Abstract Aims Epichloë infection can affect the fungal disease resistance of host grasses. However, few studies have been reported on the effects of endophyte infection on non-symbiotic neighbours. Methods We surveyed the plant diseases in natural grassland, and compared differences of total disease index between neighbor and non-neighbor plants of Achnatherum sibiricum. Then the laboratory experiments were conducted to investigate the effects of endophyte on the growth of four pathogen species as well as the brown patch of the host and its neighbor plants. Important findings Major epidemic diseases of grasses were spot blight, rust disease and powdery mildew in Hulun Buir natural grassland. Among common herbages, the total disease index of endophyte-infected A. sibiricum was the lowest. Compared with non-neighbor plants, the brown patch disease index of Leymus chinensis, Stipa baicalensis and Agropyron cristatum was significantly reduced when neighbouring with A. sibiricum. The laboratory experiments results showed that the culture filtration of both Epichloë gansuensis and Epichloë sibiricum significantly restrained the growth of Curvularia lunata, Bipolaris sorokiniana, Sclerotinia sclerotioru and Sclerotinia trifoliorum. The two species of endophytes could reduce lesion area of detached host leaves. In the intact leaves, the endophyte reduced the disease resistance of both the host and its neighbor grasses L. chinensis to C. lunata and B. sorokiniana. This study first verified that the endophytes in A. sibiricum have a positive effect on disease resistance of neighbor grasses to brown patch. Keywords:endophyte;Achnatherum sibiricum;neighbor grasses;fungal pathogens;brown patch
PDF (1562KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 石新建, 张靖歆, 秦天姿, 刘金铭, 高玉葆, 任安芝. 内生真菌感染对宿主羽茅及邻生植物抗病性的影响. 植物生态学报, 2021, 45(8): 860-869. DOI: 10.17521/cjpe.2021.0132 SHI Xin-Jian, ZHANG Jing-Xin, QIN Tian-Zi, LIU Jin-Ming, GAO Yu-Bao, REN An-Zhi. Effects of endophyte infection on fungal disease resistance of Achnatherum sibiricum and non-symbiotic neighbours. Chinese Journal of Plant Ecology, 2021, 45(8): 860-869. DOI: 10.17521/cjpe.2021.0132
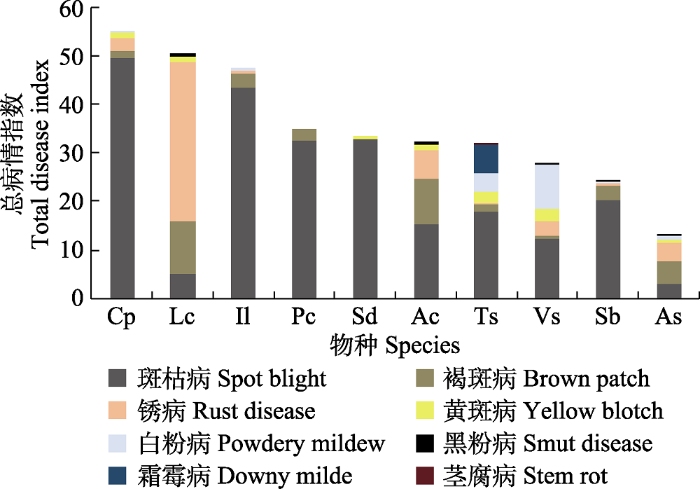
2019年8-9月, 对内蒙古呼伦贝尔草原天然草地的病害进行调查, 调查样地为中国农业科学院农业资源与农业区划研究所呼伦贝尔国家草原生态系统观测研究站的4个观察样地: 海拉尔狼针草(Stipa baicalensis)草甸草原观测样地(120.12° E, 49.35° N, 位于呼伦贝尔市陈巴尔虎旗)、伊敏狼针草草甸草原样地(119.67° E, 48.49° N, 位于呼伦贝尔市鄂温克族自治旗)、羊草(Leymus chinensis)草甸草原退化演替长期观测区(120.05° E, 49.33° N, 位于呼伦贝尔市海拉尔区)、羊草草甸草原观测样地(120.26° E, 49.19° N, 位于呼伦贝尔市海拉尔区)。每块样地面积为0.33 km2。调查样地所在的呼伦贝尔市属中温带半温润和半干旱大陆性气候。1961-2017年的年平均气温-0.59 ℃; 年降水量398 mm; 年平均相对湿度58.38%-70.24%; 年日照时间2 285-2 909 h。
新窗口打开|下载原图ZIP|生成PPT 图2呼伦贝尔草原天然草地中与羽茅邻生和非邻生常见禾草的病情指数分布。A, D, 羊草。B, E, 狼针草。C, F, 冰草。p经非参数Mann-Whitney U检验, p < 0.05表明邻生与非邻生差异显著。
Fig. 2Frequency distribution of disease index of neighbor and non-neighbor plants with Achnatherum sibiricum of the native grassland in Hulun Buir grassland. A, D, Leymus chinensis. B, E, Stipa baicalensis. C, F, Agropyron cristatum. p was tested by nonparametric Mann-Whitney U test, p < 0.05 indicate significant difference between neighbor and non-neighber.
Fig. 3Mean inhibition rate of culture filtrate of different endophytes on four species of pathogenic fungi (mean ± SE, n = 5). Different uppercase letters indicate significant difference among pathogenic fungis (p < 0.05). EG, Epichloë gansuensis; ES, Epichloë sibiricum. Bs, Bipolaris sorokiniana; Cl, Curvularia lunata; Ss, Sclerotinia sclerotioru; St, Sclerotinia trifoliorum.
Fig. 4Mean inhibition rate of disease area on detached leaves of Achnatherum sibiricum after inoculation with 4 pathogenic fungi (means ± SE, n = 5). Different uppercase letters indicate significant difference between different pathogenic fungis (p < 0.05). EF, endophyte-free A. sibiricum leaves; EG, Epichloë gansuensis-infected A. sibiricum leaves; ES, Epichloë sibiricum-infected A. sibiricum leaves; Bs, Bipolaris sorokiniana; Cl, Curvularia lunata; Ss, Sclerotinia sclerotioru; St, Sclerotinia trifoliorum.
Table 2 表2 表2内生真菌(E)、病原真菌(P)和种植方式(M)对羽茅和羊草的病斑数以及病斑长度的多因素方差分析 Table 2Multi-way ANOVA showing the effects of endophyte (E), pathogen (P) and mixture type (M) on lesion number and length of Achnatherum sibiricum and Leymus chinensis
因素 Factor
羽茅 Achnatherum sibiricum
羊草 Leymus chinensis
病斑数 Lesion number
病斑长度 Lesion length
病斑数 Lesion number
病斑长度 Lesion length
F
p
F
p
F
p
F
p
E
137.68
<0.001
65.73
<0.001
P
147.75
<0.001
3.16
0.078
82.59
<0.001
17.53
<0.001
M
1.32
0.271
1.83
0.165
4.87
0.011
0.81
0.451
P × M
0.02
0.980
0.55
0.578
1.24
0.298
0.20
0.819
E × P
4.74
0.032
2.33
0.130
E × M
0.13
0.871
0.31
0.734
E × P × M
0.26
0.774
0.15
0.861
加粗字体代表差异显著(p < 0.05)。 Significant difference (p < 0.05) are shown in bold.
新窗口打开|下载原图ZIP|生成PPT 图5病原真菌侵染下不感染内生真菌(□)和感染内生真菌(■)羽茅的病斑数(A)和病斑长度(B)(平均值±标准误, n = 10)。不同小写字母表示差异显著(p < 0.05)。
Fig. 5Mean number (A) and length of lesions (B) of endophyte-free (□) and endophyte-infected (■) Achnatherum sibiricum after inoculated by pathogenic fungi (means ± SE, n = 10). Different lowercase letters indicate significant difference (p < 0.05).
Fig. 6Mean number (A) and length of lesions (B) of neighbor Leymus chinensis inoculated by pathogenic fungi (means ± SE, n = 10). Different lowercase letters indicate significant difference (p < 0.05). Lc, L. chinensis grown in monoculture (mono); LcEF, L. chinensis grown with endophyte-free Achnatherum sibiricum; LcEI, L. chinensis grown with endophyte-infected A. sibiricum.
AntenNPR, ChenBJW (2021). Detect thy family: mechanisms, ecology and agricultural aspects of kin recognition in plants Plant, Cell & Environment, 44, 1059-1071. [本文引用: 1]
BabikovaZ, GilbertL, BruceTJA, BirkettM, CaulfieldJC, WoodcockC, PickettJA, JohnsonD (2013). Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack Ecology Letters, 16, 835-843. [本文引用: 1]
BaiKY, PengXF (2000). Current situation and countermeasure of sustainable utilization of grassland resources in Inner Mongolia Journal of China Agricultural Resources and Regional Planning, 21, 40-44. [本文引用: 1]
BastíasDA, Martínez-GhersaMA, BallaréCL, GundelPE (2017). Epichloë fungal endophytes and plant defenses: not just alkaloids Trends in Plant Science, 22, 939-948. [本文引用: 1]
BilasRD, BretmanA, BennettT (2021). Friends, neighbours and enemies: an overview of the communal and social biology of plants Plant, Cell & Environment, 44, 997-1013. [本文引用: 1]
ChenZY, GaoTD, YanDF, NiSK, LuF, ShiAB (1997). Result of controlling sheath blight of rice with antagonistic bacteria (Bacillus subtilis 916) under field conditions Chinese Journal of Biological Control, 13(2), 75-78. [本文引用: 1]
ClayK (1990). Fungal endophytes of grasses Annual Review of Ecology, Evolution, and Systematics, 21, 275-297. [本文引用: 1]
de SilvaNI, BrooksS, LumyongS, HydeKD (2019). Use of endophytes as biocontrol agents Fungal Biology Reviews, 33, 133-148. [本文引用: 1]
de WitCT (1960). On competition Verslagen Landbouwkundige Onderzoekigen, 66, 1-82. [本文引用: 1]
ErnebergM, StrandbergB, StrandbergM, JensenB, WeinerJ (2008). Effects of soil disturbance and disease on the growth and reproduction of Lolium perenne(Poaceae) introduced to semi-natural grasslands Polish Journal of Ecology, 56, 593-604. [本文引用: 1]
FiorenzaJE, FernándezPC, OmaciniM (2021). Z-3-Hexenylacetate emissions induced by the endophyte Epichloë occultans at different levels of defoliation during the host plant’s life cycle Fungal Ecology, 49, 101015. DOI: 10.1016/j.funeco.2020.101015. [本文引用: 1]
García-ParisiPA, GrimoldiAA, OmaciniM (2014). Endophytic fungi of grasses protect other plants from aphid herbivory Fungal Ecology, 9, 61-64. [本文引用: 1]
García-ParisiPA, GavilánSA, CasasC, GundelPE, OmaciniM (2021). A fungal endophyte of an annual weed reduces host competitive ability and confers associational protection to wheat Basic and Applied Ecology, 50, 16-24. [本文引用: 1]
HeilM, KarbanR (2010). Explaining evolution of plant communication by airborne signals Trends in Ecology & Evolution, 25, 137-144. [本文引用: 1]
HorwitzSB, LothsteinL, ManfrediJJ, MelladoW, ParnessJ, RoySN, SchiffPB, SorbaraL, ZehebR (1986). Taxol: mechanisms of action and resistance Annals of the New York Academy of Sciences, 466, 733-744. [本文引用: 1]
HouTJ (1987). Major herbage disease in Inner Mongolia Plant Protection, 13, 17-18. [本文引用: 1]
JamesW (1971). An illustrated series of assessment keys for plant diseases, their preparation and usage Canadian Plant Disease Survey, 51, 39-65. [本文引用: 1]
KarbanR, ShiojiriK, IshizakiS, WetzelWC, EvansRY (2013). Kin recognition affects plant communication and defence Proceedings of the Royal Society B: Biological Sciences, 280, 20123062. DOI: 10.1098/rspb.2012.3062. [本文引用: 1]
LatchGCM, ChristensenMJ (1985). Artificial infection of grasses with endophytes Annals of Applied Biology, 107, 17-24. [本文引用: 1]
LiB (1997). The rangeland degradation in north China and its preventive strategy Scientia Agricultura Sinica, 30, 1-9. [本文引用: 1]
LiuH, ChenJ, QinTZ, ShiXJ, GaoYB, RenAZ (2020). Removal of soil microbes alters interspecific competitiveness of Epichloë endophyte-infected over endophyte- free Leymus chinensis Microorganisms, 8, 219. DOI: 10.3390/microorganisms8020219. [本文引用: 1]
LiuJM, GeXY, FanXW, LiuH, GaoYB, RenAZ (2021). The inhibitory effect of endophyte-infected tall fescue on white clover can be alleviated by Glomus mosseae instead of rhizobia Microorganisms, 9, 109. DOI: 10.3390/microorganisms9010109. [本文引用: 1]
LiuY, ZhangYW, NanZB, DuanTY (2016). Progress of research into the effects of native grassland management practices on plant disease Acta Ecologica Sinica, 36, 4211-4220. [本文引用: 1]
MalinowskiDP, AlloushGA, BeleskyDP (1998). Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum Plant and Soil, 205, 1-12. [本文引用: 1]
MüllerCB, KraussJ (2005). Symbiosis between grasses and asexual fungal endophytes Current Opinion in Plant Biology, 8, 450-456. [本文引用: 1]
MurphyB, DoohanF, HodkinsonT (2018). From concept to commerce: developing a successful fungal endophyte inoculant for agricultural crops Journal of Fungi, 4, 24. DOI: 10.3390/jof4010024. [本文引用: 2]
NanZB, LiCJ (1994). The directory of forage grasses disease in China Pratacultural Science, 13(Suppl.), 1-160. [本文引用: 1]
NiuY, GaoY, LiGP, RenAZ, GaoYB (2016). Effect of different species of endophytes on fungal disease resistance of Achnatherum sibiricum Chinese Journal of Plant Ecology, 40, 925-932. [本文引用: 1]
OloweOM, AkanmuAO, AsemoloyeMD (2020). Exploration of microbial stimulants for induction of systemic resistance in plant disease management Annals of Applied Biology, 177, 282-293. [本文引用: 1]
PańkaD, PiesikD, JeskeM, Baturo-CieśniewskaA (2013a). Production of phenolics and the emission of volatile organic compounds by perennial ryegrass (Lolium perenne L.)/Neotyphodium lolii association as a response to infection by Fusarium poae Journal of Plant Physiology, 170, 1010-1019. [本文引用: 3]
PańkaD, WestCP, GuerberCA, RichardsonMD (2013b). Susceptibility of tall fescue to Rhizoctonia zeae infection as affected by endophyte symbiosis Annals of Applied Biology, 163, 257-268. [本文引用: 1]
PatchettA, NewmanJA (2021). Comparison of plant metabolites in root exudates of Lolium perenne infected with different strains of the fungal endophyte Epichloë festucae var. lolii Journal of Fungi, 7, 148. DOI: 10.3390/jof7020148. [本文引用: 1]
PélissierR, ViolleC, MorelJB (2021). Plant immunity: good fences make good neighbors? Current Opinion in Plant Biology, 62, 102045. DOI: 10.1016/j.pbi.2021.102045. [本文引用: 1]
PengD, LiSD, WangJX, ChenCJ, ZhouMG (2014). Integrated biological and chemical control of rice sheath blight by Bacillus subtilis NJ-18 and jinggangmycin Pest Management Science, 70, 258-263. [本文引用: 1]
PérezLI, GundelPE, OmaciniM (2016). Can the defensive mutualism between grasses and fungal endophytes protect non-symbiotic neighbours from soil pathogens? Plant and Soil, 405, 289-298. [本文引用: 2]
PérezLI, GundelPE, ZabalgogeazcoaI, OmaciniM (2020). An ecological framework for understanding the roles of Epichloë endophytes on plant defenses against fungal diseases Fungal Biology Reviews, 34, 115-125. [本文引用: 1]
PlattTG, BeverJD (2009). Kin competition and the evolution of cooperation Trends in Ecology & Evolution, 24, 370-377. [本文引用: 1]
QawasmehA, ObiedHK, RamanA, WheatleyW (2012). Influence of fungal endophyte infection on phenolic content and antioxidant activity in grasses: interaction between Lolium perenne and different strains of Neotyphodium lolii Journal of Agricultural and Food Chemistry, 60, 3381-3388. [本文引用: 1]
QinJH, GaoY, LiuH, ZhouY, RenAZ, GaoYB (2016). Effect of endophyte infection and clipping treatment on resistance and tolerance of Achnatherum sibiricum Frontiers in Microbiology, 7, 1988. DOI: 10.3389/fmicb.2016.01988. [本文引用: 1]
RaymaekersK, PonetL, HoltappelsD, BerckmansB, CammueBPA (2020). Screening for novel biocontrol agents applicable in plant disease management: a review Biological Control, 144, 104240. DOI: 10.1016/j.biocontrol.2020. 104240. [本文引用: 1]
ReyT, DumasB (2017). Plenty is no plague: streptomyces symbiosis with crops Trends in Plant Science, 22, 30-37. [本文引用: 1]
RiedellWE, KieckheferRE, PetroskiRJ, PowellRG (1991). Naturally-occurring and synthetic loline alkaloid derivatives: insect feeding behavior modification and Toxicity Journal of Entomological Science, 26, 122-129. [本文引用: 1]
SchardlCL, FloreaS, PanJ, NagabhyruP, BecS, CaliePJ (2013). The epichloae: alkaloid diversity and roles in symbiosis with grasses Current Opinion in Plant Biology, 16, 480-488. [本文引用: 1]
ShiXJ, QinTZ, LiuH, WuM, LiJJ, ShiYS, GaoYB, RenAZ (2020). Endophytic fungi activated similar defense strategies of Achnatherum sibiricum host to different trophic types of pathogens Frontiers in Microbiology, 11, 1607. DOI: 10.3389/fmicb.2020.01607. [本文引用: 1]
SongYY, YeM, LiCY, HeXH, Zhu-SalzmanK, WangRL, SuYJ, LuoSM, ZengRS (2014). Hijacking common mycorrhizal networks for herbivore-induced defence signal transfer between tomato plants Scientific Reports, 4, 3915. DOI: 10.1038/srep03915. [本文引用: 1]
SongYY, ZengRS, XuJF, LiJ, ShenX, YihdegoWG (2010). Interplant communication of tomato plants through underground common mycorrhizal networks PLOS ONE, 5, e13324. DOI: 10.1371/journal.pone.0013324. [本文引用: 1]
SteinebrunnerF, SchiestlFP, LeuchtmannA (2008a). Ecological role of volatiles produced by Epichloë: differences in antifungal toxicity FEMS Microbiology Ecology, 64, 307-316. [本文引用: 2]
SteinebrunnerF, TweleR, FranckeW, LeuchtmannA, SchiestlFP (2008b). Role of odour compounds in the attraction of gamete vectors in endophytic Epichloë fungi New Phytologist, 178, 401-411. [本文引用: 2]
StierleA, StrobelG, StierleD (1993). Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew Science, 260, 214-216. [本文引用: 1]
SubrahmaniamHJ, LibourelC, JournetEP, MorelJB, MuñosS, NiebelA, RaffaeleS, RouxF (2018). The genetics underlying natural variation of plant-plant interactions, a beloved but forgotten member of the family of biotic interactions The Plant Journal, 93, 747-770. [本文引用: 1]
TanakaA, TakemotoD, ChujoT, ScottB (2012). Fungal endophytes of grasses Current Opinion in Plant Biology, 15, 462-468. [本文引用: 1]
VikukV, YoungCA, LeeST, NagabhyruP, KrischkeM, MuellerMJ, KraussJ (2019). Infection rates and alkaloid patterns of different grass species with systemic Epichloë endophytes Applied and Environmenta Microbiology, 85, e00465-9. DOI: 10.1128/aem.00465-19. [本文引用: 1]
VurukondaSSKP, GiovanardiD, StefaniE (2018). Plant growth promoting and biocontrol activity of streptomyces spp. as endophytes International Journal of Molecular Sciences, 19, 952. DOI: 10.3390/ijms19040952. [本文引用: 1]
WeiYK, GaoYB, XuH, SuD, ZhangX, WangYH, LinF, ChenL, NieLY, RenAZ (2006). Occurrence of endophytes in grasses native to Northern China Grass and Forage Science, 61, 422-429. [本文引用: 1]
WeiYK, GaoYB, ZhangX, SuD, WangYH, XuH, LinF, RenAZ, ChenL, NieLY (2007). Distribution and diversity of Epichloë/Neotyphodium fungal endophytes from different populations of Achnatherum sibiricum(Poaceae) in the Inner Mongolia Steppe, China Fungal Diversity, 24, 329-345. [本文引用: 1]
XiaC, LiNN, ZhangYW, LiCJ, ZhangXX, NanZB (2018). Role of Epichloë endophytes in defense responses of cool-season grasses to pathogens: a review Plant Disease, 102, 2061-2073. [本文引用: 2]
YueQ, MillerCJ, WhiteJF, RichardsonMD (2000). Isolation and characterization of fungal inhibitors from Epichloë festucae Journal of Agricultural and Food Chemistry, 48, 4687-4692. [本文引用: 1]
ZhangX, RenAZ, WeiYK, LinF, LiC, LiuZJ, GaoYB (2009). Taxonomy, diversity and origins of symbiotic endophytes of Achnatherum sibiricum in the Inner Mongolia Steppe of China FEMS Microbiology Letters, 301, 12-20. [本文引用: 1]
ZhengRH, LiSJ, ZhangX, ZhaoCQ (2021). Biological activities of some new secondary metabolites isolated from endophytic fungi: a review study International Journal of Molecular Sciences, 22, 959. DOI: 10.3390/ijms22020959. [本文引用: 1]
ZinNA, BadaluddinNA (2020). Biological functions of Trichoderma spp. for agriculture applications Annals of Agricultural Sciences, 65, 168-178. [本文引用: 1]
Detect thy family: mechanisms, ecology and agricultural aspects of kin recognition in plants 1 2021
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
Underground signals carried through common mycelial networks warn neighbouring plants of aphid attack 1 2013
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
Z-3-Hexenylacetate emissions induced by the endophyte Epichloë occultans at different levels of defoliation during the host plant’s life cycle 1 2021
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Endophytic fungi of grasses protect other plants from aphid herbivory 1 2014
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
A fungal endophyte of an annual weed reduces host competitive ability and confers associational protection to wheat 1 2021
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
Explaining evolution of plant communication by airborne signals 1 2010
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
Taxol: mechanisms of action and resistance 1 1986
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
内蒙古的主要牧草病害 1 1987
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
内蒙古的主要牧草病害 1 1987
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
我国北方草地病害调查及主要病害防治 1 1993
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
我国北方草地病害调查及主要病害防治 1 1993
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
An illustrated series of assessment keys for plant diseases, their preparation and usage 1 1971
Epichloë endophytes alter inducible indirect defences in host grasses 1 2014
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Removal of soil microbes alters interspecific competitiveness of Epichloë endophyte-infected over endophyte- free Leymus chinensis 1 2020
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
The inhibitory effect of endophyte-infected tall fescue on white clover can be alleviated by Glomus mosseae instead of rhizobia 1 2021
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum 1 1998
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Symbiosis between grasses and asexual fungal endophytes 1 2005
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
From concept to commerce: developing a successful fungal endophyte inoculant for agricultural crops 2 2018
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
... ).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
Exploration of microbial stimulants for induction of systemic resistance in plant disease management 1 2020
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
Production of phenolics and the emission of volatile organic compounds by perennial ryegrass (Lolium perenne L.)/Neotyphodium lolii association as a response to infection by Fusarium poae 3 2013a
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
... ; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
... )、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Susceptibility of tall fescue to Rhizoctonia zeae infection as affected by endophyte symbiosis 1 2013b
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Comparison of plant metabolites in root exudates of Lolium perenne infected with different strains of the fungal endophyte Epichloë festucae var. lolii 1 2021
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Plant immunity: good fences make good neighbors? 1 2021
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
Integrated biological and chemical control of rice sheath blight by Bacillus subtilis NJ-18 and jinggangmycin 1 2014
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
Can the defensive mutualism between grasses and fungal endophytes protect non-symbiotic neighbours from soil pathogens? 2 2016
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
Kin competition and the evolution of cooperation 1 2009
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
Influence of fungal endophyte infection on phenolic content and antioxidant activity in grasses: interaction between Lolium perenne and different strains of Neotyphodium lolii 1 2012
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Effect of endophyte infection and clipping treatment on resistance and tolerance of Achnatherum sibiricum 1 2016
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Screening for novel biocontrol agents applicable in plant disease management: a review 1 2020
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
Plenty is no plague: streptomyces symbiosis with crops 1 2017
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
Naturally-occurring and synthetic loline alkaloid derivatives: insect feeding behavior modification and Toxicity 1 1991
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
Fungal endophytes help prevent weed invasions 1 2013
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
The epichloae: alkaloid diversity and roles in symbiosis with grasses 1 2013
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
Endophytic fungi activated similar defense strategies of Achnatherum sibiricum host to different trophic types of pathogens 1 2020
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Hijacking common mycorrhizal networks for herbivore-induced defence signal transfer between tomato plants 1 2014
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
Interplant communication of tomato plants through underground common mycorrhizal networks 1 2010
... 此外, 植物也可以通过共生微生物如菌根真菌网络传递信号, 激活邻生植物对生物胁迫的响应(Song et al., 2010, 2014; Babikova et al., 2013).然而内生真菌对邻生植物抗病性的研究鲜有报道.目前, 关于Epichlo-禾草互作对邻生植物影响的研究大都集中在种间竞争上(Saikkonen et al., 2013; Liu et al., 2020, 2021), 而对邻生植物抵抗生物胁迫的研究仍然很少(García-Parisi et al., 2014, 2021).在本研究中, 天然草地中羽茅Epichloë内生真菌感染率100%.我们推测羽茅对邻生禾草病害程度的影响可能与其内生真菌有关.但因自然生境中羽茅的内生真菌感染率达到100%, 为了证实内生真菌的作用, 还需进行控制实验. ...
Ecological role of volatiles produced by Epichloë: differences in antifungal toxicity 2 2008a
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Role of odour compounds in the attraction of gamete vectors in endophytic Epichloë fungi 2 2008b
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew 1 1993
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
The genetics underlying natural variation of plant-plant interactions, a beloved but forgotten member of the family of biotic interactions 1 2018
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
Fungal endophytes of grasses 1 2012
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
Infection rates and alkaloid patterns of different grass species with systemic Epichloë endophytes 1 2019
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
Plant growth promoting and biocontrol activity of streptomyces spp. as endophytes 1 2018
... 近年来, 以环境污染小、持久性强为主要特点的生物防治技术不断发展, 以芽孢杆菌(Bacillus spp.)、链霉菌(Streptomyces spp.)和木霉菌(Trichoderma spp.)等为代表的植物生防菌在农牧业病害防治中已得到广泛应用(Rey & Dumas, 2017; Murphy et al., 2018; Raymaekers et al., 2020; Zin & Badaluddin, 2020).但目前普遍存在的问题是生物防治直接施用活的微生物菌剂, 所以周围环境如温湿度、土壤酸碱度、土壤中微生物群落等因素对生物防治效果均有很大的影响(Murphy et al., 2018; Vurukonda et al., 2018; Olowe et al., 2020).植物内生菌以植物体为宿主进行生存和繁衍而不引起宿主明显病症.内生菌在与宿主植物长期地协同进化中, 会促进宿主产生或自身合成独特生物活性的代谢物, 如具有抗癌活性的紫杉醇(Horwitz et al., 1986; Stierle et al., 1993)、广谱抗菌活性的萜类和酮类化合物等(Zheng et al., 2021).一些利用植物内生菌如枯草芽胞杆菌(Bacillus subtilis)开发的生物农药“纹曲宁”已成功用于防治水稻纹枯病及伪黑穗病(陈志谊等, 1997; Peng et al., 2014).由于内生菌在植物界广泛分布, 而且受外界因素的干扰较小, 是挖掘成为新型生物农药的重要潜在资源(de Silva et al., 2019). ...
Root exudate signals in plant-plant interactions 1 2021
... 本研究调查发现, 在天然草地常见牧草中禾本科植物发病最严重, 斑枯病、锈病和白粉病是其主要病害; 在豆科植物中, 斑枯病、白粉病和褐斑病发病最严重.这与侯天爵(1987, 1993)对内蒙古草原病害调查结果基本一致.对天然草地病害状况进一步分析发现, 羽茅对邻生植物如莎草科柄状薹草、毛茛科展枝唐松草以及伞形科防风等植物发病程度无显著影响, 但显著降低了邻生禾本科植物羊草、冰草和狼针草的褐斑病病害.这可能与物种之间的亲缘关系有关(Subrahmaniam et al., 2018; Anten & Chen, 2021; Bilas et al., 2021).植物之间普遍存在化学信号交流(Heil & Karban, 2010; Wang et al., 2021).物种间更近的进化关系可能使彼此的信号交流识别更加有效(Platt & Bever, 2009; Pélissier et al., 2021).例如, 与远亲缘关系的邻生植物相比, 三齿蒿(Artemisia tridentata)植株的挥发物可以显著提高近亲缘植物对生物胁迫的抗性(Karban et al., 2013). ...
Distribution and diversity of Epichloë/Neotyphodium fungal endophytes from different populations of Achnatherum sibiricum(Poaceae) in the Inner Mongolia Steppe, China 1 2007
Isolation and characterization of fungal inhibitors from Epichloë festucae 1 2000
... 在天然草地中, 许多冷季型禾草可以与子囊菌门麦角菌科的Epichloë属内生真菌形成互利共生的关系(Clay, 1990; Müller & Krauss, 2005; Tanaka et al., 2012; Vikuk et al., 2019).国内外关于禾草内生真菌作为生物防治手段的基础研究已经取得丰富成果.例如, 内生真菌产生具有广谱抗虫活性的生物碱(Riedell et al., 1991; Schardl et al., 2013; Bastías et al., 2017)以及抗菌活性的chokol K等(Yue et al., 2000; Steinebrunner et al., 2008a, 2008b)次生代谢物, 是潜在的替代化学农药的生物防控剂.目前, 内生真菌对宿主抗病性影响的研究很多(Xia et al., 2018), 但是关于内生真菌对宿主邻生植物抗病性的研究非常少(Pérez et al., 2016).挖掘天然禾草内生真菌资源, 探索内生真菌的生物防治机制, 将对农牧业绿色防控技术的发展带来十分积极的影响. ...
Volatile compounds of endophyte-free and infected tall fescue (Festuca arundinacea Schreb.) 1 2001
... 目前, 关于内生真菌增强宿主抗病性的机制并不十分清楚, 推测可能与植物免疫相关的次生代谢物有关.例如, 内生真菌可以促进宿主产生具有抗菌活性的酚类化合物, 这些物质可能通过根系分泌(Malinowski et al., 1998; Patchett & Newman, 2021)、叶片淋溶及凋落物分解进入土壤(Qawasmeh et al., 2012; Pańka et al., 2013a, 2013b), 保护宿主以及邻近植物的根系免受土传病原真菌危害.另外, 也有研究发现内生真菌感染可以诱导宿主产生激活植物免疫系统或具有抗菌活性的挥发性有机化合物(VOCs), 如(Z)-3-己烯-1-乙酸酯(Z-3-HAC)(Yue et al., 2001; Pańka et al., 2013a; Li et al., 2014; Fiorenza et al., 2021)、水杨酸甲酯(Pańka et al., 2013a)和chokol K (Steinebrunner et al., 2008a, 2008b)等.至于内生真菌增强宿主邻生植物抗病性的原因, 既可能与内生真菌感染提高了宿主的酚类物质含量(Qin et al., 2016; Shi et al., 2020)有关, 也可能由于内生真菌诱导宿主散发的VOCs激活了邻生植物的免疫调控响应.这些推测还需要进一步的实验证据验证. ...
Taxonomy, diversity and origins of symbiotic endophytes of Achnatherum sibiricum in the Inner Mongolia Steppe of China 1 2009
 ,*南开大学生命科学学院, 天津 300071
,*南开大学生命科学学院, 天津 300071

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT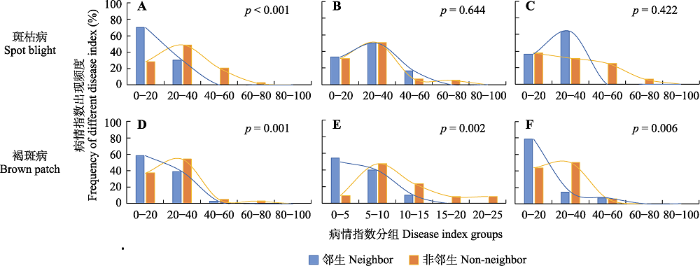 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT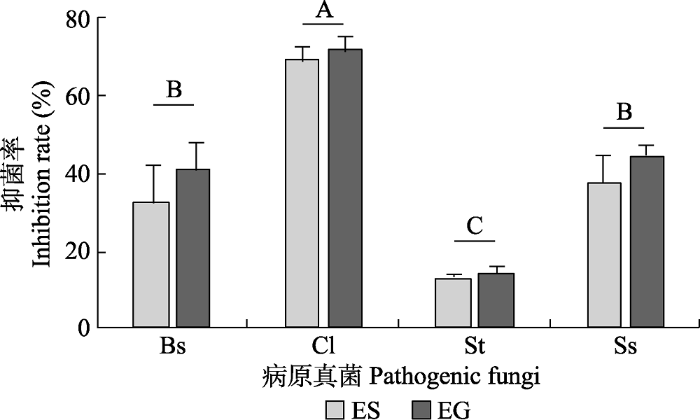 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT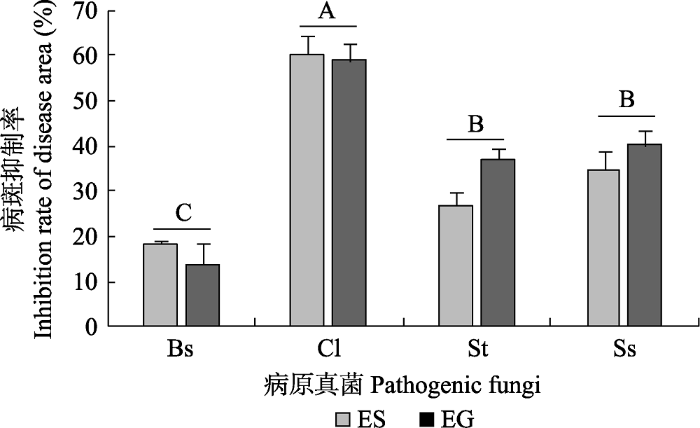 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT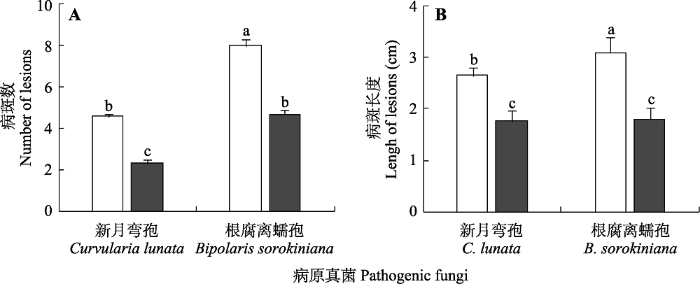 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT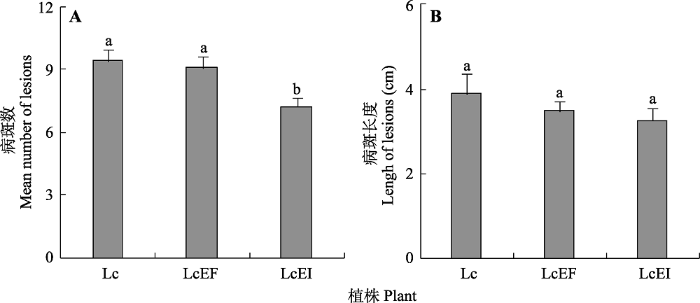 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}