Influencing factors of soil nitrous oxide emission during freeze-thaw cycles
De-Cai GAO,, E BAI,*Key Laboratory of Geographical Processes and Ecological Security of Changbai Mountains, Ministry of Education, School of Geographical Sciences, Northeast Normal University, Changchun 130024, China
National Key R&D Program of China(2019YFA0607301) National Natural Science Foundation of China(31901157) National Natural Science Foundation of China(41971058) China Postdoctoral Science Foundation(2020T130088) Natural Science Foundation of Jilin Province, China(YDZJ202101ZYTS104) Natural Science Foundation of Jilin Province, China(20180520087JH)
Abstract Aims Enhanced duration, intensity, and frequency of freeze-thaw cycles owing to global climate change may significantly affect soil nitrous oxide (N2O) emission. N2O is an important greenhouse gas, but our current understanding of soil N2O emission and its influencing factors during freeze-thaw cycles is still limited. Methods Here, we adopted the meta-analysis method and collected 30 articles on the effects of freeze-thaw cycles on soil N2O flux and cumulative emission from peer-reviewed journal articles. Our objectives were to explore the effects of freeze-thaw cycles on N2O emissions in different ecosystems and to comprehensively explore the influencing factors from the perspectives of experimental settings, soil physical and chemical properties, and the patterns of freeze-thaw cycles. Important findings Results showed that freeze-thaw cycles significantly increased N2O instantaneous emission, cumulative emission, and nitrification by 72.34%, 143.25%, and 124.63%, respectively. Freeze-thaw cycles also increased denitrification by 162.56%. Conversely, freeze-thaw cycles significantly decreased microbial biomass nitrogen by 6.39%. The effect of freeze-thaw cycles on N2O emission was significantly affected by the variations in soil microclimate and soil physical and chemical properties in different ecosystems. When the mean annual temperature (MAT) of a site exceeded 5 °C, freeze-thaw cycles could significantly enhance the N2O flux by 104.13%, which was significantly higher than that the effect at sites with MAT between 0-5 °C (25.56%) or less than 0 °C (55.29%). When soil moisture was greater than 70%, the increase of soil N2O flux caused by freeze-thaw cycles was 109.17%, which was significantly higher than that when soil moisture was between 50%-70% (65.67%) or less than 50% (20.37%). The higher soil clay and nutrient contents were, the greater the increase in N2O emission caused by freeze-thaw cycles became. Freeze-thaw cycles could significantly increase soil N2O flux by 91.21% in the presence of plants, which was higher than the effect in the absence of plants (54.43%). The impact of freeze-thaw cycles on N2O emission could be enhanced by soil sieving. In addition, soils sampled during the freeze-thaw cycling period showed more responses to freeze-thaw cycles than soils sampled during other times. The response of cumulative N2O emissions to freeze-thaw cycles was significantly improved by longer duration of thawing, higher intensity of freezing, and higher frequency of freeze-thaw cycles. When the freezing temperature was lower than -10 °C, freeze-thaw cycles could enhance soil N2O flux by 100.73%, which was significantly higher than the effect when the freezing temperature was between -10- -5 °C (47.74%) or more than -5 °C (70.25%). The main reason was that higher intensity of freezing could promote the release of more nutrients from soil microorganisms and soil structure, thereby increasing the production and emission of N2O. Overall, these results can help better understand the response of soil N2O to freeze-thaw cycles and its influencing factors, and provide scientific data for accurately predicting the impact of global climate change on N2O emission in the future. Keywords:freeze-thaw;nitrous oxide (N2O);microbial biomass;global climate change;denitrification
Fig. 1Effects of freeze-thaw cycles on soil nitrous oxide (N2O) flux, cumulative N2O emissions, microbial biomass nitrogen (MBN) content, nitrification, and denitrification rates. Error bars stand for 95% confidence intervals. Values next to the error bars represent the number of observations of the variable. Dashed line indicates that there is no effect.
Fig. 3Effects of freeze-thaw cycles on soil nitrous oxide (N2O) flux and cumulative N2O emissions. The variables are categorized into different groups according to mean annual temperature, experimental method, ecosystem type, soil treatment, presence of plant, period of sampling soil, temperature of freezing, and soil moisture. Error bars stand for 95% confidence intervals. The values next to the error bars represent the number of observations of the variable. Dashed line indicates that there is no effect. FT represents the freeze-thaw period.
2.3 土壤基本理化性质、硝化和反硝化作用速率及冻融循环格局与N2O排放的相关性
根据冻融循环对N2O通量的效应值与MAP和MAT之间的相关关系得知, 冻融循环对N2O通量的效应值与MAT之间呈现显著正相关关系(r = 0.68, p < 0.001), 而与MAP之间无相关性(图4)。
图4
新窗口打开|下载原图ZIP|生成PPT 图4冻融循环对土壤氧化亚氮(N2O)通量的效应值(ln R (N2O flux))与年降水量(MAP)(A)和年平均气温(MAT)(B)之间的相关关系。
Fig. 4Relationships between the effect size of freeze-thaw cycles on soil nitrous oxide (N2O) flux (ln R (N2O flux)) and mean annual precipitation (MAP)(A) and mean annual temperature (MAT)(B).
通过冻融循环对N2O通量的效应值与土壤基本理化性质之间的相关关系可得: 冻融循环对N2O通量的效应值与土壤质地有一定相关性, 比如与土壤黏粒含量呈正相关关系(r = 0.27, p < 0.001), 与土壤砂粒含量呈负相关关系(r = -0.22, p < 0.001), 而与土壤粉粒含量无显著相关关系(图5; 附件I)。在土壤养分中, 冻融循环对N2O通量的效应值只与土壤硝态氮含量呈现显著正相关关系(r = 0.42, p < 0.001), 而与土壤pH、有机碳含量、总碳含量、总氮含量、碳氮比、铵态氮含量、可溶性有机碳含量、可溶性有机碳与硝态氮含量比值无显著相关性。另外, 冻融循环对N2O通量的效应值也与土壤湿度呈现极好的线性正相关关系(r = 0.63, p < 0.001)。
图5
新窗口打开|下载原图ZIP|生成PPT 图5冻融循环对土壤氧化亚氮(N2O)通量的效应值(ln R (N2O flux))与土壤基本理化性质之间的相关关系。本图只列出有显著相关关系的理化性质, 没有相关关系的见附件I。
Fig. 5Relationships between the effect size of freeze-thaw cycles on soil nitrous oxide (N2O) flux (ln R (N2O flux)) and basic physical and chemical characteristics of soils. Those with significant relationships were shown in this figure, but those with non-significant relationships were shown in Appendix I.
由图6可得, 冻融循环对N2O通量的效应值与冻融循环对反硝化作用速率的效应值呈显著正相关关系(r = 0.72, p < 0.001), 而与冻融循环对硝化作用速率的效应值无明显相关性。
图6
新窗口打开|下载原图ZIP|生成PPT 图6冻融循环对土壤氧化亚氮(N2O)通量的效应值(ln R (N2O flux))与冻融循环对反硝化作用速率的效应值(ln R (Denitrification))(A)和冻融循环对硝化作用速率的效应值(ln R (Nitrification))(B)之间相关关系。
Fig. 6Relationships between the effect size of freeze-thaw cycles on soil nitrous oxide (N2O) flux (ln R (N2O flux)) and the effect size of freeze-thaw cycle on denitrification rate (ln R (Denitrification))(A) and the effect size of freeze-thaw cycles on nitrification rate (ln R (Nitrification))(B).
冻融循环时间, 温度以及频率也会影响N2O排放。根据图7可知, 冻融循环过程中土壤融化时间与冻融循环对N2O通量效应值(r = 0.15, p = 0.003)和N2O累积排放量效应值(r = 0.63, p < 0.001)均呈现显著线性正相关关系。与此相反, 冻结温度与冻融循环对N2O通量效应值(r = -0.15, p = 0.002)和N2O累积排放量效应值(r = -0.49, p < 0.001)均呈现显著线性负相关关系。另外, 冻融循环频率与冻融循环对N2O通量效应值和累积排放量效应值的相关关系相反, 即分别为显著负相关关系(r = -0.26, p < 0.001)和显著正相关关系(r = 0.76, p < 0.001)。而其他冻融循环参数与N2O排放之间并没有统计学意义上的显著相关性(附件II)。
图7
新窗口打开|下载原图ZIP|生成PPT 图7冻融循环对土壤氧化亚氮(N2O)通量的效应值(ln R (N2O flux))(A, B, C)和N2O累积排放量的效应值(ln R (Cumulative N2O emissions))(D, E, F)与冻融循环时间、温度以及频率之间的相关性。本图只列出有显著相关关系的理化性质, 没有相关关系的见附件II。
Fig. 7Relationships between the effect size of freeze-thaw cycles on soil nitrous oxide (N2O) flux (ln R (N2O flux))(A, B, C) and cumulative N2O emissions (ln R (Cumulative N2O emissions))(D, E, F) and the duration, temperature, and frequency of freeze-thaw cycles. Those with significant relationships were shown in this figure, but those with non-significant relationships were shown in Appendix II.
3 讨论
本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1)。冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019)。本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1)。这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放。在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的。
Appendix I Relationships between the effect size of freeze-thaw cycles on soil nitrous oxide (N2O) flux (ln R (N2O flux)) and basic physical and chemical characteristics of soils
附件II 冻融循环对土壤氧化亚氮(N2O)通量的效应值(ln R (N2O flux))和N2O累积量排放(ln R (Cumulative N2O emissions))与冻融循环的总时间、冻结时间以及融化温度之间相关关系
Appendix II Relationships between the effect size of freeze-thaw cycles on soil nitrous oxide (N2O) flux (ln R (N2O flux)) and the effect size of freeze-thaw cycles on cumulative N2O emissions (ln R (Cumulative N2O emissions)) and the total duration, the duration of freezing, and temperature of thawing
AbalosD, van GroenigenJW, de DeynGB, (2018). What plant functional traits can reduce nitrous oxide emissions from intensively managed grasslands Global Change Biology, 24, 248-258. [本文引用: 1]
AlvarezR, SantanatogliaOJ, GarcîaR (1995). Effect of temperature on soil microbial biomass and its metabolic quotient in situ under different tillage systems Biology and Fertility of Soils, 19, 227-230. DOI:10.1007/BF00336164URL [本文引用: 1]
BaiE, LiSL, XuWH, LiW, DaiWW, JiangP (2013). A meta-analysis of experimental warming effects on terrestrial nitrogen pools and dynamics New Phytologist, 199, 441-451. DOI:10.1111/nph.2013.199.issue-2URL [本文引用: 1]
CaiYJ, DingWX, XiangJ (2012). Mechanisms of nitrous oxide and nitric oxide production in soils: a review Soils, 44, 712-718. [本文引用: 1]
ChenWJ, GongL, LiuYT (2018). Effects of seasonal snow cover on decomposition and carbon, nitrogen and phosphorus release of Picea schrenkiana leaf litter in Mt. Tianshan, Northwest China Chinese Journal of Plant Ecology, 42, 487-497. DOI:10.17521/cjpe.2017.0298URL [本文引用: 1]
CongrevesKA, Wagner-RiddleC, SiBC, CloughTJ (2018). Nitrous oxide emissions and biogeochemical responses to soil freezing-thawing and drying-wetting Soil Biology & Biochemistry, 117, 5-15. DOI:10.1016/j.soilbio.2017.10.040URL [本文引用: 3]
CuiQ, SongCC, WangXW, ShiFX, WangLL, GuoYD (2016). Rapid N2O fluxes at high level of nitrate nitrogen addition during freeze-thaw events in boreal peatlands of Northeast China Atmospheric Environment, 135, 1-8. DOI:10.1016/j.atmosenv.2016.03.053URL
DaiZ, YuM, ChenH, ZhaoH, HuangY, SuW, XiaF, ChangSX, BrookesPC, DahlgrenRA, XuJ (2020). Elevated temperature shifts soil N cycling from microbial immobilization to enhanced mineralization, nitrification and denitrification across global terrestrial ecosystems Global Change Biology, 26, 5267-5276. DOI:10.1111/gcb.v26.9URL [本文引用: 1]
FazzolariÉ, NicolardotB, GermonJC (1998). Simultaneous effects of increasing levels of glucose and oxygen partial pressures on denitrification and dissimilatory nitrate reduction to ammonium in repacked soil cores European Journal of Soil Biology, 34, 47-52. DOI:10.1016/S1164-5563(99)80006-5URL [本文引用: 2]
FitzhughRD, DriscollCT, GroffmanPM, TierneyGL, FaheyTJ, HardyJP (2001). Effects of soil freezing disturbance on soil solution nitrogen, phosphorus, and carbon chemistry in a northern hardwood ecosystem Biogeochemistry, 56, 215-238. DOI:10.1023/A:1013076609950URL [本文引用: 1]
FreppazM, WilliamsBL, EdwardsAC, ScalengheR, ZaniniE (2007). Simulating soil freeze/thaw cycles typical of winter alpine conditions: implications for N and P availability Applied Soil Ecology, 35, 247-255. DOI:10.1016/j.apsoil.2006.03.012URL [本文引用: 3]
GaoDC, ZhangL, LiuJ, PengB, FanZZ, DaiWW, JiangP, BaiE (2018). Responses of terrestrial nitrogen pools and dynamics to different patterns of freeze-thaw cycle: a meta-analysis Global Change Biology, 24, 2377-2389. DOI:10.1111/gcb.2018.24.issue-6URL [本文引用: 4]
GaoYH, ZengXY, XieQY, MaXX (2015). Release of carbon and nitrogen from alpine soils during thawing periods in the eastern Qinghai-Tibet Plateau Water, Air & Soil Pollution, 226, 1-9.
GoldbergSD, BorkenW, GebauerG (2010). N2O emission in a Norway spruce forest due to soil frost: concentration and isotope profiles shed a new light on an old story Biogeochemistry, 97, 21-30. DOI:10.1007/s10533-009-9294-zURL
GoldbergSD, MuhrJ, BorkenW, GebauerG (2008). Fluxes of climate-relevant trace gases between a Norway spruce forest soil and atmosphere during repeated freeze-thaw cycles in mesocosms Journal of Plant Nutrition and Soil Science, 171, 729-739. DOI:10.1002/jpln.v171:5URL
GroffmanPM, HardyJP, DriscollCT, FaheyTJ (2006). Snow depth, soil freezing, and fluxes of carbon dioxide, nitrous oxide and methane in a northern hardwood forest Global Change Biology, 12, 1748-1760. DOI:10.1111/gcb.2006.12.issue-9URL
GroffmanPM, HardyJP, Fashu-KanuS, DriscollCT, CleavittNL, FaheyTJ, FiskMC (2011). Snow depth, soil freezing and nitrogen cycling in a northern hardwood forest landscape Biogeochemistry, 102, 223-238. DOI:10.1007/s10533-010-9436-3URL [本文引用: 1]
GroganP, MichelsenA, AmbusP, JonassonS (2004). Freeze-thaw regime effects on carbon and nitrogen dynamics in sub-arctic heath tundra mesocosms Soil Biology & Biochemistry, 36, 641-654. DOI:10.1016/j.soilbio.2003.12.007URL [本文引用: 3]
HanCL, GuYJ, KongM, HuLW, JiaY, LiFM, SunGJ, SiddiqueKHM (2018). Responses of soil microorganisms, carbon and nitrogen to freeze-thaw cycles in diverse land-use types Applied Soil Ecology, 124, 211-217. DOI:10.1016/j.apsoil.2017.11.012URL [本文引用: 2]
HuHW, ChenDL, HeJZ (2015). Microbial regulation of terrestrial nitrous oxide formation: understanding the biological pathways for prediction of emission rates FEMS Microbiology Reviews, 39, 729-749. DOI:10.1093/femsre/fuv021URL [本文引用: 1]
IPCC (2014). Climate Change 2014: Mitigation of Climate Change. Cambridge University Press, Cambridge. [本文引用: 1]
KatayanagiN, HatanoR (2012). N2O emissions during the freezing and thawing periods from six fields in a livestock farm, southern Hokkaido, Japan Soil Science and Plant Nutrition, 58, 261-271. DOI:10.1080/00380768.2012.670810URL
KettunenR, SaarnioS (2013). Biochar can restrict N2O emissions and the risk of nitrogen leaching from an agricultural soil during the freeze-thaw period Agricultural and Food Science, 22, 373-379. DOI:10.23986/afsci.7887URL
KoponenHT, JaakkolaT, Keinänen-ToivolaMM, KaipainenS, TuomainenJ, ServomaaK, MartikainenPJ (2006). Microbial communities, biomass, and activities in soils as affected by freeze thaw cycles Soil Biology & Biochemistry, 38, 1861-1871. DOI:10.1016/j.soilbio.2005.12.010URL [本文引用: 1]
LudwigB, WolfI, TeepeR (2004). Contribution of nitrification and denitrification to the emission of N2O in a freeze-thaw event in an agricultural soil Journal of Plant Nutrition and Soil Science, 167, 678-684. DOI:10.1002/jpln.v167:6URL [本文引用: 1]
MaljanenM, KohonenAR, VirkajaäRviP, MartikainenPJ (2007). Fluxes and production of N2O, CO2 and CH4 in boreal agricultural soil during winter as affected by snow cover Tellus B: Chemical and Physical Meteorology, 59, 853-859. DOI:10.1111/j.1600-0889.2007.00304.xURL
MaljanenM, VirkajarviP, HytonenJ, OquistM, SparrmanT, MartikainenPJ (2009). Nitrous oxide production in boreal soils with variable organic matter content at low temperature- snow manipulation experiment Biogeosciences, 6, 2461-2473. DOI:10.5194/bg-6-2461-2009URL
MatznerE, BorkenW (2008). Do freeze-thaw events enhance C and N losses from soils of different ecosystems? A review European Journal of Soil Science, 59, 274-284. DOI:10.1111/ejs.2008.59.issue-2URL [本文引用: 2]
MergelA, SchmitzO, MallmannT, BotheH (2001). Relative abundance of denitrifying and dinitrogen-fixing bacteria in layers of a forest soil FEMS Microbiology Ecology, 36, 33-42. PMID:11377771 [本文引用: 1] The populations of N(2)-fixing and denitrifying bacteria in an acid forest soil near Cologne were characterized by gene probing. The DNA isolated from the soil for this purpose was suitable for DNA-DNA hybridization using 0.4-0.7-kb probes targeting denitrification enzymes, dinitrogenase reductase (nifH) and eubacterial 16S rRNA. The densitometrical comparison of band intensities obtained in these Southern hybridizations indicated that the highest number of total bacteria, of denitrifying and N(2)-fixing microorganisms always occurred in the upper ( approximately 5 cm) soil layer. The concentration of all these organisms decreased in parallel with the soil depth. The soil investigated was rich in nitrate in all layers, and the availability of nitrate apparently did not govern the distribution of denitrifying and N(2)-fixing bacteria in this soil. Soil cores investigated in the laboratory formed N(2)O on addition of nitrate irrespective of the presence of C(2)H(2). Hybridization intensities, with a gene probe for the 16S rRNA, and MPN numbers were generally higher in soil samples taken from the roots of plants than in the bulk soil. There was no selective enrichment of denitrifying or N(2)-fixing bacteria at the roots. The data obtained by hybridizing isolated soil DNA generally matched previous results obtained with culturable bacteria.
MørkvedPT, DörschP, HenriksenTM, BakkenLR (2006). N2O emissions and product ratios of nitrification and denitrification as affected by freezing and thawing Soil Biology & Biochemistry, 38, 3411-3420. DOI:10.1016/j.soilbio.2006.05.015URL [本文引用: 1]
NeilsenCB, GroffmanPM, HamburgSP, DriscollCT, FaheyTJ, HardyJP (2001). Freezing effects on carbon and nitrogen cycling in northern hardwood forest soils Soil Science Society of America Journal, 65, 1723-1730. DOI:10.2136/sssaj2001.1723URL
PelsterDE, ChantignyMH, RochetteP, AngersDA, LaganièreJ, ZebarthB, GoyerC (2013). Crop residue incorporation alters soil nitrous oxide emissions during freeze-thaw cycles Canadian Journal of Soil Science, 93, 415-425. DOI:10.4141/cjss2012-043URL [本文引用: 3]
PelsterDE, ChantignyMH, RochetteP, BertrandN, AngersDA, ZebarthBJ, GoyerC, NaethMA (2019). Rates and intensity of freeze-thaw cycles affect nitrous oxide and carbon dioxide emissions from agricultural soils Canadian Journal of Soil Science, 99, 472-484. DOI:10.1139/cjss-2019-0058 In cool temperate regions, large emissions of nitrous oxide (N2O), an important greenhouse and ozone-depleting gas, have been observed during freeze-thaw (FT) cycles. However, it is unclear how freezing and thawing rates, freezing intensity, and freezing duration influence N2O emissions. We used a laboratory incubation to measure N2O emissions from two soils (sandy loam, silty clay) undergoing a single FT cycle of various freezing and thawing rates [rapid (0.5 degrees C h(-1)) vs. slow (0.017 degrees C h(-1))], freezing intensity (-1 vs. -3 degrees C), and freezing duration (24 vs. 48 freezing degree-days). In general, soil carbon dioxide fluxes during freezing were highest when soils were frozen slowly at -1 degrees C, whereas fluxes after thawing were highest from the soils frozen and thawed rapidly at -3 degrees C. Soil N2O emissions during both the freezing and thawing periods were greatest in the soils exposed to rapid freezing to -3 degrees C, intermediate under rapid freezing to -1 degrees C and slow freezing to -3 degrees C, and smallest under slow freezing to -1 degrees C and the control treatment (constant +1 degrees C). The similar N2O emissions between the unfrozen control and the slowly frozen -1 degrees C treatment was unexpected as previous field studies with similar freezing rates and temperatures still experienced high N2O emissions during thaw. This suggests that the physical disruptions caused by freezing and thawing of the surface soil are not the primary driver of FT-induced N2O emissions under field conditions.
PrieméA, ChristensenS (2001). Natural perturbations, drying-wetting and freezing-thawing cycles, and the emission of nitrous oxide, carbon dioxide and methane from farmed organic soils Soil Biology & Biochemistry, 33, 2083-2091. DOI:10.1016/S0038-0717(01)00140-7URL [本文引用: 1]
RavishankaraAR, DanielJS, PortmannRW (2009). Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century Science, 26, 123-125. [本文引用: 1]
ReinmannAB, TemplerPH (2016). Reduced winter snowpack and greater soil frost reduce live root biomass and stimulate radial growth and stem respiration of red maple (Acer rubrum) trees in a mixed-hardwood forest Ecosystems, 19, 129-141. DOI:10.1007/s10021-015-9923-4URL [本文引用: 3]
ReinmannAB, TemplerPH, CampbellJL (2012). Severe soil frost reduces losses of carbon and nitrogen from the forest floor during simulated snowmelt: a laboratory experiment Soil Biology & Biochemistry, 44, 65-74. DOI:10.1016/j.soilbio.2011.08.018URL
RogeljJ, MeinshausenM, KnuttiR (2012). Global warming under old and new scenarios using IPCC climate sensitivity range estimates Nature Climate Change, 2, 248-253. DOI:10.1038/nclimate1385URL [本文引用: 1]
RosenbergMS, AdamsDC, GurevitchJ (2000). MetaWin: Statistical Software for Meta-analysis Sinauer Associates, Sunderland, USA. [本文引用: 1]
RuanL, RobertsonGP (2017). Reduced snow cover increases wintertime nitrous oxide (N2O) emissions from an agricultural soil in the upper U.S. Midwest Ecosystems, 20, 917-927. DOI:10.1007/s10021-016-0077-9URL
Sánchez-GarcíaM, RoigA, Sánchez-MonederoMA, CayuelaML (2014). Biochar increases soil N2O emissions produced by nitrification-mediated pathways Frontiers in Environmental Science, 2, 25. DOI: 10.3389/fenvs.2014.00025. [本文引用: 1]
SawickaJE, RobadorA, HubertC, JørgensenBB, BrüchertV (2010). Effects of freeze-thaw cycles on anaerobic microbial processes in an Arctic intertidal mud flat The ISME Journal, 4, 585-594. DOI:10.1038/ismej.2009.140URL [本文引用: 1]
SehyU, DyckmansJ, RuserR, MunchJC (2004). Adding dissolved organic carbon to simulate freeze-thaw related N2O emissions from soil Journal of Plant Nutrition and Soil Science, 167, 471-478. DOI:10.1002/jpln.v167:4URL
SeitzingerS, HarrisonJA, BöhlkeJK, BouwmanAF, LowranceR, PetersonB, TobiasC,van Drecht G (2006). Denitrification across landscapes and waterscapes: a synthesis Ecological Applications, 16, 2064-2090. DOI:10.1890/1051-0761(2006)016[2064:DALAWA]2.0.CO;2URL [本文引用: 2]
SongY, YuXF, ZouYC, WangGP, ZhangLL (2016). Progress of freeze-thaw effects on carbon, nitrogen and phosphorus cyclings in soils Soils and Crops, 5, 78-90. [本文引用: 1]
SongY, ZouYC, WangGP, YuXF (2017). Altered soil carbon and nitrogen cycles due to the freeze-thaw effect: a meta-analysis Soil Biology & Biochemistry, 109, 35-49. DOI:10.1016/j.soilbio.2017.01.020URL [本文引用: 3]
vanBochove E, PrevostD, PelletierF (2000). Effects of freeze-thaw and soil structure on nitrous oxide produced in a clay soil Soil Science Society of America Journal, 64, 1638-1643. DOI:10.2136/sssaj2000.6451638xURL
Wagner-RiddleC, CongrevesKA, AbalosD, BergAA, BrownSE, AmbadanJT, GaoX, TenutaM (2017). Globally important nitrous oxide emissions from croplands induced by freeze-thaw cycles Nature Geoscience, 10, 279-283. DOI:10.1038/NGEO2907 [本文引用: 2] Seasonal freezing induces large thawemissions of nitrous oxide, a trace gas that contributes to stratospheric ozone destruction and atmospheric warming. Cropland soils are by far the largest anthropogenic source of nitrous oxide. However, the global contribution of seasonal freezing to nitrous oxide emissions from croplands is poorly quantified, mostly due to the lack of year-round measurements and difficulty in capturing short-lived pulses of nitrous oxide with traditional measurement methods. Here we present measurements collected with half-hourly resolution at two contrasting cropland sites in Ontario and Manitoba, Canada, over 14 and 9 years, respectively. We find that the magnitude of freeze-thaw-induced nitrous oxide emissions is related to the number of days with soil temperatures below 0 degrees C, and we validate these findings with emissions data from 11 additional sites from cold climates around the globe. Based on an estimate of cropland area experiencing seasonal freezing, reanalysis model estimates of soil temperature, and the relationship between cumulative soil freezing days and emissions that we derived from the cropland sites, we estimate that seasonally frozen cropland contributes 1.07 +/- 0.59 Tg of nitrogen as nitrous oxide annually. We conclude that neglecting freeze-thaw emissions would lead to an underestimation of global agricultural nitrous oxide emissions by 17 to 28%.
Wagner-RiddleC, RapaiJ, WarlandJ, FuronA (2010). Nitrous oxide fluxes related to soil freeze and thaw periods identified using heat pulse probes Canadian Journal of Soil Science, 90, 409-418. DOI:10.4141/CJSS09016URL [本文引用: 1]
WalkerVK, PalmerGR, VoordouwG (2006). Freeze-thaw tolerance and clues to the winter survival of a soil community Applied and Environmental Microbiology, 72, 1784-1792. DOI:10.1128/AEM.72.3.1784-1792.2006URL [本文引用: 1]
WertzS, GoyerC, ZebarthBJ, TattiE, BurtonDL, ChantignyMH, FilionM (2016). The amplitude of soil freeze-thaw cycles influences temporal dynamics of N2O emissions and denitrifier transcriptional activity and community composition Biology and Fertility of Soils, 52, 1149-1162. DOI:10.1007/s00374-016-1146-0URL
WiederWR, ClevelandCC, TownsendAR (2011). Throughfall exclusion and leaf litter addition drive higher rates of soil nitrous oxide emissions from a lowland wet tropical forest Global Change Biology, 17, 3195-3207. DOI:10.1111/gcb.2011.17.issue-10URL [本文引用: 2]
WuX, BrüeggemannN, Butterbach-BahlK, FuB, LiuG (2014a). Snow cover and soil moisture controls of freeze-thaw-related soil gas fluxes from a typical semi-arid grassland soil: a laboratory experiment Biology and Fertility of Soils, 50, 295-306. DOI:10.1007/s00374-013-0853-zURL
WuX, LiT, WangDB, WangFF, FuBJ, LiuGH, LvYH (2020a). Soil properties mediate the freeze-thaw-related soil N2O and CO2 emissions from temperate grasslands Catena, 195, 104797. DOI: 10.1016/j.catena.2020.104797. URL
WuX, LiuHF, ZhangLN, FuBJ, LiZS, WangQB, LiuGH (2014b). Effects of snow cover and soil moisture on CO2 and N2O fluxes from typical semi-arid grassland soil subjected to freeze-thaw cycles Acta Ecologica Sinica, 34, 5484-5493.
WuX, WangFF, LiT, FuBJ, LvY, LiuGH (2020b). Nitrogen additions increase N2O emissions but reduce soil respiration and CH4 uptake during freeze-thaw cycles in an alpine meadow Geoderma, 363, 114157. DOI:10.1016/j.geoderma.2019.114157URL [本文引用: 1]
XiaoL, ZhangY, LiP, XuGC, ShiP, ZhangY (2019). Effects of freeze-thaw cycles on aggregate-associated organic carbon and glomalin-related soil protein in natural- succession grassland and Chinese pine forest on the Loess Plateau Geoderma, 334, 1-8. DOI:10.1016/j.geoderma.2018.07.043 [本文引用: 5] Glomalin-related soil protein (GRSP) is a hydrophobic glycoprotein that can protect soil organic carbon (SOC) from decomposition by promoting the formation of soil aggregates. Few studies, however, have evaluated the response of aggregate-associated SOC and GRSP to freeze-thaw cycles (FTs) under different patterns of vegetation restoration. Soil samples were collected from three typical natural-succession grasslands (dominated by Artemisia capalaris, A. sacrorum, and Bothriochloa ischaemum) and a Chinese pine (Pains tabuliformis) forest before and after natural FTs during a winter-spring season on the Loess Plateau in China. The soil aggregate stability and concentrations of SOC, easily extractable GRSP (EE-GRSP) and total GRSP (T-GRSP) in different aggregate fractions were measured. The FTs decreased the proportion of > 5.0 mm wet-sieved fractions, increased the proportion of < 0.25 mm wet-sieved fractions, and decreased the mean weight diameters by 8.7, 9.5, 2.9, and 9.4% in the soils of A. capiilaris, A. sacrorum, B. ischaemum, and P. tabuliformis, respectively. The SOC, T-GRSP, and EE-GRSP concentrations in each aggregate fraction generally decreased after FTs in the four vegetation types. The SOC, T-GRSP, and EE-GRSP concentrations did not differ significantly in the larger aggregate fractions before the FTs in the soils for B. ischaemum and P. tabuliformis, but their concentrations were significantly higher for B. ischaemum than P. tabuliformis after FTs. The SOC and GRSP concentrations were significantly and linearly correlated in most aggregate fractions both before and after the FTs. These results indicated that the FTs decreased SOC by decreasing the GRSP concentration in each aggregate fraction and that SOC was more sensitive to FTs in forest than in natural-succession grassland.
YanaiY, HirotaT, IwataY, NemotoM, NagataO, KogaN (2011). Accumulation of nitrous oxide and depletion of oxygen in seasonally frozen soils in northern Japan- Snow cover manipulation experiments Soil Biology & Biochemistry, 43, 1779-1786. DOI:10.1016/j.soilbio.2010.06.009URL [本文引用: 1]
YanaiY, ToyotaK, OkazakiM (2004). Effects of successive soil freeze-thaw cycles on soil microbial biomass and organic matter decomposition potential of soils Soil Science and Plant Nutrition, 50, 821-829. DOI:10.1080/00380768.2004.10408542URL [本文引用: 2]
YanaiY, ToyotaK, OkazakiM (2007). Response of denitrifying communities to successive soil freeze-thaw cycles Biology and Fertility of Soils, 44, 113-119. DOI:10.1007/s00374-007-0185-yURL
YangHL, QinJH, SunH (2010). A review: response of soil CO2 and N2O emissions to freeze-thaw pattern change Soils, 42, 519-525. [本文引用: 1]
YangKJ, YangWQ, TanY, HeRY, ZhuangLY, LiZJ, TanB, XuZF (2017). Short-term responses of winter soil respiration to snow removal in a Picea asperata forest of western Sichuan Chinese Journal of Plant Ecology, 41, 964-971. DOI:10.17521/cjpe.2017.0015URL [本文引用: 1]
YaoZ, WuX, WolfB, DannenmannM, Butterbach-BahlK, BrueggemannN, ChenW, ZhengX (2010). Soil-atmosphere exchange potential of NO and N2O in different land use types of Inner Mongolia as affected by soil temperature, soil moisture, freeze-thaw, and drying- wetting events Journal of Geophysical Research-Atmospheres, 115, D17116. DOI: 10.1029/2009JD013528. URL
YinSX, ShenQR, TangY, ChengLM (1998). Reduction of nitrate to ammonium in selected paddy soils of China Pedosphere, 8, 221-228. [本文引用: 2]
YuXF, ZhangYX, ZhaoHM, LuXG, WangGP (2010). Freeze-thaw effects on sorption/desorption of dissolved organic carbon in wetland soils Chinese Geographical Science, 20, 209-217. DOI:10.1007/s11769-010-0209-7URL [本文引用: 2]
ZhangXY, WangW, ChenWL, ZhangNL, ZengH (2014). Comparison of seasonal soil microbial process in snow-covered temperate ecosystems of northern China PLOS ONE, 9, e92985. DOI: 10.1371/journal.pone.0092985. URL [本文引用: 3]
ZhouW, ChenH, ZhouL, LewisBJ, YeY, TianJ, LiG, DaiL (2011). Effect of freezing-thawing on nitrogen mineralization in vegetation soils of four landscape zones of Changbai Mountain Annals of Forest Science, 68, 943-951. DOI:10.1007/s13595-011-0100-4URL [本文引用: 2]
Micrometeorological measurements over 3 years reveal differences in N2O emissions between annual and perennial crops 1 2016
... 虽然在不同研究地区冻融循环均会显著促进土壤N2O的排放, 然而土壤N2O累积排放量在不同生态系统表现出明显差异, 具体表现为农田>草地>森林(图3).原因可能是农田生态系统土壤硝态氮含量高于其他生态系统, 较高的硝态氮含量可为反硝化作用产生N2O提供充足底物(Congreves et al., 2018).这点也间接被本研究结论所支持, 即土壤硝态氮含量与土壤N2O排放通量的效应值呈显著正相关关系(图5).本研究还指出, 在苔原或者湿地生态系统, 冻融循环对N2O排放通量的影响较其他生态系统下的大, 主要原因可能是这些地区含有高的有机质和铵态氮含量, 有利用异氧硝化和反硝化真菌的作用, 从而产生更多N2O (蔡延江等, 2012).一般养分含量较高的地区在冻融循环期间土壤N2O排放会偏高, 比如在农业系统中, 每年30%-90%的N2O排放归因于土壤冻融循环作用(Yanai et al., 2011; Abalos et al., 2016), 因此在未来模型模拟预测全球农田N2O排放时, 需要特别考虑冻融循环作用的影响, 否则会低估农田生态系统17%-28%的N2O排放(Wagner-Riddle et al., 2017). ...
What plant functional traits can reduce nitrous oxide emissions from intensively managed grasslands 1 2018
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
Effect of temperature on soil microbial biomass and its metabolic quotient in situ under different tillage systems 1 1995
... 在室内研究中, 土壤采样的时间和土壤的处理方式也会影响N2O排放对冻融循环响应的大小(图3).比如, 与非冻融循环期间采集的土壤相比, 在冻融循环期间采集的土壤N2O排放效应值较大.原因可能与微生物特性和养分含量有关.在冻融与非冻融期间土壤微生物群落结构和微生物适应性是不同的(Zhang et al., 2014).在秋末至初冬, 土壤反硝化细菌的数量明显高于夏季(Mergel et al., 2001).由于冻融循环期间土壤N2O的产生主要是通过反硝化作用, 因此反硝化细菌数量增多可提高反硝化作用速率, 从而增加N2O产生的量.相比于夏季, 冬季土壤微生物对于温度的适应性较强(Zhang et al., 2014), 在经历温度的降低和升高时, 仍然能保持较高的微生物活性, 从而提高N2O的产生.另外, 土壤微生物量往往是冬季大于夏季(Alvarez et al., 1995; Zhang et al., 2014), 在冻融循环过程中微生物死亡可释放更多可利用性养分, 这就为冻融循环过程中N2O的产生提供充足的底物.除了采集土壤的时间外, 土壤处理方式也会影响N2O排放对于冻融循环的响应.本研究得出, 与土壤不过筛相比, 土壤过筛会显著增加N2O排放对冻融循环的响应(图3).主要原因是过筛会破坏土壤结构.一方面土壤团聚体晶格结构的破坏会释放更多可利用性养分进入土壤(Xiao et al., 2019), 另一方面土壤过筛也会导致土壤胶体表面吸附的养分离子释放出来(Freppaz et al., 2007), 这些可利用性养分的增加可为N2O产生提供充足的底物.在非冻融循环期间采集土壤和土壤过筛处理可能会分别导致冻融循环效应被低估和高估, 因此为了得到真实的数据和探究其真正地内在机制, 今后在室内研究冻融循环试验时, 应尽量模拟原位条件.比如在真正野外冻融循环期间采集土壤样品以及野外采集的土壤样品不要过筛. ...
A meta-analysis of experimental warming effects on terrestrial nitrogen pools and dynamics 1 2013
... 为了平衡来自不同研究的权重, 权衡后的权重按照公式(5)(Bai et al., 2013)计算: ...
土壤N2O和NO产生机制研究进展 1 2012
... 虽然在不同研究地区冻融循环均会显著促进土壤N2O的排放, 然而土壤N2O累积排放量在不同生态系统表现出明显差异, 具体表现为农田>草地>森林(图3).原因可能是农田生态系统土壤硝态氮含量高于其他生态系统, 较高的硝态氮含量可为反硝化作用产生N2O提供充足底物(Congreves et al., 2018).这点也间接被本研究结论所支持, 即土壤硝态氮含量与土壤N2O排放通量的效应值呈显著正相关关系(图5).本研究还指出, 在苔原或者湿地生态系统, 冻融循环对N2O排放通量的影响较其他生态系统下的大, 主要原因可能是这些地区含有高的有机质和铵态氮含量, 有利用异氧硝化和反硝化真菌的作用, 从而产生更多N2O (蔡延江等, 2012).一般养分含量较高的地区在冻融循环期间土壤N2O排放会偏高, 比如在农业系统中, 每年30%-90%的N2O排放归因于土壤冻融循环作用(Yanai et al., 2011; Abalos et al., 2016), 因此在未来模型模拟预测全球农田N2O排放时, 需要特别考虑冻融循环作用的影响, 否则会低估农田生态系统17%-28%的N2O排放(Wagner-Riddle et al., 2017). ...
土壤N2O和NO产生机制研究进展 1 2012
... 虽然在不同研究地区冻融循环均会显著促进土壤N2O的排放, 然而土壤N2O累积排放量在不同生态系统表现出明显差异, 具体表现为农田>草地>森林(图3).原因可能是农田生态系统土壤硝态氮含量高于其他生态系统, 较高的硝态氮含量可为反硝化作用产生N2O提供充足底物(Congreves et al., 2018).这点也间接被本研究结论所支持, 即土壤硝态氮含量与土壤N2O排放通量的效应值呈显著正相关关系(图5).本研究还指出, 在苔原或者湿地生态系统, 冻融循环对N2O排放通量的影响较其他生态系统下的大, 主要原因可能是这些地区含有高的有机质和铵态氮含量, 有利用异氧硝化和反硝化真菌的作用, 从而产生更多N2O (蔡延江等, 2012).一般养分含量较高的地区在冻融循环期间土壤N2O排放会偏高, 比如在农业系统中, 每年30%-90%的N2O排放归因于土壤冻融循环作用(Yanai et al., 2011; Abalos et al., 2016), 因此在未来模型模拟预测全球农田N2O排放时, 需要特别考虑冻融循环作用的影响, 否则会低估农田生态系统17%-28%的N2O排放(Wagner-Riddle et al., 2017). ...
Increased nitrogen leaching following soil freezing is due to decreased root uptake in a northern hardwood forest 3 2014
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1).冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019).本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1).这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放.在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的. ...
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
季节性雪被对天山雪岭云杉凋落叶分解和碳氮磷释放的影响 1 2018
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
季节性雪被对天山雪岭云杉凋落叶分解和碳氮磷释放的影响 1 2018
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
Nitrous oxide emission from an agricultural soil subjected to different freeze-thaw cycles 0 1995
冻融对土壤氮素损失及有效性的影响 2 2016
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
冻融对土壤氮素损失及有效性的影响 2 2016
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
Nitrous oxide emissions and biogeochemical responses to soil freezing-thawing and drying-wetting 3 2018
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
... 虽然在不同研究地区冻融循环均会显著促进土壤N2O的排放, 然而土壤N2O累积排放量在不同生态系统表现出明显差异, 具体表现为农田>草地>森林(图3).原因可能是农田生态系统土壤硝态氮含量高于其他生态系统, 较高的硝态氮含量可为反硝化作用产生N2O提供充足底物(Congreves et al., 2018).这点也间接被本研究结论所支持, 即土壤硝态氮含量与土壤N2O排放通量的效应值呈显著正相关关系(图5).本研究还指出, 在苔原或者湿地生态系统, 冻融循环对N2O排放通量的影响较其他生态系统下的大, 主要原因可能是这些地区含有高的有机质和铵态氮含量, 有利用异氧硝化和反硝化真菌的作用, 从而产生更多N2O (蔡延江等, 2012).一般养分含量较高的地区在冻融循环期间土壤N2O排放会偏高, 比如在农业系统中, 每年30%-90%的N2O排放归因于土壤冻融循环作用(Yanai et al., 2011; Abalos et al., 2016), 因此在未来模型模拟预测全球农田N2O排放时, 需要特别考虑冻融循环作用的影响, 否则会低估农田生态系统17%-28%的N2O排放(Wagner-Riddle et al., 2017). ...
Rapid N2O fluxes at high level of nitrate nitrogen addition during freeze-thaw events in boreal peatlands of Northeast China 0 2016
Elevated temperature shifts soil N cycling from microbial immobilization to enhanced mineralization, nitrification and denitrification across global terrestrial ecosystems 1 2020
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
Simultaneous effects of increasing levels of glucose and oxygen partial pressures on denitrification and dissimilatory nitrate reduction to ammonium in repacked soil cores 2 1998
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
Effects of soil freezing disturbance on soil solution nitrogen, phosphorus, and carbon chemistry in a northern hardwood ecosystem 1 2001
... 冻融循环格局的改变也会显著影响土壤N2O对冻融循环响应的大小.土壤融化时间与N2O排放呈显著正相关关系(图7).随着土壤融化时间的增加, 反硝化细菌活性逐渐恢复, 从而可能增加反硝化强度, 进而增加N2O产生与排放.随着冻结温度降低, 土壤N2O排放的效应值随之增加(图7).当冻结温度低于-10 ℃时, 土壤N2O排放通量要高于冻结温度大于-10 ℃时(图3).主要原因是冻结强度较高会增加土壤团聚体结构和微生物破坏(Fitzhugh et al., 2001; Oztas & Fayetorbay, 2003; Zhou et al., 2011), 从而促进了可溶性有机碳和无机氮的释放, 这些可溶性碳、氮含量的增加可促进硝化作用和反硝化作用速率, 进而增加了土壤N2O产生.另外, 本研究结果表明, 冻融循环频率与土壤N2O通量呈显著负相关关系(图7).随着冻融循环频率增加, 土壤养分可通过微生物利用、淋洗损失以及土壤颗粒重吸附等方式降低(Grogan et al., 2004; Yu et al., 2010; Han et al., 2018; Xiao et al., 2019), 使氮转化过程减慢, 从而导致N2O排放通量随之减少.有趣的是, 本研究发现冻融循环频率与土壤N2O累积排放量反而呈显著正相关关系(图7).虽然随着冻融循环频率增加, N2O通量随之降低, 这种降低是相对于第一次的N2O通量值, 而相对于对照组, N2O通量仍然是增加的.只是随着冻融循环频率增加, 这种增加幅度在降低.因此, 土壤N2O累积排放量会随着冻融循环频率增加而增加.在未来气候变暖背景下, 可能会减少雪被厚度和雪被存在时间(Groffman et al., 2011), 由于雪被起到隔热层作用, 因此这些变化可能引起冻融格局发生改变, 比如冻融循环时间、强度以及频率都会增加(杨开军等, 2017), 这种改变可能会加剧土壤N2O累积排放量, 从而加剧全球气候温室效应和臭氧层破坏. ...
Simulating soil freeze/thaw cycles typical of winter alpine conditions: implications for N and P availability 3 2007
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1).冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019).本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1).这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放.在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的. ...
... 在室内研究中, 土壤采样的时间和土壤的处理方式也会影响N2O排放对冻融循环响应的大小(图3).比如, 与非冻融循环期间采集的土壤相比, 在冻融循环期间采集的土壤N2O排放效应值较大.原因可能与微生物特性和养分含量有关.在冻融与非冻融期间土壤微生物群落结构和微生物适应性是不同的(Zhang et al., 2014).在秋末至初冬, 土壤反硝化细菌的数量明显高于夏季(Mergel et al., 2001).由于冻融循环期间土壤N2O的产生主要是通过反硝化作用, 因此反硝化细菌数量增多可提高反硝化作用速率, 从而增加N2O产生的量.相比于夏季, 冬季土壤微生物对于温度的适应性较强(Zhang et al., 2014), 在经历温度的降低和升高时, 仍然能保持较高的微生物活性, 从而提高N2O的产生.另外, 土壤微生物量往往是冬季大于夏季(Alvarez et al., 1995; Zhang et al., 2014), 在冻融循环过程中微生物死亡可释放更多可利用性养分, 这就为冻融循环过程中N2O的产生提供充足的底物.除了采集土壤的时间外, 土壤处理方式也会影响N2O排放对于冻融循环的响应.本研究得出, 与土壤不过筛相比, 土壤过筛会显著增加N2O排放对冻融循环的响应(图3).主要原因是过筛会破坏土壤结构.一方面土壤团聚体晶格结构的破坏会释放更多可利用性养分进入土壤(Xiao et al., 2019), 另一方面土壤过筛也会导致土壤胶体表面吸附的养分离子释放出来(Freppaz et al., 2007), 这些可利用性养分的增加可为N2O产生提供充足的底物.在非冻融循环期间采集土壤和土壤过筛处理可能会分别导致冻融循环效应被低估和高估, 因此为了得到真实的数据和探究其真正地内在机制, 今后在室内研究冻融循环试验时, 应尽量模拟原位条件.比如在真正野外冻融循环期间采集土壤样品以及野外采集的土壤样品不要过筛. ...
Responses of terrestrial nitrogen pools and dynamics to different patterns of freeze-thaw cycle: a meta-analysis 4 2018
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... ., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1).冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019).本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1).这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放.在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的. ...
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
Rates and intensity of freeze-thaw cycles affect nitrous oxide and carbon dioxide emissions from agricultural soils 0 2019
Natural perturbations, drying-wetting and freezing-thawing cycles, and the emission of nitrous oxide, carbon dioxide and methane from farmed organic soils 1 2001
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
Nitrous oxide (N2O): the dominant ozone-depleting substance emitted in the 21st century 1 2009
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
Reduced winter snowpack and greater soil frost reduce live root biomass and stimulate radial growth and stem respiration of red maple (Acer rubrum) trees in a mixed-hardwood forest 3 2016
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1).冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019).本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1).这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放.在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的. ...
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
Severe soil frost reduces losses of carbon and nitrogen from the forest floor during simulated snowmelt: a laboratory experiment 0 2012
Global warming under old and new scenarios using IPCC climate sensitivity range estimates 1 2012
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
MetaWin: Statistical Software for Meta-analysis 1 2000
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
冻融作用对土壤碳、氮、磷循环的影响 1 2016
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
冻融作用对土壤碳、氮、磷循环的影响 1 2016
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
Altered soil carbon and nitrogen cycles due to the freeze-thaw effect: a meta-analysis 3 2017
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
Effects of freeze-thaw and soil structure on nitrous oxide produced in a clay soil 0 2000
Globally important nitrous oxide emissions from croplands induced by freeze-thaw cycles 2 2017
... 氧化亚氮(N2O)作为一种重要温室气体, 对全球变暖贡献率可达到6%, 此外它也是破坏臭氧层的主要气体之一(Ravinshakara et al., 2009; Abalos et al., 2018).全球N2O产生和排放途径很多, 其中以土壤排放为主, 可占全球排放总量的60%左右(IPCC, 2014).在土壤排放的N2O中, 以冻融循环期间的排放量相对较高, 可占到土壤年排放总量的20%-90% (陈哲等, 2016; 宋阳等, 2016; Wagner- Riddle et al., 2017; Congreves et al., 2018).随着未来全球气候变暖(Rogelj et al., 2012), 冻融循环格局可能也会随之发生改变, 比如冻融循环时间、强度和频率增加, 这些变化可能会通过影响土壤基本理化性质及微生物特性等途径来改变土壤N2O排放.因此, 研究土壤N2O在冻融循环期间的排放规律以及作用机制具有重要意义. ...
... 虽然在不同研究地区冻融循环均会显著促进土壤N2O的排放, 然而土壤N2O累积排放量在不同生态系统表现出明显差异, 具体表现为农田>草地>森林(图3).原因可能是农田生态系统土壤硝态氮含量高于其他生态系统, 较高的硝态氮含量可为反硝化作用产生N2O提供充足底物(Congreves et al., 2018).这点也间接被本研究结论所支持, 即土壤硝态氮含量与土壤N2O排放通量的效应值呈显著正相关关系(图5).本研究还指出, 在苔原或者湿地生态系统, 冻融循环对N2O排放通量的影响较其他生态系统下的大, 主要原因可能是这些地区含有高的有机质和铵态氮含量, 有利用异氧硝化和反硝化真菌的作用, 从而产生更多N2O (蔡延江等, 2012).一般养分含量较高的地区在冻融循环期间土壤N2O排放会偏高, 比如在农业系统中, 每年30%-90%的N2O排放归因于土壤冻融循环作用(Yanai et al., 2011; Abalos et al., 2016), 因此在未来模型模拟预测全球农田N2O排放时, 需要特别考虑冻融循环作用的影响, 否则会低估农田生态系统17%-28%的N2O排放(Wagner-Riddle et al., 2017). ...
Nitrous oxide fluxes related to soil freeze and thaw periods identified using heat pulse probes 1 2010
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
Freeze-thaw tolerance and clues to the winter survival of a soil community 1 2006
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
The amplitude of soil freeze-thaw cycles influences temporal dynamics of N2O emissions and denitrifier transcriptional activity and community composition 0 2016
Throughfall exclusion and leaf litter addition drive higher rates of soil nitrous oxide emissions from a lowland wet tropical forest 2 2011
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
Snow cover and soil moisture controls of freeze-thaw-related soil gas fluxes from a typical semi-arid grassland soil: a laboratory experiment 0 2014a
Soil properties mediate the freeze-thaw-related soil N2O and CO2 emissions from temperate grasslands 0 2020a
雪被和土壤水分对典型半干旱草原土壤冻融过程中CO2和N2O排放的影响 0 2014b
雪被和土壤水分对典型半干旱草原土壤冻融过程中CO2和N2O排放的影响 0 2014b
Nitrogen additions increase N2O emissions but reduce soil respiration and CH4 uptake during freeze-thaw cycles in an alpine meadow 1 2020b
... 本研究得出, 试验方法的差异并没有改变土壤N2O排放对冻融循环的响应(图3).前人在室内模拟冻融循环试验布局并不合理, 比如土壤体积较小, 冻融循环强度大, 温度变化速率较快, 采集土壤样品的时间不对等(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018), 这些与室外条件不切合的因素会导致土壤微生物在最初的冻融循环过程后超过半数的微生物死亡(Koponen et al., 2006; Walker et al., 2006; Sawicka et al., 2010), 以及土壤团聚体受到破坏较大, 从而使养分在最初的冻融阶段被大量释放出来, 进而导致在冻融循环初期N2O的排放通量陡然增加.然而随着冻融循环频率和时间的增加, 养分很快被消耗, N2O排放通量表现出明显的下降趋势, 甚至会低于对照条件下(Wu et al., 2020b).而在室外原位条件下, 虽然在冻融循环初期, N2O排放通量增加幅度没有室内那么大, 但在冻融循环过程中不断有养分供应, 比如死亡的植物根系释放(Campbell et al., 2014; Reinmann & Templer, 2016)和凋落物分解释放(Wieder et al., 2011; Pelster et al., 2013; 陈文静等, 2018).这点也被本研究所证实, 在有植物存在的条件下, 土壤N2O排放通量对冻融循环的响应大于在无植物存在的条件下(图3).因此, 随着冻融循环频率和时间的增加, 在室外条件下N2O排放通量表现为较为稳定的水平, 而在室内培养条件下N2O排放量表现为先增加后降低的趋势..最后计算两种条件在不同频率和时间的平均N2O通量时, 可能导致两种方法的平均N2O通量并没有明显差异. ...
Effects of freeze-thaw cycles on aggregate-associated organic carbon and glomalin-related soil protein in natural- succession grassland and Chinese pine forest on the Loess Plateau 5 2019
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 除了土壤本身的性质外, 冻融循环格局不同也可能会影响N2O对冻融循环的响应.前人研究结果(Oztas & Fayetorbay, 2003; Zhou et al., 2011)显示, 冻结强度增加, 可提高其对微生物和土壤结构的影响, 从而释放更多养分, 进而增加土壤N2O排放.随着冻融循环频率增加和冻融循环时间的增加, 土壤可利用养分可能会通过以下方式被降低: 养分被微生物不断消耗, 养分的淋洗损失, 养分被土壤颗粒重吸附(Grogan et al., 2004; Yu et al., 2010; Han e al., 2018; Xiao et al., 2019).这些可利用养分的减少会导致N2O对冻融循环的响应随之下降.相比于N2O排放通量, 冻融循环期间N2O累积排放量的生态学意义更大.然而前人的研究都集中在N2O排放通量, 并没有探究N2O累积排放量与冻融循环格局变化之间的关系.未来气候变化背景下, 冻融循环格局也会发生相应改变, 比如冻融时间延长, 冻融循环的强度和频率增加等, 那么这些变化会给N2O累积排放量产生什么样的影响? 前人研究并不足. ...
... 本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1).冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019).本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1).这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放.在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的. ...
... 冻融循环格局的改变也会显著影响土壤N2O对冻融循环响应的大小.土壤融化时间与N2O排放呈显著正相关关系(图7).随着土壤融化时间的增加, 反硝化细菌活性逐渐恢复, 从而可能增加反硝化强度, 进而增加N2O产生与排放.随着冻结温度降低, 土壤N2O排放的效应值随之增加(图7).当冻结温度低于-10 ℃时, 土壤N2O排放通量要高于冻结温度大于-10 ℃时(图3).主要原因是冻结强度较高会增加土壤团聚体结构和微生物破坏(Fitzhugh et al., 2001; Oztas & Fayetorbay, 2003; Zhou et al., 2011), 从而促进了可溶性有机碳和无机氮的释放, 这些可溶性碳、氮含量的增加可促进硝化作用和反硝化作用速率, 进而增加了土壤N2O产生.另外, 本研究结果表明, 冻融循环频率与土壤N2O通量呈显著负相关关系(图7).随着冻融循环频率增加, 土壤养分可通过微生物利用、淋洗损失以及土壤颗粒重吸附等方式降低(Grogan et al., 2004; Yu et al., 2010; Han et al., 2018; Xiao et al., 2019), 使氮转化过程减慢, 从而导致N2O排放通量随之减少.有趣的是, 本研究发现冻融循环频率与土壤N2O累积排放量反而呈显著正相关关系(图7).虽然随着冻融循环频率增加, N2O通量随之降低, 这种降低是相对于第一次的N2O通量值, 而相对于对照组, N2O通量仍然是增加的.只是随着冻融循环频率增加, 这种增加幅度在降低.因此, 土壤N2O累积排放量会随着冻融循环频率增加而增加.在未来气候变暖背景下, 可能会减少雪被厚度和雪被存在时间(Groffman et al., 2011), 由于雪被起到隔热层作用, 因此这些变化可能引起冻融格局发生改变, 比如冻融循环时间、强度以及频率都会增加(杨开军等, 2017), 这种改变可能会加剧土壤N2O累积排放量, 从而加剧全球气候温室效应和臭氧层破坏. ...
... 在室内研究中, 土壤采样的时间和土壤的处理方式也会影响N2O排放对冻融循环响应的大小(图3).比如, 与非冻融循环期间采集的土壤相比, 在冻融循环期间采集的土壤N2O排放效应值较大.原因可能与微生物特性和养分含量有关.在冻融与非冻融期间土壤微生物群落结构和微生物适应性是不同的(Zhang et al., 2014).在秋末至初冬, 土壤反硝化细菌的数量明显高于夏季(Mergel et al., 2001).由于冻融循环期间土壤N2O的产生主要是通过反硝化作用, 因此反硝化细菌数量增多可提高反硝化作用速率, 从而增加N2O产生的量.相比于夏季, 冬季土壤微生物对于温度的适应性较强(Zhang et al., 2014), 在经历温度的降低和升高时, 仍然能保持较高的微生物活性, 从而提高N2O的产生.另外, 土壤微生物量往往是冬季大于夏季(Alvarez et al., 1995; Zhang et al., 2014), 在冻融循环过程中微生物死亡可释放更多可利用性养分, 这就为冻融循环过程中N2O的产生提供充足的底物.除了采集土壤的时间外, 土壤处理方式也会影响N2O排放对于冻融循环的响应.本研究得出, 与土壤不过筛相比, 土壤过筛会显著增加N2O排放对冻融循环的响应(图3).主要原因是过筛会破坏土壤结构.一方面土壤团聚体晶格结构的破坏会释放更多可利用性养分进入土壤(Xiao et al., 2019), 另一方面土壤过筛也会导致土壤胶体表面吸附的养分离子释放出来(Freppaz et al., 2007), 这些可利用性养分的增加可为N2O产生提供充足的底物.在非冻融循环期间采集土壤和土壤过筛处理可能会分别导致冻融循环效应被低估和高估, 因此为了得到真实的数据和探究其真正地内在机制, 今后在室内研究冻融循环试验时, 应尽量模拟原位条件.比如在真正野外冻融循环期间采集土壤样品以及野外采集的土壤样品不要过筛. ...
Accumulation of nitrous oxide and depletion of oxygen in seasonally frozen soils in northern Japan- Snow cover manipulation experiments 1 2011
... 虽然在不同研究地区冻融循环均会显著促进土壤N2O的排放, 然而土壤N2O累积排放量在不同生态系统表现出明显差异, 具体表现为农田>草地>森林(图3).原因可能是农田生态系统土壤硝态氮含量高于其他生态系统, 较高的硝态氮含量可为反硝化作用产生N2O提供充足底物(Congreves et al., 2018).这点也间接被本研究结论所支持, 即土壤硝态氮含量与土壤N2O排放通量的效应值呈显著正相关关系(图5).本研究还指出, 在苔原或者湿地生态系统, 冻融循环对N2O排放通量的影响较其他生态系统下的大, 主要原因可能是这些地区含有高的有机质和铵态氮含量, 有利用异氧硝化和反硝化真菌的作用, 从而产生更多N2O (蔡延江等, 2012).一般养分含量较高的地区在冻融循环期间土壤N2O排放会偏高, 比如在农业系统中, 每年30%-90%的N2O排放归因于土壤冻融循环作用(Yanai et al., 2011; Abalos et al., 2016), 因此在未来模型模拟预测全球农田N2O排放时, 需要特别考虑冻融循环作用的影响, 否则会低估农田生态系统17%-28%的N2O排放(Wagner-Riddle et al., 2017). ...
Effects of successive soil freeze-thaw cycles on soil microbial biomass and organic matter decomposition potential of soils 2 2004
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 本研究结果表明, 冻融循环可显著增加N2O通量和累积排放量, 平均增幅达72.34%和143.25% (图1).冻融循环主要通过破坏土壤结构, 杀死微生物和根系细胞以及破坏凋落物等途径促进土壤可利用性养分的释放(Yanai et al., 2004; Freppaz et al., 2007; Pelster et al., 2013; Campbell et al., 2014; Reinmann & Templer, 2016; Xiao et al., 2019).本研究也证实, 冻融循环可显著减少微生物生物量氮含量, 减幅可达6.39% (图1).这些可溶性养分的增加, 一方面可为存活的微生物提供养分, 提高其活性; 另一方面这些养分可为氮转化过程提供充足的底物, 使土壤硝化作用和反硝化作用速率增强(图1), 在这些氮转化过程中会产生N2O, 从而增加了土壤N2O的排放.在不同研究区域和冻融循环格局下, 土壤N2O对冻融循环的响应有所差异, 下面探究这些差异是如何影响N2O对冻融循环响应的. ...
Response of denitrifying communities to successive soil freeze-thaw cycles 0 2007
... 冻融循环格局的改变也会显著影响土壤N2O对冻融循环响应的大小.土壤融化时间与N2O排放呈显著正相关关系(图7).随着土壤融化时间的增加, 反硝化细菌活性逐渐恢复, 从而可能增加反硝化强度, 进而增加N2O产生与排放.随着冻结温度降低, 土壤N2O排放的效应值随之增加(图7).当冻结温度低于-10 ℃时, 土壤N2O排放通量要高于冻结温度大于-10 ℃时(图3).主要原因是冻结强度较高会增加土壤团聚体结构和微生物破坏(Fitzhugh et al., 2001; Oztas & Fayetorbay, 2003; Zhou et al., 2011), 从而促进了可溶性有机碳和无机氮的释放, 这些可溶性碳、氮含量的增加可促进硝化作用和反硝化作用速率, 进而增加了土壤N2O产生.另外, 本研究结果表明, 冻融循环频率与土壤N2O通量呈显著负相关关系(图7).随着冻融循环频率增加, 土壤养分可通过微生物利用、淋洗损失以及土壤颗粒重吸附等方式降低(Grogan et al., 2004; Yu et al., 2010; Han et al., 2018; Xiao et al., 2019), 使氮转化过程减慢, 从而导致N2O排放通量随之减少.有趣的是, 本研究发现冻融循环频率与土壤N2O累积排放量反而呈显著正相关关系(图7).虽然随着冻融循环频率增加, N2O通量随之降低, 这种降低是相对于第一次的N2O通量值, 而相对于对照组, N2O通量仍然是增加的.只是随着冻融循环频率增加, 这种增加幅度在降低.因此, 土壤N2O累积排放量会随着冻融循环频率增加而增加.在未来气候变暖背景下, 可能会减少雪被厚度和雪被存在时间(Groffman et al., 2011), 由于雪被起到隔热层作用, 因此这些变化可能引起冻融格局发生改变, 比如冻融循环时间、强度以及频率都会增加(杨开军等, 2017), 这种改变可能会加剧土壤N2O累积排放量, 从而加剧全球气候温室效应和臭氧层破坏. ...
川西亚高山云杉林冬季土壤呼吸对雪被去除的短期响应 1 2017
... 冻融循环格局的改变也会显著影响土壤N2O对冻融循环响应的大小.土壤融化时间与N2O排放呈显著正相关关系(图7).随着土壤融化时间的增加, 反硝化细菌活性逐渐恢复, 从而可能增加反硝化强度, 进而增加N2O产生与排放.随着冻结温度降低, 土壤N2O排放的效应值随之增加(图7).当冻结温度低于-10 ℃时, 土壤N2O排放通量要高于冻结温度大于-10 ℃时(图3).主要原因是冻结强度较高会增加土壤团聚体结构和微生物破坏(Fitzhugh et al., 2001; Oztas & Fayetorbay, 2003; Zhou et al., 2011), 从而促进了可溶性有机碳和无机氮的释放, 这些可溶性碳、氮含量的增加可促进硝化作用和反硝化作用速率, 进而增加了土壤N2O产生.另外, 本研究结果表明, 冻融循环频率与土壤N2O通量呈显著负相关关系(图7).随着冻融循环频率增加, 土壤养分可通过微生物利用、淋洗损失以及土壤颗粒重吸附等方式降低(Grogan et al., 2004; Yu et al., 2010; Han et al., 2018; Xiao et al., 2019), 使氮转化过程减慢, 从而导致N2O排放通量随之减少.有趣的是, 本研究发现冻融循环频率与土壤N2O累积排放量反而呈显著正相关关系(图7).虽然随着冻融循环频率增加, N2O通量随之降低, 这种降低是相对于第一次的N2O通量值, 而相对于对照组, N2O通量仍然是增加的.只是随着冻融循环频率增加, 这种增加幅度在降低.因此, 土壤N2O累积排放量会随着冻融循环频率增加而增加.在未来气候变暖背景下, 可能会减少雪被厚度和雪被存在时间(Groffman et al., 2011), 由于雪被起到隔热层作用, 因此这些变化可能引起冻融格局发生改变, 比如冻融循环时间、强度以及频率都会增加(杨开军等, 2017), 这种改变可能会加剧土壤N2O累积排放量, 从而加剧全球气候温室效应和臭氧层破坏. ...
Soil-atmosphere exchange potential of NO and N2O in different land use types of Inner Mongolia as affected by soil temperature, soil moisture, freeze-thaw, and drying- wetting events 0 2010
Reduction of nitrate to ammonium in selected paddy soils of China 2 1998
... 冻融循环是由于季节或者昼夜温差引起的反复冻结和融化过程.这种现象普遍存在于自然界中, 并广泛发生在北半球高海拔、高纬度和一些温带地区, 例如, 在中国30° N以北地区中有46.3%的区域受到季节性冻融循环的影响(陈哲等, 2016).前人通过综述和整合分析等已经证实, 冻融循环可显著增加土壤N2O排放(Henry, 2007; Matzner & Borken, 2008; Song et al., 2017; Gao et al., 2018).冻融循环作用主要通过下面几个过程来显著提高土壤可利用养分含量: 土壤结构破坏可释放养分(Xiao et al., 2019); 土壤有机或无机胶体表面吸附的养分释放(Freppaz et al., 2007); 死亡的微生物可释放养分(Yanai et al., 2004); 死亡的根系可释放养分(Campbell et al., 2014; Reinmann & Templer, 2016); 凋落物分解释放养分(Wieder et al., 2011; Pelster et al., 2013).这些由于冻融循环作用增加的可利用养分可促进土壤氮转化过程, 比如硝化和反硝化作用等, 在这些氮转过程中会产生N2O, 从而增加了土壤N2O排放(Dai et al., 2020).除了硝化和反硝化外, 硝酸盐的异化还原作用也可以产生N2O, 研究得出, 可利用碳是调节硝态氮含量在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).在冻融循环过程中, 碳、氮养分的变化会不会影响N2O产生过程, 需要进一步证实.另外, 在研究冻融循环对土壤N2O排放中所采用的试验方法不尽相同, 比如采用室内模拟方法或室外原位研究方法以及在室内研究中采集土壤时间和处理方式以及冻融循环过程中是否有植物的存在等都有所差异, 这些试验方法和设置的差异究竟会给冻融循环期间N2O排放带来怎样的影响, 仍需进一步探究.在不同地区研究冻融循环的时候, 其所处的水热条件是不同的, 土壤结构、养分含量以及微生物活性和种类也是不同的.例如, 与年平均气温较高的温带地区相比, 苔原和高山草甸地区土壤微生物对冻融循环的响应较高(Gao et al., 2018), 这可能是因为在苔原和高山草甸等地区年平均气温较低, 土壤微生物对低温的适应性较强(Grogan et al., 2004).土壤本身基本理化及微生物特性差异可能也会影响土壤N2O对冻融循环响应的大小.然而, 前人研究缺乏这些因素对N2O排放的影响的综合考虑. ...
... 除了研究区域间的水热条件外, 土壤本身的理化性质也会影响土壤N2O排放对冻融循环的响应.本研究结果显示, 土壤N2O通量的效应值与土壤黏粒含量呈正比, 而与土壤砂粒含量呈反比(图5).原因可能主要归功于两个方面: 一方面, 在特定的土壤温度下, 黏粒含量较高的土壤中所含液态水含量较高.例如, 在土壤温度为-5 ℃时, 沙土中液体水含量可忽略不计, 而黏土中液体水可达0.15 cm3·cm-3 (Congreves et al., 2018).这种液态水的存在可为微生物在土壤冻结条件下继续保持一定的活性, 从而产生N2O气体.另一方面, 黏粒含量高的土壤, 其养分含量也高, 这就为N2O的产生提供充足底物.这一点也被本研究进一步证实, 即土壤硝态氮含量越高, N2O通量的效应值越大(图5).有趣的是, 本研究得出N2O排放的效应值只与硝态氮含量呈显著正相关关系, 而与铵态氮等其他有机、无机养分含量无显著相关性, 主要原因是与冻融循环期间N2O产生途径有关.土壤N2O气体产生途径包括硝化细菌的硝化作用、硝化细菌的反硝化、反硝化作用、联合反硝化作用和异化还原作用(Sánchez- García et al., 2014; Hu et al., 2015).在冻融循环期间, 前人通过同位素标记技术(Ludwig et al., 2004; Wagner-Riddle et al., 2010), 控制氧分压(Öquist et al., 2004), 乙炔抑制(Priemé & Christensen, 2001; Mørkved et al., 2006)等方法均证实了反硝化作用是冻融循环期间N2O显著增加的主要贡献过程, 其过程产生的N2O可占总排放量的80%以上.主要原因是由于雪被的融化导致冻融循环期间水分较高, 当土壤湿度超过75%时, 土壤以反硝化作用为主(Seitzinger et al., 2006), 硝态氮是反硝化作用的重要底物之一, 因此土壤N2O排放的效应值与硝态氮含量表现出显著正相关关系.另外, 反硝化的异化还原过程也可产生N2O, 可利用碳是调节硝态氮在反硝化和异化还原过程分配的关键因子(Fazzolari et al., 1998), 可溶性碳和硝态氮含量比值大于12时, 才会有更多的硝酸盐的异化还原作用发生(Yin et al., 1998).前人通过整合分析也发现, 冻融循环作用可显著增加土壤可溶性有机碳含量(Song et al., 2017), 有机碳源通过分解产生的电子被硝酸根或者亚硝酸根接受还原为N2O和N2.因此, 可溶性碳和硝态氮含量共同决定了N2O的排放.然而, 本研究并没有发现可溶性有机碳和可溶性有机碳含量与硝态氮含量比值与N2O排放有明显的相关性(附件I), 可能是相关数据报道较少限制了揭示可溶性有机碳含量与N2O排放之间的相关性或者可能是反硝化的异化还原过程产生的N2O在冻融循环过程中的贡献较小. ...
Freeze-thaw effects on sorption/desorption of dissolved organic carbon in wetland soils 2 2010
... 除了土壤本身的性质外, 冻融循环格局不同也可能会影响N2O对冻融循环的响应.前人研究结果(Oztas & Fayetorbay, 2003; Zhou et al., 2011)显示, 冻结强度增加, 可提高其对微生物和土壤结构的影响, 从而释放更多养分, 进而增加土壤N2O排放.随着冻融循环频率增加和冻融循环时间的增加, 土壤可利用养分可能会通过以下方式被降低: 养分被微生物不断消耗, 养分的淋洗损失, 养分被土壤颗粒重吸附(Grogan et al., 2004; Yu et al., 2010; Han e al., 2018; Xiao et al., 2019).这些可利用养分的减少会导致N2O对冻融循环的响应随之下降.相比于N2O排放通量, 冻融循环期间N2O累积排放量的生态学意义更大.然而前人的研究都集中在N2O排放通量, 并没有探究N2O累积排放量与冻融循环格局变化之间的关系.未来气候变化背景下, 冻融循环格局也会发生相应改变, 比如冻融时间延长, 冻融循环的强度和频率增加等, 那么这些变化会给N2O累积排放量产生什么样的影响? 前人研究并不足. ...
... 冻融循环格局的改变也会显著影响土壤N2O对冻融循环响应的大小.土壤融化时间与N2O排放呈显著正相关关系(图7).随着土壤融化时间的增加, 反硝化细菌活性逐渐恢复, 从而可能增加反硝化强度, 进而增加N2O产生与排放.随着冻结温度降低, 土壤N2O排放的效应值随之增加(图7).当冻结温度低于-10 ℃时, 土壤N2O排放通量要高于冻结温度大于-10 ℃时(图3).主要原因是冻结强度较高会增加土壤团聚体结构和微生物破坏(Fitzhugh et al., 2001; Oztas & Fayetorbay, 2003; Zhou et al., 2011), 从而促进了可溶性有机碳和无机氮的释放, 这些可溶性碳、氮含量的增加可促进硝化作用和反硝化作用速率, 进而增加了土壤N2O产生.另外, 本研究结果表明, 冻融循环频率与土壤N2O通量呈显著负相关关系(图7).随着冻融循环频率增加, 土壤养分可通过微生物利用、淋洗损失以及土壤颗粒重吸附等方式降低(Grogan et al., 2004; Yu et al., 2010; Han et al., 2018; Xiao et al., 2019), 使氮转化过程减慢, 从而导致N2O排放通量随之减少.有趣的是, 本研究发现冻融循环频率与土壤N2O累积排放量反而呈显著正相关关系(图7).虽然随着冻融循环频率增加, N2O通量随之降低, 这种降低是相对于第一次的N2O通量值, 而相对于对照组, N2O通量仍然是增加的.只是随着冻融循环频率增加, 这种增加幅度在降低.因此, 土壤N2O累积排放量会随着冻融循环频率增加而增加.在未来气候变暖背景下, 可能会减少雪被厚度和雪被存在时间(Groffman et al., 2011), 由于雪被起到隔热层作用, 因此这些变化可能引起冻融格局发生改变, 比如冻融循环时间、强度以及频率都会增加(杨开军等, 2017), 这种改变可能会加剧土壤N2O累积排放量, 从而加剧全球气候温室效应和臭氧层破坏. ...
Comparison of seasonal soil microbial process in snow-covered temperate ecosystems of northern China 3 2014
... 在室内研究中, 土壤采样的时间和土壤的处理方式也会影响N2O排放对冻融循环响应的大小(图3).比如, 与非冻融循环期间采集的土壤相比, 在冻融循环期间采集的土壤N2O排放效应值较大.原因可能与微生物特性和养分含量有关.在冻融与非冻融期间土壤微生物群落结构和微生物适应性是不同的(Zhang et al., 2014).在秋末至初冬, 土壤反硝化细菌的数量明显高于夏季(Mergel et al., 2001).由于冻融循环期间土壤N2O的产生主要是通过反硝化作用, 因此反硝化细菌数量增多可提高反硝化作用速率, 从而增加N2O产生的量.相比于夏季, 冬季土壤微生物对于温度的适应性较强(Zhang et al., 2014), 在经历温度的降低和升高时, 仍然能保持较高的微生物活性, 从而提高N2O的产生.另外, 土壤微生物量往往是冬季大于夏季(Alvarez et al., 1995; Zhang et al., 2014), 在冻融循环过程中微生物死亡可释放更多可利用性养分, 这就为冻融循环过程中N2O的产生提供充足的底物.除了采集土壤的时间外, 土壤处理方式也会影响N2O排放对于冻融循环的响应.本研究得出, 与土壤不过筛相比, 土壤过筛会显著增加N2O排放对冻融循环的响应(图3).主要原因是过筛会破坏土壤结构.一方面土壤团聚体晶格结构的破坏会释放更多可利用性养分进入土壤(Xiao et al., 2019), 另一方面土壤过筛也会导致土壤胶体表面吸附的养分离子释放出来(Freppaz et al., 2007), 这些可利用性养分的增加可为N2O产生提供充足的底物.在非冻融循环期间采集土壤和土壤过筛处理可能会分别导致冻融循环效应被低估和高估, 因此为了得到真实的数据和探究其真正地内在机制, 今后在室内研究冻融循环试验时, 应尽量模拟原位条件.比如在真正野外冻融循环期间采集土壤样品以及野外采集的土壤样品不要过筛. ...
... ., 2014), 在经历温度的降低和升高时, 仍然能保持较高的微生物活性, 从而提高N2O的产生.另外, 土壤微生物量往往是冬季大于夏季(Alvarez et al., 1995; Zhang et al., 2014), 在冻融循环过程中微生物死亡可释放更多可利用性养分, 这就为冻融循环过程中N2O的产生提供充足的底物.除了采集土壤的时间外, 土壤处理方式也会影响N2O排放对于冻融循环的响应.本研究得出, 与土壤不过筛相比, 土壤过筛会显著增加N2O排放对冻融循环的响应(图3).主要原因是过筛会破坏土壤结构.一方面土壤团聚体晶格结构的破坏会释放更多可利用性养分进入土壤(Xiao et al., 2019), 另一方面土壤过筛也会导致土壤胶体表面吸附的养分离子释放出来(Freppaz et al., 2007), 这些可利用性养分的增加可为N2O产生提供充足的底物.在非冻融循环期间采集土壤和土壤过筛处理可能会分别导致冻融循环效应被低估和高估, 因此为了得到真实的数据和探究其真正地内在机制, 今后在室内研究冻融循环试验时, 应尽量模拟原位条件.比如在真正野外冻融循环期间采集土壤样品以及野外采集的土壤样品不要过筛. ...
... ., 2014), 在冻融循环过程中微生物死亡可释放更多可利用性养分, 这就为冻融循环过程中N2O的产生提供充足的底物.除了采集土壤的时间外, 土壤处理方式也会影响N2O排放对于冻融循环的响应.本研究得出, 与土壤不过筛相比, 土壤过筛会显著增加N2O排放对冻融循环的响应(图3).主要原因是过筛会破坏土壤结构.一方面土壤团聚体晶格结构的破坏会释放更多可利用性养分进入土壤(Xiao et al., 2019), 另一方面土壤过筛也会导致土壤胶体表面吸附的养分离子释放出来(Freppaz et al., 2007), 这些可利用性养分的增加可为N2O产生提供充足的底物.在非冻融循环期间采集土壤和土壤过筛处理可能会分别导致冻融循环效应被低估和高估, 因此为了得到真实的数据和探究其真正地内在机制, 今后在室内研究冻融循环试验时, 应尽量模拟原位条件.比如在真正野外冻融循环期间采集土壤样品以及野外采集的土壤样品不要过筛. ...
Effect of freezing-thawing on nitrogen mineralization in vegetation soils of four landscape zones of Changbai Mountain 2 2011
... 除了土壤本身的性质外, 冻融循环格局不同也可能会影响N2O对冻融循环的响应.前人研究结果(Oztas & Fayetorbay, 2003; Zhou et al., 2011)显示, 冻结强度增加, 可提高其对微生物和土壤结构的影响, 从而释放更多养分, 进而增加土壤N2O排放.随着冻融循环频率增加和冻融循环时间的增加, 土壤可利用养分可能会通过以下方式被降低: 养分被微生物不断消耗, 养分的淋洗损失, 养分被土壤颗粒重吸附(Grogan et al., 2004; Yu et al., 2010; Han e al., 2018; Xiao et al., 2019).这些可利用养分的减少会导致N2O对冻融循环的响应随之下降.相比于N2O排放通量, 冻融循环期间N2O累积排放量的生态学意义更大.然而前人的研究都集中在N2O排放通量, 并没有探究N2O累积排放量与冻融循环格局变化之间的关系.未来气候变化背景下, 冻融循环格局也会发生相应改变, 比如冻融时间延长, 冻融循环的强度和频率增加等, 那么这些变化会给N2O累积排放量产生什么样的影响? 前人研究并不足. ...
... 冻融循环格局的改变也会显著影响土壤N2O对冻融循环响应的大小.土壤融化时间与N2O排放呈显著正相关关系(图7).随着土壤融化时间的增加, 反硝化细菌活性逐渐恢复, 从而可能增加反硝化强度, 进而增加N2O产生与排放.随着冻结温度降低, 土壤N2O排放的效应值随之增加(图7).当冻结温度低于-10 ℃时, 土壤N2O排放通量要高于冻结温度大于-10 ℃时(图3).主要原因是冻结强度较高会增加土壤团聚体结构和微生物破坏(Fitzhugh et al., 2001; Oztas & Fayetorbay, 2003; Zhou et al., 2011), 从而促进了可溶性有机碳和无机氮的释放, 这些可溶性碳、氮含量的增加可促进硝化作用和反硝化作用速率, 进而增加了土壤N2O产生.另外, 本研究结果表明, 冻融循环频率与土壤N2O通量呈显著负相关关系(图7).随着冻融循环频率增加, 土壤养分可通过微生物利用、淋洗损失以及土壤颗粒重吸附等方式降低(Grogan et al., 2004; Yu et al., 2010; Han et al., 2018; Xiao et al., 2019), 使氮转化过程减慢, 从而导致N2O排放通量随之减少.有趣的是, 本研究发现冻融循环频率与土壤N2O累积排放量反而呈显著正相关关系(图7).虽然随着冻融循环频率增加, N2O通量随之降低, 这种降低是相对于第一次的N2O通量值, 而相对于对照组, N2O通量仍然是增加的.只是随着冻融循环频率增加, 这种增加幅度在降低.因此, 土壤N2O累积排放量会随着冻融循环频率增加而增加.在未来气候变暖背景下, 可能会减少雪被厚度和雪被存在时间(Groffman et al., 2011), 由于雪被起到隔热层作用, 因此这些变化可能引起冻融格局发生改变, 比如冻融循环时间、强度以及频率都会增加(杨开军等, 2017), 这种改变可能会加剧土壤N2O累积排放量, 从而加剧全球气候温室效应和臭氧层破坏. ...
 ,, 白娥
,, 白娥 ,*
,*

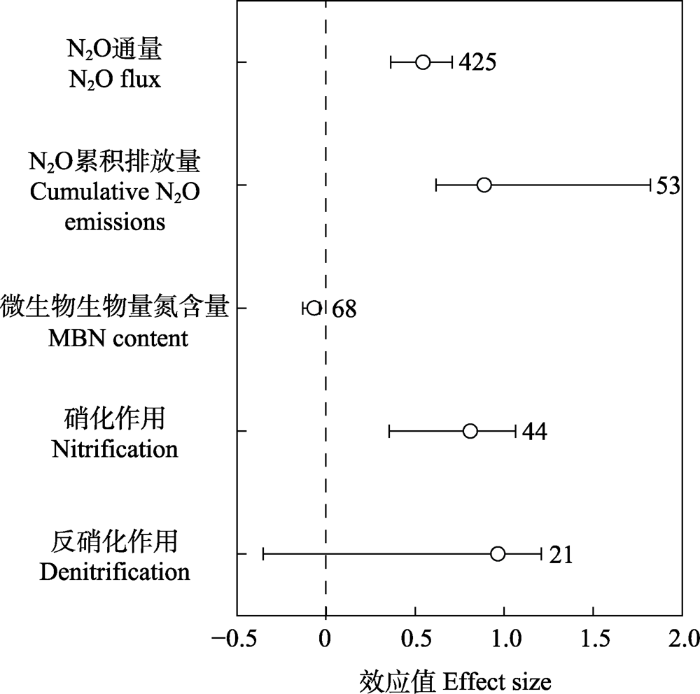 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT
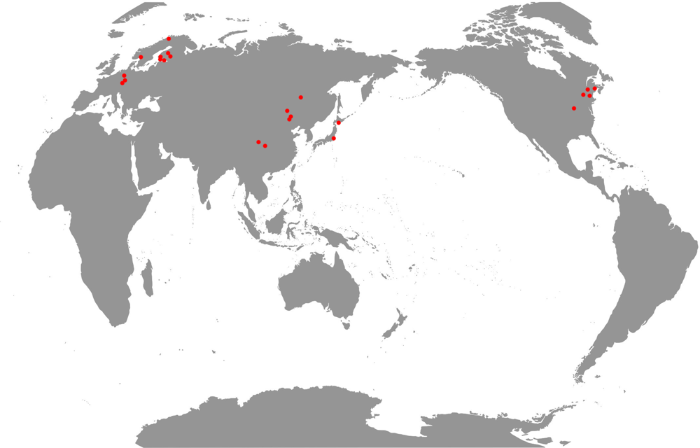 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT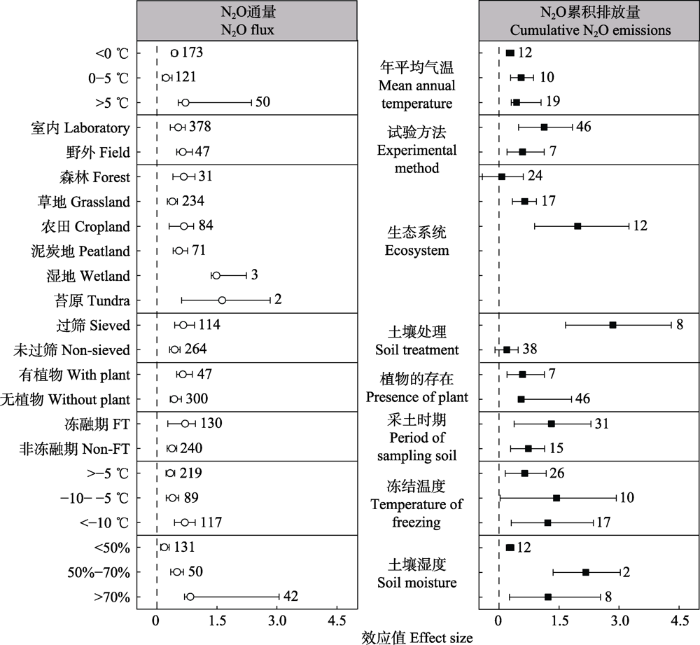 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT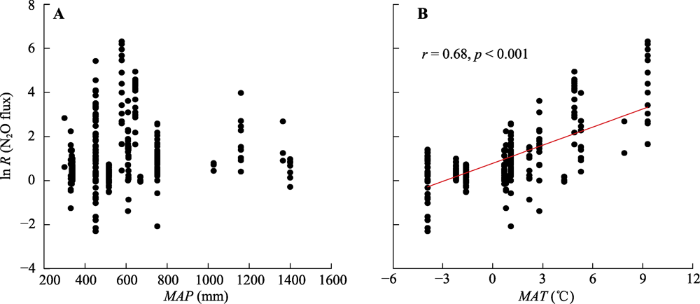 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT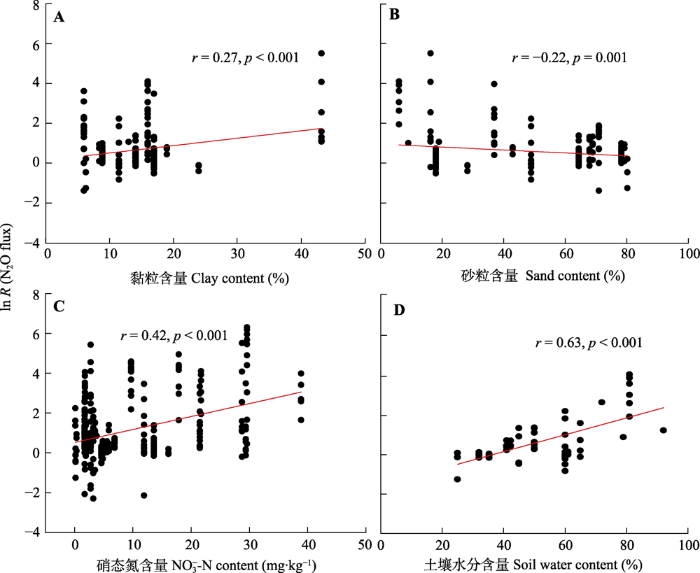 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT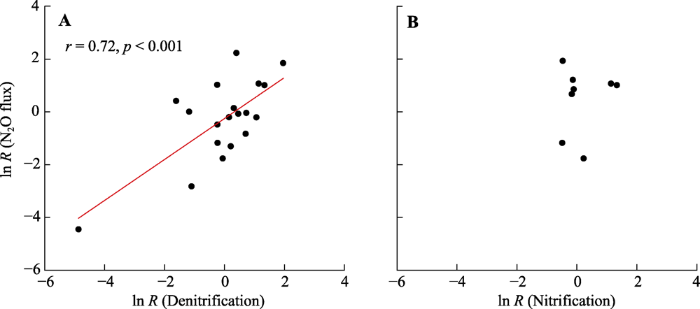 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT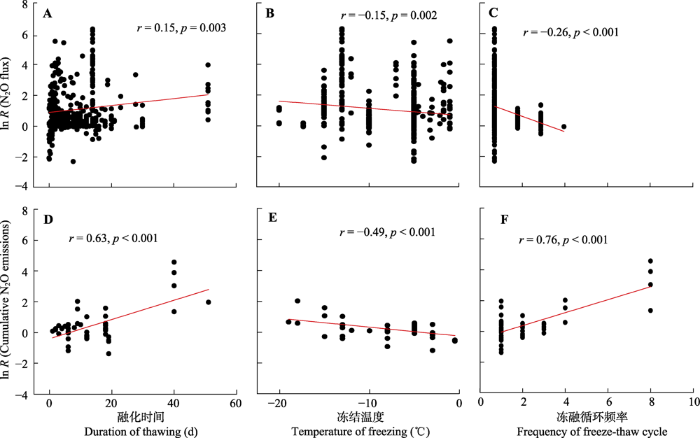 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}