p < 0.01, 表1), 土壤质地的影响不显著; 而MAOM的氮含量主要受土壤质地的影响(p < 0.01), 氮添加的作用不显著。图5
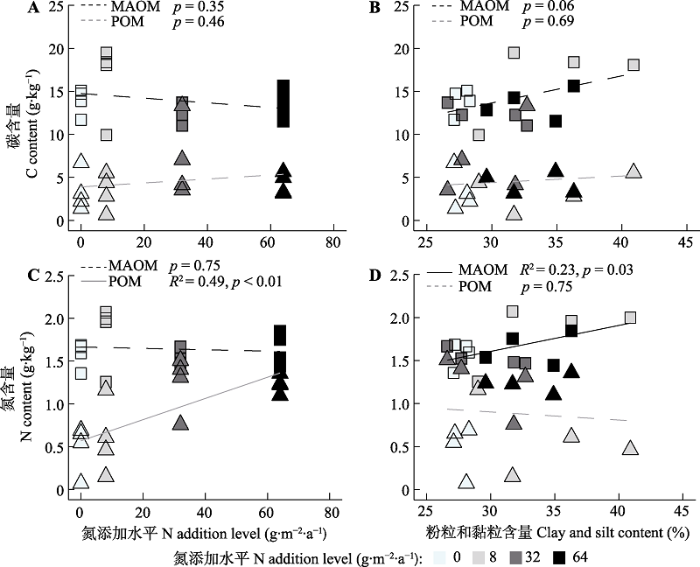 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5土壤中不同有机质组分的碳氮含量与氮添加水平(A, C)和土壤黏粒与粉粒含量(B, D)的关系。实线表示相关性显著(p < 0.05), 虚线表示相关性不显著。□, 矿质结合态有机质(MAOM); △, 颗粒态有机质(POM)。
Fig. 5Relationships between the soil carbon (C) in different SOM fractions and nitrogen (N) addition rates (A), and soil clay and silt content (B); and relationships between the soil N in different SOM fractions and N addition rates (C), and soil clay and silt content (D). Solid and dashed lines represent significant (p < 0.05) and insignificant (p > 0.05) relationships, respectively. □, mineral associated organic matter (MAOM); △, particulate organic matter (POM).
Table 1
表1
表1氮添加和土壤粉粒与黏粒含量对土壤颗粒态有机质(POM)和矿质结合态有机质(MAOM)碳、氮含量影响的广义线性混合模型(GLMM)分析结果
Table 1
| 响应变量 Response variable | 变量 Variable | 相关系数 Correlation coefficient | 标准误 Standard error | t | p |
|---|---|---|---|---|---|
| POM碳含量 C content in POM | 氮添加水平 N addition (g·m-2·a-1) | 0.022 | 0.028 | 0.793 | 0.446 |
| 黏粒与粉粒含量 Silt and sand clay content (%) | 0.013 | 0.187 | -0.068 | 0.947 | |
| MAOM碳含量 C content in MAOM | 氮添加水平 N addition (g·m-2·a-1) | -0.043 | 0.019 | -2.287 | 0.045 |
| 黏粒与粉粒含量 Silt and sand clay content (%) | 0.396 | 0.126 | 3.145 | 0.010 | |
| POM氮含量 N content in POM | 氮添加水平 N addition (g·m-2·a-1) | 0.014 | 0.003 | 4.045 | 0.002 |
| 黏粒与粉粒含量 Silt and sand clay content (%) | -0.030 | 0.021 | -2.086 | 0.182 | |
| MAOM氮含量 N content in MAOM | 氮添加水平 N addition (g·m-2·a-1) | -0.002 | 0.001 | -1.408 | 0.162 |
| 黏粒与粉粒含量 Silt and sand clay content (%) | 0.035 | 0.010 | 3.586 | 0.005 |
新窗口打开|下载CSV
模型筛选的结果表明, POM的氮含量变化主要受到地上生物量的影响, 而凋落物质量, 微生物等没有显著的影响; MAOM的氮含量变化主要受到土壤质地的显著影响, 而微生物等的作用不显著(表2)。SEM的结果也表明, 氮添加主要通过增加AGB (p < 0.01), 增加了POM的氮含量(p < 0.01, 图6A)。氮添加降低了MBC含量, 但MBC对MAOM的氮含量的影响不显著。土壤质地显著影响了土壤中MAOM的氮含量(p < 0.01, 图6B), 土壤中黏粒和粉粒含量越高, MAOM中的氮含量越高。
Table 2
表2
表2颗粒态有机质(POM)和矿质结合态有机质(MAOM)的氮(N)含量影响因子的基于赤池信息推测的最优模型筛选
Table 2
| 响应变量 Response variable | 因子 Factor | 模型Model | |||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||
| POM氮含量 N content in POM | 常数项 Constant term | 1.770 | 1.697 | 0.090 | - |
| 地上生物量 Aboveground biomass | 0.003 | 0.002 | 0.003 | - | |
| 微生物生物量碳含量 Microbial biomass carbon content | 0.002 | - | - | - | |
| C:N | -1.111 | -0.067 | - | - | |
| 赤池信息准则 Akaike information criterion | 24.90 | 21.95 | 17.66 | - | |
| MAOM氮含量 N content in MAOM | 常数项 Constant term | 1.202 | 1.451 | 0.762 | 0.702 |
| 黏粒与粉粒含量 Silt and clay content | 0.031 | 0.029 | 0.036 | 0.030 | |
| 微生物生物量碳含量 Microbial biomass carbon content | -0.002 | -0.001 | - | - | |
| 异养呼吸速率 Heterotrophic respiration rate | 0.375 | - | - | - | |
| 地上生物量 Aboveground biomass | -0.002 | -0.002 | -0.001 | - | |
| 赤池信息准则 Akaike information criterion | 5.92 | 5.17 | 3.01 | 2.30 | |
新窗口打开|下载CSV
图6
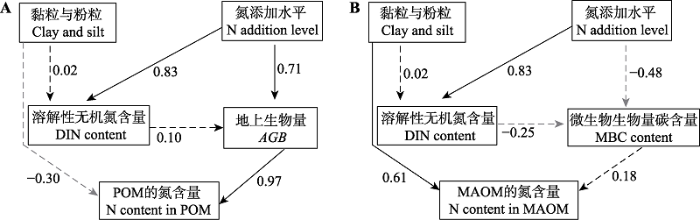 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图6氮添加对颗粒态有机质(POM)(A)和矿质结合态有机质(MAOM)(B)的氮含量影响途径的结构方程模型(SEM)分析结果。模型拟合的结果: POMN, χ2 = 0.480, p = 0.787, 相对拟合指数(CFI) = 1.000, 近似误差平方根(RMSEA) < 0.001; MAOMN, X2 = 2.856, p = 0.414, CFI = 1.000, RMSER < 0.001。实线箭头代表显著的效应(p < 0.05), 虚线箭头代表没有显著的效应(p > 0.05), 箭头附近的数字代表标准化的路径系数。
Fig. 6A schematic representation of the structural equation modeling (SEM) analysis used to identify the controls of soil nitrogen (N) content in particulate organic matter (POM)(A) and mineral associated organic matter (MAOM)(B) under N addition. Results of the model fitting were: POMN, X2 = 0.480, p = 0.787, comparative fit index (CFI) = 1.000, root-mean-square (RMSEA) < 0.001; MAOMN, X2 = 2.856, p = 0.414, CFI = 1.000, RMSEA < 0.001. Solid and dashed arrows represent significant effect (p < 0.05) and insignificant effect (p > 0.05), respectively. Number adjacent to the arrows refer to the standardized path coefficients. AGB, aboveground biomass; DIN, dissolved inorganic nitrogen; MBC, microbial biomass carbon.
3 讨论
土壤氮库是植物和微生物代谢养分的重要来源(Kuzyakov & Xu, 2013)。大气氮沉降对土壤中不同稳定性有机质组分的氮含量的影响, 可能会影响土壤氮库的储量及其稳定性。本研究结果显示, 随着氮添加水平的增加, 土壤总碳含量没有变化, 但土壤总氮含量明显增加(图3)。氮添加导致的土壤氮增加主要发生在POM组分中, 而MAOM氮含量在氮添加后没有明显变化(图4)。大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al., 2019)。长期氮添加通过缓解植物的养分限制, 提高了植物地上生物量及凋落物量(图1A)(Näsholm et al., 2009; 刁励玮等, 2018)。所以, 在凋落物分解过程中, 通过物理迁移过程形成的POM的氮含量沿施氮梯度显著增加(图4C)。同时POM的氮含量变化主要与POM的形成过程相关, 会受到凋落物输入量、凋落物质量、微生物等因素的共同影响。本研究通过模型筛选分析发现, 只有地上生物量显著影响POM的氮含量, 而凋落物质量、微生物等其他因素的作用不显著(表2)。此外, 结构方程模型的结果表明, 在进一步将POM对无机氮可能存在的物理吸附作用考虑在内的情况下, 氮添加导致的地上生物量增加是土壤中POM的氮含量增加的最主要途径, 这与我们的假设(1)一致(图6)。此外, 前期的研究表明, 长期氮添加使得此样地中低碳氮比的物种逐渐成为优势种, 增加了凋落物的氮含量(Yang et al., 2019), 从而可能进一步增加POM的氮含量。与POM氮含量的变化不同, POM的碳含量沿施氮梯度没有显著变化(图4B)。可能是因为土壤的碳库较大(Pribyl, 2010), 所以施氮增加的地上生物量, 没有显著增加POM的碳含量。
大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al., 2015; Mikutta et al., 2019)。长期氮添加导致土壤酸化(图1C), 抑制了微生物的活性(图1D), 可能通过减少形成MAOM的底物来源, 降低MAOM的含量(Frey et al., 2014; Zhang et al., 2018)。但是, 本研究的结果显示, MAOM的碳、氮含量沿施氮梯度均没有发生显著变化(图4), 这与我们的假设(2)不符。我们之前在本研究站点的工作发现, 施氮后群落水平凋落物可降解性的下降和植物多样性的丧失共同导致了微生物呼吸速率的下降, 同时减缓了凋落物层的周转, 表明微生物整体的分解能力在施氮之后减弱(Yang et al., 2019)。同时, 多年的氮添加显著降低了微生物生物量和真菌细菌比, 增加微生物对活性碳源的利用效率, 而降低了对惰性碳源的利用效率(Liu et al., 2018)。因此基于之前的研究结果, 我们认为长期施氮处理后, 微生物将植物来源的碳整合进入土壤碳库的能力较弱, 这也是本研究中长期施氮处理显著提高了植物来源的碳输入量, 但未观察到土壤碳库储量变化的可能原因。此外, MAOM的含量不仅受微生物过程影响, 还与土壤矿物含量有关(Sollins et al., 2009)。我们的结果也显示, MAOM氮含量随黏粒和粉粒含量增加而增加(图5D)。而MAOM的碳、氮含量在氮添加后未发生改变, 可能是由于土壤质地在不同的施氮水平样地之间没有显著差异(图2)。模型筛选的结果表明, 长期氮添加处理下, MAOM的氮含量主要受土壤黏粒与粉粒含量的影响, 而凋落物输入量、微生物活性的作用较弱(表2)。结构方程模型的结果也进一步确认, 在氮添加下, 土壤质地是影响MAOM含量的最主要因素(图6B)。
综上所述, 本研究结果表明, 在典型草地生态系统中, 长期氮添加显著促进了土壤氮库积累, 但土壤中增加的氮主要分布在周转速率较快的POM中, 而较为稳定的MAOM中氮含量没有显著变化。储存在POM中的氮更容易受到土地利用变化或气候变化的影响而被分解, 可能通过土壤的淋溶过程造成氮流失; 或通过硝化细菌的反硝化过程, 释放氮氧化物等温室气体, 加剧全球变暖。因此, 为了准确预测全球气候变化下陆地生态系统的氮储量变化以及碳氮循环过程, 未来研究需要考虑大气氮沉降对不同稳定性有机质组分氮库的影响。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
DOI:10.1016/S0038-0717(97)00030-8URL [本文引用: 1]
DOI:10.1016/0038-0717(85)90144-0URL [本文引用: 1]
DOI:10.1111/gcb.12982PMID:25990618 [本文引用: 1]

Labile, 'high-quality', plant litters are hypothesized to promote soil organic matter (SOM) stabilization in mineral soil fractions that are physicochemically protected from rapid mineralization. However, the effect of litter quality on SOM stabilization is inconsistent. High-quality litters, characterized by high N concentrations, low C/N ratios, and low phenol/lignin concentrations, are not consistently stabilized in SOM with greater efficiency than 'low-quality' litters characterized by low N concentrations, high C/N ratios, and high phenol/lignin concentrations. Here, we attempt to resolve these inconsistent results by developing a new conceptual model that links litter quality to the soil C saturation concept. Our model builds on the Microbial Efficiency-Matrix Stabilization framework (Cotrufo et al., 2013) by suggesting the effect of litter quality on SOM stabilization is modulated by the extent of soil C saturation such that high-quality litters are not always stabilized in SOM with greater efficiency than low-quality litters. © 2015 John Wiley & Sons Ltd.
DOI:10.1046/j.1365-2389.2001.00417.xURL [本文引用: 1]
DOI:10.17521/cjpe.2018.0056 [本文引用: 1]
Aims Plant biomass accounts for the main part of grassland productivity. The productivity of grassland regarded as one of important ecosystem function is always co-limited by nitrogen and water availability, therefore, how grasslands respond to atmosphic nitrogen (N) addition and precipitation increasing need to be systematically and quantitatively evaluated at different climate conditions and temporal scales.Methods To investigate the impact of nitrogen addition and precipitation increasing on grassland biomass over climate conditions and temproal scales, a meta-analysis was conducted based on 46 papers that were published during 1990-2017 involving 1 350 observations.Important findings Results showed that: (1) N addtion, precipitation increasing and the combinations of these two treatments significantly increased the aboveground biomass (37%, 41%, 104%), total biomass (32%, 23%, 60%) and the ratio of aboveground biomass to belowground biomass (29%, 25%, 46%) in grassland ecosystem. Belowground biomass showed no response to single N addtion, but could be significantly enhanced together with increaseing precipitation; (2) The response of grassland biomass under these N addtion and the increasing of precipitation showed obvious spatial pattern under different climate conditions. The N addition tended to increase more aboveground biomass, total biomass and the ratio of aboveground biomass to belowground biomass under high sites with high mean annual air temperature (MAT) and mean annual precipitation (MAP) while precipitation increasing tended to simulate more belowground biomass and total biomass under low MAT and MAP sites; (3) In addition, the response of grassland biomass under these two global change index showed obvious temporal pattern. With the increase of duration of N addition, the belowgound biomass tended to decrease, while the aboveground biomass, total biomass and the ratio of aboveground biomass to belowground biomass tended to increase under N addition. With the increase of duration of precipitation manipulation, the total biomass showed no response to precipitation increasing, while aboveground biomass, belowground biomass and the ratio of aboveground biomass to belowground biomass tended to be enhanced. The results indicated that aboveground biomass was more likely to be enhanced than belowground biomass under N addition or precipitation increasing in the long term.
DOI:10.17521/cjpe.2018.0056 [本文引用: 1]
生产力是草地生态系统重要的服务功能, 而生物量作为生态系统生产力的主要组成部分, 往往同时受到氮和水分两个因素的限制。在全球变化背景下, 研究草地生态系统生物量对氮沉降增加和降水变化的响应具有重要意义, 但现有研究缺乏对其在大区域空间尺度以及长时间尺度上响应的综合评估和量化。本研究搜集了1990-2017年间发表论文的有关模拟氮沉降及降水变化研究的相关数据, 进行整合分析, 探讨草地生态系统生物量对氮沉降和降水量两个因素的变化在空间和时间尺度上的响应。结果表明: (1)氮添加、增雨处理以及同时增氮增雨处理都能够显著地提高草地生态系统的地上生物量(37%, 41%, 104%)、总生物量(32%, 23%, 60%)和地上地下生物量比(29%, 25%, 46%)。单独增雨显著提高地下生物量(10%), 单独施氮对地下生物量影响不显著, 但同时增雨则能显著提高地下生物量(43%); (2)氮添加和增雨处理对草地生态系统生物量的影响存在明显的空间变异。在温暖性气候区和海洋性气候区的草地生态系统中, 氮添加对地上、总生物量及地上地下生物量比的促进作用更强, 而在寒冷性气候区和温带大陆性气候区的草地生态系统中, 则增雨处理对地下、总生物量的促进作用更强; (3)草地生态系统生物量对氮添加和增雨处理的响应也存在时间格局上的变化, 地下生物量随着氮添加年限的增加有降低的趋势, 地上、总生物量及地上地下生物量比则有增加的趋势。增雨年限的增加对总生物量没有明显的影响, 但持续促进地上生物量和地下生物量, 增加地上地下生物量比, 可见长期增氮、长期增雨对地上生物量的促进作用更明显。
DOI:10.1111/gcb.2012.18.issue-6URL [本文引用: 1]
[本文引用: 1]
DOI:10.1038/s41561-019-0484-6 [本文引用: 1]
Effective land-based solutions to climate change mitigation require actions that maximize soil carbon storage without generating surplus nitrogen. Land management for carbon sequestration is most often informed by bulk soil carbon inventories, without considering the form in which carbon is stored, its capacity, persistency and nitrogen demand. Here, we present coupling of European-wide databases with soil organic matter physical fractionation to determine continental-scale forest and grassland topsoil carbon and nitrogen stocks and their distribution between mineral-associated and particulate organic matter pools. Grasslands and arbuscular mycorrhizal forests store more soil carbon in mineral-associated organic carbon, which is more persistent but has a higher nitrogen demand and saturates. Ectomycorrhizal forests store more carbon in particulate organic matter, which is more vulnerable to disturbance but has a lower nitrogen demand and can potentially accumulate indefinitely. The share of carbon between mineral-associated and particulate organic matter and the ratio between carbon and nitrogen affect soil carbon stocks and mediate the effects of other variables on soil carbon stocks. Understanding the physical distribution of organic matter in pools of mineral-associated versus particulate organic matter can inform land management for nitrogen-efficient carbon sequestration, which should be driven by the inherent soil carbon capacity and nitrogen availability in ecosystems.
DOI:10.1038/NGEO2520 [本文引用: 1]
Cotrufo, M. Francesca; Soong, Jennifer L.; Horton, Andrew J.; Campbell, Eleanor E.; Haddix, Michelle L.; Wall, Diana H.; Parton, Andwilliam J. Colorado State Univ, Nat Resources Ecol Lab, Ft Collins, CO 80523 USA. Cotrufo, M. Francesca Colorado State Univ, Dept Soil & Crop Sci, Ft Collins, CO 80523 USA. Wall, Diana H. Colorado State Univ, Dept Biol, Ft Collins, CO 80523 USA.
DOI:10.1007/s10533-014-0004-0URL [本文引用: 1]
DOI:10.1126/science.1136674PMID:18487183 [本文引用: 1]
Humans continue to transform the global nitrogen cycle at a record pace, reflecting an increased combustion of fossil fuels, growing demand for nitrogen in agriculture and industry, and pervasive inefficiencies in its use. Much anthropogenic nitrogen is lost to air, water, and land to cause a cascade of environmental and human health problems. Simultaneously, food production in some parts of the world is nitrogen-deficient, highlighting inequities in the distribution of nitrogen-containing fertilizers. Optimizing the need for a key human resource while minimizing its negative consequences requires an integrated interdisciplinary approach and the development of strategies to decrease nitrogen-containing waste.
DOI:10.1023/A:1004213929699URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/nph.12235PMID:23521345 [本文引用: 1]
Demand of all living organisms on the same nutrients forms the basis for interspecific competition between plants and microorganisms in soils. This competition is especially strong in the rhizosphere. To evaluate competitive and mutualistic interactions between plants and microorganisms and to analyse ecological consequences of these interactions, we analysed 424 data pairs from 41 (15)N-labelling studies that investigated (15)N redistribution between roots and microorganisms. Calculated Michaelis-Menten kinetics based on K(m) (Michaelis constant) and V(max) (maximum uptake capacity) values from 77 studies on the uptake of nitrate, ammonia, and amino acids by roots and microorganisms clearly showed that, shortly after nitrogen (N) mobilization from soil organic matter and litter, microorganisms take up most N. Lower K(m) values of microorganisms suggest that they are especially efficient at low N concentrations, but can also acquire more N at higher N concentrations (V(max)) compared with roots. Because of the unidirectional flow of nutrients from soil to roots, plants are the winners for N acquisition in the long run. Therefore, despite strong competition between roots and microorganisms for N, a temporal niche differentiation reflecting their generation times leads to mutualistic relationships in the rhizosphere. This temporal niche differentiation is highly relevant ecologically because it: protects ecosystems from N losses by leaching during periods of slow or no root uptake; continuously provides roots with available N according to plant demand; and contributes to the evolutionary development of mutualistic interactions between roots and microorganisms.© 2013 The Authors. New Phytologist © 2013 New Phytologist Trust.
DOI:10.1111/gcb.14859PMID:31587451 [本文引用: 3]
Managing soil organic matter (SOM) stocks to address global change challenges requires well-substantiated knowledge of SOM behavior that can be clearly communicated between scientists, management practitioners, and policy makers. However, SOM is incredibly complex and requires separation into multiple components with contrasting behavior in order to study and predict its dynamics. Numerous diverse SOM separation schemes are currently used, making cross-study comparisons difficult and hindering broad-scale generalizations. Here, we recommend separating SOM into particulate (POM) and mineral-associated (MAOM) forms, two SOM components that are fundamentally different in terms of their formation, persistence, and functioning. We provide evidence of their highly contrasting physical and chemical properties, mean residence times in soil, and responses to land use change, plant litter inputs, warming, CO enrichment, and N fertilization. Conceptualizing SOM into POM versus MAOM is a feasible, well-supported, and useful framework that will allow scientists to move beyond studies of bulk SOM, but also use a consistent separation scheme across studies. Ultimately, we propose the POM versus MAOM framework as the best way forward to understand and predict broad-scale SOM dynamics in the context of global change challenges and provide necessary recommendations to managers and policy makers.© 2019 John Wiley & Sons Ltd.
DOI:10.1111/ele.2010.13.issue-7URL [本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1016/j.geoderma.2018.07.008URL [本文引用: 3]
DOI:10.1111/nph.2011.189.issue-4URL [本文引用: 1]
DOI:10.5194/bg-9-853-2012URL [本文引用: 1]
DOI:10.1016/j.soilbio.2011.04.012URL [本文引用: 1]
PMID:31312015 [本文引用: 1]
Formation of mineral-organic associations is a key process in the global carbon cycle. Recent concepts propose litter quality-controlled microbial assimilation and direct sorption processes as main factors in transferring carbon from plant litter into mineral-organic associations. We explored the pathways of the formation of mineral-associated organic matter (MOM) in soil profiles along a 120-ky ecosystem gradient that developed under humid climate from the retreating Franz Josef Glacier in New Zealand. We determined the stocks of particulate and mineral-associated carbon, the isotope signature and microbial decomposability of organic matter, and plant and microbial biomarkers (lignin phenols, amino sugars and acids) in MOM. Results revealed that litter quality had little effect on the accumulation of mineral-associated carbon and that plant-derived carbon bypassed microbial assimilation at all soil depths. Seemingly, MOM forms by sorption of microbial as well as plant-derived compounds to minerals. The MOM in carbon-saturated topsoil was characterized by the steady exchange of older for recent carbon, while subsoil MOM arises from retention of organic matter transported with percolating water. Overall, MOM formation is not monocausal but involves various mechanisms and processes, with reactive minerals being effective filters capable of erasing chemical differences in organic matter inputs.
DOI:10.1111/j.1469-8137.2008.02751.xPMID:19210725 [本文引用: 1]
Languishing for many years in the shadow of plant inorganic nitrogen (N) nutrition research, studies of organic N uptake have attracted increased attention during the last decade. The capacity of plants to acquire organic N, demonstrated in laboratory and field settings, has thereby been well established. Even so, the ecological significance of organic N uptake for plant N nutrition is still a matter of discussion. Several lines of evidence suggest that plants growing in various ecosystems may access organic N species. Many soils display amino acid concentrations similar to, or higher than, those of inorganic N, mainly as a result of rapid hydrolysis of soil proteins. Transporters mediating amino acid uptake have been identified both in mycorrhizal fungi and in plant roots. Studies of endogenous metabolism of absorbed amino acids suggest that L- but not D-enantiomers are efficiently utilized. Dual labelled amino acids supplied to soil have provided strong evidence for plant uptake of organic N in the field but have failed to provide information on the quantitative importance of this process. Thus, direct evidence that organic N contributes significantly to plant N nutrition is still lacking. Recent progress in our understanding of the mechanisms underlying plant organic N uptake may open new avenues for the exploration of this subject.
DOI:10.1111/ele.12591URL [本文引用: 1]
DOI:10.1016/j.geoderma.2010.02.003URL [本文引用: 1]
DOI:10.1007/s10533-015-0123-2URL [本文引用: 1]
DOI:10.1038/nature10386URL [本文引用: 1]
DOI:10.2136/sssaj2002.1981URL [本文引用: 1]
DOI:10.1007/s10533-009-9359-zURL [本文引用: 1]
DOI:10.1016/0038-0717(87)90052-6URL [本文引用: 1]
DOI:10.1016/j.soilbio.2007.03.007URL [本文引用: 1]
DOI:10.1111/gcb.12348URL [本文引用: 1]
DOI:10.1111/gcb.12957PMID:25916410 [本文引用: 1]
Organic carbon (OC) sequestration in degraded semi-arid environments by improved soil management is assumed to contribute substantially to climate change mitigation. However, information about the soil organic carbon (SOC) sequestration potential in steppe soils and their current saturation status remains unknown. In this study, we estimated the OC storage capacity of semi-arid grassland soils on the basis of remote, natural steppe fragments in northern China. Based on the maximum OC saturation of silt and clay particles <20 μm, OC sequestration potentials of degraded steppe soils (grazing land, arable land, eroded areas) were estimated. The analysis of natural grassland soils revealed a strong linear regression between the proportion of the fine fraction and its OC content, confirming the importance of silt and clay particles for OC stabilization in steppe soils. This relationship was similar to derived regressions in temperate and tropical soils but on a lower level, probably due to a lower C input and different clay mineralogy. In relation to the estimated OC storage capacity, degraded steppe soils showed a high OC saturation of 78-85% despite massive SOC losses due to unsustainable land use. As a result, the potential of degraded grassland soils to sequester additional OC was generally low. This can be related to a relatively high contribution of labile SOC, which is preferentially lost in the course of soil degradation. Moreover, wind erosion leads to substantial loss of silt and clay particles and consequently results in a direct loss of the ability to stabilize additional OC. Our findings indicate that the SOC loss in semi-arid environments induced by intensive land use is largely irreversible. Observed SOC increases after improved land management mainly result in an accumulation of labile SOC prone to land use/climate changes and therefore cannot be regarded as contribution to long-term OC sequestration. © 2015 John Wiley & Sons Ltd.
DOI:10.1111/1365-2435.13338 [本文引用: 4]
1. Nitrogen (N) deposition not only alters the physiological processes of individual plant, but also leads to world-wide biodiversity loss. However, little is known about how the hierarchical responses from individual physiological processes to plant community structure would have cascading effects on soil carbon (C) cycling. 2. Here, we assessed whether changes in plant chemical composition and community composition under increasing N input would affect the turnover rate of litter layer and soil C loss via heterotrophic respiration (R-h) in a temperate grassland. 3. We showed that more than a decade's N addition significantly decreased plant species richness, litter layer turnover rate and R-h. The C-13-NMR results showed that, for individual species, N addition either increased the abundance of recalcitrant C groups such as alkyl and methoxyl, or decreased labile C groups such as carbohydrate, resulting in decreases in carbohydrate C-to-methoxyl C ratio (CC/MC) for most species. Our data also showed that with the increase in N deposition, the abundance of relatively high degradable dominant species, such as Agropyron cristatum and Artimesia frigida declined rapidly, and the relatively recalcitrant species such as Potentilla bifurca and Leymus chinensis become dominant. Changes in individual species' chemical composition and plant community composition significantly decreased litter quality at community level, as indicated by the lower community-level CC/MC at higher N addition rates. 4. The result of step-AIC model selection further showed that plant diversity loss and the decrease in community-level CC/MC jointly explained the decrease in R-h after N addition best, and further relative importance partition result showed that these two factors respectively contributed 65.1% and 34.9% of the explained variation. 5. Overall, we demonstrated that changes in plant chemical composition and diversity loss due to N addition reduced the quality of plant C input to soil, which further slowed down litter layer turnover rate and inhibited soil heterotrophic respiration. Our study complements the intermediate links of how shifts in plant community structure regulate soil C cycle under global changes.
DOI:10.1038/s41561-019-0352-4URL [本文引用: 1]
DOI:10.1038/s41396-018-0096-yURL [本文引用: 1]
DOI:10.1016/j.atmosenv.2007.12.015URL [本文引用: 1]
An inter-laboratory comparison of ten different ways of measuring soil microbial biomass C.
1
1997
... 土壤样品在2016年8月中旬收集.在每个样方内用直径5 cm的土钻随机取3钻表层土壤(0-15 cm), 混匀为一个土壤样品.利用2 mm的网筛去除土壤样品中较大的根和石头, 冷藏保存并带回实验室.一部分样品被冷冻保存, 用于测定微生物生物量碳氮含量和土壤溶解性无机氮(DIN)含量; 一部分样品自然风干后用于测定土壤的理化性质, 如碳氮含量、机械组成、土壤 pH、有机质组分等.微生物生物量碳、氮含量采用氯仿熏蒸法测定(Beck et al.,
Chloroform fumigation and the release of soil-nitrogen-A rapid direct extraction method to measure microbial biomass nitrogen in soil
1
1985
... 土壤样品在2016年8月中旬收集.在每个样方内用直径5 cm的土钻随机取3钻表层土壤(0-15 cm), 混匀为一个土壤样品.利用2 mm的网筛去除土壤样品中较大的根和石头, 冷藏保存并带回实验室.一部分样品被冷冻保存, 用于测定微生物生物量碳氮含量和土壤溶解性无机氮(DIN)含量; 一部分样品自然风干后用于测定土壤的理化性质, 如碳氮含量、机械组成、土壤 pH、有机质组分等.微生物生物量碳、氮含量采用氯仿熏蒸法测定(Beck et al.,
Integrating plant litter quality, soil organic matter stabilization, and the carbon saturation concept
1
2015
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
Physical fractionation of soil and structural and functional complexity in organic matter turnover
1
2001
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应
1
2018
... 大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al.,
草地生态系统生物量在不同气候及多时间尺度上对氮添加和增雨处理的响应
1
2018
... 大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al.,
Soil organic matter turnover is governed by accessibility not recalcitrance
1
2012
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
Introduction to biostatistics: a guide to design, analysis, and discovery
1
1996
... 研究利用单因素方差分析和最小显著性差异(LSD)法检验氮添加对土壤碳氮含量、不同有机质组分的碳氮含量、土壤质地、土壤 pH、植物AGB和微生物生物量的影响.在统计分析中, 我们参考经典的统计方法, 仅在方差分析显著的情况下对不同处理水平间进行了多重比较, 以减小统计结果中一类错误(即原假设H0符合实际情况, 检验结果却将其否定)发生的概率(Forthofer & Lee,
Soil carbon storage informed by particulate and mineral-associated organic matter
1
2019
... 大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al.,
Formation of soil organic matter via biochemical and physical pathways of litter mass loss
1
2015
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
Chronic nitrogen additions suppress decomposition and sequester soil carbon in temperate forests
1
2014
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
Transformation of the nitrogen cycle: recent trends, questions, and potential solutions
1
2008
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
The capacity of soils to preserve organic C and N by their association with clay and silt particles
1
1997
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
Mineral-organic associations: formation, properties, and relevance in soil environments
1
2015
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance
1
2013
... 土壤氮库是植物和微生物代谢养分的重要来源(Kuzyakov & Xu,
Conceptualizing soil organic matter into particulate and mineral-associated forms to address global change in the 21st century
3
2020
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
... .,
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
A global perspective on belowground carbon dynamics under nitrogen enrichment
1
2010
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
土壤有机碳稳定机制研究进展
1
2007
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
土壤有机碳稳定机制研究进展
1
2007
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
Microbial carbon use efficiency and priming effect regulate soil carbon storage under nitrogen deposition by slowing soil organic matter decomposition
3
2018
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
... .,
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
Responses of ecosystem nitrogen cycle to nitrogen addition: a meta-analysis
1
2011
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
Free and protected soil organic carbon dynamics respond differently to abandonment of mountain grassland
1
2012
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
Organic matter bound to mineral surfaces: resistance to chemical and biological oxidation
1
2011
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
Microbial and abiotic controls on mineral-associated organic matter in soil profiles along an ecosystem gradient
1
2019
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
Uptake of organic nitrogen by plants
1
2009
... 大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al.,
Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle
1
2016
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
A critical review of the conventional SOC to SOM conversion factor
1
2010
... 大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al.,
Nitrogen addition changes grassland soil organic matter decomposition
1
2015
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
Persistence of soil organic matter as an ecosystem property
1
2011
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
Measuring and understanding carbon storage in afforested soils by physical fractionation
1
2002
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
Sequential density fractionation across soils of contrasting mineralogy: evidence for both microbial- and mineral-controlled soil organic matter stabilization
1
2009
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
An extraction method for measuring soil microbial biomass C.
1
1987
... 土壤样品在2016年8月中旬收集.在每个样方内用直径5 cm的土钻随机取3钻表层土壤(0-15 cm), 混匀为一个土壤样品.利用2 mm的网筛去除土壤样品中较大的根和石头, 冷藏保存并带回实验室.一部分样品被冷冻保存, 用于测定微生物生物量碳氮含量和土壤溶解性无机氮(DIN)含量; 一部分样品自然风干后用于测定土壤的理化性质, 如碳氮含量、机械组成、土壤 pH、有机质组分等.微生物生物量碳、氮含量采用氯仿熏蒸法测定(Beck et al.,
SOM fractionation methods: relevance to functional pools and to stabilization mechanisms
1
2007
... 土壤有机质根据其密度或粒径大小被分为颗粒态有机质(POM)和矿质结合态有机质(MAOM)(von Lüetzow et al.,
Nitrogen deposition weakens plant-microbe interactions in grassland ecosystems
1
2013
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
Carbon storage capacity of semi-arid grassland soils and sequestration potentials in Northern China
1
2015
... 为了探究不同有机质组分的质量及其氮含量的变化, 本研究通过土壤密度分级的方法将土壤有机质分为颗粒态有机质(POM)和矿质结合态有机质(MAOM).即: 称取30 g自然风干的土壤样品于250 mL的离心瓶中, 加入150 mL密度为1.8 g·cm-3的NaI溶液, 利用220 r·min-1的摇床振荡3 h, 水平静置24 h后将上层悬浮的自由态POM转移至0.22 μm孔径的滤纸上.剩余沉淀液用150 J·mL-1的能量超声处理, 释放被土壤团聚体包裹的POM, 沉淀液经过高速离心(4 250 r·min-1) 10 min后, 将上层悬浮的包裹态POM转移至0.22 μm孔径的滤纸上.两部分的POM用蒸馏水冲洗至溶液的电导率<2 μs, 重复上述所有操作3次.将剩余的沉淀液高速离心(4 250 r·min-1) 30 min后, 转移至0.22 μm孔径的滤纸上, 用蒸馏水冲洗至溶液的电导率<50 μs, 得到的组分即为MAOM.将POM和MAOM置于50 ℃烘箱中烘至恒质量, 粉碎后利用元素分析仪测定每个组分的碳氮含量.相关方法参见Wiesmeier等(
The decline in plant biodiversity slows down soil carbon turnover under increasing nitrogen deposition in a temperate steppe
4
2019
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
... 大气氮沉降可通过改变凋落物输入量及其氮含量、微生物代谢等过程, 影响土壤中POM和MAOM的氮含量(Riggs et al.,
... 大气氮沉降可通过影响植物向土壤的碳输入, 影响土壤中POM的含量(Francesca Cotrufo et al.,
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
Stabilization of atmospheric nitrogen deposition in China over the past decade
1
2019
... 氮肥使用、化石燃料燃烧和豆科作物种植等人类活动导致全球大气氮沉降量不断增加(Galloway et al.,
Global negative effects of nitrogen deposition on soil microbes
1
2018
... 大气氮沉降对MAOM含量的影响, 不仅与微生物过程相关, 同时受土壤中矿物含量的影响(Kleber et al.,
Evidence for organic N deposition and its anthropogenic sources in China
1
2008
... 本研究位于内蒙古多伦县典型草地生态系统(42.03° N, 116.28° E; 海拔1 324 m).该区域年平均气温为2.1 ℃, 年降水量为382.2 mm, 降水集中在植被生长季(5-9月).自然条件下大气氮沉降速率约为1.6 g·m-2·a-1 (Zhang et al.,

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}