Abstract: We used 21 rice (Oryza sativa) germplasm as experimental materials. Dry seeds were treated for 8 d in 1.5% NaCl solution and germination rate was measured. The seedlings with 2 green leaves were cultured by hydroponics in Petri dishes with different NaCl concentrations. After 10 days, relative shoot height, relative root length, leaf mortality, water content and the expression of 7 of 9 high-affinity K+ transporter (HKT) gene families were measured. In the adult period (30-d seedling to maturation), 3 representative germplasm were treated at different NaCl concentrations in a pot experiment. We investigated the effects of salt stress at flowering or dough stage of rice, for example, soluble sugar content. The results showed that germination rates of seed were affected significantly, with a wide range of variation, with 1.5% NaCl stress concentration. The variation in leaf mortality was the largest among the measured indicators at seedling stage. Besides OsHKT2;4, 6 members of HKT gene family were detected in the 8 germplasm genome. Soluble sugar content of germplasm showed a growth-stimulation effect with salt content stress ≤1 g∙kg-1. The biomass of CG15R was positively correlated with salt content and the biomass increased with the addition of salt. Biomass of Zhonghua9 increased with the addition of salt with salt content ≤1 g∙kg-1. Salt tolerance of rice shows the characteristic of stage development, and salt tolerance at different developmental stages is not correlated. The leaf mortality and biomass of dough stage could be the key indicators of salt tolerance for seedling and adult rice. CG15R represents a germplasm with high salt tolerance and should be further analyzed.
1 材料与方法1.1 材料供试材料为21份水稻(Oryza sativa L.)种质, 由西南科技大学植物分子遗传育种实验室提供(表1)。CG系列为新育成籼型恢复系, 其它为核心种质。 本研究在西南科技大学植物分子遗传育种实验室进行。所用智能光照培养箱为武汉瑞华仪器设备有限责任公司生产的HP2500G-C型。电热恒温培养箱为上海齐欣科学仪器有限公司生产的DHP-9082型。 表1 Table 1 表1 表1 供试材料及类型 Table 1 Materials and their type for this study
No.
Germplasm
Subspecies
No.
Germplasm
Subspecies
1
C190
Indica
12
CG131R
Indica
2
CG133R
Indica
13
CG158R
Indica
3
HD63
Indica
14
CG111R
Indica
4
CG15R
Indica
15
CG114R
Indica
5
Javanica 22
Japonica
16
CG173R
Indica
6
Shuhui527
Indica
17
CG132R
Indica
7
3301R
Indica
18
CG151R
Indica
8
Lehui188
Indica
19
CG159R
Indica
9
Zhonghua9
Japonica
20
CG240R
Indica
10
Jing925
Japonica
21
BR207-2
Indica
11
BR207-1
Indica
表1 供试材料及类型 Table 1 Materials and their type for this study
表2 Table 2 表2 表2HKT家族基因引物信息 Table 2 Primers information of HKT family genes
2 结果与讨论2.1 盐胁迫对不同种质发芽率的影响与对照相比, 经过1.5%NaCl溶液处理后, 21份水稻种质的发芽率受到显著影响, 各材料在未处理下的平均发芽率变化范围为60.02%-99.33% (表3)。而经盐处理后各材料平均发芽率变化范围是3.33%-81.67%。 以上结果表明种质间发芽率差异显著。其中10号种质在盐处理下的相对盐害率最低, 为9.26%; 该种质耐盐性极强, 达到1级。其次是16号, 耐盐性为强, 耐盐级别为3级。而相对盐害率大于60%的有16个种质, 说明这些种质在1.5%盐溶液处理下发芽率受到明显的影响, 盐胁迫对其伤害较大。 表3 Table 3 表3 表3 不同水稻种质的发芽率及相对盐害率 Table 3 Germination rate of different varieties and their relative salt damage rate
No.
Average germination rate (%)
Relative salt damage rate (%)
Salt tolerance
Level
CK
NaCl (1.5%)
1
99.33±1.15 aA
36.67±0.01 cC
63.08
Weak
7
2
99.33±1.15 aA
7.78±1.92 ghFG
92.17
Very weak
9
3
83.33±0.01 gG
13.33±0.01 efgEFG
84.00
Very weak
9
4
60.20±0.34 lL
8.84±3.08 ghFG
85.97
Very weak
9
5
73.33±0.01 jJ
11.67±1.67 fghEFG
84.08
Very weak
9
6
99.33±1.15 aA
28.00±15.62 dCD
71.68
Weak
7
7
83.33±0.01 gG
14.00±6 efgEFG
83.20
Very weak
9
8
86.67±0.01 fF
13.34±8.80 efgEFG
84.60
Very weak
9
9
93.34±0.01 dD
15.00±5 efgEF
83.93
Very weak
9
10
90.00±0.02 eE
81.67±1.67 aA
9.26
Very strong
1
11
73.33±0.01 jJ
16.89±3.00 efDEF
71.96
Weak
7
12
93.34±0.02 dD
3.33±0.01 hG
96.43
Very weak
9
13
93.34±0.02 dD
26.67±3.34 dCD
71.43
Weak
7
14
80.00±0.02 hH
10.00±2 fghFG
87.50
Very weak
9
15
86.68±0.02 fF
21.67±1.67 deDE
75.00
Weak
7
16
66.68±0.02 kK
50.00±0.02 bB
25.02
Strong
3
17
98.02±0.02 bB
13.33±7.02 efgEFG
86.39
Very weak
9
18
96.67±0.01 cC
7.33±3.05 ghFG
92.41
Very weak
9
19
76.67±0.01 iI
8.89±5.09 fghFG
88.41
Very weak
9
20
99.33±1.15 aA
8.89±5.09 fghFG
91.09
Very weak
9
21
93.34±0.01 dD
16.89±3.06 efDEF
81.90
Very weak
9
No. 1-21 see Table 1. Different lowercase letters indicate significant differences at the 0.05 level; and different capital letters indicate significant differences at the 0.01 level. 编号1-21同表1。不同小写字母表示在0.05水平差异显著; 不同大写字母表示在0.01水平差异显著。
表3 不同水稻种质的发芽率及相对盐害率 Table 3 Germination rate of different varieties and their relative salt damage rate
2.2 盐胁迫对不同种质苗期理化指标的影响2.2.1 盐胁迫对不同种质苗高和根长的影响 在盐胁迫下, 供试水稻幼苗的苗高和根长在低浓度盐处理下表现为刺激生长, 在高浓度下则受到抑制, 且不同种质受抑制的程度不尽相同。表4显示, 试供水稻种质的相对根长和相对苗高一般随盐浓度的增加而逐渐降低。其中在0.3%和0.5%的盐浓度下, 种质7、8、9和11号的相对苗高都大于1; 种质4、6、8、9、11、12和20号的相对根长也都大于或等于1, 说明0.3%和0.5%盐浓度促进这些种质的苗高和根长的生长。此外, 0.7%盐浓度也促进种质4、8、9和17号苗高的生长和4、9、10、11、12和17号根长的生长。从相对根长来看, 1、3和5号种质的相对根长降低较为明显, 平均值偏小, 而4、6、9和20号4个种质的相对根长降低较少, 平均值较高; 而对相对苗高来说, 种质7、9、11、14和17号降低很少, 总平均值较高。0.7%盐浓度还能促进部分种质苗高和根长的生长, 究其原因可能是遗传机制的差异。从图1可以明显看出, 11号在0.5%盐处理下生长较好, 与对照相比苗高较高; 随盐浓度的增加, 苗高逐渐降低且叶片的死亡面积也逐渐增加, 在0.9%盐处理下叶片几乎全部死亡。这说明同一种质对不同盐浓度有不同的抗性反应, 低浓度能刺激生长, 高浓度则抑制生长甚至致死。 表4 Table 4 表4 表4 不同浓度盐处理下部分水稻种质的相对苗高与相对根长 Table 4 Relative seedling height and relative root length of part rice germplasms under different salt concentration treatments
表4 不同浓度盐处理下部分水稻种质的相对苗高与相对根长 Table 4 Relative seedling height and relative root length of part rice germplasms under different salt concentration treatments
图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-77/img_1.png<b>图1</b> 11号水稻种质在不同浓度盐处理下的生长形态变化<br/><b>Figure 1</b> Growth morphology of No.11 germplasm of rice under different salt concentration treatments Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-77/img_1.png<b>图1</b> 11号水稻种质在不同浓度盐处理下的生长形态变化<br/><b>Figure 1</b> Growth morphology of No.11 germplasm of rice under different salt concentration treatments
图1 11号水稻种质在不同浓度盐处理下的生长形态变化 Figure 1 Growth morphology of No.11 germplasm of rice under different salt concentration treatments
2.2.2 盐胁迫对不同种质叶片死亡率和含水量的影响 盐胁迫下, 不同种质的含水量和叶片死亡率情况见表5。各种质在盐处理10天后, 含水量表现趋势不一致。种质16、17和21号经不同浓度盐处理后的含水量与对照相比呈较低幅度的上升。单因素方差分析表明, 各种质的平均含水量间差异达到极显著水平, 表明这些种质在盐处理下能吸收水分。但是, 16、17和21号种质的平均含水量仅排名9、10和13位, 而19、10和9号种质的平均含水量分别排名前3位。5、6和8号种质经盐处理后失水量最多。各种质的平均叶片死亡率变幅为52.59%-95.41%; 未处理组叶片死亡率变幅为4.87%-43.64%。单因素方差分析表明, 各种质的叶片死亡率差异达到极显著水平, 说明盐胁迫对水稻苗期叶片的生长有显著影响。1、19和20号种质的叶片平均死亡率与其对照相减后, 分别为39.22%、33.67%和34.08%, 低于其它种质。根据平均叶片死亡率分级标准来看, 这3个种质的耐盐性都属于强, 耐盐级别为3级。而在发芽率实验中耐盐性鉴定为极强的10号和鉴定为强的16号种质在不同浓度盐处理下叶片的死亡率却很高, 分别达83.22%和86.57%。16、17和21号种质的叶片平均死亡率分别位列3、2和15, 说明盐处理后, 种质含水量的变化与耐盐性关系不大。叶片死亡率在耐盐种质所测量指标中变化幅度最明显, 不同种质间在相同盐浓度下的变化范围差异较大, 同一种质在不同盐浓度下变化也较明显。因此, 在苗期进行耐盐筛选时叶片死亡率可作为关键指标进行考查。但是, 从上述指标的变化可以看出水稻耐盐机制比较复杂, 故鉴定指标必须慎重选择。不同生育期的鉴定结果只能表示本阶段的耐盐状况, 不能作为全生育期是否耐盐的依据。 表5 Table 5 表5 表5 不同浓度盐处理下部分水稻种质的叶片死亡率和含水量 Table 5 Leaf mortality and water content of part rice germplasms under different salt concentration treatments
No.
NaCl concentration
Average
0
0.3%
0.5%
0.7%
0.9%
1.1%
Water content (%)
Leaf mortality (%)
Water content (%)
Leaf mortality (%)
Water content (%)
Leaf mortality (%)
Water content (%)
Leaf mortality (%)
Water content (%)
Leaf mortality (%)
Water content (%)
Leaf mortality (%)
Water content (%)
Leaf mortal- ity (%)
1
71.60
13.37
69.37
39.50
71.93
24.06
68.13
53.68
63.25
56.66
66.72
89.04
67.88 eE
52.59 qQ
3
72.54
37.44
68.31
57.27
67.33
65.58
64.50
89.79
65.15
90.05
61.47
94.35
65.35 hH
79.41 gG
4
65.21
43.64
60.9
86.26
62.25
97.12
63.94
93.69
54.00
100.00
58.90
100.00
60.02 kK
95.41 aA
5
58.99
32.17
58.32
49.48
44.24
61.40
45.09
81.67
49.40
99.20
41.81
100.00
47.77 pP
78.35 hH
6
65.96
28.14
55.80
64.53
54.95
59.88
55.27
78.24
53.96
91.49
64.07
93.80
56.81 lL
77.59 iI
7
51.76
34.30
59.98
56.32
50.71
84.40
48.66
86.47
51.56
100.00
63.01
100.00
54.78 oO
85.44 dD
8
61.72
15.12
51.85
42.46
53.95
87.83
42.69
84.49
41.18
100.00
42.93
100.00
46.52 qQ
82.96 fF
9
74.42
29.95
75.18
26.63
70.04
71.48
69.25
84.12
73.10
97.96
67.05
100.00
70.92 cC
76.04 kK
10
75.25
33.66
72.65
53.65
73.23
84.40
73.17
81.00
73.96
97.07
72.16
100.00
73.03 bB
83.22 eE
11
67.27
19.64
65.62
42.87
68.42
44.97
68.54
47.71
65.62
91.48
68.66
100.00
67.37 fF
61.41 oO
12
69.14
13.52
70.96
44.81
68.32
30.92
65.46
70.89
65.19
93.70
63.54
99.49
66.70 gG
67.96 mM
14
71.15
20.27
68.51
55.58
70.12
60.57
70.75
81.07
70.76
88.92
70.22
99.60
70.07 dD
77.15 jJ
16
61.67
14.11
64.32
50.08
63.70
87.71
65.50
95.07
63.77
100.00
61.30
100.00
63.72 iI
86.57 cC
17
52.96
4.87
55.11
75.41
58.78
90.19
56.18
100.00
53.77
100.00
56.23
100.00
56.02 mM
93.12 bB
19
76.00
40.14
75.79
54.37
75.74
73.99
75.14
74.93
71.70
81.24
70.60
84.52
73.79 aA
73.81 lL
20
60.01
33.42
55.99
79.19
50.96
53.49
59.31
83.03
59.75
50.41
52.32
71.36
55.67 nN
67.50 nN
21
58.84
18.89
64.62
14.35
67.90
47.86
64.07
42.02
61.10
92.95
58.82
100.00
63.30 jJ
59.44 pP
No. 1, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 14, 16, 17, 19, 20, and 21 see Table 1. The means were the average value of results came from treatments under different salt concentration treatments. Different lowercase letters indicate significant differences at the 0.05 level; and different capital letters indicate significant differences at the 0.01 level. 编号1、3、4、5、6、7、8、9、10、11、12、14、16、17、19、20和21同表1。平均值是指不同浓度盐处理结果的算术平均。不同小写字母表示在0.05水平差异显著; 不同大写字母表示在0.01水平差异显著。
表5 不同浓度盐处理下部分水稻种质的叶片死亡率和含水量 Table 5 Leaf mortality and water content of part rice germplasms under different salt concentration treatments
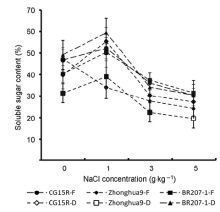
图3 盐胁迫下水稻开花期和蜡熟期可溶性糖含量的比较^F: 开花期; D: 蜡熟期 Figure 3 Comparison of soluble sugar content in flowering and dough period of rice under salt stress^F: Flowering stage; D: Dough stage
2.4.2 盐胁迫对游离脯氨酸含量的影响 脯氨酸是植物蛋白质的最重要组分之一, 在植物细胞中以游离态广泛存在。在干旱、盐碱、冷和热等逆境胁迫下, 植物体内游离脯氨酸(Pro)含量显著增加, 因此游离脯氨酸含量可作为耐盐育种的重要生理指标。从图4A可知, 随盐浓度的不断增加, 3份种质的游离脯氨酸含量均明显增加, 对照组各种质间的游离脯氨酸含量差距很小, 当用5 g∙kg-1盐处理时, 游离脯氨酸含量相差最大, 具体表现为BR207-1>中花9号>CG- 15R。CG15R的游离脯氨酸含量明显低于中花9号。 2.4.3 盐胁迫对丙二醛含量的影响 水稻在盐胁迫等逆境中, 会发生膜脂过氧化反应产生丙二醛(MDA), 因此可以用丙二醛含量来表示膜质过氧化程度, 以此判断植物细胞膜对逆境反应的强弱关系。如图4B所示, 种质BR207-1随着盐处理浓度的增加, 叶片MDA含量呈增加趋势, 且盐浓度在0-1 g∙kg-1范围内增加比较明显。随着盐浓度的增加, CG15R和中花9号的MDA含量不断下降, CG15R的下降趋势明显, 说明随着盐浓度的增加, CG15R和中花9号对盐伤害的防御机制不断完善, 其膜脂过氧化程度不断降低。 2.4.4 盐胁迫对过氧化氢酶活性的影响 在盐胁迫等逆境条件下, 植物体内产生的活性氧对植物细胞产生严重的伤害, 所以过氧化氢酶(CAT)及时有效地清除多余的活性氧具有重要意义。如图4C所示, 3个种质在CAT活性上表现出明显的差异。BR207-1随盐浓度的增加, CAT活性呈下降趋势; 而中花9号在盐浓度为1 g∙kg-1时, CAT活性达到最大值(0.375 μg-1·min-1), 以后逐渐降低; CG15R的CAT活性表现为先减后增的状态, 在1 g∙kg-1时为最低值(0.1375 μg-1·min-1), 在5 g∙kg-1时达最大值(0.775 μg-1·min-1)。以上结果表明, CG15R在较高盐浓度下有较好的防御机制, 通过产生大量的CAT及时清除活性氧, 避免细胞受到伤害。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-77/img_4.png<b>图4</b> 盐胁迫对水稻成株期不同生理指标的影响^(A) 脯氨酸含量; (B) 丙二醛含量; (C) 过氧化氢酶活性; (D) 蜡熟期干物质量<br/><b>Figure 4</b> Effects of salt stress on different physiological indices of rice in adult period^(A) Proline content; (B) Malondialdehyde (MDA) content; (C) Catalase activity; (D) Accumulation of dry matter in dough stage Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-77/img_4.png<b>图4</b> 盐胁迫对水稻成株期不同生理指标的影响^(A) 脯氨酸含量; (B) 丙二醛含量; (C) 过氧化氢酶活性; (D) 蜡熟期干物质量<br/><b>Figure 4</b> Effects of salt stress on different physiological indices of rice in adult period^(A) Proline content; (B) Malondialdehyde (MDA) content; (C) Catalase activity; (D) Accumulation of dry matter in dough stage
图4 盐胁迫对水稻成株期不同生理指标的影响^(A) 脯氨酸含量; (B) 丙二醛含量; (C) 过氧化氢酶活性; (D) 蜡熟期干物质量 Figure 4 Effects of salt stress on different physiological indices of rice in adult period^(A) Proline content; (B) Malondialdehyde (MDA) content; (C) Catalase activity; (D) Accumulation of dry matter in dough stage
AndrewD (2012). The short- and medium- term impacts of rises in staple food prices.Food Secur 4, 633-645. [本文引用: 1]
[19]
BerthomieuP, ConéjéroG, NublatA, BrackenburyWJ, LambertC, SavioC, UozumiN, OikiS, YamadaK, CellierF, GostiF, SimonneauT, EssahPA, TesterM, VéryAA, SentenacH, CasseF (2003). Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is cddrucial for salt tolerance.EMBO J 22, 2004-2014. [本文引用: 1]
[20]
ByrtCS, PlattenJD, SpielmeyerW, JamesRA, LagudahES, DennisES, TesterM, MunnsR (2007). HKT1;5-like cation transporters linked to Na+ exclusion loci in wheat, Nax2 and Kna1. Plant Physiol 143, 1918-1928. [本文引用: 1]
[21]
FooladMR, LinGY (1997). Absence of a relationship bet- ween salt tolerance during germination and vegetative growth in tomato.Plant Breed 116, 363-367. [本文引用: 1]
JabnouneM, EspeoutS, MieuletD, FizamesC, VerdeilJL, ConéjéroG, Rodríguez-NavarroA, SentenacH, Guid- erdoniE, AbdellyC, VéryAA (2009). Diversity in expression patterns and functional properties in the rice HKT transporter family.Plant Physiol 150, 1955-1971.
[24]
JohnsonDW, SmithSE, DobrenzAK (1992). Genetic and phenotypic relationships in response to NaCl at different developmental stages in alfalfa.Theor Appl Genet 83, 833-838. [本文引用: 1]
[25]
KarstenU, FranklinLA, LuningK, WienckeC (1998). Natural ultraviolet radiation and photosynthetically active radiation induce formation of mycosporine- line amino aci- ds in the marine macroalga Chondrus crispus (Phodophyta).Planta 205, 257-262. [本文引用: 1]
[26]
LiXJ, YangMF, ZhuY, LiangY, ShenSH (2011). Proteomic analysis of salt stress responses in rice shoot.J Plant Biol 54, 384-395. [本文引用: 1]
[27]
LinHX, ZhuMZ, YanoM, GaoJP, LiangZW, SuWA, HuXH, RenZH, ChaoDY (2004). QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance.Theor Appl Genet 108, 253-260. [本文引用: 1]
[28]
MäserP, HosooY, GoshimaS, HorieT, EckelmanB, YamadaK, YoshidaK, BakkerEP, ShinmyoA, OikiS, SchroederJI, UozumiN (2002). Glycine residues in potassium channel-like selectivity filters determine potassium selectivity in four-loop-per-subunit HKT transporters from plants.Proc Natl Acad Sci USA 99, 6428-6433. [本文引用: 1]
[29]
MunnsR, JamesRA, XuB, AthmanA, ConnSJ, JordansC, ByrtCS, HareRA, TyermanSD, TesterM, PlettD, GillihamM (2012). Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene.Nature Biotechnol 30, 360-364. [本文引用: 1]
[30]
PlettD, SafwatG, GillihamM, Skrumsager MøllerI, RoyS, ShirleyN, JacobsA, JohnsonA, TesterM (2010). Improved salinity tolerance of rice through cell type-specific expression of AtHKT1;1.PLoS One 5, e12571. [本文引用: 1]
[31]
RenZ, GaoJ, LiL, CaiX, HuangW, ChaoD, ZhuM, WangZ, LuanS, LinH (2005). A rice quantitative trait locus for salt tolerance encodes a sodium transporter.Nat Genet 37, 1141-1146.
[32]
ShereenA, MumtazS, RazaS, KhanMA, SolangiS (2014). Salinity effects on seedling growth and yield components of different inbred rice lines.Pakistan J Bot 37, 131-139.
[33]
Sunarpi, HorieT, MotodaJ, KuboM, YangH, YodaK, HorieR, ChanWY, LeungHY, HattoriK, KonomiM, OsumiM, YamagamiM, SchroederJI, UozumiN (2005). Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na+ unloading from xylem vessels to xylem parenchyma cells.Plant J 44, 928-938. [本文引用: 1]
[34]
VinocurB, AhmanA (2005). Recent advances in engineering plant tolerance to a biotic stress: achievements and limitations.Curr Opin Biotechnol 16, 123-132. [本文引用: 1]
 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}