Abstract Aims Epichlo? infection can play an important role in the growth and development of host plants. However, few studies have been reported on the effects of endophyte infection on leaf shape and area of host plants. Methods Our study used Achnatherum sibiricum as experimental material, and explored the effects of endophyte species and host plant genotype on the leaf shape and area of A. sibiricum-endophyte symbionts through the combination of the length and width method with the revised correction coefficient and the scanning calculation method. Important findings Endophyte infection status, endophyte species and host plant genotype had no significant influences on the correction coefficient of leaf size calculation, leaf length, leaf width and leaf length-width ratio reflecting leaf shape. By calculation and verification, we determined that the correction coefficient of leaf size calculation of A. sibiricum was 0.594 9. There was no significant difference between the leaf areas measured by length and width method with the revised correction coefficient and the scanning calculation method. In addition, the leaf areas measured by two methods were not significantly affected by endophyte infection, endophyte species and plant maternal genotype. Keywords:fungal endophyte;Achnatherum sibiricum;leaf area;maternal genotype;correction coefficient
PDF (933KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 秦天姿, 任安芝, 樊晓雯, 高玉葆. 内生真菌种类和母本基因型对内生真菌-禾草共生体叶形状和叶面积的影响. 植物生态学报, 2020, 44(6): 654-660. DOI: 10.17521/cjpe.2020.0037 QIN Tian-Zi, REN An-Zhi, FAN Xiao-Wen, GAO Yu-Bao. Effects of endophyte fungal species and host plant genotype on the leaf shape and leaf area of endophyte-grass symbionts. Chinese Journal of Plant Ecology, 2020, 44(6): 654-660. DOI: 10.17521/cjpe.2020.0037
Table 1 表1 表1不同内生真菌-羽茅共生体中, 内生真菌感染及宿主母本基因型对羽茅叶片长度、宽度和长宽比影响的方差分析 Table 1Analysis of variance for the effects of endophyte infection and host plant genotype on leaf length, width and length-width ratio of Achnatherum sibiricum-endophyte symbionts
叶长 Leaf length (cm)
叶宽 Leaf width (cm)
长宽比 Length-width ratio
F
p
F
p
F
p
E
3.503
0.068
0.563
0.457
1.970
0.167
PG (EG)
0.372
0.893
1.955
0.095
1.596
0.171
E × PG (EG)
0.349
0.906
0.801
0.532
1.728
0.143
E, 内生真菌感染; PG (EG), 植物母本基因型(嵌套于不同内生真菌)。 E, endophyte infection; PG (EG), plant maternal genotype (nested within endophyte species).
Table 2 表2 表2内生真菌种类对羽茅叶片长度、宽度和长宽比影响的单因素方差分析 Table 2One-way analysis of variance for the effects of endophyte species on leaf length, width and leaf length-width ratio of Achnatherum sibiricum
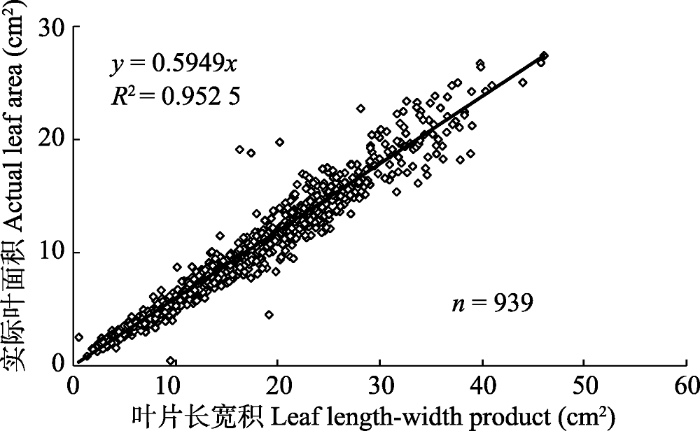
本研究利用广义线性混合模型(GLMM)探究不同处理对羽茅叶片校正系数的影响, 实际是对叶片长宽积与实际叶面积拟合方程斜率的影响。结果表明, 内生真菌感染、植物母本基因型以及二者与长宽积的交互作用对叶面积均无显著影响, 但叶片长宽积与羽茅叶面积存在显著线性关系(表3)。固定系数结果表明只有叶片长宽积与叶面积存在显著线性关系(固定系数为0.596, p < 0.001)。该结果表明叶片长宽积和叶面积之间的线性关系不受内生真菌感染与否、不同羽茅母本基因型的影响。
Table 3 表3 表3不同内生真菌-羽茅共生体中, 内生真菌感染及宿主母本基因型对羽茅叶片长宽积和叶面积影响的广义线性混合模型分析 Table 3Analysis of generalized linear mixed model for the effects of endophyte infection and host plant genotype on leaf length-width product and leaf area of different Achnatherum sibiricum-endophyte symbionts
羽茅叶面积 Leaf area of A. sibiricum
F
p
校正模型 Corrected Model
595.131
<0.001
E
1.112
0.292
PG (EG)
0.843
0.537
LWP
15 233.050
<0.001
E × PG (EG)
0.683
0.707
E × LWP
0.707
0.401
PG (EG) × LWP
1.567
0.131
E × PG (EG) × LWP
1.858
0.063
E, 内生真菌感染; PG (EG), 植物母本基因型(嵌套于不同内生真菌); LWP, 叶片长宽积。 E, endophyte infection; PG (EG), plant genotype (nested within endophyte species); LWP, leaf length-width product.
Es和Eg1是不同种类内生真菌, 但它们具有相同的传播方式, Eg1和Eg2是具有不同传播方式的同种内生真菌, GLMM结果表明羽茅叶片长宽积和叶面积的线性关系不受内生真菌种类的影响; 且传播方式不同也未影响羽茅叶片长宽积和叶面积的线性关系(表4)。固定系数结果表明只有叶片长宽积与叶面积存在显著线性关系(固定系数为0.591, p < 0.001)。
Table 4 表4 表4内生真菌种类对羽茅叶片长宽积和叶面积的广义线性混合模型分析 Table 4The generalized linear mixed model analysis for the effects of endophyte species on leaf length-width product and area of Achnatherum sibiricum
Table 5 表5 表5不同内生真菌-羽茅共生体中, 内生真菌感染及宿主母本基因型对羽茅平均叶面积影响的方差分析 Table 5Analysis of variance for the effects of endophyte infection and host plant genotype on average leaf area of different Achnatherum sibiricum-endophyte symbionts
实际叶面积 Actual leaf area (cm2)
长宽法测得叶面积 Leaf area measured by length-width method (cm2)
F
p
F
p
E
1.677
0.202
2.324
0.135
PG (EG)
1.101
0.377
1.256
0.297
E × PG (EG)
0.291
0.937
0.205
0.973
E, 内生真菌感染; PG (EG), 植物母本基因型(嵌套于不同内生真菌)。 E, endophyte infection; PG (EG), plant maternal genotype (nested within endophyte species).
Table 6 表6 表6内生真菌种类对羽茅平均叶面积影响的单因素方差分析 Table 6One-way analysis of variance for the effects of endophyte species on average leaf area of Achnatherum sibiricum
实际叶面积 Actual leaf area (cm2)
长宽法测得叶面积 Leaf area measured by length-width method (cm2)
BittlestonLS, BrockmannF, WcisloW, van BaelSA (2011). Endophytic fungi reduce leaf-cutting ant damage to seedlings Biology Letters, 7, 30-32. DOI:10.1098/rsbl.2010.0456URLPMID:20610420 [本文引用: 1] Our study examines how the mutualism between Atta colombica leaf-cutting ants and their cultivated fungus is influenced by the presence of diverse foliar endophytic fungi (endophytes) at high densities in tropical leaf tissues. We conducted laboratory choice trials in which ant colonies chose between Cordia alliodora seedlings with high (E(high)) or low (E(low)) densities of endophytes. The E(high) seedlings contained 5.5 times higher endophyte content and a greater diversity of fungal morphospecies than the E(low) treatment, and endophyte content was not correlated with leaf toughness or thickness. Leaf-cutting ants cut over 2.5 times the leaf area from E(low) relative to E(high) seedlings and had a tendency to recruit more ants to E(low) plants. Our findings suggest that leaf-cutting ants may incur costs from cutting and processing leaves with high endophyte loads, which could impact Neotropical forests by causing variable damage rates within plant communities.
CarrollG (1988). Fungal endophytes in stems and leaves: from latent pathogen to mutualistic symbiont Ecology, 69, 2-9. [本文引用: 1]
CheplickGP (1997). Effects of endophytic fungi on the phenotypic plasticity of Lolium perenne(Poaceae) American Journal of Botany, 84, 34-40. [本文引用: 2]
ClayK (1987). Effects of fungal endophytes on the seed and seedling biology of Lolium perenne and Festuca arundinacea Oecologia, 73, 358-362. [本文引用: 3]
DuJX, WangXF, ZhangGJ (2007). Leaf shape based plant species recognition Applied Mathematics and Computation, 185, 883-893. [本文引用: 1]
DuYJ, SunXB, HanLB (2010). Effect ofNeotyphodium starrii infected on photosynthetic and orphological characteristics of tall fescue under high temperature Journal of Central South University of Forestry & Technology, 30, 41-47. [本文引用: 1]
EstradaC, RojasEI, WcisloWT, van BaelSA (2014). Fungal endophyte effects on leaf chemistry alter the in vitro growth rates of leaf-cutting ants’ fungal mutualist, Leucocoprinus gongylophorus Fungal Ecology, 8, 37-45. [本文引用: 1]
IannoneLJ, PingetAD, NagabhyruP, SchardlCL, de BattistaJP (2012). Beneficial effects of Neotyphodium tembladerae and Neotyphodium pampeanum on a wild forage grass Grass and Forage Science, 67, 382-390. [本文引用: 2]
JiaT, RenAZ, WangS, GaoYB (2011). Effect of Endophytic fungi on growth and photosynthetic characteristics of Achnatherum sibiricum Acta Ecologica Sinica, 31, 4811-4817. [本文引用: 1]
KannadanS, RudgersJA (2008). Endophyte symbiosis benefits a rare grass under low water availability Functional Ecology, 22, 706-713. [本文引用: 4]
KnappAK (2001). Variation among biomes in temporal dynamics of aboveground primary production Science, 291, 481-484. DOI:10.1126/science.291.5503.481URLPMID:11161201 [本文引用: 1] Interannual variability in aboveground net primary production (ANPP) was assessed with long-term (mean = 12 years) data from 11 Long Term Ecological Research sites across North America. The greatest interannual variability in ANPP occurred in grasslands and old fields, with forests the least variable. At a continental scale, ANPP was strongly correlated with annual precipitation. However, interannual variability in ANPP was not related to variability in precipitation. Instead, maximum variability in ANPP occurred in biomes where high potential growth rates of herbaceous vegetation were combined with moderate variability in precipitation. In the most dynamic biomes, ANPP responded more strongly to wet than to dry years. Recognition of the fourfold range in ANPP dynamics across biomes and of the factors that constrain this variability is critical for detecting the biotic impacts of global change phenomena.
LatchGCM, ChristensenMJ, SamuelsGJ (1984). Five endophytes of Lolium and Festuca in New Zealand Mycotaxon, 20, 535-550. [本文引用: 1]
LiHR, LiHL, WangHG, LiDX, LiRQ, LiYM (2018). Further study on the method of leaf area calculation in winter wheat Journal of Triticeae Crops, 38, 455-459. [本文引用: 1]
LiX, ZhouY, ZhuMJ, QinJH, RenAZ, GaoYB (2015). Stroma-bearing endophyte and its potential horizontal transmission ability in Achnatherum sibiricum Mycologia, 107, 21-31. [本文引用: 1]
LiYM (1993a). A preliminary study on the correction value (k) of leaf area of Sorghum bicolor(L.) Moench Journal of Agricultural University of Hebei, 16, 32-36. [本文引用: 1]
LiYM (1993b). Preliminary study on the method of leaf area measurement in oats (Avena spp.) Journal of Agricultural University of Hebei, 16, 25-28. [本文引用: 1]
LiuH, ChenW, ZhouY, LiX, RenAZ, GaoYB (2015). Effects of endophyte and arbuscular mycorrhizal fungi on growth of Leymus chinensis Chinese Journal of Plant Ecology, 39, 477-485. [本文引用: 2]
LuoJ, ZhangH, DengZH, XuLN, GaoSJ, ChenRK, ChenYQ (2005). Relationship between canopy characters and leaf morphology at different positions of sugarcane Chinese Journal of Applied and Environmental Biology, 11, 28-31. [本文引用: 1]
NovasMV, GentileA, CabralD (2003). Comparative study of growth parameters on diaspores and seedlings between populations of Bromus setifolius from Patagonia, differing in Neotyphodium endophyte infection Flora, 198, 421-426. [本文引用: 1]
OberhoferM, GüsewellS, LeuchtmannA (2014). Effects of natural hybrid and non-hybrid Epichlo? endophytes on the response of Hordelymus europaeus to drought stress New Phytologist, 201, 242-253. [本文引用: 2]
RenAZ, ClayK (2009). Impact of a horizontally transmitted endophyte, Balansia henningsiana, on growth and drought tolerance of Panicum rigidulum International Journal of Plant Sciences, 170, 599-608. [本文引用: 2]
RenAZ, GaoYB (2003). Growth characteristics of endophyte- infected and endophyte-freeLolium perenne L. seedlings under osmotic stress conditions Acta Ecologica Sinica, 23, 2307-2317. [本文引用: 1]
RenAZ, LiX, HanR, YinLJ, WeiMY, GaoYB (2011). Benefits of a symbiotic association with endophytic fungi are subject to water and nutrient availability in Achnatherum sibiricum Plant and Soil, 346, 363-373. [本文引用: 2]
RymbaiH, LaxmanRH, DineshMR, SunojVSJ, RavishankarKV, JhaAK (2014). Diversity in leaf morphology and physiological characteristics among mango (Mangifera indica) cultivars popular in different agro-climatic regions of India Scientia Horticulturae, 176, 189-193. [本文引用: 1]
SimpsonWR, SchmidJ, SinghJ, FavilleMJ, JohnsonRD (2012). A morphological change in the fungal symbiont Neotyphodium lolii induces dwarfing in its host plant Lolium perenne Fungal Biology, 116, 234-240. [本文引用: 2]
SongML, ChaiQ, LiXZ, YaoX, LiCJ, ChristensenMJ, NanZB (2015). An asexual Epichlo? endophyte modifies the nutrient stoichiometry of wild barley(Hordeum brevisubulatum) under salt stress Plant and Soil, 387, 153-165. [本文引用: 2]
WangJF, NanZB, ChristensenMJ, ZhangXX, TianP, ZhangZX, NiuXL, GaoP, ChenT, MaLX (2018). Effect of Epichlo? gansuensis endophyte on the nitrogen metabolism, nitrogen use efficiency, and stoichiometry of Achnatherum inebrians under nitrogen limitation Journal of Agricultural and Food Chemistry, 66, 4022-4031. [本文引用: 1]
WangX, DaiTB, JiangD, JingQ, CaoWX (2005). Yield- formation and source-sink characteristics of rice genotypes under two different eco-environments Chinese Journal of Applied Ecology, 16, 615-619. URLPMID:16011154 [本文引用: 1] With nine Japanese varieties and two local cultivars of rice as test materials, this paper studied the characteristics of their yield-formation and source-sink characteristics under different eco-environments of Nanjing and Lijiang. The results showed that eco-environment had an obvious effect on rice grain yield and dry matter accumulation. High-yielding rice genotypes could accumulate more dry matter, especially in their late growth period. They had higher total spikelet number, leaf area index (LAI) and crop growth rate (CGR), as compared with other genotypes. The grain yield increased with increasing total dry matter accumulation, and significantly positively correlated with dry matter accumulation after heading, total spikelet number, LAI, and spikelet/leaf area ratio. The results suggested that the lower grain yield in Nanjing than in Lijiang was mainly resulted from the lower LAI, total spikelet, dry matter accumulation and CGR after heading. [ 王勋, 戴廷波, 姜东, 荆奇, 曹卫星 (2005). 不同生态环境下水稻基因型产量形成与源库特性的比较研究 应用生态学报, 16, 615-619.] PMID:16011154 [本文引用: 1]
WeiYK, GaoYB, XuH, SuD, ZhangX, WangYH, LinF, ChenL, NieLY, RenAZ (2006). Occurrence of endophytes in grasses native to Northern China Grass and Forage Science, 61, 422-429. [本文引用: 1]
YuJY, HeY, ZhaoZF, WangD (2007). Study on the correction coefficient of measuring crop leaf area by length- width method Jiangsu Agricultural Sciences, 35, 37-39. [本文引用: 1]
ZhangX, RenAZ, WeiYK, LinF, LiC, LiuZJ, GaoYB (2009). Taxonomy, diversity and origins of symbiotic endophytes of Achnatherum sibiricum in the Inner Mongolia Steppe of China FEMS Microbiology Letters, 301, 12-20. [本文引用: 1]
ZhouYG, LiHL, JiangTC, DouZH, LiuJ, WuSF, FengH, ZhangTB, HeJQ (2016). Temporal and spatial variations of leaf shape coefficients of summer maize Scientia Agricultura Sinica, 49, 4520-4530. [本文引用: 1]
Endophytic fungi reduce leaf-cutting ant damage to seedlings 1 2011
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
Fungal endophytes in stems and leaves: from latent pathogen to mutualistic symbiont 1 1988
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
Effects of endophytic fungi on the phenotypic plasticity of Lolium perenne(Poaceae) 2 1997
Fungal endophyte effects on leaf chemistry alter the in vitro growth rates of leaf-cutting ants’ fungal mutualist, Leucocoprinus gongylophorus 1 2014
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
Beneficial effects of Neotyphodium tembladerae and Neotyphodium pampeanum on a wild forage grass 2 2012
... 已有研究表明, 内生真菌对宿主植物生长的影响不仅与内生真菌的存在与否有关, 也与内生真菌的种类有关(Iannone et al., 2012; Simpson et al., 2012).Simpson等(2012)在黑麦草(Lolium perenne)中分离到两种不同形态的内生真菌, 其中, 菌落呈棉絮状的菌株对植物生长无显著影响; 但菌落呈黏液状的菌株使得宿主植株矮化.雀麦属(Bromus)的一种野生牧草Bromus auleticus可与两种内生真菌Epichlo? tembladerae和Epichlo? pampeanum共生, E. tembladerae感染显著降低宿主植物种子产量, 而E. pampeanum对宿主种子产量无显著影响(Iannone et al., 2012).那么内生真菌种类是否会影响宿主植物叶面积, 目前尚无相关报道. ...
... 对宿主种子产量无显著影响(Iannone et al., 2012).那么内生真菌种类是否会影响宿主植物叶面积, 目前尚无相关报道. ...
Endophyte symbiosis benefits a rare grass under low water availability 4 2008
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
Comparative study of growth parameters on diaspores and seedlings between populations of Bromus setifolius from Patagonia, differing in Neotyphodium endophyte infection 1 2003
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
Effects of natural hybrid and non-hybrid Epichlo? endophytes on the response of Hordelymus europaeus to drought stress 2 2014
... ), 而本实验进一步发现内生真菌感染与否、内生真菌种类和植物母本基因型均对羽茅的叶长、叶宽、叶片长宽比、叶校正系数和叶面积无显著影响, 因此, 内生真菌对羽茅生长的促进作用与促进分蘖生长(Ren et al., 2011)有关, 而与宿主叶面积和叶形状之间没有明显的相关性. ...
Diversity in leaf morphology and physiological characteristics among mango (Mangifera indica) cultivars popular in different agro-climatic regions of India 1 2014
A morphological change in the fungal symbiont Neotyphodium lolii induces dwarfing in its host plant Lolium perenne 2 2012
... 已有研究表明, 内生真菌对宿主植物生长的影响不仅与内生真菌的存在与否有关, 也与内生真菌的种类有关(Iannone et al., 2012; Simpson et al., 2012).Simpson等(2012)在黑麦草(Lolium perenne)中分离到两种不同形态的内生真菌, 其中, 菌落呈棉絮状的菌株对植物生长无显著影响; 但菌落呈黏液状的菌株使得宿主植株矮化.雀麦属(Bromus)的一种野生牧草Bromus auleticus可与两种内生真菌Epichlo? tembladerae和Epichlo? pampeanum共生, E. tembladerae感染显著降低宿主植物种子产量, 而E. pampeanum对宿主种子产量无显著影响(Iannone et al., 2012).那么内生真菌种类是否会影响宿主植物叶面积, 目前尚无相关报道. ...
... ).Simpson等(2012)在黑麦草(Lolium perenne)中分离到两种不同形态的内生真菌, 其中, 菌落呈棉絮状的菌株对植物生长无显著影响; 但菌落呈黏液状的菌株使得宿主植株矮化.雀麦属(Bromus)的一种野生牧草Bromus auleticus可与两种内生真菌Epichlo? tembladerae和Epichlo? pampeanum共生, E. tembladerae感染显著降低宿主植物种子产量, 而E. pampeanum对宿主种子产量无显著影响(Iannone et al., 2012).那么内生真菌种类是否会影响宿主植物叶面积, 目前尚无相关报道. ...
An asexual Epichlo? endophyte modifies the nutrient stoichiometry of wild barley(Hordeum brevisubulatum) under salt stress 2 2015
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
Effect of Epichlo? gansuensis endophyte on the nitrogen metabolism, nitrogen use efficiency, and stoichiometry of Achnatherum inebrians under nitrogen limitation 1 2018
... 内生真菌是指分布在健康植物组织中、在植物中度过大部分或全部生活史, 但又不会对宿主植物产生明显病症的一类真菌(Carroll, 1988).大量研究表明, 内生真菌感染不仅可以促进宿主植物的生长(Novas et al., 2003; Vila-Aiub et al., 2005; Kannadan & Rudgers, 2008; Song et al., 2015), 而且还可以提高宿主植物对生物干扰和非生物胁迫的抵抗能力(Bittleston et al., 2011; Estrada et al., 2014; Song et al., 2015; Wang et al., 2018). ...
 ,
, ,*, 樊晓雯, 高玉葆南开大学生命科学学院, 天津 300071
,*, 樊晓雯, 高玉葆南开大学生命科学学院, 天津 300071
 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT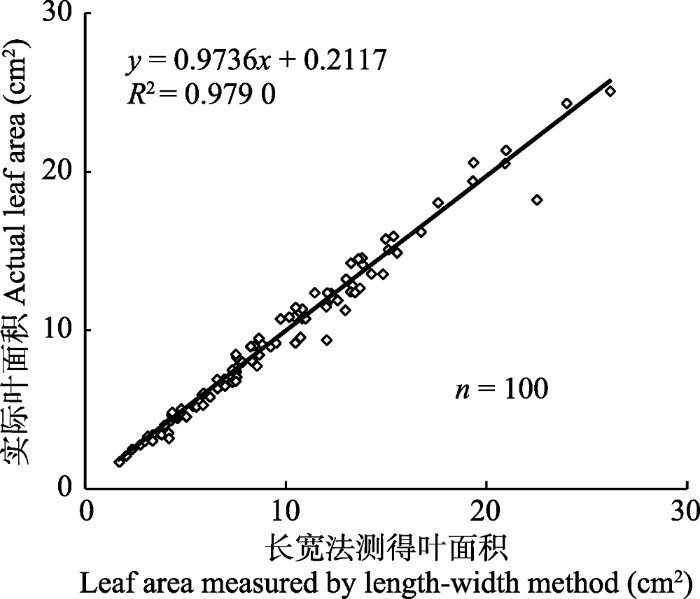 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT


{kind=link}
{kind=link}
{kind=link}
{kind=link}