张景慧
1 , 2 , 3 , * , 王铮
1 , * , 黄永梅
, 4 , * , 陈慧颖
5 , 李智勇
1 , 2 , 3 , 梁存柱
1 , 2 , 3 1 内蒙古大学生态与环境学院, 呼和浩特 0100212 蒙古高原生态学与资源利用教育部重点实验室, 呼和浩特 0100213 省部共建草地生态学国家重点实验室培育基地, 呼和浩特 0100214 北京师范大学地理科学学部, 地表过程与资源生态国家重点实验室, 北京 1008755 人民教育出版社课程教材研究所, 北京 100081Effects of grassland utilization on the functional traits of dominant plants in a temperate typical steppe Jing-Hui ZHANG
1 , 2 , 3 , * , Zheng WANG
1 , * , Yong-Mei HUANG
, 4 , * , Hui-Ying CHEN
5 , Zhi-Yong LI
1 , 2 , 3 , Cun-Zhu LIANG
1 , 2 , 3 1 School of Ecology and Environment, Inner Mongolia University, Hohhot 010021, China2 Key Laboratory of Mongolian Plateau Ecology and Resource Utilization, Ministry of Education, Hohhot 010021, China3 Provincial and Ministry Jointly Build the Cultivation Base of State Key Laboratory of Grassland Ecology, Hohhot 010021, China4 Faculty of Geographical Science, State Key Laboratory of Earth Surface Processes and Resource Ecology, Beijing Normal University, Beijing 100875, China5 Curriculum and Teaching Materials Research Institute, Peopleʼs Education Press, Beijing 100081, China通讯作者: **
ymhuang@bnu.edu.cn * 同等贡献Contributed equally to this work编委: 王德利
责任编辑: 赵航
收稿日期: 2020-11-12
修回日期: 2021-04-2
基金资助: 国家重点研发计划 (2016YFC0500503 )国家自然科学基金 (31960243 )内蒙古自然科学基金 (2018BS03006 )
Corresponding authors: **
ymhuang@bnu.edu.cn Received: 2020-11-12
Revised: 2021-04-2
Fund supported: National Key R&D Program of China (2016YFC0500503 )National Natural Science Foundation of China (31960243 )Natural Science Foundation of Nei Mongol (2018BS03006 )
摘要 当外界环境发生变化后植物能够改变自身功能性状及时调整适应策略, 因此植物功能性状能够有效地反映植物对草地利用变化的响应, 然而在内蒙古草原从植物功能性状角度开展草地利用方式影响的研究略少。该研究以内蒙古典型草原大针茅(
Stipa grandis )、羊草(
Leymus chinensis )、糙隐子草(
Cleistogenes squarrosa )和冷蒿(
Artemisia frigida ) 4种主要优势种为研究对象, 探讨在长期自由放牧、割草、短期围封和长期无干扰的影响下优势种植物功能性状的差异, 以期从功能性状视角, 揭示植物在受到外界干扰后的适应策略, 旨为天然草地的可持续管理提供基础数据支持和科学依据。结果表明: 1)除糙隐子草外, 在长期放牧后内蒙古典型草原优势植物植株高度、根长和植物碳氮含量降低, 这些性状的变化能够使植物个体小型化, 适口性降低, 表明植物通过逃避放牧的策略适应长期自由放牧的干扰; 在割草管理方式下, 优势种的高度和比叶面积有增加的趋势, 其中冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低; 围封和长期无干扰处理下植物的碳氮含量增加, 表明在干扰强度降低后, 植物通过功能性状的改变从资源获取策略向资源储藏策略转变。2)对优势种功能性状集合分析表明, 糙隐子草具有较低的植株高度和较高的比叶面积, 冷蒿具有较高的木质素含量和氮含量, 这些性状能够使两种植物被家畜采食量减少, 并保证其具有较强的再生能力, 这可能是糙隐子草和冷蒿耐牧的原因; 大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 说明大针茅是非常典型的竞争物种, 在干扰较低的条件下, 大针茅采取竞争策略对其他物种产生较大的竞争压力可能是其占优势的重要原因。
关键词: 植物功能性状 ;
草地利用方式 ;
典型草原 ;
优势种 ;
元素性状 Abstract Aims When the external environment changes, plants can change their own functional traits and adjust adaptation strategies in a timely manner. Therefore, plant functional traits can effectively reflect the response of plants to the change of grassland use. However, there are few studies on the effects of grassland use patterns from the perspective of plant functional traits in the Nei Mongol grassland. Therefore, from the perspective of functional traits, this paper reveals the adaptation strategies of plants after external disturbances, aiming to provide basic data support and scientific basis for the sustainable management of natural grasslands.
Methods In this study, four important dominant species of
Stipa grandis, Leymus chinensis, Cleistogenes squarrosa and
Artemisia frigida in the typical grasslands of Nei Mongol were selected as the research objects to explore the differences in the functional traits of dominant species under the influence of long-term free grazing, mowing, short-term enclosed and long-term reservation.
Important findings We found that: 1) Vegetation height, root length and carbon and nitrogen concentration of dominant plants in Nei Mongol typical grasslands are reduced under long-term grazing. Changes in these traits can miniaturize individual plants and reduce their palatability, indicating that plants may adopt avoiding grazing strategy to adjust to the interference of long-term free grazing. Under the mowing management, the vegetation height and specific leaf area of the dominant species tend to increase. Among them, the nitrogen content of
A. frigida is the most sensitive to the response of mowing. The nitrogen concentration in its roots, stems, leaves are the lowest in the mowing sites; the carbon and nitrogen concentration of plants increases under enclosure and long-term non-disturbance treatments, indicating that the plant transforms from resource acquisition strategy to resource storage strategy through changes in functional traits when the disturbance intensity is reduced. 2) The analysis of the assemblage of functional traits of dominant species shows that
C. squarrosa has a small plant height and a high specific leaf area, and
A. frigida has a high lignin and nitrogen concentration. The two species thus can adopt some avoidance (escaping from grazers) and tolerance (regrowth capacity after defoliation) strategies to improve their grazing resistance;
S. grandis has the highest plant height, the largest leaf dry matter concentration, the highest stem and leaf cellulose concentration, which indicates that
S. grandis is a very typical competitive species. Under management conditions with low interference intensity, the competitive pressure of
S. grandis (Competitive strategy) against other species may be an important reason for its high dominance.
Keywords: plant functional traits ;
grassland utilization patterns ;
typical grasslands ;
dominant species ;
element traits PDF (1260KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 张景慧, 王铮, 黄永梅, 陈慧颖, 李智勇, 梁存柱. 草地利用方式对温性典型草原优势种植物功能性状的影响. 植物生态学报, 2021, 45(8): 818-833. DOI:
10.17521/cjpe.2020.0373 ZHANG Jing-Hui, WANG Zheng, HUANG Yong-Mei, CHEN Hui-Ying, LI Zhi-Yong, LIANG Cun-Zhu.
Effects of grassland utilization on the functional traits of dominant plants in a temperate typical steppe .
Chinese Journal of Plant Ecology , 2021, 45(8): 818-833. DOI:
10.17521/cjpe.2020.0373 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 )。植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 )。在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 )。
植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 )。在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 )。其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 )。叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 )。植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 )。植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 )。
放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响。其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 )。此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 )。与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少。已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 )。
内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 )。内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 )。近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 )。尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱。鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律?
1 材料和方法 1.1 研究区概况 研究区位于内蒙古自治区锡林浩特市以东的毛登牧场(44.18° N, 116.37° E), 总面积为5.85 × 10
4 hm
2 。该区域地貌为波状高平原, 海拔910- 1 377 m。气候为典型的温带大陆性气候, 年平均气温为2.36 ℃, 最热月7月平均气温为21.2 ℃, 最冷月1月平均气温为-19.4 ℃, 年温差35-42 ℃, 日温差12-16 ℃; 年降水量281 mm, 由东南向西北逐渐递减, 降水量年际和季节变化较大, 主要集中在5-9月, 占全年总降水量的80%以上(1953-2013年)。实验期间研究区月降水量和月平均气温的变化如
图1 所示。研究区属于中国温带典型草原区, 地带性植被为大针茅、克氏针茅(
Stipa krylovii )和羊草草原。物种组成主要以禾本科、菊科、藜科、百合科和豆科等科的植物为主, 主要优势种有大针茅、克氏针茅和羊草, 常见物种有糙隐子草、冷蒿和冰草(
Agropyron cristatum )等。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图1内蒙古温带典型草原研究区实验期间月降水量和月平均气温的变化。 Fig. 1Variations of mean monthly air temperature and monthly precipitation of the study site area in temperate typical grasslands of Nei Mongol in 2011-2014. 1.2 样地设置 (1)长期自由放牧样地(G): 放牧是毛登牧场草地利用的主要方式, 我们在天然草场选取3个长期自由放牧样地, 样地放牧家畜包括牛和绵羊, 近几十年来放牧强度可折算为1.08只·hm
-2 绵羊(
陈慧颖等, 2014 ;
张景慧等, 2016 )。该放牧样地与居民点的距离大于15 km。
(2)割草样地(M): 从2008年毛登牧场选择2.73 × 10
4 hm
2 草原作为割草场, 割草场内每年8月进行1次打草且禁止放牧, 我们在割草场内选择3个割草样地, 2008年之前割草样地与长期自由放牧样地的利用方式和强度完全相同。
(3)短期围封样地(E): 短期围封样地以生态保护为主, 与割草样地同处一个区域, 2008年之前与长期自由放牧样地的利用方式和强度完全相同。2008年以来禁止放牧和割草。设置3个短期围封样地, 本试验开始在此取样时, 该地已围封4年。割草样地、短期围封样地和长期自由放牧样地彼此相临(最小距离不超过1 km)。
(4)长期无干扰样地(R): 在距离其他3个样地15 km左右的天然草地布设了3个长期无干扰样地。该片草原属于毛登牧场的草原生态保护区, 为以大针茅占绝对优势的大针茅草原。因为离居民点较远, 从1956年以来人为干扰一直较少。
上述供试样地共计12个(100 m × 100 m), 两样地间距均大于500 m (
Fischer et al ., 2010 ;
Socher et al ., 2013 )。在每个样地内随机选择3个小样区(10 m × 10 m), 用PVC管标定好边界范围。
1.3 试验方法 1.3.1 优势种的选择
本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象。选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )。为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象。
Table 1 表1 表1 测定植物功能性状前(2011年)各样地优势物种相对生物量(平均值±标准误)
Table 1
Relative biomass of dominant species in each sites before measuring plant functional traits (mean ± SE ) 物种 Species 相对生物量 Relative biomass p 长期自由放牧 割草 短期围封 长期无干扰 大针茅 Stipa grandis 9.89 ± 2.58b 7.20 ± 2.70b 5.84 ± 1.99b 77.80 ± 4.77a <0.001 羊草 Leymus chinensis 36.00 ± 6.88a 12.27 ± 2.63ab 29.21 ± 15.40a 2.56 ± 1.93b 0.004 糙隐子草 Cleistogenes squarrosa 32.55 ± 6.65a 35.13 ± 2.26a 21.39 ± 5.26a 3.53 ± 1.95b <0.001 冷蒿 Artemisia frigida 0.31 ± 0.31b 24.94 ± 8.24a 32.98 ± 11.02a 3.32 ± 1.65b 0.002
不同小写字母表示在0.05水平上具有显著差异。 Different lowercase letters mean significant difference at 0.05 level. 新窗口打开 |
下载CSV 1.3.2 植物功能性状测定
2012年8月中旬, 对每个样地中4种优势物种进行采样, 对每个物种在每个样地中选取没有遮挡的、长势良好的成熟植株10株(丛), 首先对植物高度(
VH )进行测量。然后挖取目标植物, 尽量保证根系的完整性以及与地上部分的连接性。将植物样品装入塑封袋尽快带入实验室, 观测分蘖数, 分解根、茎、叶、生殖部分, 在65 ℃条件下烘干至恒质量, 测定植株各构件部分的质量, 计算植株的根冠比(
R /
S )和茎叶比(
S /
L )。
于2013和2014年8月中旬, 在日出2-3 h后, 日落3-4 h前, 对样地中4种优势物种进行采样。在每个样地中, 每个物种选取健康、完整的成熟个体10株(丛), 迅速放入黑暗的容器内储藏, 保证容器湿润且内部温度<5 ℃, 回到室内将植株放入水中, 在5 ℃的黑暗环境中储藏超过6 h (
Cornelissen et al ., 2003 )。在水分达到饱和后, 每株(丛)植株选择完全伸展的、无病虫害的叶片2-4枚, 用吸水纸迅速吸去叶片表面的水分, 在1/1 000电子天平上称量饱和鲜质量。然后用叶面积仪(LI-3000, LI-COR, Lincoln, USA)测量叶面积。最后将叶片放入烘箱, 在65 ℃条件下烘干至恒质量, 测定叶片干质量。
SLA 和
LDMC 分别用以下公式计算:
SLA =叶片面积(cm
2 )/叶片干质量(g)
LDMC =叶片干质量(mg)/叶片饱和鲜质量(g)
于2011和2012年8月, 对每种草地利用方式下草原中的4个优势物种进行采样。每个物种挖取没有遮挡的、长势良好的成熟植株10株(丛), 汇合成为3份, 将植株分为根、茎、叶、生殖部分, 在65 ℃条件下烘干至恒质量。用旋风磨磨碎后, 采用全自动元素分析仪(CHN-600, LECO, Joseph, USA)测量每种植物各构件部分的总碳、总氮含量。本研究的元素功能性状包括根碳含量(
RCC ), 根氮含量(
RNC ); 茎碳含量(
SCC ), 茎氮含量(
SNC ); 叶碳含量(
LCC ), 叶氮含量(
LNC )。参照
AOAC (2000) 的操作标准, 使用
van Soest (1963) 的方法, 对2012年各个样地共有物种的根、茎、叶进行木质素和纤维素含量的测定。
1.4 统计分析 使用R 4.0.2和SPSS 19.0进行统计分析。采用比较均值中的单因素方差分析, 并进行0.05和0.01水平显著性检验, 分析优势种不同植物功能性状在利用方式间的差异, 以及物种之间性状的差异。用可塑性指数来表达各功能性状的可塑性对不同草地利用方式的响应程度, 参考
Valladares等(2000) 的方法来计算, 即G、M、E 3种样地中的某一性状的可塑性指数(
PI )为: R样地的数值减去G、M、E样地的数值, 再除以R样地的数值, 用环状条形图展示不同物种不同性状的可塑性。在性状×物种矩阵上进行主成分分析(PCA), 以分析物种内植物功能性状在物种间的差异, 并探索植物功能性状之间的相关性。用R 4.0.2和Origin 8.5进行绘图。
2 结果和分析 2.1 不同草地利用方式对4种植物形态功能性状的影响 4种植物形态功能性状对草地利用方式的响应敏感, 其中大针茅、羊草和糙隐子草的
SLA 在割草利用下增加, 而在放牧后降低(
p < 0.001)。冷蒿的响应不同, 其
SLA 在割草利用下最低, 而在长期无干扰条件下最高(
图2A )。草地利用方式对4种植物
LDMC 的影响没有一致性的规律。总体上, 放牧使大针茅、羊草和冷蒿的
VH 降低, 使羊草和冷蒿的根长(
RL )变短, 但对糙隐子草
VH 的影响规律相反。糙隐子草的
R /
S 在长期无干扰后显著增加而在放牧和短期围封后明显降低。冷蒿的
S /
L 在割草后增加, 说明割草后冷蒿地上部分中茎的比例增加明显(
图2 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2不同土地利用方式下4种植物形态植物功能性状(平均值±标准误)。E, 短期围封; G, 长期自由放牧; M, 割草; R, 长期无干扰。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 2Comparisons of morphological traits of different species under different grassland uses (mean ± SE ). E, short-term enclosed; G, long-term free grazing; M, mowing; R, long-term reservation. Different lowercase letters indicate significant differences among treatments (p < 0.05). 2.2 不同草地利用方式对4种植物元素功能性状的影响 放牧和割草对大针茅、糙隐子草和冷蒿
RCC 的影响明显, 其中放牧使大针茅和糙隐子草的
RCC 降低, 割草使糙隐子草和冷蒿的
RCC 降低; 相似地, 总体上4种优势物种的
RNC 在短期围封和长期无干扰条件下有明显增加的趋势(
图3 )。草地利用方式对羊草和糙隐子草茎中碳氮含量无显著影响, 但对冷蒿茎碳氮含量的影响较明显, 主要表现为放牧和长期无干扰使冷蒿
SCC 降低, 割草使冷蒿
SNC 降低。草地利用方式对大针茅、羊草和糙隐子草的影响比较相似, 长期无干扰条件下3种植物的
LCC 最高, 但3种植物的
LNC 在不同草地利用条件下无变化(
图3 )。冷蒿的
LCC 与
SCC 变化规律一致, 均在割草和短期围封样地较高, 而在长期放牧样地和长期无干扰样地较低(
图3 )。
图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3不同土地利用方式下4种植物碳氮含量(平均值±标准误)。E, 短期围封; G, 长期自由放牧; M, 割草; R, 长期无干扰。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 3Comparisons of carbon and nitrogen concentration of different species under different grassland uses (mean ± SE ). E, short-term enclosed; G, long-term free grazing; M, mowing; R, long-term reservation. Different lowercase letters indicate significant differences among treatments (p < 0.05). 2.3 不同草地利用方式对4种植物纤维素和木质素的影响 纤维素和木质素是重要的结构性碳水化合物, 不同物种的木质素和纤维素对草地利用方式的响应不同。如
图4A 所示, 草地利用方式对大针茅、羊草和糙隐子草的根纤维素含量(
RCE )无显著影响(
F 2,8 = 1.192,
p = 0.373;
F 2,8 = 1.484,
p = 0.291和
F 2,8 = 0.213,
p = 0.884), 长期自由放牧使冷蒿的
RCE 最高为30.17%。草地利用方式对大针茅和羊草的根木质素含量(
RLI )无影响(
F 2,8 = 1.222,
p = 0.363和
F 2,8 = 0.173,
p = 0.912,
图4B ), 糙隐子草和冷蒿的
RLI 则均在长期自由放牧样地较低。不同物种的茎纤维素含量(
SCE )对草地利用方式的响应不同, 大针茅的
SCE 在长期自由放牧利用方式最低, 羊草的
SCE 在长期无干扰样地中最低, 而糙隐子草的
SCE 在长期无干扰利用方式下最高, 显著高于长期自由放牧、割草和短期围封几种利用方式, 冷蒿的
SCE 则对草地利用方式响应不敏感(
F 2,8 = 2.221,
p = 0.163,
图4C )。4种主要物种茎木质素含量(
SLI )对草地利用方式的响应比较敏感, 总体上长期自由放牧使4种优势植物
SLI 降低(
图4D )。除糙隐子草外, 大针茅、羊草和冷蒿的叶纤维素含量(
LCE )对草地利用方式的响应均不敏感(
F 2,8 = 0.899,
p = 0.483;
F 2,8 = 1.243,
p = 0.357和
F 2,8 = 2.040,
p = 0.187), 糙隐子草的
LCE 在长期无干扰样地显著高于长期自由放牧样地、割草样地和短期围封样地(
p = 0.007、
p = 0.009和
p = 0.016,
图4E )。4种优势物种叶木质素含量(
LLI )在不同草地利用方式下差异比较明显, 放牧和割草使大针茅和羊草的
LLI 降低, 糙隐子草和冷蒿的
LLI 则在放牧条件下较低(
图4F )。
图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4不同土地利用方式下4种植物纤维素和木质素含量(平均值±标准误)。E, 短期围封; G, 长期自由放牧; M, 割草; R, 长期无干扰。不同小写字母表示处理间差异显著(p < 0.05)。 Fig. 4Comparisons of cellulose and lignin concentration of different species under different grassland uses (mean ± SE ). E, short-term enclosed; G, long-term free grazing; M, mowing; R, long-term reservation. Different lowercase letters indicate significant differences among treatments (p < 0.05). 2.4 4种优势植物不同性状指标可塑性变化的敏感度 以长期无干扰样地为参照系, 分析了各植物功能性状的可塑性指数,
图5 对放牧、割草和短期围封样地的4种研究对象的18种植物功能性状的可塑性指数大小进行了排序。对于大针茅来说, 在3个样地中的
R /
S 、
S /
L 等的可塑性幅度较大, 为对利用方式响应敏感的性状; 而
SCC 和
RL 的可塑性变化最不敏感, 为对利用方式响应不敏感的惰性性状。羊草的各功能性状在不同利用方式的样地中可塑性程度不同。羊草的
VH 、
SLA 、
SLI 和
RL 在放牧样地中为较敏感性状, 在割草样地中则表现为
VH 、
R /
S 、
RNC 和
LLI 较敏感, 在短期围封样地中
S /
L 为敏感性状; 而在放牧和割草样地中, 羊草的
LCE 的可塑性幅度较小, 短期围封样地中
RLI 为惰性性状。对于糙隐子草来说,
RLI 、
R /
S 、
SLA 和
RL 均为对利用方式响应敏感的性状, 其可塑性幅度较大; 而放牧和割草样地中的
LCC 和短期围封样地中的
RNC 可塑性变化不敏感。冰草的
S /
L 和
VH 在3种样地中的可塑性变化幅度较大, 对利用方式响应敏感; 而
RCC 、
SCC 和
LCC 的可塑性幅度较小, 是对利用方式响应不敏感的惰性性状。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5长期自由放牧(G)、割草(M)与短期围封(E)下大针茅(Stipa grandis )、羊草(Leymus chinensis )、糙隐子草(Cleistogenes squarrosa )、冷蒿(Artemisia frigida )功能性状可塑性指数(PI )变化程度。LCC, 叶碳含量; LCE, 叶纤维素含量; LDMC, 叶片干物质含量; LLI, 叶木质素含量; LNC, 叶氮含量; RCC, 根碳含量; RCE, 根纤维素含量; RL, 根长; RLI, 根木质素含量; RNC, 根氮含量; R /S, 根冠比; SCC, 茎碳含量; SCE, 茎纤维素含量; S /L, 茎叶比; SLA, 比叶面积; SLI, 茎木质素含量; SNC, 茎氮含量; VH, 植株高度。 Fig. 5Changes in the plasticity index (PI ) of functional traits of Stipa grandis, Leymus chinensis, Cleistogenes squarrosa and Artemisia frigida under long-term free grazing (G), mowing (M) and short-term enclosure (E) treatments. LCC, leaf carbon concentration; LCE, leaf cellulose concentration; LDMC, leaf dry matter concentration; LLI, leaf lignin concentration; LNC, leaf nitrogen concentration; RCC, root carbon concentration; RCE, root cellulose concentration; RL, root length concentration; RLI, root lignin concentration; RNC, root nitrogen concentration; R /S, root:shoot ratio; SCC, stem carbon concentration; SCE, stem cellulose concentration; S /L, stem:leaf ratio; SLA, specific leaf area; SLI, stem lignin concentration; SNC, stem nitrogen concentration; VH, vegetation height. 2.5 4种优势植物功能性状变异性统计学分析 在4种优势植物功能性状的PCA中, 前两个轴分别解释了37.89%和19.6%的方差。植物性状集合结果表明, 大针茅具有最高的
LDMC 、
VH 以及
S /
L, 而具有较低的
SLA 。糙隐子草的
SLA 和
R /
S 在4种植物中最大, 而
VH 最低。冷蒿的
LDMC 最低(
图6 ;
表2 )。在植物的碳氮含量中, 大针茅的
SNC 和
RNC 最低。羊草碳氮含量在4种植物中均为中等水平。糙隐子草具有最低的
SCC 和
LCC, 较高的
SNC 。冷蒿的
RCC 、
SCC 、
LCC 以及
RNC 、
LNC 均显著高于其他3种植物。大针茅具有最高的
SCE 和最低的
RCE 。糙隐子草的
LLI 最低。冷蒿根、茎、叶中的木质素含量在4种植物中均最高(
图6 ;
表2 )。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6大针茅(Stipa grandis )、羊草(Leymus chinensis )、糙隐子草(Cleistogenes squarrosa )、冷蒿(Artemisia frigida )功能性状的主成分分析(PCA)。LCC, 叶碳含量; LCE, 叶纤维素含量; LDMC, 叶片干物质含量; LLI, 叶木质素含量; LNC, 叶氮含量; RCC, 根碳含量; RCE, 根纤维素含量; RL, 根长; RLI , 根木质素含量; RNC, 根氮含量; R /S, 根冠比; SCC, 茎碳含量; SCE, 茎纤维素含量; S /L, 茎叶比; SLA, 比叶面积; SLI, 茎木质素含量; SNC, 茎氮含量; VH, 植株高度。 Fig. 6Principal component analysis (PCA) of functional traits of Stipa grandis, Leymus chinensis, Cleistogenes squarrosa and Artemisia frigida . LCC, leaf carbon concentration; LCE, leaf cellulose concentration; LDMC, leaf dry matter concentration; LLI, leaf lignin concentration; LNC, leaf nitrogen concentration; RCC, root carbon concentration; RCE, root cellulose concentration; RL, root length; RLI, root lignin concentration; RNC, root nitrogen concentration; R /S, root:shoot ratio; SCC, stem carbon concentration; SCE, stem cellulose concentration; S /L, stem:leaf ratio; SLA, specific leaf area; SLI, stem lignin concentration; SNC, stem nitrogen concentration; VH, vegetation height. Table 2 表2 表2 4种优势植物功能性状统计分析(平均值±标准误)
Table 2
Values of different functional traits of four dominant species (mean ± SE ) 植物功能性状 大针茅 Stipa grandis 羊草 Leymus chinensis 糙隐子草 Cleistogenes squarrosa 冷蒿 Artemisia frigida F p 比叶面积 SLA (m2 ·kg-1 ) 8.04 ± 0.23c 9.14 ± 0.41c 15.80 ± 0.83a 14.02 ± 0.40b 42.85 <0.001 叶片干物质含量 LDMC (mg·g-1 ) 498.11 ± 4.79a 437.91 ± 7.55b 446.91 ± 5.61b 337.67 ± 6.70c 115.03 <0.001 植株高度 VH (cm) 90.69 ± 6.57a 32.74 ± 3.08b 15.63 ± 0.83c 29.27 ± 2.59b 73.51 <0.001 根长 RL (cm) 15.36 ± 0.92ab 13.50 ± 1.47b 17.44 ± 1.00a 11.62 ± 1.85b 3.35 0.031 根冠比 R /S 1.74 ± 0.37b 0.72 ± 0.13b 3.40 ± 0.63a 0.98 ± 0.12b 10.32 <0.001 茎叶比 S /L 0.69 ± 0.41b 0.34 ± 0.06c 0.97 ± 0.16a 0.94 ± 0.16a 2.32 0.094 根碳含量 RCC (mg·g-1 ) 213.91 ± 18.41d 327.48 ± 15.84b 285.26 ± 10.01c 382.30 ± 9.58a 25.81 <0.001 茎碳含量 SCC (mg·g-1 ) 430.74 ± 6.61b 420.99 ± 4.37bc 409.31 ± 4.90c 452.30 ± 2.01a 14.63 <0.001 叶碳含量 LCC (mg·g-1 ) 443.07 ± 1.81ab 433.98 ± 3.34b 415.68 ± 8.34c 449.56 ± 2.72a 9.48 <0.001 根氮含量 RNC (mg·g-1 ) 5.43 ± 0.56b 5.95 ± 0.52ab 6.11 ± 0.29ab 6.95 ± 0.51a 1.70 0.190 茎氮含量 SNC (mg·g-1 ) 5.28 ± 0.45c 6.75 ± 0.51bc 10.87 ± 0.71a 6.99 ± 0.45b 19.50 <0.001 叶氮含量 LNC (mg·g-1 ) 14.21 ± 0.62c 17.61 ± 0.91b 16.05 ± 0.99bc 23.02 ± 0.88a 19.32 <0.001 根纤维素含量 RCE (%) 12.43 ± 1.20c 25.17 ± 0.96a 16.75 ± 1.22b 24.70 ± 2.01a 19.67 <0.001 茎纤维素含量 SCE (%) 34.17 ± 1.61a 31.25 ± 0.69ab 27.86 ± 0.52b 29.93 ± 2.30ab 2.30 0.096 叶纤维素含量 LCE (%) 25.31 ± 0.29a 25.76 ± 0.62a 25.94 ± 0.51a 19.74 ± 1.05b 18.41 <0.001 根木质素含量 RLI (%) 11.47 ± 1.38b 11.64 ± 0.64b 14.00 ± 1.38b 21.42 ± 1.62a 12.71 <0.001 茎木质素含量 SLI (%) 8.24 ± 0.45b 5.06 ± 0.58c 5.38 ± 0.70c 21.59 ± 2.54a 29.43 <0.001 叶木质素含量 LLI (%) 5.47 ± 0.28b 4.27 ± 0.29c 3.01 ± 0.26d 10.64 ± 1.17a 35.53 <0.001
不同小写字母表示在0.05水平上差异显著。 Different lowercase letters mean significant difference at the 0.05 level. LCC, leaf carbon concentration; LCE, leaf cellulose concentration; LDMC, leaf dry matter concentration; LLI, leaf lignin concentration; LNC, leaf nitrogen concentration; RCC, root carbon concentration; RCE, root cellulose concentration; RL, root length; RLI, root lignin concentration; RNC, root nitrogen concentration; R /S, root:shoot ratio; SCC, stem carbon concentration; SCE, stem cellulose concentration; S /L, stem:leaf ratio; SLA, specific leaf area; SLI, stem lignin concentration; SNC, stem nitrogen concentration; VH, vegetation height. 新窗口打开 |
下载CSV 3 讨论 3.1 植物功能性状对放牧的响应机制 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应。植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 )。从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 )。植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 )。此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似。在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因。
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 )。
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标。在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因。
从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 )。植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量。牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一。植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果。
此外, 放牧条件下除养分含量有下降的趋势外, 羊草的茎木质素以及糙隐子草、冷蒿的木质素含量在放牧条件下明显降低(
图4 )。植物碳氮、纤维素、木质素含量等决定着凋落物本身的性质, 直接影响着凋落物分解的快慢, 木质素含量低的植株死亡以后, 凋落物更易分解, 植物体在系统中的周转速率更加迅速(
Taylor et al ., 1989 )。本研究中, 优势物种碳氮含量、木质素含量等植物功能性状的降低, 将会使优势物种凋落物质量下降, 从而加速植物体在生态系统的周转速率, 使植物分解后的养分及时补充到生态系统中去, 补充植物在放牧生态系统中生存生长所需的养分。
3.2 割草和围封对植物功能性状的影响 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 )。导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长。割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 )。但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论。此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低。连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值。
围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累。同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小。我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 )。在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变。
3.3 从优势种植物功能性状对管理方式的响应探讨草原演替机制 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 )。这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律。首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )。从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 )。植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强。而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致。相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应。此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 )。总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 )。
其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段。而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 )。因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原。C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种)。在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势。竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 )。通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 )。此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致。通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势。羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定。
3.4 关键植物功能性状的选择 植物生态学理论的一个重要研究内容是理解植物对气候和土地利用变化的响应, 寻找关键的植物功能性状是解决这一问题的重要途径(
Pérez- Harguindeguy et al ., 2013 ), 其中两类植物功能性状得到生态学工作者的广泛关注, 其一是植物的形态特征, 另一类是与植物获取资源效率相关的功能性状(
Díaz et al ., 2004 )。植物的高度和根长与植株的表型最密切相关, 通过可塑性分析, 我们发现高度是一个可塑性较强的功能性状(
图5 ), 高度可以反映植物对光资源的获取能力, 与植物竞争能力相关(
Hodgson et al ., 1999 )。Díaz等(2007)对全球范围植物功能性状对放牧响应的综合分析发现, 植株高度是反映植物对放牧响应的有效植物功能性状。与植株高度相比, 大部分情况下根长的敏感性均低于植株高度(
图5 ), 鉴于地下根系采集的难度较大, 对植物破坏较严重且其属于惰性性状, 我们建议生态学工作者可以更关注植株高度。
在本研究中我们分器官测量了不同草地利用条件下优势物种的元素含量, 前人的研究表明植物的叶片性状对环境变化的反应更为敏感, 能够有效地反映植物对环境变化的适应状况(
Meziane & Shipley, 1999 )。其中
SLA 、
LNC 和
LDMC 一直备受关注(
Reich et al ., 1999 ;
Wilson et al ., 1999 )。在叶片水平,
SLA 和
LNC 与叶片的寿命有关, 二者结合可以准确预测许多植物的最大净光合速率(
Reich et al ., 1997 ), 随后
Ryser和Urbas (2000) 发现叶片干物质含量也具有相似的功能。但在我们的研究中, 我们发现叶片干物质含量的可塑性指数非常低, 属于惰性性状, 在很多情况下其对草地利用方式的响应并不敏感。与此同时, 我们发现
RNC 对草地利用方式的敏感性在很多情况下高于
SNC 和
LNC (
图5 )。这可能是因为根与土壤密切接触, 草地利用方式引起的环境中氮含量的改变及时反映到植物
RNC 中来。且有时不同器官氮含量对草地利用方式的响应不一致(例如冷蒿对放牧和短期围封的响应,
图5 ), 这体现了元素含量分配在器官中的权衡。因此在植物功能性状的研究中, 在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状。
4 结论 内蒙古典型草原优势植物通过降低植株高度、根长和植物碳氮含量来实现植物个体小型化和降低适口性, 通过采取逃避放牧的策略适应长期自由放牧的干扰。优势种的高度和
SLA 在割草处理下有增加的趋势, 冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低。围封和长期无干扰有助于植物体中碳氮营养物质的贮藏积累, 说明植物通过功能性状的改变从资源获取策略向资源储藏策略转变。糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧。大针茅具有最高的植株高度、最大的叶片干物质含量, 和最高的
SCE 、
LCE, 说明大针茅是非常典型的竞争物种, 在长期无干扰草原中占绝对优势地位。与植株高度相比, 根长测量困难且属于惰性性状, 建议更关注植株高度。
RNC 对草地利用方式响应敏感, 说明在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状。
[1] Ansquer P Duru M Theau JP Cruz P 2009 ). Functional traits as indicators of fodder provision over a short time scale in species-rich grasslandsAnnals of Botany 117 -126 . [本文引用: 1] [2] AOAC (2000 ). AOAC official method 973.18 fiber (acid detergent) and lignin (H2 SO4 ) in animal feed//Horwitz, Wed. Official Methods of Analysis of AOAC International . 17th edAssociation of Official Analytical Chemists, Gaithersburg, USA . [本文引用: 1] [3] Austin AT Ballaré CL 2010 ). Dual role of lignin in plant litter decomposition in terrestrial ecosystemsProceedings of the National Academy of Sciences of the United States of America 4618 -4622 . [本文引用: 1] [4] Bai YF Wu JG Pan QM Huang JH Wang QB Li FS Buyantuyev A Han XG 2007 ). Positive linear relationship between productivity and diversity: evidence from the Eurasian SteppeJournal of Applied Ecology 1023 -1034 . [本文引用: 2] [5] Bao YJ Cao M Li ZH Guo P Zhang J Qin J 2019 ). A comparative study of the response of Leymus chinensis and Stipa grandis root characteristics to moisture gradientsActa Ecologica Sinica 1063 -1070 . [本文引用: 1] [ 鲍雅静 , 曹明 , 李政海 , 郭鹏 , 张靖 , 秦洁 (2019 ). 羊草与大针茅根系构型对水分梯度响应的比较研究生态学报 , 39, 1063 -1070 .] [本文引用: 1] [6] Chen DM Zheng SX Shan YM Taube F Bai YF 2013 ). Vertebrate herbivore-induced changes in plants and soils: linkages to ecosystem functioning in a semi-arid steppeFunctional Ecology 273 -281 . [本文引用: 1] [7] Chen HY Zhang JH Huang YM Gong JR 2014 ). Traits related to carbon sequestration of common plant species in a Stipa grandis steppe in Nei Mongol under different land-usesChinese Journal of Plant Ecology 821 -832 . [本文引用: 3] [ 陈慧颖 , 张景慧 , 黄永梅 , 龚吉蕊 (2014 ). 内蒙古大针茅草原常见植物在不同土地利用方式下的固碳相关属性植物生态学报 , 38, 821 -832 .] [本文引用: 3] [8] Cingolani AM Posse G Collantes MB 2005 ). Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslandsJournal of Applied Ecology 4250 -4259 . [本文引用: 3] [9] Collins SL Knapp AK Briggs JM Blair JM Steinauer EM 1998 ). Modulation of diversity by grazing and mowing in native tallgrass prairieScience 745 -747 . [本文引用: 1] [10] Cornelissen JHC Diez PC Hunt R 1996 ). Seedling growth, allocation and leaf attributes in a wide range of woody plant species and typesJournal of Ecology 755 -765 . [本文引用: 2] [11] Cornelissen JHC Lavorel S Garnier E Díaz S Buchmann N Gurvich DE Reich PB Steege HT Morgan HD Heijden MGAVD Pausas JG Poorter H 2003 ). A handbook of protocols for standardised and easy measurement of plant functional traits worldwideAustralian Journal of Botany 335 -380 . [本文引用: 3] [12] Daorina , Song YT Wu yunna Huo GW Wang XM Xu ZC 2016 ). Response of plant leaf traits to grazing intensity in Stipa krylovii steppeChinese Journal of Applied Ecology 2231 -2238 . [本文引用: 1] [ 道日娜 , 宋彦涛 , 乌云娜 , 霍光伟 , 王晓朦 , 徐志超 (2016 ). 克氏针茅草原植物叶片性状对放牧强度的响应应用生态学报 , 27, 2231 -2238 .] [本文引用: 1] [13] Díaz S Cabido M 1997 ). Plant functional types and ecosystem function in relation to global changeJournal of Vegetation Science 463 -474 . [本文引用: 2] [14] Díaz S Hodgson JG Thompson K Cabido M Cornelissen JHC Jalili A Montserrat-Martí G Grime JP Zarrinkamar F Asri Y Band SR Basconcelo S Castro-Díez P Funes G Hamzehee B et al 2004 ). The plant traits that drive ecosystems: evidence from three continentsJournal of Vegetation Science 295 -304 . [本文引用: 1] [15] Díaz S Kattge J Cornelissen JHC Wright IJ Lavorel S Dray S Reu B Kleyer M Wirth C Prentice IC Garnier E Bönisch G Westoby M Poorter H Reich PB et al 2016 ). The global spectrum of plant form and functionNature 167 -171 . [本文引用: 1] [16] Díaz S Lavorel S McIntyre S Falczuk V Casanoves F Milchunas DG Skarpe C Rusch G Sternberg M Noy-Meir I Landsberg J Zhang W Clark H Campbell BD 2007 ). Plant trait responses to grazing—A global synthesisGlobal Change Biology 313 -341 . [本文引用: 1] [17] Díaz S Noy-Meir I Cabido M 2001 ). Can grazing response of herbaceous plants be predicted from simple vegetative traits?Journal of Applied Ecology 497 -508 . [本文引用: 1] [18] Fan YJ 2013 ). Effects of Fencing and Grazing on Plant Morphology, Community Characteristics and Carbon Balance of Kobresia pygmaea Meadow in the Three Headwater Resource Regions PhD dissertation, Gansu Agricultural University, Lanzhou . [本文引用: 1] [ 范月君 (2013 ). 围栏与放牧对三江源区高山嵩草草甸植物形态、群落特征及碳平衡的影响甘肃农业大学, 兰州 .] [本文引用: 1] [19] Fischer M Bossdorf O Gockel S Hänsel F Hemp A Hessenmöller D Korte G Nieschulze J Pfeiffer S Prati D Renner S Schöning I Schumacher U Wells K Buscot F et al 2010 ). Implementing large-scale and long-term functional biodiversity research: the biodiversity exploratoriesBasic and Applied Ecology 473 -485 . [本文引用: 1] [20] Frenette-Dussault C Shipley B Léger JF Meziane D Hingrat Y 2012 ). Functional structure of an arid steppe plant community reveals similarities with Grimeʼs C-S-R theoryJournal of Vegetation Science 208 -222 . [本文引用: 1] [21] Garnier E Cortez J Billès G Navas ML Roumet C Debussche M Laurent G Blanchard A Aubry D Bellmann A Neill C Toussaint JP 2004 ). Plant functional markers capture ecosystem properties during secondary successionEcology 2630 -2637 . [本文引用: 3] [22] Garnier E Laurent G Bellmann A Debain S Berthelier P Ducout B Roumet C Navas M-L 2001 ). Consistency of species ranking based on functional leaf traitsNew Phytologist 69 -83 . [本文引用: 1] [23] Garnier E Lavorel S Ansquer P Castro H Cruz P Dolezal J Eriksson O Fortunel C Freitas H Golodets C Grigulis K Jouany C Kazakou E Kigel J Kleyer M et al 2007 ). Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: a standardized methodology and lessons from an application to 11 European sitesAnnals of Botany 967 -985 . [本文引用: 3] [24] Grime JP 1974 ). Vegetation classification by reference to strategiesNature 26 -31 . [本文引用: 2] [25] Grime JP 1977 ). Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theoryThe American Naturalist 1169 -1194 . [本文引用: 1] [26] He NP Liu CC Tian M Li ML Yang H Yu GR Guo DL Smith MD Yu Q Hou JH 2018 ). Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functionsFunctional Ecology 10 -19 . [本文引用: 1] [27] He NP Liu CC Zhang JH Xu L Yu GR 2018 ). Perspectives and challenges in plant traits: from organs to communitiesActa Ecologica Sinica 6787 -6796 . [本文引用: 1] [ 何念鹏 , 刘聪聪 , 张佳慧 , 徐丽 , 于贵瑞 (2018 ). 植物性状研究的机遇与挑战: 从器官到群落生态学报 , 38, 6787 -6796 .] [本文引用: 1] [28] Hodgson JG Wilson PJ Hunt R Grime JP Thompson K 1999 ). Allocating C-S-R plant functional types: a soft approach to a hard problem Oikos , 85, 282 -294 . [本文引用: 2] [29] Hou DJ Lu SZ Wang J Guo K 2020 ). Response of aboveground plant functional traits of Leymus chinensis to litter accumulation in fenced grasslandsActa Ecologica Sinica 6522 -6531 . [本文引用: 1] [ 侯东杰 , 陆帅志 , 王静 , 郭柯 (2020 ). 典型草原围封后羊草地上功能性状对枯落物累积的响应生态学报 , 40, 6522 -6531 .] [本文引用: 1] [30] Hou LL Yan RR Zhang Y Xin XP 2020 ). Effects of grazing intensity on functional traits of Leymus chinensis in meadow steppeScientia Agricultura Sinica 2562 -2572 . [本文引用: 1] [ 侯路路 , 闫瑞瑞 , 张宇 , 辛晓平 (2020 ). 放牧强度对草甸草原羊草功能性状的影响中国农业科学 , 53, 2562 -2572 .] [本文引用: 1] [31] Ilmarinen K Mikola J Nissinen K Vestberg M 2009 ). Role of soil organisms in the maintenance of species-rich seminatural grasslands through mowingRestoration Ecology 78 -88 . [本文引用: 1] [32] Jin Y Wang CK 2015 ). Trade-offs between plant leaf hydraulic and economic traitsChinese Journal of Plant Ecology 1021 -1032 . [本文引用: 1] [ 金鹰 , 王传宽 (2015 ). 植物叶片水力与经济性状权衡关系的研究进展植物生态学报 , 39, 1021 -1032 .] [本文引用: 1] [33] Kattge J Diaz S Lavorel S Prentice C Leadley P Boenisch G Garnier E Westoby M Reich PB Wright IJ Cornelissen JHC Violle C Harrison SP van Bodegom PM Reichstein M et al 2011 ). TRY—A global database of plant traitsGlobal Change Biology 2905 -2935 . [本文引用: 1] [34] Lavorel S Grigulis K Lamarque P Colace MP Garden D Girel J Pellet G Douzet R 2011 ). Using plant functional traits to understand the landscape distribution of multiple ecosystem servicesJournal of Ecology 135 -147 . [本文引用: 2] [35] Li JH Li ZQ Wang G 2003 ). Effect of different grazing intensities on the nutrient contents of Artemisia frigida and Potentilla acaulis Acta Prataculturae Sinica 30 -35 . [本文引用: 1] [ 李金花 , 李镇清 , 王刚 (2003 ). 不同放牧强度对冷蒿和星毛委陵菜养分含量的影响草业学报 , 12, 30 -35 .] [本文引用: 1] [36] Li XL Hou XY Wu XH Sarula , Ji L Chen HJ Liu ZY Ding Y 2014 ). Plastic responses of stem and leaf functional traits in Leymus chinensis to long-term grazing in a meadow steppeChinese Journal of Plant Ecology 440 -451 . [本文引用: 1] [ 李西良 , 侯向阳 , 吴新宏 , 萨茹拉 , 纪磊 , 陈海军 , 刘志英 , 丁勇 (2014 ). 草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应植物生态学报 , 38, 440 -451 .] [本文引用: 1] [37] Li YH 1994 ). Research on the grazing degradation model of the main steppe rangelands in Inner Mongolia and some considerations for the establishment of a comput-erized rangeland monitoring systemActa Phytoecologica Sinica 68 -79 . [本文引用: 4] [ 李永宏 (1994 ). 内蒙古草原草场放牧退化模式研究及退化监测专家系统雏议植物生态学报 , 18, 68 -79 .] [本文引用: 4] [38] Liu WT Wei ZJ Lv SJ Wang TL Zhang S Hai S 2017 ). Effect of grazing on Stipa breviflora age dynamics and leaf traitsActa Prataculturae Sinica 63 -71 . [本文引用: 1] [ 刘文亭 , 卫智军 , 吕世杰 , 王天乐 , 张爽 , 海松 (2017 ). 放牧调控对短花针茅种群年龄及叶性状的影响草业学报 , 26, 63 -71 .] [本文引用: 1] [39] Liu XJ Ma KP 2015 ). Plant functional traits—Concepts, applications and future directionsScientia Sinica Vitae 325 -339 . [本文引用: 1] [ 刘晓娟 , 马克平 (2015 ). 植物功能性状研究进展中国科学: 生命科学 , 45, 325 -339 .] [本文引用: 1] [40] Liu ZL Wang W Hao DY Liang CZ 2002 ). Probes on the degeneration and recovery succession mechanisms of Inner Mongolia steppeJournal of Arid Land Resources and Environment 84 -91 . [本文引用: 4] [ 刘钟龄 , 王炜 , 郝敦元 , 梁存柱 (2002 ). 内蒙古草原退化与恢复演替机理的探讨干旱区资源与环境 , 16, 84 -91 .] [本文引用: 4] [41] Ma JJ Yao H Feng ZY Zhang SL 2012 ). Changes in plant functional groups and species diversity under three grassland using modes in typical grassland area of Inner Mongolia, ChinaChinese Journal of Plant Ecology 1 -9 . [本文引用: 1] [ 马建军 , 姚虹 , 冯朝阳 , 张树礼 (2012 ). 内蒙古典型草原区3种不同草地利用模式下植物功能群及其多样性的变化植物生态学报 , 36, 1 -9 .] [本文引用: 1] [42] Melillo JM Aber JD Muratore JF 1982 ). Nitrogen and lignin control of hardwood leaf litter decomposition dynamicsEcology 621 -626 . [本文引用: 1] [43] Meziane D Shipley B 1999 ). Interacting determinants of specific leaf area in 22 herbaceous species: effects of irradiance and nutrient availabilityPlant, Cell & Environment 447 -459 . [本文引用: 1] [44] Mooney KA Halitschke R Kessler A Agrawal AA 2010 ). Evolutionary trade-offs in plants mediate the strength of trophic cascadesScience 1642 -1644 . [本文引用: 1] [45] Odum EP 1969 ). The strategy of ecosystem developmentScience 262 -270 . [本文引用: 1] [46] Pérez-Harguindeguy N Díaz S Garnier E Jaureguiberry P Poorter L ter Steege H Cornelissen JHC 2013 ). New handbook for standardised measurement of plant functional traits worldwideAustralian Journal of Botany 167 -234 . [本文引用: 1] [47] Pérez-Harguindeguy N Díaz S Vendramini F Cornelissen JHC Gurvich DE Cabido M 2003 ). Leaf traits and herbivore selection in the field and in cafeteria experimentsAustral Ecology 642 -650 . [本文引用: 1] [48] Poorter H Garnier E 2007 ). Ecological significance of inherent variation in relative growth rate and its components //Pugnatre FI, Valladares F. Functional Plant Ecology. 2nd ed. CRC Press, London. [本文引用: 1] [49] Reich PB Ellsworth DS Walters MB Vose JM Gresham C Volin JC Bowman WD 1999 ). Generality of leaf trait relationships: a test across six biomesEcology 1955 -1969 . [本文引用: 1] [50] Reich PB Walters MB Ellsworth DS 1992 ). Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystemsEcological Monographs 365 -392 . [本文引用: 1] [51] Reich PB Walters MB Ellsworth DS 1997 ). From tropics to tundra: global convergence in plant functioningProceedings of the National Academy of Sciences of the United States of America 13730 -13734 . [本文引用: 3] [52] Ryser P Urbas P 2000 ). Ecological significance of leaf life span among Central European grass speciesOikos 41 -50 . [本文引用: 1] [53] Sabaté S Sala A Gracia CA 1995 ). Nutrient content in Quercus ilex canopies: seasonal and spatial variation within a catchmentPlant and Soil 297 -304 . [本文引用: 1] [54] Sage RF Wedin DA Li M 1999 ). The biogeography of C4 photosynthesis: patterns and controlling factors //Sage RF, Monson RK. C4 Plant Biology . Academic Press, San Diego, USA. 313 -373 . [本文引用: 1] [55] Saugier B 2001 ). Plant strategies, vegetation processes, and ecosystem propertiesPlant Science 813 . [本文引用: 1] [56] Saura-Mas S Lloret F 2007 ). Leaf and shoot water content and leaf dry matter content of Mediterranean woody species with different post-fire regenerative strategiesAnnals of Botany 545 -554 . [本文引用: 1] [57] Shi SB Li HM Wang XY Yue XG Xu WH Chen GC 2006 ). Comparative studies of photosynthetic characteristics in typical alpine plants of the Qinghai-Tibet PlateauJournal of Plant Ecology (Chinese Version )40 -46 . [本文引用: 1] [ 师生波 , 李惠梅 , 王学英 , 岳向国 , 徐文华 , 陈桂琛 (2006 ). 青藏高原几种典型高山植物的光合特性比较植物生态学报 , 30, 40 -46 .] [本文引用: 1] [58] Socher SA Prati D Boch S Müller J Baumbach H Gockel S Hemp A Schöening I Wells K Buscot F Kalko EKV Linsenmair KE Schulze ED Weisser WW Fischer M 2013 ). Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regionsBasic and Applied Ecology 126 -136 . [本文引用: 1] [59] Sun M Tian K Zhang Y Wang H Guan DX Yue HT 2017 ). Research on leaf functional traits and their environmental adaptationPlant Science Journal 940 -949 . [本文引用: 1] [ 孙梅 , 田昆 , 张贇 , 王行 , 管东旭 , 岳海涛 (2017 ). 植物叶片功能性状及其环境适应研究植物科学学报 , 35, 940 -949 .] [本文引用: 1] [60] Talbot JM Treseder KK 2012 ). Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationshipsEcology 345 -354 . [本文引用: 1] [61] Taylor BR Parsons WFJ Parkinson D 1989 ). Decomposition of Populus tremuloides leaf litter accelerated by addition of Alnuscrispa litterCanadian Journal of Forest Research 674 -679 . [本文引用: 1] [62] Tong C Wu J Yong S Yang J Yong W 2004 ). A landscape- scale assessment of steppe degradation in the Xilin River Basin, Inner Mongolia, ChinaJournal of Arid Environments 133 -149 . [本文引用: 1] [63] Valladares F Wright SJ Lasso E Kitajima K Pearcy RW 2000 ). Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforestEcology 1925 -1936 . [本文引用: 1] [64] van Soest PJ 1963 ). Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and ligninJournal of the Association of Official Analytical Chemists 829 -835 . [本文引用: 1] [65] Violle C Navas ML Vile D Kazakou E Fortunel C Hummel I Garnier E 2007 ). Let the concept of trait be functional!Oikos 882 -892 . [本文引用: 1] [66] Wan H Bai Y Schönbach P Gierus M Taube F 2011 ). Effects of grazing management system on plant community structure and functioning in a semiarid steppe: scaling from species to communityPlant and Soil 215 -226 . [本文引用: 1] [67] Wang W Liang CZ Liu ZL Hao DY 2000 ). Analysis of the palnt individual behaviour during the degradation and restoring succession in steppe communityActa Phytoecologica Sinica 268 -274 . [本文引用: 1] [ 王炜 , 梁存柱 , 刘钟龄 , 郝敦元 (2000 ). 草原群落退化与恢复演替中的植物个体行为分析植物生态学报 , 24, 268 -274 .] [本文引用: 1] [68] Wang W Liu ZL Hao DY Liang CZ 1996 ). Research on the restoring succession of the degenerated grassland in Inner Mongolia—I. Basic characteristics and driving force for restoration of the degenerated grasslandActa Phytoecologica Sinica 449 -459 . [本文引用: 6] [ 王炜 , 刘钟龄 , 郝敦元 , 梁存柱 (1996 ). 内蒙古草原退化群落恢复演替的研究——I. 退化草原的基本特征与恢复演替动力植物生态学报 , 20, 449 -459 .] [本文引用: 6] [69] Westoby M 1999 ). The LHS strategy scheme in relation to grazing and firePeople and Rangelands Building the Future . VI International Rangeland Congress, Townsville, Australia . [本文引用: 1] [70] Wilson PJ Thompson K Hodgson JG 1999 ). Specific leaf area and leaf dry matter content as alternative predictors of plant strategiesNew Phytologist 155 -162 . [本文引用: 1] [71] Wright IA Jones JR Parsons AJ 2001 ). Effects of grazing by sheep or cattle on sward structure and subsequent performance of weaned lambsGrass and Forage Science 138 -150 . [本文引用: 1] [72] Wright IJ Reich PB Westoby M Ackerly DD Baruch Z Bongers F Cavender-Bares J Chapin T Cornelissen JHC Diemer M Flexas J Garnier E Groom PK Gulias J Hikosaka K et al 2004 ). The worldwide leaf economics spectrumNature 821 -827 . [本文引用: 6] [73] Wu GL Du GZ Liu ZH Thirgood S 2009 ). Effect of fencing and grazing on a Kobresia -dominated meadow in the Qinghai-Tibetan PlateauPlant and Soil 115 -126 . [本文引用: 1] [74] Xing XQ Hao BT Qi LX Zhang L Baoyin TGT 2019 ). Effect of grazing on functional traits of Stipa krylovii Ecology and Environmental Sciences 57 -64 . [ 邢小青 , 郝匕台 , 齐丽雪 , 张璐 , 宝音陶格涛 (2019 ). 放牧对克氏针茅功能性状的影响生态环境学报 , 28, 57 -64 .] [75] Xu XY Cao JJ Yang L Yang SR Gong YF Li MT 2018 ). Effects of grazing and enclosure on foliar and soil stoichiometry of grassland on the Qinghai-Tibetan PlateauChinese Journal of Ecology 1349 -1355 . [本文引用: 1] [ 许雪贇 , 曹建军 , 杨淋 , 杨书荣 , 龚毅帆 , 李梦天 (2018 ). 放牧与围封对青藏高原草地土壤和植物叶片化学计量学特征的影响生态学杂志 , 37, 1349 -1355 .] [本文引用: 1] [76] Zhang F Yang Y Qiao JR Jia LX Zhao TQ Wang X Wang YT Zhao ML 2018 ). Effect of utilization on community functional diversity in Stipa grandis steppeGrassland and Prataculture 36 -43 . [本文引用: 3] [ 张峰 , 杨阳 , 乔荠瑢 , 贾丽欣 , 赵天启 , 王玺 , 王亚婷 , 赵萌莉 (2018 ). 利用方式对大针茅草原群落功能多样性的影响草原与草业 , 30, 36 -43 .] [本文引用: 3] [77] Zhang JH Huang YM Chen HY Gong JR Qi Y Li EG Wu XC 2018 ). Response of plant functional traits at species and community levels to grazing exclusion on Inner Mongolian steppe, ChinaRangeland Journal , 40, 179 -189 . [78] Zhang JH Huang YM Chen HY Yang HY 2016 ). Effect of disturbance removal on leaf functional traits of plants in the Inner Mongolia steppeActa Ecologica Sinica 5902 -5911 . [本文引用: 2] [ 张景慧 , 黄永梅 , 陈慧颖 , 杨涵越 (2016 ). 去除干扰对内蒙古典型草原植物叶片功能属性的影响生态学报 , 36, 5902 -5911 .] [本文引用: 2] [79] Zhao N Zhao XQ Zhao L Xu SX Zou XY 2016 ). Progress in researches of the response of plant functional traits to grazing disturbanceChinese Journal of Ecology 1916 -1926 . [本文引用: 2] [ 赵娜 , 赵新全 , 赵亮 , 徐世晓 , 邹小艳 (2016 ). 植物功能性状对放牧干扰的响应生态学杂志 , 35, 1916 -1926 .] [本文引用: 2] [80] Zhao W Chen SP Han XG Lin GH 2009 ). Effects of long-term grazing on the morphological and functional traits of Leymus chinensis in the semiarid grassland of Inner Mongolia, ChinaEcological Research 99 -108 . [本文引用: 2] [81] Zheng SX Lan ZC Li WH Shao RX Shan YM Wan HW Taube F Bai YF 2011 ). Differential responses of plant functional trait to grazing between two contrasting dominant C3 and C4 species in a typical steppe of Inner Mongolia, ChinaPlant and Soil , 340, 141 -155 . [本文引用: 2] [82] Zheng SX Ren HY Lan ZC Li WH Wang KB Bai YF 2010 ). Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: scaling from species to communityBiogeosciences 1117 -1132 . [本文引用: 4] [83] Zhu JT Zhang YJ Wang WF Yang X Chen N Shen RN Wang L Jiang L 2020 ). Species turnover drives grassland community to phylogenetic clustering over long-term grazing disturbanceJournal of Plant Ecology 157 -164 . [本文引用: 1] [84] Zirbel CR Bassett T Grman E Brudvig LA 2017 ). Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restorationJournal of Applied Ecology 1070 -1079 . [本文引用: 1] Functional traits as indicators of fodder provision over a short time scale in species-rich grasslands
1
2009
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
AOAC official method 973.18 fiber (acid detergent) and lignin (H
2 SO
4 ) in animal feed//Horwitz, Wed.
Official Methods of Analysis of AOAC International . 17th ed
1
2000
... 于2011和2012年8月, 对每种草地利用方式下草原中的4个优势物种进行采样.每个物种挖取没有遮挡的、长势良好的成熟植株10株(丛), 汇合成为3份, 将植株分为根、茎、叶、生殖部分, 在65 ℃条件下烘干至恒质量.用旋风磨磨碎后, 采用全自动元素分析仪(CHN-600, LECO, Joseph, USA)测量每种植物各构件部分的总碳、总氮含量.本研究的元素功能性状包括根碳含量(
RCC ), 根氮含量(
RNC ); 茎碳含量(
SCC ), 茎氮含量(
SNC ); 叶碳含量(
LCC ), 叶氮含量(
LNC ).参照
AOAC (2000) 的操作标准, 使用
van Soest (1963) 的方法, 对2012年各个样地共有物种的根、茎、叶进行木质素和纤维素含量的测定. ...
Dual role of lignin in plant litter decomposition in terrestrial ecosystems
1
2010
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
Positive linear relationship between productivity and diversity: evidence from the Eurasian Steppe
2
2007
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
羊草与大针茅根系构型对水分梯度响应的比较研究
1
2019
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
羊草与大针茅根系构型对水分梯度响应的比较研究
1
2019
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Vertebrate herbivore-induced changes in plants and soils: linkages to ecosystem functioning in a semi-arid steppe
1
2013
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
内蒙古大针茅草原常见植物在不同土地利用方式下的固碳相关属性
3
2014
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... (1)长期自由放牧样地(G): 放牧是毛登牧场草地利用的主要方式, 我们在天然草场选取3个长期自由放牧样地, 样地放牧家畜包括牛和绵羊, 近几十年来放牧强度可折算为1.08只·hm
-2 绵羊(
陈慧颖等, 2014 ;
张景慧等, 2016 ).该放牧样地与居民点的距离大于15 km. ...
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
内蒙古大针茅草原常见植物在不同土地利用方式下的固碳相关属性
3
2014
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... (1)长期自由放牧样地(G): 放牧是毛登牧场草地利用的主要方式, 我们在天然草场选取3个长期自由放牧样地, 样地放牧家畜包括牛和绵羊, 近几十年来放牧强度可折算为1.08只·hm
-2 绵羊(
陈慧颖等, 2014 ;
张景慧等, 2016 ).该放牧样地与居民点的距离大于15 km. ...
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands
3
2005
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
Modulation of diversity by grazing and mowing in native tallgrass prairie
1
1998
... 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 ).导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长.割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 ).但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论.此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低.连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值. ...
Seedling growth, allocation and leaf attributes in a wide range of woody plant species and types
2
1996
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
... 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
A handbook of protocols for standardised and easy measurement of plant functional traits worldwide
3
2003
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 于2013和2014年8月中旬, 在日出2-3 h后, 日落3-4 h前, 对样地中4种优势物种进行采样.在每个样地中, 每个物种选取健康、完整的成熟个体10株(丛), 迅速放入黑暗的容器内储藏, 保证容器湿润且内部温度<5 ℃, 回到室内将植株放入水中, 在5 ℃的黑暗环境中储藏超过6 h (
Cornelissen et al ., 2003 ).在水分达到饱和后, 每株(丛)植株选择完全伸展的、无病虫害的叶片2-4枚, 用吸水纸迅速吸去叶片表面的水分, 在1/1 000电子天平上称量饱和鲜质量.然后用叶面积仪(LI-3000, LI-COR, Lincoln, USA)测量叶面积.最后将叶片放入烘箱, 在65 ℃条件下烘干至恒质量, 测定叶片干质量.
SLA 和
LDMC 分别用以下公式计算: ...
克氏针茅草原植物叶片性状对放牧强度的响应
1
2016
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
克氏针茅草原植物叶片性状对放牧强度的响应
1
2016
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
Plant functional types and ecosystem function in relation to global change
2
1997
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
... ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
The plant traits that drive ecosystems: evidence from three continents
1
2004
... 植物生态学理论的一个重要研究内容是理解植物对气候和土地利用变化的响应, 寻找关键的植物功能性状是解决这一问题的重要途径(
Pérez- Harguindeguy et al ., 2013 ), 其中两类植物功能性状得到生态学工作者的广泛关注, 其一是植物的形态特征, 另一类是与植物获取资源效率相关的功能性状(
Díaz et al ., 2004 ).植物的高度和根长与植株的表型最密切相关, 通过可塑性分析, 我们发现高度是一个可塑性较强的功能性状(
图5 ), 高度可以反映植物对光资源的获取能力, 与植物竞争能力相关(
Hodgson et al ., 1999 ).Díaz等(2007)对全球范围植物功能性状对放牧响应的综合分析发现, 植株高度是反映植物对放牧响应的有效植物功能性状.与植株高度相比, 大部分情况下根长的敏感性均低于植株高度(
图5 ), 鉴于地下根系采集的难度较大, 对植物破坏较严重且其属于惰性性状, 我们建议生态学工作者可以更关注植株高度. ...
The global spectrum of plant form and function
1
2016
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
Plant trait responses to grazing—A global synthesis
1
2007
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
Can grazing response of herbaceous plants be predicted from simple vegetative traits?
1
2001
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
围栏与放牧对三江源区高山嵩草草甸植物形态、群落特征及碳平衡的影响
1
2013
... 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 ).导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长.割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 ).但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论.此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低.连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值. ...
围栏与放牧对三江源区高山嵩草草甸植物形态、群落特征及碳平衡的影响
1
2013
... 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 ).导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长.割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 ).但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论.此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低.连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值. ...
Implementing large-scale and long-term functional biodiversity research: the biodiversity exploratories
1
2010
... 上述供试样地共计12个(100 m × 100 m), 两样地间距均大于500 m (
Fischer et al ., 2010 ;
Socher et al ., 2013 ).在每个样地内随机选择3个小样区(10 m × 10 m), 用PVC管标定好边界范围. ...
Functional structure of an arid steppe plant community reveals similarities with Grimeʼs C-S-R theory
1
2012
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Plant functional markers capture ecosystem properties during secondary succession
3
2004
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
... 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
Consistency of species ranking based on functional leaf traits
1
2001
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
Assessing the effects of land-use change on plant traits, communities and ecosystem functioning in grasslands: a standardized methodology and lessons from an application to 11 European sites
3
2007
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Vegetation classification by reference to strategies
2
1974
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory
1
1977
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Variation in leaf anatomical traits from tropical to cold-temperate forests and linkage to ecosystem functions
1
2018
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
植物性状研究的机遇与挑战: 从器官到群落
1
2018
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
植物性状研究的机遇与挑战: 从器官到群落
1
2018
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
Allocating C-S-R plant functional types: a soft approach to a hard problem 2
1999
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 植物生态学理论的一个重要研究内容是理解植物对气候和土地利用变化的响应, 寻找关键的植物功能性状是解决这一问题的重要途径(
Pérez- Harguindeguy et al ., 2013 ), 其中两类植物功能性状得到生态学工作者的广泛关注, 其一是植物的形态特征, 另一类是与植物获取资源效率相关的功能性状(
Díaz et al ., 2004 ).植物的高度和根长与植株的表型最密切相关, 通过可塑性分析, 我们发现高度是一个可塑性较强的功能性状(
图5 ), 高度可以反映植物对光资源的获取能力, 与植物竞争能力相关(
Hodgson et al ., 1999 ).Díaz等(2007)对全球范围植物功能性状对放牧响应的综合分析发现, 植株高度是反映植物对放牧响应的有效植物功能性状.与植株高度相比, 大部分情况下根长的敏感性均低于植株高度(
图5 ), 鉴于地下根系采集的难度较大, 对植物破坏较严重且其属于惰性性状, 我们建议生态学工作者可以更关注植株高度. ...
典型草原围封后羊草地上功能性状对枯落物累积的响应
1
2020
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
典型草原围封后羊草地上功能性状对枯落物累积的响应
1
2020
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
放牧强度对草甸草原羊草功能性状的影响
1
2020
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
放牧强度对草甸草原羊草功能性状的影响
1
2020
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
Role of soil organisms in the maintenance of species-rich seminatural grasslands through mowing
1
2009
... 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 ).导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长.割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 ).但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论.此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低.连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值. ...
植物叶片水力与经济性状权衡关系的研究进展
1
2015
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
植物叶片水力与经济性状权衡关系的研究进展
1
2015
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
TRY—A global database of plant traits
1
2011
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
Using plant functional traits to understand the landscape distribution of multiple ecosystem services
2
2011
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
不同放牧强度对冷蒿和星毛委陵菜养分含量的影响
1
2003
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
不同放牧强度对冷蒿和星毛委陵菜养分含量的影响
1
2003
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应
1
2014
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
草甸草原羊草茎叶功能性状对长期过度放牧的可塑性响应
1
2014
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
内蒙古草原草场放牧退化模式研究及退化监测专家系统雏议
4
1994
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象.选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
内蒙古草原草场放牧退化模式研究及退化监测专家系统雏议
4
1994
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象.选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
放牧调控对短花针茅种群年龄及叶性状的影响
1
2017
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
放牧调控对短花针茅种群年龄及叶性状的影响
1
2017
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
植物功能性状研究进展
1
2015
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
植物功能性状研究进展
1
2015
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
内蒙古草原退化与恢复演替机理的探讨
4
2002
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象.选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
内蒙古草原退化与恢复演替机理的探讨
4
2002
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象.选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
内蒙古典型草原区3种不同草地利用模式下植物功能群及其多样性的变化
1
2012
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
内蒙古典型草原区3种不同草地利用模式下植物功能群及其多样性的变化
1
2012
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
Nitrogen and lignin control of hardwood leaf litter decomposition dynamics
1
1982
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
Interacting determinants of specific leaf area in 22 herbaceous species: effects of irradiance and nutrient availability
1
1999
... 在本研究中我们分器官测量了不同草地利用条件下优势物种的元素含量, 前人的研究表明植物的叶片性状对环境变化的反应更为敏感, 能够有效地反映植物对环境变化的适应状况(
Meziane & Shipley, 1999 ).其中
SLA 、
LNC 和
LDMC 一直备受关注(
Reich et al ., 1999 ;
Wilson et al ., 1999 ).在叶片水平,
SLA 和
LNC 与叶片的寿命有关, 二者结合可以准确预测许多植物的最大净光合速率(
Reich et al ., 1997 ), 随后
Ryser和Urbas (2000) 发现叶片干物质含量也具有相似的功能.但在我们的研究中, 我们发现叶片干物质含量的可塑性指数非常低, 属于惰性性状, 在很多情况下其对草地利用方式的响应并不敏感.与此同时, 我们发现
RNC 对草地利用方式的敏感性在很多情况下高于
SNC 和
LNC (
图5 ).这可能是因为根与土壤密切接触, 草地利用方式引起的环境中氮含量的改变及时反映到植物
RNC 中来.且有时不同器官氮含量对草地利用方式的响应不一致(例如冷蒿对放牧和短期围封的响应,
图5 ), 这体现了元素含量分配在器官中的权衡.因此在植物功能性状的研究中, 在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状. ...
Evolutionary trade-offs in plants mediate the strength of trophic cascades
1
2010
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
The strategy of ecosystem development
1
1969
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
New handbook for standardised measurement of plant functional traits worldwide
1
2013
... 植物生态学理论的一个重要研究内容是理解植物对气候和土地利用变化的响应, 寻找关键的植物功能性状是解决这一问题的重要途径(
Pérez- Harguindeguy et al ., 2013 ), 其中两类植物功能性状得到生态学工作者的广泛关注, 其一是植物的形态特征, 另一类是与植物获取资源效率相关的功能性状(
Díaz et al ., 2004 ).植物的高度和根长与植株的表型最密切相关, 通过可塑性分析, 我们发现高度是一个可塑性较强的功能性状(
图5 ), 高度可以反映植物对光资源的获取能力, 与植物竞争能力相关(
Hodgson et al ., 1999 ).Díaz等(2007)对全球范围植物功能性状对放牧响应的综合分析发现, 植株高度是反映植物对放牧响应的有效植物功能性状.与植株高度相比, 大部分情况下根长的敏感性均低于植株高度(
图5 ), 鉴于地下根系采集的难度较大, 对植物破坏较严重且其属于惰性性状, 我们建议生态学工作者可以更关注植株高度. ...
Leaf traits and herbivore selection in the field and in cafeteria experiments
1
2003
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
1
2007
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
Generality of leaf trait relationships: a test across six biomes
1
1999
... 在本研究中我们分器官测量了不同草地利用条件下优势物种的元素含量, 前人的研究表明植物的叶片性状对环境变化的反应更为敏感, 能够有效地反映植物对环境变化的适应状况(
Meziane & Shipley, 1999 ).其中
SLA 、
LNC 和
LDMC 一直备受关注(
Reich et al ., 1999 ;
Wilson et al ., 1999 ).在叶片水平,
SLA 和
LNC 与叶片的寿命有关, 二者结合可以准确预测许多植物的最大净光合速率(
Reich et al ., 1997 ), 随后
Ryser和Urbas (2000) 发现叶片干物质含量也具有相似的功能.但在我们的研究中, 我们发现叶片干物质含量的可塑性指数非常低, 属于惰性性状, 在很多情况下其对草地利用方式的响应并不敏感.与此同时, 我们发现
RNC 对草地利用方式的敏感性在很多情况下高于
SNC 和
LNC (
图5 ).这可能是因为根与土壤密切接触, 草地利用方式引起的环境中氮含量的改变及时反映到植物
RNC 中来.且有时不同器官氮含量对草地利用方式的响应不一致(例如冷蒿对放牧和短期围封的响应,
图5 ), 这体现了元素含量分配在器官中的权衡.因此在植物功能性状的研究中, 在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状. ...
Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems
1
1992
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
From tropics to tundra: global convergence in plant functioning
3
1997
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... 在本研究中我们分器官测量了不同草地利用条件下优势物种的元素含量, 前人的研究表明植物的叶片性状对环境变化的反应更为敏感, 能够有效地反映植物对环境变化的适应状况(
Meziane & Shipley, 1999 ).其中
SLA 、
LNC 和
LDMC 一直备受关注(
Reich et al ., 1999 ;
Wilson et al ., 1999 ).在叶片水平,
SLA 和
LNC 与叶片的寿命有关, 二者结合可以准确预测许多植物的最大净光合速率(
Reich et al ., 1997 ), 随后
Ryser和Urbas (2000) 发现叶片干物质含量也具有相似的功能.但在我们的研究中, 我们发现叶片干物质含量的可塑性指数非常低, 属于惰性性状, 在很多情况下其对草地利用方式的响应并不敏感.与此同时, 我们发现
RNC 对草地利用方式的敏感性在很多情况下高于
SNC 和
LNC (
图5 ).这可能是因为根与土壤密切接触, 草地利用方式引起的环境中氮含量的改变及时反映到植物
RNC 中来.且有时不同器官氮含量对草地利用方式的响应不一致(例如冷蒿对放牧和短期围封的响应,
图5 ), 这体现了元素含量分配在器官中的权衡.因此在植物功能性状的研究中, 在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状. ...
Ecological significance of leaf life span among Central European grass species
1
2000
... 在本研究中我们分器官测量了不同草地利用条件下优势物种的元素含量, 前人的研究表明植物的叶片性状对环境变化的反应更为敏感, 能够有效地反映植物对环境变化的适应状况(
Meziane & Shipley, 1999 ).其中
SLA 、
LNC 和
LDMC 一直备受关注(
Reich et al ., 1999 ;
Wilson et al ., 1999 ).在叶片水平,
SLA 和
LNC 与叶片的寿命有关, 二者结合可以准确预测许多植物的最大净光合速率(
Reich et al ., 1997 ), 随后
Ryser和Urbas (2000) 发现叶片干物质含量也具有相似的功能.但在我们的研究中, 我们发现叶片干物质含量的可塑性指数非常低, 属于惰性性状, 在很多情况下其对草地利用方式的响应并不敏感.与此同时, 我们发现
RNC 对草地利用方式的敏感性在很多情况下高于
SNC 和
LNC (
图5 ).这可能是因为根与土壤密切接触, 草地利用方式引起的环境中氮含量的改变及时反映到植物
RNC 中来.且有时不同器官氮含量对草地利用方式的响应不一致(例如冷蒿对放牧和短期围封的响应,
图5 ), 这体现了元素含量分配在器官中的权衡.因此在植物功能性状的研究中, 在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状. ...
Nutrient content in
Quercus ilex canopies: seasonal and spatial variation within a catchment
1
1995
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
1
1999
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
Plant strategies, vegetation processes, and ecosystem properties
1
2001
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Leaf and shoot water content and leaf dry matter content of Mediterranean woody species with different post-fire regenerative strategies
1
2007
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
青藏高原几种典型高山植物的光合特性比较
1
2006
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
青藏高原几种典型高山植物的光合特性比较
1
2006
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
Interacting effects of fertilization, mowing and grazing on plant species diversity of 1500 grasslands in Germany differ between regions
1
2013
... 上述供试样地共计12个(100 m × 100 m), 两样地间距均大于500 m (
Fischer et al ., 2010 ;
Socher et al ., 2013 ).在每个样地内随机选择3个小样区(10 m × 10 m), 用PVC管标定好边界范围. ...
植物叶片功能性状及其环境适应研究
1
2017
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
植物叶片功能性状及其环境适应研究
1
2017
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
Interactions among lignin, cellulose, and nitrogen drive litter chemistry-decay relationships
1
2012
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
Decomposition of
Populus tremuloides leaf litter accelerated by addition of
Alnuscrispa litter
1
1989
... 此外, 放牧条件下除养分含量有下降的趋势外, 羊草的茎木质素以及糙隐子草、冷蒿的木质素含量在放牧条件下明显降低(
图4 ).植物碳氮、纤维素、木质素含量等决定着凋落物本身的性质, 直接影响着凋落物分解的快慢, 木质素含量低的植株死亡以后, 凋落物更易分解, 植物体在系统中的周转速率更加迅速(
Taylor et al ., 1989 ).本研究中, 优势物种碳氮含量、木质素含量等植物功能性状的降低, 将会使优势物种凋落物质量下降, 从而加速植物体在生态系统的周转速率, 使植物分解后的养分及时补充到生态系统中去, 补充植物在放牧生态系统中生存生长所需的养分. ...
A landscape- scale assessment of steppe degradation in the Xilin River Basin, Inner Mongolia, China
1
2004
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
Plastic phenotypic response to light of 16 congeneric shrubs from a Panamanian rainforest
1
2000
... 使用R 4.0.2和SPSS 19.0进行统计分析.采用比较均值中的单因素方差分析, 并进行0.05和0.01水平显著性检验, 分析优势种不同植物功能性状在利用方式间的差异, 以及物种之间性状的差异.用可塑性指数来表达各功能性状的可塑性对不同草地利用方式的响应程度, 参考
Valladares等(2000) 的方法来计算, 即G、M、E 3种样地中的某一性状的可塑性指数(
PI )为: R样地的数值减去G、M、E样地的数值, 再除以R样地的数值, 用环状条形图展示不同物种不同性状的可塑性.在性状×物种矩阵上进行主成分分析(PCA), 以分析物种内植物功能性状在物种间的差异, 并探索植物功能性状之间的相关性.用R 4.0.2和Origin 8.5进行绘图. ...
Use of detergents in the analysis of fibrous feeds. II. A rapid method for the determination of fiber and lignin
1
1963
... 于2011和2012年8月, 对每种草地利用方式下草原中的4个优势物种进行采样.每个物种挖取没有遮挡的、长势良好的成熟植株10株(丛), 汇合成为3份, 将植株分为根、茎、叶、生殖部分, 在65 ℃条件下烘干至恒质量.用旋风磨磨碎后, 采用全自动元素分析仪(CHN-600, LECO, Joseph, USA)测量每种植物各构件部分的总碳、总氮含量.本研究的元素功能性状包括根碳含量(
RCC ), 根氮含量(
RNC ); 茎碳含量(
SCC ), 茎氮含量(
SNC ); 叶碳含量(
LCC ), 叶氮含量(
LNC ).参照
AOAC (2000) 的操作标准, 使用
van Soest (1963) 的方法, 对2012年各个样地共有物种的根、茎、叶进行木质素和纤维素含量的测定. ...
Let the concept of trait be functional!
1
2007
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
Effects of grazing management system on plant community structure and functioning in a semiarid steppe: scaling from species to community
1
2011
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
草原群落退化与恢复演替中的植物个体行为分析
1
2000
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
草原群落退化与恢复演替中的植物个体行为分析
1
2000
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
内蒙古草原退化群落恢复演替的研究——I. 退化草原的基本特征与恢复演替动力
6
1996
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象.选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
内蒙古草原退化群落恢复演替的研究——I. 退化草原的基本特征与恢复演替动力
6
1996
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 本研究我们选取大针茅、羊草、冷蒿和糙隐子草4种植物作为研究对象.选取的原因包括以下两点: (1)在本研究中, 大针茅、羊草、冷蒿和糙隐子草在各样地物种组成中占有重要的比例(
表1 ); (2)大针茅和羊草是温性典型草原分布最广泛的两个物种, 其中羊草为多年生根茎禾草, 大针茅为多年生丛生禾草, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(半灌木状多年生杂类草)和糙隐子草(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).为了厘清不同草地利用方式下草原演替规律, 我们选取这4个温性典型草原物种作为研究对象. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
The LHS strategy scheme in relation to grazing and fire
1
1999
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
Specific leaf area and leaf dry matter content as alternative predictors of plant strategies
1
1999
... 在本研究中我们分器官测量了不同草地利用条件下优势物种的元素含量, 前人的研究表明植物的叶片性状对环境变化的反应更为敏感, 能够有效地反映植物对环境变化的适应状况(
Meziane & Shipley, 1999 ).其中
SLA 、
LNC 和
LDMC 一直备受关注(
Reich et al ., 1999 ;
Wilson et al ., 1999 ).在叶片水平,
SLA 和
LNC 与叶片的寿命有关, 二者结合可以准确预测许多植物的最大净光合速率(
Reich et al ., 1997 ), 随后
Ryser和Urbas (2000) 发现叶片干物质含量也具有相似的功能.但在我们的研究中, 我们发现叶片干物质含量的可塑性指数非常低, 属于惰性性状, 在很多情况下其对草地利用方式的响应并不敏感.与此同时, 我们发现
RNC 对草地利用方式的敏感性在很多情况下高于
SNC 和
LNC (
图5 ).这可能是因为根与土壤密切接触, 草地利用方式引起的环境中氮含量的改变及时反映到植物
RNC 中来.且有时不同器官氮含量对草地利用方式的响应不一致(例如冷蒿对放牧和短期围封的响应,
图5 ), 这体现了元素含量分配在器官中的权衡.因此在植物功能性状的研究中, 在关注植物叶片的功能性状的同时, 还应关注植物其他器官的功能性状. ...
Effects of grazing by sheep or cattle on sward structure and subsequent performance of weaned lambs
1
2001
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
The worldwide leaf economics spectrum
6
2004
... 植物性状是指影响植物生存生长、繁殖和最终适合度的植物属性(
Violle et al ., 2007 ;
Mooney et al ., 2010 ), 其中, 能够反映生态系统功能的植物性状称为植物功能性状(
Cornelissen et al ., 2003 ;
Díaz et al ., 2016 ).植物功能性状决定植物生存、生长和繁殖, 既能够对生态系统过程产生强烈影响, 也能够反映生态系统对环境变化的响应过程(
Wright et al ., 2004 ;
刘晓娟和马克平, 2015 ).在气候、地形、土壤及干扰等环境因子改变后, 植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰, 因此, 植物功能性状具有很强的可塑性, 能够起到良好的生态指示作用(
Lavorel et al ., 2011 ;
孙梅等, 2017 ). ...
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Effect of fencing and grazing on a
Kobresia -dominated meadow in the Qinghai-Tibetan Plateau
1
2009
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
放牧对克氏针茅功能性状的影响
0
2019
放牧对克氏针茅功能性状的影响
0
2019
放牧与围封对青藏高原草地土壤和植物叶片化学计量学特征的影响
1
2018
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
放牧与围封对青藏高原草地土壤和植物叶片化学计量学特征的影响
1
2018
... 围封样地和长期无干扰样地中大针茅和羊草的
LCC 显著增加, 其中4种植物根系的氮含量, 以及糙隐子草和冷蒿的
RLI 均在围封和长期无干扰后有增加的趋势, 这与
许雪贇等(2018) 的研究结果有相同趋势, 说明围封有助于营养物质的贮藏积累.同样地,
Garnier等(2004) 对12个弃耕年限不同的样地植物功能性状和生态系统过程进行比较研究, 发现随着弃耕年限的增加, 植物从资源获取策略向资源储藏策略转变, 植物的
SLA 随着弃耕年限的增加而减小.我们的结果以及前人的研究说明植物功能性状能够决定“资源获取-储藏的权衡” (
Grime, 1974 ;
Wright et al ., 2004 ): 一般而言, 在养分状况好且存在干扰的系统, 植物通过增加
SLA, 降低叶片干物质含量来促进获取资源的效率, 反之, 在排除放牧或停止其他干扰之后, 植物和生态系统倾向于更有效地保护内部资源(
Garnier et al ., 2004 ,
2007 ), 这时, 植物将具有更厚更密实且C:N较高的叶片, 植物纤维化和木质化的程度较高, 寿命更长(
Lavorel et al ., 2011 ).在围封和长期无干扰的条件下, 植物功能性状倾向于向资源贮藏的策略转变. ...
利用方式对大针茅草原群落功能多样性的影响
3
2018
... 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 ).导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长.割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 ).但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论.此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低.连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
利用方式对大针茅草原群落功能多样性的影响
3
2018
... 本研究发现优势种的高度和
SLA 在割草样地有增加的趋势(
图2 ,
图3 ).导致这种结果的原因可能是: 1)禁止放牧为牧草休养生息提供了机会, 使植物可以充分生长.割草样地自2008年以来禁止放牧, 在没有家畜的干扰后, 植物从个体小型化中恢复过来(
范月君, 2013 ); 2)刈割会去除立枯和高大植物的遮挡, 改善光照环境, 促进植物的生长和更新(
Collins et al ., 1998 ).但
张峰等(2018) 认为刈割相较于围封会显著降低大针茅的植株高度, 这可能是由于植株的恢复需要一段时间, 而两个研究中植株恢复的时间不同, 因此得出不同的结论.此外, 我们还发现冷蒿的氮含量对割草响应最敏感, 其根、茎、叶中的氮含量均在割草样地最低.连续打草会造成生态系统中凋落物持续减少, 使草地中的营养元素流出, 除留茬部分和根系凋落物之外, 没有牲畜排泄物等物质对草地生态系统进行营养返还, 从而导致土壤养分降低, 进一步导致植物体中的氮含量降低(
Ilmarinen et al ., 2009 ), 冷蒿对土壤养分含量变化的响应可能更加敏感, 植物体内氮含量的下降会降低冷蒿的饲用价值. ...
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... 其次,
王炜等(1996) 的研究表明, 围封13年的冷蒿小禾草草原从冷蒿占优势阶段演替为羊草占优势阶段.而在本研究中我们发现在长期无干扰的条件下, 草原变为大针茅占绝对优势的群落(
表1 ).因此, 我们比较关注为何在干扰相对较小的条件下群落再次演替为大针茅或羊草占优势的草原.C-S-R模型可能是关于植物生态策略最经典的一个模型(
Grime, 1974 ,
1977 ;
Saugier, 2001 ), 该策略模型基于与压力和干扰相关的功能性状划分出3个主要的生态策略: C策略(竞争物种), S策略(耐压力物种)和R策略(杂草物种).在低压力和低干扰地区采取C策略的竞争物种往往占绝对优势.竞争物种(C策略)的特征是具有更大的枝条和根系, 更大的植株高度以及很高的凋落物量, 它们可以有效地利用资源(
Frenette-Dussault et al ., 2012 ).通过性状集合我们可以发现大针茅具有最高的植株高度、最大的叶片干物质含量, 以及最高的茎、叶纤维素含量, 这些性状说明大针茅是非常典型的竞争物种, 决定大针茅在生存中主要采取C策略, 可产生大量的生物量, 在长期无干扰或围封的区域中对其他物种产生很大的竞争压力, 快速替代其他物种使草原演替成为大针茅占绝对优势的草原(
Zhang et al ., 2018 ).此外, 从植物获取-储藏资源策略角度分析, 我们发现大针茅除具有最高的叶片干物质含量外还具有最低的
SLA 、
SNC 和
LNC, 在植物叶片经济型谱中(
Wright et al ., 2004 ), 这些性状决定大针茅在资源贮藏的一端, 属于典型的资源贮藏类物种,
Garnier等(2007) 发现, 随着弃耕年限的增加, 具有资源贮藏性状的植物多度增加, 这一总体情况与
Odum (1969) 预测的植物演替模式一致.通过对羊草与大针茅根系构型对水分梯度响应的比较研究,
鲍雅静等(2019) 发现干旱条件将导致大针茅种群在群落中的优势地位增加, 湿润条件下羊草种群则会占据更大的优势.羊草的性状集合与大针茅相近(
图6 ), 在长期无干扰或围封的条件下, 群落是否演替成为大针茅草原或羊草草原可能在很大程度上由生境的水分条件决定. ...
Response of plant functional traits at species and community levels to grazing exclusion on Inner Mongolian steppe, China
0
2018
去除干扰对内蒙古典型草原植物叶片功能属性的影响
2
2016
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... (1)长期自由放牧样地(G): 放牧是毛登牧场草地利用的主要方式, 我们在天然草场选取3个长期自由放牧样地, 样地放牧家畜包括牛和绵羊, 近几十年来放牧强度可折算为1.08只·hm
-2 绵羊(
陈慧颖等, 2014 ;
张景慧等, 2016 ).该放牧样地与居民点的距离大于15 km. ...
去除干扰对内蒙古典型草原植物叶片功能属性的影响
2
2016
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... (1)长期自由放牧样地(G): 放牧是毛登牧场草地利用的主要方式, 我们在天然草场选取3个长期自由放牧样地, 样地放牧家畜包括牛和绵羊, 近几十年来放牧强度可折算为1.08只·hm
-2 绵羊(
陈慧颖等, 2014 ;
张景慧等, 2016 ).该放牧样地与居民点的距离大于15 km. ...
植物功能性状对放牧干扰的响应
2
2016
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
植物功能性状对放牧干扰的响应
2
2016
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
Effects of long-term grazing on the morphological and functional traits of
Leymus chinensis in the semiarid grassland of Inner Mongolia, China
2
2009
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
... ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
Differential responses of plant functional trait to grazing between two contrasting dominant C
3 and C
4 species in a typical steppe of Inner Mongolia, China
2
2011
... 植物功能性状能够反映物种的生态策略并决定植物如何响应环境因素的变化(
Kattge et al ., 2011 ).这里我们试图从优势种功能性状对管理方式的响应探讨草原演替机制, 由于我们样地的放牧强度仅为1.08只·hm
-2 绵羊, 放牧强度较轻, 且围封年限与前人研究结果(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 )相比较短, 因此在讨论演替规律时, 我们主要讨论被广泛验证且经典的内蒙古草原群落演替规律.首先, 我们试图回答为何内蒙古高原中东部大针茅和羊草为优势种的草原发生退化后形成冷蒿+糙隐子草小禾草草原(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ).从植物功能性状的集合看, 糙隐子草的
SLA 最大而植株最为矮小(
图6 ;
表2 ).植株矮小能够使糙隐子草具有更大几率躲避家畜的啃食, 而其较高的
SLA 也决定其相对生长速率较高(
Wright et al ., 2004 ); 此外, 糙隐子草为C
4 植物, 与C
3 植物相比, 它们特殊的固定CO
2 机制使其在高辐射生境中具有更高的资源利用效率和光合速率(
Sage et al ., 1999 ;
Zheng et al ., 2011 ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
... ), 因此在被啃食后其叶片的再生能力较强.而在本研究中, 糙隐子草的高度在放牧后有增加的趋势, 其根长也未出现变短的趋势, 也就是其未出现小型化现象, 说明糙隐子草更加耐牧, 这一结果与
Zheng等(2011) 的结果一致.相似地, 冷蒿为多年生杂类草, 与其他3种多年生禾草属于不同的功能群, 通过对优势物种功能性状集合的分析, 我们发现冷蒿根、茎、叶中的木质素含量显著高于其他物种, 特别是冷蒿属于半灌木, 其根和茎高度木质化(
图6 ;
表2 ), 这能够保证冷蒿的根和茎免受或少受家畜的啃食, 从而达到逃避放牧的效应.此外, 我们还发现冷蒿的
RNC 和
LNC 在几种优势植物里最高(
图6 ;
表2 ), 植物体中的氮含量与植物的最大净光合速率密切相关(
Reich et al ., 1997 ), 氮含量越高, 植物的净光合速率也越高, 将会导致冷蒿的植物体被啃食后有较强的再生能力, 这可能是因为冷蒿属于多年生杂类草, 其在叶片经济型谱中比多年生禾草的位置更加偏向于资源获取的一端(
Wright et al ., 2004 ;
Zhang et al ., 2018 ).总之, 糙隐子草和冷蒿的功能性状决定其能够通过采取逃避放牧策略(低的植株高度或高的木质素含量减少被采食几率)和适应放牧策略(高
SLA 、氮含量和净光合速率保证高的再生能力)使其更加耐牧, 在持续放牧的条件下, 挤占大针茅和羊草等优势种的生态位, 使大针茅草原和羊草草原演替成为冷蒿+糙隐子草的小型禾草草原成为可能(
马建军等, 2012 ). ...
Effects of grazing on leaf traits and ecosystem functioning in Inner Mongolia grasslands: scaling from species to community
4
2010
... 放牧、割草和围封等草地利用方式能够对植物功能性状产生明显影响.其中放牧对植物功能性状的研究比较充分(
Díaz et al ., 2007 ), 研究表明对放牧适应的物种在放牧后
SLA 和叶氮含量(
LNC )增加而
LDMC 降低, 这些性状的改变说明植物在放牧后生长速度加快, 适口性变好, 家畜与植物表现出正反馈; 而对放牧逃避的植物则通过降低植株高度和叶面积来减少家畜的单口采食量, 增加
LDMC 和降低植物营养含量来降低适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ).此外, 放牧强度能够显著改变羊草(
Leymus chinensis )和短花针茅(
Stipa breviflora )等草原优势种的叶片性状(
刘文亭等, 2017 ;
侯路路等, 2020 ), 影响植物养分利用策略(
道日娜等, 2016 ).与放牧相比, 割草、围封等草地利用方式对植物功能性状影响的研究相对较少.已有的研究发现, 由于常年缺乏动物排泄物补给, 割草区域植物叶片氮含量降低, 与固碳功能相关的植物功能性状对割草和围封响应敏感(
陈慧颖等, 2014 ;
张景慧等, 2016 ); 围封后由于枯落物的积累, 典型草原羊草的单株质量、茎叶比、叶面积、叶片长度、叶片质量和茎干质量等功能性状显著增加(
侯东杰等, 2020 ). ...
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
... 在本研究中, 我们使用了基于性状的方法来检验温带草原优势物种对家畜放牧的响应.植物形态特征的变化是其适应环境条件变化的重要策略(
师生波等, 2006 ).从植物的形态性状来看, 高度和根长对放牧的响应比较一致, 除糙隐子草外, 其他3种优势植物总体呈现高度变矮、根长变短的趋势(
图2 ), 这在一定程度上体现了植物通过改变自身功能性状来实现植物个体小型化(
Westoby, 1999 ;
王炜等, 2000 ), 除本研究涉及的性状外, 植物个体小型化的其他特征还包括叶面积变小、分蘖数减少、茎节间变短等(
Zhao et al ., 2009 ;
李西良等, 2014 ).植物个体的小型化会使家畜单口采食量降低, 进而增加家畜的采食难度, 从而使物种在一定程度上达到了逃避放牧的效果(
Zheng et al ., 2010 ), 如果放牧家畜为牛, 这种逃避放牧的效果将会更加明显, 因为牛通过舌头卷食植物, 对高度小于12 cm的植物无法达到很好的采食效率(
Wright et al ., 2001 ).此外, 我们发现在长期放牧处理下优势种
SLA 通常较低(
图2 ), 这与邢小青等(2019)的研究结果相似.在资源相对贫瘠的生境中, 植物通常具有较低的
SLA (
Poorter & Garnier, 2007 ), 常年的连续放牧使资源不间断地从系统中流失, 使该区域资源相对贫瘠可能是植物
SLA 降低的重要原因.
SLA 与生态系统功能具有直接关系(
Reich et al ., 1992 ;
Garnier et al ., 2001 ), 与植物的叶片净光合速率和相对生长速率正相关(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ).
SLA 降低说明植物的相对生长速率和净光合速率降低(
Cornelissen et al ., 1996 ;
Díaz & Cabido, 1997 ),
Zhao等(2009) 发现放牧显著降低羊草的净光合速率、表观量子效率、PSII光化学效率和水分利用效率等生理指标.在本研究中,
SLA 降低引起植物相对生长速率的降低可能是植物出现小型化的重要原因. ...
... 从植物养分含量的角度, 我们发现在长期放牧条件下大针茅根和茎, 冷蒿茎和叶中的碳含量明显降低, 4种植物的
RNC 相对较低(
图3 ).植物碳含量是衡量植物碳密度的一种有效度量指标(
陈慧颖等, 2014 ),
Saura-Mas和Lloret (2007) 发现受到破坏的植物体新产生的体细胞壁较薄, 会降低植物的碳含量.牲畜连续不间断的啃食和践踏会破坏植物体的原有组织, 使植物产生新的植物体, 这可能是大针茅和冷蒿碳含量降低的原因之一.植物碳氮含量的降低在一定程度上反映了植物营养成分的降低, 这会影响植物的适口性(
Cingolani et al ., 2005 ;
Zheng et al ., 2010 ;
赵娜等, 2016 ), 从而使植物进一步产生逃避放牧的效果. ...
Species turnover drives grassland community to phylogenetic clustering over long-term grazing disturbance
1
2020
... 内蒙古典型草原是欧亚温带草原的代表类型(
Bai et al ., 2007 ), 放牧、打草是该区域的主要利用方式, 由于长期过度利用导致典型草原生产力下降(
Tong et al ., 2004 ), 改变了草原生态系统的结构和功能(
Zhu et al ., 2020 ).内蒙古草原在不同草地利用方式下能够呈现较有规律的群落演替, 例如, 羊草和大针茅(
Stipa grandis )是温性典型草原分布最广泛的两个物种, 二者占优势的群落在被过度利用后的演替具有一定的趋同性, 即都会退化为以冷蒿(
Artemisia frigida )(半灌木状多年生杂类草)和糙隐子草(
Cleistogenes squarrosa )(多年生丛生小禾草)占优势的群落(
李永宏, 1994 ;
王炜等, 1996 ;
刘钟龄等, 2002 ), 相反地, 围封13年的冷蒿小禾草草原又从冷蒿占优势阶段演替为羊草占优势阶段(
王炜等, 1996 ).近年来, 许多研究者从生产力和生物多样性的角度研究草地利用方式对温带典型草原结构和功能的影响(
Bai et al ., 2007 ;
Wu et al ., 2009 ;
Wan et al ., 2011 ), 相较而言, 从植物功能性状角度开展草地利用方式影响的研究略少, 且已有的研究多集中在放牧对植物功能性状影响方面(
Zheng et al ., 2010 ;
Chen et al ., 2013 ).尽管如此, 放牧、割草和围封等草地利用方式对植物功能性状的影响仍需进一步较为系统地研究, 针对植物碳氮含量以及木质素和纤维素含量等元素计量特征的研究需要加强, 尤其从植物功能性状角度系统地探讨不同草地利用方式下草原演替规律的研究略显薄弱.鉴于此, 本研究以内蒙古锡林郭勒典型草原为例, 选取4个优势种, 测定其形态特征、碳氮含量以及木质素和纤维素含量等18种植物功能性状, 在植物个体水平分析植物功能性状对自由放牧、割草、短期围封和长期无干扰4种草地利用方式的响应, 以期解决以下两个科学问题: (1)草原优势物种如何通过改变自身功能性状适应不同草地利用方式的干扰? (2)是否能够通过草原优势物种功能性状对草地利用方式的响应揭示内蒙古草原群落演替规律? ...
Plant functional traits and environmental conditions shape community assembly and ecosystem functioning during restoration
1
2017
... 植物功能性状在个体水平上可以揭示群落甚至更大尺度上的生态学问题, 对于进一步研究植物群落和生态系统功能具有重要的意义(
Garnier et al ., 2004 ,
2007 ).在诸多植物功能性状中, 影响植物生长、繁殖、存活及资源利用的植物功能性状备受关注(
He et al ., 2018 ).其中, 植株的高度是反映物种竞争力的一个重要功能性状(
Díaz et al ., 2001 ), 高大的植物可以获取更多的光照, 在群落中具有很强的竞争能力(
Hodgson et al ., 1999 ).叶片性状不仅与植物生产力和资源利用密切相关(
Zirbel et al ., 2017 ), 也是实现植物性状由器官到群落推导的关键性状(
何念鹏等, 2018 ), 在一定程度上决定了叶片的生理活性, 能够反映植物叶片对所吸收光能的利用水平(
金鹰和王传宽, 2015 ), 其中, 比叶面积(
SLA )与植物光合作用的净同化速率密切相关,
SLA 大的植物往往相对生长速率更高(
Reich et al ., 1997 ;
Wright et al ., 2004 ); 叶片干物质含量(
LDMC )是对植物叶片组织密度的估计,
LDMC 高的植物通常具有较高的资源储存能力(
Cornelissen et al ., 2003 ;
Ansquer et al ., 2009 ).植物体中化学元素的循环是能量流动和物质循环的重要组成部分, 其中碳氮比被认为会影响植物体的循环速率, 进而影响整个生态系统的生物地球化学循环(
李金花等, 2003 ), 纤维素和木质素是重要的结构性碳水化合物, 与凋落物的分解有密切关系(
Melillo et al ., 1982 ;
Talbot & Treseder, 2012 ), 进而对生态系统的碳循环过程产生重要影响(
Austin & Ballaré, 2010 ).植物氮含量可以很好地反映土壤的肥力状况(
Sabaté et al ., 1995 ), 是植物营养价值和喜食性的指示指标(
Pérez-Harguindeguy et al ., 2003 ;
Cingolani et al ., 2005 ). ...
 ,4,*, 陈慧颖5, 李智勇1,2,3, 梁存柱1,2,3
,4,*, 陈慧颖5, 李智勇1,2,3, 梁存柱1,2,3

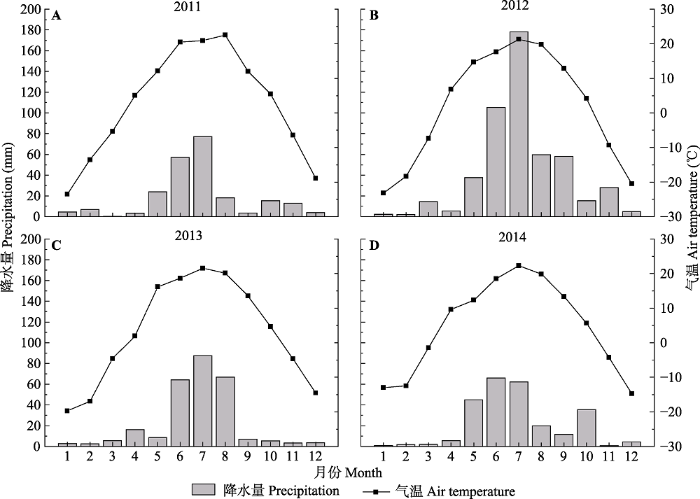 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT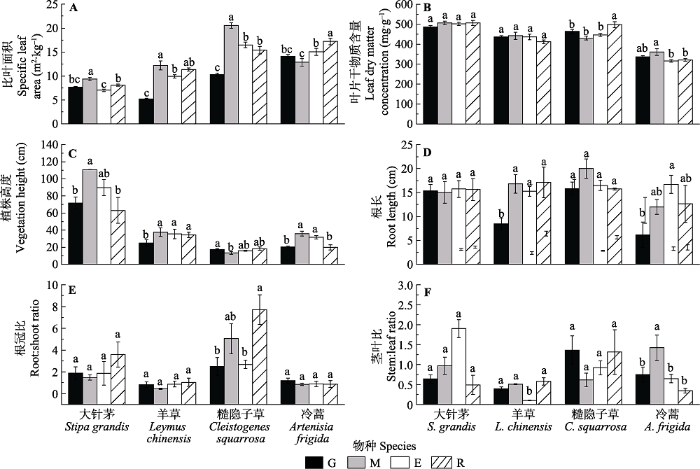 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT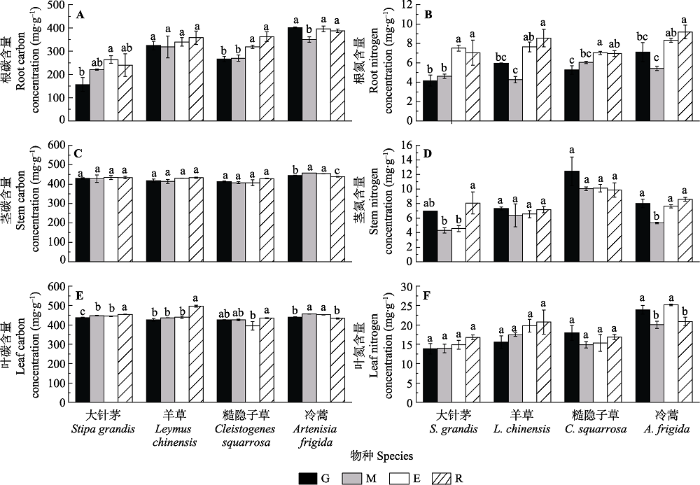 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT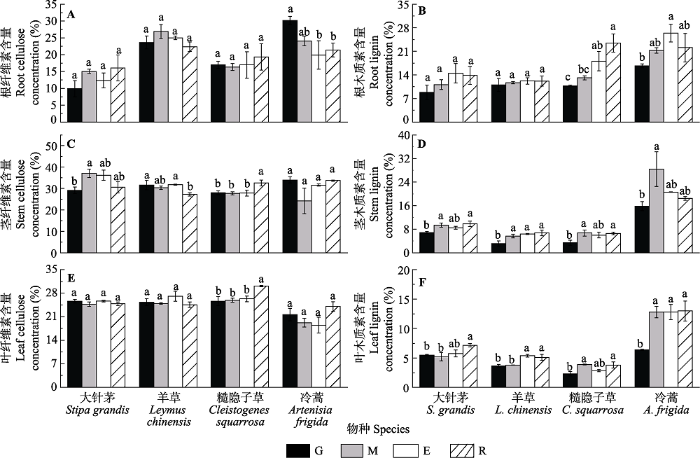 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT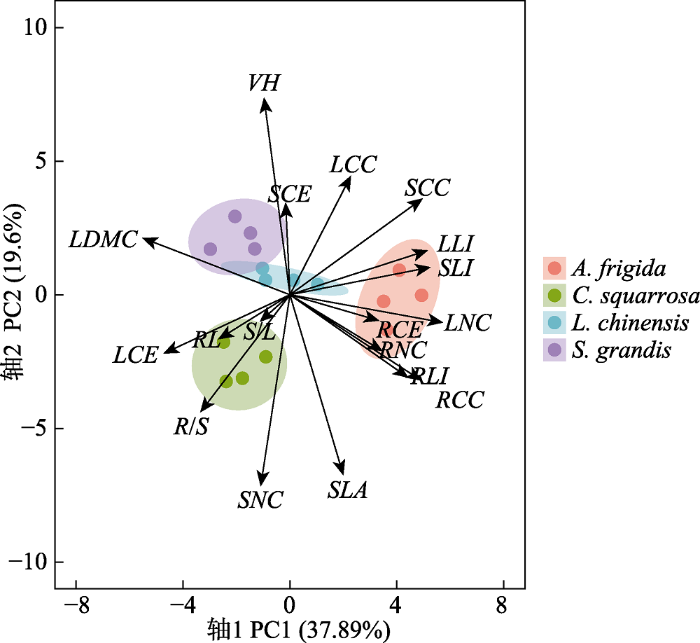 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}