Variations of nitrogen and phosphorus limitation along the environmental gradient in alpine grasslands on the Northern Xizang Plateau
Ning ZONG1, Pei-Li SHI,,1,2,*, Guang-Shuai ZHAO3, Li-Li ZHENG1,2, Ben NIU1, Tian-Cai ZHOU1,2, Ge HOU1,21Lhasa Plateau Ecological Research Station, Key Laboratory of Ecosystem Network Observation and Modelling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China 2University of Chinese Academy of Sciences, Beijing 100049, China 3Economics and Development Research Center, National Forestry and Grassland Administration, Beijing 100714, China
the National Key R&D Program of China(2017YFA0604802) the National Key R&D Program of China(2016YFC0501803) National Natural Science Foundation of China(31870406) National Natural Science Foundation of China(41703079) the Second Tibetan Plateau Scientific Expedition and Research (STEP) Program(2019QZKK0302)
Abstract Aims Understanding the mode of nutrient limitation on ecosystem net primary production is an important issue of modern ecology. Nutrient availability is a key determinant of ecosystem dynamics, but the relationship between soil resource availability and ecosystem nutrient limitation is still unclear. Methods A series of nitrogen and phosphorus nutrient addition experiments were set up in four types of alpine grasslands (alpine meadow, alpine meadow-steppe, alpine steppe and alpine desert-steppe) along the precipitation gradient on the Northern Xizang, to systematically study the effects of nitrogen and phosphorus addition on different types of alpine grasslands, and to explore the nitrogen and phosphorus limitation models of different alpine grasslands. Important findings The results showed that: (1) The effects of nitrogen and phosphorus addition on different alpine grasslands varied. Nitrogen addition significantly increased the aboveground biomass of alpine meadows and alpine meadow grasslands, but had no effect on alpine meadows and alpine desert grasslands. The addition of phosphorus alone had no significant effect on the four alpine grasslands, while the addition of nitrogen and phosphorus had a promoting effect on the aboveground biomass of the four alpine grasslands. (2) With the decrease of precipitation, the nitrogen limitation index of the alpine grasslands gradually decreased from 1.18 to 0.52-0.64, and the nutrient limitation mode transitioned from nitrogen limitation to co-limitation by nitrogen and phosphorus; the phosphorus limitation index was negative in the alpine meadow-steppe and alpine steppe, indicating that phosphorus addition alone has side effects on these two grassland types. These results suggest that alpine meadow is mainly limited by nitrogen availability, and phosphorus addition alone has side effects; the alpine meadow-steppe is between the nitrogen limit and the joint nitrogen and phosphorus limitation, and phosphorus addition alone also has side effects; the alpine steppe is limited by both nitrogen and phosphorus availability, and the addition of phosphorus has side effects; the alpine desert-steppe is jointly limited by nitrogen and phosphorus availability. These results show that nutrient limitation mode transits from nitrogen limitation to nitrogen and phosphorus co-limitation with the decrease of precipitation. This study implies that the impacts of increasing nitrogen deposition under future climate change on different types of alpine grasslands may be different. Additionally, the differences in nitrogen and phosphorus limitation mode should also be taken into consideration when nutrient addition is used to restore different types of degraded alpine grasslands. Keywords:control experiment;nitrogen and phosphorus limitation;grassland type;plant functional group
PDF (1210KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 宗宁, 石培礼, 赵广帅, 郑莉莉, 牛犇, 周天财, 侯阁. 降水量变化对藏北高寒草地养分限制的影响. 植物生态学报, 2021, 45(5): 444-455. DOI: 10.17521/cjpe.2020.0135 ZONG Ning, SHI Pei-Li, ZHAO Guang-Shuai, ZHENG Li-Li, NIU Ben, ZHOU Tian-Cai, HOU Ge. Variations of nitrogen and phosphorus limitation along the environmental gradient in alpine grasslands on the Northern Xizang Plateau. Chinese Journal of Plant Ecology, 2021, 45(5): 444-455. DOI: 10.17521/cjpe.2020.0135
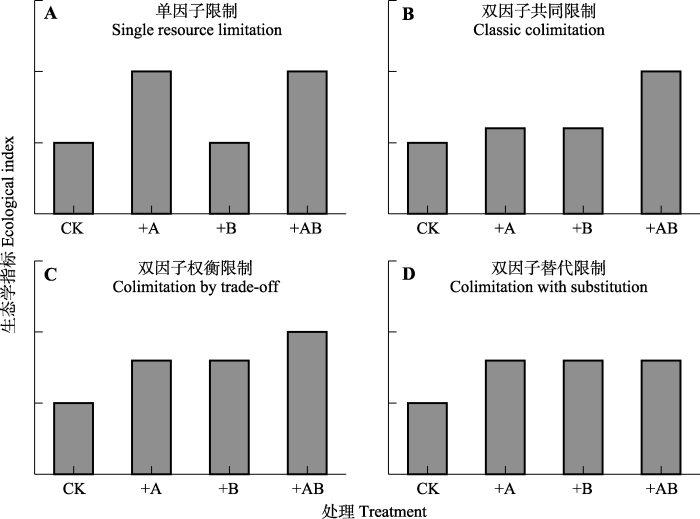
Fig. 1Biomass response patterns of the four main types of nitrogen and phosphorus co-limitation. Assuming that nutritional additions have no negative effects. CK, control (no resource addition); +A, adding resource A; +B, adding resource B; +AB, adding A and B together.
Fig. 2Inter-annual variations of precipitation in different types of alpine grasslands and the relationship between soil nitrogen (N) and phosphorus (P) content and precipitation on the Northern Xizang Plateau. ADS, alpine desert-steppe; AM, alpine meadow; AMS, alpine meadow-steppe; AS, alpine steppe.
随着降水量的减少, 降水梯度带上土壤氮含量背景值呈现显著降低的趋势(图2B, p < 0.001), 而磷含量背景值整体呈现逐渐升高的趋势(图2B, p = 0.02)。两者之间关系分析表明, 虽然土壤氮磷含量背景值沿降水量变化呈现相反的趋势, 但两者之间的相关关系并不显著(图2C, p = 0.12)。土壤氮磷比随降水量增加呈现显著增加趋势(图2D, p = 0.02)。
2.2 不同类型草地地上生产力变化
氮磷添加对不同高寒草地地上生产力的影响存在差异(表2, p < 0.001), 同时也存在年际间差异(表2, p < 0.001)。氮添加和氮磷共同添加只在施肥第3年(2015年)显著提高高寒草甸地上生产力, 提高比例分别是49%和72% (p < 0.05), 在其他年份无显著影响(图3A-3C)。而对于高寒草甸草原, 氮添加和氮磷共同添加只在施肥第4年(2016年)显著提高高寒草甸草原地上生产力, 提高比例分别是78%和113% (图3D-3F, p < 0.05), 在其他年份无显著影响(图3D-3F, p > 0.05)。对于高寒草原, 氮磷共同添加在所有年份均显著提高地上生产力, 2014-2016年提高比例分别是48%、101%和102% (图3G-3I, p < 0.05), 而单独氮添加和磷添加处理无显著影响(图3G-3I, p > 0.05)。氮磷共同添加只在施肥第4年(2016年)显著提高了149%的高寒荒漠草原地上生产力(图3J-3L, p < 0.05), 在其他年份无显著影响, 单独氮添加无显著影响(图3J-3L, p > 0.05)。磷添加在所有年份对各种类型高寒草地均无显著影响(图3)。
Table 2 表2 表2以年份为重复因子, 利用重复测定方差分析氮磷添加对不同草地类型高寒草地植物功能群地上生产力的影响 Table 2Using year as the repeated factor, Repeated Measure ANOVA analysis of the effects of nitrogen and phosphorus addition on the aboveground biomass of different types of alpine grasslands
Fig. 3Effects of nitrogen (N) and phosphorus (P) addition on the aboveground biomass of different plant functional groups of alpine grasslands. Different uppercase letters in the same year represent significant differences among fertilization treatments (p < 0.05). F and P represent the differences in different functional groups among fertilization treatments. A-C, Alpine meadow. D-F, Alpine meadow-steppe. G-I, Alpine steppe. J-L, Alpine desert-steppe.
氮磷添加对禾草植物地上生产力的影响存在年际间和草地类型间的差异(表2; 图3)。总体来讲, 与对照相比较, 氮添加提高了高寒草甸和高寒草甸草原禾草植物地上生产力, 增加比例分别是254%- 491%和53%-209% (图3A-3F), 而在高寒草原和高寒荒漠草原对禾草植物无显著影响(图3G-3L, p > 0.05)。磷添加在所有草地中均对禾草植物无显著影响(图3, p > 0.05)。氮磷添加对高寒草甸和高寒草原禾草植物无显著影响, 而显著提高高寒草甸草原和高寒荒漠草原禾草植物地上生产力, 增加比例分别是13%- 370%和67%-239% (图3)。
Fig. 4Relative co-limitation index of nitrogen and phosphorus for different types of alpine grasslands. ADS, alpine desert-steppe; AM, alpine meadow; AMS, alpine meadow-steppe; AS, alpine steppe. RCIN, nitrogen limitation index; RCIP, phosphorus limitation index. Different uppercase letters in the same year represent significant differences among grassland types (p < 0.05).
Fig. 5Relationships between relative co-limitation index and precipitation as well as soil nutrient content in alpine grasslands on the Northern Xizang Plateau. RCIN, nitrogen limitation index; RCIP, phosphorus limitation index.
BowmanWD,TheodoseTA,SchardtJC,ConantRT(1993).Constraints of nutrient availability on primary production in two alpine tundra communities .Ecology,74, 2085-2097. DOI:10.2307/1940854URL [本文引用: 3]
BrantAN,ChenHYH(2015).Patterns and mechanisms of nutrient resorption in plants .Critical Reviews in Plant Sciences,34, 471-486. DOI:10.1080/07352689.2015.1078611URL [本文引用: 1]
ClarkCM,TilmanD(2008).Loss of plant species after chronic low-level nitrogen deposition to prairie grasslands .Nature,451, 712-715. DOI:10.1038/nature06503URL [本文引用: 1]
CraineJM,JacksonRD(2010).Plant nitrogen and phosphorus limitation in 98 North American grassland soils .Plant and Soil,334, 73-84. DOI:10.1007/s11104-009-0237-1URL [本文引用: 5]
CraineJM,MorrowC,StockWD(2008).Nutrient concentration ratios and co-limitation in South African grasslands .New Phytologist,179, 829-836. DOI:10.1111/nph.2008.179.issue-3URL [本文引用: 1]
CrowleyKF,McNeilBE,LovettGM,CanhamCD,DriscollCT,RustadLE,DennyE,HallettRA,ArthurMA,BoggsJL,GoodaleCL,KahlJS,McNultySG,OllingerSV,PardoLH,SchabergPG,StoddardJL,WeandMP,WeathersKC(2012).Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the Northeastern United States? Ecosystems,15, 940-957. DOI:10.1007/s10021-012-9550-2URL [本文引用: 1]
DuEZ,TerrerC,PellegriniAFA,AhlstromA,van LissaCJ,ZhaoX,XiaN,WuXH,JacksonRB(2020).Global patterns of terrestrial nitrogen and phosphorus limitation .Nature Geoscience,13, 221-226. DOI:10.1038/s41561-019-0530-4URL [本文引用: 2]
ElserJJ,BrackenMES,ClelandEE,GrunerDS,HarpoleWS,HillebrandH,NgaiJT,SeabloomEW,ShurinJB,SmithJE(2007).Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems .Ecology Letters,10, 1135-1142. DOI:10.1111/ele.2007.10.issue-12URL [本文引用: 3]
GaoQZ,LiYE,WanYF(2006).Grassland degradation in Northern Tibet based on remote sensing data .Journal of Geographical Sciences,16, 165-173. DOI:10.1007/s11442-006-0204-1URL [本文引用: 1]
GaoYH,CooperDJ,MaXX(2016).Phosphorus additions have no impact on plant biomass or soil nitrogen in an alpine meadow on the Qinghai-Tibetan Plateau, China .Applied Soil Ecology,106, 18-23. DOI:10.1016/j.apsoil.2016.04.020URL [本文引用: 1]
GaoYH,CooperDJ,ZengXY(2018).Nitrogen, not phosphorus, enrichment controls biomass production in alpine wetlands on the Tibetan Plateau, China .Ecological Engineering,116, 31-34. DOI:10.1016/j.ecoleng.2018.02.016URL [本文引用: 1]
GleesonSK,TilmanD(1992).Plant allocation and the multiple limitation hypothesis .The American Naturalist,139, 1322-1343. DOI:10.1086/285389URL [本文引用: 1]
GusewellS(2004).N:P ratios in terrestrial plants: variation and functional significance .New Phytologist,164, 243-266. DOI:10.1111/nph.2004.164.issue-2URL
HanWX,TangLY,ChenYH,FangJY(2013).Relationship between the relative limitation and resorption efficiency of nitrogen vs phosphorus in woody plants .PLOS ONE,8, e83366. DOI:10.1371/journal.pone.0083366. DOI:10.1371/journal.pone.0083366URL
HautierY,NiklausPA,HectorA(2009).Competition for light causes plant biodiversity loss after eutrophication .Science,324, 636-638. DOI:10.1126/science.1169640PMID:19407202 [本文引用: 1] Human activities have increased the availability of nutrients in terrestrial and aquatic ecosystems. In grasslands, this eutrophication causes loss of plant species diversity, but the mechanism of this loss has been difficult to determine. Using experimental grassland plant communities, we found that addition of light to the grassland understory prevented the loss of biodiversity caused by eutrophication. There was no detectable role for competition for soil resources in diversity loss. Thus, competition for light is a major mechanism of plant diversity loss after eutrophication and explains the particular threat of eutrophication to plant diversity. Our conclusions have implications for grassland management and conservation policy and underscore the need to control nutrient enrichment if plant diversity is to be preserved.
HuangJ,WangGF,AnSZ,YunJ,LiH,ZhangRH(2009).Effect of nitrogen fertilization on the vegetation structure and biomass of degraded meadow and soil fertility .Pratacultural Science,26, 75-78. [本文引用: 1]
LamarqueJF,HessP,EmmonsL,BujaL,WashingtonW,GranierC(2005).Tropospheric ozone evolution between 1890 and 1990 .Journal of Geophysical Research,110, D08304. DOI:10.1029/2004jd005537. DOI:10.1029/2004jd005537 [本文引用: 1]
LeBauerDS,TresederKK(2008).Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed .Ecology,89, 371-379. PMID:18409427 [本文引用: 1] Our meta-analysis of 126 nitrogen addition experiments evaluated nitrogen (N) limitation of net primary production (NPP) in terrestrial ecosystems. We tested the hypothesis that N limitation is widespread among biomes and influenced by geography and climate. We used the response ratio (R approximately equal ANPP(N)/ANPP(ctrl)) of aboveground plant growth in fertilized to control plots and found that most ecosystems are nitrogen limited with an average 29% growth response to nitrogen (i.e., R = 1.29). The response ratio was significant within temperate forests (R = 1.19), tropical forests (R = 1.60), temperate grasslands (R = 1.53), tropical grasslands (R = 1.26), wetlands (R = 1.16), and tundra (R = 1.35), but not deserts. Eight tropical forest studies had been conducted on very young volcanic soils in Hawaii, and this subgroup was strongly N limited (R = 2.13), which resulted in a negative correlation between forest R and latitude. The degree of N limitation in the remainder of the tropical forest studies (R = 1.20) was comparable to that of temperate forests, and when the young Hawaiian subgroup was excluded, forest R did not vary with latitude. Grassland response increased with latitude, but was independent of temperature and precipitation. These results suggest that the global N and C cycles interact strongly and that geography can mediate ecosystem response to N within certain biome types.
LiMS(2000).Rational exploitation of grassland resources in the Northern Xizang Plateau .Journal of Natural Resources,15, 335-339. [本文引用: 1]
LiebigJ(1855).Principles of Agricultural Chemistry. 2nd ed.Walton and Maberly,London. [本文引用: 1]
OdumEP,BarrettGW(2005).Fundamentals of Ecology. 5th ed.Thomson Brooks/Cole, Belmont,USA. [本文引用: 1]
PeñuelasJ,PoulterB,SardansJ,CiaisP,van der VeldeM,BoppL,BoucherO,GodderisY,HinsingerP,LlusiaJ,NardinE,ViccaS,ObersteinerM,JanssensIA(2013).Human-induced nitrogen-phosphorus imbalances alter natural and managed ecosystems across the globe .Nature Communications,4, 2934. DOI:10.1038/ncomms3934. DOI:10.1038/ncomms3934PMID:24343268 [本文引用: 1] The availability of carbon from rising atmospheric carbon dioxide levels and of nitrogen from various human-induced inputs to ecosystems is continuously increasing; however, these increases are not paralleled by a similar increase in phosphorus inputs. The inexorable change in the stoichiometry of carbon and nitrogen relative to phosphorus has no equivalent in Earth's history. Here we report the profound and yet uncertain consequences of the human imprint on the phosphorus cycle and nitrogen: phosphorus stoichiometry for the structure, functioning and diversity of terrestrial and aquatic organisms and ecosystems. A mass balance approach is used to show that limited phosphorus and nitrogen availability are likely to jointly reduce future carbon storage by natural ecosystems during this century. Further, if phosphorus fertilizers cannot be made increasingly accessible, the crop yields projections of the Millennium Ecosystem Assessment imply an increase of the nutrient deficit in developing regions.
ShenZX,ZhouXM,ChenZZ,ZhouHK(2002).Response of plant groups to simulated rainfall and nitrogen supply in alpine Kobresia humilis meadow .Acta Phytoecologica Sinica,26, 288-294. [本文引用: 2]
SongMH,YuFH,OuyangH,CaoGM,XuXL,CornelissenJHC(2012).Different inter-annual responses to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing .Global Change Biology,18, 3100-3111. DOI:10.1111/gcb.2012.18.issue-10URL [本文引用: 1]
SongMH,ZongN,JiangJ,ShiPL,ZhangXZ,GaoJQ,ZhouHK,LiYK,LoreauM(2019).Nutrient-induced shifts of dominant species reduce ecosystem stability via increases in species synchrony and population variability .Science of the Total Environment,692, 441-449. DOI:10.1016/j.scitotenv.2019.07.266URL [本文引用: 2]
SullivanBW,Alvarez-ClareS,CastleSC,PorderS,ReedSC,SchreegL,TownsendAR,ClevelandCC(2014).Assessing nutrient limitation in complex forested ecosystems: alternatives to large-scale fertilization experiments .Ecology,95, 668-681. DOI:10.1890/13-0825.1URL [本文引用: 2]
TilmanD(1982).Resource Competition and Community Structure.Princeton University Press,Princeton. [本文引用: 2]
VitousekPM(2004).Nutrient Cycling and Limitation: Hawai'i as a Model System.Princeton University Press,Princeton. [本文引用: 1]
WiederWR,ClevelandCC,SmithWK,Todd-BrownK(2015).Future productivity and carbon storage limited by terrestrial nutrient availability .Nature Geoscience,8, 441-444. DOI:10.1038/ngeo2413URL [本文引用: 1]
WuJS,ZhangXZ,ShenZX,ShiPL,XuXL,LiXJ(2013).Grazing-exclusion effects on aboveground biomass and water-use efficiency of alpine grasslands on the Northern Tibetan Plateau .Rangeland Ecology and Management,66, 454-461. DOI:10.2111/REM-D-12-00051.1URL [本文引用: 2]
YanZB,TianD,HanWX,TangZY,FangJY(2017).An assessment on the uncertainty of the nitrogen to phosphorus ratio as a threshold for nutrient limitation in plants .Annals of Botany,120, 937-942. DOI:10.1093/aob/mcx106URL
YangXX,RenF,ZhouHK,HeJS(2014).Responses of plant community biomass to nitrogen and phosphorus additions in an alpine meadow on the Qinghai-Xizang Plateau .Chinese Journal of Plant Ecology,38, 159-166. DOI:10.3724/SP.J.1258.2014.00014URL [本文引用: 3]
YuanZY,ChenHYH(2009).Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation .Global Ecology and Biogeography,18, 11-18. DOI:10.1111/geb.2009.18.issue-1URL [本文引用: 2]
ZhaoGS,ShiPL,WuJS,XiongDP,ZongN,ZhangXZ(2017).Foliar nutrient resorption patterns of four functional plants along a precipitation gradient on the Tibetan Changtang Plateau .Ecology and Evolution,7, 7201-7212. DOI:10.1002/ece3.2017.7.issue-18URL [本文引用: 7]
ZongN,ShiPL,NiuB,JiangJ,SongMH,ZhangXZ,HeYT(2014).Effects of nitrogen and phosphorous fertilization on community structure and productivity of degraded alpine meadows in northern Tibet, China .Chinese Journal of Applied Ecology,25, 3458-3468. [本文引用: 4]
ZongN,SongMH,ZhaoGS,ShiPL(2020).Nitrogen economy of alpine plants on the north Tibetan Plateau: nitrogen conservation by resorption rather than open sources through biological symbiotic fixation .Ecology and Evolution,10, 2051-2061. DOI:10.1002/ece3.v10.4URL [本文引用: 1]
Constraints of nutrient availability on primary production in two alpine tundra communities 3 1993
... 目前有几种方法被用于推断生态系统养分限制模式, 包括土壤养分含量、氮磷比、养分回收效率等(Du et al., 2020).资源比率类型的限制性假说认为, 土壤氮磷的相对有效性决定了植物是否受到氮、磷或氮和磷的共同限制(Tilman, 1982; Vitousek, 2004), 即不同生态系统的养分限制模式应与资源可用性平行: 磷可用性相对较低的地点应主要受磷限制, 氮可用性相对较低的地点应主要受氮限制, 而氮和磷中等可用性的地点受两种元素共同限制.尽管土壤养分含量是生态系统养分限制的主要决定因素, 但土壤养分可用性与生态系统养分限制之间的关系尚不清楚(Craine & Jackson, 2010), 故以养分多寡作为评价生态系统养分限制的依据存在较大不确定性.同时, 植物、土壤微生物和资源可利用性之间的相互作用非常复杂, 以至于很难预测大多数草原生态系统中特定的养分限制模式(Craine et al., 2008; Craine & Jackson, 2010).施肥试验可以直接确定限制养分的类型与程度, 利用植物对养分添加的响应来评价生态系统养分限制模式是一种重要方法(Sullivan et al., 2014).美国西部落基山高寒草甸氮磷添加试验表明, 干草甸群落生产主要受氮限制, 而湿草甸受氮磷共同限制(Bowman et al., 1993).对青藏高原湿润草甸的研究发现, 氮添加对高寒草甸植物总生产力无显著影响, 而磷添加后植物总生产力显著增加, 这表明高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制(杨晓霞等, 2014).作者前期在草原化高寒草甸的研究发现, 长期单独施氮对轻度和重度退化高寒草甸群落生产均无显著影响, 而氮磷配施显著提高群落盖度及地上、地下生产力, 这种类型高寒草地生产受到氮和磷的共同限制(宗宁等, 2014; Song et al., 2019).氮磷添加试验已开展很多, 但已有试验多在单点开展, 目前对不同类型草地施肥来探究氮磷限制模式的研究仍然较少. ...
Nutrient concentration ratios and co-limitation in South African grasslands 1 2008
... 目前有几种方法被用于推断生态系统养分限制模式, 包括土壤养分含量、氮磷比、养分回收效率等(Du et al., 2020).资源比率类型的限制性假说认为, 土壤氮磷的相对有效性决定了植物是否受到氮、磷或氮和磷的共同限制(Tilman, 1982; Vitousek, 2004), 即不同生态系统的养分限制模式应与资源可用性平行: 磷可用性相对较低的地点应主要受磷限制, 氮可用性相对较低的地点应主要受氮限制, 而氮和磷中等可用性的地点受两种元素共同限制.尽管土壤养分含量是生态系统养分限制的主要决定因素, 但土壤养分可用性与生态系统养分限制之间的关系尚不清楚(Craine & Jackson, 2010), 故以养分多寡作为评价生态系统养分限制的依据存在较大不确定性.同时, 植物、土壤微生物和资源可利用性之间的相互作用非常复杂, 以至于很难预测大多数草原生态系统中特定的养分限制模式(Craine et al., 2008; Craine & Jackson, 2010).施肥试验可以直接确定限制养分的类型与程度, 利用植物对养分添加的响应来评价生态系统养分限制模式是一种重要方法(Sullivan et al., 2014).美国西部落基山高寒草甸氮磷添加试验表明, 干草甸群落生产主要受氮限制, 而湿草甸受氮磷共同限制(Bowman et al., 1993).对青藏高原湿润草甸的研究发现, 氮添加对高寒草甸植物总生产力无显著影响, 而磷添加后植物总生产力显著增加, 这表明高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制(杨晓霞等, 2014).作者前期在草原化高寒草甸的研究发现, 长期单独施氮对轻度和重度退化高寒草甸群落生产均无显著影响, 而氮磷配施显著提高群落盖度及地上、地下生产力, 这种类型高寒草地生产受到氮和磷的共同限制(宗宁等, 2014; Song et al., 2019).氮磷添加试验已开展很多, 但已有试验多在单点开展, 目前对不同类型草地施肥来探究氮磷限制模式的研究仍然较少. ...
Do nutrient limitation patterns shift from nitrogen toward phosphorus with increasing nitrogen deposition across the Northeastern United States? 1 2012
... 陆生生态系统初级生产力受必需营养元素的限制, 尤其是受氮和磷限制(Elser et al., 2007), 氮磷在植物生长、发育和繁殖等过程中起着重要的作用.生态系统净初级生产养分限制的性质和程度, 是现代生态学关注的重要科学问题.自然状态下, 很多生态系统生产力都表现为氮限制或者磷限制或者氮磷共同限制(Elser et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010; Harpole et al., 2011).大尺度的综合分析表明, 陆地上自然生态系统约18%面积的植被受氮限制, 约43%受磷限制, 其余39%可能受氮和磷的共同限制(Du et al., 2020).20世纪初以来, 人类活动导致全球大气氮沉降量增加了3倍以上(Galloway et al., 2004; Lamarque et al., 2005), 氮沉降增加已经极大地影响了某些自然生态系统的稳定性和功能, 同时还会增加某些区域的磷限制程度(Crowley et al., 2012; Peñuelas et al., 2013).在全球范围内, 氮和磷的限制可能会影响生态系统对当前和未来CO2浓度升高和气候变化的反应(Wieder et al., 2015).因此, 了解养分限制状况对于理解生态系统对未来气候变化的响应和适应至关重要.同时, 受气候变化和放牧干扰的共同影响, 高寒草地呈现普遍退化趋势, 生态系统结构与功能受损, 草地质量日趋下降(Gao et al., 2006).合理施肥已成为改善草原生态系统养分状况和恢复退化草地的重要管理措施(黄军等, 2009; 宗宁等, 2014).合理的草地施肥能增加土壤肥力, 改善草地植物群落组成, 有助于草地生产力的恢复.但目前由于气候条件、环境因子与群落结构组成的不同, 不同类型草地的氮磷限制模式并不清楚, 这严重妨碍了施肥措施在不同类型高寒草地的应用. ...
Global patterns of terrestrial nitrogen and phosphorus limitation 2 2020
... 陆生生态系统初级生产力受必需营养元素的限制, 尤其是受氮和磷限制(Elser et al., 2007), 氮磷在植物生长、发育和繁殖等过程中起着重要的作用.生态系统净初级生产养分限制的性质和程度, 是现代生态学关注的重要科学问题.自然状态下, 很多生态系统生产力都表现为氮限制或者磷限制或者氮磷共同限制(Elser et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010; Harpole et al., 2011).大尺度的综合分析表明, 陆地上自然生态系统约18%面积的植被受氮限制, 约43%受磷限制, 其余39%可能受氮和磷的共同限制(Du et al., 2020).20世纪初以来, 人类活动导致全球大气氮沉降量增加了3倍以上(Galloway et al., 2004; Lamarque et al., 2005), 氮沉降增加已经极大地影响了某些自然生态系统的稳定性和功能, 同时还会增加某些区域的磷限制程度(Crowley et al., 2012; Peñuelas et al., 2013).在全球范围内, 氮和磷的限制可能会影响生态系统对当前和未来CO2浓度升高和气候变化的反应(Wieder et al., 2015).因此, 了解养分限制状况对于理解生态系统对未来气候变化的响应和适应至关重要.同时, 受气候变化和放牧干扰的共同影响, 高寒草地呈现普遍退化趋势, 生态系统结构与功能受损, 草地质量日趋下降(Gao et al., 2006).合理施肥已成为改善草原生态系统养分状况和恢复退化草地的重要管理措施(黄军等, 2009; 宗宁等, 2014).合理的草地施肥能增加土壤肥力, 改善草地植物群落组成, 有助于草地生产力的恢复.但目前由于气候条件、环境因子与群落结构组成的不同, 不同类型草地的氮磷限制模式并不清楚, 这严重妨碍了施肥措施在不同类型高寒草地的应用. ...
... 目前有几种方法被用于推断生态系统养分限制模式, 包括土壤养分含量、氮磷比、养分回收效率等(Du et al., 2020).资源比率类型的限制性假说认为, 土壤氮磷的相对有效性决定了植物是否受到氮、磷或氮和磷的共同限制(Tilman, 1982; Vitousek, 2004), 即不同生态系统的养分限制模式应与资源可用性平行: 磷可用性相对较低的地点应主要受磷限制, 氮可用性相对较低的地点应主要受氮限制, 而氮和磷中等可用性的地点受两种元素共同限制.尽管土壤养分含量是生态系统养分限制的主要决定因素, 但土壤养分可用性与生态系统养分限制之间的关系尚不清楚(Craine & Jackson, 2010), 故以养分多寡作为评价生态系统养分限制的依据存在较大不确定性.同时, 植物、土壤微生物和资源可利用性之间的相互作用非常复杂, 以至于很难预测大多数草原生态系统中特定的养分限制模式(Craine et al., 2008; Craine & Jackson, 2010).施肥试验可以直接确定限制养分的类型与程度, 利用植物对养分添加的响应来评价生态系统养分限制模式是一种重要方法(Sullivan et al., 2014).美国西部落基山高寒草甸氮磷添加试验表明, 干草甸群落生产主要受氮限制, 而湿草甸受氮磷共同限制(Bowman et al., 1993).对青藏高原湿润草甸的研究发现, 氮添加对高寒草甸植物总生产力无显著影响, 而磷添加后植物总生产力显著增加, 这表明高寒草甸植物的生长极有可能更受土壤中可利用磷含量的限制(杨晓霞等, 2014).作者前期在草原化高寒草甸的研究发现, 长期单独施氮对轻度和重度退化高寒草甸群落生产均无显著影响, 而氮磷配施显著提高群落盖度及地上、地下生产力, 这种类型高寒草地生产受到氮和磷的共同限制(宗宁等, 2014; Song et al., 2019).氮磷添加试验已开展很多, 但已有试验多在单点开展, 目前对不同类型草地施肥来探究氮磷限制模式的研究仍然较少. ...
Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems 3 2007
... 陆生生态系统初级生产力受必需营养元素的限制, 尤其是受氮和磷限制(Elser et al., 2007), 氮磷在植物生长、发育和繁殖等过程中起着重要的作用.生态系统净初级生产养分限制的性质和程度, 是现代生态学关注的重要科学问题.自然状态下, 很多生态系统生产力都表现为氮限制或者磷限制或者氮磷共同限制(Elser et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010; Harpole et al., 2011).大尺度的综合分析表明, 陆地上自然生态系统约18%面积的植被受氮限制, 约43%受磷限制, 其余39%可能受氮和磷的共同限制(Du et al., 2020).20世纪初以来, 人类活动导致全球大气氮沉降量增加了3倍以上(Galloway et al., 2004; Lamarque et al., 2005), 氮沉降增加已经极大地影响了某些自然生态系统的稳定性和功能, 同时还会增加某些区域的磷限制程度(Crowley et al., 2012; Peñuelas et al., 2013).在全球范围内, 氮和磷的限制可能会影响生态系统对当前和未来CO2浓度升高和气候变化的反应(Wieder et al., 2015).因此, 了解养分限制状况对于理解生态系统对未来气候变化的响应和适应至关重要.同时, 受气候变化和放牧干扰的共同影响, 高寒草地呈现普遍退化趋势, 生态系统结构与功能受损, 草地质量日趋下降(Gao et al., 2006).合理施肥已成为改善草原生态系统养分状况和恢复退化草地的重要管理措施(黄军等, 2009; 宗宁等, 2014).合理的草地施肥能增加土壤肥力, 改善草地植物群落组成, 有助于草地生产力的恢复.但目前由于气候条件、环境因子与群落结构组成的不同, 不同类型草地的氮磷限制模式并不清楚, 这严重妨碍了施肥措施在不同类型高寒草地的应用. ...
... ), 氮磷在植物生长、发育和繁殖等过程中起着重要的作用.生态系统净初级生产养分限制的性质和程度, 是现代生态学关注的重要科学问题.自然状态下, 很多生态系统生产力都表现为氮限制或者磷限制或者氮磷共同限制(Elser et al., 2007; LeBauer & Treseder, 2008; Vitousek et al., 2010; Harpole et al., 2011).大尺度的综合分析表明, 陆地上自然生态系统约18%面积的植被受氮限制, 约43%受磷限制, 其余39%可能受氮和磷的共同限制(Du et al., 2020).20世纪初以来, 人类活动导致全球大气氮沉降量增加了3倍以上(Galloway et al., 2004; Lamarque et al., 2005), 氮沉降增加已经极大地影响了某些自然生态系统的稳定性和功能, 同时还会增加某些区域的磷限制程度(Crowley et al., 2012; Peñuelas et al., 2013).在全球范围内, 氮和磷的限制可能会影响生态系统对当前和未来CO2浓度升高和气候变化的反应(Wieder et al., 2015).因此, 了解养分限制状况对于理解生态系统对未来气候变化的响应和适应至关重要.同时, 受气候变化和放牧干扰的共同影响, 高寒草地呈现普遍退化趋势, 生态系统结构与功能受损, 草地质量日趋下降(Gao et al., 2006).合理施肥已成为改善草原生态系统养分状况和恢复退化草地的重要管理措施(黄军等, 2009; 宗宁等, 2014).合理的草地施肥能增加土壤肥力, 改善草地植物群落组成, 有助于草地生产力的恢复.但目前由于气候条件、环境因子与群落结构组成的不同, 不同类型草地的氮磷限制模式并不清楚, 这严重妨碍了施肥措施在不同类型高寒草地的应用. ...
Different inter-annual responses to availability and form of nitrogen explain species coexistence in an alpine meadow community after release from grazing 1 2012
Nitrogen economy of alpine plants on the north Tibetan Plateau: nitrogen conservation by resorption rather than open sources through biological symbiotic fixation 1 2020
 ,
, ,1,2,*, 赵广帅3, 郑莉莉1,2, 牛犇1, 周天财1,2, 侯阁1,2
,1,2,*, 赵广帅3, 郑莉莉1,2, 牛犇1, 周天财1,2, 侯阁1,2

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT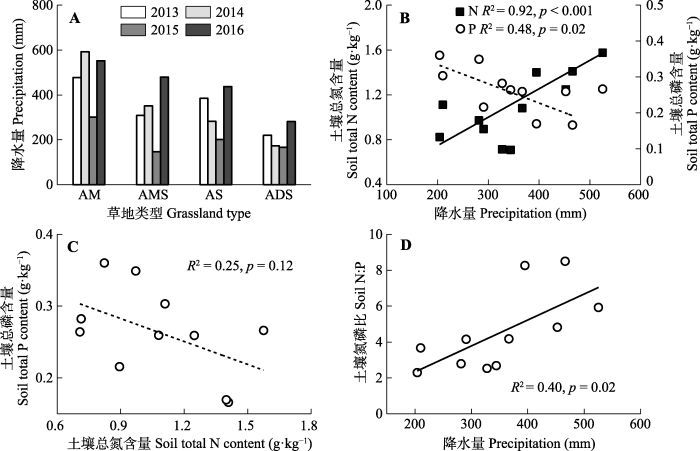 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT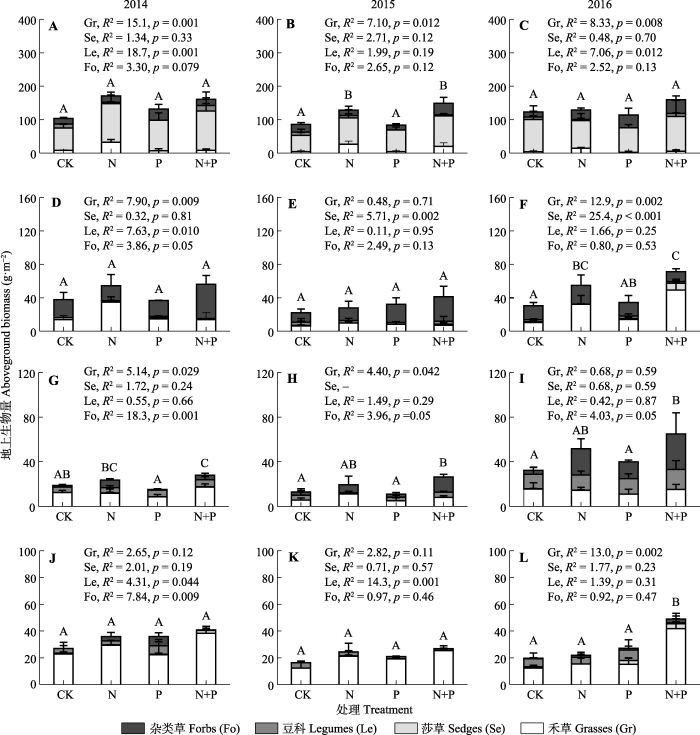 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT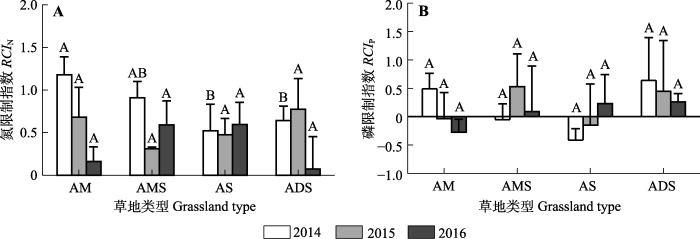 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT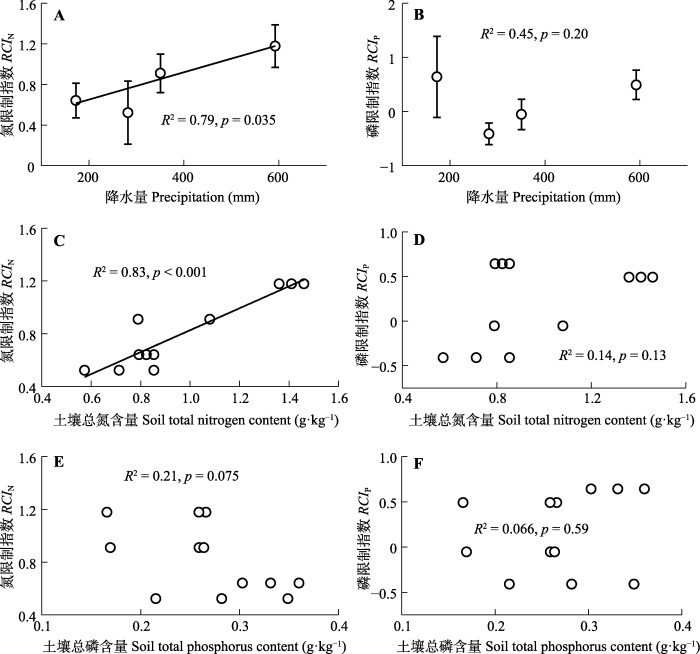 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}