Influences of warming and snow reduction in winter on soil nutrients and bacterial communities composition in a typical grassland of the Loess Plateau
Jin MAO1, Ying DUO1, Jun DENG2, Jie CHENG3, Ji-Min CHENG4, Chang-Hui PENG4,5, Liang GUO,4,6,*1College of Grassland Agriculture, Northwest A&F University, Yangling, Shaanxi 712100, China 2Administration Bureau of Ningxia Yunwushan National Nature Reserve, Guyuan, Ningxia 756000, China 3Northwest Surveying Planning and Designing Institute of National Forestry and Grassland Administration, Xiʼan 710048, China 4State Key Laboratory of Soil Erosion and Dryland Farming on the Loess Plateau, Northwest A&F University, Yangling, Shaanxi 712100, China 5Institute of Environmental Sciences, University of Quebec at Montreal, Montreal H3C 3P8, Canada 6Institute of Soil and Water Conservation, Chinese Academy of Sciences & Ministry of Water Resources, Yangling, Shaanxi 712100, China
National Key R&D Program of China(2016YFC0500700) National Natural Science Foundation of China(41701606) Fundamental Research Funds for the Central Universities(2452020009)
Abstract Aims Variations in temperature and snow accumulations in winter will change the structure and function of the soil-microbial system. As a key biological factor in the terrestrial ecosystem, microorganisms play an important role in regulating soil nutrient cycles. However, they are very sensitive to environmental disturbances, especially to winter climate changes. It is in great need to study the response of soil nutrients and microbial properties of typical semi-arid grasslands to climate change in winter, in order to predict the ecological process and functional changes of grassland ecosystem in the long term. Methods In the present study, the semi-arid grassland in the Yunwushan National Nature Reserve in Ningxia Province was taken as the research object. The four treatments including warming (W), snow reduction (S), interaction of warming and snow reduction (WS), and control (CK) were set to explore the responses of soil nutrients, enzyme activities and soil bacterial communities in the 0-5 cm soil layer of the typical grassland of the Loess Plateau to variations in winter temperature and snow cover. Important findings Our results indicated that: (1) Warming, snow reduction and their interaction in winter increased the 0-5 cm soil temperature, lowered the relative humidity of the soil, but significantly increased the number of soil freeze-thaw cycles. (2) Compared with the control, other different treatments generally reduced the microbial biomass and bacterial diversity, which led to reduced activity of soil β-1,4-glucosidase (BG), β-1,4-N-acetylglucosaminidase (NAG) and alkaline phosphatase (AKP). The content of soil organic carbon, total nitrogen, available phosphorus, and nitrate nitrogen in the soil increased, while the content of nitrate nitrogen decreased. (3) The soil bacterial species in the study area were mainly Acidobacteria, Proteobacteria, Actinobacteria and Gemmatimonadetes. The dominant bacteria at the class level included Acidobacteria, γ-Proteobacteria, Thermophiles and σ-Proteobacteria. Redundancy analysis (RDA) results showed that available phosphorus (AP) content had the most significant impact on the bacterial community composition, with an explanation rate of 21.3% for the community variation. In conclusion, winter climate change can significantly affect soil temperature and humidity, especially the freezing and thawing cycles, which might further influence soil nutrients cycles, enzyme activities, and soil bacterial diversity. These results are of great significance for enriching and expanding the understanding of the process and mechanism of climate change on grassland ecosystem, as well as predicting the mid and long-term dynamic changes of typical grassland ecosystems. Keywords:soil nutrients;soil microorganisms;winter;warming;changes in snowfall
PDF (2637KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 毛瑾, 朵莹, 邓军, 程杰, 程积民, 彭长辉, 郭梁. 冬季增温和减雪对黄土高原典型草原土壤养分和细菌群落组成的影响. 植物生态学报, 2021, 45(8): 891-902. DOI: 10.17521/cjpe.2021.0085 MAO Jin, DUO Ying, DENG Jun, CHENG Jie, CHENG Ji-Min, PENG Chang-Hui, GUO Liang. Influences of warming and snow reduction in winter on soil nutrients and bacterial communities composition in a typical grassland of the Loess Plateau. Chinese Journal of Plant Ecology, 2021, 45(8): 891-902. DOI: 10.17521/cjpe.2021.0085
Fig. 2Influence of different treatments on the soil temperature and relative humidity underground 5 cm from November 2018 to March 2019. CK, control; S, snow reduction; W, warming; WS, interaction of warming and snow reduction.
Table 1 表1 表1冬季土壤冻融循环次数在不同处理下的比较 Table 1Comparison of the numbers of soil freeze-thaw cycles under different treatments in winter
处理 Treatment
年份-月份 Year-month
冻融循环次数 Numbers of freeze-thaw cycle
总计 Sum
对照 CK
2018-11
2
29
2018-12
0
2019-01
0
2019-02
8
2019-03
19
增温 W
2018-11
7
53
2018-12
22
2019-01
0
2019-02
24
2019-03
0
减雪 S
2018-11
8
51
2018-12
21
2019-01
9
2019-02
13
2019-03
0
互作 WS
2018-11
2
61
2018-12
16
2019-01
14
2019-02
23
2019-03
6
CK, control; S, snow reduction; W, warming; WS, interaction of warming and snow reduction.
Table 2 表2 表2土壤理化性质和酶活性在不同处理下的比较(平均值±标准差, n = 4) Table 2Comparison of soil physical and chemical properties and enzyme activities under different treatments (means ± SD, n = 4)
土壤指标 Soil indicator
对照 CK
增温 W
减雪 S
互作 WS
F
p
土壤有机碳含量 SOC content (g·kg-1)
23.66 ± 1.87b
30.92 ± 0.28a
25.00 ± 0.94b
25.15 ± 1.09b
7.39
*
全氮含量 TN content (g·kg-1)
2.47 ± 0.06c
3.03 ± 0.03a
2.97 ± 0.03a
2.71 ± 0.02b
47.28
***
速效磷含量 AP content (mg·kg-1)
3.30 ± 0.15c
3.97 ± 0.15ab
4.17 ± 0.03a
3.53 ± 0.2bc
7.23
*
硝态氮含量 NO- 3-N content (mg·kg-1)
6.33 ± 0.41a
4.37 ± 0.12b
6.40 ± 0.55a
4.83 ± 0.24b
7.93
**
铵态氮含量 NH+ 4-N content (mg·kg-1)
3.90 ± 0.41b
4.03 ± 0.12b
3.33 ± 0.55b
5.83 ± 0.24a
4.73
*
pH
7.51 ± 0.02a
7.51 ± 0.01a
7.48 ± 0a
7.52 ± 0.02a
1.54
NS
微生物生物量碳含量 MBC content (mg·kg-1)
820.08 ± 23.97b
951.73 ± 6.91a
807.58 ± 48.58b
696.02 ± 17.49c
13.33
**
微生物生物量氮含量 MBN content (mg·kg-1)
63.48 ± 1.51a
67.57 ± 2.57a
61.39 ± 9.10a
54.60 ± 1.26a
1.26
NS
β-1,4-葡萄糖苷酶活性 BG activity (nmol·g-1·h-1)
203.37 ± 16.45a
211.55 ± 13.82a
197.87 ± 16.04ab
157.08 ± 4.32b
3.19
NS
β-1,4-N-乙酰基氨基葡萄糖苷酶活性 NAG activity (nmol·g-1·h-1)
76.85 ± 1.67a
70.94 ± 2.33a
74.33 ± 5.31a
56.33 ± 1.63b
8.64
**
碱性磷酸酶活性 AKP activity (nmol·g-1·h-1)
656.61 ± 14.04a
642.68 ± 46.49a
586.07 ± 30.72ab
539.64 ± 4.12b
3.60
NS
同一行中不同字母表示差异显著。*, p < 0.05; **, p < 0.01; ***, p < 0.001; NS, p > 0.05。 AKP, alkaline phosphatase; AP, available phosphorus; BG, β-1,4-glucosidase; MBC, microbial biomass carbon; MBN, microbial biomass nitrogen; NAG, β-1,4-N-acetylglucosaminidase; NH+ 4-N, ammonium nitrogen; NO- 3-N, nitrate nitrogen; SOC, soil organic carbon; TN, total nitrogen. Different letters in the same row indicate significant differences. *, p < 0.05; **, p < 0.01; ***, p < 0.001; NS, p > 0.05.
Fig. 3Comparison of soil bacterial diversity under different treatments. CK, control; S, snow reduction; W, warming; WS, interaction of warming and snow reduction. Different lowercase letters indicate significant differences among different treatments (p < 0.05).
Fig. 4Comparison of the relative abundance of soil bacteria at different taxonomic levels under different experimental treatments. A, Relative abundance of bacteria at phylum level. B, Relative abundance of bacteria at class level. CK, control; S, snow reduction; W, warming; WS, interaction of warming and snow reduction. Different lowercase letters of the same taxon indicate significant differences in abundance between different treatments (p < 0.05). The lowercase letters are marked in the center of the corresponding bar graph of taxon relative abundance.
酸杆菌门的相对丰度在增温处理和增温减雪互作下显著减少(p < 0.05), 在减雪处理中相对丰度有降低但不显著, 不同处理下酸杆菌门相对丰度大小为: CK > S > WS > W。放线菌门的相对丰度变化表现为: W > S > CK > WS, 在增温减雪互作下相对丰度最小。与放线菌门类似, 芽单胞菌门的相对丰度也在互作处理下最低, 其相对丰度CK > W > S > WS, 而变形菌门相对丰度在不同处理间未发现显著差异(图4A)。各处理下细菌纲水平的变化如图4B, 酸杆菌纲的相对丰度变化表现为: CK > W > S > WS, 增温减雪互作下酸杆菌纲丰度与对照相比有下降趋势(0.13% vs. 0.15%, 图4B), 这与嗜热油菌纲相对丰度变化一致。与变形菌门水平响应类似, γ-变形杆菌纲和σ-变形菌纲在不同处理间亦不存在显著差异。
Fig. 5Redundancy analysis (RDA) of soil bacterial communities and soil physical and chemical properties. CK, control; S, snow reduction; W, warming; WS, interaction of warming and snow reduction. AP, available phosphorus content; MBC, microbial biomass carbon content; MBN, microbial biomass nitrogen content; NH4+-N, ammonium nitrogen content; NO3--N, nitrate nitrogen content; SOC, soil organic carbon content; TN, total nitrogen content. The blue arrows indicate the distribution of soil bacterial populations at the phylum level, and the red arrows indicate environmental factors.
Table 3 表3 表3土壤细菌多样性和各分类群丰度与土壤理化性质和酶活性的Pearson相关性分析 Table 3Pearson correlation coefficients of soil bacterial diversity and abundance of various taxon with soil physical and chemical properties and enzyme activities
Fig. 6Potential influencing paths of soil warming, snow reduction and interactions between the above two treatments on soil microbial community, enzyme activity, and nutrient availability. SOC, soil organic carbon; TN, soil total nitrogen. The thick red arrow in the figure represents the change in the content or frequency of the index.
冻融频发导致土壤微生物大量死亡的报道较多(Larsen et al., 2002; Sulkava & Huhta, 2003; Sjursen et al., 2005), 微生物死亡可导致微生物生物量和微生物多样性下降(图6), 此假设在本研究中也得到了证实: 互作处理下细菌种类最少, 多样性下降最显著, 其3种多样性指数分别下降了5.9%、10.5%和8.7% (图3); 门分类水平上, 细菌群落组成的前4门细菌, 除变形菌门相对丰度在不同处理间未发现显著差异外, 增温、减雪及互作处理下其余3种细菌(酸杆菌门、放线菌门、芽单胞菌门)相对丰度均显著低于对照(图4A)。微生物大量死亡及其多样性的下降, 会造成微生物生物量下降。本研究中互作处理下, 土壤MBC含量显著降低, 这与Freppaz等(2007)的结果一致, 后者发现一半以上的微生物可在第一个冻融周期中被杀死, 进而导致微生物生物量迅速减少。冻融加剧导致土壤MBN含量下降亦见诸报道, 如Song等(2017)发现冻融过程使土壤MBN含量下降12.2%, 本研究中MBN含量有所下降, 但并不显著(表2), 推测可能与微生物组成、抗逆性及恢复力不同有关(Tierney et al., 2003; Schmidt et al., 2007)。
土壤酶的产生大多来自动植物和微生物的分泌物, 酶促反应可以催化土壤内部多种化学元素的转化及生理过程(王楠等, 2014)。土壤酶活性受多种因素的影响, 例如土壤温湿度、养分可利用性、微生物生物量和群落组成及环境因子等(Weintraub et al., 2013)。本研究中BG、NAG、AKP 3种胞外酶活性在增温和减雪互作的处理下显著下降(表2),究其原因, 一方面土壤冻融频发导致微生物大量死亡, 分泌胞外酶含量随之减少, 从而直接或间接影响土壤酶活性(Torres et al., 2015; López-Aizpún et al., 2018)(图6)。另一方面, 增温、减雪和互作处理下土壤湿度下降(图2B)也可能是造成酶催化反应效率下降的原因。Chaer等(2009)对美国安德鲁实验森林土壤分析表明, β-葡萄糖苷酶活性在1、2、4和8个冻融循环之后降低; DeForest等(2012)发现磷酸酶活性在未冻结土壤中比冻结土壤中高两倍, 冻结环境会降低磷酸酶活性, 也与本研究结果一致。然而, 微生物群落组成与分泌酶种类、含量及其驱动的土壤碳氮磷循环过程和机制研究, 目前仍较为匮乏。已有研究表明, 酸杆菌可产生β-葡萄糖苷酶、纤维二糖水解酶及NAG等碳氮分解关键酶(Lladó et al., 2016), 放线菌在有机质周转、碳循环中也具有重要作用(Chinnadurai et al., 2014)。酸杆菌和放线菌作为研究区土壤中重要菌门, 在互作处理下相对丰度下降, 可导致土壤碳氮循环相关酶活性的降低, 也可以部分解释土壤中有机质积累及NO- 3-N含量的下降(表2), 但其具体作用机制仍不清楚。
冬季温度和降雪变化不仅直接影响冬季土壤养分循环、微生物多样性及关键酶活性, 其滞后效应还会对生长季生态过程和功能产生持续影响(Durán et al., 2014)。如本研究中, 冬季增温、减雪及其互作一方面提高了土壤养分含量(图6), 对生长季植物生长具有一定促进作用; 但另一方面, 冬季增温减雪引致的土壤频繁冻融又降低了微生物多样性、丰度及胞外酶活性, 减缓了有机物质的循环周转, 对生长季植物生长具有抑制效应。冬季气候变化(增温及降雪减少)对生长季草地生态系统的影响究竟是正效应还是负效应, 作用机制如何, 其滞后效应是否随时间尺度而改变(Luo et al., 2001; Melillo et al., 2002), 都是亟待解决的关键科学问题, 需开展深入研究。
BardgettRD, FreemanC, OstleNJ (2008). Microbial contributions to climate change through carbon cycle feedbacks The ISME Journal, 2, 805-814. [本文引用: 1]
BlankinshipJC, HartSC (2012). Consequences of manipulated snow cover on soil gaseous emission and N retention in the growing season: a meta-analysis Ecosphere, 3, 1-20. [本文引用: 1]
CaporasoJG, KuczynskiJ, StombaughJ, BittingerK, BushmanFD, CostelloEK, FiererN, PeñaAG, GoodrichJK, GordonJI, HuttleyGA, KelleyST, KnightsD, KoenigJE, LeyRE, et al. (2010). QIIME allows analysis of high-throughput community sequencing data Nature Methods, 7, 335-336. [本文引用: 1]
ChaerGM, MyroldDD, BottomleyPJ (2009). A soil quality index based on the equilibrium between soil organic matter and biochemical properties of undisturbed coniferous forest soils of the Pacific Northwest Soil Biology & Biochemistry, 41, 822-830. [本文引用: 1]
ChengJM (2014). Grassland Ecosystem of the Loess Plateau in China—Yunwushan National Nature Reserve. Science Press, Beijing. [本文引用: 1]
ChinnaduraiC, GopalaswamyG, BalachandarD (2014). Impact of long-term organic and inorganic nutrient managements on the biological properties and eubacterial community diversity of the Indian semi-arid Alfisol Archives of Agronomy and Soil Science, 60, 531-548. [本文引用: 1]
DeForestJL, SmemoKA, BurkeDJ, ElliottHL, BeckerJC (2012). Soil microbial responses to elevated phosphorus and pH in acidic temperate deciduous forests Biogeochemistry, 109, 189-202. [本文引用: 1]
DuránJ, MorseJL, GroffmanPM, CampbellJL, ChristensonLM, DriscollCT, FaheyTJ, FiskMC, MitchellMJ, TemplerPH (2014). Winter climate change affects growing- season soil microbial biomass and activity in northern hardwood forests Global Change Biology, 20, 3568-3577. [本文引用: 1]
EdgarRC (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads Nature Methods, 10, 996-998. [本文引用: 1]
EisenhauerN, DobiesT, CesarzS, HobbieSE, MeyerRJ, WormK, ReichPB (2013). Plant diversity effects on soil food webs are stronger than those of elevated CO2 and N deposition in a long-term grassland experiment Proceedings of the National Academy of Sciences of the United States of America, 110, 6889-6894. [本文引用: 1]
FreppazM, WilliamsBL, EdwardsAC, ScalengheR, ZaniniE (2007). Simulating soil freeze/thaw cycles typical of winter alpine conditions: implications for N and P availability Applied Soil Ecology, 35, 247-255. [本文引用: 1]
GrimmNB, Chapin IIIFS, BierwagenB, GonzalezP, GroffmanPM, LuoYQ, MeltonF, NadelhofferK, PairisA, RaymondPA, SchimelJ, WilliamsonCE (2013). The impacts of climate change on ecosystem structure and function Frontiers in Ecology and the Environment, 11, 474-482. [本文引用: 1]
GuoL, ChengJ, LuedelingE, KoernerSE, HeJS, XuJ, GangC, LiW, LuoR, PengC (2017). Critical climate periods for grassland productivity on China’s Loess Plateau Agricultural and Forest Meteorology, 233, 101-109. [本文引用: 3]
HardyJP, GroffmanPM, FitzhughRD, HenryKS, WelmanAT, DemersJD, FaheyTJ, DriscollCT, TierneyGL, NolanS (2001). Snow depth manipulation and its influence on soil frost and water dynamics in a northern hardwood forest Biogeochemistry, 56, 151-174. [本文引用: 1]
HudsonJMG, HenryGHR (2009). Increased plant biomass in a High Arctic heath community from 1981 to 2008 Ecology, 90, 2657-2663. [本文引用: 1]
IPCC (2014). The Physical Science Basis—Summary for Policymakers. Contribution of WG1 to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, UK. [本文引用: 2]
KnappAK, SmithMD (2001). Variation among biomes in temporal dynamics of aboveground primary production Science, 291, 481-484. [本文引用: 1]
KreylingJ, BeierkuhnleinC, JentschA (2010). Effects of soil freeze-thaw cycles differ between experimental plant communities Basic and Applied Ecology, 11, 65-75. [本文引用: 3]
LarsenKS, JonassonS, MichelsenA (2002). Repeated freeze- thaw cycles and their effects on biological processes in two arctic ecosystem types Applied Soil Ecology, 21, 187-195. [本文引用: 1]
LiW, WuJ, BaiE, GuanD, WangA, YuanF, WangS, JinC (2016). Response of terrestrial nitrogen dynamics to snow cover change: a meta-analysis of experimental manipulation Soil Biology & Biochemistry, 100, 51-58. [本文引用: 1]
LladóS, ŽifčákováL, VětrovskýT, EichlerováI, BaldrianP (2016). Functional screening of abundant bacteria from acidic forest soil indicates the metabolic potential of Acidobacteria subdivision 1 for polysaccharide decomposition Biology and Fertility of Soils, 52, 251-260. [本文引用: 1]
López-AizpúnM, Arango-MoraC, SantamaríaC, LasherasE, SantamaríaJM, CigandaVS, CárdenasLM, ElustondoD (2018). Atmospheric ammonia concentration modulates soil enzyme and microbial activity in an oak forest affecting soil microbial biomass Soil Biology & Biochemistry, 116, 378-387. [本文引用: 1]
LuoY, WanS, HuiD, WallaceLL (2001). Acclimatization of soil respiration to warming in a tall grass prairie Nature, 413, 622-625. [本文引用: 1]
MagočT, SalzbergSL (2011). FLASH: fast length adjustment of short reads to improve genome assemblies Bioinformatics, 27, 2957-2963. [本文引用: 1]
MarxMC, WoodM, JarvisSC (2001). A microplate fluorimetric assay for the study of enzyme diversity in soils Soil Biology & Biochemistry, 33, 1633-1640. [本文引用: 1]
MelilloJM, SteudlerPA, AberJD, NewkirkK, LuxH, BowlesFP, CatricalaC, MagillA, AhrensT, MorrisseauS (2002). Soil warming and carbon-cycle feedbacks to the climate system Science, 298, 2173-2176. [本文引用: 1]
NiuSL, HanXG, MaKP, WanSQ (2007). Field facilities in global warming and terrestrial ecosystem research Journal of Plant Ecology (Chinese Version), 31, 262-271. [本文引用: 2]
PengSS, PiaoSL, CiaisP, FangJY, WangXH (2010). Change in winter snow depth and its impacts on vegetation in China Global Change Biology, 16, 3004-3013. [本文引用: 1]
QinDH (2014). Climate change science and sustainable development Progress in Geography, 33, 874-883. [本文引用: 1]
ReinmannAB, SusserJR, DemariaEMC, TemplerPH (2019). Declines in northern forest tree growth following snowpack decline and soil freezing Global Change Biology, 25, 420-430. [本文引用: 1]
RognesT, FlouriT, NicholsB, QuinceC, MahéF (2016). VSEARCH: a versatile open source tool for metagenomics PeerJ, 4, e2584. DOI: 10.7717/peerj.2584. [本文引用: 1]
Romero-OlivaresAL, AllisonSD, TresederKK (2017). Soil microbes and their response to experimental warming over time: a meta-analysis of field studies Soil Biology & Biochemistry, 107, 32-40. [本文引用: 1]
SchmidtSK, CostelloEK, NemergutDR, ClevelandCC, ReedSC, WeintraubMN, MeyerAF, MartinAM (2007). Biogeochemical consequences of rapid microbial turnover and seasonal succession in soil Ecology, 88, 1379-1385. [本文引用: 1]
SjursenH, MichelsenA, HolmstrupM (2005). Effects of freeze-thaw cycles on microarthropods and nutrient availability in a sub-Arctic soil Applied Soil Ecology, 28, 79-93. [本文引用: 1]
SongY, ZouYC, WangGP, YuXF (2017). Altered soil carbon and nitrogen cycles due to the freeze-thaw effect: a meta-analysis Soil Biology & Biochemistry, 109, 35-49. [本文引用: 2]
SorensenPO, FinziAC, GiassonMA, ReinmannAB, Sanders- DeMottR, TemplerPH, Biochemistry (2018). Winter soil freeze-thaw cycles lead to reductions in soil microbial biomass and activity not compensated for by soil warming Soil Biology & Biochemistry, 116, 39-47. [本文引用: 2]
SulkavaP, HuhtaV (2003). Effects of hard frost and freeze-thaw cycles on decomposer communities and N mineralisation in boreal forest soil Applied Soil Ecology, 22, 225-239. [本文引用: 1]
TanB, WuFZ, YangWQ, YangYL, WangA, KangLN (2011). Effects of snow pack removal on the dynamics of winter-time soil temperature, carbon, nitrogen, and phosphorus in alpine forests of west Sichuan Chinese Journal of Applied Ecology, 22, 2553-2559. [本文引用: 1]
TierneyGL, FaheyTJ, GroffmanPM, HardyJP, FitzhughRD, DriscollCT, YavittJB (2003). Environmental control of fine root dynamics in a northern hardwood forest Global Change Biology, 9, 670-679. [本文引用: 1]
TierneyGL, FaheyTJ, GroffmanPM, HardyJP, FitzhughRD, DriscollCT (2001). Soil freezing alters fine root dynamics in a northern hardwood forest Biogeochemistry, 56, 175-190. [本文引用: 1]
TorresIF, BastidaF, HernándezT, AlbaladejoJ, GarcíaC (2015). Enzyme activity, microbial biomass and community structure in a long-term restored soil under semi-arid conditions Soil Research, 53, 553-560. [本文引用: 1]
VanceED, BrookesPC, JenkinsonDS (1987). An extraction method for measuring soil microbial biomass C Soil Biology & Biochemistry, 19, 703-707. [本文引用: 1]
WangN, WangS, GaoQ, ZhaoLP, TianT, ZhangJJ (2014). Effect of nitrogen application levels on microbiological characteristics of soils with different fertility basics Journal of Soil and Water Conservation, 28, 148-152. [本文引用: 1]
WangQ, GarrityGM, TiedjeJM, ColeJR (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy Applied and Environmental Microbiology, 73, 5261-5267. [本文引用: 1]
WangY, LiuJS, WangGP, ZhouWM (2007). Study on the effect of freezing and thawing action to soil physical and chemical characteristics Geography and Geo-information Science, 23, 91-96. [本文引用: 1]
WeintraubSR, WiederWR, ClevelandCC, TownsendAR (2013). Organic matter inputs shift soil enzyme activity and allocation patterns in a wet tropical forest Biogeochemistry, 114, 313-326. [本文引用: 1]
XiaoW, ChenX, JingX, ZhuBA (2018). A meta-analysis of soil extracellular enzyme activities in response to global change Soil Biology & Biochemistry, 123, 21-32. [本文引用: 1]
YuXJ, YangJS, WangET, LiBZ, YuanHL (2015). Effects of growth stage and fulvic acid on the diversity and dynamics of endophytic bacterial community in Stevia rebaudiana Bertoni leaves Frontiers in Microbiology, 6, 867. DOI: 10.3389/fmicb.2015.00867. [本文引用: 1]
ZhouW, ChenH, ZhouL, LewisBJ, YeY, TianJ, LiG, DaiL (2011). Effect of freezing-thawing on nitrogen mineralization in vegetation soils of four landscape zones of Changbai Mountain Annals of Forest Science, 68, 943-951. [本文引用: 1]
A meta- analysis of experimental warming effects on terrestrial nitrogen pools and dynamics 2 2013
... 全球变化背景下土壤微生物群落组成和多样性、土壤酶活性及碳氮磷等养分变化的响应研究一直是生态学研究的热点问题.近年来, 已有大量控制试验及meta分析文章针对上述科学问题进行了较为系统与全面的阐述(Bai et al., 2013; Romero- Olivares et al., 2017; Xiao et al., 2018), 但以往研究大都侧重于全年增温或生长季增温的生态效应探索(Guo et al., 2017), 如Bai等(2013)汇总分析了全球陆地生态系统氮循环对气候变化响应的528个观测数据, 发现生长季和全年增温实验数量占全部样本的85.4%, 冬季增温实验不仅个案数量少, 其测定指标更为匮乏.然而, 大量研究指出冬季土壤微生物活动仍较为频繁, 养分物质循环及能量代谢依旧进行(Kreyling et al., 2010; Sorensen et al., 2018), 显著的冬季气候变化(增温及降雪减少)不仅影响植被休眠期内土壤微生物和养分循环, 对生长季植被生长发育及群落构建、稳定性等亦具有持续影响(Hudson & Henry, 2009).因此, 探究冬季温度及积雪变化下土壤理化性质, 以及微生物群落组成、多样性的变化规律, 对丰富气候变化的生态效应研究内涵、准确预测未来气候情景下生态系统响应规律, 进而促进植被资源管理能力提升具有重要意义. ...
... 采用重铬酸盐氧化法测定样品中土壤有机碳(SOC)含量, 凯式定氮仪(Kjeltec 8400, FOSS Corporation, Hillerod, Denmark)测定总氮(TN)含量.速效磷(AP)含量用0.5 mol·L-1 NaHCO3浸提-钼锑抗比色法测定.铵态氮(NH+ 4-N)和硝态氮(NO- 3-N)含量利用流动分析仪(Alpkem, OI Analytical, College Station, USA)测定.土壤pH用酸度计进行分析.微生物生物量碳(MBC)和氮(MBN)含量采用氯仿熏蒸-K2SO4浸提法测定(Vance et al., 1987).选取参与C、N、P循环的3种关键酶, 即β-1,4-葡萄糖苷酶(BG), β-1,4-N-乙酰氨基葡萄糖苷酶(NAG)和碱性磷酸酶(AKP), 使用微孔板荧光法检测其活性(Marx et al., 2001).以上土壤样品理化性质详细测定步骤参考《土壤农化分析》(鲍士旦, 2000). ...
1 2000
... 采用重铬酸盐氧化法测定样品中土壤有机碳(SOC)含量, 凯式定氮仪(Kjeltec 8400, FOSS Corporation, Hillerod, Denmark)测定总氮(TN)含量.速效磷(AP)含量用0.5 mol·L-1 NaHCO3浸提-钼锑抗比色法测定.铵态氮(NH+ 4-N)和硝态氮(NO- 3-N)含量利用流动分析仪(Alpkem, OI Analytical, College Station, USA)测定.土壤pH用酸度计进行分析.微生物生物量碳(MBC)和氮(MBN)含量采用氯仿熏蒸-K2SO4浸提法测定(Vance et al., 1987).选取参与C、N、P循环的3种关键酶, 即β-1,4-葡萄糖苷酶(BG), β-1,4-N-乙酰氨基葡萄糖苷酶(NAG)和碱性磷酸酶(AKP), 使用微孔板荧光法检测其活性(Marx et al., 2001).以上土壤样品理化性质详细测定步骤参考《土壤农化分析》(鲍士旦, 2000). ...
Microbial contributions to climate change through carbon cycle feedbacks 1 2008
... 土壤微生物和参与碳(C)、氮(N)、磷(P)循环的关键酶活性是衡量土壤质量的重要指标, 是参与土壤有机物质积累、矿化作用及调控土壤养分循环的关键因子, 受生物和非生物因素综合作用(沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016), 并且对环境扰动敏感度较高.大量生长季开展的研究表明, 温度升高可增强微生物和酶活性, 加速土壤有机质分解, 增强土壤呼吸, 对全球变化产生正反馈作用(Bardgett et al., 2008; Yue et al., 2015); 然而, 冬季增温和降雪变化对土壤微生物群落结构功能、关键酶活性及地上生态过程的影响研究相对较少, 且彼此间结论存在较大差异(Guo et al., 2017).有研究指出, 冬季升温可提高土壤酶和微生物活性, 促进有机质的分解和氮矿化速率, 提高土壤养分有效性(Grimm et al., 2013); 但也有报道发现, 增温降低了积雪厚度与持续时间, 积雪绝热保温作用的降低导致土壤冻融交替频发, 导致微生物大量裂解死亡(Schuerings et al., 2014), 进而降低了土壤养分循环速率(Blankinship & Hart, 2012; Li et al., 2016).此外, 降雪对陆地生态系统影响的研究主要集中于北半球高纬度寒区或冻区, 以及中低纬度高海拔地区, 温带干旱半干旱区冬季积雪变化及其影响效应的研究较少(Kreyling et al., 2010).因此, 本研究以黄土高原半干旱草原为研究对象, 探究冬季气候变化(增温和积雪减少)对草地生态系统土壤微生物、酶活性及养分循环影响特征, 不仅能丰富和拓展气候变化对生态系统影响过程与机制的认识, 也对提高草地资源管理能力和水平, 特别是在全球冬季显著增温和降雪减少背景下, 准确预测草地生态系统中长期动态变化具有重要意义. ...
Consequences of manipulated snow cover on soil gaseous emission and N retention in the growing season: a meta-analysis 1 2012
... 土壤微生物和参与碳(C)、氮(N)、磷(P)循环的关键酶活性是衡量土壤质量的重要指标, 是参与土壤有机物质积累、矿化作用及调控土壤养分循环的关键因子, 受生物和非生物因素综合作用(沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016), 并且对环境扰动敏感度较高.大量生长季开展的研究表明, 温度升高可增强微生物和酶活性, 加速土壤有机质分解, 增强土壤呼吸, 对全球变化产生正反馈作用(Bardgett et al., 2008; Yue et al., 2015); 然而, 冬季增温和降雪变化对土壤微生物群落结构功能、关键酶活性及地上生态过程的影响研究相对较少, 且彼此间结论存在较大差异(Guo et al., 2017).有研究指出, 冬季升温可提高土壤酶和微生物活性, 促进有机质的分解和氮矿化速率, 提高土壤养分有效性(Grimm et al., 2013); 但也有报道发现, 增温降低了积雪厚度与持续时间, 积雪绝热保温作用的降低导致土壤冻融交替频发, 导致微生物大量裂解死亡(Schuerings et al., 2014), 进而降低了土壤养分循环速率(Blankinship & Hart, 2012; Li et al., 2016).此外, 降雪对陆地生态系统影响的研究主要集中于北半球高纬度寒区或冻区, 以及中低纬度高海拔地区, 温带干旱半干旱区冬季积雪变化及其影响效应的研究较少(Kreyling et al., 2010).因此, 本研究以黄土高原半干旱草原为研究对象, 探究冬季气候变化(增温和积雪减少)对草地生态系统土壤微生物、酶活性及养分循环影响特征, 不仅能丰富和拓展气候变化对生态系统影响过程与机制的认识, 也对提高草地资源管理能力和水平, 特别是在全球冬季显著增温和降雪减少背景下, 准确预测草地生态系统中长期动态变化具有重要意义. ...
QIIME allows analysis of high-throughput community sequencing data 1 2010
... 使用TruSeq® DNA PCR-Free Sample Preparation Kit建库试剂盒进行文库构建, 构建好的文库经过Qubit和Q-PCR定量, 文库合格后, 使用HiSeq2500 PE250进行上机测序.根据Barcode序列和PCR扩增引物序列从下机数据中拆分出各样本数据, 对每个样本的reads进行拼接(Magoč & Salzberg, 2011), 得到原始Tags数据(Raw Tags), 原始数据在Quantitative Insights Into Microbial Ecology (QIIME)平台进行过滤(Caporaso et al., 2010)、去除嵌合体序列(Rognes et al., 2016)最终得到有效数据(Effective Tags).对所有样本的有效数据以97%的一致性进行操作分类单元(Operational Taxonomic Units, OTU)聚类, 用Mothur方法与SILVA132 (http://www.arb- silva.de/)(Edgar, 2013)的SSUrRNA数据库(Wang et al., 2007)对OTU序列进行物种注释(设定阈值为0.8-1), 最后获得OTU丰度表. ...
A soil quality index based on the equilibrium between soil organic matter and biochemical properties of undisturbed coniferous forest soils of the Pacific Northwest 1 2009
... 土壤酶的产生大多来自动植物和微生物的分泌物, 酶促反应可以催化土壤内部多种化学元素的转化及生理过程(王楠等, 2014).土壤酶活性受多种因素的影响, 例如土壤温湿度、养分可利用性、微生物生物量和群落组成及环境因子等(Weintraub et al., 2013).本研究中BG、NAG、AKP 3种胞外酶活性在增温和减雪互作的处理下显著下降(表2),究其原因, 一方面土壤冻融频发导致微生物大量死亡, 分泌胞外酶含量随之减少, 从而直接或间接影响土壤酶活性(Torres et al., 2015; López-Aizpún et al., 2018)(图6).另一方面, 增温、减雪和互作处理下土壤湿度下降(图2B)也可能是造成酶催化反应效率下降的原因.Chaer等(2009)对美国安德鲁实验森林土壤分析表明, β-葡萄糖苷酶活性在1、2、4和8个冻融循环之后降低; DeForest等(2012)发现磷酸酶活性在未冻结土壤中比冻结土壤中高两倍, 冻结环境会降低磷酸酶活性, 也与本研究结果一致.然而, 微生物群落组成与分泌酶种类、含量及其驱动的土壤碳氮磷循环过程和机制研究, 目前仍较为匮乏.已有研究表明, 酸杆菌可产生β-葡萄糖苷酶、纤维二糖水解酶及NAG等碳氮分解关键酶(Lladó et al., 2016), 放线菌在有机质周转、碳循环中也具有重要作用(Chinnadurai et al., 2014).酸杆菌和放线菌作为研究区土壤中重要菌门, 在互作处理下相对丰度下降, 可导致土壤碳氮循环相关酶活性的降低, 也可以部分解释土壤中有机质积累及NO- 3-N含量的下降(表2), 但其具体作用机制仍不清楚. ...
1 2014
... 研究区位于宁夏云雾山国家级自然保护区(36.17°-36.28° N, 106.35°-106.45° E, 海拔1 800-?2 100 m), 该区地处黄土高原中间地带, 具有典型温带半干旱气候特征.年平均气温7 ℃, 年降水量425 mm, 降水主要集中于6-8月.冬季降雨较少, 降雪为冬季主要降水形式, 11月初开始降雪, 2月底3月初积雪深度达到最大, 随后迅速下降, 4月初基本融化(Guo et al., 2017).土壤类型多为黄绵土、山地灰褐土与黑垆土.研究区优势物种以长芒草(Stipa bungeana)、大针茅(S. grandis)、甘青针茅(S. przewalskyi)、百里香(Thymus mongolicus)、白莲蒿(Artemisia stechmanniana)以及星毛委陵菜(Potentilla acaulis)为主(程积民, 2014). ...
1 2014
... 研究区位于宁夏云雾山国家级自然保护区(36.17°-36.28° N, 106.35°-106.45° E, 海拔1 800-?2 100 m), 该区地处黄土高原中间地带, 具有典型温带半干旱气候特征.年平均气温7 ℃, 年降水量425 mm, 降水主要集中于6-8月.冬季降雨较少, 降雪为冬季主要降水形式, 11月初开始降雪, 2月底3月初积雪深度达到最大, 随后迅速下降, 4月初基本融化(Guo et al., 2017).土壤类型多为黄绵土、山地灰褐土与黑垆土.研究区优势物种以长芒草(Stipa bungeana)、大针茅(S. grandis)、甘青针茅(S. przewalskyi)、百里香(Thymus mongolicus)、白莲蒿(Artemisia stechmanniana)以及星毛委陵菜(Potentilla acaulis)为主(程积民, 2014). ...
Impact of long-term organic and inorganic nutrient managements on the biological properties and eubacterial community diversity of the Indian semi-arid Alfisol 1 2014
... 土壤酶的产生大多来自动植物和微生物的分泌物, 酶促反应可以催化土壤内部多种化学元素的转化及生理过程(王楠等, 2014).土壤酶活性受多种因素的影响, 例如土壤温湿度、养分可利用性、微生物生物量和群落组成及环境因子等(Weintraub et al., 2013).本研究中BG、NAG、AKP 3种胞外酶活性在增温和减雪互作的处理下显著下降(表2),究其原因, 一方面土壤冻融频发导致微生物大量死亡, 分泌胞外酶含量随之减少, 从而直接或间接影响土壤酶活性(Torres et al., 2015; López-Aizpún et al., 2018)(图6).另一方面, 增温、减雪和互作处理下土壤湿度下降(图2B)也可能是造成酶催化反应效率下降的原因.Chaer等(2009)对美国安德鲁实验森林土壤分析表明, β-葡萄糖苷酶活性在1、2、4和8个冻融循环之后降低; DeForest等(2012)发现磷酸酶活性在未冻结土壤中比冻结土壤中高两倍, 冻结环境会降低磷酸酶活性, 也与本研究结果一致.然而, 微生物群落组成与分泌酶种类、含量及其驱动的土壤碳氮磷循环过程和机制研究, 目前仍较为匮乏.已有研究表明, 酸杆菌可产生β-葡萄糖苷酶、纤维二糖水解酶及NAG等碳氮分解关键酶(Lladó et al., 2016), 放线菌在有机质周转、碳循环中也具有重要作用(Chinnadurai et al., 2014).酸杆菌和放线菌作为研究区土壤中重要菌门, 在互作处理下相对丰度下降, 可导致土壤碳氮循环相关酶活性的降低, 也可以部分解释土壤中有机质积累及NO- 3-N含量的下降(表2), 但其具体作用机制仍不清楚. ...
Soil microbial responses to elevated phosphorus and pH in acidic temperate deciduous forests 1 2012
... 土壤酶的产生大多来自动植物和微生物的分泌物, 酶促反应可以催化土壤内部多种化学元素的转化及生理过程(王楠等, 2014).土壤酶活性受多种因素的影响, 例如土壤温湿度、养分可利用性、微生物生物量和群落组成及环境因子等(Weintraub et al., 2013).本研究中BG、NAG、AKP 3种胞外酶活性在增温和减雪互作的处理下显著下降(表2),究其原因, 一方面土壤冻融频发导致微生物大量死亡, 分泌胞外酶含量随之减少, 从而直接或间接影响土壤酶活性(Torres et al., 2015; López-Aizpún et al., 2018)(图6).另一方面, 增温、减雪和互作处理下土壤湿度下降(图2B)也可能是造成酶催化反应效率下降的原因.Chaer等(2009)对美国安德鲁实验森林土壤分析表明, β-葡萄糖苷酶活性在1、2、4和8个冻融循环之后降低; DeForest等(2012)发现磷酸酶活性在未冻结土壤中比冻结土壤中高两倍, 冻结环境会降低磷酸酶活性, 也与本研究结果一致.然而, 微生物群落组成与分泌酶种类、含量及其驱动的土壤碳氮磷循环过程和机制研究, 目前仍较为匮乏.已有研究表明, 酸杆菌可产生β-葡萄糖苷酶、纤维二糖水解酶及NAG等碳氮分解关键酶(Lladó et al., 2016), 放线菌在有机质周转、碳循环中也具有重要作用(Chinnadurai et al., 2014).酸杆菌和放线菌作为研究区土壤中重要菌门, 在互作处理下相对丰度下降, 可导致土壤碳氮循环相关酶活性的降低, 也可以部分解释土壤中有机质积累及NO- 3-N含量的下降(表2), 但其具体作用机制仍不清楚. ...
Microbial diversity drives multifunctionality in terrestrial ecosystems 1 2016
... 土壤微生物和参与碳(C)、氮(N)、磷(P)循环的关键酶活性是衡量土壤质量的重要指标, 是参与土壤有机物质积累、矿化作用及调控土壤养分循环的关键因子, 受生物和非生物因素综合作用(沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016), 并且对环境扰动敏感度较高.大量生长季开展的研究表明, 温度升高可增强微生物和酶活性, 加速土壤有机质分解, 增强土壤呼吸, 对全球变化产生正反馈作用(Bardgett et al., 2008; Yue et al., 2015); 然而, 冬季增温和降雪变化对土壤微生物群落结构功能、关键酶活性及地上生态过程的影响研究相对较少, 且彼此间结论存在较大差异(Guo et al., 2017).有研究指出, 冬季升温可提高土壤酶和微生物活性, 促进有机质的分解和氮矿化速率, 提高土壤养分有效性(Grimm et al., 2013); 但也有报道发现, 增温降低了积雪厚度与持续时间, 积雪绝热保温作用的降低导致土壤冻融交替频发, 导致微生物大量裂解死亡(Schuerings et al., 2014), 进而降低了土壤养分循环速率(Blankinship & Hart, 2012; Li et al., 2016).此外, 降雪对陆地生态系统影响的研究主要集中于北半球高纬度寒区或冻区, 以及中低纬度高海拔地区, 温带干旱半干旱区冬季积雪变化及其影响效应的研究较少(Kreyling et al., 2010).因此, 本研究以黄土高原半干旱草原为研究对象, 探究冬季气候变化(增温和积雪减少)对草地生态系统土壤微生物、酶活性及养分循环影响特征, 不仅能丰富和拓展气候变化对生态系统影响过程与机制的认识, 也对提高草地资源管理能力和水平, 特别是在全球冬季显著增温和降雪减少背景下, 准确预测草地生态系统中长期动态变化具有重要意义. ...
Winter climate change affects growing- season soil microbial biomass and activity in northern hardwood forests 1 2014
... 冬季温度和降雪变化不仅直接影响冬季土壤养分循环、微生物多样性及关键酶活性, 其滞后效应还会对生长季生态过程和功能产生持续影响(Durán et al., 2014).如本研究中, 冬季增温、减雪及其互作一方面提高了土壤养分含量(图6), 对生长季植物生长具有一定促进作用; 但另一方面, 冬季增温减雪引致的土壤频繁冻融又降低了微生物多样性、丰度及胞外酶活性, 减缓了有机物质的循环周转, 对生长季植物生长具有抑制效应.冬季气候变化(增温及降雪减少)对生长季草地生态系统的影响究竟是正效应还是负效应, 作用机制如何, 其滞后效应是否随时间尺度而改变(Luo et al., 2001; Melillo et al., 2002), 都是亟待解决的关键科学问题, 需开展深入研究. ...
UPARSE: highly accurate OTU sequences from microbial amplicon reads 1 2013
... 使用TruSeq® DNA PCR-Free Sample Preparation Kit建库试剂盒进行文库构建, 构建好的文库经过Qubit和Q-PCR定量, 文库合格后, 使用HiSeq2500 PE250进行上机测序.根据Barcode序列和PCR扩增引物序列从下机数据中拆分出各样本数据, 对每个样本的reads进行拼接(Magoč & Salzberg, 2011), 得到原始Tags数据(Raw Tags), 原始数据在Quantitative Insights Into Microbial Ecology (QIIME)平台进行过滤(Caporaso et al., 2010)、去除嵌合体序列(Rognes et al., 2016)最终得到有效数据(Effective Tags).对所有样本的有效数据以97%的一致性进行操作分类单元(Operational Taxonomic Units, OTU)聚类, 用Mothur方法与SILVA132 (http://www.arb- silva.de/)(Edgar, 2013)的SSUrRNA数据库(Wang et al., 2007)对OTU序列进行物种注释(设定阈值为0.8-1), 最后获得OTU丰度表. ...
Plant diversity effects on soil food webs are stronger than those of elevated CO2 and N deposition in a long-term grassland experiment 1 2013
... 已有研究大多从生长季或年平均气温变化角度, 阐述气候变暖对草地生态系统影响规律(Knapp & Smith, 2001).冬季作为植被躲避恶劣环境、微生物活动较为频繁的关键期却较少受到关注(Kreyling et al., 2010; Sorensen et al., 2018).此外, 气候变暖存在明显的季节差异, 冬季升温速率远大于生长季(牛书丽等, 2007; IPCC, 2014).显著的冬季增温使北半球大部分地区季节性雪被减少或消失(秦大河, 2014), 导致土壤经受更为广泛和频繁的冻融交替作用(Peng et al., 2010), 严重影响土壤理化性质、养分循环、生物多样性和生态系统生产力(Schuerings et al., 2014).因此, 仅从生长季或全年尺度分析气候变化影响效应, 显然无法全面且准确揭示全球气候变化下草地生态系统变化特征, 系统深入分析冬季气候变化特征(温度及积雪变化)及其对草地生态系统影响机制具有重要研究价值. ...
Variation among biomes in temporal dynamics of aboveground primary production 1 2001
... 已有研究大多从生长季或年平均气温变化角度, 阐述气候变暖对草地生态系统影响规律(Knapp & Smith, 2001).冬季作为植被躲避恶劣环境、微生物活动较为频繁的关键期却较少受到关注(Kreyling et al., 2010; Sorensen et al., 2018).此外, 气候变暖存在明显的季节差异, 冬季升温速率远大于生长季(牛书丽等, 2007; IPCC, 2014).显著的冬季增温使北半球大部分地区季节性雪被减少或消失(秦大河, 2014), 导致土壤经受更为广泛和频繁的冻融交替作用(Peng et al., 2010), 严重影响土壤理化性质、养分循环、生物多样性和生态系统生产力(Schuerings et al., 2014).因此, 仅从生长季或全年尺度分析气候变化影响效应, 显然无法全面且准确揭示全球气候变化下草地生态系统变化特征, 系统深入分析冬季气候变化特征(温度及积雪变化)及其对草地生态系统影响机制具有重要研究价值. ...
Effects of soil freeze-thaw cycles differ between experimental plant communities 3 2010
... 已有研究大多从生长季或年平均气温变化角度, 阐述气候变暖对草地生态系统影响规律(Knapp & Smith, 2001).冬季作为植被躲避恶劣环境、微生物活动较为频繁的关键期却较少受到关注(Kreyling et al., 2010; Sorensen et al., 2018).此外, 气候变暖存在明显的季节差异, 冬季升温速率远大于生长季(牛书丽等, 2007; IPCC, 2014).显著的冬季增温使北半球大部分地区季节性雪被减少或消失(秦大河, 2014), 导致土壤经受更为广泛和频繁的冻融交替作用(Peng et al., 2010), 严重影响土壤理化性质、养分循环、生物多样性和生态系统生产力(Schuerings et al., 2014).因此, 仅从生长季或全年尺度分析气候变化影响效应, 显然无法全面且准确揭示全球气候变化下草地生态系统变化特征, 系统深入分析冬季气候变化特征(温度及积雪变化)及其对草地生态系统影响机制具有重要研究价值. ...
... 土壤微生物和参与碳(C)、氮(N)、磷(P)循环的关键酶活性是衡量土壤质量的重要指标, 是参与土壤有机物质积累、矿化作用及调控土壤养分循环的关键因子, 受生物和非生物因素综合作用(沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016), 并且对环境扰动敏感度较高.大量生长季开展的研究表明, 温度升高可增强微生物和酶活性, 加速土壤有机质分解, 增强土壤呼吸, 对全球变化产生正反馈作用(Bardgett et al., 2008; Yue et al., 2015); 然而, 冬季增温和降雪变化对土壤微生物群落结构功能、关键酶活性及地上生态过程的影响研究相对较少, 且彼此间结论存在较大差异(Guo et al., 2017).有研究指出, 冬季升温可提高土壤酶和微生物活性, 促进有机质的分解和氮矿化速率, 提高土壤养分有效性(Grimm et al., 2013); 但也有报道发现, 增温降低了积雪厚度与持续时间, 积雪绝热保温作用的降低导致土壤冻融交替频发, 导致微生物大量裂解死亡(Schuerings et al., 2014), 进而降低了土壤养分循环速率(Blankinship & Hart, 2012; Li et al., 2016).此外, 降雪对陆地生态系统影响的研究主要集中于北半球高纬度寒区或冻区, 以及中低纬度高海拔地区, 温带干旱半干旱区冬季积雪变化及其影响效应的研究较少(Kreyling et al., 2010).因此, 本研究以黄土高原半干旱草原为研究对象, 探究冬季气候变化(增温和积雪减少)对草地生态系统土壤微生物、酶活性及养分循环影响特征, 不仅能丰富和拓展气候变化对生态系统影响过程与机制的认识, 也对提高草地资源管理能力和水平, 特别是在全球冬季显著增温和降雪减少背景下, 准确预测草地生态系统中长期动态变化具有重要意义. ...
... 全球变化背景下土壤微生物群落组成和多样性、土壤酶活性及碳氮磷等养分变化的响应研究一直是生态学研究的热点问题.近年来, 已有大量控制试验及meta分析文章针对上述科学问题进行了较为系统与全面的阐述(Bai et al., 2013; Romero- Olivares et al., 2017; Xiao et al., 2018), 但以往研究大都侧重于全年增温或生长季增温的生态效应探索(Guo et al., 2017), 如Bai等(2013)汇总分析了全球陆地生态系统氮循环对气候变化响应的528个观测数据, 发现生长季和全年增温实验数量占全部样本的85.4%, 冬季增温实验不仅个案数量少, 其测定指标更为匮乏.然而, 大量研究指出冬季土壤微生物活动仍较为频繁, 养分物质循环及能量代谢依旧进行(Kreyling et al., 2010; Sorensen et al., 2018), 显著的冬季气候变化(增温及降雪减少)不仅影响植被休眠期内土壤微生物和养分循环, 对生长季植被生长发育及群落构建、稳定性等亦具有持续影响(Hudson & Henry, 2009).因此, 探究冬季温度及积雪变化下土壤理化性质, 以及微生物群落组成、多样性的变化规律, 对丰富气候变化的生态效应研究内涵、准确预测未来气候情景下生态系统响应规律, 进而促进植被资源管理能力提升具有重要意义. ...
Repeated freeze- thaw cycles and their effects on biological processes in two arctic ecosystem types 1 2002
... 冻融频发导致土壤微生物大量死亡的报道较多(Larsen et al., 2002; Sulkava & Huhta, 2003; Sjursen et al., 2005), 微生物死亡可导致微生物生物量和微生物多样性下降(图6), 此假设在本研究中也得到了证实: 互作处理下细菌种类最少, 多样性下降最显著, 其3种多样性指数分别下降了5.9%、10.5%和8.7% (图3); 门分类水平上, 细菌群落组成的前4门细菌, 除变形菌门相对丰度在不同处理间未发现显著差异外, 增温、减雪及互作处理下其余3种细菌(酸杆菌门、放线菌门、芽单胞菌门)相对丰度均显著低于对照(图4A).微生物大量死亡及其多样性的下降, 会造成微生物生物量下降.本研究中互作处理下, 土壤MBC含量显著降低, 这与Freppaz等(2007)的结果一致, 后者发现一半以上的微生物可在第一个冻融周期中被杀死, 进而导致微生物生物量迅速减少.冻融加剧导致土壤MBN含量下降亦见诸报道, 如Song等(2017)发现冻融过程使土壤MBN含量下降12.2%, 本研究中MBN含量有所下降, 但并不显著(表2), 推测可能与微生物组成、抗逆性及恢复力不同有关(Tierney et al., 2003; Schmidt et al., 2007). ...
Response of terrestrial nitrogen dynamics to snow cover change: a meta-analysis of experimental manipulation 1 2016
... 土壤微生物和参与碳(C)、氮(N)、磷(P)循环的关键酶活性是衡量土壤质量的重要指标, 是参与土壤有机物质积累、矿化作用及调控土壤养分循环的关键因子, 受生物和非生物因素综合作用(沈菊培和贺纪正, 2011; Delgado-Baquerizo et al., 2016), 并且对环境扰动敏感度较高.大量生长季开展的研究表明, 温度升高可增强微生物和酶活性, 加速土壤有机质分解, 增强土壤呼吸, 对全球变化产生正反馈作用(Bardgett et al., 2008; Yue et al., 2015); 然而, 冬季增温和降雪变化对土壤微生物群落结构功能、关键酶活性及地上生态过程的影响研究相对较少, 且彼此间结论存在较大差异(Guo et al., 2017).有研究指出, 冬季升温可提高土壤酶和微生物活性, 促进有机质的分解和氮矿化速率, 提高土壤养分有效性(Grimm et al., 2013); 但也有报道发现, 增温降低了积雪厚度与持续时间, 积雪绝热保温作用的降低导致土壤冻融交替频发, 导致微生物大量裂解死亡(Schuerings et al., 2014), 进而降低了土壤养分循环速率(Blankinship & Hart, 2012; Li et al., 2016).此外, 降雪对陆地生态系统影响的研究主要集中于北半球高纬度寒区或冻区, 以及中低纬度高海拔地区, 温带干旱半干旱区冬季积雪变化及其影响效应的研究较少(Kreyling et al., 2010).因此, 本研究以黄土高原半干旱草原为研究对象, 探究冬季气候变化(增温和积雪减少)对草地生态系统土壤微生物、酶活性及养分循环影响特征, 不仅能丰富和拓展气候变化对生态系统影响过程与机制的认识, 也对提高草地资源管理能力和水平, 特别是在全球冬季显著增温和降雪减少背景下, 准确预测草地生态系统中长期动态变化具有重要意义. ...
Functional screening of abundant bacteria from acidic forest soil indicates the metabolic potential of Acidobacteria subdivision 1 for polysaccharide decomposition 1 2016
... 土壤酶的产生大多来自动植物和微生物的分泌物, 酶促反应可以催化土壤内部多种化学元素的转化及生理过程(王楠等, 2014).土壤酶活性受多种因素的影响, 例如土壤温湿度、养分可利用性、微生物生物量和群落组成及环境因子等(Weintraub et al., 2013).本研究中BG、NAG、AKP 3种胞外酶活性在增温和减雪互作的处理下显著下降(表2),究其原因, 一方面土壤冻融频发导致微生物大量死亡, 分泌胞外酶含量随之减少, 从而直接或间接影响土壤酶活性(Torres et al., 2015; López-Aizpún et al., 2018)(图6).另一方面, 增温、减雪和互作处理下土壤湿度下降(图2B)也可能是造成酶催化反应效率下降的原因.Chaer等(2009)对美国安德鲁实验森林土壤分析表明, β-葡萄糖苷酶活性在1、2、4和8个冻融循环之后降低; DeForest等(2012)发现磷酸酶活性在未冻结土壤中比冻结土壤中高两倍, 冻结环境会降低磷酸酶活性, 也与本研究结果一致.然而, 微生物群落组成与分泌酶种类、含量及其驱动的土壤碳氮磷循环过程和机制研究, 目前仍较为匮乏.已有研究表明, 酸杆菌可产生β-葡萄糖苷酶、纤维二糖水解酶及NAG等碳氮分解关键酶(Lladó et al., 2016), 放线菌在有机质周转、碳循环中也具有重要作用(Chinnadurai et al., 2014).酸杆菌和放线菌作为研究区土壤中重要菌门, 在互作处理下相对丰度下降, 可导致土壤碳氮循环相关酶活性的降低, 也可以部分解释土壤中有机质积累及NO- 3-N含量的下降(表2), 但其具体作用机制仍不清楚. ...
Atmospheric ammonia concentration modulates soil enzyme and microbial activity in an oak forest affecting soil microbial biomass 1 2018
... 土壤酶的产生大多来自动植物和微生物的分泌物, 酶促反应可以催化土壤内部多种化学元素的转化及生理过程(王楠等, 2014).土壤酶活性受多种因素的影响, 例如土壤温湿度、养分可利用性、微生物生物量和群落组成及环境因子等(Weintraub et al., 2013).本研究中BG、NAG、AKP 3种胞外酶活性在增温和减雪互作的处理下显著下降(表2),究其原因, 一方面土壤冻融频发导致微生物大量死亡, 分泌胞外酶含量随之减少, 从而直接或间接影响土壤酶活性(Torres et al., 2015; López-Aizpún et al., 2018)(图6).另一方面, 增温、减雪和互作处理下土壤湿度下降(图2B)也可能是造成酶催化反应效率下降的原因.Chaer等(2009)对美国安德鲁实验森林土壤分析表明, β-葡萄糖苷酶活性在1、2、4和8个冻融循环之后降低; DeForest等(2012)发现磷酸酶活性在未冻结土壤中比冻结土壤中高两倍, 冻结环境会降低磷酸酶活性, 也与本研究结果一致.然而, 微生物群落组成与分泌酶种类、含量及其驱动的土壤碳氮磷循环过程和机制研究, 目前仍较为匮乏.已有研究表明, 酸杆菌可产生β-葡萄糖苷酶、纤维二糖水解酶及NAG等碳氮分解关键酶(Lladó et al., 2016), 放线菌在有机质周转、碳循环中也具有重要作用(Chinnadurai et al., 2014).酸杆菌和放线菌作为研究区土壤中重要菌门, 在互作处理下相对丰度下降, 可导致土壤碳氮循环相关酶活性的降低, 也可以部分解释土壤中有机质积累及NO- 3-N含量的下降(表2), 但其具体作用机制仍不清楚. ...
Acclimatization of soil respiration to warming in a tall grass prairie 1 2001
... 冬季温度和降雪变化不仅直接影响冬季土壤养分循环、微生物多样性及关键酶活性, 其滞后效应还会对生长季生态过程和功能产生持续影响(Durán et al., 2014).如本研究中, 冬季增温、减雪及其互作一方面提高了土壤养分含量(图6), 对生长季植物生长具有一定促进作用; 但另一方面, 冬季增温减雪引致的土壤频繁冻融又降低了微生物多样性、丰度及胞外酶活性, 减缓了有机物质的循环周转, 对生长季植物生长具有抑制效应.冬季气候变化(增温及降雪减少)对生长季草地生态系统的影响究竟是正效应还是负效应, 作用机制如何, 其滞后效应是否随时间尺度而改变(Luo et al., 2001; Melillo et al., 2002), 都是亟待解决的关键科学问题, 需开展深入研究. ...
FLASH: fast length adjustment of short reads to improve genome assemblies 1 2011
... 使用TruSeq® DNA PCR-Free Sample Preparation Kit建库试剂盒进行文库构建, 构建好的文库经过Qubit和Q-PCR定量, 文库合格后, 使用HiSeq2500 PE250进行上机测序.根据Barcode序列和PCR扩增引物序列从下机数据中拆分出各样本数据, 对每个样本的reads进行拼接(Magoč & Salzberg, 2011), 得到原始Tags数据(Raw Tags), 原始数据在Quantitative Insights Into Microbial Ecology (QIIME)平台进行过滤(Caporaso et al., 2010)、去除嵌合体序列(Rognes et al., 2016)最终得到有效数据(Effective Tags).对所有样本的有效数据以97%的一致性进行操作分类单元(Operational Taxonomic Units, OTU)聚类, 用Mothur方法与SILVA132 (http://www.arb- silva.de/)(Edgar, 2013)的SSUrRNA数据库(Wang et al., 2007)对OTU序列进行物种注释(设定阈值为0.8-1), 最后获得OTU丰度表. ...
A microplate fluorimetric assay for the study of enzyme diversity in soils 1 2001
... 采用重铬酸盐氧化法测定样品中土壤有机碳(SOC)含量, 凯式定氮仪(Kjeltec 8400, FOSS Corporation, Hillerod, Denmark)测定总氮(TN)含量.速效磷(AP)含量用0.5 mol·L-1 NaHCO3浸提-钼锑抗比色法测定.铵态氮(NH+ 4-N)和硝态氮(NO- 3-N)含量利用流动分析仪(Alpkem, OI Analytical, College Station, USA)测定.土壤pH用酸度计进行分析.微生物生物量碳(MBC)和氮(MBN)含量采用氯仿熏蒸-K2SO4浸提法测定(Vance et al., 1987).选取参与C、N、P循环的3种关键酶, 即β-1,4-葡萄糖苷酶(BG), β-1,4-N-乙酰氨基葡萄糖苷酶(NAG)和碱性磷酸酶(AKP), 使用微孔板荧光法检测其活性(Marx et al., 2001).以上土壤样品理化性质详细测定步骤参考《土壤农化分析》(鲍士旦, 2000). ...
Soil warming and carbon-cycle feedbacks to the climate system 1 2002
... 冬季温度和降雪变化不仅直接影响冬季土壤养分循环、微生物多样性及关键酶活性, 其滞后效应还会对生长季生态过程和功能产生持续影响(Durán et al., 2014).如本研究中, 冬季增温、减雪及其互作一方面提高了土壤养分含量(图6), 对生长季植物生长具有一定促进作用; 但另一方面, 冬季增温减雪引致的土壤频繁冻融又降低了微生物多样性、丰度及胞外酶活性, 减缓了有机物质的循环周转, 对生长季植物生长具有抑制效应.冬季气候变化(增温及降雪减少)对生长季草地生态系统的影响究竟是正效应还是负效应, 作用机制如何, 其滞后效应是否随时间尺度而改变(Luo et al., 2001; Melillo et al., 2002), 都是亟待解决的关键科学问题, 需开展深入研究. ...
 ,4,6,*
,4,6,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT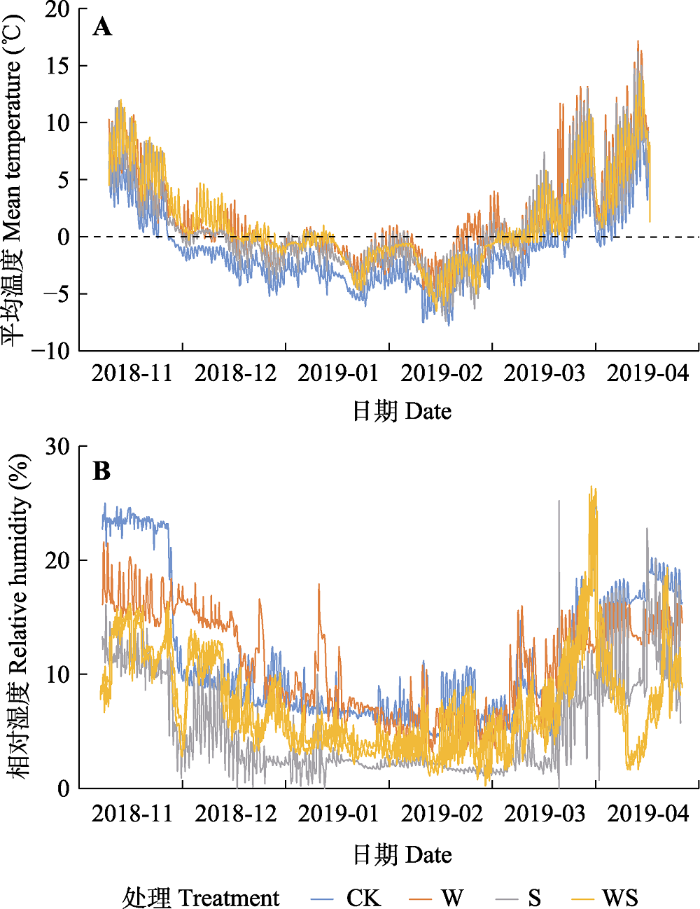 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT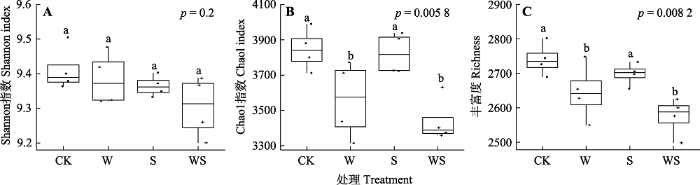 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT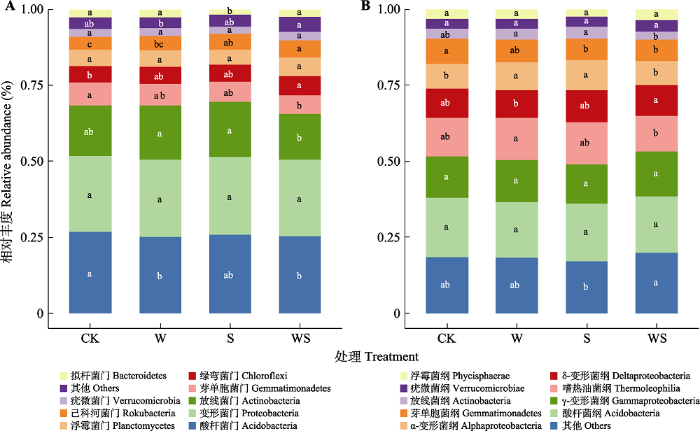 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT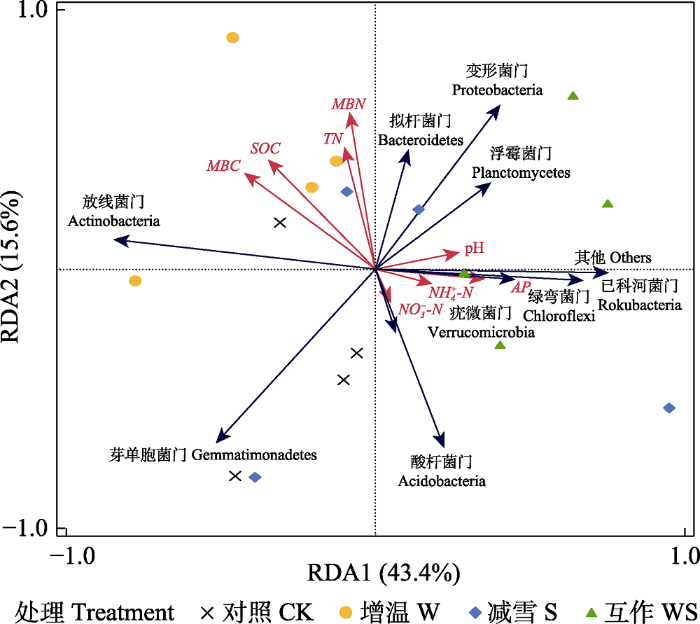 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT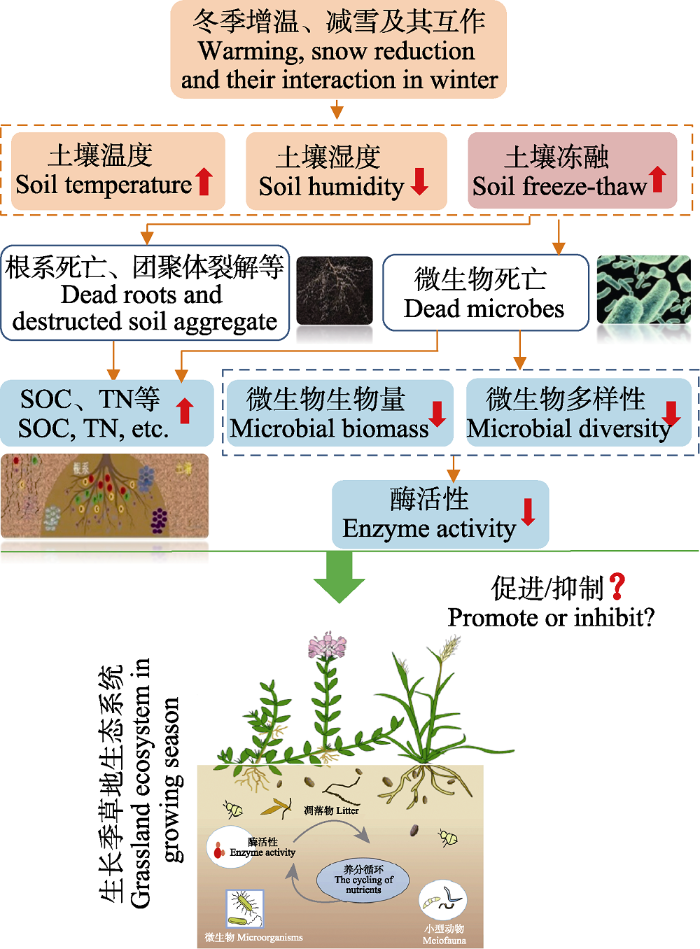 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}