胡朝芹 ,
刘剑宇 ,
王韵茜 ,
杨睿 ,
汪秉琨 ,
何月秋 ,
曾千春 ,
罗琼 * , 云南农业大学农业生物多样性与病虫害控制教育部重点实验室
Hu Chaoqin ,
,
Liu Jianyu ,
,
Wang Yunqian ,
Yang Rui ,
Wang Bingkun ,
He Yueqiu ,
Zeng Qianchun ,
Luo Qiong * , State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan 引用本文 胡朝芹 ,
刘剑宇 ,
王韵茜 ,
杨睿 ,
汪秉琨 ,
何月秋 ,
曾千春 ,
罗琼 . 粳稻子预44抗LP11稻瘟病菌基因()的定位. , 2017, 52(1): 61-69
贡献者
*通讯作者。E-mail: qiongbf@aliyun.com 基金资助
国家重点研发计划(No.2016YFD0100601)和国家自然科学基金(No ;
U1302261, No.31160223) ;
接受日期:2016-06-7接受日期:2016-07-29网络出版日期:2017-01-15
-->Copyright
20172010 《植物学报》编辑部
History
Received :
Accepted :
Online :
摘要: 稻瘟病是世界范围内严重威胁水稻(Oryza sativa )生产可持续发展的主要病害之一, 每年造成10%-30%的水稻产量损失。抗瘟水稻品种的培育和育种利用是解决稻瘟病危害最经济有效的方法。对新的致病性菌株进行分离和筛选是定位与克隆抗病新基因及抗病育种的基础。选择分离自不同稻瘟病发生重灾区的单孢菌株, 对广谱抗瘟水稻子预44和感病水稻江南香糯进行致病性鉴定, 筛选出两材料间致病性差异明显的5个菌株; 进一步利用子预44、湘资3150、9311、日本晴、丽江新团黑谷、中花11、TP309和江南香糯8个抗瘟性不同的水稻材料, 对筛选的菌株进行致病性鉴定。结果显示, LP11能使广谱抗瘟籼稻湘资3150严重发病, 推测其很可能是新进化出来的强致病菌株。利用子预44和江南香糯杂交构建的F2 群体进行抗性遗传分析, 结果表明子预44对LP11菌株的抗性是由单显性基因控制。利用SSR分子标记和图位克隆方法在子预44中定位了1个抗稻瘟病基因Pizy6 (t )。研究结果不仅为抗病相关研究提供了有价值的新菌株, 而且为子预44中抗稻瘟病基因Pizy6 (t )的克隆奠定了基础。 关键词 :
水稻 ;
稻瘟病 ;
抗病基因 ;
基因定位 Abstract :
Rice blast, caused by Magnaporthe oryzae infection, is one of the major diseases threatening the sustainable development of rice (Oryza sativa ) production worldwide. Between 10% and 30% of the annual rice harvest is lost due to infection by the fungus. Development and breeding utilization of rice-resistant resources are considered the most effective and economic means to control rice blast. Isolating new virulent blast strains from the pathotype Oryza is the basis for cloning novel optimal genes resistant to rice blast and rice breeding for disease resistance. In the present study, we isolated and screened blast strains and identified new genes resistant to rice blast. Overall, 5 strains with significantly different pathogenicity between Ziyu44 and Jiangnanxiangnuo (JNXN) were identified. Eight rice varieties, including Ziyu44, Xiangzi3150, 9311, Nipponbare, Lijiangxintuanheigu (LTH), Zhonghua11, JNXN and TP309, were challenged with the 5 blast strains by spray-inoculation at seedling stage. Xiangzi3150, a variety with broad-spectrum blast resistance, showed obvious attacks after inoculation with LP11, so LP11 is probably a newly emerged strong virulent strain. The F2 progenies derived from the cross between Ziyu44 and JNXN were inoculated with the LP11; genetic analysis and gene mapping indicated that the resistance phenotype of Ziyu44 to LP11 was controlled by a single dominant gene, and the resistance gene Pizy6 (t ) was located on rice chromosome 6. These results provided valuable blast strains for genetic studies of disease resistance and laid the foundation for Pizy6 (t ) gene cloning in Ziyu44. Key words :
rice ;
rice blast ;
resistance gene ;
gene mapping 中国有近三分之二的人口以稻米为主食, 水稻(
Oryza sativa )作为中国第一大粮食作物, 在确保中国粮食生产安全中占有重要地位(
邹征欧等, 2016 )。人口持续增长、全球气候变暖等因素致使各种病虫害发生增加, 严重威胁水稻生产及粮食安全(
Cheng et al., 2007 )。稻瘟病是由稻瘟病菌(
Magnaporthe oryzae )引起的水稻真菌病害, 发生在水稻生长的各个时期, 流行年份造成减产10%-20%, 严重时产量损失达到40%-50%, 甚至颗粒无收(
温小红等, 2013 ;
赵家铭等, 2014 ), 严重威胁水稻生产的持续发展。
抗稻瘟病基因的鉴定和育种利用是防治稻瘟病经济环保的措施。但由于稻瘟病菌生理小种复杂且变异快, 一般抗病品种连续种植3-5年后, 其抗性就会减弱, 甚至消失(
Waller and Qu, 1985 )。因此, 发掘新的具有广谱持久抗性的抗病基因对稻瘟病的防治具有重要意义。然而, 无论是鉴定新抗性基因, 还是通过抗谱测定评估一个抗性基因的能效, 或是推测其持久性和抗性变异程度, 都需要大量的稻瘟病菌株作为基础。稻瘟病菌株单胞分离不仅是稻瘟病研究中一项重要的技术, 而且也是获得新稻瘟病菌致病菌株的重要基础工作(
袁桂荣和甘贤友, 1987 ;
张书建和何月秋, 2003 )。
子预44是一份具有广谱持久抗瘟性的云南地方水稻材料(
张锦文等, 2009 ,
2011 ;
李书等, 2014 ;
樊琳琳等, 2015 ;
周镕等, 2015 )。为了鉴定和克隆子预44中的优异抗瘟基因, 揭示子预44的广谱抗瘟分子基础, 我们通过在稻瘟病发病严重的水稻种植区采集穗颈瘟病标样, 采用适当改进的单孢分离方法进行稻瘟病菌株的单孢分离, 在利用广谱持久抗病水稻子预44和谱感水稻江南香糯对分离菌株进行初步的致病性鉴定和筛选的基础上, 用分离自云南罗平的菌株LP11对子预44和江南香糯杂交构建的F
2 群体进行了抗性遗传分析和相关抗瘟基因的初步定位。本研究为抗性遗传研究提供了有价值的新菌株, 为子预44中抗LP11菌株基因的克隆奠定了基础。
1 材料与方法1.1 材料水稻(
Oryza sativa L.)材料子预44、湘资3150、9311、日本晴、丽江新团黑谷、中花11、江南香糯和TP309及子预44与江南香糯杂交的F
2 遗传群体均由本课题组提供。水稻病样于2012-2015年采集自云南宜良、腾冲和罗平、黑龙江哈尔滨及江苏盐城稻瘟病重发地区。36个抗病单基因系材料由云南农业大学何月秋教授提供。
1.2 方法1.2.1 单孢分离和产孢培养
本研究参照张书建和何月秋(2003) 涂平板单孢分离法, 并稍作改进。采用番茄燕麦培养基进行产孢培养。挑取少量的单孢菌丝, 先用PDA培养基对其进行传代培养和纯培养, 得到纯培养物后, 挑取少量菌丝于马铃薯葡萄糖液体培养基中, 26°C、每分钟180转摇菌5-7天。取出菌液, 涂在番茄燕麦培养基上, 每皿吸取500 µL, 在光照培养箱中培养, 26°C昼夜交替光照, 2-3天即可产孢。在超净工作台上, 向培养皿表面注入少量无菌水, 用消毒棉棒刮去菌丝, 经3层擦镜纸过滤, 即可得到大量孢子。
1.2.2 水稻的种植和管理
以腐殖土与云南红土按照1:1 (v/v)的比例混合, 经121°C高温高压灭菌40分钟后备用。选取饱满的种子, 用75%乙醇消毒40秒, 用无菌水浸泡1分钟, 放置在铺1层滤纸片的培养皿中, 加适量无菌水, 35°C催芽2天。待胚芽长至0.5-1 cm, 播于装有种植土的塑料筐中, 注意水肥管理。待水稻长至三叶一心至四叶期, 用喷雾方式接种稻瘟病菌。同时将另一批长至三叶期的水稻移栽到大塑料框中, 待水稻长至抽穗期, 用注射方式接种稻瘟病菌。
1.2.3 稻瘟病菌孢子悬浮液的准备及苗期接种鉴定
在超净工作台上, 用灭菌镊子在酒精灯前挑取-80°C保存的菌株, 置于马铃薯葡萄糖培养基平板上, 放置在28°C恒温箱内黑暗培养。待菌株活化生长后, 用接菌针挑取少量菌丝接种于马铃薯葡萄糖液体培养基中, 在28°C、每分钟180转恒温摇床摇菌培养3-4天。用番茄燕麦培养基产孢法产孢, 在普通光学显微镜下镜检, 将孢子悬浮液的终浓度调至2×10
5 个孢子·mL
-1 , 加5‰明胶配成孢子悬浮液。
当水稻生长至三叶一心时, 用喷雾接种的方式将稻瘟病菌孢子悬浮液均匀接种于水稻叶片上, 将其放入26°C、相对湿度为95%的人工气候箱中, 黑暗处理24小时。保持温度和湿度不变, 光照培育12小时, 然后再黑暗培育12小时。如此循环, 培育7-10天。依据《云南省水稻区域性试验材料稻瘟病抗性鉴定分级标准》调查发病情况。
1.2.4 幼苗基因组DNA提取
根据抗性调查结果, 收集F
2 全部感病单株和20株抗病单株以及适量亲本材料。参照Murray和Thompson (1980) CTAB法分单株提取水稻幼苗基因组DNA, 经1%琼脂糖凝胶电泳检测备用。
1.2.5 PCR引物设计与合成
根据Gramene (http://www.gramene.org/)和Interna- tional Rice Genome Sequencing Project (
International Rice Genome Sequencing Project, 2005 )提供的序列由华大基因公司或硕擎生物科技有限公司合成SSR引物。
Indel引物根据NCBI (https://www.ncbi.nlm.nih. gov/)提供的籼稻和粳稻的基因组序列进行BLAST比对分析, 查找两基因组之间存在插入缺失差异的位点, 根据这些差异位点利用DNASTAR软件设计PCR引物, 由华大基因公司或硕擎生物科技有限公司合成。
1.2.6 PCR扩增及产物检测
PCR扩增体系: 2.0 µL 10×PCR Buffer, 1.6 µL dNTP (2.5 mmol·L
-1 ), 0.2 µL Easy
Taq 酶(TransGen, 5 U·µL
-1 ), 2.0 µL基因组DNA (10 ng), 2.0 µL引物(10 ng·µL
-1 ), 加ddH
2 O补足至15 µL。PCR反应程序: 94 °C预变性5分钟; 94°C变性40秒, 55-60°C (依据引物的Tm值稍作调整)退火40秒, 72°C延伸40秒, 30个循环; 72°C延伸10分钟。PCR产物用3%-4%的琼脂糖凝胶以及12%聚丙烯酰胺凝胶电泳检测。
1.2.7 稻瘟病菌株LP11和LP33的基因组DNA提取及无毒基因分析
将稻瘟病菌株LP11和LP33分别接种于PDA液体培养基中, 于26°C、每分钟180转振荡培养3天后, 用滤纸过滤收集菌体。用无菌水洗涤菌丝体2次, 尽量压干水分, 然后转移到30°C烘箱烘干。取烘干的菌丝体用液氮研成粉末, 分装于2 mL的EP管中。采用CTAB法提取稻瘟病菌株LP11和LP33的基因组DNA。
无毒基因及PCR扩增引物参考Huang等(2014) 的方法(
表1 )。PCR扩增体系: 10 μL 2xGC Buffer I (Takara), 2 μL dNTP (2.5 mmol·L
-1 , Takara), 0.2 μL Easy
Taq 酶(TransGen), 1.0 μL引物(10 μmol·µL
-1 ), 2 μL DNA模板(10 ng), 加ddH
2 O补足至20 μL。PCR反应程序: 94°C预变性5分钟; 94°C30秒, 55°C (依据引物的Tm值稍作调整)30秒, 72°C1分钟, 30个循环; 72°C延伸10分钟。PCR产物用1%琼脂糖凝胶电泳检测。
表1 Table 1 表1 表1 无毒基因及其PCR引物
Table 1 Avr genes and primers used in this study
Avr genesPrimers Sequence (5'-3') AVR_Pita F GACCCGTTTCCGCCTTTATT R GATTCCCTCCATTCCAACAC AVR_PWL2 F CTCCGCCACTTTTCTCATTC R GCCCTCTTCTCGCTGTTCAC AVR_Pik F GTCAACCAAGCGTAAACCTC R CGATTCAGAAGTTAGGCATT AVR_pii F GAGGCCGATATGTTACGATT R CTCTGCTCTCACGCTTTACC AVR_Pia F GCCGCTAGCTGTATAGACAA R TCATCGTCGAGTGGTGTAGG AVR_ACE1 F GAGGTGCCAGATATGTCGTC R GGATGAGCAGATGAGCAACA
表1
无毒基因及其PCR引物
Table 1
Avr genes and primers used in this study
2 结果与讨论2.1 稻瘟病菌株的分离和筛选
菌株分离参照张书建和何月秋(2003) 的涂平板单孢分离法, 并作适当改进。把滤纸折成扇形, 以便更好地控制病组织表面的湿度和减少杂菌的生长。每个培养皿(直径9 cm)的水琼脂精确控制在5-7 mL, 使水琼脂平板的厚度控制在2-3 mm, 便于涂板, 同时透明度好, 能很清楚地在体视显微镜下看到孢子, 便于进行单孢挑取。采用改进后的孢子涂板单孢分离法, 利用采集自黑龙江哈尔滨、江苏盐城和云南宜良的病组织进行单孢分离, 成功地从上述不同稻区的病组织中分离约500份单孢, 经过进一步活化培养, 保存63份有效单孢菌株。
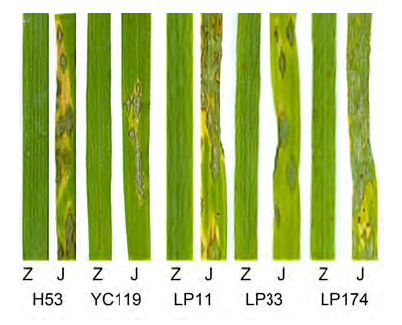
在云南农业大学工程中心温室, 分期播种水稻子预44和江南香糯。随机选取滤纸片保存的黑龙江哈尔滨、江苏盐城、云南宜良、罗平和腾冲(何月秋教授提供)单孢菌株各5个, 共25个菌株, 进行分批活化和产孢培养, 制备孢子悬浮液, 喷雾接种于3-4叶期子预44和江南香糯水稻幼苗。最后成功筛选出在子预44和江南香糯间致病性差异明显的菌株5株, 其中来自黑龙江哈尔滨、江苏盐城、云南罗平分别为1株、1株和3株(
图1 )。
图1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_1.png <b>图1</b> 子预44和江南香糯水稻接种不同稻瘟病菌株^Z: 子预44; J: 江南香糯; H53: 哈尔滨53号菌株; YC119: 盐城119号菌株; LP11: 罗平11号菌株; LP33: 罗平33号菌株; LP174: 罗平174号菌株<br/><b>Figure 1</b> Ziyu44 and JNXN were inoculated with different rice blast strains^Z: Ziyu44; J: JNXN; H53: No.53 strain from Harbin; YC119: No.119 strain from Yancheng; LP11: No.11 strain from Luo- ping; LP33: No.33 strain from Luoping; LP174: No.174 strain from Luoping Figure 1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_1.png <b>图1</b> 子预44和江南香糯水稻接种不同稻瘟病菌株^Z: 子预44; J: 江南香糯; H53: 哈尔滨53号菌株; YC119: 盐城119号菌株; LP11: 罗平11号菌株; LP33: 罗平33号菌株; LP174: 罗平174号菌株<br/><b>Figure 1</b> Ziyu44 and JNXN were inoculated with different rice blast strains^Z: Ziyu44; J: JNXN; H53: No.53 strain from Harbin; YC119: No.119 strain from Yancheng; LP11: No.11 strain from Luo- ping; LP33: No.33 strain from Luoping; LP174: No.174 strain from Luoping 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
子预44和江南香糯水稻接种不同稻瘟病菌株^Z: 子预44; J: 江南香糯; H53: 哈尔滨53号菌株; YC119: 盐城119号菌株; LP11: 罗平11号菌株; LP33: 罗平33号菌株; LP174: 罗平174号菌株
Figure 1
Ziyu44 and JNXN were inoculated with different rice blast strains^Z: Ziyu44; J: JNXN; H53: No.53 strain from Harbin; YC119: No.119 strain from Yancheng; LP11: No.11 strain from Luo- ping; LP33: No.33 strain from Luoping; LP174: No.174 strain from Luoping
2.2 菌株致病性进一步鉴定子预44、湘资3150和9311是抗谱广、抗性稳定且持久的抗稻瘟品种(
胡标林等, 2006 ;
张锦文等, 2009 ,
2011 ;
黄红梅等, 2011 ;
李书等, 2014 ;
樊琳琳等, 2015 )。丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(
伍尚忠等, 2004 )。中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(
朱旭东等, 2003 )。利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(
图2 )。结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机。
图2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_2.png <b>图2</b> 8个水稻材料接种稻瘟病菌株LP11^Z: 子预44; J: 江南香糯; 9: 9311; H: 中花11; N: 日本晴; X: 湘资3150; L: LTH; T: TP309<br/><b>Figure 2</b> Eight rice varieties were inoculated with <i>Magnaporthe oryzae</i> LP11^Z: Ziyu44; J: JNXN; 9: 9311; H: Zhonghua11; N: Nipponbare; X: Xiangzi3150; L: LTH; T: TP309 Figure 2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_2.png <b>图2</b> 8个水稻材料接种稻瘟病菌株LP11^Z: 子预44; J: 江南香糯; 9: 9311; H: 中花11; N: 日本晴; X: 湘资3150; L: LTH; T: TP309<br/><b>Figure 2</b> Eight rice varieties were inoculated with <i>Magnaporthe oryzae</i> LP11^Z: Ziyu44; J: JNXN; 9: 9311; H: Zhonghua11; N: Nipponbare; X: Xiangzi3150; L: LTH; T: TP309 在新窗口打开 下载 下载原图ZIP 生成PPT 图2
8个水稻材料接种稻瘟病菌株LP11^Z: 子预44; J: 江南香糯; 9: 9311; H: 中花11; N: 日本晴; X: 湘资3150; L: LTH; T: TP309
Figure 2
Eight rice varieties were inoculated with
Magnaporthe oryzae LP11^Z: Ziyu44; J: JNXN; 9: 9311; H: Zhonghua11; N: Nipponbare; X: Xiangzi3150; L: LTH; T: TP309
2.3 子预44对菌株LP11的抗性遗传分析为了进一步鉴定子预44中抗稻瘟病菌株LP11的抗瘟基因, 我们进一步利用子预44与江南香糯杂交构建的F
2 群体, 对子预44抗LP11菌株的抗性遗传进行了分析。在水稻幼苗3-4叶期利用稻瘟病菌株LP11孢子悬浮液喷雾接种日本晴、9311、亲本子预44、江南香糯及其杂交F
2 代群体。结果表明, 亲本子预44 (25株)和9311 (10株)全部表现高抗, 亲本江南香糯(25株)和日本晴(14株)全部表现高感, 与之前的致病性鉴定结果一致(
图3 )。F
2 代群体共489株, 其中抗病单株370株, 感病单株119株; X
2 c<X
2 0.05(1)=3.84, 符合3:1的抗感分离比(
表2 )。结果表明, 子预44对稻瘟病菌株LP11的抗性属于单基因控制的显性遗传。前期本研究组通过图位克隆方法已在子预44中鉴定了
Pizy (
t )/
Pizy1 (
t ) (
张锦文等, 2009 )、
Pizy2 (
t ) (待发表)、
Pizy3 (
t ) (
樊琳琳等, 2015 )、
Pizy4 (
t ) (待发表)及
Pizy5 (
t ) (待发表), 因此将该基因定名为
Pizy6 (
t )。
图3 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_3.png <b>图3</b> 子预44、江南香糯及其F<sub>2</sub>群体单株喷雾接种稻瘟病菌LP11^Z: 子预44; J: 江南香糯; N: 日本晴; 9: 9311; R: F<sub>2</sub>群体中的抗病单株; S: F<sub>2</sub>群体中的感病单株<br/><b>Figure 3</b> Ziyu44, JNXN and the F<sub>2 </sub>progenies were inoculated with <i>Magnaporthe oryzae</i> LP11 by spraying^Z: Ziyu44; J: JNXN; N: Nipponbare; 9: 9311; R: The resistant individual plants of F<sub>2</sub> population; S: The susceptible individu- al plants of F<sub>2 </sub>population Figure 3 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_3.png <b>图3</b> 子预44、江南香糯及其F<sub>2</sub>群体单株喷雾接种稻瘟病菌LP11^Z: 子预44; J: 江南香糯; N: 日本晴; 9: 9311; R: F<sub>2</sub>群体中的抗病单株; S: F<sub>2</sub>群体中的感病单株<br/><b>Figure 3</b> Ziyu44, JNXN and the F<sub>2 </sub>progenies were inoculated with <i>Magnaporthe oryzae</i> LP11 by spraying^Z: Ziyu44; J: JNXN; N: Nipponbare; 9: 9311; R: The resistant individual plants of F<sub>2</sub> population; S: The susceptible individu- al plants of F<sub>2 </sub>population 在新窗口打开 下载 下载原图ZIP 生成PPT 图3
子预44、江南香糯及其F
2 群体单株喷雾接种稻瘟病菌LP11^Z: 子预44; J: 江南香糯; N: 日本晴; 9: 9311; R: F
2 群体中的抗病单株; S: F
2 群体中的感病单株
Figure 3
Ziyu44, JNXN and the F
2 progenies were inoculated with
Magnaporthe oryzae LP11 by spraying^Z: Ziyu44; J: JNXN; N: Nipponbare; 9: 9311; R: The resistant individual plants of F
2 population; S: The susceptible individu- al plants of F
2 population
表2 Table 2 表2 表2 子预44/江南香糯F
2 群体对LP11的抗性分离
Table 2 Segregation ratios of resistance to blast isolate LP11 in the F
2 population of Ziyu44/JNXN
Parents and F2 population Number of total plants Number of R plants Number of S plants Expratio (R:S) Χ2 X2 0.05(1) JNXN 25 0 25 9311 10 10 0 Nipponbare 14 0 14 Ziyu44/JNXN F2 489 370 119 3:1 0.012 3.84
R: Resistant; S: Susceptible R: 抗病; S: 感病 表2
子预44/江南香糯F
2 群体对LP11的抗性分离
Table 2
Segregation ratios of resistance to blast isolate LP11 in the F
2 population of Ziyu44/JNXN
2.4
Pizy6 (
t )基因的染色体定位遗传分析结果表明, 子预44对稻瘟病菌株LP11的抗性属于单基因控制的显性遗传。我们进一步利用SSR分子标记和图位克隆法对子预44中
Pizy6 (
t )基因进行了定位。首先在水稻12条染色体的长臂和短臂上各选取1对多态性较好的引物对子预44、江南香糯及其F
2 代群体中的感病单株进行性状连锁分析。结果表明, 位于第6号染色体短臂上的RM276 (物理位置: 6.23 Mb) (
图4 )和长臂上的RM3628 (物理位置: 24.61 Mb) (
图5 )与抗感性状存在明显的连锁关系。
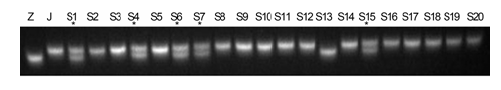
图4 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_4.png <b>图4</b> RM276在接种LP11的水稻F<sub>2</sub>群体中的分离^Z: 子预44; J: 江南香糯; S1-S20: F<sub>2</sub>群体中的感病单株; * : 重组单株<br/><b>Figure 4</b> Segregation of RM276 in F<sub>2</sub> population of rice treated by LP11^Z: Ziyu44; J: JNXN; S1-S20: Susceptible plants of F<sub>2</sub> population; * : Recombinant plants Figure 4 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_4.png <b>图4</b> RM276在接种LP11的水稻F<sub>2</sub>群体中的分离^Z: 子预44; J: 江南香糯; S1-S20: F<sub>2</sub>群体中的感病单株; * : 重组单株<br/><b>Figure 4</b> Segregation of RM276 in F<sub>2</sub> population of rice treated by LP11^Z: Ziyu44; J: JNXN; S1-S20: Susceptible plants of F<sub>2</sub> population; * : Recombinant plants 在新窗口打开 下载 下载原图ZIP 生成PPT 图4
RM276在接种LP11的水稻F
2 群体中的分离^Z: 子预44; J: 江南香糯; S1-S20: F
2 群体中的感病单株; * : 重组单株
Figure 4
Segregation of RM276 in F
2 population of rice treated by LP11^Z: Ziyu44; J: JNXN; S1-S20: Susceptible plants of F
2 population; * : Recombinant plants
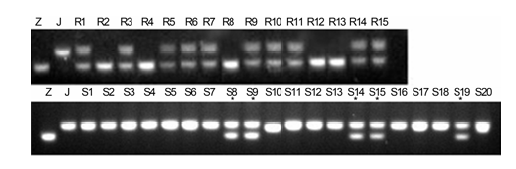
图5 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_5.png <b>图5</b> RM3628在接种LP11的水稻F<sub>2</sub>群体中的分离^Z: 子预44; J: 江南香糯; R1-R15: F<sub>2</sub>群体中的抗病单株; S1-S20: F<sub>2</sub>群体中的感病单株; * : 重组单株<br/><b>Figure 5</b> Segregation of RM3628 in F<sub>2</sub> population of rice treated by LP11^Z: Ziyu44; J: JNXN; R1-R15: Resistant plants of F<sub>2</sub> population; S1-S20: Susceptible plants of F<sub>2</sub> population; * : Recombinant plants Figure 5 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_5.png <b>图5</b> RM3628在接种LP11的水稻F<sub>2</sub>群体中的分离^Z: 子预44; J: 江南香糯; R1-R15: F<sub>2</sub>群体中的抗病单株; S1-S20: F<sub>2</sub>群体中的感病单株; * : 重组单株<br/><b>Figure 5</b> Segregation of RM3628 in F<sub>2</sub> population of rice treated by LP11^Z: Ziyu44; J: JNXN; R1-R15: Resistant plants of F<sub>2</sub> population; S1-S20: Susceptible plants of F<sub>2</sub> population; * : Recombinant plants 在新窗口打开 下载 下载原图ZIP 生成PPT 图5
RM3628在接种LP11的水稻F
2 群体中的分离^Z: 子预44; J: 江南香糯; R1-R15: F
2 群体中的抗病单株; S1-S20: F
2 群体中的感病单株; * : 重组单株
Figure 5
Segregation of RM3628 in F
2 population of rice treated by LP11^Z: Ziyu44; J: JNXN; R1-R15: Resistant plants of F
2 population; S1-S20: Susceptible plants of F
2 population; * : Recombinant plants
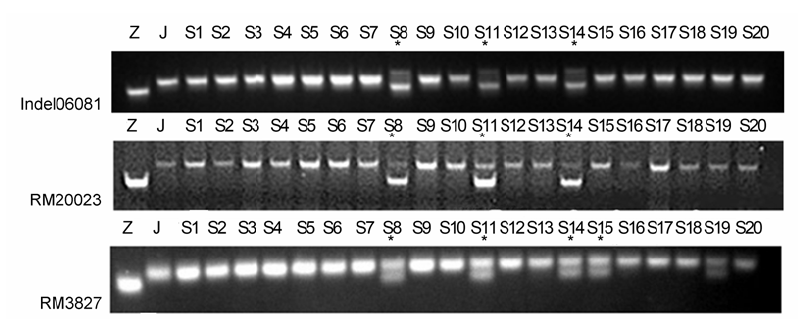
进一步利用第6号染色体RM276与RM3628之间的Indel (Indel06081: F-GGCCTATACATCCAGCT- GAGT; R-ACAGGTGTGTACGGAAATTATCT; 物理位置: 8.73 Mb)及SSR多态性引物RM20023 (物理位置: 16.71 Mb)、RM3827 (物理位置: 23.17 Mb) (
图6 )对F
2 群体的感病单株进行连锁分析, 结果表明, 从染色体长臂SSR标记RM3827到短臂Indel06081, 交换单株趋势一致且数量逐渐减少, 但是Indel06081与RM276的交换单株不一致。而染色体短臂的SSR标记RM3805 (2.85 Mb)到RM276交换单株趋势也一致且数量渐减。由此可将
Pizy6 (
t )基因定位在水稻第6号染色体短臂RM276与Indel06081之间大约2.5 Mb范围内。
图6 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_6.png <b>图6</b> Indel06081、RM20023和RM3827在接种LP11的水稻F<sub>2</sub>群体中的分离^Z: 子预44; J: 江南香糯; S1-S20: F<sub>2</sub>群体中的感病单株; *: 重组单株<br/><b>Figure 6</b> Segregation of Indel06081, RM20023 and RM3827 in F<sub>2</sub> population of rice treated by LP11^Z: Ziyu44; J: JNXN; S1-S20: Susceptible plants of F<sub>2</sub> population; * : Recombinant plants Figure 6 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_6.png <b>图6</b> Indel06081、RM20023和RM3827在接种LP11的水稻F<sub>2</sub>群体中的分离^Z: 子预44; J: 江南香糯; S1-S20: F<sub>2</sub>群体中的感病单株; *: 重组单株<br/><b>Figure 6</b> Segregation of Indel06081, RM20023 and RM3827 in F<sub>2</sub> population of rice treated by LP11^Z: Ziyu44; J: JNXN; S1-S20: Susceptible plants of F<sub>2</sub> population; * : Recombinant plants 在新窗口打开 下载 下载原图ZIP 生成PPT 图6
Indel06081、RM20023和RM3827在接种LP11的水稻F
2 群体中的分离^Z: 子预44; J: 江南香糯; S1-S20: F
2 群体中的感病单株; *: 重组单株
Figure 6
Segregation of Indel06081, RM20023 and RM3827 in F
2 population of rice treated by LP11^Z: Ziyu44; J: JNXN; S1-S20: Susceptible plants of F
2 population; * : Recombinant plants
2.5
Pizy6 (
t )的定位区间与已知的抗稻瘟病位点的关系利用SSR分子标记初步将
Pizy6 (
t )基因定位在水稻第6号染色体短臂RM276 (物理位置: 6.23 Mb)与Indel06081 (物理位置: 8.73 Mb)之间大约2.5 Mb范围内。该基因与通过分子标记鉴定的位于水稻第6号染色体短臂上其它6个位点上的10个抗瘟主效基因在染色体上的位置不同, 它们是
Pi22 (4.89-6.02 Mb)、
Pi26 (8.75-11.67 Mb)、
Pi27 (
t ) (0.74-5.55 Mb)、
Pi40 (16.27-17.53 Mb)、
Pid3 /
Pi25 (13.05 Mb)和
Piz (10.15-10.51 Mb)位点上的复等位基因
Pi2 /
Pizt /
Pi9 /
Pigm (10.36-10.42 Mb) (
Zhou et al., 2006 ;
Chen et al., 2011 ;
Sharma et al., 2012 ;
Tanweer et al., 2015 )。由此可见,
Pizy6 (
t )可能是一个新的抗稻瘟病基因。
表3 Table 3 表3 表3 LP11和LP33菌株的36个抗病单基因系鉴定
Table 3 Pathogenic identification of LP11 and LP33 with 36 single gene lines of blast resistance
Variety/R- gene LP11 LP33 Variety/R- gene LP11 LP33 LTH/- 4 3 IRBL-7/Pi-kp 4 1 F80-1/Pi-k 4 2 IRBL-8/Pi-kh 3 1 F98-7/Pi-km 4 2 IRBL-9/Pi-z 3 4 F124-1/Pi-ta 4 1 IRBL-10/Pi-z5 4 3 F128-1/Pi-ta2 2 0 IRBL-11/Pi-zt 4 2 F129/Pi-kp 4 3 IRBL-12/Pi-ta 4 4 F145-2/Pi-b 4 2 IRBL-13/Pi-ta 4 4 C039/Pi-CO39 4 0 IRBL-14/Pi-b 3 1 C101A51/Pi-2 3 1 IRBL-15/Pi-t 4 2 C101LAC/Pi-1 3 1 IRBL-16/Pi-sh 3 1 C101PKT/Pi-4a 2 1 IRBL-17/Pi-sh 4 2 C104PKT/Pi-3 4 2 IRBL-18/Pi-1 3 1 IRBL-1/Pi-a (1 ) 4 1 IRBL-19/Pi-3 3 2 IRBL-2/Pi-a (2 ) 4 2 IRBL-20/Pi-5 (t ) 3 1 IRBL-3/Pi-i 3 1 IRBL-21/Pi-7 (t ) 4 1 IRBL-4/Pi-ks 3 0 IRBL-22/Pi-9 (t ) 2 1 IRBL-5/Pi-ks 4 1 IRBL-23/Pi-12 (t ) 2 0 IRBL-6/Pi-k 4 2 IRBL-24/Pi-19 (t ) 3 1
The identification result was provided by professor He Yueqiu in May 2013. LTH: Lijiangxintuanheigu, a susceptible material without any R genes. Disease severity was scored based on 5 classes: 0 represents without lesion; 1 represents lesions seldom (less than 5) and small (less than 1 cm); 2 represents multiple small lesions (more than 5); 3 represents multiple big lesions; 4 represents death of whole plant. 数据为2013年5月温室鉴定结果, 由何月秋教授提供。LTH: 丽江新团黑谷, 不含任何抗病基因的感病材料。不同材料发病情况分为5级: 0代表无病斑; 1代表叶片病斑少(少于5个)而小(小于1 cm); 2代表叶片病斑小而多(多于5个); 3代表叶片病斑大而多; 4代表全叶病枯。 表3
LP11和LP33菌株的36个抗病单基因系鉴定
Table 3
Pathogenic identification of LP11 and LP33 with 36 single gene lines of blast resistance
本课题组曾利用稻瘟病菌株LP33在子预44中定位了第6号染色体RM276与RM3827之间(即6.23- 23.17 Mb)的主效抗稻瘟病基因
Pizy3 (
t ) (
樊琳琳等, 2015 ),
Pizy6 (
t )基因的定位区间与
Pizy3 (
t )基因部分重叠。我们前期的研究结果显示, LP11和LP33菌株对36个抗病单基因系的致病性存在明显差异(
表3 ), 推测这2个菌株很可能携带不同的无毒基因。参照Yang等(2013)、Huang等(2014)和Guo等(2016)的方法, 我们利用6个已克隆的无毒基因特异引物对2个菌株进行分析, 证实2个菌株携带的无毒基因确实存在差异(
图7 )。同时, 我们利用子预44、湘资3150、9311、中花11、日本晴、TP309、丽江新团黑谷和江南香糯8个水稻材料, 通过苗期喷雾接种对LP11和LP33的致病性进行比较研究, 结果表明, LP11和LP33对抗病水稻材料9311的致病性明显不同(
图8 )。我们将利用2个致病性不同的稻瘟病菌株LP11和LP33对含目标基因的群体材料进行深入研究以明确
Pizy3 (
t )和
Pizy6 (
t )的关系。
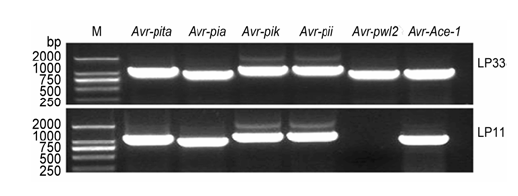
图7 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_7.png <b>图7</b> 不同无毒基因位点在稻瘟病菌株LP33和LP11中的PCR扩增<br/><b>Figure 7</b> PCR amplification of <i>Avr</i> genes in LP33 and LP11 strains Figure 7 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_7.png <b>图7</b> 不同无毒基因位点在稻瘟病菌株LP33和LP11中的PCR扩增<br/><b>Figure 7</b> PCR amplification of <i>Avr</i> genes in LP33 and LP11 strains 在新窗口打开 下载 下载原图ZIP 生成PPT 图7
不同无毒基因位点在稻瘟病菌株LP33和LP11中的PCR扩增
Figure 7
PCR amplification of
Avr genes in LP33 and LP11 strains
图8 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_8.png <b>图8</b> LP11和LP33对8个水稻品种的致病性^Z、9、X、H、N、T、L和J同<xref ref-type="fig" rid="F2-1674-3466-52-1-61">图2</xref>。<br/><b>Figure 8</b> The pathogenicity of LP11 and LP33 to 8 rice varieties^Z, 9, X, H, N, T, L and J see <xref ref-type="fig" rid="F2-1674-3466-52-1-61">Figure 2</xref>. Figure 8 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-61/img_8.png <b>图8</b> LP11和LP33对8个水稻品种的致病性^Z、9、X、H、N、T、L和J同<xref ref-type="fig" rid="F2-1674-3466-52-1-61">图2</xref>。<br/><b>Figure 8</b> The pathogenicity of LP11 and LP33 to 8 rice varieties^Z, 9, X, H, N, T, L and J see <xref ref-type="fig" rid="F2-1674-3466-52-1-61">Figure 2</xref>. 在新窗口打开 下载 下载原图ZIP 生成PPT 图8
LP11和LP33对8个水稻品种的致病性^Z、9、X、H、N、T、L和J同
图2。Figure 2.Pib 被克隆, 到2006年7年间只克隆了1个抗稻瘟病基因Pita 。从2006年开始, 抗稻瘟病基因的克隆数量大规模增长, 然而从已经克隆的抗稻瘟病基因位点我们发现, 从2009年开始, 尽管抗稻瘟病基因的数目在增加, 但真正的抗稻瘟病基因位点数却增加很少, 后来克隆的抗稻瘟病基因大多数都是已经克隆的等位或是直系同源基因(Xu et al., 2014 )。由于稻瘟病菌群体具复杂性和易变性, 我们每年从感病田块采集新的稻瘟病标样, 分离新的强致病性菌株, 并定位和克隆到具有最佳时效性的抗病新基因。黑龙江哈尔滨、江苏盐城、云南宜良、罗平和腾冲是近年来稻瘟病发生较严重的稻作区, 我们采用稻瘟病抗性不同的水稻材料, 对新分离自这些稻瘟病流行稻作区病样的稻瘟病菌单孢菌株进行致病性鉴定和新出现的强致病菌株筛选, 并利用筛选出的强致病菌株LP33和LP11, 利用图位克隆法在广谱抗瘟水稻子预44中定位了Pizy3 (t ) (樊琳琳等, 2015 )和Pizy6 (t )两个不同的稻瘟病抗性基因。上述研究结果表明, 从栽培品种多样、利于稻瘟病菌繁殖活动、田间稻瘟病菌组成复杂且信息流强度大的稻作区分离和筛选新的致病性菌株用于基因定位克隆, 是鉴定抗瘟新基因的有效方法。Pib 、Pi9 和Pid3 (李书等, 2014 )。利用不同稻瘟病菌株, 通过图位克隆法鉴定了位于第11号染色体的Pizy (t )/Pizy1 (t ) (张锦文等, 2009 )、位于第6号染色体的Pizy2 (t ) (待发表)、Pizy3 (t ) (樊琳琳等, 2015 )、Pizy4 (t )和Pizy5 (t ) (待发表)以及本研究中定位的Pizy6 (t )。上述研究结果表明, 广谱持久抗瘟水稻子预44携带多个主效和微效抗瘟基因, 其中主效抗稻瘟病基因具有集中成簇分布在第6号染色体上的特征, 他们对维持子预44的广谱持久抗性发挥着重要作用。此外, 根据基因预测的结 果, 水稻第6号染色体上还有许多未被鉴定的抗病 基因(http://rice.plantbiology.msu.edu/)。相关基因 的进一步鉴定和分子标记的发展, 对利用子预44进行广谱持久抗瘟水稻品种的分子模块育种具有重要意义。The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 樊琳琳 , 姚波 , 刘志涛 , 曾千春 , 罗琼 (2015 ). 子预44中抗稻瘟病基因Pi-zy3(t)的定位. 分子植物育种 13, 961 -967 .[2] 胡标林 , 谢建坤 , 包劲松 , 余守武 , 张铮 (2006 ). 籼稻9311辐射突变体的分离与鉴定. 分子植物育种 4, 181 -188 .[3] 黄红梅 , 肖应辉 , 黄玲 , 奉光平 , 燕玮婷 , 戴良英 , 王国梁 , 刘二明 (2011 ). 湘资3150微效抗瘟性基因鉴定. 植物病理学报 41, 509 -515 .[4] 李书 , 李权 , 樊琳琳 , 沙莎 , 曾千春 , 罗琼 (2014 ). 高原粳稻子预44中三个稻瘟病抗性基因的假基因化分子标记鉴定. 分子植物育种 12, 219 -225 .[5] 温小红 , 谢明杰 , 姜健 , 杨宝灵 , 邵艳龙 , 何伟 , 刘丽 , 赵毅 (2013 ). 水稻稻瘟病防治方法研究进展. 中国农学通报 29, 190 -195 .[6] 伍尚忠 , 朱小源 , 刘斌 , 杨祁云 , 张少红 , Leung H 2004 ). 籼稻品种三黄占2号的稻瘟病持久抗性评价与遗传分析. 中国农业科学 37, 528 -534 .[7] 袁桂荣 , 甘贤友 (1987 ). 一种有效的真菌单孢分离法. 河南职技师院学报 15, 70 -71 .[8] 张锦文 , 洪汝科 , 范静华 , 张祎颖 , 曾千春 , 罗琼 (2011 ). 一份云南地方稻广谱持久抗稻瘟病初步分析. 西南农业学报 24, 1323 -1326 .[9] 张锦文 , 谭亚玲 , 洪汝科 , 范静华 , 罗琼 , 曾千春 (2009 ). 高原粳稻子预44抗稻瘟病基因遗传分析和定位. 中国水稻科学 23, 31 -35 .[10] 张书建 , 何月秋 (2003 ). 介绍一种简单的真菌单孢子分离法. 云南农业大学学报 18, 315 -316 .[11] 赵家铭 , 张丽霞 , 郑文静 (2014 ). 稻瘟病抗病基因定位及克隆研究进展. 辽宁农业科学 2, 47 -49 .[12] 周镕 , 王波 , 杨睿 , 李书 , 樊琳琳 , 曾千春 , 罗琼 (2015 ). 粳稻子预44中稻瘟病数量抗性位点分析. 植物学报 50, 691 -698 .[13] 朱旭东 , 陈红旗 , 罗达 , 张建军 , 方红民 , 闵绍楷 (2003 ). 水稻中花11辐射突变体的分离与鉴定. 中国水稻科学 17, 205 -210 .[14] 邹征欧 , 阳春瑜 , 李闻名 , 杨立明 (2016 ). 衡阳市水稻生产的现状与发展对策. 作物研究 2, 186 -189 .[15] Chen J Shi YF Liu WZ Chai RY Fu YP Zhuang JY Wu [16] JL (2011 ). A Pid3 allele from rice cultivar Gumei2 confers resistance to Magnaporthe oryzae. J Genet Genomics 38, 209 -216 .[17] Cheng SH Cao LY Zhuang JY Chen SG Zhan XD Fan YY Zhu DF Peng SB 2007 ). Super hybrid rice breeding in China: achievements and prospects. J Integr Plant Biol 49, 805 -810 .[18] Guo CJ Sun XG Chen X Yang SH Li J Wang L Zhang XH 2016 ). Cloning of novel rice blast resistance genes from two rapidly evolving NBS-LRR gene families in rice.Plant Mol Biol 90, 95 -105 .[19] Huang J Si W Deng QM Li P Yang SH 2014 ). Rapid evolution of avirulence genes in rice blast fungus Magnaporthe oryzae.BMC Genet 15, 45 .[20] International Rice Genome Sequencing Project (2005 ). The map-based sequence of the rice genome.Nature 436, 793 -800 .[21] Murray MG Thompson WF 1980 ). Rapid isolation of high molecular weight plant DNA.Nucleic Acids Res 8, 4321 -4325 .[22] Sharma TR Rai AK Gupta SK Vijayan J Devanna BN Ray S 2012 ). Rice blast management through host-plant resistance: retrospect and prospects.Agric Res 1, 37 -52 .[23] Tanweer FA Rafii MY Sijam K Rahim HA Ahmed F Latif MA 2015 ). Current advance methods for the identification of blast resistance genes in rice.Plant Biol Pathol 338, 321 -334 .[24] Waller JM Qu SH 1985 ). Rice Diseases (2nd). London: Common Wealth Agricultural Bureau. pp.109 -201 .[25] Xu X Lv QM Shang JJ Pang ZQ Zhou ZZ Wang J Jiang GH Tao Y Xu Q Li XB Zhao XF Li SG Xu JC Zhu LH 2014 ). Excavation of Pid3 orthologs with differential resistance spectra to Magnaporthe oryzae in rice resource. PLoS One 9, e93275 .[26] Yang SH Li J Zhang XH Zhang QJ Huang J Chen JQ Hartl DL Tian DC 2013 ). Rapidly evolving R genes in diverse grass species confer resistance to rice blast disease.Proc Natl Acad Sci USA 110, 18572 -18577 .[27] Zhou B Qu SH Liu GF Dolan M Sakai H Lu GD Bellizzi M Wang GL 2006 ). The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea.Mol Plant Microbe Interact 19, 1216 -1228 .
张锦文等, 2009 , 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ; 周镕等, 2015 ).为了鉴定和克隆子预44中的优异抗瘟基因, 揭示子预44的广谱抗瘟分子基础, 我们通过在稻瘟病发病严重的水稻种植区采集穗颈瘟病标样, 采用适当改进的单孢分离方法进行稻瘟病菌株的单孢分离, 在利用广谱持久抗病水稻子预44和谱感水稻江南香糯对分离菌株进行初步的致病性鉴定和筛选的基础上, 用分离自云南罗平的菌株LP11对子预44和江南香糯杂交构建的F2 群体进行了抗性遗传分析和相关抗瘟基因的初步定位.本研究为抗性遗传研究提供了有价值的新菌株, 为子预44中抗LP11菌株基因的克隆奠定了基础. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...2 群体, 对子预44抗LP11菌株的抗性遗传进行了分析.在水稻幼苗3-4叶期利用稻瘟病菌株LP11孢子悬浮液喷雾接种日本晴、9311、亲本子预44、江南香糯及其杂交F2 代群体.结果表明, 亲本子预44 (25株)和9311 (10株)全部表现高抗, 亲本江南香糯(25株)和日本晴(14株)全部表现高感, 与之前的致病性鉴定结果一致(图3 ).F2 代群体共489株, 其中抗病单株370株, 感病单株119株; X2 c<X2 0.05(1)=3.84, 符合3:1的抗感分离比(表2 ).结果表明, 子预44对稻瘟病菌株LP11的抗性属于单基因控制的显性遗传.前期本研究组通过图位克隆方法已在子预44中鉴定了Pizy (t )/Pizy1 (t ) (张锦文等, 2009 )、Pizy2 (t ) (待发表)、Pizy3 (t ) (樊琳琳等, 2015 )、Pizy4 (t ) (待发表)及Pizy5 (t ) (待发表), 因此将该基因定名为Pizy6 (t ). ...Pizy3 (t ) (樊琳琳等, 2015 ), Pizy6 (t )基因的定位区间与Pizy3 (t )基因部分重叠.我们前期的研究结果显示, LP11和LP33菌株对36个抗病单基因系的致病性存在明显差异(表3 ), 推测这2个菌株很可能携带不同的无毒基因.参照Yang等(2013)、Huang等(2014)和Guo等(2016)的方法, 我们利用6个已克隆的无毒基因特异引物对2个菌株进行分析, 证实2个菌株携带的无毒基因确实存在差异(图7 ).同时, 我们利用子预44、湘资3150、9311、中花11、日本晴、TP309、丽江新团黑谷和江南香糯8个水稻材料, 通过苗期喷雾接种对LP11和LP33的致病性进行比较研究, 结果表明, LP11和LP33对抗病水稻材料9311的致病性明显不同(图8 ).我们将利用2个致病性不同的稻瘟病菌株LP11和LP33对含目标基因的群体材料进行深入研究以明确Pizy3 (t )和Pizy6 (t )的关系. ...Pib 被克隆, 到2006年7年间只克隆了1个抗稻瘟病基因Pita .从2006年开始, 抗稻瘟病基因的克隆数量大规模增长, 然而从已经克隆的抗稻瘟病基因位点我们发现, 从2009年开始, 尽管抗稻瘟病基因的数目在增加, 但真正的抗稻瘟病基因位点数却增加很少, 后来克隆的抗稻瘟病基因大多数都是已经克隆的等位或是直系同源基因(Xu et al., 2014 ).由于稻瘟病菌群体具复杂性和易变性, 我们每年从感病田块采集新的稻瘟病标样, 分离新的强致病性菌株, 并定位和克隆到具有最佳时效性的抗病新基因.黑龙江哈尔滨、江苏盐城、云南宜良、罗平和腾冲是近年来稻瘟病发生较严重的稻作区, 我们采用稻瘟病抗性不同的水稻材料, 对新分离自这些稻瘟病流行稻作区病样的稻瘟病菌单孢菌株进行致病性鉴定和新出现的强致病菌株筛选, 并利用筛选出的强致病菌株LP33和LP11, 利用图位克隆法在广谱抗瘟水稻子预44中定位了Pizy3 (t ) (樊琳琳等, 2015 )和Pizy6 (t )两个不同的稻瘟病抗性基因.上述研究结果表明, 从栽培品种多样、利于稻瘟病菌繁殖活动、田间稻瘟病菌组成复杂且信息流强度大的稻作区分离和筛选新的致病性菌株用于基因定位克隆, 是鉴定抗瘟新基因的有效方法. ...Pib 、Pi9 和Pid3 (李书等, 2014 ).利用不同稻瘟病菌株, 通过图位克隆法鉴定了位于第11号染色体的Pizy (t )/Pizy1 (t ) (张锦文等, 2009 )、位于第6号染色体的Pizy2 (t ) (待发表)、Pizy3 (t ) (樊琳琳等, 2015 )、Pizy4 (t )和Pizy5 (t ) (待发表)以及本研究中定位的Pizy6 (t ).上述研究结果表明, 广谱持久抗瘟水稻子预44携带多个主效和微效抗瘟基因, 其中主效抗稻瘟病基因具有集中成簇分布在第6号染色体上的特征, 他们对维持子预44的广谱持久抗性发挥着重要作用.此外, 根据基因预测的结 果, 水稻第6号染色体上还有许多未被鉴定的抗病 基因(http://rice.plantbiology.msu.edu/).相关基因 的进一步鉴定和分子标记的发展, 对利用子预44进行广谱持久抗瘟水稻品种的分子模块育种具有重要意义. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...张锦文等, 2009 , 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ; 周镕等, 2015 ).为了鉴定和克隆子预44中的优异抗瘟基因, 揭示子预44的广谱抗瘟分子基础, 我们通过在稻瘟病发病严重的水稻种植区采集穗颈瘟病标样, 采用适当改进的单孢分离方法进行稻瘟病菌株的单孢分离, 在利用广谱持久抗病水稻子预44和谱感水稻江南香糯对分离菌株进行初步的致病性鉴定和筛选的基础上, 用分离自云南罗平的菌株LP11对子预44和江南香糯杂交构建的F2 群体进行了抗性遗传分析和相关抗瘟基因的初步定位.本研究为抗性遗传研究提供了有价值的新菌株, 为子预44中抗LP11菌株基因的克隆奠定了基础. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...Pib 、Pi9 和Pid3 (李书等, 2014 ).利用不同稻瘟病菌株, 通过图位克隆法鉴定了位于第11号染色体的Pizy (t )/Pizy1 (t ) (张锦文等, 2009 )、位于第6号染色体的Pizy2 (t ) (待发表)、Pizy3 (t ) (樊琳琳等, 2015 )、Pizy4 (t )和Pizy5 (t ) (待发表)以及本研究中定位的Pizy6 (t ).上述研究结果表明, 广谱持久抗瘟水稻子预44携带多个主效和微效抗瘟基因, 其中主效抗稻瘟病基因具有集中成簇分布在第6号染色体上的特征, 他们对维持子预44的广谱持久抗性发挥着重要作用.此外, 根据基因预测的结 果, 水稻第6号染色体上还有许多未被鉴定的抗病 基因(http://rice.plantbiology.msu.edu/).相关基因 的进一步鉴定和分子标记的发展, 对利用子预44进行广谱持久抗瘟水稻品种的分子模块育种具有重要意义. ...Oryza sativa )作为中国第一大粮食作物, 在确保中国粮食生产安全中占有重要地位(邹征欧等, 2016 ).人口持续增长、全球气候变暖等因素致使各种病虫害发生增加, 严重威胁水稻生产及粮食安全(Cheng et al., 2007 ).稻瘟病是由稻瘟病菌(Magnaporthe oryzae )引起的水稻真菌病害, 发生在水稻生长的各个时期, 流行年份造成减产10%-20%, 严重时产量损失达到40%-50%, 甚至颗粒无收(温小红等, 2013 ; 赵家铭等, 2014 ), 严重威胁水稻生产的持续发展. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...Waller and Qu, 1985 ).因此, 发掘新的具有广谱持久抗性的抗病基因对稻瘟病的防治具有重要意义.然而, 无论是鉴定新抗性基因, 还是通过抗谱测定评估一个抗性基因的能效, 或是推测其持久性和抗性变异程度, 都需要大量的稻瘟病菌株作为基础.稻瘟病菌株单胞分离不仅是稻瘟病研究中一项重要的技术, 而且也是获得新稻瘟病菌致病菌株的重要基础工作(袁桂荣和甘贤友, 1987 ; 张书建和何月秋, 2003 ). ...张锦文等, 2009 , 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ; 周镕等, 2015 ).为了鉴定和克隆子预44中的优异抗瘟基因, 揭示子预44的广谱抗瘟分子基础, 我们通过在稻瘟病发病严重的水稻种植区采集穗颈瘟病标样, 采用适当改进的单孢分离方法进行稻瘟病菌株的单孢分离, 在利用广谱持久抗病水稻子预44和谱感水稻江南香糯对分离菌株进行初步的致病性鉴定和筛选的基础上, 用分离自云南罗平的菌株LP11对子预44和江南香糯杂交构建的F2 群体进行了抗性遗传分析和相关抗瘟基因的初步定位.本研究为抗性遗传研究提供了有价值的新菌株, 为子预44中抗LP11菌株基因的克隆奠定了基础. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...张锦文等, 2009 , 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ; 周镕等, 2015 ).为了鉴定和克隆子预44中的优异抗瘟基因, 揭示子预44的广谱抗瘟分子基础, 我们通过在稻瘟病发病严重的水稻种植区采集穗颈瘟病标样, 采用适当改进的单孢分离方法进行稻瘟病菌株的单孢分离, 在利用广谱持久抗病水稻子预44和谱感水稻江南香糯对分离菌株进行初步的致病性鉴定和筛选的基础上, 用分离自云南罗平的菌株LP11对子预44和江南香糯杂交构建的F2 群体进行了抗性遗传分析和相关抗瘟基因的初步定位.本研究为抗性遗传研究提供了有价值的新菌株, 为子预44中抗LP11菌株基因的克隆奠定了基础. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...2 群体, 对子预44抗LP11菌株的抗性遗传进行了分析.在水稻幼苗3-4叶期利用稻瘟病菌株LP11孢子悬浮液喷雾接种日本晴、9311、亲本子预44、江南香糯及其杂交F2 代群体.结果表明, 亲本子预44 (25株)和9311 (10株)全部表现高抗, 亲本江南香糯(25株)和日本晴(14株)全部表现高感, 与之前的致病性鉴定结果一致(图3 ).F2 代群体共489株, 其中抗病单株370株, 感病单株119株; X2 c<X2 0.05(1)=3.84, 符合3:1的抗感分离比(表2 ).结果表明, 子预44对稻瘟病菌株LP11的抗性属于单基因控制的显性遗传.前期本研究组通过图位克隆方法已在子预44中鉴定了Pizy (t )/Pizy1 (t ) (张锦文等, 2009 )、Pizy2 (t ) (待发表)、Pizy3 (t ) (樊琳琳等, 2015 )、Pizy4 (t ) (待发表)及Pizy5 (t ) (待发表), 因此将该基因定名为Pizy6 (t ). ...Pib 、Pi9 和Pid3 (李书等, 2014 ).利用不同稻瘟病菌株, 通过图位克隆法鉴定了位于第11号染色体的Pizy (t )/Pizy1 (t ) (张锦文等, 2009 )、位于第6号染色体的Pizy2 (t ) (待发表)、Pizy3 (t ) (樊琳琳等, 2015 )、Pizy4 (t )和Pizy5 (t ) (待发表)以及本研究中定位的Pizy6 (t ).上述研究结果表明, 广谱持久抗瘟水稻子预44携带多个主效和微效抗瘟基因, 其中主效抗稻瘟病基因具有集中成簇分布在第6号染色体上的特征, 他们对维持子预44的广谱持久抗性发挥着重要作用.此外, 根据基因预测的结 果, 水稻第6号染色体上还有许多未被鉴定的抗病 基因(http://rice.plantbiology.msu.edu/).相关基因 的进一步鉴定和分子标记的发展, 对利用子预44进行广谱持久抗瘟水稻品种的分子模块育种具有重要意义. ...Waller and Qu, 1985 ).因此, 发掘新的具有广谱持久抗性的抗病基因对稻瘟病的防治具有重要意义.然而, 无论是鉴定新抗性基因, 还是通过抗谱测定评估一个抗性基因的能效, 或是推测其持久性和抗性变异程度, 都需要大量的稻瘟病菌株作为基础.稻瘟病菌株单胞分离不仅是稻瘟病研究中一项重要的技术, 而且也是获得新稻瘟病菌致病菌株的重要基础工作(袁桂荣和甘贤友, 1987 ; 张书建和何月秋, 2003 ). ...本研究参照张书建和何月秋(2003) 涂平板单孢分离法, 并稍作改进.采用番茄燕麦培养基进行产孢培养.挑取少量的单孢菌丝, 先用PDA培养基对其进行传代培养和纯培养, 得到纯培养物后, 挑取少量菌丝于马铃薯葡萄糖液体培养基中, 26°C、每分钟180转摇菌5-7天.取出菌液, 涂在番茄燕麦培养基上, 每皿吸取500 µL, 在光照培养箱中培养, 26°C昼夜交替光照, 2-3天即可产孢.在超净工作台上, 向培养皿表面注入少量无菌水, 用消毒棉棒刮去菌丝, 经3层擦镜纸过滤, 即可得到大量孢子. ...菌株分离参照张书建和何月秋(2003) 的涂平板单孢分离法, 并作适当改进.把滤纸折成扇形, 以便更好地控制病组织表面的湿度和减少杂菌的生长.每个培养皿(直径9 cm)的水琼脂精确控制在5-7 mL, 使水琼脂平板的厚度控制在2-3 mm, 便于涂板, 同时透明度好, 能很清楚地在体视显微镜下看到孢子, 便于进行单孢挑取.采用改进后的孢子涂板单孢分离法, 利用采集自黑龙江哈尔滨、江苏盐城和云南宜良的病组织进行单孢分离, 成功地从上述不同稻区的病组织中分离约500份单孢, 经过进一步活化培养, 保存63份有效单孢菌株. ...Oryza sativa )作为中国第一大粮食作物, 在确保中国粮食生产安全中占有重要地位(邹征欧等, 2016 ).人口持续增长、全球气候变暖等因素致使各种病虫害发生增加, 严重威胁水稻生产及粮食安全(Cheng et al., 2007 ).稻瘟病是由稻瘟病菌(Magnaporthe oryzae )引起的水稻真菌病害, 发生在水稻生长的各个时期, 流行年份造成减产10%-20%, 严重时产量损失达到40%-50%, 甚至颗粒无收(温小红等, 2013 ; 赵家铭等, 2014 ), 严重威胁水稻生产的持续发展. ...张锦文等, 2009 , 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ; 周镕等, 2015 ).为了鉴定和克隆子预44中的优异抗瘟基因, 揭示子预44的广谱抗瘟分子基础, 我们通过在稻瘟病发病严重的水稻种植区采集穗颈瘟病标样, 采用适当改进的单孢分离方法进行稻瘟病菌株的单孢分离, 在利用广谱持久抗病水稻子预44和谱感水稻江南香糯对分离菌株进行初步的致病性鉴定和筛选的基础上, 用分离自云南罗平的菌株LP11对子预44和江南香糯杂交构建的F2 群体进行了抗性遗传分析和相关抗瘟基因的初步定位.本研究为抗性遗传研究提供了有价值的新菌株, 为子预44中抗LP11菌株基因的克隆奠定了基础. ...胡标林等, 2006 ; 张锦文等, 2009 , 2011 ; 黄红梅等, 2011 ; 李书等, 2014 ; 樊琳琳等, 2015 ).丽江新团黑谷、江南香糯、日本晴和TP309是抗病基因研究中常用的感病对照材料(伍尚忠等, 2004 ).中花11是一个含有广谱抗性材料特普血缘的常规粳稻品种(朱旭东等, 2003 ).利用上述8个抗感性状有差异的水稻材料, 通过苗期喷雾接种, 对H53、YC119、LP11、LP33和LP174的致病性进行了进一步鉴定, 发现接种稻瘟病菌株LP11后, 除子预44、9311和中花11表现抗性外, 其余5个材料, 包括具有广谱抗瘟性的籼稻湘资3150均表现高感(图2 ).结果表明, LP11很可能是新进化出来的强致病菌株, 为优良抗瘟新基因鉴定带来了契机. ...Oryza sativa )作为中国第一大粮食作物, 在确保中国粮食生产安全中占有重要地位(邹征欧等, 2016 ).人口持续增长、全球气候变暖等因素致使各种病虫害发生增加, 严重威胁水稻生产及粮食安全(Cheng et al., 2007 ).稻瘟病是由稻瘟病菌(Magnaporthe oryzae )引起的水稻真菌病害, 发生在水稻生长的各个时期, 流行年份造成减产10%-20%, 严重时产量损失达到40%-50%, 甚至颗粒无收(温小红等, 2013 ; 赵家铭等, 2014 ), 严重威胁水稻生产的持续发展. ...Pizy6 (t )基因定位在水稻第6号染色体短臂RM276 (物理位置: 6.23 Mb)与Indel06081 (物理位置: 8.73 Mb)之间大约2.5 Mb范围内.该基因与通过分子标记鉴定的位于水稻第6号染色体短臂上其它6个位点上的10个抗瘟主效基因在染色体上的位置不同, 它们是Pi22 (4.89-6.02 Mb)、Pi26 (8.75-11.67 Mb)、Pi27 (t ) (0.74-5.55 Mb)、Pi40 (16.27-17.53 Mb)、Pid3 /Pi25 (13.05 Mb)和Piz (10.15-10.51 Mb)位点上的复等位基因Pi2 /Pizt / Pi9 /Pigm (10.36-10.42 Mb) (Zhou et al., 2006 ; Chen et al., 2011 ; Sharma et al., 2012 ; Tanweer et al., 2015 ).由此可见, Pizy6 (t )可能是一个新的抗稻瘟病基因. ...Oryza sativa )作为中国第一大粮食作物, 在确保中国粮食生产安全中占有重要地位(邹征欧等, 2016 ).人口持续增长、全球气候变暖等因素致使各种病虫害发生增加, 严重威胁水稻生产及粮食安全(Cheng et al., 2007 ).稻瘟病是由稻瘟病菌(Magnaporthe oryzae )引起的水稻真菌病害, 发生在水稻生长的各个时期, 流行年份造成减产10%-20%, 严重时产量损失达到40%-50%, 甚至颗粒无收(温小红等, 2013 ; 赵家铭等, 2014 ), 严重威胁水稻生产的持续发展. ...无毒基因及PCR扩增引物参考Huang等(2014) 的方法(表1 ).PCR扩增体系: 10 μL 2xGC Buffer I (Takara), 2 μL dNTP (2.5 mmol·L-1 , Takara), 0.2 μL Easy Taq 酶(TransGen), 1.0 μL引物(10 μmol·µL-1 ), 2 μL DNA模板(10 ng), 加ddH2 O补足至20 μL.PCR反应程序: 94°C预变性5分钟; 94°C30秒, 55°C (依据引物的Tm值稍作调整)30秒, 72°C1分钟, 30个循环; 72°C延伸10分钟.PCR产物用1%琼脂糖凝胶电泳检测. ...International Rice Genome Sequencing Project, 2005 )提供的序列由华大基因公司或硕擎生物科技有限公司合成SSR引物. ...Pizy6 (t )基因定位在水稻第6号染色体短臂RM276 (物理位置: 6.23 Mb)与Indel06081 (物理位置: 8.73 Mb)之间大约2.5 Mb范围内.该基因与通过分子标记鉴定的位于水稻第6号染色体短臂上其它6个位点上的10个抗瘟主效基因在染色体上的位置不同, 它们是Pi22 (4.89-6.02 Mb)、Pi26 (8.75-11.67 Mb)、Pi27 (t ) (0.74-5.55 Mb)、Pi40 (16.27-17.53 Mb)、Pid3 /Pi25 (13.05 Mb)和Piz (10.15-10.51 Mb)位点上的复等位基因Pi2 /Pizt / Pi9 /Pigm (10.36-10.42 Mb) (Zhou et al., 2006 ; Chen et al., 2011 ; Sharma et al., 2012 ; Tanweer et al., 2015 ).由此可见, Pizy6 (t )可能是一个新的抗稻瘟病基因. ...Pizy6 (t )基因定位在水稻第6号染色体短臂RM276 (物理位置: 6.23 Mb)与Indel06081 (物理位置: 8.73 Mb)之间大约2.5 Mb范围内.该基因与通过分子标记鉴定的位于水稻第6号染色体短臂上其它6个位点上的10个抗瘟主效基因在染色体上的位置不同, 它们是Pi22 (4.89-6.02 Mb)、Pi26 (8.75-11.67 Mb)、Pi27 (t ) (0.74-5.55 Mb)、Pi40 (16.27-17.53 Mb)、Pid3 /Pi25 (13.05 Mb)和Piz (10.15-10.51 Mb)位点上的复等位基因Pi2 /Pizt / Pi9 /Pigm (10.36-10.42 Mb) (Zhou et al., 2006 ; Chen et al., 2011 ; Sharma et al., 2012 ; Tanweer et al., 2015 ).由此可见, Pizy6 (t )可能是一个新的抗稻瘟病基因. ...Waller and Qu, 1985 ).因此, 发掘新的具有广谱持久抗性的抗病基因对稻瘟病的防治具有重要意义.然而, 无论是鉴定新抗性基因, 还是通过抗谱测定评估一个抗性基因的能效, 或是推测其持久性和抗性变异程度, 都需要大量的稻瘟病菌株作为基础.稻瘟病菌株单胞分离不仅是稻瘟病研究中一项重要的技术, 而且也是获得新稻瘟病菌致病菌株的重要基础工作(袁桂荣和甘贤友, 1987 ; 张书建和何月秋, 2003 ). ...Pib 被克隆, 到2006年7年间只克隆了1个抗稻瘟病基因Pita .从2006年开始, 抗稻瘟病基因的克隆数量大规模增长, 然而从已经克隆的抗稻瘟病基因位点我们发现, 从2009年开始, 尽管抗稻瘟病基因的数目在增加, 但真正的抗稻瘟病基因位点数却增加很少, 后来克隆的抗稻瘟病基因大多数都是已经克隆的等位或是直系同源基因(Xu et al., 2014 ).由于稻瘟病菌群体具复杂性和易变性, 我们每年从感病田块采集新的稻瘟病标样, 分离新的强致病性菌株, 并定位和克隆到具有最佳时效性的抗病新基因.黑龙江哈尔滨、江苏盐城、云南宜良、罗平和腾冲是近年来稻瘟病发生较严重的稻作区, 我们采用稻瘟病抗性不同的水稻材料, 对新分离自这些稻瘟病流行稻作区病样的稻瘟病菌单孢菌株进行致病性鉴定和新出现的强致病菌株筛选, 并利用筛选出的强致病菌株LP33和LP11, 利用图位克隆法在广谱抗瘟水稻子预44中定位了Pizy3 (t ) (樊琳琳等, 2015 )和Pizy6 (t )两个不同的稻瘟病抗性基因.上述研究结果表明, 从栽培品种多样、利于稻瘟病菌繁殖活动、田间稻瘟病菌组成复杂且信息流强度大的稻作区分离和筛选新的致病性菌株用于基因定位克隆, 是鉴定抗瘟新基因的有效方法. ...Pizy6 (t )基因定位在水稻第6号染色体短臂RM276 (物理位置: 6.23 Mb)与Indel06081 (物理位置: 8.73 Mb)之间大约2.5 Mb范围内.该基因与通过分子标记鉴定的位于水稻第6号染色体短臂上其它6个位点上的10个抗瘟主效基因在染色体上的位置不同, 它们是Pi22 (4.89-6.02 Mb)、Pi26 (8.75-11.67 Mb)、Pi27 (t ) (0.74-5.55 Mb)、Pi40 (16.27-17.53 Mb)、Pid3 /Pi25 (13.05 Mb)和Piz (10.15-10.51 Mb)位点上的复等位基因Pi2 /Pizt / Pi9 /Pigm (10.36-10.42 Mb) (Zhou et al., 2006 ; Chen et al., 2011 ; Sharma et al., 2012 ; Tanweer et al., 2015 ).由此可见, Pizy6 (t )可能是一个新的抗稻瘟病基因. ... 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}