Effects of arbuscular mycorrhizal fungi and nitrogen addition on nitrogen uptake of rice genotypes with different root morphologies
Ju-Feng MA1,2, Min XIN1,2, Chen-Chao XU1,2, Wan-Ying ZHU1,2, Chuan-Zao MAO2, Xin CHEN2, Lei CHENG,1,2,*1MOE Key Laboratory of Biosystems Homeostasis & Protection, Zhejiang University, Hangzhou 310058, China 2College of Life Sciences, Zhejiang University, Hangzhou 310058, China
National Natural Science Foundation of China(91951107) National Natural Science Foundation of China(32025024) Fundamental Research Funds for the Central Universities(K20200026)
Abstract Aims Plants absorb mineral nutrients such as nitrogen (N) mainly through their roots. The nutrient uptake of plants with different root morphologies differs. Many studies have shown that arbuscular mycorrhizal fungi (AMF) can help their symbiotic associates absorb mineral N. However, there is little research on whether the effect of AMF on nutrient uptake of plant roots is affected by root morphology. Methods In this study, we selected three rice mutants and one wild type (root hairless (rhl1), lateral rootless (iaa11), adventitious rootless (arl1) and wild type (Kas)) to investigate the role of root morphology in plant nutrient uptake. Subsequently, we used the 15N isotope labeling method to explore the effects of arbuscular mycorrhizal fungi and N addition (low N: 20 mg·kg-1 NH4+-N; high N: 100 mg·kg-1 NH4+-N) on N uptake of rice mutants with different root morphologies. Important findings The results show that the leaf 15N concentrations of rhl1,Kas, iaa11 and arl1 were increased by 60%, 72%, 128% and 118%, respectively, under the high N compared to the low N treatment. This result indicates that the addition of N significantly promoted rice N uptake with the most evident effect occurring in iaa11 and arl1. The average effect sizes of AMF on rhl1, Kas, iaa11 and arl1 were 17%, 31%, 42% and 51% under the low N level, indicating that root morphology can alter the effect of AMF on plant N uptake. Compared to the low N treatment, high N significantly downregulated the AMF effect on N uptake by rice plants with different root morphologies, indicating that N addition may mediate the complementary effect of AMF and root morphology on plant nutrient uptake. In conclusion, our data provide direct experimental evidence of funcitonal complementarity of mycrrohzal fungi and their associated roots with different root morphogy. Keywords:arbuscular mycorrhizal fungi;nitrogen addition;root morphology;nitrogen uptake;functional complementarity
PDF (3108KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 马炬峰, 辛敏, 徐陈超, 祝琬莹, 毛传澡, 陈欣, 程磊. 丛枝菌根真菌与氮添加对不同根形态基因型水稻氮吸收的影响. 植物生态学报, 2021, 45(7): 728-737. DOI: 10.17521/cjpe.2021.0155 MA Ju-Feng, XIN Min, XU Chen-Chao, ZHU Wan-Ying, MAO Chuan-Zao, CHEN Xin, CHENG Lei. Effects of arbuscular mycorrhizal fungi and nitrogen addition on nitrogen uptake of rice genotypes with different root morphologies. Chinese Journal of Plant Ecology, 2021, 45(7): 728-737. DOI: 10.17521/cjpe.2021.0155
植物在生长的过程中需要从土壤中获取矿质养分, 比如氮。为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020)。植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系。养分竞争型根系根径细、根分支密度高、根毛密集。这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019)。而保守型根系根径粗、根分支密度小、根毛稀疏。这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019)。此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮。AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015)。研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005)。精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011)。相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003)。不同根形态植物对养分吸收存在较大差异。但是, AMF如何影响不同根形态植物的养分吸收仍不清楚。
新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016)。由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略。Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收。Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大。此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017)。这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态。另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生。研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021)。这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚。对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少。
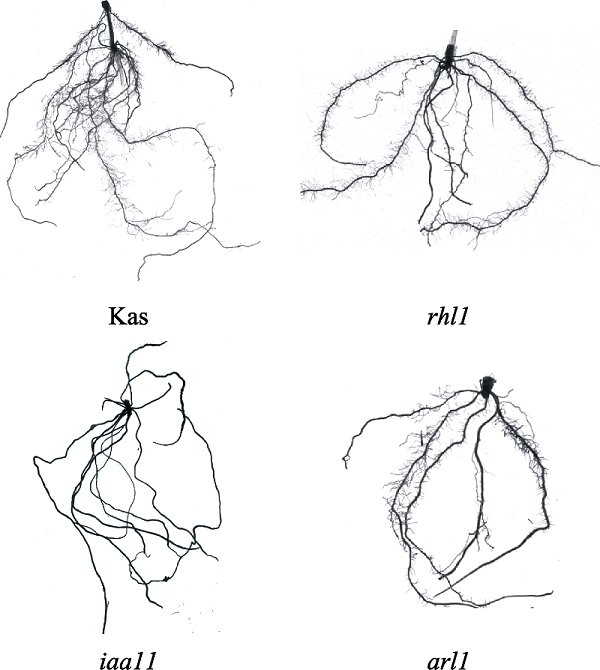
新窗口打开|下载原图ZIP|生成PPT 图1不同根形态基因型水稻根系扫描图。材料来自种植7周的水稻。arl1, 不定根缺失突变体水稻; iaa11, 侧根缺陷型突变体水稻; Kas, 野生型水稻; rhl1, 根毛缺陷型突变体水稻。
Fig. 1Root scanning images of rice with different genotypes. The material was obtained from rice was planted for 7 weeks. arl1, adventitious rootless mutant; iaa11, lateral rootless mutant; Kas, wild type; rhl1, root hairless mutant.
新窗口打开|下载原图ZIP|生成PPT 图2不同根形态基因型水稻根系不定根数目、平均根径、总根长和根尖数(平均值±标准误, n = 10)。arl1, 不定根缺失突变体水稻; iaa11, 侧根缺陷型突变体水稻; Kas, 野生型水稻; rhl1, 根毛缺陷型突变体水稻。柱形图上方不同小写字母表示处理间差异显著(p < 0.05)。
Fig. 2Number of adventitious roots, root diameter, root length and root tips of rice with different genotypes (mean ± SE, n = 10). arl1, adventitious rootless mutant; iaa11, lateral rootless mutant; Kas, wild type; rhl1, root hairless mutant. Different lowercase letters above the columns represent significant differences between treatments according to least significant difference test (p < 0.05).
2.2 氮处理对不同基因型水稻氮吸收的影响
为了探究不同土壤氮浓度对不同基因型水稻氮吸收的影响, 实验二设置了两种不同的氮养分条件, 并且测定了水稻茎叶含氮量。陈晨等(2016)研究发现茎叶的累积氮含量要高于根的累积氮含量。因此, 本实验通过测定水稻地上部分15N浓度来表征植物根系对矿质氮的吸收能力(Awaydul et al., 2019)。结果发现, 氮处理对水稻氮吸收有显著影响(p < 0.001),不同基因型水稻对氮吸收也有显著影响(p < 0.05), 但两者的交互作用对氮吸收的影响并不显著(图3, p > 0.05)。
图3
新窗口打开|下载原图ZIP|生成PPT 图3不同氮处理对不同根形态基因型水稻地上部分15N浓度的影响(平均值±标准误, n = 5)。arl1, 不定根缺失突变体水稻; iaa11, 侧根缺陷型突变体水稻; Kas, 野生型水稻; rhl1, 根毛缺陷型突变体水稻。柱形图上方不同小写字母表示处理间差异显著( p < 0.05)。neu, 氮处理效应; st, 基因型效应; neu × st, 氮处理与基因型交互效应。ns, 没有显著差异; *, p < 0.05; ***, p < 0.001。
Fig. 3Effect of different nitrogen treatments on 15N concentrations of rice shoots with different genotypes (mean ± SE, n = 10). arl1, adventitious rootless mutant; iaa11, lateral rootless mutant; Kas, wild type; rhl1, root hairless mutant. Different lowercase letters above the columns represent significant differences between treatments according to least significant difference test (p < 0.05). neu, nitrogen treatment effect; st, genotype effect; neu × st, interaction effect between nitrogen treatment and genotype. ns, not significant; *, p < 0.05; ***, p < 0.001.
在低氮水平下, Kas与rhl1地上部分15N浓度显著高于iaa11与arl1 (图3, p < 0.05), 表明侧根与不定根的缺失对水稻根系养分吸收功能的影响较大, 而根毛缺失对水稻根系养分吸收功能的影响较小。高氮水平下, rhl1、Kas、iaa11与arl1的茎叶15N浓度分别提高了60%、72%、128%与118% (图3, p < 0.05), 表明高氮水平下, 水稻对氮的吸收显著增强, 并且iaa11与arl1对氮添加的响应更强烈。但是, 在高氮水平下, 4种水稻地上部分15N浓度并没有显著差异(图3, p > 0.05), 表明环境养分浓度升高能抵消侧根与不定根缺失对水稻氮吸收的抑制效应。
Table 1 表1 表1不同氮养分条件下不同基因型水稻菌根侵染率(平均值±标准误, n = 5) Table 1Mycorrhizal colonization rate of rice with different genotypes under different nitrogen nutrient conditions (mean ± SE, n = 5)
氮处理 Nitrogen treatment
AMF处理 AMF treatment
菌根侵染率 Mycorrhizal colonization rate (%)
Kas
rhl1
iaa11
arl1
低氮 Low nitrogen
Non-AMF
0
0
0
0
AMF
40 ± 4b
50 ± 2ab
62 ± 4a
43 ± 1b
高氮 High nitrogen
Non-AMF
0
0
0
0
AMF
45 ± 3ab
49 ± 3ab
55 ± 4a
40 ± 3b
arl1, 不定根缺失突变体水稻; iaa11, 侧根缺陷型突变体水稻; Kas, 野生型水稻; rhl1, 根毛缺陷型突变体水稻。每行中不同小写字母表示差异显著(p < 0.05)。 arl1, adventitious rootless mutant; iaa11, lateral rootless mutant; Kas, wild type; rhl1, root hairless mutant. Different lowercase letters within each line mean significant difference (p < 0.05).
低氮处理下, 与对照相比, Kas、iaa11和arl1接种AMF后茎叶15N浓度显著提高(图4A, p < 0.05), 其中AMF对于氮吸收能力较弱的arl1与iaa11氮吸收的促进效应比其他两种基因型水稻更加明显(图5), 表明根系养分吸收能力弱的植物更依赖于AMF吸收土壤养分。相较于低氮水平, 高氮水平下, 4种基因型水稻对AMF的响应均显著降低(图5, p < 0.05), 表明养分浓度的增加改善了植物养分吸收的环境, 植物依靠自身根系便可吸收足够养分, 故而AMF对植物的养分吸收促进作用逐渐减弱。另外,在高氮处理下, 只有iaa11接种AMF后茎叶15N浓度显著提高(图4B, p < 0.05), arl1与iaa11对AMF的响应仍显著高于其他2种基因型水稻(图5, p < 0.05), 表明侧根的缺失对水稻氮吸收存在较大的影响, 即使在高氮水平下仍需与大量AMF共生来提高养分吸收。
图4
新窗口打开|下载原图ZIP|生成PPT 图4不同氮处理下菌根真菌(AMF)处理对不同基因型水稻氮吸收的影响(平均值±标准误, n = 5)。柱形图上方不同小写字母表示处理间差异显著(p < 0.05)。amf, AMF处理效应; st, 基因型效应; amf × st, AMF处理与基因型交互效应。ns, 没有显著差异; *, p < 0.05, ***, p < 0.001。
Fig. 4Effect of arbuscular mycorrhizal fungi (AMF) on nitrogen uptake in rice with different genotypes under different nitrogen treatments (mean ± SE, n = 5). Different lowercase letters above the columns represent significant differences between treatments according to least significant difference test (p < 0.05). amf, AMF treatment effect; st, genotype effect; amf × st, interaction effect between AMF treatment and genotype, there was no significant difference. ns, not significant; *, p < 0.05; ***, p < 0.001.
图5
新窗口打开|下载原图ZIP|生成PPT 图5不同氮处理下菌根真菌(AMF)处理对不同基因型水稻氮吸收的平均效应值(平均值±标准误, n = 5)。平均效应值: AMF处理下地上部分15N浓度变化百分量。柱形图上方不同小写字母表示处理间差异显著(p < 0.05)。neu, 氮处理效应; st, 基因型效应; neu × st, 氮处理与基因型交互效应。ns, 没有显著差异; **, p < 0.01, ***, p < 0.001。
Fig. 5Effect size of arbuscular mycorrhizal fungi (AMF) treatment on nitrogen uptake in rice with different genotypes under different nitrogen treatments (mean ± SE, n = 5). Effect size: proportion of change of 15N concentration of rice shoots by AMF treatment. Different lowercase letters above the columns represent significant differences between treatments according to least significant difference test (p < 0.05). neu, nitrogen treatment effect; st, genotype effect; neu × st, interaction effect between nitrogen treatment and genotype, there was no significant difference. ns, not significant; **, p < 0.01; ***, p < 0.001.
此外, 本研究还发现侧根缺陷突变株水稻根系的养分吸收能力显著小于根毛缺陷突变株水稻。该结果与Paszkowski和Boller (2002)对不同根形态玉米突变株的研究一致, 两者都证明了侧根对于植物养分吸收的作用更大。根系分枝密度与植物养分吸收呈正相关关系, 是反映养分吸收能力的重要指标(陈晨等, 2016), 而侧根的缺失会导致植物根系的根分枝密度显著减小, 因此, 侧根突变株iaa11养分吸收能力比其他基因型水稻更弱。本研究使用水稻地上部分15N浓度反映水稻根系氮吸收的情况, 而缺少水稻根系与水稻整体氮含量数据。陈晨等(2016)的研究发现水稻整株氮含量、茎叶氮含量、根系氮含量的均值分别为4.22%、4.42%、3.75%, 说明水稻地上部分茎叶氮含量与水稻整株氮含量更接近, 水稻地上部分氮含量也能够反映水稻的氮吸收情况。与此同时, 前人也有很多研究通过测定植物地上部分氮含量来反映植物对氮吸收的情况(Wang et al., 2018a; Awaydul et al., 2019; Tateno et al., 2020)。
另一方面, 本研究发现不同基因型水稻对氮处理的响应不同, 该结果与宋智勇等(2012)的研究一致。不同基因型水稻对氮的敏感度存在一定差异, 不同氮敏感度水稻对氮浓度增加的响应不同。同时, 本研究发现除了iaa11与arl1外, 其他基因型水稻的菌根侵染率没有显著差异, 而之前的研究表明根形态对菌根侵染率存在影响(Ma et al., 2018)。本研究每盆水稻接种约1 000个孢子, 充足的AMF密度使得根被侵染的概率增大(Ohtomo et al., 2018), 进而导致不同根形态基因型水稻菌根侵染率差异减小。然而, 本研究只在两个氮浓度水平下研究了氮处理对不同基因型水稻氮吸收的影响, 想要更具体地分析4种基因型水稻对氮添加的响应有何不同, 需要设计多种养分处理。
3.2 不同氮水平处理下4种基因型水稻对AMF的响应特征
前人的研究已证实了在养分匮乏的环境下植物更依赖于AMF吸收养分(Corkidi et al., 2002; Bakhshandeh et al., 2017; Lu et al., 2020), 本文的研究结果与之一致, 共同证明了AMF对植物养分吸收的促进效应受土壤养分影响。在养分匮乏的条件下, 植物的生长更受养分吸收的限制。因此, 为了获取更多的养分, 植物需要通过与AMF共生获取更多的养分(Bonneau et al., 2013); 而在土壤养分充足的情况下, 植物依靠根系便可吸收足够养分, 对AMF的依赖性减弱(Schalamuk et al., 2011)。
相对于菌根植物, 根系与根外菌丝都具有养分吸收的功能, 为了避免养分吸收功能的冗余, 植物根系与AMF在养分吸收上往往存在功能互补的关系(Koide, 2000; Chen et al., 2016; Cheng et al., 2016)。本文研究结果也验证了这一观点, 养分竞争型根系植物具有较强的养分吸收能力, 因此, 主要依赖自身根系进行养分吸收。相反, 保守型根系植物养分吸收能力较弱, 故主要通过AMF进行养分吸收(Liu et al., 2015)。平均根径较粗的侧根缺陷突变体水稻对AMF的依赖性与菌根侵染率要高于其他水稻。在自然界中, 土壤养分的有效性在空间与时间上存在高度异质性(王庆成和程云环, 2004), 因此, 增殖快、周转速率高的细根更适宜于这样的养分环境; 而增殖慢、寿命长的粗根为了提高自身的养分吸收能力选择与AMF共生。同时, 由于粗根寿命长的特点, AMF也更倾向于与粗根共生(Ma et al., 2018)。本研究结果说明不同根形态基因型水稻与AMF在氮吸收方面存在互补关系, 该结论支持植物根系与AMF在养分吸收上存在功能互补的理论框架。然而, 本研究没有测定土壤中菌丝体的15N浓度, 故无法在根外菌丝氮吸收方面提供更直接的证据, 还需要在下一步研究中加以验证。
AwaydulA, ZhuWY, YuanYG, XiaoJ, HuH, ChenX, KoideRT, ChengL (2019). Common mycorrhizal networks influence the distribution of mineral nutrients between an invasive plant, Solidago canadensis, and a native plant, Kummerowa striata Mycorrhiza, 29, 29-38. DOI:10.1007/s00572-018-0873-5PMID:30421153 [本文引用: 4] Invasive species often reduce ecosystem services and lead to a serious threat to native biodiversity. Roots of invasive plants are often linked to roots of native plants by common mycorrhizal networks (CMNs) of arbuscular mycorrhizal (AM) fungi, but whether and how CMNs mediate interactions between invasive and native plant species remains largely uninvestigated. We conducted two microcosm experiments, one in which we amended the soil with mineral N and another in which we amended the soil with mineral P. In each experiment, we grew a pair of test plants consisting of Kummerowia striata (native to our research site) and Solidago canadensis (an invasive species). CMNs were established between the plants, and these were either left intact or severed. Intact CMNs increased growth and nutrient acquisition by S. canadensis while they decreased nutrient acquisition by K. striata in comparison with severed CMNs. N and P analyses indicated that compared to severed CMNs, intact CMNs preferentially transferred mineral nutrients to S. canadensis. CMNs produced by different species of AM fungi had slightly different effects on the interaction between these two plant species. These results highlight the role of CMNs in the understanding of interactions between the invasive species S. canadensis and its native neighbor.
BaiJJ, PiaoZZ, ZengW, LiGX, YangRF (2019). Effects of different lateral root densities on growth, development and main agronomic characters of rice Molecular Plant Breeding, 17, 1624-1630. [本文引用: 1]
BakhshandehS, CorneoPE, MariotteP, KerteszMA, DijkstraFA (2017). Effect of crop rotation on mycorrhizal colonization and wheat yield under different fertilizer treatments Agriculture, Ecosystems & Environment, 247, 130-136. DOI:10.1016/j.agee.2017.06.027URL [本文引用: 1]
BatesTR, LynchJP (2001). Root hairs confer a competitive advantage under low phosphorus availability Plant and Soil, 236, 243-250. DOI:10.1023/A:1012791706800URL [本文引用: 1]
BonneauL, HuguetS, WipfD, PaulyN, TruongHN (2013). Combined phosphate and nitrogen limitation generates a nutrient stress transcriptome favorable for arbuscular mycorrhizal symbiosis in Medicago truncatula New Phytologist, 199, 188-202. DOI:10.1111/nph.2013.199.issue-1URL [本文引用: 2]
BrundrettMC (2002). Coevolution of roots and mycorrhizas of land plants New Phytologist, 154, 275-304. DOI:10.1046/j.1469-8137.2002.00397.xPMID:33873429 [本文引用: 1] Here, the coevolution of mycorrhizal fungi and roots is assessed in the light of evidence now available, from palaeobotanical and morphological studies and the analysis of DNA-based phylogenies. The first bryophyte-like land plants, in the early Devonian (400 million years ago), had endophytic associations resembling vesicular-arbuscular mycorrhizas (VAM) even before roots evolved. Mycorrhizal evolution would have progressed from endophytic hyphae towards balanced associations where partners were interdependent due to the exchange of limiting energy and nutrient resources. Most mycorrhizas are mutualistic, but in some cases the trend for increasing plant control of fungi culminates in the exploitative mycorrhizas of achlorophyllous, mycoheterotrophic plants. Ectomycorrhizal, ericoid and orchid mycorrhizas, as well as nonmycorrhizal roots, evolved during the period of rapid angiosperm radiation in the Cretaceous. It is hypothesised that roots gradually evolved from rhizomes to provide more suitable habitats for mycorrhizal fungi and provide plants with complex branching and leaves with water and nutrients. Selection pressures have caused the morphological divergence of roots with different types of mycorrizas. Root cortex thickness and exodermis suberization are greatest in obllgately mycorrhizal plants, while nonmycorrhizal plants tend to have fine roots, with more roots hairs and relatively advanced chemical defences. Major coevolutionary trends and the relative success of plants with different root types are discussed. Contents Summary 275 I. Introduction 276 II. Mycorrhizal Fungi 276 III. The Dawn of Mycorrhizas 279 IV. Mycorrhizal Associations of Living and Extinct Plants 282 V. Evolution of Roots 288 VI. The Root as a Habitat for Fungi 290 VII. Mycorrhizal Evolution Trends 295 Acknowledgements 298 References 298.
ChenC, GongHQ, ZhangJZ, XuYJ, GaoHJ (2016). Evaluation of nitrogen nutrition characteristics of different rice cultivars at seedling stage Chinese Journal of Eco- Agriculture, 24, 1347-1355. [本文引用: 5]
ChenWL, KoideRT, AdamsTS, DeForestJL, ChengL, EissenstatDM (2016). Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees Proceedings of the National Academy of Sciences of the United States of America, 113, 8741-8746. [本文引用: 1]
ChengL, BookerFL, TuC, BurkeyKO, ZhouLS, ShewHD, RuftyTW, HuSJ (2012). Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2 Science, 337, 1084-1087. DOI:10.1126/science.1224304PMID:22936776 [本文引用: 1] The extent to which terrestrial ecosystems can sequester carbon to mitigate climate change is a matter of debate. The stimulation of arbuscular mycorrhizal fungi (AMF) by elevated atmospheric carbon dioxide (CO(2)) has been assumed to be a major mechanism facilitating soil carbon sequestration by increasing carbon inputs to soil and by protecting organic carbon from decomposition via aggregation. We present evidence from four independent microcosm and field experiments demonstrating that CO(2) enhancement of AMF results in considerable soil carbon losses. Our findings challenge the assumption that AMF protect against degradation of organic carbon in soil and raise questions about the current prediction of terrestrial ecosystem carbon balance under future climate-change scenarios.
CorkidiL, RowlandDL, JohnsonNC, AllenEB (2002). Nitrogen fertilization alters the functioning of arbuscular mycorrhizas at two semiarid grasslands Plant and Soil, 240, 299-310. [本文引用: 1]
CoutinhoBG, MeversE, SchaeferAL, PelletierDA, HarwoodCS, ClardyJ, GreenbergEP (2018). A plant-responsive bacterial-signaling system senses an ethanolamine derivative Proceedings of the National Academy of Sciences of the United States of America, 115, 9785-9790. [本文引用: 1]
DingWN (2009). Cloning and Functional Analysis of OsRHL1 Controlling Root Hair Development in Rice (Orzya sativa). PhD dissertation, Zhejiang University, Hangzhou. 10-15. [本文引用: 1]
GenevaMP, StanchevaIV, BoychinovaMM, MinchevaNH, YonovaPA (2010). Effects of foliar fertilization and arbuscular mycorrhizal colonization on Salvia officinalis L. growth, antioxidant capacity, and essential oil composition Journal of the Science of Food and Agriculture, 90, 696-702. DOI:10.1002/jsfa.3871PMID:20355101 [本文引用: 1] The effect of foliar fertilization and Glomus intraradices inoculation on the growth, qualitative and quantitative pattern of essential oil in Salvia officinalis was determined. Sage plants were grown in a glass house on a soil/sand mixture (w/w = 3:1). Agroleaf total, N:P:K = 20:20:20 + microelements, was used at the whole vegetative growth stage as a 0.3% solution. Inoculation with Glomus intraradices was done at the sowing stage.Application of foliar fertilization and/or mycorrhizal colonization improved dry biomass accumulation and increased the content of antioxidant metabolites (ascorbate and reduced glutathione). Applied treatments lowered the activities of the antioxidants enzymes catalase, ascorbate peroxidase and superoxide dismutase, while guaiacol peroxidase increased. The relative quantity of essential oil pattern was also altered as a result of the applied treatments. Combined application (FF + Gi) significantly promoted 1,8-cineole and alpha-thujone, mycorrhizal colonization enhanced bornyl acetate, 1,8-cineole, alpha- and beta-thujones, while foliar fertilization increased bornyl acetate and camphor. The favorable effect of root colonization by Glomus intraradices was determined both on quantitative and qualitative pattern of sage essential oil.We conclude that inoculation with Glomus intraradices resulted in improved essential oil yield and quality, while combined application of foliar fertilizer and mycorrhizal fungi predominantly enhanced shoot biomass accumulation.
GiehlRFH, GruberBD, vonWirén N (2014). It’s time to make changes: modulation of root system architecture by nutrient signals Journal of Experimental Botany, 65, 769-778. DOI:10.1093/jxb/ert421URL [本文引用: 2]
GovindarajuluM, PfefferPE, JinHR, AbubakerJ, DoudsDD, AllenJW, BückingH, LammersPJ, Shachar-HillY (2005). Nitrogen transfer in the arbuscular mycorrhizal symbiosis Nature, 435, 819-823. DOI:10.1038/nature03610URL [本文引用: 1]
HawkinsHJ, JohansenA, GeorgeE (2000). Uptake and transport of organic and inorganic nitrogen by arbuscular mycorrhizal fungi Plant and Soil, 226, 275-285. DOI:10.1023/A:1026500810385URL [本文引用: 1]
HodgeA, StorerK (2015). Arbuscular mycorrhiza and nitrogen: implications for individual plants through to ecosystems Plant and Soil, 386, 1-19. DOI:10.1007/s11104-014-2162-1URL [本文引用: 1]
JansaJ, MozafarA, FrossardE (2003). Long-distance transport of P and Zn through the hyphae of an arbuscular mycorrhizal fungus in symbiosis with maize Agronomie, 23, 481-488. DOI:10.1051/agro:2003013URL [本文引用: 1]
JinH, PfefferPE, DoudsDD, PiotrowskiE, LammersPJ, Shachar-HillY (2005). The uptake, metabolism, transport and transfer of nitrogen in an arbuscular mycorrhizal symbiosis New Phytologist, 168, 687-696. PMID:16313650 [本文引用: 1] Nitrogen (N) is known to be transferred from fungus to plant in the arbuscular mycorrhizal (AM) symbiosis, yet its metabolism, storage and transport are poorly understood. In vitro mycorrhizas of Glomus intra-radices and Ri T-DNA-transformed carrot roots were grown in two-compartment Petri dishes. (15)N- and/or (13)C-labeled substrates were supplied to either the fungal compartment or to separate dishes containing uncolonized roots. The levels and labeling of free amino acids (AAs) in the extra-radical mycelium (ERM) in mycorrhizal roots and in uncolonized roots were measured by gas chromatography/mass spectrometry (GC-MS) and high-performance liquid chromatography (HPLC). Arginine (Arg) was the predominant free AA in the ERM, and almost all Arg molecules became labeled within 3 wk of supplying (15)NH(4) (+) to the fungal compartment. Labeling in Arg represented > 90% of the total (15)N in the free AAs of the ERM. [Guanido-2-(15)N]Arg taken up by the ERM and transported to the intra-radical mycelium (IRM) gave rise to (15)N-labeled AAs. [U-(13)C]Arg added to the fungal compartment did not produce any (13)C labeling of other AAs in the mycorrhizal root. Arg is the major form of N synthesized and stored in the ERM and transported to the IRM. However, NH(4) (+) is the most likely form of N transferred to host cells following its generation from Arg breakdown.
JohnsonNC (2010). Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales New Phytologist, 185, 631-647. DOI:10.1111/nph.2010.185.issue-3URL [本文引用: 1]
LiHB, LiuBT, McCormackML, MaZQ, GuoDL (2017). Diverse belowground resource strategies underlie plant species coexistence and spatial distribution in three grasslands along a precipitation gradient New Phytologist, 216, 1140-1150. DOI:10.1111/nph.2017.216.issue-4URL [本文引用: 1]
LiuBT, LiHB, ZhuB, KoideRT, EissenstatDM, GuoDL (2015). Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species New Phytologist, 208, 125-136. DOI:10.1111/nph.2015.208.issue-1URL [本文引用: 2]
LiuHJ (2005). Cloning and Functional Analysis of ARL1 Gene Required for Adventitious Root Formation in Rice. PhD dissertation, Zhejiang University, Hangzhou. 12-16. [本文引用: 2]
LiuMM, LiYS, SunJ, HeCX (2018). Comparison of propagation of two arbuscular mycorrhizal fungi and their effects on maize growth Bulletin of Agricultural Science and Technology, 556(4), 65-69. [本文引用: 1]
LuYW, LiuX, ChenF, ZhouSR (2020). Shifts in plant community composition weaken the negative effect of nitrogen addition on community-level arbuscular mycorrhizal fungi colonization Proceedings of the Royal Society B: Biological Sciences, 287, 20200483. DOI: 10.1098/rspb.2020.0483. URL [本文引用: 1]
MaXC, GengQH, ZhangHG, BianCY, ChenHYH, JiangDL, XuX (2021). Global negative effects of nutrient enrichment on arbuscular mycorrhizal fungi, plant diversity and ecosystem multifunctionality New Phytologist, 229, 2957-2969. DOI:10.1111/nph.v229.5URL [本文引用: 1]
MaZQ, GuoDL, XuXL, LuMZ, BardgettRD, EissenstatDM, McCormackML, HedinLO (2018). Evolutionary history resolves global organization of root functional traits Nature, 555, 94-97. DOI:10.1038/nature25783URL [本文引用: 2]
MergemannH, SauterM (2000). Ethylene induces epidermal cell death at the site of adventitious root emergence in rice Plant Physiology, 124, 609-614. PMID:11027711 [本文引用: 1] In deepwater rice (Oryza sativa), adventitious root primordia initiate at the nodes as part of normal development. Emergence of the roots is dependent on flooding of the plant and is mediated by ethylene action. Root growth was preceded by the induced death of epidermal cells of the node external to the tip of the root primordium. Cell death proceeded until the epidermis split open. Through this crack the root eventually emerged. Induced death was confined to nodal epidermal cells covering the tip of the primordia. Our results suggest that this process facilitates adventitious root emergence and prevents injury to the growing root. Cell death was inducible not only by submergence but also by application of 1-aminocyclopropane-1-carboxylic acid, the natural precursor of ethylene and it was suppressed in the presence of 2,5-norbornadiene (bicyclo[2.2.1]hepta-2,5-diene), an inhibitor of ethylene action. Adventitious root growth and epidermal cell death are therefore linked to the ethylene signaling pathway, which is activated in response to low oxygen stress.
OhtomoR, KobaeY, MorimotoS, OkaN (2018). Infection unit density as an index of infection potential of arbuscular mycorrhizal fungi Microbes and Environments, 33, 34-39. DOI:10.1264/jsme2.ME17098PMID:29415903 [本文引用: 1] The effective use of arbuscular mycorrhizal (AM) fungal function to promote host plant phosphate uptake in agricultural practice requires the accurate quantitative evaluation of AM fungal infection potential in field soil or AM fungal inoculation material. The number of infection units (IUs), intraradical fungal structures derived from single root entries formed after a short cultivation period, may reflect the number of propagules in soil when pot soil is completely permeated by the host root. However, the original IU method, in which all AM propagules in a pot are counted, requires the fine tuning of plant growing conditions and is considered to be laborious. The objective of the present study was to test whether IU density, not the total count of IU, but the number of IUs per unit root length, reflects the density of AM fungal propagules in soil. IU density assessed after 12 d of host plant cultivation and 3,3'-diaminobenzidine (DAB) staining showed a stronger linear correlation with propagule density than the mean infection percentage (MIP). In addition, IU density was affected less by the host plant species than MIP. We suggest that IU density provides a more rapid and reliable quantitation of the propagule density of AM fungi than MIP or the original IU method. Thus, IU density may be a more robust index of AM fungal infection potential for research and practical applications.
ParniskeM (2008). Arbuscular mycorrhiza: the mother of plant root endosymbioses Nature Reviews Microbiology, 6, 763-775. DOI:10.1038/nrmicro1987PMID:18794914 [本文引用: 1] Arbuscular mycorrhiza (AM), a symbiosis between plants and members of an ancient phylum of fungi, the Glomeromycota, improves the supply of water and nutrients, such as phosphate and nitrogen, to the host plant. In return, up to 20% of plant-fixed carbon is transferred to the fungus. Nutrient transport occurs through symbiotic structures inside plant root cells known as arbuscules. AM development is accompanied by an exchange of signalling molecules between the symbionts. A novel class of plant hormones known as strigolactones are exuded by the plant roots. On the one hand, strigolactones stimulate fungal metabolism and branching. On the other hand, they also trigger seed germination of parasitic plants. Fungi release signalling molecules, in the form of 'Myc factors' that trigger symbiotic root responses. Plant genes required for AM development have been characterized. During evolution, the genetic programme for AM has been recruited for other plant root symbioses: functional adaptation of a plant receptor kinase that is essential for AM symbiosis paved the way for nitrogen-fixing bacteria to form intracellular symbioses with plant cells.
PaszkowskiU, BollerT (2002). The growth defect of lrt1, a maize mutant lacking lateral roots, can be complemented by symbiotic fungi or high phosphate nutrition Planta, 214, 584-590. PMID:11925041 [本文引用: 1] The growth of three maize (Zea mays L.) mutants, each impaired in the formation of one individual element of its root system, was compared under "natural" limiting phosphate conditions (0.1 mM). Mutant plants exhibiting a reduction in root hairs (rth3-1) or a depletion of crown and brace roots (rtcs) grew as well as the corresponding wild-type plants. However, mutant plants lacking lateral roots (lrt1) showed a strong reduction in plant growth. The growth defect of lrt1 was overcome when it was grown in association with an arbuscular mycorrhizal fungus, Glomus mosseae. Establishment of symbiosis was associated with the occurrence of a new type of lateral root. These new lateral roots were stunted and highly branched, giving rise to a bush-like structure. Supply of high phosphate (1 microM) ameliorated the growth of lrt1 plants too, but less efficiently than the symbiosis did. Hence, arbuscular mycorrhizal fungi as well as phosphate functionally complemented the lrt1 mutation.
ReinhardtDR, MillerRM (1990). Size classes of root diameter and mycorrhizal fungal colonization in two temperate grassland communities New Phytologist, 116, 129-136. DOI:10.1111/nph.1990.116.issue-1URL [本文引用: 1]
SchalamukS, CabelloMN, ChidichimoH, GolikS (2011). Effects of inoculation with Glomus mosseae in conventionally tilled and nontilled soils with different levels of nitrogen fertilization on wheat growth, arbuscular mycorrhizal colonization, and nitrogen nutrition Communications in Soil Science and Plant Analysis, 42, 586-598. DOI:10.1080/00103624.2011.546924URL [本文引用: 2]
SmithSE, SmithFA (2011). Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales Annual Review of Plant Biology, 62, 227-250. DOI:10.1146/arplant.2011.62.issue-1URL [本文引用: 1]
SongZY, LüK, LuoF, LianXM (2012). Effect of nitrogen application on nitrogen uptaking and utilization in ten different rice varieties Journal of Huazhong Agricultural University, 31, 165-170. [本文引用: 1]
TatenoR, NakayamaM, YanoM, FukuzawaK, InagakiY, KobaK, UgawaS (2020). Nitrogen source utilization in co-existing canopy tree and dwarf bamboo in a northern hardwood forest in Japan Trees, 34, 1047-1057. DOI:10.1007/s00468-020-01980-1URL [本文引用: 1]
Valenzuela-EstradaLR, Vera-CaraballoV, RuthLE, EissenstatDM (2008). Root anatomy, morphology, and longevity among root orders in Vaccinium corymbosum (Ericaceae) American Journal of Botany, 95, 1506-1514. DOI:10.3732/ajb.0800092PMID:21628158 [本文引用: 1] Understanding root processes at the whole-plant or ecosystem scales requires an accounting of the range of functions within a root system. Studying root traits based on their branching order can be a powerful approach to understanding this complex system. The current study examined the highly branched root system of the ericoid plant, Vaccinium corymbosum L. (highbush blueberry) by classifying its root orders with a modified version of the morphometric approach similar to that used in hydrology for stream classification. Root anatomy provided valuable insight into variation in root function across orders. The more permanent portion of the root system occurred in 4th- and higher-order roots. Roots in these orders had radial growth; the lowest specific root length, N:C ratios, and mycorrhizal colonization; the highest tissue density and vessel number; and the coarsest root diameter. The ephemeral portion of the root system was mainly in the first three root orders. First- and 2nd-order roots were nearly anatomically identical, with similar mycorrhizal colonization and diameter, and also, despite being extremely fine, median lifespans were not very short (115-120 d; estimated with minirhizotrons). Our research underscores the value of examining root traits by root order and its implications to understanding belowground processes.
WangQC, ChengYH (2004). Response of fine roots to soil nutrient spatial heterogeneity Chinese Journal of Applied Ecology, 15, 1063-1068. [本文引用: 1]
WenZH, LiHB, ShenQ, TangXM, XiongCY, LiHG, PangJY, RyanMH, LambersH, ShenJB (2019). Tradeoffs among root morphology, exudation and mycorrhizal symbioses for phosphorus-acquisition strategies of 16 crop species New Phytologist, 223, 882-895. DOI:10.1111/nph.2019.223.issue-2URL [本文引用: 1]
YaharaH, TanikawaN, OkamotoM, MakitaN (2019). Characterizing fine-root traits by species phylogeny and microbial symbiosis in 11 co-existing woody species Oecologia, 191, 983-993. DOI:10.1007/s00442-019-04546-2URL [本文引用: 1]
Fine-root morphological trait variation in tropical forest ecosystems: an evidence synthesis 1 2020
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Common mycorrhizal networks influence the distribution of mineral nutrients between an invasive plant, Solidago canadensis, and a native plant, Kummerowa striata 4 2019
Effect of crop rotation on mycorrhizal colonization and wheat yield under different fertilizer treatments 1 2017
... 前人的研究已证实了在养分匮乏的环境下植物更依赖于AMF吸收养分(Corkidi et al., 2002; Bakhshandeh et al., 2017; Lu et al., 2020), 本文的研究结果与之一致, 共同证明了AMF对植物养分吸收的促进效应受土壤养分影响.在养分匮乏的条件下, 植物的生长更受养分吸收的限制.因此, 为了获取更多的养分, 植物需要通过与AMF共生获取更多的养分(Bonneau et al., 2013); 而在土壤养分充足的情况下, 植物依靠根系便可吸收足够养分, 对AMF的依赖性减弱(Schalamuk et al., 2011). ...
Root hairs confer a competitive advantage under low phosphorus availability 1 2001
Combined phosphate and nitrogen limitation generates a nutrient stress transcriptome favorable for arbuscular mycorrhizal symbiosis in Medicago truncatula 2 2013
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... 前人的研究已证实了在养分匮乏的环境下植物更依赖于AMF吸收养分(Corkidi et al., 2002; Bakhshandeh et al., 2017; Lu et al., 2020), 本文的研究结果与之一致, 共同证明了AMF对植物养分吸收的促进效应受土壤养分影响.在养分匮乏的条件下, 植物的生长更受养分吸收的限制.因此, 为了获取更多的养分, 植物需要通过与AMF共生获取更多的养分(Bonneau et al., 2013); 而在土壤养分充足的情况下, 植物依靠根系便可吸收足够养分, 对AMF的依赖性减弱(Schalamuk et al., 2011). ...
Coevolution of roots and mycorrhizas of land plants 1 2002
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
不同基因型水稻苗期氮营养特性差异及综合评价 5 2016
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... 为了探究不同土壤氮浓度对不同基因型水稻氮吸收的影响, 实验二设置了两种不同的氮养分条件, 并且测定了水稻茎叶含氮量.陈晨等(2016)研究发现茎叶的累积氮含量要高于根的累积氮含量.因此, 本实验通过测定水稻地上部分15N浓度来表征植物根系对矿质氮的吸收能力(Awaydul et al., 2019).结果发现, 氮处理对水稻氮吸收有显著影响(p < 0.001),不同基因型水稻对氮吸收也有显著影响(p < 0.05), 但两者的交互作用对氮吸收的影响并不显著(图3, p > 0.05). ...
... 此外, 本研究还发现侧根缺陷突变株水稻根系的养分吸收能力显著小于根毛缺陷突变株水稻.该结果与Paszkowski和Boller (2002)对不同根形态玉米突变株的研究一致, 两者都证明了侧根对于植物养分吸收的作用更大.根系分枝密度与植物养分吸收呈正相关关系, 是反映养分吸收能力的重要指标(陈晨等, 2016), 而侧根的缺失会导致植物根系的根分枝密度显著减小, 因此, 侧根突变株iaa11养分吸收能力比其他基因型水稻更弱.本研究使用水稻地上部分15N浓度反映水稻根系氮吸收的情况, 而缺少水稻根系与水稻整体氮含量数据.陈晨等(2016)的研究发现水稻整株氮含量、茎叶氮含量、根系氮含量的均值分别为4.22%、4.42%、3.75%, 说明水稻地上部分茎叶氮含量与水稻整株氮含量更接近, 水稻地上部分氮含量也能够反映水稻的氮吸收情况.与此同时, 前人也有很多研究通过测定植物地上部分氮含量来反映植物对氮吸收的情况(Wang et al., 2018a; Awaydul et al., 2019; Tateno et al., 2020). ...
... N浓度反映水稻根系氮吸收的情况, 而缺少水稻根系与水稻整体氮含量数据.陈晨等(2016)的研究发现水稻整株氮含量、茎叶氮含量、根系氮含量的均值分别为4.22%、4.42%、3.75%, 说明水稻地上部分茎叶氮含量与水稻整株氮含量更接近, 水稻地上部分氮含量也能够反映水稻的氮吸收情况.与此同时, 前人也有很多研究通过测定植物地上部分氮含量来反映植物对氮吸收的情况(Wang et al., 2018a; Awaydul et al., 2019; Tateno et al., 2020). ...
Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees 1 2016
... 相对于菌根植物, 根系与根外菌丝都具有养分吸收的功能, 为了避免养分吸收功能的冗余, 植物根系与AMF在养分吸收上往往存在功能互补的关系(Koide, 2000; Chen et al., 2016; Cheng et al., 2016).本文研究结果也验证了这一观点, 养分竞争型根系植物具有较强的养分吸收能力, 因此, 主要依赖自身根系进行养分吸收.相反, 保守型根系植物养分吸收能力较弱, 故主要通过AMF进行养分吸收(Liu et al., 2015).平均根径较粗的侧根缺陷突变体水稻对AMF的依赖性与菌根侵染率要高于其他水稻.在自然界中, 土壤养分的有效性在空间与时间上存在高度异质性(王庆成和程云环, 2004), 因此, 增殖快、周转速率高的细根更适宜于这样的养分环境; 而增殖慢、寿命长的粗根为了提高自身的养分吸收能力选择与AMF共生.同时, 由于粗根寿命长的特点, AMF也更倾向于与粗根共生(Ma et al., 2018).本研究结果说明不同根形态基因型水稻与AMF在氮吸收方面存在互补关系, 该结论支持植物根系与AMF在养分吸收上存在功能互补的理论框架.然而, 本研究没有测定土壤中菌丝体的15N浓度, 故无法在根外菌丝氮吸收方面提供更直接的证据, 还需要在下一步研究中加以验证. ...
Arbuscular mycorrhizal fungi increase organic carbon decomposition under elevated CO2 1 2012
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Mycorrhizal fungi and roots are complementary in foraging within nutrient patches 3 2016
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... ).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... 相对于菌根植物, 根系与根外菌丝都具有养分吸收的功能, 为了避免养分吸收功能的冗余, 植物根系与AMF在养分吸收上往往存在功能互补的关系(Koide, 2000; Chen et al., 2016; Cheng et al., 2016).本文研究结果也验证了这一观点, 养分竞争型根系植物具有较强的养分吸收能力, 因此, 主要依赖自身根系进行养分吸收.相反, 保守型根系植物养分吸收能力较弱, 故主要通过AMF进行养分吸收(Liu et al., 2015).平均根径较粗的侧根缺陷突变体水稻对AMF的依赖性与菌根侵染率要高于其他水稻.在自然界中, 土壤养分的有效性在空间与时间上存在高度异质性(王庆成和程云环, 2004), 因此, 增殖快、周转速率高的细根更适宜于这样的养分环境; 而增殖慢、寿命长的粗根为了提高自身的养分吸收能力选择与AMF共生.同时, 由于粗根寿命长的特点, AMF也更倾向于与粗根共生(Ma et al., 2018).本研究结果说明不同根形态基因型水稻与AMF在氮吸收方面存在互补关系, 该结论支持植物根系与AMF在养分吸收上存在功能互补的理论框架.然而, 本研究没有测定土壤中菌丝体的15N浓度, 故无法在根外菌丝氮吸收方面提供更直接的证据, 还需要在下一步研究中加以验证. ...
Nitrogen fertilization alters the functioning of arbuscular mycorrhizas at two semiarid grasslands 1 2002
... 前人的研究已证实了在养分匮乏的环境下植物更依赖于AMF吸收养分(Corkidi et al., 2002; Bakhshandeh et al., 2017; Lu et al., 2020), 本文的研究结果与之一致, 共同证明了AMF对植物养分吸收的促进效应受土壤养分影响.在养分匮乏的条件下, 植物的生长更受养分吸收的限制.因此, 为了获取更多的养分, 植物需要通过与AMF共生获取更多的养分(Bonneau et al., 2013); 而在土壤养分充足的情况下, 植物依靠根系便可吸收足够养分, 对AMF的依赖性减弱(Schalamuk et al., 2011). ...
A plant-responsive bacterial-signaling system senses an ethanolamine derivative 1 2018
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Linking root traits to nutrient foraging in arbuscular mycorrhizal trees in a temperate forest 1 2015
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Effects of foliar fertilization and arbuscular mycorrhizal colonization on Salvia officinalis L. growth, antioxidant capacity, and essential oil composition 1 2010
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
It’s time to make changes: modulation of root system architecture by nutrient signals 2 2014
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Arbuscular mycorrhiza and nitrogen: implications for individual plants through to ecosystems 1 2015
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Long-distance transport of P and Zn through the hyphae of an arbuscular mycorrhizal fungus in symbiosis with maize 1 2003
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
The uptake, metabolism, transport and transfer of nitrogen in an arbuscular mycorrhizal symbiosis 1 2005
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Resource stoichiometry elucidates the structure and function of arbuscular mycorrhizas across scales 1 2010
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
Functional complementarity in the arbuscular mycorrhizal symbiosis 2 2000
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... 相对于菌根植物, 根系与根外菌丝都具有养分吸收的功能, 为了避免养分吸收功能的冗余, 植物根系与AMF在养分吸收上往往存在功能互补的关系(Koide, 2000; Chen et al., 2016; Cheng et al., 2016).本文研究结果也验证了这一观点, 养分竞争型根系植物具有较强的养分吸收能力, 因此, 主要依赖自身根系进行养分吸收.相反, 保守型根系植物养分吸收能力较弱, 故主要通过AMF进行养分吸收(Liu et al., 2015).平均根径较粗的侧根缺陷突变体水稻对AMF的依赖性与菌根侵染率要高于其他水稻.在自然界中, 土壤养分的有效性在空间与时间上存在高度异质性(王庆成和程云环, 2004), 因此, 增殖快、周转速率高的细根更适宜于这样的养分环境; 而增殖慢、寿命长的粗根为了提高自身的养分吸收能力选择与AMF共生.同时, 由于粗根寿命长的特点, AMF也更倾向于与粗根共生(Ma et al., 2018).本研究结果说明不同根形态基因型水稻与AMF在氮吸收方面存在互补关系, 该结论支持植物根系与AMF在养分吸收上存在功能互补的理论框架.然而, 本研究没有测定土壤中菌丝体的15N浓度, 故无法在根外菌丝氮吸收方面提供更直接的证据, 还需要在下一步研究中加以验证. ...
Diverse belowground resource strategies underlie plant species coexistence and spatial distribution in three grasslands along a precipitation gradient 1 2017
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species 2 2015
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... 相对于菌根植物, 根系与根外菌丝都具有养分吸收的功能, 为了避免养分吸收功能的冗余, 植物根系与AMF在养分吸收上往往存在功能互补的关系(Koide, 2000; Chen et al., 2016; Cheng et al., 2016).本文研究结果也验证了这一观点, 养分竞争型根系植物具有较强的养分吸收能力, 因此, 主要依赖自身根系进行养分吸收.相反, 保守型根系植物养分吸收能力较弱, 故主要通过AMF进行养分吸收(Liu et al., 2015).平均根径较粗的侧根缺陷突变体水稻对AMF的依赖性与菌根侵染率要高于其他水稻.在自然界中, 土壤养分的有效性在空间与时间上存在高度异质性(王庆成和程云环, 2004), 因此, 增殖快、周转速率高的细根更适宜于这样的养分环境; 而增殖慢、寿命长的粗根为了提高自身的养分吸收能力选择与AMF共生.同时, 由于粗根寿命长的特点, AMF也更倾向于与粗根共生(Ma et al., 2018).本研究结果说明不同根形态基因型水稻与AMF在氮吸收方面存在互补关系, 该结论支持植物根系与AMF在养分吸收上存在功能互补的理论框架.然而, 本研究没有测定土壤中菌丝体的15N浓度, 故无法在根外菌丝氮吸收方面提供更直接的证据, 还需要在下一步研究中加以验证. ...
Shifts in plant community composition weaken the negative effect of nitrogen addition on community-level arbuscular mycorrhizal fungi colonization 1 2020
... 前人的研究已证实了在养分匮乏的环境下植物更依赖于AMF吸收养分(Corkidi et al., 2002; Bakhshandeh et al., 2017; Lu et al., 2020), 本文的研究结果与之一致, 共同证明了AMF对植物养分吸收的促进效应受土壤养分影响.在养分匮乏的条件下, 植物的生长更受养分吸收的限制.因此, 为了获取更多的养分, 植物需要通过与AMF共生获取更多的养分(Bonneau et al., 2013); 而在土壤养分充足的情况下, 植物依靠根系便可吸收足够养分, 对AMF的依赖性减弱(Schalamuk et al., 2011). ...
Predicting fine root lifespan from plant functional traits in temperate trees 1 2012
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
Global negative effects of nutrient enrichment on arbuscular mycorrhizal fungi, plant diversity and ecosystem multifunctionality 1 2021
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
Evolutionary history resolves global organization of root functional traits 2 2018
Effects of inoculation with Glomus mosseae in conventionally tilled and nontilled soils with different levels of nitrogen fertilization on wheat growth, arbuscular mycorrhizal colonization, and nitrogen nutrition 2 2011
... 新近提出的一个理论假设认为, 在长期共同进化过程中, 不同根形态植物与其共生菌根真菌之间在养分获取策略上形成互补(Koide, 2000; Cheng et al., 2016).由于绝大多数(>90%)植物的根系与AMF或外生菌根形成共生关系, 理解根形态和菌根真菌之间的关系, 才能够更加准确地预测植物地下部分的养分获取策略.Cheng等(2016)通过野外同质园实验发现植物根和菌根真菌在营养斑块内的觅食是互补的, 即不同根形态树种在营养斑块中的觅食策略不同; 细根树种倾向于通过快速增殖吸收根的方式来增加养分吸收的效率, 而粗根树种选择增殖更多的菌根真菌来促进对养分的吸收.Chen等(2016)对不同根形态温带树种的养分觅食策略进行了研究, 发现随植物根径的增加, 在营养斑块内, 植物根的觅食精度逐渐减小; 但是, AMF根外菌丝的觅食精度变化不大.此外, 很多研究也提出保守型根系植物比养分竞争型根系植物更依赖于AMF吸收养分(Liu et al., 2015; Li et al., 2017).这说明植物根系与AMF在养分吸收上存在互补关系, 在研究不同植物与AMF的互作上需要考虑植物根形态.另一方面, 环境的养分浓度对植物与AMF的互作也存在一定影响, Schalamuk等(2011)发现氮肥添加降低了AMF对根的定殖, 并减弱了小麦(Triticum aestivum)对AMF的响应; Bonneau等(2013)发现氮、磷共同限制的条件更有利于菌根共生.研究表明, 随着土壤养分的降低, 植物的生长对AMF的响应增强(Geneva et al., 2010; Johnson, 2010; Ma et al., 2021).这说明植物与AMF的养分吸收策略受根形态与土壤养分的影响, 而AMF对于植物根系养分吸收的促进效应是否会受根形态与土壤养分的影响并不清楚.对于不同根形态禾本科植物与AMF在养分吸收上互作的研究尤为稀少. ...
... 前人的研究已证实了在养分匮乏的环境下植物更依赖于AMF吸收养分(Corkidi et al., 2002; Bakhshandeh et al., 2017; Lu et al., 2020), 本文的研究结果与之一致, 共同证明了AMF对植物养分吸收的促进效应受土壤养分影响.在养分匮乏的条件下, 植物的生长更受养分吸收的限制.因此, 为了获取更多的养分, 植物需要通过与AMF共生获取更多的养分(Bonneau et al., 2013); 而在土壤养分充足的情况下, 植物依靠根系便可吸收足够养分, 对AMF的依赖性减弱(Schalamuk et al., 2011). ...
Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular to ecosystem scales 1 2011
... 植物在生长的过程中需要从土壤中获取矿质养分, 比如氮.为了适应不同的环境, 植物发展出不同的养分吸收策略, 在不同的环境背景下发育出不同的根形态是策略之一(Giehl et al., 2014; Weemstra et al., 2016; Addo-Danso et al., 2020).植物的根系按养分吸收能力大小可分为两类: 养分竞争型根系和保守型根系.养分竞争型根系根径细、根分支密度高、根毛密集.这些形态特征使植物根系具有更强的养分吸收能力(Eissenstat et al., 2015; Coutinho et al., 2018; Wen et al., 2019).而保守型根系根径粗、根分支密度小、根毛稀疏.这些形态特征使植物根系的养分吸收能力减弱(Valenzuela-Estrada et al., 2008; Luck McCormack et al., 2012; Yahara et al., 2019).此外, 植物还可以与丛枝菌根真菌(AMF)共生, 通过根外菌丝吸收氮.AMF通过大量根外菌丝增加根系与土壤的接触面积, 吸收土壤中的铵态氮(NH4+-N)、硝态氮(NO3--N)以及氨基酸, 并转运给植物, 进而增加植物对氮的吸收(Hawkins et al., 2000; Parniske, 2008; Cheng et al., 2012; Hodge & Storer, 2015).研究发现, 根外菌丝吸收土壤氮后, 迅速通过GS/GOGAT途径合成氨基酸, 如精氨酸(Govindarajulu et al., 2005; Jin et al., 2005).精氨酸从根外菌丝转运至根内菌丝, 并在根内菌丝中被精氨酸酶催化水解生成尿素, 尿素进一步被尿酶分解生成NH4+, 再转运至根细胞(Smith & Smith, 2011).相较于植物根系, AMF的菌丝直径小、数量多, 可以拓展延伸到很多根系无法到达的土壤缝隙中, 进而更有效地帮助植物吸收矿质养分(Brundrett, 2002; Jansa et al., 2003).不同根形态植物对养分吸收存在较大差异.但是, AMF如何影响不同根形态植物的养分吸收仍不清楚. ...
 ,1,2,*
,1,2,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT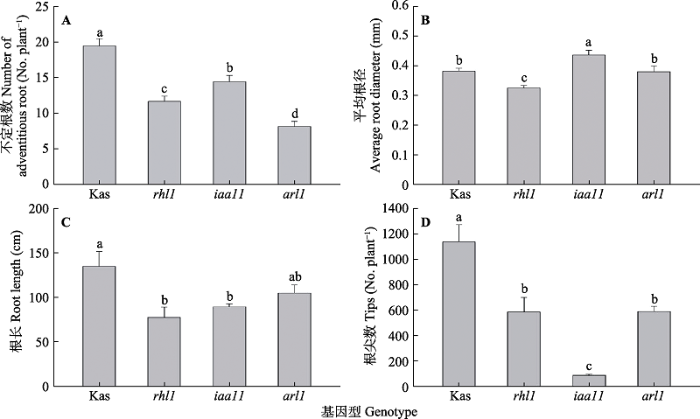 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}