Abstract: Potassium ion (K+) is the most abundant cation in plants, and it plays an essential role in a number of biochemical and physiological processes during plant growth and development. Potassium deficiency in plants confines root growth, decreases the root to shoot ratio, and limits the transport of assimilates via the phloem. Limited photoassimilate production coupled with transport inhibition can lead to reduction in plant biomass. Therefore, increasing the efficiency of K+ uptake, transport and utilization in plants is important for improving crop productivity. In this review, we summarize the major advances in understanding the physiological responses of plants to K+ deficiency and propose 4 strategies to enhance K+ use efficiency in plants. Further more, We discuss the prospects for increasing K+ acquisition and use efficiency by plants.
植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部。多项研究表明, 众多钾通道参与了这一过程(图2)。拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006)。KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能。在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2)。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-89/img_2.png<b>图2</b> 拟南芥与水稻中参与钾吸收和维管组织钾分配的转运体系(<xref ref-type="bibr" rid="b25-1674-3466-52-1-89">改自Chérel et al., 2014</xref>; <xref ref-type="bibr" rid="b23-1674-3466-52-1-89">Chen et al., 2015b</xref>)<br/><b>Figure 2</b> Transport systems involved in K<sup>+</sup> uptake and its distribution in the plant vasculature of Arabidopsis and rice (<xref ref-type="bibr" rid="b25-1674-3466-52-1-89">modified from Chérel et al., 2014</xref>; <xref ref-type="bibr" rid="b23-1674-3466-52-1-89">Chen et al., 2015b</xref>) Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-89/img_2.png<b>图2</b> 拟南芥与水稻中参与钾吸收和维管组织钾分配的转运体系(<xref ref-type="bibr" rid="b25-1674-3466-52-1-89">改自Chérel et al., 2014</xref>; <xref ref-type="bibr" rid="b23-1674-3466-52-1-89">Chen et al., 2015b</xref>)<br/><b>Figure 2</b> Transport systems involved in K<sup>+</sup> uptake and its distribution in the plant vasculature of Arabidopsis and rice (<xref ref-type="bibr" rid="b25-1674-3466-52-1-89">modified from Chérel et al., 2014</xref>; <xref ref-type="bibr" rid="b23-1674-3466-52-1-89">Chen et al., 2015b</xref>)
AdamsE, ShinR (2014). Transport, signaling, and homeo- stasis of potassium and sodium in plants.J Integr Plant Biol 56, 231-249. [本文引用: 1]
[7]
AhnSJ, ShinR, SchachtmanDP (2004). Expression of KT/KUP genes in Arabidopsis and the role of root hairs in K+ uptake.Plant Physiol 134, 1135-1145. [本文引用: 1]
[8]
AlemánF, Nieves-CordonesM, MartínezV, RubioF (2009). Differential regulation of the HAK5 genes encod- ing the high-affinity K+ transporters of Thellungiella halophila and Arabidopsis thaliana.Environ Exp Bot 65, 263-269. [本文引用: 1]
AmtmannA, HammondJP, ArmengaudP, WhitePJ (2005). Nutrient sensing and signaling in plants: potas- sium and phosphorus.Adv Bot Res 43, 209-257. [本文引用: 1]
[11]
Aranda-SiciliaMN, CagnacO, ChanrojS, SzeH, Rodrí- guez-RosalesMP, VenemaK (2012). Arabidopsis KEA2, a homolog of bacterial KefC, encodes a K+/H+ antiporter with a chloroplast transit peptide.BBA-Biomembranes 1818, 2362-2371. [本文引用: 1]
[12]
ArmengaudP, BreitlingR, AmtmannA (2004). The potassium-dependent transcriptome of Arabidopsis reveals a prominent role of jasmonic acid in nutrient signaling.Plant Physiol 136, 2556-2576. [本文引用: 3]
[13]
AshleyMK, GrantM, GrabovA (2006). Plant responses to potassium deficiencies: a role for potassium transport proteins.J Exp Bot 57, 425-436. [本文引用: 2]
[14]
BañuelosMA, GarciadeblasB, CuberoB, Rodrı?guez- NavarroA (2002). Inventory and functional characterization of the HAK potassium transporters of rice.Plant Physiol 130, 784-795. [本文引用: 1]
[15]
BerthomieuP, ConéjéroG, NublatA, BrackenburyWJ, LambertC, SavioC, UozumiN, OikiS, YamadaK, CellierF, GostiF, SimonneauT, EssahPA, TesterM, VéryAA, SentenacH, CasseF (2003). Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is crucial for salt tolerance.EMBO J 22, 2004-2014. [本文引用: 1]
[16]
BreuningerH, RikirschE, HermannM, UedaM, LauxT (2008). Differential expression of WOX genes mediates apical-basal axis formation in the Arabidopsis embryo.Dev Cell 14, 867-876. [本文引用: 1]
[17]
BrittoDT, KronzuckerHJ (2008). Cellular mechanisms of potassium transport in plants.Physiol Plantarum 133, 637-650. [本文引用: 1]
[18]
CaiJ, ChenL, QuH, LianJ, LiuW, HuY, XuG (2012). Alteration of nutrient allocation and transporter genes expression in rice under N, P, K, and Mg deficiencies.Acta Physiol Plant 34, 939-946. [本文引用: 3]
[19]
CakmakI, HengelerC, MarschnerH (1994a). Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency.J Exp Bot 45, 1245-1250. [本文引用: 4]
[20]
CakmakI, HengelerC, MarschnerH (1994b). Changes in phloem export of sucrose in leaves in response to phosphorus, potassium and magnesium deficiency in bean plants.J Exp Bot 45, 1251-1257. [本文引用: 3]
ChenG, HuQ, LuoL, YangT, ZhangS, HuY, YuL, XuG (2015b). Rice potassium transporter OsHAK1 is essential for maintaining potassium-mediated growth and functions in salt tolerance over low and high potassium concentration ranges.Plant Cell Environ 38, 2747-2765. [本文引用: 6]
[24]
ChengS, HuangY, ZhuN, ZhaoY (2014). The rice WUSCHEL-related homeobox genes are involved in reproductive organ development, hormone signaling and abiotic stress response.Gene 549, 266-274. [本文引用: 1]
ChérelI, MichardE, PlatetN, MoulineK, AlconC, SentenacH, ThibaudJB (2002). Physical and functional interaction of the Arabidopsis K+ channel AKT2 and phosphatase AtPP2CA.Plant Cell 14, 1133-1146. [本文引用: 1]
[27]
ChoSH, YooSC, ZhangH, PandeyaD, KohHJ, HwangJY, KimGT, PaekNC (2013). The rice narrow leaf2 and narrow leaf3 loci encode WUSCHEL-related homeobox 3A (OsWOX3A) and function in leaf, spikelet, tiller and lateral root development.New Phytol 198, 1071-1084. [本文引用: 2]
[28]
Corratgé-FaillieC, JabnouneM, ZimmermannS, VéryAA, FizamesC, SentenacH (2010). Potassium and so- dium transport in non-animal cells: the Trk/Ktr/HKT trans- porter family.Cell Mol Life Sci 67, 2511-2532. [本文引用: 1]
[29]
DamasJM, OliveiraASF, BaptistaAM, SoaresCM (2011). Structural consequences of ATP hydrolysis on the ABC transporter NBD dimer: molecular dynamics studies of HlyB.Protein Sci 20, 1220-1230. [本文引用: 1]
[30]
DaramP, UrbachS, GaymardF, SentenacH, ChérelI (1997). Tetramerization of the AKT1 plant potassium channel involves its C-terminal cytoplasmic domain.EMBO J 16, 3455-3463. [本文引用: 1]
[31]
DavenportRJ, Muñoz-mayorAlicia, JhaD, EssahPA, RusANA, TesterM (2007). The Na+ transporter AtHKT1;1 controls retrieval of Na+ from the xylem in Arabidopsis.Plant Cell Environ 30, 497-507. [本文引用: 1]
[32]
DeanG, CassonS, LindseyK (2004). KNAT6 gene of Arabidopsis is expressed in roots and is required for correct lateral root formation.Plant Mol Biol 54, 71-84. [本文引用: 1]
[33]
DeekenR, GeigerD, FrommJ, KorolevaO, AcheP, Langenfeld-HeyserR, SauerN, MayST, HedrichR (2002). Loss of the AKT2/3 potassium channel affects sugar loading into the phloem of Arabidopsis.Planta 216, 334-344. [本文引用: 3]
[34]
DeekenR, IvashikinaN, CzirjakT, PhilipparK, BeckerD, AcheP, HedrichR (2003). Tumour development in Arabidopsis thaliana involves the Shaker-like K+ channels AKT1 and AKT2/3.Plant J 34, 778-787. [本文引用: 1]
[35]
DesbrossesG, JosefssonC, RigasS, HatzopoulosP, DolanL (2003). AKT1 and TRH1 are required during root hair elongation in Arabidopsis.J Exp Bot 54, 781-788. [本文引用: 2]
[36]
DrewMC, SakerLR, BarberSA, JenkinsW (1984). Changes in the kinetics of phosphate and potassium absorption in nutrient-deficient barley roots measured by a solution-depletion technique.Planta 160, 490-499. [本文引用: 1]
[37]
DubyG, HosyE, FizamesC, AlconC, CostaA, SentenacH, ThibaudJB (2008). AtKC1, a conditionally targeted Shaker-type subunit, regulates the activity of plant K+ channels.Plant J 53, 115-123. [本文引用: 1]
[38]
EpsteinE, RainsDW, ElzamOE (1963). Resolution of dual mechanisms of potassium absorption by barley roots.Proc Natl Acad Sci USA 49, 684-692. [本文引用: 2]
[39]
FuHH, LuanS (1998). AtKUP1: a dual-affinity K+ transporter from Arabidopsis.Plant Cell 10, 63-73. [本文引用: 1]
[40]
FulgenziFR, PeraltaML, ManganoS, DannaCH, VallejoAJ, PuigdomenechP, Santa-MaríaGE (2008). The ionic environment controls the contribution of the barley Hv- HAK1 transporter to potassium acquisition.Plant Physiol 147, 252-262. [本文引用: 1]
[41]
GarciadeblasB, SennME, BanuelosMA, Rodríguez- NavarroA (2003). Sodium transport and HKT transporters: the rice model.Plant J 34, 788-801. [本文引用: 1]
[42]
GattwardJN, AlmeidaAAF, SouzaJO, GomesFP, KronzuckerHJ (2012). Sodium-potassium synergism in Theobroma cacao: stimulation of photosynthesis, water- use efficiency and mineral nutrition.Physiol Plantarum 146, 350-362. [本文引用: 1]
HorieT, BrodskyDE, CostaA, KanekoT, SchiavoFL, KatsuharaM, SchroederJI (2011a). K+ transport by the OsHKT2;4 transporter from rice with a typical Na+ transport properties and competition in permeation of K+ over Mg2+ and Ca2+ ions.Plant Physiol 156, 1493-1507.
[55]
HorieT, CostaA, KimTH, HanMJ, HorieR, LeungHY, MiyaoA, HirochikaH, AnG, SchroederJI (2007). Rice OsHKT2; 1 transporter mediates large Na+ influx component into K+-starved roots for growth.EMBO J 26, 3003-3014. [本文引用: 1]
HorieT, YoshidaK, NakayamaH, YamadaK, OikiS, ShinmyoA (2001). Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa.Plant J 27, 129-138. [本文引用: 2]
[58]
HuertasR, RubioL, CagnacO, García-sánchezMJ, AlchéJDD, VenemaK, FernándezJA, Rodríguez- RosalesMP (2013). The K+/H+ antiporter LeNHX2 increases salt tolerance by improving K+ homeostasis in transgenic tomato.Plant Cell Environ 36, 2135-2149. [本文引用: 1]
[59]
IvashikinaN, BeckerD, AcheP, MeyerhoffO, FelleHH, HedrichR (2001). K+ channel profile and electrical pro- perties of Arabidopsis root hairs.FEBS Lett 508, 463-469. [本文引用: 1]
KimMJ, CianiS, SchachtmanDP (2010). A peroxidase contributes to ROS production during Arabidopsis root response to potassium deficiency.Mol Plant 3, 420-427. [本文引用: 1]
KirikV, SimonM, HuelskampM, SchiefelbeinJ (2004). The enhancer of TRY and CPC1 gene acts redundantly with TRIPTYCHON and CAPRICE in trichome and root hair cell patterning in Arabidopsis.Dev Biol 268, 506-513. [本文引用: 1]
[68]
KochianLV, LucasWJ (1983). Potassium transport in corn roots II. The significance of the root periphery.Plant Physiol 73, 208-215. [本文引用: 1]
[69]
KoyamaML, LevesleyA, KoebnerR, FlowersTJ, YeoAR (2001). Quantitative trait loci for component physiological traits determining salt tolerance in rice.Plant Physiol 125, 406-422. [本文引用: 1]
[70]
LanWZ, LeeSC, CheYF, JiangYQ, LuanS (2011). Mechanistic analysis of AKT1 regulation by the CBL- CIPK-PP2CA interactions.Mol Plant 4, 527-536. [本文引用: 1]
[71]
LanWZ, WangW, WangSM, LiLG, BuchananBB, LinHX, GaoJP, LuanS (2010). A rice high-affinity potassium transporter (HKT) conceals a calcium-permeable cation channel.Proc Natl Acad Sci USA 107, 7089-7094. [本文引用: 1]
[72]
LangA (1983). Turgor-regulated translocation.Plant Cell Environ 6, 683-689. [本文引用: 1]
[73]
LeighRA, Wyn JonesRG (1984). A hypothesis relating critical potassium concentrations for growth to the distribution and functions of this ion in the plant cell.New Phytol 97, 1-13. [本文引用: 1]
[74]
LiescheJ, SchulzA, KrügelU, GrimmB, KühnC (2008). Dimerization and endocytosis of the sucrose transporter StSUT1 in mature sieve elements.Plant Signal Behav 3, 1136-1137. [本文引用: 1]
[75]
LinHX, ZhuMZ, YanoM, GaoJP, LiangZW, SuWA, HuXH, RenZH, ChaoDY (2004). QTLs for Na+ and K+ uptake of the shoots and roots controlling rice salt tolerance.Theor Appl Genet 108, 253-260. [本文引用: 1]
[76]
LiuK, LiL, LuanS (2006). Intracellular K+ sensing of SKOR, a Shaker-type K+ channel from Arabidopsis.Plant J 46, 260-268. [本文引用: 2]
[77]
LuY, ChanrojS, ZulkifliL, JohnsonMA, UozumiN, CheungA, SzeH (2011). Pollen tubes lacking a pair of K+ transporters fail to target ovules in Arabidopsis.Plant Cell 23, 81-93. [本文引用: 1]
[78]
MaTL, WuWH, WangY (2012). Transcriptome analysis of rice root responses to potassium deficiency.BMC Plant Biol 12, 161. [本文引用: 1]
MaathuisFJ, SandersD (1993). Energization of potassium uptake in Arabidopsis thaliana.Planta 191, 302-307. [本文引用: 1]
[82]
MartinT, FrommerWB, SalanoubatM, WillmitzerL (1993). Expression of an Arabidopsis sucrose synthase gene indicates a role in metabolization of sucrose both during phloem loading and in sink organs. Plant J 4, 367-377. [本文引用: 1]
[83]
MäserP, HosooY, GoshimaS, HorieT, EckelmanB, YamadaK, YoshidaK, BakkerEP, ShinmyoA, OikiS, SchroederJI, UozumiN (2002). Glycine residues in potassium channel-like selectivity filters determine potassium selectivity in four-loop-per-subunit HKT transporters from plants.Proc Natl Acad Sci USA 99, 6428-6433. [本文引用: 1]
MontielG, GantetP, Jay-AllemandC, BretonC (2004). Transcription factor networks. Pathways to the knowledge of root development.Plant Physiol 136, 3478-3485. [本文引用: 2]
[86]
Nieves-CordonesM, AlemánF, MartínezV, RubioF (2010). The Arabidopsis thaliana HAK5 K+ transporter is required for plant growth and K+ acquisition from low K+ solutions under saline conditions.Mol Plant 3, 326-333. [本文引用: 1]
[87]
Nieves-CordonesM, Martínez-CorderoM, MartínezV, RubioF (2007). An NH4+-sensitive component dominates high-affinity K+ uptake in tomato plants.Plant Sci 172, 273-280. [本文引用: 3]
[88]
Nieves-CordonesM, MillerAJ, AlemánF, MartínezV, RubioF (2008). A putative role for the plasma membrane potential in the control of the expression of the gene encoding the tomato high-affinity potassium transporter HAK5.Plant Mol Biol 68, 521-532. [本文引用: 1]
[89]
PanditA, RaiV, BalS, SinhaS, KumarV, ChauhanM, GautamRK, SinghR, SharmaPC, SinghAK, GaikwadK, SharmaTR, MohapatraT, SinghNK (2010). Combining QTL mapping and transcriptome profiling of bulked RILs for identification of functional polymorphism for salt tolerance genes in rice (Oryza sativa L.).Mol Genet Genomics 284, 121-136. [本文引用: 1]
[90]
PeelAJ, RogersS (1982). Stimulation of sugar loading into sieve elements of willow by potassium and sodium salts.Planta 154, 94-96. [本文引用: 1]
[91]
PettigrewWT (2008). Potassium influences on yield and quality production for maize, wheat, soybean and cotton.Physiol Plantarum 133, 670-681. [本文引用: 2]
[92]
PhilipparK, IvashikinaN, AcheP, ChristianM, LüthenH, PalmeK, HedrichR (2004). Auxin activates KAT1 and KAT2, two K+-channel genes expressed in seedlings of Arabidopsis thaliana.Plant J 37, 815-827. [本文引用: 1]
[93]
PilotG, GaymardF, MoulineK, ChérelI, SentenacH (2003). Regulated expression of Arabidopsis Shaker K+ channel genes involved in K+ uptake and distribution in the plant.Plant Mol Biol 51, 773-787. [本文引用: 2]
[94]
QiZ, HamptonCR, ShinR, BarklaBJ, WhitePJ, SchachtmanDP (2008). The high affinity K+ transporter AtHAK5 plays a physiological role in planta at very low K+ concentrations and provides a caesium uptake pathway in Arabidopsis.J Exp Bot 59, 595-607. [本文引用: 4]
[95]
QiZ, SpaldingEP (2004). Protection of plasma membrane K+ transport by the salt overly sensitive1 Na+-H+ antiporter during salinity stress.Plant Physiol 136, 2548-2555. [本文引用: 1]
[96]
RenXL, QiGN, FengHQ, ZhaoS, ZhaoSS, WangY, WuWH (2013). Calcineurin B-like protein CBL10 directly interacts with AKT1 and modulates K+ homeostasis in Ara- bidopsis.Plant J 74, 258-266. [本文引用: 1]
[97]
RengelZ, DamonPM (2008). Crops and genotypes differ in efficiency of potassium uptake and use.Physiol Plantarum 133, 624-636. [本文引用: 2]
[98]
RigasS, DebrossesG, HaralampidisK, Vicente-AgulloF, FeldmannKA, GrabovA, DolanL, HatzopoulosP (2001). TRH1 encodes a potassium transporter required for tip growth in Arabidopsis root hairs.Plant Cell 13, 139-151. [本文引用: 1]
[99]
Rodríguez-RosalesMP, JiangX, GálvezFJ, ArandaMN, CuberoB, VenemaK (2008). Overexpression of the tomato K+/H+ antiporter LeNHX2 confers salt tolerance by improving potassium compartmentalization.New Phytol 179, 366-377. [本文引用: 1]
[100]
RosR, LemailletG, FonrougeAG, DaramP, EnjutoM, SalmonJM, ThibaudJB, SentenacH (1999). Molecular determinants of the Arabidopsis AKT1 K+ channel ionic selectivity investigated by expression in yeast of randomly mutated channels.Physiol Plantarum 105, 459-468. [本文引用: 1]
[101]
RubioF, Santa-MaríaGE, Rodríguez-NavarroA (2000). Cloning of Arabidopsis and barley cDNAs encoding HAK potassium transporters in root and shoot cells.Physiol Plantarum 109, 34-43. [本文引用: 1]
[102]
Santa-MaríaGE, RubioF, DubcovskyJ, Rodríguez- NavarroA (1997). The HAK1 gene of barley is a member of a large gene family and encodes a high-affinity potassium transporter.Plant Cell 9, 2281-2289. [本文引用: 1]
SchellmannS, SchnittgerA, KirikV, WadaT, OkadaK, BeermannA, ThumfahrtJ, JürgensG, HülskampM (2002). TRIPTYCHON and CAPRICE mediate lateral inhibition during trichome and root hair patterning in Arabidopsis.EMBO J 21, 5036-5046. [本文引用: 1]
[106]
SharmaT, DreyerI, RiedelsbergerJ (2013). The role of K+ channels in uptake and redistribution of potassium in the model plant Arabidopsis thaliana.Front Plant Sci 4, 224. [本文引用: 1]
[107]
ShengXF, HeLY (2006). Solubilization of potassium- bearing minerals by a wild-type strain of Bacillus eda- phicus and its mutants and increased potassium uptake by wheat.Can J Microbiol 52, 66-72. [本文引用: 1]
[108]
ShimizuA, GuertaCQ, GregorioGB, KawasakiS, IkehashiH (2005). QTLs for nutritional contents of rice seedlings (Oryza sativa L.) in solution cultures and its implication to tolerance to iron-toxicity.Plant Soil 275, 57-66. [本文引用: 1]
[109]
ShinR (2014). Strategies for improving potassium use efficiency in plants.Mol Cells 37, 575-584. [本文引用: 3]
SongZ, YangS, ZhuH, JinM, SuY (2014). Heterologous expression of an alligatorweed high-affinity potassium transporter gene enhances salinity tolerance in Arabidopsis thaliana.Am J Bot 101, 840-850. [本文引用: 1]
[113]
SpaldingEP, HirschRE, LewisDR, QiZ, SussmanMR, LewisBD (1999). Potassium uptake supporting plant growth in the absence of AKT1 channel activity inhibition by ammonium and stimulation by sodium.J Gen Physiol 113, 909-918. [本文引用: 3]
[114]
SunJ, BankstonJR, PayandehJ, HindsTR, ZagottaWN, ZhengN (2014). Crystal structure of the plant dual-affinity nitrate transporter NRT1.1.Nature 507, 73-77. [本文引用: 1]
[115]
SzczerbaMW, BrittoDT, KronzuckerHJ (2009). K+ transport in plants: physiology and molecular biology.J Plant Physiol 166, 447-466. [本文引用: 1]
[116]
Tominaga-WadaR, IwataM, NukumizuY, SanoR, WadaT (2012). A full-length R-like basic-helix-loop-helix trans- cription factor is required for anthocyanin upregulation whereas the N-terminal region regulates epidermal hair formation. Plant Sci 183, 115-122. [本文引用: 1]
[117]
VéryAA, SentenacH (2003). Molecular mechanisms and regulation of K+ transport in higher plants.Annu Rev Plant Biol 54, 575-603. [本文引用: 1]
[118]
Vicente-AgulloF, RigasS, DesbrossesG, DolanL, HatzopoulosP, GrabovA (2004). Potassium carrier TRH1 is required for auxin transport in Arabidopsis roots.Plant J 40, 523-535. [本文引用: 1]
[119]
VoelkerC, SchmidtD, Mueller-RoeberB, CzempinskiK (2006). Members of the Arabidopsis AtTPK/KCO family form homomeric vacuolar channels in planta.Plant J 48, 296-306. [本文引用: 1]
[120]
WadaT, KurataT, TominagaR, Koshino-KimuraY, TachibanaT, GotoK, MarksMD, ShimuraY, OkadaK (2002). Role of a positive regulator of root hair development, CAPRICE, in Arabidopsis root epidermal cell differentiation.Development 129, 5409-5419. [本文引用: 1]
[121]
WalkerDJ, LeighRA, MillerAJ (1996). Potassium homeostasis in vacuolate plant cells.Proc Natl Acad Sci USA 93, 10510-10514. [本文引用: 3]
[122]
WangJG, ZhangFS, ZhangXL, CaoYP (2000). Release of potassium from K-bearing minerals: effect of plant roots under P deficiency.Nutr Cycl Agroecosys 56, 45-52. [本文引用: 1]
[123]
WangTB, GassmannW, RubioF, SchroederJI, GlassAD (1998). Rapid up-regulation of HKT1, a high-affinity potassium transporter gene, in roots of barley and wheat following withdrawal of potassium.Plant Physiol 118, 651-659. [本文引用: 1]
[124]
WangW, LiG, ZhaoJ, ChuH, LinW, ZhangD, WangZ, LiangW (2014). DWARF TILLER1, a WUSCHEL-related homeobox transcription factor, is required for tiller growth in rice.PLoS Genet 10, e1004154. [本文引用: 2]
[125]
WangY, WuWH (2013). Potassium transport and signaling in higher plants.Annu Rev Plant Biol 64, 451-476. [本文引用: 1]
[126]
WangY, WuWH (2015). Genetic approaches for improvement of the crop potassium acquisition and utilization efficiency.Curr Opin Plant Biol 25, 46-52. [本文引用: 1]
[127]
WangYH, GarvinDF, KochianLV (2002). Rapid induction of regulatory and transporter genes in response to phosphorus, potassium, and iron deficiencies in tomato roots. Evidence for cross talk and root/rhizosphere-mediated signals. Plant Physiol 130, 1361-1370. [本文引用: 1]
WhitePJ, GeorgeTS, GregoryPJ, BengoughAG, HallettPD, McKenzieBM (2013b). Matching roots to their environment.Ann Bot 112, 207-222. [本文引用: 1]
[130]
WhitePJ, HammondJP, KingGJ, BowenHC, HaydenRM, MeachamMC, SpracklenWP, BroadleyMR (2010). Genetic analysis of potassium use efficiency in Brassica oleracea.Ann Bot 105, 1199-1210. [本文引用: 1]
[131]
WuP, NiJJ, LuoAC (1998). QTLs underlying rice tolerance to low-potassium stress in rice seedlings.Crop Sci 38, 1458-1462. [本文引用: 1]
[132]
WuX, DabiT, WeigelD (2005). Requirement of homeobox gene STIMPY/WOX9 for Arabidopsis meristem growth and maintenance.Curr Biol 15, 436-440. [本文引用: 1]
[133]
XieQ, FrugisG, ColganD, ChuaNH (2000). Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development.Gene Dev 14, 3024-3036. [本文引用: 1]
[134]
XuJ, LiHD, ChenLQ, WangY, LiuLL, HeL, WuWH (2006). A protein kinase, interacting with two calcineurin B-like proteins, regulates K+ transporter AKT1 in Arabidopsis.Cell 125, 1347-1360. [本文引用: 1]
[135]
YangT, ZhangS, HuY, WuF, HuQ, ChenG, CaiJ, WuT, MoranN, YuL, XuG (2014). The role of a potassium transporter OsHAK5 in potassium acquisition and transport from roots to shoots in rice at low potassium supply levels.Plant Physiol 166, 945-959. [本文引用: 5]
[136]
YangZ, GaoQ, SunC, LiW, GuS, XuC (2009). Molecular evolution and functional divergence of HAK potassium transporter gene family in rice (Oryza sativa L.).J Genet Genomics 36, 161-172. [本文引用: 1]
[137]
YaoWZ, HadjebN, BerkowitzGA (1997). Molecular cloning and characterization of the first plant K (Na)/proton antiporter.Plant Physiol 114S, 200. [本文引用: 2]
[138]
YongZ, KoturZ, GlassAD (2010). Characterization of an intact two-component high-affinity nitrate transporter from Arabidopsis roots. Plant J 63, 739-748. [本文引用: 1]
[139]
YooSC, ChoSH, PaekNC (2013). Rice WUSCHEL-related homeobox 3A (OsWOX3A) modulates auxin-transport gene expression in lateral root and root hair development.Plant Signal Behav 8, e25929. [本文引用: 2]
[140]
ZhangH, FordeBG (1998). An Arabidopsis MADS box gene that controls nutrient-induced changes in root architecture.Science 279, 407-409. [本文引用: 1]
[141]
ZhaoF, GuoXQ, WangP, HeLY, HuangZ, ShengXF (2013). Dyella jiangningensis sp. nov., a γ-proteobacter- ium isolated from the surface of potassium-bearing rock. Int J Syst Evol Micr 63, 3154-3157. [本文引用: 1]
[142]
ZhaoY, HuY, DaiM, HuangL, ZhouDX (2009). The WUSCHEL-related homeobox gene WOX11 is required to activate shoot-borne crown root development in rice.Plant Cell 21, 736-748. [本文引用: 2]
[143]
ZörbC, SenbayramM, PeiterE (2014). Potassium in agriculture—status and perspectives.J Plant Physiol 171, 656-669. [本文引用: 2]
水稻耐低钾胁迫研究进展 1 2012
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
植物钾营养高效分子遗传机制 1 2009
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
盐、碱胁迫条件下粳稻Na+、K+浓度的QTL分析 1 2015
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
不同生长环境下水稻氮、磷、钾利用相关性状的QTL定位分析 1 2015
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 2001
... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2014
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2004
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2009
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2009
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2005
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ...
1 2012
... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ...
3 2004
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
2 2006
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ... ... ; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ...
1 2002
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2003
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 2008
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2008
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
3 2012
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... )中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
4 1994
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... ), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... )中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
3 1994
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... , 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... , 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2013
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
4 2015
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ... ... 培育新型作物品种的主要目标就是提高钾的吸收和利用效率.虽然理论上通过增加根系生物量以及活化钾吸收和转运系统可以增强植物对钾的获取, 但是钾高效利用的植株表型是多种复杂代谢机制的综合体现.许多实验证实, 组成型超表达编码钾转运体或根发育相关转录因子基因并未改善植株体内钾的积累, 甚至会导致负面的生长发育表型.例如, 过量表达钾转运蛋白基因或高亲和钾吸收组分的上游调控元件, 有时虽能促进植株的生长或提高耐盐性, 但并未增加钾的吸收或积累(Song et al., 2014); 组成型超表达OsWOX11不仅使水稻根系大量生长, 而且在茎节和小穗基部也产生不定根, 最终导致地上部发育受到显著抑制, 产量显著降低(Zhao et al., 2009).这可能是由于这些基因的功能冗余或是其时空表达受精密调控所致, 通过利用根系特异的或是缺钾诱导的启动子可以解决上述问题(Chen et al., 2015a, 2015b). ...
6 2015
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ... ... 拟南芥与水稻中参与钾吸收和维管组织钾分配的转运体系(改自Chérel et al., 2014; Chen et al., 2015b) ... ... Transport systems involved in K+ uptake and its distribution in the plant vasculature of Arabidopsis and rice (modified from Chérel et al., 2014; Chen et al., 2015b) ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ... ... ; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ... ... 培育新型作物品种的主要目标就是提高钾的吸收和利用效率.虽然理论上通过增加根系生物量以及活化钾吸收和转运系统可以增强植物对钾的获取, 但是钾高效利用的植株表型是多种复杂代谢机制的综合体现.许多实验证实, 组成型超表达编码钾转运体或根发育相关转录因子基因并未改善植株体内钾的积累, 甚至会导致负面的生长发育表型.例如, 过量表达钾转运蛋白基因或高亲和钾吸收组分的上游调控元件, 有时虽能促进植株的生长或提高耐盐性, 但并未增加钾的吸收或积累(Song et al., 2014); 组成型超表达OsWOX11不仅使水稻根系大量生长, 而且在茎节和小穗基部也产生不定根, 最终导致地上部发育受到显著抑制, 产量显著降低(Zhao et al., 2009).这可能是由于这些基因的功能冗余或是其时空表达受精密调控所致, 通过利用根系特异的或是缺钾诱导的启动子可以解决上述问题(Chen et al., 2015a, 2015b). ...
1 2014
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
7 2014
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ... ... 维持细胞质钾浓度的稳态平衡是确保钾离子参与的很多酶保持生物活性的基础.通过三管微电极对pH值、钾活性和膜电位的测定表明, 缺钾同时造成大麦(Hordeum vulgare)根系表皮和皮层细胞中胞质和液泡的钾活性降低.进一步研究发现, 整个植株钾含量减少的同时, 液泡中钾活性也在下降, 而在根系中, 在较宽的钾浓度范围内, 胞浆中钾活性维持恒定, 在低于25 mmol·L-1 (K+)这一阈值时, 胞质中钾活性才下降(Walker et al., 1996).实验结果表明, 植株通过释放贮存在液泡中的钾, 优先维持胞质中的钾稳态以保证细胞的正常代谢, 这也可以解释为何在缺钾的前几天干物质依旧正常积累.此外, 伴随着胞质中钾的减少, 胞质pH值也随之下降, 表明胞质钾和pH值稳态之间存在生理相关性(Chérel et al., 2014). ... ... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ... ... 植物中参与土壤高、低亲和钾吸收的转运体系(改自Chérel et al., 2014) ... ... 拟南芥与水稻中参与钾吸收和维管组织钾分配的转运体系(改自Chérel et al., 2014; Chen et al., 2015b) ... ... Transport systems involved in K+ uptake and its distribution in the plant vasculature of Arabidopsis and rice (modified from Chérel et al., 2014; Chen et al., 2015b) ...
1 2002
... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ...
2 2013
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... 对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2010
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 2011
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 1997
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2007
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 2004
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
3 2002
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2003
... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
2 2003
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ...
1 1984
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ...
1 2008
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
2 1963
... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ... ... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 1998
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2008
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2003
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 2012
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ...
1 2009
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2010
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
4 2005
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... ; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
4 2007
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... ; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... ), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2008
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2012
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
2 1978
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
3 2006
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... ).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
2 1998
... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ... ... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
2003
3 2013
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
2011
1 2007
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 2011
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
2 2001
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 2013
... 值得一提的是, 维持细胞质中钾离子浓度的稳态平衡可能也是提高钾利用效率的重要措施.已有研究结果表明, 一些属于钾(钠)氢反向转运蛋白(NHX)的成员(如LeNHX2)有助于维持番茄的钾稳态, 该基因的缺失可抑制植株的生长和果实的形成, 而过量表达则显著提高钾耗竭速率以及根系、地上部和木质部伤流液中的钾含量(Rodríguez-Rosales et al., 2008; Huertas et al., 2013). ...
1 2001
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ...
1 2005
... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ...
1 2011
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
4 2009
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... ), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... ; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2007
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2013
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2010
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2012
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2004
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 1983
... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ...
1 2001
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 2011
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2010
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
1 1983
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 1984
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2008
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2004
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
2 2006
... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2011
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ...
1 2012
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ...
1 1997
... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ...
2 1992
... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 1993
... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ...
1 1993
... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2002
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
2 2001
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
2 2004
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... Transport systems involved in high- and low-affinity K+ uptake from soil in plant (modified from Chérel et al., 2014) ...
1 2010
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
3 2007
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2008
... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ...
1 2010
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 1982
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
2 2008
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2004
... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
2 2003
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
4 2008
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... ; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... ; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... 的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2004
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ...
1 2013
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
2 2008
... 增加土壤中钾的移动性或促进钾从土壤含钾矿物的非交换性形态释放成为土壤胶体颗粒表面的阳离子交换性钾, 可提高植物根系对土壤钾的有效吸收(Rengel and Damon, 2008).适度提高土壤含水量、增强土壤通透性可以促进钾在土壤中的移动.根系分泌物有利于钾移动性的提高, 植物释放诸如苹果酸和酒石酸之类的有机酸, 可提高土壤中钾的移动性(Wang et al., 2000).植物根系接种芽孢杆菌(Bacillus spp.)可以增加根系分泌物, 提高玉米(Zea mays)对钾的吸收并促进植株生长(Sheng and He, 2006; Rengel and Damon, 2008; Zhao et al., 2013).因此, 增加土壤中的有益微生物也是提高钾可利用性的有效方法. ... ... ; Rengel and Damon, 2008; Zhao et al., 2013).因此, 增加土壤中的有益微生物也是提高钾可利用性的有效方法. ...
1 2001
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ...
1 2008
... 值得一提的是, 维持细胞质中钾离子浓度的稳态平衡可能也是提高钾利用效率的重要措施.已有研究结果表明, 一些属于钾(钠)氢反向转运蛋白(NHX)的成员(如LeNHX2)有助于维持番茄的钾稳态, 该基因的缺失可抑制植株的生长和果实的形成, 而过量表达则显著提高钾耗竭速率以及根系、地上部和木质部伤流液中的钾含量(Rodríguez-Rosales et al., 2008; Huertas et al., 2013). ...
1 1999
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2000
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 1997
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2012
... Shaker型钾通道和KT/KUP/HAK转运蛋白普遍存在于双子叶和单子叶植物中(Chérel et al., 2014).在单子叶植物中, 除了上述两大家族以外, 其它膜蛋白也具有钾转运功能(图1).HKT属于H+-Na+/K+共转运体, 与原核生物的TrkH转运蛋白和酵母的Trk转运蛋白类似, 普遍存在于所有植物中(Corratgé-Faillie et al., 2010).单子叶植物的HKT转运蛋白参与钾和钠的吸收, 但是双子叶植物的HKT只行使钠的转运功能(Horie et al., 2001, 2007; Berthomieu et al., 2003; Garciadeblas et al., 2003; Davenport et al., 2007).OsHKT2;4在水稻根外周细胞的细胞膜上具有钾和钠的共转运功能(Lan et al., 2010; Horie et al., 2011a), 且更倾向于转运钾(Sassi et al., 2012).对HKT转运蛋白的分子解析表明, 将双子叶植物中的HKT氨基酸序列进行取代(丝氨酸变为甘氨酸)后, 便能转运钾(Mäser et al., 2002). ...
2 2007
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2002
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2013
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2006
... 增加土壤中钾的移动性或促进钾从土壤含钾矿物的非交换性形态释放成为土壤胶体颗粒表面的阳离子交换性钾, 可提高植物根系对土壤钾的有效吸收(Rengel and Damon, 2008).适度提高土壤含水量、增强土壤通透性可以促进钾在土壤中的移动.根系分泌物有利于钾移动性的提高, 植物释放诸如苹果酸和酒石酸之类的有机酸, 可提高土壤中钾的移动性(Wang et al., 2000).植物根系接种芽孢杆菌(Bacillus spp.)可以增加根系分泌物, 提高玉米(Zea mays)对钾的吸收并促进植株生长(Sheng and He, 2006; Rengel and Damon, 2008; Zhao et al., 2013).因此, 增加土壤中的有益微生物也是提高钾可利用性的有效方法. ...
1 2005
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
3 2014
... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ... ... ).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ... ... ; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
2 2007
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... ).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
2 2004
... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ... ... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2014
... 培育新型作物品种的主要目标就是提高钾的吸收和利用效率.虽然理论上通过增加根系生物量以及活化钾吸收和转运系统可以增强植物对钾的获取, 但是钾高效利用的植株表型是多种复杂代谢机制的综合体现.许多实验证实, 组成型超表达编码钾转运体或根发育相关转录因子基因并未改善植株体内钾的积累, 甚至会导致负面的生长发育表型.例如, 过量表达钾转运蛋白基因或高亲和钾吸收组分的上游调控元件, 有时虽能促进植株的生长或提高耐盐性, 但并未增加钾的吸收或积累(Song et al., 2014); 组成型超表达OsWOX11不仅使水稻根系大量生长, 而且在茎节和小穗基部也产生不定根, 最终导致地上部发育受到显著抑制, 产量显著降低(Zhao et al., 2009).这可能是由于这些基因的功能冗余或是其时空表达受精密调控所致, 通过利用根系特异的或是缺钾诱导的启动子可以解决上述问题(Chen et al., 2015a, 2015b). ...
3 1999
... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ... ... 根系钾的吸收呈现出典型的双相动力学特征.根据Epstein模型, 植物对钾的转运有2种不同的机制, 一种在低钾 (0.001-0.2 mmol·L-1)条件下起作用; 而另一种在高钾(1-10 mmol·L-1)条件下起作用(Epstein et al., 1963).通常认为, 高亲和钾的吸收多是由同向转运蛋白(H+-K+同向转运体)介导的主动运输; 低亲和钾吸收是由通道介导的被动运输(Kochian and Lucas, 1983; Maathuis and Sanders, 1992).然而, 当钾通道蛋白AKT1被证实能在低于100 µmol·L-1的培养介质下参与钾的吸收以后, 转运体和通道蛋白对钾吸收亲和性的划分理论被重新修正(Hirsch et al., 1998; Spalding et al., 1999).钾饥饿增强了植株对低浓度钾的吸收.Drew等(1984)在植株缺钾1天后进行钾耗竭实验, 显示钾最大吸收速率不变, 而钾转运体系的亲和性显著增加(Km值降低为原来的25%).在拟南芥(Arabidopsis thaliana)中, 缺钾仅6小时就已显著改变钾的吸收动力学(Shin and Schachtman, 2004).在缺钾胁迫下, 番茄(Solanum lycopersicon)中高亲和钾吸收蛋白被强烈诱导表达(Nieves-Cordones et al., 2007). ... ... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2014
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2009
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2012
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2003
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2004
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
1 2006
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2002
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
3 1996
... 钾在维持离子稳态、渗透调节、蛋白代谢、酶活、膜极化和各种代谢过程中起关键作用(Hastings and Gutknecht, 1978; Walker et al., 1996; Amtmann et al., 2005; Schachtman and Shin, 2007).植物重要的生理过程, 如光合作用、光呼吸以及生长发育, 均受到钾的影响(Pettogrew, 2008; Gattward et al., 2012).土壤中水溶性钾浓度范围大多为0.1-1 mmol·L-1, 由于局部耗竭, 植物根际土壤中的钾浓度会显著低于上述水平(Chérel et al., 2014).植物的生长环境会限制钾的利用, 钾缺乏可导致植物体内总钾量降低, 在根部的降低尤为迅速(Drew et al., 1984; Pilot et al., 2003; Gierth et al., 2005; Nieves-Cordones et al., 2007; Ma et al., 2012).植物长期缺钾会出现生长受抑制和黄化病, 并首先在老叶中显现(Armeng- aud et al., 2004).在农业生产中, 发展中国家常采取施用化肥来提高作物的产量和质量.然而, 大量使用化肥不仅不能提高产量, 反而会导致土壤中营养元素的流失.因此, 通过有效的生物技术手段, 提高植物自身对土壤中有效钾的利用, 减少与完善钾肥的使用与管理, 对于生态环境的保护和资源的可持续利用有重要意义. ... ... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ... ... 维持细胞质钾浓度的稳态平衡是确保钾离子参与的很多酶保持生物活性的基础.通过三管微电极对pH值、钾活性和膜电位的测定表明, 缺钾同时造成大麦(Hordeum vulgare)根系表皮和皮层细胞中胞质和液泡的钾活性降低.进一步研究发现, 整个植株钾含量减少的同时, 液泡中钾活性也在下降, 而在根系中, 在较宽的钾浓度范围内, 胞浆中钾活性维持恒定, 在低于25 mmol·L-1 (K+)这一阈值时, 胞质中钾活性才下降(Walker et al., 1996).实验结果表明, 植株通过释放贮存在液泡中的钾, 优先维持胞质中的钾稳态以保证细胞的正常代谢, 这也可以解释为何在缺钾的前几天干物质依旧正常积累.此外, 伴随着胞质中钾的减少, 胞质pH值也随之下降, 表明胞质钾和pH值稳态之间存在生理相关性(Chérel et al., 2014). ...
1 2000
... 增加土壤中钾的移动性或促进钾从土壤含钾矿物的非交换性形态释放成为土壤胶体颗粒表面的阳离子交换性钾, 可提高植物根系对土壤钾的有效吸收(Rengel and Damon, 2008).适度提高土壤含水量、增强土壤通透性可以促进钾在土壤中的移动.根系分泌物有利于钾移动性的提高, 植物释放诸如苹果酸和酒石酸之类的有机酸, 可提高土壤中钾的移动性(Wang et al., 2000).植物根系接种芽孢杆菌(Bacillus spp.)可以增加根系分泌物, 提高玉米(Zea mays)对钾的吸收并促进植株生长(Sheng and He, 2006; Rengel and Damon, 2008; Zhao et al., 2013).因此, 增加土壤中的有益微生物也是提高钾可利用性的有效方法. ...
1 1998
... 植物不论处在何种供钾条件下, 细胞的膜电位均会受到外界钾浓度的影响而快速且可逆地发生变化, 降低钾浓度使膜电位更趋于负值.这一普遍现象归因于钾转运体系对细胞膜电位建立的显著作用, 与H+-ATP酶质子泵介导的H+外排共同导致细胞膜的超极化(Maathuis and Sanders, 1993; Maathuis et al., 1997; Wang et al., 1998; Spalding et al., 1999).随着缺钾胁迫时间的延长, 低钾植株较正常供钾植株表现出膜电位负值的绝对值更大(Walker et al., 1996; Nieves-Cordones et al., 2008).细胞膜的电化学极化可能参与感受外界钾的可利用性或者转导植株钾营养状况信号(Chérel et al., 2014). ...
2 2014
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... ; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2013
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
1 2015
... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2002
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
1 2013
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 2013
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 2010
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 1998
... 提高营养元素吸收和利用效率是以增产为目的的育种尝试的焦点.不同作物品种之间钾的吸收和利用效率差异很大, 并且根系性状显著影响产量(White et al., 2013a, 2013b).不同的研究者采用不同的群体在不同的环境下定位了一些水稻耐低钾胁迫相关性状的QTL (Wu et al., 1998; Koyama et al., 2001; Lin et al., 2004; Shimizu et al., 2005; Pandit et al., 2010; 陈艳丽等, 2012; 邢军等, 2015; 杨树明等, 2015).拟南芥中与地上部钾积累有关的QTLs (如KUP2、ATK2、KAT2和TPK3)均定位在第4号染色体上(White et al., 2010), 利用这些QTLs可以进行分子标记辅助育种.四倍体拟南芥较二倍体或单倍体具有更高的钾积累, 说明染色体倍性水平也可用于提高钾的积累(Chao et al., 2013). ...
1 2005
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2000
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2006
... 为了维持植株体内适宜浓度的钾, 植物细胞要监测根际钾的有效浓度.尽管植物感受可利用钾的具体机制尚未被透彻阐明, 但钾通道和钾载体仍被普遍认为是关键的参与者(Hastings and Gutknecht, 1978; Leigh and Wyn Jones, 1984; Fu and Luan, 1998; Gierth et al., 2005; Voelker et al., 2006; Grabov, 2007; Schachtman and Shin, 2007; Amtmann and Blatt, 2009; Wang and Wu, 2013; Adams and Shin, 2014).高等植物具有双亲和钾吸收系统(Epstein et al., 1963).当土壤含钾充足时(>100 µmol·L-1), 植物主要通过内向Shaker型钾通道介导低亲和的钾吸收.在拟南芥中, 定位于根外层细胞质膜上的内向Sha- ker型钾通道AKT1, 在低亲和钾吸收系统中发挥主要作用(Hirsch et al., 1998; Ros et al., 1999; Spalding et al., 1999).植物钾通道之间可以形成复合物, 这些复合物可被视为钾通道的功能性亚基, 并且每个亚基都能响应较宽范围的钾转导(Véry and Sentenac, 2003).AKT1可以与另一钾通道蛋白KC1形成同源或异源四聚体, 这一异源复合物可以作为一个功能性亚基, KC1单独并不能行使钾内流通道的功能, AKT1自身可以形成一个有功能的同源复合物, 具有与AKT1- KC1不同的特性(Daram et al., 1997; Duby et al., 2008; Geiger et al., 2009; Jeanguenin et al., 2011).而AKT1-KC1复合物受CBL蛋白、CIPK和SNARE蛋白转录后水平的调控(Xu et al., 2006; 王毅和武维华, 2009; Lan et al., 2011; Ren et al., 2013). ...
5 2014
... 尽管植物可以吸收土壤溶液中的钾, 但是土壤中的多数钾被固定或以结晶的形式存在, 因此可以利用的有效钾含量很低(Ashley et al., 2006).此外, 土壤中其它组分的干扰也会限制钾的有效吸收.如高浓度的Na+和NH4+可阻碍根系钾的吸收(Qi and Spalding, 2004; Ashley et al., 2006; Yang et al., 2014; Chen et al., 2015b), 土壤含水量也影响植物对钾的获取.钾的有效性是调控诸如根毛发育(Ivashikina et al., 2001; Rigas et al., 2001; Desbrosses et al., 2003)和花粉管伸长(Lu et al., 2011)等细胞生长过程的关键因子之一.因此, 在外界钾含量不足的条件下, 植物的生长发育显著受阻.植物至少具备两大机制来适应缺钾胁迫: 一方面通过激活高亲和钾吸收系统; 另一方面用增加根系体积来提高与土壤的接触面积, 从而增强对钾的获取.植株在缺钾条件下如果不能提高对外源钾的吸收或内源钾(如液泡中贮存的钾)的再分配利用, 便会减缓自身代谢并逐渐停止生长.为了解决作物因缺钾胁迫而造成的减产问题, 可以通过增施钾肥或找到能更高效吸收、转运和利用钾的方法来实现.基于目前的理论, 我们总结有四大策略可用于提高植株钾的利用效率. ... ... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ... ... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ... ... ; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2009
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
2 1997
... 植物通过上述不同的钾通道和载体从土壤中吸收钾, 之后需要将钾长距离运输到地上部.多项研究表明, 众多钾通道参与了这一过程(图2).拟南芥外流钾通道SKOR起到将钾从根系薄壁细胞外运到木质部的关键作用, 该过程是钾长距离运输的重要步骤(Liu et al., 2006).KEAs (Yao et al., 1997; Aranda- Sicilia et al., 2012)和AKT2/3 (Chérel et al., 2002; Deeken et al., 2002; Ivashikina et al., 2005)已被证实分别具有钾的木质部装载和韧皮部运输功能.在低钾条件下, OsHAK1 (Chen et al., 2015b)和OsHAK5 (Yang et al., 2014)均可促进钾从水稻根系向地上部的转运(图2). ... ... 钾的吸收和转运是精密调控的复杂而独立的过程.增加钾的吸收和提高钾的转运效率, 对作物的增产具有显著的促进作用.进一步研究钾转运蛋白和通道的功能以及调控通路将有助于促进植物对钾的吸收效率(Shin, 2014; Wang and Wu, 2015).增强钾的吸收动力学是提高钾吸收效率最直接的途径之一, 可以通过激活诸如钾载体、钾通道和钾载体/通道互作蛋白之类的钾吸收组分及其上游调控元件来实现(Shin, 2014).在低钾逆境下活化高亲和钾吸收体系有利于钾的吸收(Yang et al., 2014; Chen et al., 2015b).过量表达高亲和钾转运蛋白基因AtHAK5的正调控转录因子(DDF2、JLO、bHLH121和TFII_A)可以提高拟南芥对低钾的耐受性(Hong et al., 2013).除了增强钾的吸收, 促进钾在体内的转运也可有效提高钾的利用效率.钾在韧皮部的转运效率与植株地上部的生长直接相关(Martin et al., 1993; Ache et al., 2001; Deeken et al., 2002).SKOR在拟南芥中介导钾从根到地上部长距离运输的第1步(Liu et al., 2006); KAT2、AKT2/3、KEAs、OsHAK5和OsHAK1均参与体内钾的转运(Yao et al., 1997; Mäser et al., 2001; Deeken et al., 2003; Philippar et al., 2004; Adams and Shin, 2014; Yang et al., 2014; Chen et al., 2015b).用适当方法调控这些基因的表达可以有效提高植物对钾的利用效率. ...
1 2010
... 与低亲和钾吸收系统不同, 由电化学势驱动的载体组成的高亲和钾吸收系统在外界钾浓度较低(<100 µmol·L-1)的情况下被激活(Maathuis and Sanders, 1992; Grabov, 2007; Britto and Kronzucker, 2008).KT/KUP/HAK转运蛋白介导H+-K+共转运, 在高等植物中被证实具有高亲和钾转运的作用.大麦HvHAK1是在植物中第1个被鉴定的KT/KUP/HAK型钾转运蛋白, 编码该蛋白的基因受钾饥饿诱导表达并介导高亲和钾吸收(Santa-Maria et al., 1997).在高等植物中, 已发现大量的KT/KUP/HAK转运蛋白基因.在拟南芥和水稻中, KT/KUP/HAK转运蛋白家族分别由13个和27个成员组成, 分为4个主要集团(Rubio et al., 2000; Mäser et al., 2001; Grabov, 2007; Gupta et al., 2008; Yang et al., 2009), 已证实其中位于I集团的KT/KUP/HAK转运蛋白属于高亲和钾转运蛋白(Grabov, 2007; Sharma et al., 2013).在I集团转运蛋白中, 对AtHAK5的研究最为透彻(Gierth et al., 2005; Qi et al., 2008; Nieves-Cordones et al., 2010; Hong et al., 2013).AtHAK5受缺钾诱导表达增强, 而在恢复供钾后表达又降低(Ahn et al., 2004; Qi et al., 2008), 其它属于I集团的转运蛋白基因, 如CaHAK1、LeHAK5、ThHAK5、OsHAK5和OsHAK1, 也被证实受缺钾诱导上调表达, 并且在酵母或大肠异源体系中介导高亲和钾吸收(Bañuelos et al., 2002; Wang et al., 2002; Gierth et al., 2005; Nieves-Cordones et al., 2007; Fulgenzi et al., 2008; Qi et al., 2008; Alemán et al., 2009; Horie et al., 2011b; Yang et al., 2014; Chen et al., 2015b).KT/KUP/HAK转运蛋白与内向Shaker型钾通道在离子选择方面的不同在于它们不能快速区别诸如Na+、Rb+和Cs+之类的碱金属离子.此外, 由KT/KUP/HAK转运蛋白介导的高亲和钾转运受到NH4+的强烈抑制(Qi et al., 2008; Szczerba et al., 2009).Chen等(2015b)的研究表明, OsHAK1在高及低钾条件下均可介导根系对钾的吸收, NH4+显著抑制纯硝酸钾溶液中OsHAK1介导的高亲和钾吸收速率, 但对其介导的低亲和钾吸收无显著抑制作用.KT/KUP/HAK转运蛋白复合物在植物体中尚未被发现, 而在高等植物中, 很多种类的共转运体都是以二聚体的形式存在, 如硝酸盐转运蛋白NRT1;1 (Yong et al., 2010; Sun et al., 2014)、ABC转运蛋白(Damas et al., 2011)和蔗糖转运蛋白StSUT1 (Liesche et al., 2008).因此推测KT/KUP/HAK转运蛋白可能也是二聚体, 但有待进一步证实. ...
2 2013
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... ; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 1998
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ...
1 2013
... 增加土壤中钾的移动性或促进钾从土壤含钾矿物的非交换性形态释放成为土壤胶体颗粒表面的阳离子交换性钾, 可提高植物根系对土壤钾的有效吸收(Rengel and Damon, 2008).适度提高土壤含水量、增强土壤通透性可以促进钾在土壤中的移动.根系分泌物有利于钾移动性的提高, 植物释放诸如苹果酸和酒石酸之类的有机酸, 可提高土壤中钾的移动性(Wang et al., 2000).植物根系接种芽孢杆菌(Bacillus spp.)可以增加根系分泌物, 提高玉米(Zea mays)对钾的吸收并促进植株生长(Sheng and He, 2006; Rengel and Damon, 2008; Zhao et al., 2013).因此, 增加土壤中的有益微生物也是提高钾可利用性的有效方法. ...
2 2009
... 将拟南芥置于缺钾环境下28小时即可促进根毛的伸长, 这被视为植株适应缺钾环境的响应, 即在短时间内通过增加根系体积以获得更多可利用的钾(Jung et al., 2009).延长钾饥饿时间, 会导致地上部生物量降低, 并且主根和侧根发育均受到抑制(Armengaud et al., 2004; Shin et al., 2007).低钾胁迫下根毛长度增加的现象, 在豌豆(Pisum sativum)、红三叶(Trifolium pratense)、紫苜蓿(Medicago sativa)、大麦、黑麦(Secale cereale)、黑麦草(Lolium perenne)和欧洲油菜(Brassica napus)等植物中均有发现(Høgh-Jen- sen and Pedersen, 2003).因此, 植物可能通过增加根系体积以应对短期的土壤低钾胁迫.然而, 与缺氮、缺磷相比, 在长时间缺钾情况下, 植物根系的生长受到强烈的抑制, 根冠比反而会显著下降(Hermans et al., 2006; Cai et al., 2012; Chen et al., 2015a).通过超表达直接参与根毛发育过程的转录因子, 可以达到增加根系体积的目的.MYB转录因子CPC是根毛分化的正调控因子(Schellmann et al., 2002; Wada et al., 2002; Kirik et al., 2004).拟南芥CPC启动子启动的番茄SITRY基因(类似CPC的MYB基因)在拟南芥CPC突变体cpc-2中过量表达时, 可增加根毛的数量(Tominaga-Wada et al., 2012).过量表达低钾诱导的AP2/ERF转录因子11 (RAP2.11)可增加根毛长度(Kim et al., 2012).在拟南芥中超表达棉花bHLH结构域(含TCP转录因子)可正调控根毛的发生和伸长(Hao et al., 2012).据报道, 包括MYB77 (Shin et al., 2007)、NAC1 (Xie et al., 2000)、KNAT6 (Dean et al., 2004)和ANR1 (Zhang and Forde, 1998; Montiel et al., 2004)在内的一些拟南芥转录因子, 都对侧根发生具有正调控作用.WUSCHEL相关的同源异形盒(homeobox)基因WOX是发育的调控因子(Wang et al., 2014).该基因家族在拟南芥和水稻中至少分别由15和13个成员组成, 其中一些成员参与调控植物根系发育过程中细胞的分裂和分化(Cho et al., 2013; Yoo et al., 2013; Cheng et al., 2014).例如, 拟南芥WOX2和WOX8调控胚根形成过程中细胞的分化(Breuninger et al., 2008), WOX9参与根尖早期的胚性生长(Wu et al., 2005; Wang et al., 2014); 水稻OsWOX3A对侧根和根毛的发育均起作用(Cho et al., 2013; Yoo et al., 2013), OsWOX11通过直接抑制不定根原基中A型细胞分裂素响应因子RR2的表达, 对不定根的发生和生长起关键作用(Zhao et al., 2009).Chen等(2015a)选用OsHAK16基因(根系中表达量高且受缺钾诱导表达显著增强)的启动子启动OsWOX- 11在根系中表达, 显著促进了根系的生长、总钾的积累和缺钾逆境下的单株产量.因此, 用低钾诱导上调表达基因的启动子(如OsHAK16)启动上述转录因子, 使其在根中特异高表达, 是提高钾利用效率的可行策略之一. ... ... 培育新型作物品种的主要目标就是提高钾的吸收和利用效率.虽然理论上通过增加根系生物量以及活化钾吸收和转运系统可以增强植物对钾的获取, 但是钾高效利用的植株表型是多种复杂代谢机制的综合体现.许多实验证实, 组成型超表达编码钾转运体或根发育相关转录因子基因并未改善植株体内钾的积累, 甚至会导致负面的生长发育表型.例如, 过量表达钾转运蛋白基因或高亲和钾吸收组分的上游调控元件, 有时虽能促进植株的生长或提高耐盐性, 但并未增加钾的吸收或积累(Song et al., 2014); 组成型超表达OsWOX11不仅使水稻根系大量生长, 而且在茎节和小穗基部也产生不定根, 最终导致地上部发育受到显著抑制, 产量显著降低(Zhao et al., 2009).这可能是由于这些基因的功能冗余或是其时空表达受精密调控所致, 通过利用根系特异的或是缺钾诱导的启动子可以解决上述问题(Chen et al., 2015a, 2015b). ...
2 2014
... 钾对蔗糖的装载和质流推动的溶质在韧皮部筛管中的运输起重要作用(Peel and Rogers, 1982; Lang, 1983).缺钾植物中, 韧皮部的蔗糖装载受到抑制(Deeken et al., 2002; Pilot et al., 2003), 向根部的转运量显著减少, 导致根的蔗糖浓度显著降低(Cakmak et al., 1994a, 1994b; Hermans et al., 2006).在钾充足供应的甘蔗(Saccharum sinense)中, 半数以上14C标记的光合产物在90分钟内就从源叶转运到其它器官, 其中20%转运至茎秆这一主要贮存器官; 而在缺钾胁迫下, 光合产物的外运速率显著降低(Cakmak et al., 1994a, 1994b).缺钾胁迫使光合产物在成熟叶片(源叶)中的含量显著增加(糖转运速率显著降低) (Hermans et al., 2006; Cai et al., 2012), 而向生长的根系(Cakmak et al., 1994a, 1994b)、嫩芽(Pet- tigrew, 2008)、发育的叶片(Gerardeaux et al., 2010)和果实(Kanai et al., 2007)中的输送量显著减少.因此, 缺钾影响植物地上部和根系的生长发育(Cakmak et al., 1994a; Jung et al., 2009).在根中, 缺钾阻碍侧根的起始生长和发育(Armengaud et al., 2004; Shin and Schachtman, 2004), 抑制主根的生长(Jung et al., 2009; Kim et al., 2010), 促进根毛的伸长(Desbrosses et al., 2003; Jung et al., 2009), 根呈现出轻微的背重力性生长(Vicente-Agullo et al., 2004).拟南芥主根和侧根对缺钾胁迫的响应不一致, 导致根系构型发生显著变化(Kellermeier et al., 2013).在水稻(Oryza sativa)中, 缺钾胁迫可抑制根系生长并降低植株的根冠比(Cai et al., 2012).在地上部, 缺钾的早期表型是老叶叶尖失绿(Zörb et al., 2014).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ... ... ).随着缺钾时间的延长, 失绿现象会从老叶转移到新叶.当缺钾进一步加剧, 植株叶片则从叶尖到整个叶缘都呈现出失绿直至坏死, 随着叶片中腐胺的积累, 失绿的同时还伴有褐斑的大量出现.严重缺钾植株会呈现萎蔫的症状, 这主要是由于体内的水分平衡被破坏, 同时细胞壁的木质化过程受阻.因此, 细胞会对真菌侵染表现出更高的敏感性, 最终导致产量减少和品质下降(Zörb et al., 2014). ...
 ,
, 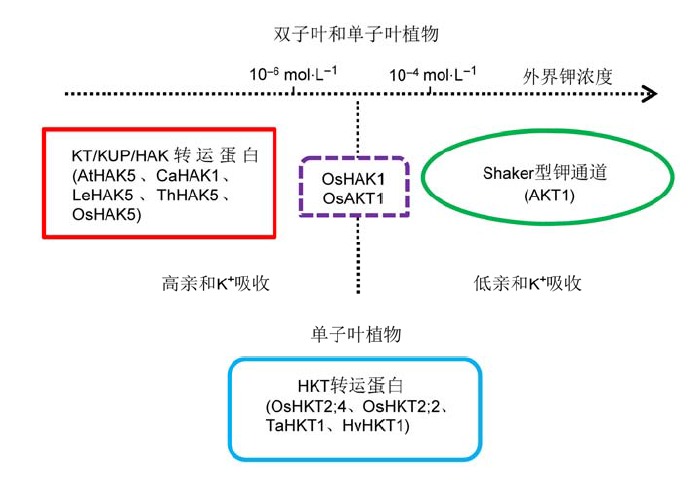
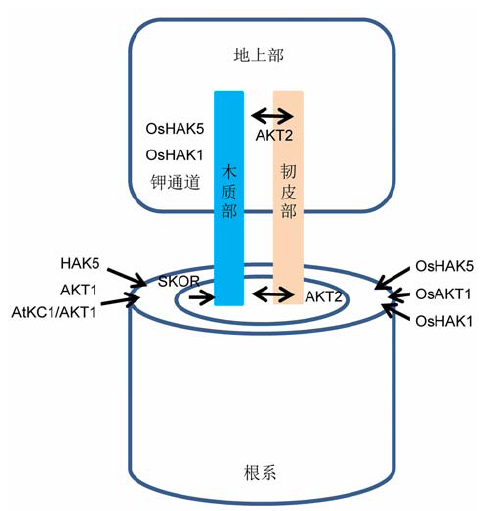

{kind=link}
{kind=link}
{kind=link}
{kind=link}