Fundamental Research Funds for the Central Universities(2572018AA07) National Key R&D Program of China(2016YFD0600201) Program for Changjiang Scholars and Innovative Research Team in University(IRT_15R09)
Abstract Drought-related tree mortality profoundly impacts the ecosystem functions and carbon budgets, in which one of the principal mechanisms involved is the catastrophic failure of the hydraulic systems. However, our understanding of tree hydraulic systems and the mechanisms of tree death under extreme drought conditions are limited because the responses of trees to drought stress are multi-dimensional and complex. In this review, we first expounded the indexes of measuring plant drought resistance, and focused on the stomatal safety margin (SSM) that can be used to comprehensively evaluate the drought tolerance of plants. A larger positive value of SSM indicates a stronger coordination between stomata and hydraulic traits, a lower possibility of xylem embolization, and a more conservative hydraulic strategy adopted. Second, we integrated general response processes of woody plants to drought stress. Third, we introduced response mechanisms of different plant organs (leaf, stem and root) to drought stress. The probability of reaching the critical threshold and the duration of tree death are determined by interactions between physiological and morphological traits. Finally, we discussed hydraulic recovery mechanisms of woody plants, and put forward three research priorities in the future: (1) to improve the methodology for measuring leaf hydraulic conductance, especially the xylem and outside-xylem hydraulic conductance, and quantify the relative contributions of the four water transport pathways in mesophyll tissues; (2) to quantify variations in the epidermal permeability for better understanding plant water-use strategies; and (3) to deepen the understanding of the water-carbon coupling mechanisms, and link individual-level structural and physiological traits with patterns and processes at the community and landscape levels, so as to better assessing and monitoring the potential risk of drought-induced tree mortality. Keywords:drought resistance;xylem embolism;stomatal adjustment;tree mortality;hydraulic trait
随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019)。其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017)。土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率。因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016)。然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018)。因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018)。
干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017)。但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011)。大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011)。因此, 植物对干旱具有复杂的多维响应机制。鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018)。因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用。
1 衡量植物抗旱性的水力指标
植物维持自身水力系统完整性的能力, 主要取决于其维管系统的物理极限以及植物维持自身水势的能力, 进而决定了植物在干旱胁迫期间的脱水速度以及相应的死亡阈值(Choat et al., 2018)。叶膨压损失点水势(ΨTLP), 通常被用以直接量化植物叶片的耐旱性, 因为细胞膨压损失影响了细胞结构完整性、代谢和整个植物的性能, 是衡量植物生理耐旱性的经典指标。低ΨTLP的植物, 可以在较低叶水势(Ψleaf)的情况下仍然维持叶片膨压及其功能, 如光合作用和生长。此外, ΨTLP是与物种的相对等水程度关系最为密切的一个性状, ΨTLP越高, 植物越趋向于等水调节。Meinzer等(2016)建议使用ΨTLP作为衡量等水调节程度的指标, 并已经被大量物种的研究所支持(Fu & Meinzer, 2019; Li et al., 2019)。Bartlett等(2016)发现ΨTLP与叶和枝水力导度下降50%的水势(P50)呈正相关关系。Johnson等(2018)发现, 在干旱季节中, 非等水的物种能够更有效地调节其ΨTLP。这种ΨTLP的变化也与叶P50的下降有关。
在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率。随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988)。因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012)。当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损。在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012)。P50和P88值越负, 木质部抵抗栓塞能力越强。研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012)。空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势。然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021)。研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017)。但在相似的WD下, 裸子植物的致死水势要比被子植物更低。这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021)。目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性。
单一的指标无法准确表征植物在干旱胁迫下的生存力。例如, 对美国加州南部灌木的研究发现, 具有高栓塞抗性的浅根系灌木在干旱胁迫下的死亡率较高, 而木质部栓塞脆弱性高的深根物种死亡水平则较低(Pivovaroff et al., 2018), 这表明空穴化脆弱性和干旱在某些情况下是解耦的(Blackman et al., 2012)。这种解耦现象是由于某些植物的避旱策略所致, 例如较深的根系、干旱落叶和气孔调节等。由于植物对空穴化的脆弱性不能决定其自身的在干旱环境下的生存能力, 因此, 近期通过水力安全阈值(HSM)来预测物种死亡率(Anderegg et al., 2016; Adams et al., 2017)。HSM是指一个物种在自然条件下经历的最小水势(Ψmin)与引起木质部功能障碍的水势(如P50或P88)之间的差值, 用来描述植物水力策略的保守程度(Skelton et al., 2015), 其值越大, 表明植物经历栓塞的可能性越低, 意味着越安全(Choat et al.,2012)。这里的Ψmin被假定为植物气孔调节对Ψx的影响。植物通过气孔的开闭来限制蒸腾速率、维持水势相对恒定, 是一种重要的水力调节策略。因此, 干旱胁迫下的气孔响应被用来指示植物的抗旱性。但是, 事实上Ψmin集成体现了植物结构特征(如根深)和生理特征(例如气孔行为)对环境胁迫的多重响应(Choat et al., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019)。
为了直接评估干旱胁迫下的气孔响应, Skelton等(2015)通过拟合气孔导度(gs)随Ψx变化的响应曲线来定义标准化的临界点。其中, 最大气孔导度下降50%时的切线与Ψx轴的交点被定义为气孔闭合点, 且该点对应的gs约损失了最大气孔导度的88%, 因此Ψgs88被定义为气孔关闭所对应的水势, 而Ψgs88与P50 (或P88)的差值被称为气孔安全阈值(SSM), 用于评价气孔和水力之间的协调能力(Skelton et al., 2015)。该值越高, 表明植物气孔和水力之间的协调能力越强(Creek et al., 2018)。有研究用SSM50 (Ψgs88与P50之差)作为指示植物等水调节程度的指标, 该值为正, 表明气孔关闭发生在水力导度下降到50%之前, 定义为等水调节行为; 而该值为负时则表明发生在P50之后, 定义为非等水调节行为。并认为等水调节物种比非等水调节物种具有更大的水力和气孔安全阈值(Skelton et al., 2015)。此外, 有研究表明, 只有少数物种气孔关闭与木质部栓塞形成初期的水势相等, 且随植物栓塞抗性的增加, SSM增加(Martin-Stpaul et al., 2017)。Chen等(2019)在对温带阔叶树种死亡率预测的研究中, 发现相比空穴化脆弱性(P50和P88)和HSM, SSM能更好地预测干旱胁迫下植物的死亡风险。但该指标目前使用并不广泛, 需要更多的研究加以验证。
2 整株植物对干旱胁迫的水力响应过程
植物对干旱胁迫的响应过程主要分为两个阶段(图1)。第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017)。在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降。植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017)。随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度。气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019)。在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014)。尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018)。
Fig. 1Phases of drought response to drought stress in plants. With the increase of drought stress, the dotted curve represents the trend of stomatal and cuticular conductance, while the solid curve represents the loss of hydraulic conductance. Ψgs88, the water potential at 88% loss of stomatal conductance; P50 and P88, the water potential at 50% and 88% loss of hydraulic function, respectively; SSM50 and SSM88, the margins between Ψgs88 and P50 or P88, respectively.
气孔关闭后, 水仍会通过气孔渗漏, 以及表皮和树皮等其他组织流失, 使植物水势持续缓慢下降。而植物也会释放内部储存水来进一步缓冲Ψx的下降。与此同时, 植物水力导度会通过一系列生物物理和生理机制的改变而下降, 包括叶脉塌缩、植物水通道蛋白调节、细根皮层腔隙的形成等(Choat et al., 2018)。当水势下降到一个临界点(P50)时, 栓塞便会在木质部中迅速增长, 这一过程会发生在包括根茎叶在内的整个植物水力系统中(Creek et al., 2018; Rodriguez-Dominguez et al., 2018)。由于气孔长期关闭, 光合作用受到抑制, 而呼吸作用还在继续, 从而消耗大量储存的碳水化合物。此外, 植物需要利用碳水化合物来驱动韧皮部运输、维持膨压, 以及重新填充栓塞的木质部, 因此维持呼吸所需的碳水化合物进一步减少。而水和碳的限制又逐渐增加了木质部栓塞的比例, 限制了对冠层的水分输送, 导致分枝斑块性死亡, 冠层叶片面积显著减少(McDowell et al., 2011)。随着栓塞逐渐遍布整个输水网络, 造成植物水力系统不可逆的损伤(P88), 最终导致整个植株死亡(Anderegg et al., 2012; Choat et al., 2018)。事实上, 在干旱胁迫下, 植物达到死亡临界阈值的概率和时间最终取决于相关生理和形态学特征的相互作用(图2)。这种相互作用的多维性是维管植物能够生长在地球上几乎所有的陆地生态系统的有效策略, 并为抗旱提供了大量可能的形态和生理解决方案。
不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同。树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低。在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008)。有两种假设解释了植物的这种保护机制。一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018)。而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降。两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018)。此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角。
有研究表明, 物种的脆弱性分割程度受气候、树种等的影响。在半干旱、温带和季节性干旱环境中, 植物具有较强的脆弱性分割现象(Johnson et al., 2016), 而在湿润环境中缺少该机制(Zhu et al., 2016)。Jin等(2019)在自然条件下的研究发现, 温带森林中针叶树种存在脆弱性分割机制, 而被子植物中不存在; 但是我们进一步的干旱控制试验研究发现, 温带阔叶树种水曲柳(Fraxinus mandshurica)存在脆弱性分割现象(未发表数据); 这可能与试验环境条件有关。在干旱环境下, 脆弱性分割有利于树木长期生存。
此外, 叶、茎和根性状的协调, 决定树木应对干旱胁迫的水力策略和耐受范围。Creek等(2018)发现, 导致Eucalyptus coolabah叶片气孔关闭的水势与根水力下降水势相吻合, 表明其中可能存在一种与气孔关闭相关的水力信号。López等(2021)的研究表明, 叶和茎的耐旱性既关联又相对独立, 其功能权衡取决于系统发育阶段和水分有效性。其中, 在茎叶性状变异轴上, 一端是具有较低的栓塞脆弱性、较高的水分利用效率和胡伯尔值(Hv, 茎边材面积与叶面积之比)以及气孔关闭较晚的“慢”对策物种; 而另一端是具有高栓塞脆弱性、低水分利用效率和Hv的“快”对策物种。这种生存对策的差异是由区域水分有效性所致。此外, 植物性状变异是多维的(Powell et al., 2017), 不同的维度允许不同功能性状组合, 因而解释了一些功能性状权衡未能反映在较低系统发育水平上的原因(Pivovaroff et al., 2014; López et al., 2021)。
4 水力恢复机制
许多水力安全阈值较窄的木本植物经常处于接近栓塞临界阈值的水势下, 当可利用水分增加时, 其水势和水分运输能力会迅速恢复(Ogasa et al., 2013), 表明很多植物具有水力恢复策略(Trifilò et al., 2015; Klein et al., 2018)。有研究表明, 高水力风险的物种具有较高的水力恢复能力, 而水力对策较保守的物种可能并不依赖水力恢复对策(Ogasa et al., 2013)。因此Klein等(2018)将恢复力作为一个额外的维度, 加入“安全-效率”权衡中, 形成一个新的“安全-效率-恢复”框架。了解在干旱胁迫下存活树木的水力恢复过程同样重要。
在未致死的前提下, 当干旱造成植物相对严重的水力功能障碍时, 水力恢复可通过两种机制进行。一是形成新木质部, 二是木质部栓塞的修复。木质部的再生似乎是树木在干旱后恢复水力能力的主要途径(Brodribb et al., 2010; Creek et al., 2018)。通过维管束形成层的活动, 在木质部外部区域形成新的导管(或管胞), 以替代之前被树胶或侵填体永久堵塞并失去导水能力的部分。而在干旱造成植物地上部分大量死亡的情况下, 可通过芽或木质块茎的重新萌生促进恢复。在热带和温带森林物种中, 木质部再生的发生程度是高度可变的, 最终取决于植物分生组织的存活程度(Zeppel et al., 2015)。对于栓塞修复, 某些植物可通过根压在木质部中形成“正压”以重填栓塞木质部的观点已被普遍接受(Nardini et al., 2018); 而关于在负压下木质部再充水的机制以及用于检测和量化的方法仍然存在争论。
4.1 叶表面吸水
植物受干旱影响后的水力恢复速率在很大程度上取决于其水势下降的程度, 以及经历极限低水势的时间。轻度干旱胁迫通常不会导致高水平空穴化栓塞, 但叶脉中的导管塌缩和水通道蛋白对细胞膜通透性的调节机制可能导致植物水力运输能力瞬时且可逆的下降(Zhang et al., 2016)。此时, 水力恢复会在降水后通过叶表面吸水(FWU)在很短的时间内发生(Binks et al., 2019; Fuenzalida et al., 2019)。FWU是一种几乎在所有植物中都存在的水分获取机制(Dawson & Goldsmith, 2018; Berry et al., 2019a)。
叶片吸水以液相和气相两种形式出现。如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017)。由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020)。当气孔关闭时, 表皮是FWU的主要途径。不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a)。通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动。在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015)。对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生。之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中。Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水。Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况。但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H)。此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a)。Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释。因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高。但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU。Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱。此外, Cleaf也会改变FWU的持续时间。在FWU条件下, Cleaf会影响水势的增长。高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的。Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压。而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险。但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019)。
AllenCD, BreshearsDD, McDowellNG (2015). On under- estimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene Ecosphere, 6, 1-55. [本文引用: 1]
AndereggWRL, BerryJA, SmithDD, SperryJS, AndereggLDL, FieldCB (2012). The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off Proceedings of the National Academy of Sciences of the United States of America, 109, 233-237. DOI:10.1073/pnas.1107891109PMID:22167807 [本文引用: 2] Forest ecosystems store approximately 45% of the carbon found in terrestrial ecosystems, but they are sensitive to climate-induced dieback. Forest die-off constitutes a large uncertainty in projections of climate impacts on terrestrial ecosystems, climate-ecosystem interactions, and carbon-cycle feedbacks. Current understanding of the physiological mechanisms mediating climate-induced forest mortality limits the ability to model or project these threshold events. We report here a direct and in situ study of the mechanisms underlying recent widespread and climate-induced trembling aspen (Populus tremuloides) forest mortality in western North America. We find substantial evidence of hydraulic failure of roots and branches linked to landscape patterns of canopy and root mortality in this species. On the contrary, we find no evidence that drought stress led to depletion of carbohydrate reserves. Our results illuminate proximate mechanisms underpinning recent aspen forest mortality and provide guidance for understanding and projecting forest die-offs under climate change.
AndereggWRL, KleinT, BartlettM, SackL, PellegriniAF, ChoatB, JansenS (2016). Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought- induced tree mortality across the globe Proceedings of the National Academy of Sciences of the United States of America, 113, 5024-5029. DOI:10.1073/pnas.1525678113PMID:27091965 [本文引用: 3] Drought-induced tree mortality has been observed globally and is expected to increase under climate change scenarios, with large potential consequences for the terrestrial carbon sink. Predicting mortality across species is crucial for assessing the effects of climate extremes on forest community biodiversity, composition, and carbon sequestration. However, the physiological traits associated with elevated risk of mortality in diverse ecosystems remain unknown, although these traits could greatly improve understanding and prediction of tree mortality in forests. We performed a meta-analysis on species' mortality rates across 475 species from 33 studies around the globe to assess which traits determine a species' mortality risk. We found that species-specific mortality anomalies from community mortality rate in a given drought were associated with plant hydraulic traits. Across all species, mortality was best predicted by a low hydraulic safety margin-the difference between typical minimum xylem water potential and that causing xylem dysfunction-and xylem vulnerability to embolism. Angiosperms and gymnosperms experienced roughly equal mortality risks. Our results provide broad support for the hypothesis that hydraulic traits capture key mechanisms determining tree death and highlight that physiological traits can improve vegetation model prediction of tree mortality during climate extremes.
ÁvilaE, HerreraA, TezaraW (2014). Contribution of stem CO2 fixation to whole-plant carbon balance in nonsucculent species Photosynthetica, 52, 3-15. DOI:10.1007/s11099-014-0004-2URL [本文引用: 2]
BarnardDM, MeinzerFC, LachenbruchB, McCullohKA, JohnsonDM, WoodruffDR (2011). Climate-related trends in sapwood biophysical properties in two conifers: avoidance of hydraulic dysfunction through coordinated adjustments in xylem efficiency, safety and capacitance Plant, Cell & Environment, 34, 643-654. [本文引用: 1]
BartlettMK, KleinT, JansenS, ChoatB, SackL (2016). The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought Proceedings of the National Academy of Sciences of the United States of America, 113, 13098-13103. PMID:27807136 [本文引用: 2] Climate change is expected to exacerbate drought for many plants, making drought tolerance a key driver of species and ecosystem responses. Plant drought tolerance is determined by multiple traits, but the relationships among traits, either within individual plants or across species, have not been evaluated for general patterns across plant diversity. We synthesized the published data for stomatal closure, wilting, declines in hydraulic conductivity in the leaves, stems, and roots, and plant mortality for 262 woody angiosperm and 48 gymnosperm species. We evaluated the correlations among the drought tolerance traits across species, and the general sequence of water potential thresholds for these traits within individual plants. The trait correlations across species provide a framework for predicting plant responses to a wide range of water stress from one or two sampled traits, increasing the ability to rapidly characterize drought tolerance across diverse species. Analyzing these correlations also identified correlations among the leaf and stem hydraulic traits and the wilting point, or turgor loss point, beyond those expected from shared ancestry or independent associations with water stress alone. Further, on average, the angiosperm species generally exhibited a sequence of drought tolerance traits that is expected to limit severe tissue damage during drought, such as wilting and substantial stem embolism. This synthesis of the relationships among the drought tolerance traits provides crucial, empirically supported insight into representing variation in multiple traits in models of plant and ecosystem responses to drought.
BerryZC, EmeryNC, GotschSG, GoldsmithGR (2019a). Foliar water uptake: processes, pathways, and integration into plant water budgets Plant, Cell & Environment, 42, 410-423. [本文引用: 4]
BerryZC, EspejelX, Williams-LineraG, AsbjornsenH (2019b). Linking coordinated hydraulic traits to drought and recovery responses in a tropical montane cloud forest American Journal of Botany, 106, 1316-1326. DOI:10.1002/ajb2.v106.10URL [本文引用: 1]
BerryZC, WhiteJC, SmithWK (2014). Foliar uptake, carbon fluxes and water status are affected by the timing of daily fog in saplings from a threatened cloud forest Tree Physiology, 34, 459-470. DOI:10.1093/treephys/tpu032URL [本文引用: 1]
BinksO, CoughlinI, MencucciniM, MeirP (2020). Equivalence of foliar water uptake and stomatal conductance Plant, Cell & Environment, 43, 524-528. [本文引用: 1]
BinksO, MencucciniM, RowlandL, da CostaACL, de CarvalhoCJR, BittencourtP, EllerC, TeodoroGS, CarvalhoEJM, SozaA, FerreiraL, VasconcelosSS, OliveiraR, MeirP,(2019). Foliar water uptake in Amazonian trees: evidence and consequences Global Change Biology, 25, 2678-2690. DOI:10.1111/gcb.2019.25.issue-8URL [本文引用: 1]
BlackmanCJ, BrodribbTJ, JordanGJ (2012). Leaf hydraulic vulnerability influences species' bioclimatic limits in a diverse group of woody angiosperms Oecologia, 168, 1-10. DOI:10.1007/s00442-011-2064-3PMID:21744163 [本文引用: 1] The ability of plants to maintain water flow through leaves under water stress-induced tension (assessed as the leaf hydraulic vulnerability; P50(leaf)) is intimately linked with survival. We examined the significance of P50(leaf) as an adaptive trait in influencing the dry-end distributional limits of cool temperate woody angiosperm species. We also examined differences in within-site variability in P50(leaf) between two high-rainfall montane rainforest sites in Tasmania and Peru, respectively. A significant relationship between P50(leaf) and the 5th percentile of mean annual rainfall across each species distribution was found in Tasmania, suggesting that P50(leaf) influences species climatic limits. Furthermore, a strong correlation between P50(leaf) and the minimum rainfall availability was found using five phylogenetically independent species pairs in wet and dry evergreen tree species, suggesting that rainfall is an important selective agent in the evolution of leaf hydraulic vulnerability. Greater within-site variability in P50(leaf) was found among dominant montane rainforest species in Tasmania than in Peru and this result is discussed within the context of differences in spatial and temporal environmental heterogeneity and parochial historical ecology.
BrodersenCR, McElroneAJ (2013). Maintenance of xylem network transport capacity: a review of embolism repair in vascular plants Frontiers in Plant Science, 4, 108. DOI: 10.3389/fpls.2013.00108. PMID:23630539 [本文引用: 1] Maintenance of long distance water transport in xylem is essential to plant health and productivity. Both biotic and abiotic environmental conditions lead to embolism formation within the xylem resulting in lost transport capacity and ultimately death. Plants exhibit a variety of strategies to either prevent or restore hydraulic capacity through cavitation resistance with specialized anatomy, replacement of compromised conduits with new growth, and a metabolically active embolism repair mechanism. In recent years, mounting evidence suggests that metabolically active cells surrounding the xylem conduits in some, but not all, species are capable of restoring hydraulic conductivity. This review summarizes our current understanding of the osmotically driven embolism repair mechanism, the known genetic and anatomical components related to embolism repair, rehydration pathways through the xylem, and the role of capacitance. Anatomical differences between functional plant groups may be one of the limiting factors that allow some plants to refill while others do not, but further investigations are necessary to fully understand this dynamic process. Finally, xylem networks should no longer be considered an assemblage of dead, empty conduits, but instead a metabolically active tissue finely tuned to respond to ever changing environmental cues.
BrodribbTJ, CochardH (2009). Hydraulic failure defines the recovery and point of death in water-stressed conifers Plant Physiology, 149, 575-584. DOI:10.1104/pp.108.129783PMID:19011001 [本文引用: 1] This study combines existing hydraulic principles with recently developed methods for probing leaf hydraulic function to determine whether xylem physiology can explain the dynamic response of gas exchange both during drought and in the recovery phase after rewatering. Four conifer species from wet and dry forests were exposed to a range of water stresses by withholding water and then rewatering to observe the recovery process. During both phases midday transpiration and leaf water potential (Psileaf) were monitored. Stomatal responses to Psileaf were established for each species and these relationships used to evaluate whether the recovery of gas exchange after drought was limited by postembolism hydraulic repair in leaves. Furthermore, the timing of gas-exchange recovery was used to determine the maximum survivable water stress for each species and this index compared with data for both leaf and stem vulnerability to water-stress-induced dysfunction measured for each species. Recovery of gas exchange after water stress took between 1 and >100 d and during this period all species showed strong 1:1 conformity to a combined hydraulic-stomatal limitation model (r2 = 0.70 across all plants). Gas-exchange recovery time showed two distinct phases, a rapid overnight recovery in plants stressed to <50% loss of leaf hydraulic conductance (Kleaf) and a highly Psileaf-dependent phase in plants stressed to >50% loss of Kleaf. Maximum recoverable water stress (Psimin) corresponded to a 95% loss of Kleaf. Thus, we conclude that xylem hydraulics represents a direct limit to the drought tolerance of these conifer species.
BrodribbTJ, FeildTS, JordanGJ (2007). Leaf maximum photosynthetic rate and venation are linked by hydraulics Plant Physiology, 144, 1890-1898. PMID:17556506 [本文引用: 1] Leaf veins are almost ubiquitous across the range of terrestrial plant diversity, yet their influence on leaf photosynthetic performance remains uncertain. We show here that specific physical attributes of the vascular plumbing network are key limiters of the hydraulic and photosynthetic proficiency of any leaf. Following the logic that leaf veins evolved to bypass inefficient water transport through living mesophyll tissue, we examined the hydraulic pathway beyond the distal ends of the vein system as a possible limiter of water transport in leaves. We tested a mechanistic hypothesis that the length of this final traverse, as water moves from veins across the mesophyll to where it evaporates from the leaf, governs the hydraulic efficiency and photosynthetic carbon assimilation of any leaf. Sampling 43 species across the breadth of plant diversity from mosses to flowering plants, we found that the post-vein traverse as determined by characters such as vein density, leaf thickness, and cell shape, was strongly correlated with the hydraulic conductivity and maximum photosynthetic rate of foliage. The shape of this correlation provided clear support for the a priori hypothesis that vein positioning limits photosynthesis via its influence on leaf hydraulic efficiency.
BrodribbTJ, McAdamSAM (2011). Passive origins of stomatal control in vascular plants Science, 331, 582-585. DOI:10.1126/science.1197985PMID:21163966 [本文引用: 2] Carbon and water flow between plants and the atmosphere is regulated by the opening and closing of minute stomatal pores in surfaces of leaves. By changing the aperture of stomata, plants regulate water loss and photosynthetic carbon gain in response to many environmental stimuli, but stomatal movements cannot yet be reliably predicted. We found that the complexity that characterizes stomatal control in seed plants is absent in early-diverging vascular plant lineages. Lycophyte and fern stomata are shown to lack key responses to abscisic acid and epidermal cell turgor, making their behavior highly predictable. These results indicate that a fundamental transition from passive to active metabolic control of plant water balance occurred after the divergence of ferns about 360 million years ago.
BrodribbTJ, McAdamSAM (2017). Evolution of the stomatal regulation of plant water content Plant Physiology, 174, 639-649. DOI:10.1104/pp.17.00078PMID:28404725 [本文引用: 1]
BrodribbTJ, McAdamSAM, JordanGJ, MartinsSCV (2014). Conifer species adapt to low-rainfall climates by following one of two divergent pathways Proceedings of the National Academy of Sciences of the United States of America, 111, 14489-14493. DOI:10.1073/pnas.1407930111PMID:25246559 [本文引用: 1] Water stress is one of the primary selective forces in plant evolution. There are characters often cited as adaptations to water stress, but links between the function of these traits and adaptation to drying climates are tenuous. Here we combine distributional, climatic, and physiological evidence from 42 species of conifers to show that the evolution of drought resistance follows two distinct pathways, both involving the coordinated evolution of tissues regulating water supply (xylem) and water loss (stomatal pores) in leaves. Only species with very efficient stomatal closure, and hence low minimum rates of water loss, inhabit dry habitats, but species diverged in their apparent mechanism for maintaining closed stomata during drought. An ancestral mechanism found in Pinaceae and Araucariaceae species relies on high levels of the hormone abscisic acid (ABA) to close stomata during water stress. A second mechanism, found in the majority of Cupressaceae species, uses leaf desiccation rather than high ABA levels to close stomata during sustained water stress. Species in the latter group were characterized by xylem tissues with extreme resistance to embolism but low levels of foliar ABA after 30 d without water. The combination of low levels of ABA under stress with cavitation-resistant xylem enables these species to prolong stomatal opening during drought, potentially extending their photosynthetic activity between rainfall events. Our data demonstrate a surprising simplicity in the way conifers evolved to cope with water shortage, indicating a critical interaction between xylem and stomatal tissues during the process of evolution to dry climates.
CassanaFF, EllerCB, OliveiraRS, DillenburgLR (2016). Effects of soil water availability on foliar water uptake of Araucaria angustifolia. Plant and Soil, 399, 147-157. [本文引用: 1]
CernusakLA, CheesmanAW (2015). The benefits of recycling: How photosynthetic bark can increase drought tolerance New Phytologist, 208, 995-997. DOI:10.1111/nph.13723PMID:26536151 [本文引用: 2]
ChenWL, KoideRT, AdamsTS, DeForestJL, ChengL, EissenstatDM (2016). Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees Proceedings of the National Academy of Sciences of the United States of America, 113, 8741-8746. [本文引用: 1]
ChenZC, LiS, LuanJW, ZhangYT, ZhuSD, WanXC, LiuSR (2019). Prediction of temperate broadleaf tree species mortality in arid limestone habitats with stomatal safety margins Tree Physiology, 39, 1428-1437. DOI:10.1093/treephys/tpz045URL [本文引用: 2]
ChoatB, BrodribbTJ, BrodersenCR, DuursmaRA, LópezR, MedlynBE (2018). Triggers of tree mortality under drought Nature, 558, 531-539. DOI:10.1038/s41586-018-0240-xURL [本文引用: 8]
ChoatB, JansenS, BrodribbTJ, CochardH, DelzonS, BhaskarR, BucciSJ, FeildTS, GleasonSM, HackeUG, JacobsenAL, LensF, MaheraliH, Martínez-VilaltaJ, MayrS, et al. (2012). Global convergence in the vulnerability of forests to drought Nature, 491, 752-755. DOI:10.1038/nature11688URL [本文引用: 5]
ClarkJS, IversonL, WoodallCW, AllenCD, BellDM, BraggDC, D'AmatoAW, DavisFW, HershMH, IbanezI, JacksonST, MatthewsS, PedersonN, PetersM, SchwartzMWet al. (2016). The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States Global Change Biology, 22, 2329-2352. DOI:10.1111/gcb.2016.22.issue-7URL [本文引用: 1]
CreekD, BlackmanC, BrodribbTJ, ChoatB, TissueDT (2018). Coordination between leaf, stem and root hydraulics and gas exchange in three arid-zone angiosperms during severe drought and recovery Plant, Cell & Environment, 41, 2869- 2881. [本文引用: 3]
CreekD, LamarqueLJ, Torres-RuizJM, PariseC, BurlettR, TissueDT, DelzonS (2020). Xylem embolism in leaves does not occur with open stomata: evidence from direct observations using the optical visualization technique Journal of Experimental Botany, 71, 1151-1159. DOI:10.1093/jxb/erz474URL [本文引用: 1]
DeGuzman ME, Acosta-RangelA, WinterK, MeinzerFC, BonalD, SantiagoLS (2021). Hydraulic traits of Neotropical canopy liana and tree species across a broad range of wood density: implications for predicting drought mortality with models Tree Physiology, 41, 24-34. DOI:10.1093/treephys/tpaa106URL [本文引用: 1]
DeRoo L, SalomónRL, OleksynJ, SteppeK (2020a). Woody tissue photosynthesis delays drought stress in Populus tremula trees and maintains starch reserves in branch xylem tissues New Phytologist, 228, 70-81. DOI:10.1111/nph.v228.1URL [本文引用: 2]
DeRoo L, SalomónRL, SteppeK (2020b). Woody tissue photosynthesis reduces stem CO2 efflux by half and remains unaffected by drought stress in young Populus tremula trees Plant, Cell & Environment, 43, 981-991. [本文引用: 1]
DomecJC, WarrenJM, MeinzerFC, BrooksJR, CoulombeR (2004). Native root xylem embolism and stomatal closure in stands of Douglas-fir and ponderosa pine: mitigation by hydraulic redistribution Oecologia, 141, 7-16. PMID:15338263 [本文引用: 2] Hydraulic redistribution (HR), the passive movement of water via roots from moist to drier portions of the soil, occurs in many ecosystems, influencing both plant and ecosystem-water use. We examined the effects of HR on root hydraulic functioning during drought in young and old-growth Douglas-fir [ Pseudotsuga menziesii (Mirb.) Franco] and ponderosa pine ( Pinus ponderosa Dougl. Ex Laws) trees growing in four sites. During the 2002 growing season, in situ xylem embolism, water deficit and xylem vulnerability to embolism were measured on medium roots (2-4-mm diameter) collected at 20-30 cm depth. Soil water content and water potentials were monitored concurrently to determine the extent of HR. Additionally, the water potential and stomatal conductance ( g(s)) of upper canopy leaves were measured throughout the growing season. In the site with young Douglas-fir trees, root embolism increased from 20 to 55 percent loss of conductivity (PLC) as the dry season progressed. In young ponderosa pine, root embolism increased from 45 to 75 PLC. In contrast, roots of old-growth Douglas-fir and ponderosa pine trees never experienced more than 30 and 40 PLC, respectively. HR kept soil water potential at 20-30 cm depth above -0.5 MPa in the old-growth Douglas-fir site and -1.8 MPa in the old-growth ponderosa pine site, which significantly reduced loss of shallow root function. In the young ponderosa pine stand, where little HR occurred, the water potential in the upper soil layers fell to about -2.8 MPa, which severely impaired root functioning and limited recovery when the fall rains returned. In both species, daily maximum g(s) decreased linearly with increasing root PLC, suggesting that root xylem embolism acted in concert with stomata to limit water loss, thereby maintaining minimum leaf water potential above critical values. HR appears to be an important mechanism for maintaining shallow root function during drought and preventing total stomatal closure.
EllerCB, LimaAL, OliveiraRS (2016). Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change New Phytologist, 211, 489-501. DOI:10.1111/nph.2016.211.issue-2URL [本文引用: 2]
FlexasJ, CarriquíM, NadalM (2018). Gas exchange and hydraulics during drought in crops: Who drives whom Journal of Experimental Botany, 69, 3791-3795. DOI:10.1093/jxb/ery235PMID:30032258 [本文引用: 1]
FortF, CruzP, LeclouxE, Bittencourtde Oliveira L, StroiaC, TheauJP, JouanyC, PugnaireF (2016). Grassland root functional parameters vary according to a community-level resource acquisition-conservation trade-off Journal of Vegetation Science, 27, 749-758. DOI:10.1111/jvs.12405URL [本文引用: 2]
FortF, VolaireF, GuilioniL, BarkaouiK, NavasML, RoumetC (2017). Root traits are related to plant water-use among rangeland Mediterranean species Functional Ecology, 31, 1700-1709. DOI:10.1111/fec.2017.31.issue-9URL [本文引用: 2]
FuXL, MeinzerFC (2019). Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): a global data set reveals coordination and trade-offs among water transport traits Tree Physiology, 39, 122-134. DOI:10.1093/treephys/tpy087URL [本文引用: 3]
FuXL, MeinzerFC, WoodruffDR, LiuYY, SmithDD, McCullohKA, HowardAR (2019). Coordination and trade-offs between leaf and stem hydraulic traits and stomatal regulation along a spectrum of isohydry to anisohydry Plant, Cell & Environment, 42, 2245-2258. [本文引用: 2]
FuenzalidaTI, BryantCJ, OvingtonLI, YoonHJ, OliveiraRS, SackL, BallMC (2019). Shoot surface water uptake enables leaf hydraulic recovery in Avicennia marina. New Phytologist, 224, 1504-1511. [本文引用: 2]
GoldsmithGR, LehmannMM, CernusakLA, ArendM, SiegwolfRTW (2017). Inferring foliar water uptake using stable isotopes of water Oecologia, 184, 763-766. DOI:10.1007/s00442-017-3917-1PMID:28735456 [本文引用: 1] A growing number of studies have described the direct absorption of water into leaves, a phenomenon known as foliar water uptake. The resultant increase in the amount of water in the leaf can be important for plant function. Exposing leaves to isotopically enriched or depleted water sources has become a common method for establishing whether or not a plant is capable of carrying out foliar water uptake. However, a careful inspection of our understanding of the fluxes of water isotopes between leaves and the atmosphere under high humidity conditions shows that there can clearly be isotopic exchange between the two pools even in the absence of a change in the mass of water in the leaf. We provide experimental evidence that while leaf water isotope ratios may change following exposure to a fog event using water with a depleted oxygen isotope ratio, leaf mass only changes when leaves are experiencing a water deficit that creates a driving gradient for the uptake of water by the leaf. Studies that rely on stable isotopes of water as a means of studying plant water use, particularly with respect to foliar water uptake, must consider the effects of these isotopic exchange processes.
GotschSG, NadkarniN, DarbyA, GlunkA, DixM, DavidsonK, DawsonTE (2015). Life in the treetops: ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest Ecological Monographs, 85, 393-412. DOI:10.1890/14-1076.1URL [本文引用: 1]
HartmannH, LinkRM, SchuldtB (2021). A whole-plant perspective of isohydry: stem-level support for leaf-level plant water regulation Tree Physiology, 41, 901-905. DOI:10.1093/treephys/tpab011PMID:33594416 [本文引用: 1]
HoffmannWA, MarchinRM, AbitP, LauOL (2011). Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought Global Change Biology, 17, 2731-2742. DOI:10.1111/j.1365-2486.2011.02401.xURL [本文引用: 1]
IshidaA, NakanoT, YazakiK, MatsukiS, KoikeN, LauensteinDL, ShimizuM, YamashitaN (2008). Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms Oecologia, 156, 193-202. DOI:10.1007/s00442-008-0965-6URL [本文引用: 2]
JiangPP, MeinzerFC, FuXL, KouL, DaiXQ, WangHM (2021). Trade-offs between xylem water and carbohydrate storage among 24 coexisting subtropical understory shrub species spanning a spectrum of isohydry Tree Physiology, 41, 403-415. DOI:10.1093/treephys/tpaa138URL [本文引用: 2]
JinY, WangCK, ZhouZH (2016). Mechanisms of xylem embolism repair in woody plants: research progress and questions Chinese Journal of Plant Ecology, 40, 834-846. DOI:10.17521/cjpe.2016.0118URL [本文引用: 2]
JinY, WangCK, ZhouZH (2019). Conifers but not angiosperms exhibit vulnerability segmentation between leaves and branches in a temperate forest Tree Physiology, 39, 454-462. DOI:10.1093/treephys/tpy111URL [本文引用: 1]
JinY, WangCK, ZhouZH, LiZM (2016). Co-ordinated performance of leaf hydraulics and economics in 10 Chinese temperate tree species Functional Plant Biology, 43, 1082- 1090. DOI:10.1071/FP16097URL [本文引用: 1]
JohnsonDM, DomecJC, CarterBerry Z, SchwantesAM, McCullohKA, WoodruffDR, WaynePolley H, WortemannR, SwensonJJ, ScottMackay D, McDowellNG, JacksonRB (2018). Co-occurring woody species have diverse hydraulic strategies and mortality rates during an extreme drought Plant, Cell & Environment, 41, 576-588. [本文引用: 3]
KleinT (2014). The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours Functional Ecology, 28, 1313-1320. DOI:10.1111/1365-2435.12289URL [本文引用: 1]
KleinT, ZeppelMJB, AndereggWRL, BloemenJ, KauweMG, HudsonP, RuehrNK, PowellTL, ArxG, NardiniA (2018). Xylem embolism refilling and resilience against drought- induced mortality in woody plants: processes and trade- offs Ecological Research, 33, 839-855. [本文引用: 2]
LanningM, WangLX, NovickKA (2020). The importance of cuticular permeance in assessing plant water-use strategies Tree Physiology, 40, 425-432. DOI:10.1093/treephys/tpaa020URL [本文引用: 1]
LaurJ, HackeUG (2014). Exploring Picea glauca aquaporins in the context of needle water uptake and xylem refilling New Phytologist, 203, 388-400. DOI:10.1111/nph.2014.203.issue-2URL [本文引用: 2]
LeighA, SevantoS, CloseJD, NicotraAB (2017). The influence of leaf size and shape on leaf thermal dynamics: Does theory hold up under natural conditions Plant, Cell & Environment, 40, 237-248. [本文引用: 1]
LensF, SperryJS, ChristmanMA, ChoatB, RabaeyD, JansenS (2011). Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer New Phytologist, 190, 709-723. DOI:10.1111/nph.2011.190.issue-3URL [本文引用: 1]
LiXM, BlackmanCJ, PetersJMR, ChoatB, RymerPD, MedlynBE, TissueDT (2019). More than iso/anisohydry: hydroscapes integrate plant water use and drought tolerance traits in 10 eucalypt species from contrasting climates Functional Ecology, 33, 1035-1049. DOI:10.1111/fec.2019.33.issue-6URL [本文引用: 1]
LiangXY, YeQ, LiuH, BrodribbTJ (2021). Wood density predicts mortality threshold for diverse trees New Phytologist, 229, 3053-3057. DOI:10.1111/nph.v229.6URL [本文引用: 2]
LiuJX, GuL, YuYC, HuangP, WuZG, ZhangQ, QianYQ, WanXC, SunZY (2019a). Corticular photosynthesis drives bark water uptake to refill embolized vessels in dehydrated branches of Salix matsudana Plant, Cell & Environment, 42, 2584-2596. [本文引用: 1]
LiuXR, LiuH, GleasonSM, GoldsteinG, ZhuSD, HePC, HouH, LiRH, YeQ (2019b). Water transport from stem to stomata: the coordination of hydraulic and gas exchange traits across 33 subtropical woody species Tree Physiology, 39, 1665-1674. DOI:10.1093/treephys/tpz076URL [本文引用: 2]
LópezR, CanoFJ, Martin-StPaulNK, CochardH, ChoatB (2021). Coordination of stem and leaf traits define different strategies to regulate water loss and tolerance ranges to aridity New Phytologist, 230, 497-509. DOI:10.1111/nph.v230.2URL [本文引用: 3]
LuoDD, WangCK, JinY (2017). Plant water-regulation strategies: isohydric versus anisohydric behavior Chinese Journal of Plant Ecology, 41, 1021-1032. [本文引用: 1]
Martínez-VilaltaJ, AndereggWRL, SapesG, SalaAN (2019). Greater focus on water pools may improve our ability to understand and anticipate drought-induced mortality in plants New Phytologist, 223, 22-32. DOI:10.1111/nph.2019.223.issue-1URL [本文引用: 2]
Martínez-VilaltaJ, Garcia-FornerN (2017). Water potential regulation, stomatal behaviour and hydraulic transport under drought: deconstructing the iso/anisohydric concept Plant, Cell & Environment, 40, 962-976. [本文引用: 2]
MartinsSCV, McAdamSAM, DeansRM, DaMattaFM, BrodribbTJ (2016). Stomatal dynamics are limited by leaf hydraulics in ferns and conifers: results from simultaneous measurements of liquid and vapour fluxes in leaves Plant, Cell & Environment, 39, 694-705. [本文引用: 2]
McDowellNG, BeerlingDJ, BreshearsDD, FisherRA, RaffaKF, StittM (2011). The interdependence of mechanisms underlying climate-driven vegetation mortality Trends in Ecology & Evolution, 26, 523-532. DOI:10.1016/j.tree.2011.06.003URL [本文引用: 5]
McDowellNG, PockmanWT, AllenCD, BreshearsDD, CobbN, KolbT, PlautJ, SperryJ, WestA, WilliamsDG, YepezEA (2008). Mechanisms of plant survival and mortality during drought: Why do some plants survive while others succumb to drought New Phytologist, 178, 719-739. DOI:10.1111/j.1469-8137.2008.02436.xPMID:18422905 [本文引用: 4] Severe droughts have been associated with regional-scale forest mortality worldwide. Climate change is expected to exacerbate regional mortality events; however, prediction remains difficult because the physiological mechanisms underlying drought survival and mortality are poorly understood. We developed a hydraulically based theory considering carbon balance and insect resistance that allowed development and examination of hypotheses regarding survival and mortality. Multiple mechanisms may cause mortality during drought. A common mechanism for plants with isohydric regulation of water status results from avoidance of drought-induced hydraulic failure via stomatal closure, resulting in carbon starvation and a cascade of downstream effects such as reduced resistance to biotic agents. Mortality by hydraulic failure per se may occur for isohydric seedlings or trees near their maximum height. Although anisohydric plants are relatively drought-tolerant, they are predisposed to hydraulic failure because they operate with narrower hydraulic safety margins during drought. Elevated temperatures should exacerbate carbon starvation and hydraulic failure. Biotic agents may amplify and be amplified by drought-induced plant stress. Wet multidecadal climate oscillations may increase plant susceptibility to drought-induced mortality by stimulating shifts in hydraulic architecture, effectively predisposing plants to water stress. Climate warming and increased frequency of extreme events will probably cause increased regional mortality episodes. Isohydric and anisohydric water potential regulation may partition species between survival and mortality, and, as such, incorporating this hydraulic framework may be effective for modeling plant survival and mortality under future climate conditions.
McDowellNG, RyanMG, ZeppelMJB, TissueDT (2013). Feature: improving our knowledge of drought-induced forest mortality through experiments, observations, and modeling New Phytologist, 200, 289-293. DOI:10.1111/nph.12502PMID:24050629 [本文引用: 1]
MeinzerFC, JohnsonDM, LachenbruchB, McCullohKA, WoodruffDR (2009). Xylem hydraulic safety margins in woody plants: coordination of stomatal control of xylem tension with hydraulic capacitance Functional Ecology, 23, 922-930. DOI:10.1111/fec.2009.23.issue-5URL [本文引用: 2]
MeinzerFC, SmithDD, WoodruffDR, MariasDE, McCullohKA, HowardAR, MagedmanAL (2017). Stomatal kinetics and photosynthetic gas exchange along a continuum of isohydric to anisohydric regulation of plant water status Plant, Cell & Environment, 40, 1618-1628. [本文引用: 1]
MeinzerFC, WoodruffDR, MariasDE, SmithDD, McCullohKA, HowardAR, MagedmanAL (2016). Mapping “hydroscapes” along the iso- to anisohydric continuum of stomatal regulation of plant water status Ecology Letters, 19, 1343-1352. DOI:10.1111/ele.2016.19.issue-11URL [本文引用: 1]
NardiniA, SaviT, TrifilòP, Lo GulloMA,(2018). Drought stress and the recovery from xylem embolism in woody plants Progress in Botany, 79, 197-231. [本文引用: 3]
OgasaM, MikiNH, MurakamiY, YoshikawaK (2013). Recovery performance in xylem hydraulic conductivity is correlated with cavitation resistance for temperate deciduous tree species Tree Physiology, 33, 335-344. DOI:10.1093/treephys/tpt010URL [本文引用: 3]
PivovaroffAL, CookVMW, SantiagoLS (2018). Stomatal behaviour and stem xylem traits are coordinated for woody plant species under exceptional drought conditions Plant, Cell & Environment, 41, 2617-2626. [本文引用: 2]
PivovaroffAL, PasquiniSC, DeGuzman ME, AlstadKP, StemkeJS, SantiagoLS (2016). Multiple strategies for drought survival among woody plant species Functional Ecology, 30, 517-526. DOI:10.1111/fec.2016.30.issue-4URL
PivovaroffAL, SackL, SantiagoLS (2014). Coordination of stem and leaf hydraulic conductance in southern California shrubs: a test of the hydraulic segmentation hypothesis New Phytologist, 203, 842-850. DOI:10.1111/nph.2014.203.issue-3URL [本文引用: 1]
PowellTL, WheelerJK, de OliveiraAAR, da CostaACL, SaleskaSR, MeirP, MoorcroftPR,(2017). Differences in xylem and leaf hydraulic traits explain differences in drought tolerance among mature Amazon rainforest trees Global Change Biology, 23, 4280-4293. DOI:10.1111/gcb.2017.23.issue-10URL [本文引用: 1]
PrattRB, JacobsenAL (2017). Conflicting demands on angiosperm xylem: tradeoffs among storage, transport and biomechanics Plant, Cell & Environment, 40, 897-913. [本文引用: 4]
RoddyAB, SimoninKA, McCullohKA, BrodersenCR, DawsonTE (2018). Water relations of Calycanthus flowers: hydraulic conductance, capacitance, and embolism resistance Plant, Cell & Environment, 41, 2250-2262. [本文引用: 1]
RomanDT, NovickKA, BrzostekER, DragoniD, RahmanF, PhillipsRP (2015). The role of isohydric and anisohydric species in determining ecosystem-scale response to severe drought Oecologia, 179, 641-654. DOI:10.1007/s00442-015-3380-9PMID:26130023 [本文引用: 1] Ongoing shifts in the species composition of Eastern US forests necessitate the development of frameworks to explore how species-specific water-use strategies influence ecosystem-scale carbon (C) cycling during drought. Here, we develop a diagnostic framework to classify plant drought-response strategies along a continuum of isohydric to anisohydric regulation of leaf water potential (Ψ(L)). The framework is applied to a 3-year record of weekly leaf-level gas exchange and Ψ measurements collected in the Morgan-Monroe State Forest (Indiana, USA), where continuous observations of the net ecosystem exchange of CO2 (NEE) have been ongoing since 1999. A severe drought that occurred in the middle of the study period reduced the absolute magnitude of NEE by 55%, though species-specific responses to drought conditions varied. Oak species were characterized by anisohydric regulation of Ψ(L) that promoted static gas exchange throughout the study period. In contrast, Ψ(L) of the other canopy dominant species was more isohydric, which limited gas exchange during the drought. Ecosystem-scale estimates of NEE and gross ecosystem productivity derived by upscaling the leaf-level data agreed well with tower-based observations, and highlight how the fraction of isohydric and anisohydric species in forests can mediate net ecosystem C balance.
SackL, BallMC, BrodersenC, DavisSD, DesMarais DL, DonovanLA, GivnishTJ, HackeUG, HuxmanT, JansenS, JacobsenAL, JohnsonDM, KochGW, MaurelC, McCullohKA, et al. (2016). Plant hydraulics as a central hub integrating plant and ecosystem function: meeting report for “Emerging Frontiers in Plant Hydraulics” (Washington DC, May 2015). Plant, Cell & Environment, 39, 2085-2094. [本文引用: 1]
SalomónRL, SteppeK, OurcivalJM, VillersS, Rodríguez-Calcerrada J, SchapmanR, LimousinJM (2020). Hydraulic acclimation in a Mediterranean oak subjected to permanent throughfall exclusion results in increased stem hydraulic capacitance Plant, Cell & Environment, 43, 1528-1544. [本文引用: 1]
SaveynA, SteppeK, UbiernaN, DawsonTE (2010). Woody tissue photosynthesis and its contribution to trunk growth and bud development in young plants Plant, Cell & Environment, 33, 1949-1958. [本文引用: 1]
SaviT, CasoloV, LuglioJ, BertuzziS, TrifiloP, LoGullo MA, NardiniA (2016). Species-specific reversal of stem xylem embolism after a prolonged drought correlates to endpoint concentration of soluble sugars Plant Physiology Biochemistry, 106, 198-207. DOI:10.1016/j.plaphy.2016.04.051URL [本文引用: 2]
SevantoS, McDowellNG, DickmanLT, PangleR, PockmanWT (2014). How do trees die? A test of the hydraulic failure and carbon starvation hypotheses Plant, Cell & Environment, 37, 153-161. [本文引用: 3]
SkeltonRP, WestAG, DawsonTE (2015). Predicting plant vulnerability to drought in biodiverse regions using functional traits Proceedings of the National Academy of Sciences of the United States of America, 112, 5744-5749. DOI:10.1073/pnas.1503376112PMID:25902534 [本文引用: 7] Attempts to understand mechanisms underlying plant mortality during drought have led to the emergence of a hydraulic framework describing distinct hydraulic strategies among coexisting species. This framework distinguishes species that rapidly decrease stomatal conductance (gs), thereby maintaining high water potential (Px; isohydric), from those species that maintain relatively high gs at low Px, thereby maintaining carbon assimilation, albeit at the cost of loss of hydraulic conductivity (anisohydric). This framework is yet to be tested in biodiverse communities, potentially due to a lack of standardized reference values upon which hydraulic strategies can be defined. We developed a system of quantifying hydraulic strategy using indices from vulnerability curves and stomatal dehydration response curves and tested it in a speciose community from South Africa's Cape Floristic Region. Degree of stomatal regulation over cavitation was defined as the margin between Px at stomatal closure (Pg12) and Px at 50% loss of conductivity. To assess relationships between hydraulic strategy and mortality mechanisms, we developed proxies for carbon limitation and hydraulic failure using time since Pg12 and loss of conductivity at minimum seasonal Px, respectively. Our approach captured continuous variation along an isohydry/anisohydry axis and showed that this variation was linearly related to xylem safety margin. Degree of isohydry/anisohydry was associated with contrasting predictions for mortality during drought. Merging stomatal regulation strategies that represent an index of water use behavior with xylem vulnerability facilitates a more comprehensive framework with which to characterize plant response to drought, thus opening up an avenue for predicting the response of diverse communities to future droughts.
SperryJS, MeinzerFC, McCullonKA (2008). Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees Plant, Cell & Environment, 31, 632-645. [本文引用: 3]
SteppeK, VandegehuchteMW, vande Wal BAE, HosteP, GuyotA, LovelockCE, LockingtonDA (2018). Direct uptake of canopy rainwater causes turgor-driven growth spurts in the mangrove Avicennia marina Tree Physiology, 38, 979-991. DOI:10.1093/treephys/tpy024PMID:29562244 [本文引用: 1] Mangrove forests depend on a dense structure of sufficiently large trees to fulfil their essential functions as providers of food and wood for animals and people, CO2 sinks and protection from storms. Growth of these forests is known to be dependent on the salinity of soil water, but the influence of foliar uptake of rainwater as a freshwater source, additional to soil water, has hardly been investigated. Under field conditions in Australia, stem diameter variation, sap flow and stem water potential of the grey mangrove (Avicennia marina (Forssk.) Vierh.) were simultaneously measured during alternating dry and rainy periods. We found that sap flow in A. marina was reversed, from canopy to roots, during and shortly after rainfall events. Simultaneously, stem diameters rapidly increased with growth rates up to 70 μm h-1, which is about 25-75 times the normal growth rate reported in temperate trees. A mechanistic tree model was applied to provide evidence that A. marina trees take up water through their leaves, and that this water contributes to turgor-driven stem growth. Our results indicate that direct uptake of freshwater by the canopy during rainfall supports mangrove tree growth and serve as a call to consider this water uptake pathway if we aspire to correctly assess influences of changing rainfall patterns on mangrove tree growth.
TanedaH, SperryJS (2008). A case-study of water transport in co-occurring ring- versus diffuse-porous trees: contrasts in water-status, conducting capacity, cavitation and vessel refilling Tree Physiology, 28, 1641-1651. DOI:10.1093/treephys/28.11.1641URL [本文引用: 1]
TardieuF, SimonneauT (1998). Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours Journal of Experimental Botany, 49, 419-432. DOI:10.1093/jxb/49.Special_Issue.419URL [本文引用: 1]
TomasellaM, CasoloV, AichnerN, PetruzzellisF, SaviT, TrifilòP, NardiniA (2019a). Non-structural carbohydrate and hydraulic dynamics during drought and recovery in Fraxinus ornus and Ostrya carpinifolia saplings Plant Physiology Biochemistry, 145, 1-9. DOI:10.1016/j.plaphy.2019.10.024URL [本文引用: 3]
TomasellaM, HäberleKH, NardiniA, HesseB, MachletA, MatyssekR (2017). Post-drought hydraulic recovery is accompanied by non-structural carbohydrate depletion in the stem wood of Norway spruce saplings Scientific Reports, 7, 14308. DOI: 10.1038/s41598-017-14645-w. PMID:29085007 [本文引用: 2] Hydraulic failure and carbon starvation are recognized as main causes of drought-induced forest decline. As water transport and carbon dynamics are strictly interdependent, it is necessary to clarify how dehydration-rehydration cycles are affecting the relations between stem embolism and non-structural carbohydrates (NSC). This is particularly needed for conifers whose embolism repair capability is still controversial. Potted Norway spruce saplings underwent two drought-re-irrigation cycles of same intensity, but performed in two consecutive summers. During the second cycle, stem percent loss of hydraulic conductivity (PLC) and NSC content showed no carry-over effects from the previous drought, indicating complete long-term recovery. The second drought treatment induced moderate PLC (20%) and did not affect total NSCs content, while starch was converted to soluble sugars in the bark. After one week of re-irrigation, PLC recovered to pre-stress values (0%) and NSCs were depleted, only in the wood, by about 30%. Our data suggest that spruce can repair xylem embolism and that, when water is newly available, NSCs stored in xylem parenchyma can be mobilized over short term to sustain respiration and/or for processes involved in xylem transport restoration. This, however, might imply dependency on sapwood NSC reserves for survival, especially if frequent drought spells occur.
TomasellaM, NardiniA, HesseBD, MachletA, MatyssekR, HäberleKH (2019b). Close to the edge: effects of repeated severe drought on stem hydraulics and non-structural carbohydrates in European beech saplings Tree Physiology, 39, 717-728. [本文引用: 2]
TombesiS, NardiniA, FarinelliD, PalliottiA (2014). Relationships between stomatal behavior, xylem vulnerability to cavitation and leaf water relations in two cultivars of Vitis vinifera Physiologia Plantarum, 152, 453-464. DOI:10.1111/ppl.2014.152.issue-3URL [本文引用: 1]
TombesiS, NardiniA, FrioniT, SoccoliniM, ZadraC, FarinelliD, PoniS, PalliottiA (2015). Stomatal closure is induced by hydraulic signals and maintained by ABA in drought- stressed grapevine Scientific Reports, 5, 12449. DOI: 10.1038/srep12449. PMID:26207993 [本文引用: 1] Water saving under drought stress is assured by stomatal closure driven by active (ABA-mediated) and/or passive (hydraulic-mediated) mechanisms. There is currently no comprehensive model nor any general consensus about the actual contribution and relative importance of each of the above factors in modulating stomatal closure in planta. In the present study, we assessed the contribution of passive (hydraulic) vs active (ABA mediated) mechanisms of stomatal closure in V. vinifera plants facing drought stress. Leaf gas exchange decreased progressively to zero during drought, and embolism-induced loss of hydraulic conductance in petioles peaked to similar to 50% in correspondence with strong daily limitation of stomatal conductance. Foliar ABA significantly increased only after complete stomatal closure had already occurred. Rewatering plants after complete stomatal closure and after foliar ABA reached maximum values did not induced stomatal re-opening, despite embolism recovery and water potential rise. Our data suggest that in grapevine stomatal conductance is primarily regulated by passive hydraulic mechanisms. Foliar ABA apparently limits leaf gas exchange over long-term, also preventing recovery of stomatal aperture upon rewatering, suggesting the occurrence of a mechanism of long-term down-regulation of transpiration to favor embolism repair and preserve water under conditions of fluctuating water availability and repeated drought events.
TrifilòP, CasoloV, RaimondoF, PetrussaE, BoscuttiF, LoGullo MA, NardiniA (2017). Effects of prolonged drought on stem non-structural carbohydrates content and post- drought hydraulic recovery in Laurus nobilis L.: the possible link between carbon starvation and hydraulic failure Plant Physiology Biochemistry, 120, 232-241. DOI:10.1016/j.plaphy.2017.10.003URL [本文引用: 2]
TrifilòP, KiorapostolouN, PetruzzellisF, VittiS, PetitG, LoGullo MA, NardiniA, CasoloV (2019). Hydraulic recovery from xylem embolism in excised branches of twelve woody species: relationships with parenchyma cells and non-structural carbohydrates Plant Physiology and Biochemistry, 139, 513-520. DOI:10.1016/j.plaphy.2019.04.013URL [本文引用: 3]
TrifilòP, NardiniA, LoGullo MA, BarberaPM, SaviT, RaimondoF (2015). Diurnal changes in embolism rate in nine dry forest trees: relationships with species-specific xylem vulnerability, hydraulic strategy and wood traits Tree Physiology, 35, 694-705. DOI:10.1093/treephys/tpv049URL [本文引用: 2]
TrifilóP, RaimondoF, SaviT, LoGullo MA, NardiniA (2016). The contribution of vascular and extra-vascular water pathways to drought-induced decline of leaf hydraulic conductance Journal of Experimental Botany, 67, 5029-5039. DOI:10.1093/jxb/erw268URL [本文引用: 2]
TyreeMT, SperryJS (1988). Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress? Answers from a model Plant physiology, 88, 574-580. PMID:16666351 [本文引用: 1] We discuss the relationship between the dynamically changing tension gradients required to move water rapidly through the xylem conduits of plants and the proportion of conduits lost through embolism as a result of water tension. We consider the implications of this relationship to the water relations of trees. We have compiled quantitative data on the water relations, hydraulic architecture and vulnerability of embolism of four widely different species: Rhizophora mangle, Cassipourea elliptica, Acer saccharum, and Thuja occidentalis. Using these data, we modeled the dynamics of water flow and xylem blockage for these species. The model is specifically focused on the conditions required to generate ;runaway embolism,' whereby the blockage of xylem conduits through embolism leads to reduced hydraulic conductance causing increased tension in the remaining vessels and generating more tension in a vicious circle. The model predicted that all species operate near the point of catastrophic xylem failure due to dynamic water stress. The model supports Zimmermann's plant segmentation hypothesis. Zimmermann suggested that plants are designed hydraulically to sacrifice highly vulnerable minor branches and thus improve the water balance of remaining parts. The model results are discussed in terms of the morphology, hydraulic architecture, eco-physiology, and evolution of woody plants.
UrliM, PortéAJ, CochardH, GuengantY, BurlettR, DelzonS (2013). Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees Tree Physiology, 33, 672-683. [本文引用: 1]
VandegehuchteMW, BloemenJ, VergeynstLL, SteppeK (2015). Woody tissue photosynthesis in trees: salve on the wounds of drought New Phytologist, 208, 998-1002. DOI:10.1111/nph.13599PMID:26226885 [本文引用: 1]
VenturasMD, SperryJS, HackeUG (2017). Plant xylem hydraulics: What we understand, current research, and future challenges Journal of Integrative Plant Biology, 59, 356- 389. DOI:10.1111/jipb.12534 [本文引用: 1] Herein we review the current state-of-the-art of plant hydraulics in the context of plant physiology, ecology, and evolution, focusing on current and future research opportunities. We explain the physics of water transport in plants and the limits of this transport system, highlighting the relationships between xylem structure and function. We describe the great variety of techniques existing for evaluating xylem resistance to cavitation. We address several methodological issues and their connection with current debates on conduit refilling and exponentially shaped vulnerability curves. We analyze the trade-offs existing between water transport safety and efficiency. We also stress how little information is available on molecular biology of cavitation and the potential role of aquaporins in conduit refilling. Finally, we draw attention to how plant hydraulic traits can be used for modeling stomatal responses to environmental variables and climate change, including drought mortality.
VesalaT, SevantoS, GrönholmT, SalmonY, NikinmaaE, HariP, HölttäT (2017). Effect of leaf water potential on internal humidity and CO2 dissolution: reverse transpiration and improved water use efficiency under negative pressure Frontiers in Plant Science, 8, 54. DOI: 10.3389/ fpls.2017.00054. PMID:28220128 [本文引用: 2] The pull of water from the soil to the leaves causes water in the transpiration stream to be under negative pressure decreasing the water potential below zero. The osmotic concentration also contributes to the decrease in leaf water potential but withmuch lesser extent. Thus, the surface tension force is approximately balanced by a force induced by negative water potential resulting in concavely curved water-air interfaces in leaves. The lowered water potential causes a reduction in the equilibrium water vapor pressure in internal (sub-stomatal/ intercellular) cavities in relation to that over water with the potential of zero, i.e., over the flat surface. The curved surface causes a reduction also in the equilibrium vapor pressure of dissolved CO2, thus enhancing its physical solubility to water. Although the water vapor reduction is acknowledged by plant physiologists its consequences for water vapor exchange at low water potential values have received very little attention. Consequences of the enhanced CO2 solubility to a leaf water-carbon budget have not been considered at all before this study. We use theoretical calculations and modeling to show how the reduction in the vapor pressures affects transpiration and carbon assimilation rates. Our results indicate that the reduction in vapor pressures of water and CO2 could enhance plant water use efficiency up to about 10% at a leaf water potential of -2 MPa, and much more when water potential decreases further. The low water potential allows for a direct stomatal water vapor uptake from the ambient air even at sub-100% relative humidity values. This alone could explain the observed rates of foliar water uptake by e.g., the coastal redwood in the fog belt region of coastal California provided the stomata are sufficiently open. The omission of the reduction in the water vapor pressure causes a bias in the estimates of the stomatal conductance and leaf internal CO2 concentration based on leaf gas exchange measurements. Manufactures of leaf gas exchange measurement systems should incorporate leaf water potentials in measurement set-ups.
WangAY, HanSJ, ZhangJH, WangM, YinXH, FangLD, YangD, HaoGY (2018a). The interaction between nonstructural carbohydrate reserves and xylem hydraulics in Korean pine trees across an altitudinal gradient Tree physiology, 38, 1792-1804. DOI:10.1093/treephys/tpy119URL [本文引用: 1]
WangXX, DuTT, HuangJL, PengSB, XiongDL (2018b). Leaf hydraulic vulnerability triggers the decline in stomatal and mesophyll conductance during drought in rice Journal of Experimental Botany, 69, 4033-4045. DOI:10.1093/jxb/ery188URL [本文引用: 1]
WheelerJK, HuggettBA, TofteAN, RockwellFE, HolbrookNM (2013). Cutting xylem under tension or supersaturated with gas can generate PLC and the appearance of rapid recovery from embolism Plant, Cell & Environment, 36, 1938-1949. [本文引用: 1]
YangB, WenXF, SunXM (2015). Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region Agricultural and Forest Meteorology, 201, 218-228. DOI:10.1016/j.agrformet.2014.11.020URL [本文引用: 1]
YoshimuraK, SaikiST, YazakiK, OgasaMY, ShiraiM, NakanoT, YoshimuraJ, IshidaA (2016). The dynamics of carbon stored in xylem sapwood to drought-induced hydraulic stress in mature trees Scientific Reports, 6, 24513. DOI: 10.1038/srep24513. PMID:27079677 [本文引用: 1] Climate-induced forest die-off is widespread in multiple biomes, strongly affecting the species composition, function and primary production in forest ecosystems. Hydraulic failure and carbon starvation in xylem sapwood are major hypotheses to explain drought-induced tree mortality. Because it is difficult to obtain enough field observations on drought-induced mortality in adult trees, the current understanding of the physiological mechanisms for tree die-offs is still controversial. However, the simultaneous examination of water and carbon uses throughout dehydration and rehydration processes in adult trees will contribute to clarify the roles of hydraulic failure and carbon starvation in tree wilting. Here we show the processes of the percent loss of hydraulic conductivity (PLC) and the content of nonstructural carbohydrates (NSCs) of distal branches in woody plants with contrasting water use strategy. Starch was converted to soluble sugar during PLC progression under drought, and the hydraulic conductivity recovered following water supply. The conversion of NSCs is strongly associated with PLC variations during dehydration and rehydration processes, indicating that stored carbon contributes to tree survival under drought; further carbon starvation can advance hydraulic failure. We predict that even slow-progressing drought degrades forest ecosystems via carbon starvation, causing more frequent catastrophic forest die-offs than the present projection.
ZhuSD, LiuH, XuQY, CaoKF, YeQ (2016). Are leaves more vulnerable to cavitation than branches Functional Ecology, 30, 1740-1744. DOI:10.1111/fec.2016.30.issue-11URL [本文引用: 1]
ZwienieckiMA, HolbrookNM (2009). Confronting Maxwell's demon: biophysics of xylem embolism repair Trends in Plant Science, 14, 530-534. DOI:10.1016/j.tplants.2009.07.002PMID:19726217 [本文引用: 1] Embolism results in a dramatic loss of xylem hydraulic transport capacity that can lead to decreased plant productivity and even death. The ability to refill embolized conduits despite the presence of tension in the xylem seems to be widespread, but how this occurs is not known. To promote discussion and future research on this topic, we describe how we believe refilling under tension might take place. Our scenario includes: (i) an osmotic role for low-molecular weight sugars; (ii) an apoplastic sugar-sensing mechanism to activate refilling; (iii) the contribution of vapor transport in both the influx of water and removal of entrapped gases; and (iv) the need for a mechanism that can synchronize reconnection to the transpiration stream through multiple bordered pits. A multi-species synthesis of physiological mechanisms in drought-induced tree mortality 4 2017
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
... 单一的指标无法准确表征植物在干旱胁迫下的生存力.例如, 对美国加州南部灌木的研究发现, 具有高栓塞抗性的浅根系灌木在干旱胁迫下的死亡率较高, 而木质部栓塞脆弱性高的深根物种死亡水平则较低(Pivovaroff et al., 2018), 这表明空穴化脆弱性和干旱在某些情况下是解耦的(Blackman et al., 2012).这种解耦现象是由于某些植物的避旱策略所致, 例如较深的根系、干旱落叶和气孔调节等.由于植物对空穴化的脆弱性不能决定其自身的在干旱环境下的生存能力, 因此, 近期通过水力安全阈值(HSM)来预测物种死亡率(Anderegg et al., 2016; Adams et al., 2017).HSM是指一个物种在自然条件下经历的最小水势(Ψmin)与引起木质部功能障碍的水势(如P50或P88)之间的差值, 用来描述植物水力策略的保守程度(Skelton et al., 2015), 其值越大, 表明植物经历栓塞的可能性越低, 意味着越安全(Choat et al.,2012).这里的Ψmin被假定为植物气孔调节对Ψx的影响.植物通过气孔的开闭来限制蒸腾速率、维持水势相对恒定, 是一种重要的水力调节策略.因此, 干旱胁迫下的气孔响应被用来指示植物的抗旱性.但是, 事实上Ψmin集成体现了植物结构特征(如根深)和生理特征(例如气孔行为)对环境胁迫的多重响应(Choat et al., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019). ...
... 水和碳的交互作用(Garcia-Forner et al., 2016), 以及干旱响应的时空格局(Martin-Stpaul et al., 2017)使植物死亡机制异常复杂.McDowell等(2008)的经典死亡机制模型认为等水调节物种更易死于碳饥饿.但在Adams等(2017)的研究中, 部分植物在干旱期间NSCs并不下降, 且有研究发现, 干旱期间等水调节程度更高的物种NSCs浓度更高, 而非等水调节物种的NSCs多用于渗透势的调节(Jiang et al.,2021).因此, 等水调节植物是否会受到更高的碳限制? 在水势值为多少时淀粉开始转化为可溶性糖?干旱期间有多少碳用于渗透调节, 又有多少碳用于呼吸作用? 这些问题都需要我们更深入地了解.此外, 植物水分生理学中对不同器官的研究有助于检验水力特征的协调与权衡, 而综合分析整株植物的水力学特征, 可以更全面地理解水对植物生长和生存的限制过程(Martinez-Vilalta et al., 2019; López et al., 2021).为了更好地理解植物水力学是如何影响生态系统功能的, 我们还需要将个体的结构和生理特征与群落和景观联系起来, 通过基于过程和 特征的模型提高评估和监测干旱诱导死亡风险的 能力. ...
Coordinated decline of leaf hydraulic and stomatal conductances under drought is not linked to leaf xylem embolism for different grapevine cultivars 1 2020
... 有研究发现, Kx和Kox均与gs和蒸腾速率(E)呈正相关关系, 但Kox与gs (或E)之间的相关性很弱, 茎水力导度(Kstem)与叶Kx呈正相关关系, 但与Kox不相关, 表明叶片Kox减弱了叶片水分运输和气体交换之间的协调能力以及茎和叶之间的水力联系, 提出木质部水分运输效率是gs和光合速率的重要决定因素(Liu et al., 2019b).叶柄和叶蒸发部位之间的水势梯度代表了水通过叶片输送的驱动力.一般来说, 由于叶木质部外水力阻力较高, 所以植物所需的木质部外水分运输驱动力(ΔΨox)比木质部水力运输驱动力(ΔΨx)更大.高ΔΨox能够补偿低Kox, 以达到所需的gs或E.此外, 在干旱胁迫条件下, 具有高ΔΨox的物种气孔关闭更早, 以防止Ψleaf过度下降和叶肉组织过度脱水(Liu et al., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
On under- estimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene 1 2015
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
The roles of hydraulic and carbon stress in a widespread climate-induced forest die-off 2 2012
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
... 气孔关闭后, 水仍会通过气孔渗漏, 以及表皮和树皮等其他组织流失, 使植物水势持续缓慢下降.而植物也会释放内部储存水来进一步缓冲Ψx的下降.与此同时, 植物水力导度会通过一系列生物物理和生理机制的改变而下降, 包括叶脉塌缩、植物水通道蛋白调节、细根皮层腔隙的形成等(Choat et al., 2018).当水势下降到一个临界点(P50)时, 栓塞便会在木质部中迅速增长, 这一过程会发生在包括根茎叶在内的整个植物水力系统中(Creek et al., 2018; Rodriguez-Dominguez et al., 2018).由于气孔长期关闭, 光合作用受到抑制, 而呼吸作用还在继续, 从而消耗大量储存的碳水化合物.此外, 植物需要利用碳水化合物来驱动韧皮部运输、维持膨压, 以及重新填充栓塞的木质部, 因此维持呼吸所需的碳水化合物进一步减少.而水和碳的限制又逐渐增加了木质部栓塞的比例, 限制了对冠层的水分输送, 导致分枝斑块性死亡, 冠层叶片面积显著减少(McDowell et al., 2011).随着栓塞逐渐遍布整个输水网络, 造成植物水力系统不可逆的损伤(P88), 最终导致整个植株死亡(Anderegg et al., 2012; Choat et al., 2018).事实上, 在干旱胁迫下, 植物达到死亡临界阈值的概率和时间最终取决于相关生理和形态学特征的相互作用(图2).这种相互作用的多维性是维管植物能够生长在地球上几乎所有的陆地生态系统的有效策略, 并为抗旱提供了大量可能的形态和生理解决方案. ...
Tree mortality predicted from drought-induced vascular damage 2 2015
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
Meta-analysis reveals that hydraulic traits explain cross-species patterns of drought- induced tree mortality across the globe 3 2016
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
... 单一的指标无法准确表征植物在干旱胁迫下的生存力.例如, 对美国加州南部灌木的研究发现, 具有高栓塞抗性的浅根系灌木在干旱胁迫下的死亡率较高, 而木质部栓塞脆弱性高的深根物种死亡水平则较低(Pivovaroff et al., 2018), 这表明空穴化脆弱性和干旱在某些情况下是解耦的(Blackman et al., 2012).这种解耦现象是由于某些植物的避旱策略所致, 例如较深的根系、干旱落叶和气孔调节等.由于植物对空穴化的脆弱性不能决定其自身的在干旱环境下的生存能力, 因此, 近期通过水力安全阈值(HSM)来预测物种死亡率(Anderegg et al., 2016; Adams et al., 2017).HSM是指一个物种在自然条件下经历的最小水势(Ψmin)与引起木质部功能障碍的水势(如P50或P88)之间的差值, 用来描述植物水力策略的保守程度(Skelton et al., 2015), 其值越大, 表明植物经历栓塞的可能性越低, 意味着越安全(Choat et al.,2012).这里的Ψmin被假定为植物气孔调节对Ψx的影响.植物通过气孔的开闭来限制蒸腾速率、维持水势相对恒定, 是一种重要的水力调节策略.因此, 干旱胁迫下的气孔响应被用来指示植物的抗旱性.但是, 事实上Ψmin集成体现了植物结构特征(如根深)和生理特征(例如气孔行为)对环境胁迫的多重响应(Choat et al., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019). ...
Contribution of stem CO2 fixation to whole-plant carbon balance in nonsucculent species 2 2014
... 当叶片C同化和韧皮部C运输受限时, 茎光合作用成为一个重要的替代C源, 对树木在干旱胁迫下的生存至关重要(Cernusak & Cheesman, 2015; Vandegehuchte et al., 2015; De Roo et al., 2020a).Ávila等(2014)将茎表皮上通过气孔或皮孔固定大气CO2的净光合作用称为茎净光合作用(SNP), 而将非光合组织呼吸释放的CO2通过茎皮层组织再同化的过程称为茎循环光合作用(SRP).SNP通常需要植物茎的表皮上存在气孔或皮孔, 以促进CO2从大气扩散到茎皮层的光合细胞中, 一般存在于一些沙漠灌木和早期演替豆科植物中, 且与叶光合作用有相似的特征, 如相似的环境响应曲线, 包括光量子通量密度、胞间CO2浓度和VPD等(Ávila et al., 2014).SRP又称内部再同化作用, 是茎光呼吸速率和茎暗呼吸速率之差.茎内CO2部分源于活细胞的局部呼吸作用, 其中包括地下根呼吸产物通过蒸腾流向上输送.树木通过光合作用将这些CO2重新固定在含有叶绿素的细胞中, 这些细胞存在于树皮、木质部薄壁组织射线甚至茎髓中.Saveyn等(2010)将这个过程称为木质组织光合作用(Pwt), 以表达CO2被整个茎吸收的概念. ...
Climate-related trends in sapwood biophysical properties in two conifers: avoidance of hydraulic dysfunction through coordinated adjustments in xylem efficiency, safety and capacitance 1 2011
... 茎水容(Cstem)为给定水势区间内水含量变化的比率, 是由水分释放曲线的斜率确定的(Sperry et al., 2008).茎水分释放曲线理论上分为3个阶段.第一阶段植物木质部水势相对较高, 释放的水来自细胞间隙和开放的导管、管胞和纤维中, 被称为毛细储水.毛细储水量主要取决于管腔空间, 尤其是纤维腔.纤维腔越大, 木质部密度越低, 因此毛细储水量与木质部密度成反比.第二阶段中, 水主要从薄壁组织、形成层和周皮组织等弹性活细胞中释放, 被称为弹性储水.此时曲线的斜率相对平缓, 且木质组织中弹性储水的体积一般很小.第三阶段的水主要来自栓塞的导管或管胞腔内, 所释放的水量主要取决于栓塞导管或管胞的数量和大小, 被称为空化储水(Pratt & Jacobsen, 2017).Cstem的范围在20- 500 kg·m-3·MPa-1, 主要受边材密度和含水量的影响, 反映了木质部水分运输受阻时蒸腾有效水分的来源(Pratt & Jacobsen, 2017).此外, Cstem还取决于树的大小.树越大, 边材体积越大, 储水空间越大, 且大树相对更依赖于储存水来满足蒸腾需求.表明Cstem是一种与生物量分配模式相关的可塑性特征(McCulloh et al., 2014).Cstem的缓冲作用可以避免植物水势大幅度波动达到不可逆值.高Cstem物种具有更高的Ψmin, 且木质部栓塞抗性与Cstem呈反比(Meinzer et al., 2009).因此, Cstem可能是与木质部安全-效率权衡相关的重要特征(Sperry et al., 2008; McCulloh et al., 2014; Pratt & Jacobsen, 2017).有研究认为, “避旱”树种WD低、Cstem高、栓塞脆弱性高, 而“抗旱”树种WD高、Cstem低, 且在低水势下仍能维持木质部功能(Meinzer et al., 2009; McCulloh et al., 2014).然而这种模式并不完全适用于种内水平.Barnard等(2011)的研究发现, 生长在干旱地区的花旗松(Pseudotsuga menziesii)和黄松(Pinus ponderosa)具有更高的Cstem和栓塞脆弱性, 并认为在较干燥的条件下Cstem的增加一方面可能部分解释了与木质部栓塞相关结构特征的适应性的不足, 另一方面则表明生物量分配的结构调整(如降低叶与边材面积比)是优化用水的关键策略.Salomón等(2020)在对Quercus ilex进行穿透雨排除实验后, 其水分释放曲线第一阶段的Cstem比对照组高, 而在其他阶段差异不显著.表明对于Q. ilex这样的耐旱树种而言, Cstem对水分亏缺的响应以优化碳增益为重. ...
The correlations and sequence of plant stomatal, hydraulic, and wilting responses to drought 2 2016
... 植物维持自身水力系统完整性的能力, 主要取决于其维管系统的物理极限以及植物维持自身水势的能力, 进而决定了植物在干旱胁迫期间的脱水速度以及相应的死亡阈值(Choat et al., 2018).叶膨压损失点水势(ΨTLP), 通常被用以直接量化植物叶片的耐旱性, 因为细胞膨压损失影响了细胞结构完整性、代谢和整个植物的性能, 是衡量植物生理耐旱性的经典指标.低ΨTLP的植物, 可以在较低叶水势(Ψleaf)的情况下仍然维持叶片膨压及其功能, 如光合作用和生长.此外, ΨTLP是与物种的相对等水程度关系最为密切的一个性状, ΨTLP越高, 植物越趋向于等水调节.Meinzer等(2016)建议使用ΨTLP作为衡量等水调节程度的指标, 并已经被大量物种的研究所支持(Fu & Meinzer, 2019; Li et al., 2019).Bartlett等(2016)发现ΨTLP与叶和枝水力导度下降50%的水势(P50)呈正相关关系.Johnson等(2018)发现, 在干旱季节中, 非等水的物种能够更有效地调节其ΨTLP.这种ΨTLP的变化也与叶P50的下降有关. ...
... 不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同.树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低.在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008).有两种假设解释了植物的这种保护机制.一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018).而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降.两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018).此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角. ...
Foliar water uptake: processes, pathways, and integration into plant water budgets 4 2019a
... 植物受干旱影响后的水力恢复速率在很大程度上取决于其水势下降的程度, 以及经历极限低水势的时间.轻度干旱胁迫通常不会导致高水平空穴化栓塞, 但叶脉中的导管塌缩和水通道蛋白对细胞膜通透性的调节机制可能导致植物水力运输能力瞬时且可逆的下降(Zhang et al., 2016).此时, 水力恢复会在降水后通过叶表面吸水(FWU)在很短的时间内发生(Binks et al., 2019; Fuenzalida et al., 2019).FWU是一种几乎在所有植物中都存在的水分获取机制(Dawson & Goldsmith, 2018; Berry et al., 2019a). ...
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
Xylem function and growth rate interact to determine recovery rates after exposure to extreme water deficit 1 2010
... 在未致死的前提下, 当干旱造成植物相对严重的水力功能障碍时, 水力恢复可通过两种机制进行.一是形成新木质部, 二是木质部栓塞的修复.木质部的再生似乎是树木在干旱后恢复水力能力的主要途径(Brodribb et al., 2010; Creek et al., 2018).通过维管束形成层的活动, 在木质部外部区域形成新的导管(或管胞), 以替代之前被树胶或侵填体永久堵塞并失去导水能力的部分.而在干旱造成植物地上部分大量死亡的情况下, 可通过芽或木质块茎的重新萌生促进恢复.在热带和温带森林物种中, 木质部再生的发生程度是高度可变的, 最终取决于植物分生组织的存活程度(Zeppel et al., 2015).对于栓塞修复, 某些植物可通过根压在木质部中形成“正压”以重填栓塞木质部的观点已被普遍接受(Nardini et al., 2018); 而关于在负压下木质部再充水的机制以及用于检测和量化的方法仍然存在争论. ...
Hydraulic failure defines the recovery and point of death in water-stressed conifers 1 2009
... 在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率.随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988).因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012).当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损.在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
Leaf maximum photosynthetic rate and venation are linked by hydraulics 1 2007
... 部分植物同时表现出低水势和低Cstem, 这是由于植物器官栓塞抗性的强度与WD和细胞壁厚度相关.因为在一定的茎体积下, 较厚的导管(或管胞)壁和纤维壁会导致储水空间较小, 使得低水势物种具有较低的Cstem.因此, 这种机械联系导致P50和Cstem之间有很强的正相关关系(McCulloh et al., 2014).对很多物种来说, Cstem与由气孔调节所决定的日最小枝水势之间也存在着功能联系.高Cstem的物种往往更偏向等水调节, 但Fu等(2019)发现更偏向非等水的物种比等水调节物种有更大的Cleaf.因此, 叶和茎的水容可能呈负相关关系(Fu et al., 2019), 这可能是因为高Cstem的物种一般具有高Kstem, Kleaf和Kstem一般呈正相关关系, 而Kleaf越高的物种, 叶片越薄(Brodribb et al., 2007), 而较薄的叶片限制了其自身的Cleaf. ...
Passive origins of stomatal control in vascular plants 2 2011
... Cleaf的缓冲作用可以在不同层次上体现.在针叶树和蕨类植物中, 随VPD增加, Cleaf越低, 气孔关闭越快.这些植物类群一般表现为被动气孔控制(Brodribb & McAdam, 2011; Brodribb et al., 2014), 气孔直接响应于叶片的水分状况.因此, 单位蒸腾损失增加时, 低Cleaf可反映出较低的Ψleaf和高VPD诱发的气孔迅速关闭(Cardoso et al., 2019).Martins等(2016)发现, 在Cleaf较低的植物中, 气孔迅速关闭并没有阻止Ψleaf下降, 这里暗示了一种受低Cleaf的低缓冲能力所驱动的被动水力响应.而在被子植物中, gs下降50%所需的时间(T50)均高于针叶树和蕨类植物, 但T50和Cleaf之间的关系仍随其等水调节程度而不同(Fu et al., 2019).被子植物对水分亏缺的主动调节机制使其对Ψleaf下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
... 下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
Evolution of the stomatal regulation of plant water content 1 2017
... Cleaf的缓冲作用可以在不同层次上体现.在针叶树和蕨类植物中, 随VPD增加, Cleaf越低, 气孔关闭越快.这些植物类群一般表现为被动气孔控制(Brodribb & McAdam, 2011; Brodribb et al., 2014), 气孔直接响应于叶片的水分状况.因此, 单位蒸腾损失增加时, 低Cleaf可反映出较低的Ψleaf和高VPD诱发的气孔迅速关闭(Cardoso et al., 2019).Martins等(2016)发现, 在Cleaf较低的植物中, 气孔迅速关闭并没有阻止Ψleaf下降, 这里暗示了一种受低Cleaf的低缓冲能力所驱动的被动水力响应.而在被子植物中, gs下降50%所需的时间(T50)均高于针叶树和蕨类植物, 但T50和Cleaf之间的关系仍随其等水调节程度而不同(Fu et al., 2019).被子植物对水分亏缺的主动调节机制使其对Ψleaf下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
Conifer species adapt to low-rainfall climates by following one of two divergent pathways 1 2014
... Cleaf的缓冲作用可以在不同层次上体现.在针叶树和蕨类植物中, 随VPD增加, Cleaf越低, 气孔关闭越快.这些植物类群一般表现为被动气孔控制(Brodribb & McAdam, 2011; Brodribb et al., 2014), 气孔直接响应于叶片的水分状况.因此, 单位蒸腾损失增加时, 低Cleaf可反映出较低的Ψleaf和高VPD诱发的气孔迅速关闭(Cardoso et al., 2019).Martins等(2016)发现, 在Cleaf较低的植物中, 气孔迅速关闭并没有阻止Ψleaf下降, 这里暗示了一种受低Cleaf的低缓冲能力所驱动的被动水力响应.而在被子植物中, gs下降50%所需的时间(T50)均高于针叶树和蕨类植物, 但T50和Cleaf之间的关系仍随其等水调节程度而不同(Fu et al., 2019).被子植物对水分亏缺的主动调节机制使其对Ψleaf下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
How do stomata respond to water status 1 2019
... 植物对干旱胁迫的响应过程主要分为两个阶段(图1).第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017).在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降.植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017).随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度.气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019).在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014).尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018). ...
How does leaf anatomy influence water transport outside the xylem 2 2015
... 维管植物的叶片中, 水通过木质部导管在叶柄和整个叶脉网络中运动, 之后穿过维管束鞘、叶肉和表皮细胞, 扩散到细胞间隙, 最后通过气孔扩散到叶片外(Buckley et al., 2015).叶水力导度(Kleaf)代表了叶片中水分的运输效率, 包括叶木质部水力导度(Kx)和木质部外水力导度(Kox).木质部水分运输是水在内聚力-张力机制下通过负压在木质部中移动的过程.Kx主要由叶脉密度、排列方式、形状和导管直径和长度等结构特征所决定(Venturas et al., 2017).在干旱胁迫下, 木质部张力增加, 形成空穴化栓塞, 则导致Kx下降.木质部外水分运输路径较为复杂, 是水通过质外体或共质体途径在活细胞内移动的过程.该水分运输途径涉及多种组织, 而这些组织的导水阻力在很大程度上仍是未知的. ...
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
Hydraulics regulate stomatal responses to changes in leaf water status in the fern Athyrium filix-femina 1 2019
... Cleaf的缓冲作用可以在不同层次上体现.在针叶树和蕨类植物中, 随VPD增加, Cleaf越低, 气孔关闭越快.这些植物类群一般表现为被动气孔控制(Brodribb & McAdam, 2011; Brodribb et al., 2014), 气孔直接响应于叶片的水分状况.因此, 单位蒸腾损失增加时, 低Cleaf可反映出较低的Ψleaf和高VPD诱发的气孔迅速关闭(Cardoso et al., 2019).Martins等(2016)发现, 在Cleaf较低的植物中, 气孔迅速关闭并没有阻止Ψleaf下降, 这里暗示了一种受低Cleaf的低缓冲能力所驱动的被动水力响应.而在被子植物中, gs下降50%所需的时间(T50)均高于针叶树和蕨类植物, 但T50和Cleaf之间的关系仍随其等水调节程度而不同(Fu et al., 2019).被子植物对水分亏缺的主动调节机制使其对Ψleaf下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
1 2016
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
The benefits of recycling: How photosynthetic bark can increase drought tolerance 2 2015
... 当叶片C同化和韧皮部C运输受限时, 茎光合作用成为一个重要的替代C源, 对树木在干旱胁迫下的生存至关重要(Cernusak & Cheesman, 2015; Vandegehuchte et al., 2015; De Roo et al., 2020a).Ávila等(2014)将茎表皮上通过气孔或皮孔固定大气CO2的净光合作用称为茎净光合作用(SNP), 而将非光合组织呼吸释放的CO2通过茎皮层组织再同化的过程称为茎循环光合作用(SRP).SNP通常需要植物茎的表皮上存在气孔或皮孔, 以促进CO2从大气扩散到茎皮层的光合细胞中, 一般存在于一些沙漠灌木和早期演替豆科植物中, 且与叶光合作用有相似的特征, 如相似的环境响应曲线, 包括光量子通量密度、胞间CO2浓度和VPD等(Ávila et al., 2014).SRP又称内部再同化作用, 是茎光呼吸速率和茎暗呼吸速率之差.茎内CO2部分源于活细胞的局部呼吸作用, 其中包括地下根呼吸产物通过蒸腾流向上输送.树木通过光合作用将这些CO2重新固定在含有叶绿素的细胞中, 这些细胞存在于树皮、木质部薄壁组织射线甚至茎髓中.Saveyn等(2010)将这个过程称为木质组织光合作用(Pwt), 以表达CO2被整个茎吸收的概念. ...
... 长时间的干旱通常伴随着高温.在高温下, SRP比叶光合作用更有优势, 因为木质部和树皮组织对气体扩散的阻碍导致茎内部CO2浓度通常处于较高水平.这种CO2与O2的高比例补偿了核酮糖-1,5-二磷酸羧化酶随温度升高而下降的专一性.因此, 在高温下, 叶光合作用几乎停止时, SRP仍可继续提供C源(Cernusak & Cheesman, 2015; De Baerdemaeker et al., 2017).干旱期间, 当大气CO2吸收有限时, Pwt将更加重要.干旱会导致叶片到木质组织的韧皮部运输停滞, 因此, 即使叶片中仍有碳水化合物, 植物也难以将其输送到木质组织.此时, Pwt产生局部可用的碳水化合物, 可用于维持植物水力完整性(Sevanto et al., 2014; De Baerdemaeker et al., 2017; De Roo et al., 2020a).除局部呼吸CO2外, 木质部运输CO2的内部循环也对全碳同化有显著贡献, 这取决于小气候条件和控制蒸腾、呼吸和光合作用的生态生理因子.只要植物呼吸作用持续, 内部的CO2底物便可用于Pwt (Mitchell et al., 2013).因此, Pwt可以被认为是干旱期间碳循环的重要来源(De Roo et al., 2020b).Pwt提高了非结构性碳水化合物(NSCs)的有效性, 可使木质部的结构特征具有更强的抗栓塞能力.De Baerdemaeker等(2017)研究表明, Pwt产生的糖降低了木质部空穴化栓塞脆弱性.Schmitz等(2012)对红树林中的13种植物进行研究, 发现当Pwt受阻时, Kstem显著降低. ...
Root morphology and mycorrhizal symbioses together shape nutrient foraging strategies of temperate trees 1 2016
... 根系负责养分和水分的吸收, 并在缓解营养和水分胁迫对植物生长和物种间相互作用的影响方面发挥重要作用(Fort et al., 2016).根系深浅可以影响植物的抗旱性.一般来说, 在同一地区共存木本植物中, 根系分布较浅树种其栓塞抗性较强; 而根系分布较深的植物可以通过获取深层土壤水来缓冲植物水势的下降, 从而避免干旱损伤(Johnson et al., 2018).不同土壤水分利用对策对生长在干旱环境中植物的生长和生存至关重要(Chen et al., 2016).此外, 一些树种的主要水源在雨季为浅层土壤水, 在旱季则转而利用深层土壤水, 这是由于其根系特征的二态性决定的(Fort et al., 2017).浅层土壤中的细根一般直径较小, 根组织密度较低, 以适应较难预测的水分有效性, 而深层土壤中的细根的中柱较粗, 木质部管道较宽, 以减少水力阻力(Fort et al., 2016, 2017).这种物种间的水文生态位划分对减少竞争、提高生态系统中群落结构的稳定性起着至关重要的作用(Yang et al., 2015). ...
Prediction of temperate broadleaf tree species mortality in arid limestone habitats with stomatal safety margins 2 2019
... 单一的指标无法准确表征植物在干旱胁迫下的生存力.例如, 对美国加州南部灌木的研究发现, 具有高栓塞抗性的浅根系灌木在干旱胁迫下的死亡率较高, 而木质部栓塞脆弱性高的深根物种死亡水平则较低(Pivovaroff et al., 2018), 这表明空穴化脆弱性和干旱在某些情况下是解耦的(Blackman et al., 2012).这种解耦现象是由于某些植物的避旱策略所致, 例如较深的根系、干旱落叶和气孔调节等.由于植物对空穴化的脆弱性不能决定其自身的在干旱环境下的生存能力, 因此, 近期通过水力安全阈值(HSM)来预测物种死亡率(Anderegg et al., 2016; Adams et al., 2017).HSM是指一个物种在自然条件下经历的最小水势(Ψmin)与引起木质部功能障碍的水势(如P50或P88)之间的差值, 用来描述植物水力策略的保守程度(Skelton et al., 2015), 其值越大, 表明植物经历栓塞的可能性越低, 意味着越安全(Choat et al.,2012).这里的Ψmin被假定为植物气孔调节对Ψx的影响.植物通过气孔的开闭来限制蒸腾速率、维持水势相对恒定, 是一种重要的水力调节策略.因此, 干旱胁迫下的气孔响应被用来指示植物的抗旱性.但是, 事实上Ψmin集成体现了植物结构特征(如根深)和生理特征(例如气孔行为)对环境胁迫的多重响应(Choat et al., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019). ...
... 为了直接评估干旱胁迫下的气孔响应, Skelton等(2015)通过拟合气孔导度(gs)随Ψx变化的响应曲线来定义标准化的临界点.其中, 最大气孔导度下降50%时的切线与Ψx轴的交点被定义为气孔闭合点, 且该点对应的gs约损失了最大气孔导度的88%, 因此Ψgs88被定义为气孔关闭所对应的水势, 而Ψgs88与P50 (或P88)的差值被称为气孔安全阈值(SSM), 用于评价气孔和水力之间的协调能力(Skelton et al., 2015).该值越高, 表明植物气孔和水力之间的协调能力越强(Creek et al., 2018).有研究用SSM50 (Ψgs88与P50之差)作为指示植物等水调节程度的指标, 该值为正, 表明气孔关闭发生在水力导度下降到50%之前, 定义为等水调节行为; 而该值为负时则表明发生在P50之后, 定义为非等水调节行为.并认为等水调节物种比非等水调节物种具有更大的水力和气孔安全阈值(Skelton et al., 2015).此外, 有研究表明, 只有少数物种气孔关闭与木质部栓塞形成初期的水势相等, 且随植物栓塞抗性的增加, SSM增加(Martin-Stpaul et al., 2017).Chen等(2019)在对温带阔叶树种死亡率预测的研究中, 发现相比空穴化脆弱性(P50和P88)和HSM, SSM能更好地预测干旱胁迫下植物的死亡风险.但该指标目前使用并不广泛, 需要更多的研究加以验证. ...
Triggers of tree mortality under drought 8 2018
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
... ., 2018). ...
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
... 植物维持自身水力系统完整性的能力, 主要取决于其维管系统的物理极限以及植物维持自身水势的能力, 进而决定了植物在干旱胁迫期间的脱水速度以及相应的死亡阈值(Choat et al., 2018).叶膨压损失点水势(ΨTLP), 通常被用以直接量化植物叶片的耐旱性, 因为细胞膨压损失影响了细胞结构完整性、代谢和整个植物的性能, 是衡量植物生理耐旱性的经典指标.低ΨTLP的植物, 可以在较低叶水势(Ψleaf)的情况下仍然维持叶片膨压及其功能, 如光合作用和生长.此外, ΨTLP是与物种的相对等水程度关系最为密切的一个性状, ΨTLP越高, 植物越趋向于等水调节.Meinzer等(2016)建议使用ΨTLP作为衡量等水调节程度的指标, 并已经被大量物种的研究所支持(Fu & Meinzer, 2019; Li et al., 2019).Bartlett等(2016)发现ΨTLP与叶和枝水力导度下降50%的水势(P50)呈正相关关系.Johnson等(2018)发现, 在干旱季节中, 非等水的物种能够更有效地调节其ΨTLP.这种ΨTLP的变化也与叶P50的下降有关. ...
... 单一的指标无法准确表征植物在干旱胁迫下的生存力.例如, 对美国加州南部灌木的研究发现, 具有高栓塞抗性的浅根系灌木在干旱胁迫下的死亡率较高, 而木质部栓塞脆弱性高的深根物种死亡水平则较低(Pivovaroff et al., 2018), 这表明空穴化脆弱性和干旱在某些情况下是解耦的(Blackman et al., 2012).这种解耦现象是由于某些植物的避旱策略所致, 例如较深的根系、干旱落叶和气孔调节等.由于植物对空穴化的脆弱性不能决定其自身的在干旱环境下的生存能力, 因此, 近期通过水力安全阈值(HSM)来预测物种死亡率(Anderegg et al., 2016; Adams et al., 2017).HSM是指一个物种在自然条件下经历的最小水势(Ψmin)与引起木质部功能障碍的水势(如P50或P88)之间的差值, 用来描述植物水力策略的保守程度(Skelton et al., 2015), 其值越大, 表明植物经历栓塞的可能性越低, 意味着越安全(Choat et al.,2012).这里的Ψmin被假定为植物气孔调节对Ψx的影响.植物通过气孔的开闭来限制蒸腾速率、维持水势相对恒定, 是一种重要的水力调节策略.因此, 干旱胁迫下的气孔响应被用来指示植物的抗旱性.但是, 事实上Ψmin集成体现了植物结构特征(如根深)和生理特征(例如气孔行为)对环境胁迫的多重响应(Choat et al., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019). ...
... 植物对干旱胁迫的响应过程主要分为两个阶段(图1).第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017).在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降.植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017).随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度.气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019).在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014).尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018). ...
... 气孔关闭后, 水仍会通过气孔渗漏, 以及表皮和树皮等其他组织流失, 使植物水势持续缓慢下降.而植物也会释放内部储存水来进一步缓冲Ψx的下降.与此同时, 植物水力导度会通过一系列生物物理和生理机制的改变而下降, 包括叶脉塌缩、植物水通道蛋白调节、细根皮层腔隙的形成等(Choat et al., 2018).当水势下降到一个临界点(P50)时, 栓塞便会在木质部中迅速增长, 这一过程会发生在包括根茎叶在内的整个植物水力系统中(Creek et al., 2018; Rodriguez-Dominguez et al., 2018).由于气孔长期关闭, 光合作用受到抑制, 而呼吸作用还在继续, 从而消耗大量储存的碳水化合物.此外, 植物需要利用碳水化合物来驱动韧皮部运输、维持膨压, 以及重新填充栓塞的木质部, 因此维持呼吸所需的碳水化合物进一步减少.而水和碳的限制又逐渐增加了木质部栓塞的比例, 限制了对冠层的水分输送, 导致分枝斑块性死亡, 冠层叶片面积显著减少(McDowell et al., 2011).随着栓塞逐渐遍布整个输水网络, 造成植物水力系统不可逆的损伤(P88), 最终导致整个植株死亡(Anderegg et al., 2012; Choat et al., 2018).事实上, 在干旱胁迫下, 植物达到死亡临界阈值的概率和时间最终取决于相关生理和形态学特征的相互作用(图2).这种相互作用的多维性是维管植物能够生长在地球上几乎所有的陆地生态系统的有效策略, 并为抗旱提供了大量可能的形态和生理解决方案. ...
Global convergence in the vulnerability of forests to drought 5 2012
... 在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率.随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988).因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012).当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损.在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
... ., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
... ., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
... 单一的指标无法准确表征植物在干旱胁迫下的生存力.例如, 对美国加州南部灌木的研究发现, 具有高栓塞抗性的浅根系灌木在干旱胁迫下的死亡率较高, 而木质部栓塞脆弱性高的深根物种死亡水平则较低(Pivovaroff et al., 2018), 这表明空穴化脆弱性和干旱在某些情况下是解耦的(Blackman et al., 2012).这种解耦现象是由于某些植物的避旱策略所致, 例如较深的根系、干旱落叶和气孔调节等.由于植物对空穴化的脆弱性不能决定其自身的在干旱环境下的生存能力, 因此, 近期通过水力安全阈值(HSM)来预测物种死亡率(Anderegg et al., 2016; Adams et al., 2017).HSM是指一个物种在自然条件下经历的最小水势(Ψmin)与引起木质部功能障碍的水势(如P50或P88)之间的差值, 用来描述植物水力策略的保守程度(Skelton et al., 2015), 其值越大, 表明植物经历栓塞的可能性越低, 意味着越安全(Choat et al.,2012).这里的Ψmin被假定为植物气孔调节对Ψx的影响.植物通过气孔的开闭来限制蒸腾速率、维持水势相对恒定, 是一种重要的水力调节策略.因此, 干旱胁迫下的气孔响应被用来指示植物的抗旱性.但是, 事实上Ψmin集成体现了植物结构特征(如根深)和生理特征(例如气孔行为)对环境胁迫的多重响应(Choat et al., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019). ...
... ., 2012, 2018), 在经历不同程度干旱时的数据也不尽相同, 因此在自然条件下很难准确测量, 从而导致HSM预测能力的不确定性(Skelton et al., 2015; Chen et al., 2019). ...
The impacts of increasing drought on forest dynamics, structure, and biodiversity in the United States 1 2016
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
Coordination between leaf, stem and root hydraulics and gas exchange in three arid-zone angiosperms during severe drought and recovery 3 2018
... 气孔关闭后, 水仍会通过气孔渗漏, 以及表皮和树皮等其他组织流失, 使植物水势持续缓慢下降.而植物也会释放内部储存水来进一步缓冲Ψx的下降.与此同时, 植物水力导度会通过一系列生物物理和生理机制的改变而下降, 包括叶脉塌缩、植物水通道蛋白调节、细根皮层腔隙的形成等(Choat et al., 2018).当水势下降到一个临界点(P50)时, 栓塞便会在木质部中迅速增长, 这一过程会发生在包括根茎叶在内的整个植物水力系统中(Creek et al., 2018; Rodriguez-Dominguez et al., 2018).由于气孔长期关闭, 光合作用受到抑制, 而呼吸作用还在继续, 从而消耗大量储存的碳水化合物.此外, 植物需要利用碳水化合物来驱动韧皮部运输、维持膨压, 以及重新填充栓塞的木质部, 因此维持呼吸所需的碳水化合物进一步减少.而水和碳的限制又逐渐增加了木质部栓塞的比例, 限制了对冠层的水分输送, 导致分枝斑块性死亡, 冠层叶片面积显著减少(McDowell et al., 2011).随着栓塞逐渐遍布整个输水网络, 造成植物水力系统不可逆的损伤(P88), 最终导致整个植株死亡(Anderegg et al., 2012; Choat et al., 2018).事实上, 在干旱胁迫下, 植物达到死亡临界阈值的概率和时间最终取决于相关生理和形态学特征的相互作用(图2).这种相互作用的多维性是维管植物能够生长在地球上几乎所有的陆地生态系统的有效策略, 并为抗旱提供了大量可能的形态和生理解决方案. ...
... 此外, 叶、茎和根性状的协调, 决定树木应对干旱胁迫的水力策略和耐受范围.Creek等(2018)发现, 导致Eucalyptus coolabah叶片气孔关闭的水势与根水力下降水势相吻合, 表明其中可能存在一种与气孔关闭相关的水力信号.López等(2021)的研究表明, 叶和茎的耐旱性既关联又相对独立, 其功能权衡取决于系统发育阶段和水分有效性.其中, 在茎叶性状变异轴上, 一端是具有较低的栓塞脆弱性、较高的水分利用效率和胡伯尔值(Hv, 茎边材面积与叶面积之比)以及气孔关闭较晚的“慢”对策物种; 而另一端是具有高栓塞脆弱性、低水分利用效率和Hv的“快”对策物种.这种生存对策的差异是由区域水分有效性所致.此外, 植物性状变异是多维的(Powell et al., 2017), 不同的维度允许不同功能性状组合, 因而解释了一些功能性状权衡未能反映在较低系统发育水平上的原因(Pivovaroff et al., 2014; López et al., 2021). ...
... 在未致死的前提下, 当干旱造成植物相对严重的水力功能障碍时, 水力恢复可通过两种机制进行.一是形成新木质部, 二是木质部栓塞的修复.木质部的再生似乎是树木在干旱后恢复水力能力的主要途径(Brodribb et al., 2010; Creek et al., 2018).通过维管束形成层的活动, 在木质部外部区域形成新的导管(或管胞), 以替代之前被树胶或侵填体永久堵塞并失去导水能力的部分.而在干旱造成植物地上部分大量死亡的情况下, 可通过芽或木质块茎的重新萌生促进恢复.在热带和温带森林物种中, 木质部再生的发生程度是高度可变的, 最终取决于植物分生组织的存活程度(Zeppel et al., 2015).对于栓塞修复, 某些植物可通过根压在木质部中形成“正压”以重填栓塞木质部的观点已被普遍接受(Nardini et al., 2018); 而关于在负压下木质部再充水的机制以及用于检测和量化的方法仍然存在争论. ...
Xylem embolism in leaves does not occur with open stomata: evidence from direct observations using the optical visualization technique 1 2020
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
The value of wet leaves 1 2018
... 植物受干旱影响后的水力恢复速率在很大程度上取决于其水势下降的程度, 以及经历极限低水势的时间.轻度干旱胁迫通常不会导致高水平空穴化栓塞, 但叶脉中的导管塌缩和水通道蛋白对细胞膜通透性的调节机制可能导致植物水力运输能力瞬时且可逆的下降(Zhang et al., 2016).此时, 水力恢复会在降水后通过叶表面吸水(FWU)在很短的时间内发生(Binks et al., 2019; Fuenzalida et al., 2019).FWU是一种几乎在所有植物中都存在的水分获取机制(Dawson & Goldsmith, 2018; Berry et al., 2019a). ...
Sugars from woody tissue photosynthesis reduce xylem vulnerability to cavitation 3 2017
... 长时间的干旱通常伴随着高温.在高温下, SRP比叶光合作用更有优势, 因为木质部和树皮组织对气体扩散的阻碍导致茎内部CO2浓度通常处于较高水平.这种CO2与O2的高比例补偿了核酮糖-1,5-二磷酸羧化酶随温度升高而下降的专一性.因此, 在高温下, 叶光合作用几乎停止时, SRP仍可继续提供C源(Cernusak & Cheesman, 2015; De Baerdemaeker et al., 2017).干旱期间, 当大气CO2吸收有限时, Pwt将更加重要.干旱会导致叶片到木质组织的韧皮部运输停滞, 因此, 即使叶片中仍有碳水化合物, 植物也难以将其输送到木质组织.此时, Pwt产生局部可用的碳水化合物, 可用于维持植物水力完整性(Sevanto et al., 2014; De Baerdemaeker et al., 2017; De Roo et al., 2020a).除局部呼吸CO2外, 木质部运输CO2的内部循环也对全碳同化有显著贡献, 这取决于小气候条件和控制蒸腾、呼吸和光合作用的生态生理因子.只要植物呼吸作用持续, 内部的CO2底物便可用于Pwt (Mitchell et al., 2013).因此, Pwt可以被认为是干旱期间碳循环的重要来源(De Roo et al., 2020b).Pwt提高了非结构性碳水化合物(NSCs)的有效性, 可使木质部的结构特征具有更强的抗栓塞能力.De Baerdemaeker等(2017)研究表明, Pwt产生的糖降低了木质部空穴化栓塞脆弱性.Schmitz等(2012)对红树林中的13种植物进行研究, 发现当Pwt受阻时, Kstem显著降低. ...
... ., 2017; De Roo et al., 2020a).除局部呼吸CO2外, 木质部运输CO2的内部循环也对全碳同化有显著贡献, 这取决于小气候条件和控制蒸腾、呼吸和光合作用的生态生理因子.只要植物呼吸作用持续, 内部的CO2底物便可用于Pwt (Mitchell et al., 2013).因此, Pwt可以被认为是干旱期间碳循环的重要来源(De Roo et al., 2020b).Pwt提高了非结构性碳水化合物(NSCs)的有效性, 可使木质部的结构特征具有更强的抗栓塞能力.De Baerdemaeker等(2017)研究表明, Pwt产生的糖降低了木质部空穴化栓塞脆弱性.Schmitz等(2012)对红树林中的13种植物进行研究, 发现当Pwt受阻时, Kstem显著降低. ...
Hydraulic traits of Neotropical canopy liana and tree species across a broad range of wood density: implications for predicting drought mortality with models 1 2021
... 在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率.随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988).因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012).当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损.在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
Woody tissue photosynthesis delays drought stress in Populus tremula trees and maintains starch reserves in branch xylem tissues 2 2020a
... 当叶片C同化和韧皮部C运输受限时, 茎光合作用成为一个重要的替代C源, 对树木在干旱胁迫下的生存至关重要(Cernusak & Cheesman, 2015; Vandegehuchte et al., 2015; De Roo et al., 2020a).Ávila等(2014)将茎表皮上通过气孔或皮孔固定大气CO2的净光合作用称为茎净光合作用(SNP), 而将非光合组织呼吸释放的CO2通过茎皮层组织再同化的过程称为茎循环光合作用(SRP).SNP通常需要植物茎的表皮上存在气孔或皮孔, 以促进CO2从大气扩散到茎皮层的光合细胞中, 一般存在于一些沙漠灌木和早期演替豆科植物中, 且与叶光合作用有相似的特征, 如相似的环境响应曲线, 包括光量子通量密度、胞间CO2浓度和VPD等(Ávila et al., 2014).SRP又称内部再同化作用, 是茎光呼吸速率和茎暗呼吸速率之差.茎内CO2部分源于活细胞的局部呼吸作用, 其中包括地下根呼吸产物通过蒸腾流向上输送.树木通过光合作用将这些CO2重新固定在含有叶绿素的细胞中, 这些细胞存在于树皮、木质部薄壁组织射线甚至茎髓中.Saveyn等(2010)将这个过程称为木质组织光合作用(Pwt), 以表达CO2被整个茎吸收的概念. ...
... 长时间的干旱通常伴随着高温.在高温下, SRP比叶光合作用更有优势, 因为木质部和树皮组织对气体扩散的阻碍导致茎内部CO2浓度通常处于较高水平.这种CO2与O2的高比例补偿了核酮糖-1,5-二磷酸羧化酶随温度升高而下降的专一性.因此, 在高温下, 叶光合作用几乎停止时, SRP仍可继续提供C源(Cernusak & Cheesman, 2015; De Baerdemaeker et al., 2017).干旱期间, 当大气CO2吸收有限时, Pwt将更加重要.干旱会导致叶片到木质组织的韧皮部运输停滞, 因此, 即使叶片中仍有碳水化合物, 植物也难以将其输送到木质组织.此时, Pwt产生局部可用的碳水化合物, 可用于维持植物水力完整性(Sevanto et al., 2014; De Baerdemaeker et al., 2017; De Roo et al., 2020a).除局部呼吸CO2外, 木质部运输CO2的内部循环也对全碳同化有显著贡献, 这取决于小气候条件和控制蒸腾、呼吸和光合作用的生态生理因子.只要植物呼吸作用持续, 内部的CO2底物便可用于Pwt (Mitchell et al., 2013).因此, Pwt可以被认为是干旱期间碳循环的重要来源(De Roo et al., 2020b).Pwt提高了非结构性碳水化合物(NSCs)的有效性, 可使木质部的结构特征具有更强的抗栓塞能力.De Baerdemaeker等(2017)研究表明, Pwt产生的糖降低了木质部空穴化栓塞脆弱性.Schmitz等(2012)对红树林中的13种植物进行研究, 发现当Pwt受阻时, Kstem显著降低. ...
Woody tissue photosynthesis reduces stem CO2 efflux by half and remains unaffected by drought stress in young Populus tremula trees 1 2020b
... 长时间的干旱通常伴随着高温.在高温下, SRP比叶光合作用更有优势, 因为木质部和树皮组织对气体扩散的阻碍导致茎内部CO2浓度通常处于较高水平.这种CO2与O2的高比例补偿了核酮糖-1,5-二磷酸羧化酶随温度升高而下降的专一性.因此, 在高温下, 叶光合作用几乎停止时, SRP仍可继续提供C源(Cernusak & Cheesman, 2015; De Baerdemaeker et al., 2017).干旱期间, 当大气CO2吸收有限时, Pwt将更加重要.干旱会导致叶片到木质组织的韧皮部运输停滞, 因此, 即使叶片中仍有碳水化合物, 植物也难以将其输送到木质组织.此时, Pwt产生局部可用的碳水化合物, 可用于维持植物水力完整性(Sevanto et al., 2014; De Baerdemaeker et al., 2017; De Roo et al., 2020a).除局部呼吸CO2外, 木质部运输CO2的内部循环也对全碳同化有显著贡献, 这取决于小气候条件和控制蒸腾、呼吸和光合作用的生态生理因子.只要植物呼吸作用持续, 内部的CO2底物便可用于Pwt (Mitchell et al., 2013).因此, Pwt可以被认为是干旱期间碳循环的重要来源(De Roo et al., 2020b).Pwt提高了非结构性碳水化合物(NSCs)的有效性, 可使木质部的结构特征具有更强的抗栓塞能力.De Baerdemaeker等(2017)研究表明, Pwt产生的糖降低了木质部空穴化栓塞脆弱性.Schmitz等(2012)对红树林中的13种植物进行研究, 发现当Pwt受阻时, Kstem显著降低. ...
Native root xylem embolism and stomatal closure in stands of Douglas-fir and ponderosa pine: mitigation by hydraulic redistribution 2 2004
On the minimum leaf conductance: Its role in models of plant water use, and ecological and environmental controls 1 2019
... 叶片表皮导度在植物干旱响应模型中对冠层水通量的估算有重要影响, 目前已有研究讨论了已发表文献中不同方法所测量的叶片表皮导度在植物水分利用模型中的作用(Duursma et al., 2019; Lanning et al., 2020).但是环境条件的变化可能会导致表皮导度的变异, 这意味着目前的研究结果和模型可能低估了表皮导度在未来气候中的影响.因此, 在之后的研究中, 需要将表皮导度的测量和估算纳入研究范畴, 以更好地理解植物水分利用策略, 并更准确地预测森林对干旱胁迫的响应. ...
Cloud forest trees with higher foliar water uptake capacity and anisohydric behavior are more vulnerable to drought and climate change 2 2016
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
... ., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
Gas exchange and hydraulics during drought in crops: Who drives whom 1 2018
... 有研究发现, Kx和Kox均与gs和蒸腾速率(E)呈正相关关系, 但Kox与gs (或E)之间的相关性很弱, 茎水力导度(Kstem)与叶Kx呈正相关关系, 但与Kox不相关, 表明叶片Kox减弱了叶片水分运输和气体交换之间的协调能力以及茎和叶之间的水力联系, 提出木质部水分运输效率是gs和光合速率的重要决定因素(Liu et al., 2019b).叶柄和叶蒸发部位之间的水势梯度代表了水通过叶片输送的驱动力.一般来说, 由于叶木质部外水力阻力较高, 所以植物所需的木质部外水分运输驱动力(ΔΨox)比木质部水力运输驱动力(ΔΨx)更大.高ΔΨox能够补偿低Kox, 以达到所需的gs或E.此外, 在干旱胁迫条件下, 具有高ΔΨox的物种气孔关闭更早, 以防止Ψleaf过度下降和叶肉组织过度脱水(Liu et al., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
Grassland root functional parameters vary according to a community-level resource acquisition-conservation trade-off 2 2016
... 根系负责养分和水分的吸收, 并在缓解营养和水分胁迫对植物生长和物种间相互作用的影响方面发挥重要作用(Fort et al., 2016).根系深浅可以影响植物的抗旱性.一般来说, 在同一地区共存木本植物中, 根系分布较浅树种其栓塞抗性较强; 而根系分布较深的植物可以通过获取深层土壤水来缓冲植物水势的下降, 从而避免干旱损伤(Johnson et al., 2018).不同土壤水分利用对策对生长在干旱环境中植物的生长和生存至关重要(Chen et al., 2016).此外, 一些树种的主要水源在雨季为浅层土壤水, 在旱季则转而利用深层土壤水, 这是由于其根系特征的二态性决定的(Fort et al., 2017).浅层土壤中的细根一般直径较小, 根组织密度较低, 以适应较难预测的水分有效性, 而深层土壤中的细根的中柱较粗, 木质部管道较宽, 以减少水力阻力(Fort et al., 2016, 2017).这种物种间的水文生态位划分对减少竞争、提高生态系统中群落结构的稳定性起着至关重要的作用(Yang et al., 2015). ...
... ., 2016, 2017).这种物种间的水文生态位划分对减少竞争、提高生态系统中群落结构的稳定性起着至关重要的作用(Yang et al., 2015). ...
Root traits are related to plant water-use among rangeland Mediterranean species 2 2017
... 根系负责养分和水分的吸收, 并在缓解营养和水分胁迫对植物生长和物种间相互作用的影响方面发挥重要作用(Fort et al., 2016).根系深浅可以影响植物的抗旱性.一般来说, 在同一地区共存木本植物中, 根系分布较浅树种其栓塞抗性较强; 而根系分布较深的植物可以通过获取深层土壤水来缓冲植物水势的下降, 从而避免干旱损伤(Johnson et al., 2018).不同土壤水分利用对策对生长在干旱环境中植物的生长和生存至关重要(Chen et al., 2016).此外, 一些树种的主要水源在雨季为浅层土壤水, 在旱季则转而利用深层土壤水, 这是由于其根系特征的二态性决定的(Fort et al., 2017).浅层土壤中的细根一般直径较小, 根组织密度较低, 以适应较难预测的水分有效性, 而深层土壤中的细根的中柱较粗, 木质部管道较宽, 以减少水力阻力(Fort et al., 2016, 2017).这种物种间的水文生态位划分对减少竞争、提高生态系统中群落结构的稳定性起着至关重要的作用(Yang et al., 2015). ...
... , 2017).这种物种间的水文生态位划分对减少竞争、提高生态系统中群落结构的稳定性起着至关重要的作用(Yang et al., 2015). ...
Metrics and proxies for stringency of regulation of plant water status (iso/anisohydry): a global data set reveals coordination and trade-offs among water transport traits 3 2019
... 植物维持自身水力系统完整性的能力, 主要取决于其维管系统的物理极限以及植物维持自身水势的能力, 进而决定了植物在干旱胁迫期间的脱水速度以及相应的死亡阈值(Choat et al., 2018).叶膨压损失点水势(ΨTLP), 通常被用以直接量化植物叶片的耐旱性, 因为细胞膨压损失影响了细胞结构完整性、代谢和整个植物的性能, 是衡量植物生理耐旱性的经典指标.低ΨTLP的植物, 可以在较低叶水势(Ψleaf)的情况下仍然维持叶片膨压及其功能, 如光合作用和生长.此外, ΨTLP是与物种的相对等水程度关系最为密切的一个性状, ΨTLP越高, 植物越趋向于等水调节.Meinzer等(2016)建议使用ΨTLP作为衡量等水调节程度的指标, 并已经被大量物种的研究所支持(Fu & Meinzer, 2019; Li et al., 2019).Bartlett等(2016)发现ΨTLP与叶和枝水力导度下降50%的水势(P50)呈正相关关系.Johnson等(2018)发现, 在干旱季节中, 非等水的物种能够更有效地调节其ΨTLP.这种ΨTLP的变化也与叶P50的下降有关. ...
... 在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率.随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988).因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012).当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损.在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
... 部分植物同时表现出低水势和低Cstem, 这是由于植物器官栓塞抗性的强度与WD和细胞壁厚度相关.因为在一定的茎体积下, 较厚的导管(或管胞)壁和纤维壁会导致储水空间较小, 使得低水势物种具有较低的Cstem.因此, 这种机械联系导致P50和Cstem之间有很强的正相关关系(McCulloh et al., 2014).对很多物种来说, Cstem与由气孔调节所决定的日最小枝水势之间也存在着功能联系.高Cstem的物种往往更偏向等水调节, 但Fu等(2019)发现更偏向非等水的物种比等水调节物种有更大的Cleaf.因此, 叶和茎的水容可能呈负相关关系(Fu et al., 2019), 这可能是因为高Cstem的物种一般具有高Kstem, Kleaf和Kstem一般呈正相关关系, 而Kleaf越高的物种, 叶片越薄(Brodribb et al., 2007), 而较薄的叶片限制了其自身的Cleaf. ...
Coordination and trade-offs between leaf and stem hydraulic traits and stomatal regulation along a spectrum of isohydry to anisohydry 2 2019
... Cleaf的缓冲作用可以在不同层次上体现.在针叶树和蕨类植物中, 随VPD增加, Cleaf越低, 气孔关闭越快.这些植物类群一般表现为被动气孔控制(Brodribb & McAdam, 2011; Brodribb et al., 2014), 气孔直接响应于叶片的水分状况.因此, 单位蒸腾损失增加时, 低Cleaf可反映出较低的Ψleaf和高VPD诱发的气孔迅速关闭(Cardoso et al., 2019).Martins等(2016)发现, 在Cleaf较低的植物中, 气孔迅速关闭并没有阻止Ψleaf下降, 这里暗示了一种受低Cleaf的低缓冲能力所驱动的被动水力响应.而在被子植物中, gs下降50%所需的时间(T50)均高于针叶树和蕨类植物, 但T50和Cleaf之间的关系仍随其等水调节程度而不同(Fu et al., 2019).被子植物对水分亏缺的主动调节机制使其对Ψleaf下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
... 部分植物同时表现出低水势和低Cstem, 这是由于植物器官栓塞抗性的强度与WD和细胞壁厚度相关.因为在一定的茎体积下, 较厚的导管(或管胞)壁和纤维壁会导致储水空间较小, 使得低水势物种具有较低的Cstem.因此, 这种机械联系导致P50和Cstem之间有很强的正相关关系(McCulloh et al., 2014).对很多物种来说, Cstem与由气孔调节所决定的日最小枝水势之间也存在着功能联系.高Cstem的物种往往更偏向等水调节, 但Fu等(2019)发现更偏向非等水的物种比等水调节物种有更大的Cleaf.因此, 叶和茎的水容可能呈负相关关系(Fu et al., 2019), 这可能是因为高Cstem的物种一般具有高Kstem, Kleaf和Kstem一般呈正相关关系, 而Kleaf越高的物种, 叶片越薄(Brodribb et al., 2007), 而较薄的叶片限制了其自身的Cleaf. ...
2 2019
... 植物受干旱影响后的水力恢复速率在很大程度上取决于其水势下降的程度, 以及经历极限低水势的时间.轻度干旱胁迫通常不会导致高水平空穴化栓塞, 但叶脉中的导管塌缩和水通道蛋白对细胞膜通透性的调节机制可能导致植物水力运输能力瞬时且可逆的下降(Zhang et al., 2016).此时, 水力恢复会在降水后通过叶表面吸水(FWU)在很短的时间内发生(Binks et al., 2019; Fuenzalida et al., 2019).FWU是一种几乎在所有植物中都存在的水分获取机制(Dawson & Goldsmith, 2018; Berry et al., 2019a). ...
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
Individual traits as determinants of time to death under extreme drought in Pinus sylvestris L. 1 2016
... 水和碳的交互作用(Garcia-Forner et al., 2016), 以及干旱响应的时空格局(Martin-Stpaul et al., 2017)使植物死亡机制异常复杂.McDowell等(2008)的经典死亡机制模型认为等水调节物种更易死于碳饥饿.但在Adams等(2017)的研究中, 部分植物在干旱期间NSCs并不下降, 且有研究发现, 干旱期间等水调节程度更高的物种NSCs浓度更高, 而非等水调节物种的NSCs多用于渗透势的调节(Jiang et al.,2021).因此, 等水调节植物是否会受到更高的碳限制? 在水势值为多少时淀粉开始转化为可溶性糖?干旱期间有多少碳用于渗透调节, 又有多少碳用于呼吸作用? 这些问题都需要我们更深入地了解.此外, 植物水分生理学中对不同器官的研究有助于检验水力特征的协调与权衡, 而综合分析整株植物的水力学特征, 可以更全面地理解水对植物生长和生存的限制过程(Martinez-Vilalta et al., 2019; López et al., 2021).为了更好地理解植物水力学是如何影响生态系统功能的, 我们还需要将个体的结构和生理特征与群落和景观联系起来, 通过基于过程和 特征的模型提高评估和监测干旱诱导死亡风险的 能力. ...
Inferring foliar water uptake using stable isotopes of water 1 2017
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
Life in the treetops: ecophysiological strategies of canopy epiphytes in a tropical montane cloud forest 1 2015
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
Tree mortality across biomes is promoted by drought intensity, lower wood density and higher specific leaf area 1 2017
... 在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率.随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988).因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012).当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损.在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
A whole-plant perspective of isohydry: stem-level support for leaf-level plant water regulation 1 2021
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
Stomatal closure, basal leaf embolism, and shedding protect the hydraulic integrity of grape stems 2 2017
... 有研究发现, Kx和Kox均与gs和蒸腾速率(E)呈正相关关系, 但Kox与gs (或E)之间的相关性很弱, 茎水力导度(Kstem)与叶Kx呈正相关关系, 但与Kox不相关, 表明叶片Kox减弱了叶片水分运输和气体交换之间的协调能力以及茎和叶之间的水力联系, 提出木质部水分运输效率是gs和光合速率的重要决定因素(Liu et al., 2019b).叶柄和叶蒸发部位之间的水势梯度代表了水通过叶片输送的驱动力.一般来说, 由于叶木质部外水力阻力较高, 所以植物所需的木质部外水分运输驱动力(ΔΨox)比木质部水力运输驱动力(ΔΨx)更大.高ΔΨox能够补偿低Kox, 以达到所需的gs或E.此外, 在干旱胁迫条件下, 具有高ΔΨox的物种气孔关闭更早, 以防止Ψleaf过度下降和叶肉组织过度脱水(Liu et al., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
... 不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同.树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低.在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008).有两种假设解释了植物的这种保护机制.一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018).而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降.两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018).此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角. ...
Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought 1 2011
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
Coordination between leaf and stem traits related to leaf carbon gain and hydraulics across 32 drought-tolerant angiosperms 2 2008
... 植物对干旱胁迫的响应过程主要分为两个阶段(图1).第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017).在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降.植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017).随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度.气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019).在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014).尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018). ...
... 不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同.树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低.在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008).有两种假设解释了植物的这种保护机制.一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018).而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降.两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018).此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角. ...
Trade-offs between xylem water and carbohydrate storage among 24 coexisting subtropical understory shrub species spanning a spectrum of isohydry 2 2021
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
... 水和碳的交互作用(Garcia-Forner et al., 2016), 以及干旱响应的时空格局(Martin-Stpaul et al., 2017)使植物死亡机制异常复杂.McDowell等(2008)的经典死亡机制模型认为等水调节物种更易死于碳饥饿.但在Adams等(2017)的研究中, 部分植物在干旱期间NSCs并不下降, 且有研究发现, 干旱期间等水调节程度更高的物种NSCs浓度更高, 而非等水调节物种的NSCs多用于渗透势的调节(Jiang et al.,2021).因此, 等水调节植物是否会受到更高的碳限制? 在水势值为多少时淀粉开始转化为可溶性糖?干旱期间有多少碳用于渗透调节, 又有多少碳用于呼吸作用? 这些问题都需要我们更深入地了解.此外, 植物水分生理学中对不同器官的研究有助于检验水力特征的协调与权衡, 而综合分析整株植物的水力学特征, 可以更全面地理解水对植物生长和生存的限制过程(Martinez-Vilalta et al., 2019; López et al., 2021).为了更好地理解植物水力学是如何影响生态系统功能的, 我们还需要将个体的结构和生理特征与群落和景观联系起来, 通过基于过程和 特征的模型提高评估和监测干旱诱导死亡风险的 能力. ...
Conifers but not angiosperms exhibit vulnerability segmentation between leaves and branches in a temperate forest 1 2019
... 有研究表明, 物种的脆弱性分割程度受气候、树种等的影响.在半干旱、温带和季节性干旱环境中, 植物具有较强的脆弱性分割现象(Johnson et al., 2016), 而在湿润环境中缺少该机制(Zhu et al., 2016).Jin等(2019)在自然条件下的研究发现, 温带森林中针叶树种存在脆弱性分割机制, 而被子植物中不存在; 但是我们进一步的干旱控制试验研究发现, 温带阔叶树种水曲柳(Fraxinus mandshurica)存在脆弱性分割现象(未发表数据); 这可能与试验环境条件有关.在干旱环境下, 脆弱性分割有利于树木长期生存. ...
Co-ordinated performance of leaf hydraulics and economics in 10 Chinese temperate tree species 1 2016
... 不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同.树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低.在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008).有两种假设解释了植物的这种保护机制.一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018).而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降.两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018).此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角. ...
Co-occurring woody species have diverse hydraulic strategies and mortality rates during an extreme drought 3 2018
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
... 植物维持自身水力系统完整性的能力, 主要取决于其维管系统的物理极限以及植物维持自身水势的能力, 进而决定了植物在干旱胁迫期间的脱水速度以及相应的死亡阈值(Choat et al., 2018).叶膨压损失点水势(ΨTLP), 通常被用以直接量化植物叶片的耐旱性, 因为细胞膨压损失影响了细胞结构完整性、代谢和整个植物的性能, 是衡量植物生理耐旱性的经典指标.低ΨTLP的植物, 可以在较低叶水势(Ψleaf)的情况下仍然维持叶片膨压及其功能, 如光合作用和生长.此外, ΨTLP是与物种的相对等水程度关系最为密切的一个性状, ΨTLP越高, 植物越趋向于等水调节.Meinzer等(2016)建议使用ΨTLP作为衡量等水调节程度的指标, 并已经被大量物种的研究所支持(Fu & Meinzer, 2019; Li et al., 2019).Bartlett等(2016)发现ΨTLP与叶和枝水力导度下降50%的水势(P50)呈正相关关系.Johnson等(2018)发现, 在干旱季节中, 非等水的物种能够更有效地调节其ΨTLP.这种ΨTLP的变化也与叶P50的下降有关. ...
... 根系负责养分和水分的吸收, 并在缓解营养和水分胁迫对植物生长和物种间相互作用的影响方面发挥重要作用(Fort et al., 2016).根系深浅可以影响植物的抗旱性.一般来说, 在同一地区共存木本植物中, 根系分布较浅树种其栓塞抗性较强; 而根系分布较深的植物可以通过获取深层土壤水来缓冲植物水势的下降, 从而避免干旱损伤(Johnson et al., 2018).不同土壤水分利用对策对生长在干旱环境中植物的生长和生存至关重要(Chen et al., 2016).此外, 一些树种的主要水源在雨季为浅层土壤水, 在旱季则转而利用深层土壤水, 这是由于其根系特征的二态性决定的(Fort et al., 2017).浅层土壤中的细根一般直径较小, 根组织密度较低, 以适应较难预测的水分有效性, 而深层土壤中的细根的中柱较粗, 木质部管道较宽, 以减少水力阻力(Fort et al., 2016, 2017).这种物种间的水文生态位划分对减少竞争、提高生态系统中群落结构的稳定性起着至关重要的作用(Yang et al., 2015). ...
A test of the hydraulic vulnerability segmentation hypothesis in angiosperm and conifer tree species 1 2016
... 有研究表明, 物种的脆弱性分割程度受气候、树种等的影响.在半干旱、温带和季节性干旱环境中, 植物具有较强的脆弱性分割现象(Johnson et al., 2016), 而在湿润环境中缺少该机制(Zhu et al., 2016).Jin等(2019)在自然条件下的研究发现, 温带森林中针叶树种存在脆弱性分割机制, 而被子植物中不存在; 但是我们进一步的干旱控制试验研究发现, 温带阔叶树种水曲柳(Fraxinus mandshurica)存在脆弱性分割现象(未发表数据); 这可能与试验环境条件有关.在干旱环境下, 脆弱性分割有利于树木长期生存. ...
The variability of stomatal sensitivity to leaf water potential across tree species indicates a continuum between isohydric and anisohydric behaviours 1 2014
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
Xylem embolism refilling and resilience against drought- induced mortality in woody plants: processes and trade- offs 2 2018
... 许多水力安全阈值较窄的木本植物经常处于接近栓塞临界阈值的水势下, 当可利用水分增加时, 其水势和水分运输能力会迅速恢复(Ogasa et al., 2013), 表明很多植物具有水力恢复策略(Trifilò et al., 2015; Klein et al., 2018).有研究表明, 高水力风险的物种具有较高的水力恢复能力, 而水力对策较保守的物种可能并不依赖水力恢复对策(Ogasa et al., 2013).因此Klein等(2018)将恢复力作为一个额外的维度, 加入“安全-效率”权衡中, 形成一个新的“安全-效率-恢复”框架.了解在干旱胁迫下存活树木的水力恢复过程同样重要. ...
Water transport from stem to stomata: the coordination of hydraulic and gas exchange traits across 33 subtropical woody species 2 2019b
... 有研究发现, Kx和Kox均与gs和蒸腾速率(E)呈正相关关系, 但Kox与gs (或E)之间的相关性很弱, 茎水力导度(Kstem)与叶Kx呈正相关关系, 但与Kox不相关, 表明叶片Kox减弱了叶片水分运输和气体交换之间的协调能力以及茎和叶之间的水力联系, 提出木质部水分运输效率是gs和光合速率的重要决定因素(Liu et al., 2019b).叶柄和叶蒸发部位之间的水势梯度代表了水通过叶片输送的驱动力.一般来说, 由于叶木质部外水力阻力较高, 所以植物所需的木质部外水分运输驱动力(ΔΨox)比木质部水力运输驱动力(ΔΨx)更大.高ΔΨox能够补偿低Kox, 以达到所需的gs或E.此外, 在干旱胁迫条件下, 具有高ΔΨox的物种气孔关闭更早, 以防止Ψleaf过度下降和叶肉组织过度脱水(Liu et al., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
... ., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
Coordination of stem and leaf traits define different strategies to regulate water loss and tolerance ranges to aridity 3 2021
... 此外, 叶、茎和根性状的协调, 决定树木应对干旱胁迫的水力策略和耐受范围.Creek等(2018)发现, 导致Eucalyptus coolabah叶片气孔关闭的水势与根水力下降水势相吻合, 表明其中可能存在一种与气孔关闭相关的水力信号.López等(2021)的研究表明, 叶和茎的耐旱性既关联又相对独立, 其功能权衡取决于系统发育阶段和水分有效性.其中, 在茎叶性状变异轴上, 一端是具有较低的栓塞脆弱性、较高的水分利用效率和胡伯尔值(Hv, 茎边材面积与叶面积之比)以及气孔关闭较晚的“慢”对策物种; 而另一端是具有高栓塞脆弱性、低水分利用效率和Hv的“快”对策物种.这种生存对策的差异是由区域水分有效性所致.此外, 植物性状变异是多维的(Powell et al., 2017), 不同的维度允许不同功能性状组合, 因而解释了一些功能性状权衡未能反映在较低系统发育水平上的原因(Pivovaroff et al., 2014; López et al., 2021). ...
... ., 2021). ...
... 水和碳的交互作用(Garcia-Forner et al., 2016), 以及干旱响应的时空格局(Martin-Stpaul et al., 2017)使植物死亡机制异常复杂.McDowell等(2008)的经典死亡机制模型认为等水调节物种更易死于碳饥饿.但在Adams等(2017)的研究中, 部分植物在干旱期间NSCs并不下降, 且有研究发现, 干旱期间等水调节程度更高的物种NSCs浓度更高, 而非等水调节物种的NSCs多用于渗透势的调节(Jiang et al.,2021).因此, 等水调节植物是否会受到更高的碳限制? 在水势值为多少时淀粉开始转化为可溶性糖?干旱期间有多少碳用于渗透调节, 又有多少碳用于呼吸作用? 这些问题都需要我们更深入地了解.此外, 植物水分生理学中对不同器官的研究有助于检验水力特征的协调与权衡, 而综合分析整株植物的水力学特征, 可以更全面地理解水对植物生长和生存的限制过程(Martinez-Vilalta et al., 2019; López et al., 2021).为了更好地理解植物水力学是如何影响生态系统功能的, 我们还需要将个体的结构和生理特征与群落和景观联系起来, 通过基于过程和 特征的模型提高评估和监测干旱诱导死亡风险的 能力. ...
植物水分调节对策: 等水与非等水行为 1 2017
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
植物水分调节对策: 等水与非等水行为 1 2017
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
植物应对干旱胁迫的气孔调节 1 2019
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
植物应对干旱胁迫的气孔调节 1 2019
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
Plant resistance to drought depends on timely stomatal closure 4 2017
... 为了直接评估干旱胁迫下的气孔响应, Skelton等(2015)通过拟合气孔导度(gs)随Ψx变化的响应曲线来定义标准化的临界点.其中, 最大气孔导度下降50%时的切线与Ψx轴的交点被定义为气孔闭合点, 且该点对应的gs约损失了最大气孔导度的88%, 因此Ψgs88被定义为气孔关闭所对应的水势, 而Ψgs88与P50 (或P88)的差值被称为气孔安全阈值(SSM), 用于评价气孔和水力之间的协调能力(Skelton et al., 2015).该值越高, 表明植物气孔和水力之间的协调能力越强(Creek et al., 2018).有研究用SSM50 (Ψgs88与P50之差)作为指示植物等水调节程度的指标, 该值为正, 表明气孔关闭发生在水力导度下降到50%之前, 定义为等水调节行为; 而该值为负时则表明发生在P50之后, 定义为非等水调节行为.并认为等水调节物种比非等水调节物种具有更大的水力和气孔安全阈值(Skelton et al., 2015).此外, 有研究表明, 只有少数物种气孔关闭与木质部栓塞形成初期的水势相等, 且随植物栓塞抗性的增加, SSM增加(Martin-Stpaul et al., 2017).Chen等(2019)在对温带阔叶树种死亡率预测的研究中, 发现相比空穴化脆弱性(P50和P88)和HSM, SSM能更好地预测干旱胁迫下植物的死亡风险.但该指标目前使用并不广泛, 需要更多的研究加以验证. ...
... 植物对干旱胁迫的响应过程主要分为两个阶段(图1).第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017).在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降.植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017).随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度.气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019).在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014).尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018). ...
... ., 2017; Choat et al., 2018). ...
... 水和碳的交互作用(Garcia-Forner et al., 2016), 以及干旱响应的时空格局(Martin-Stpaul et al., 2017)使植物死亡机制异常复杂.McDowell等(2008)的经典死亡机制模型认为等水调节物种更易死于碳饥饿.但在Adams等(2017)的研究中, 部分植物在干旱期间NSCs并不下降, 且有研究发现, 干旱期间等水调节程度更高的物种NSCs浓度更高, 而非等水调节物种的NSCs多用于渗透势的调节(Jiang et al.,2021).因此, 等水调节植物是否会受到更高的碳限制? 在水势值为多少时淀粉开始转化为可溶性糖?干旱期间有多少碳用于渗透调节, 又有多少碳用于呼吸作用? 这些问题都需要我们更深入地了解.此外, 植物水分生理学中对不同器官的研究有助于检验水力特征的协调与权衡, 而综合分析整株植物的水力学特征, 可以更全面地理解水对植物生长和生存的限制过程(Martinez-Vilalta et al., 2019; López et al., 2021).为了更好地理解植物水力学是如何影响生态系统功能的, 我们还需要将个体的结构和生理特征与群落和景观联系起来, 通过基于过程和 特征的模型提高评估和监测干旱诱导死亡风险的 能力. ...
Greater focus on water pools may improve our ability to understand and anticipate drought-induced mortality in plants 2 2019
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
... 水和碳的交互作用(Garcia-Forner et al., 2016), 以及干旱响应的时空格局(Martin-Stpaul et al., 2017)使植物死亡机制异常复杂.McDowell等(2008)的经典死亡机制模型认为等水调节物种更易死于碳饥饿.但在Adams等(2017)的研究中, 部分植物在干旱期间NSCs并不下降, 且有研究发现, 干旱期间等水调节程度更高的物种NSCs浓度更高, 而非等水调节物种的NSCs多用于渗透势的调节(Jiang et al.,2021).因此, 等水调节植物是否会受到更高的碳限制? 在水势值为多少时淀粉开始转化为可溶性糖?干旱期间有多少碳用于渗透调节, 又有多少碳用于呼吸作用? 这些问题都需要我们更深入地了解.此外, 植物水分生理学中对不同器官的研究有助于检验水力特征的协调与权衡, 而综合分析整株植物的水力学特征, 可以更全面地理解水对植物生长和生存的限制过程(Martinez-Vilalta et al., 2019; López et al., 2021).为了更好地理解植物水力学是如何影响生态系统功能的, 我们还需要将个体的结构和生理特征与群落和景观联系起来, 通过基于过程和 特征的模型提高评估和监测干旱诱导死亡风险的 能力. ...
Water potential regulation, stomatal behaviour and hydraulic transport under drought: deconstructing the iso/anisohydric concept 2 2017
... 植物对干旱胁迫的响应过程主要分为两个阶段(图1).第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017).在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降.植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017).随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度.气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019).在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014).尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018). ...
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
Stomatal dynamics are limited by leaf hydraulics in ferns and conifers: results from simultaneous measurements of liquid and vapour fluxes in leaves 2 2016
... 除了gs和Kleaf之外, 叶水容(Cleaf)被认为是决定Ψleaf的第3个关键特征.水容(C)是植物组织储存水分并将其释放到蒸腾流的能力, 在蒸腾速率增加后, 可以暂时缓冲水势的变化(Tyree & Ewers, 1991; Sperry et al., 2008).Cleaf可以通过压力-体积曲线来确定.C对木质部压力的短暂缓冲效应可以用时间常数(τ)来量化, 该值为水力阻力和水容的乘积, 是VPD增加引起蒸腾速率增加后达到最终恒定水势值63%所需的时间(McCulloh et al., 2019).因此, 在VPD增加时, 高水容物种的τ较高, 缓冲效应持续时间较长, 并可通过降低气孔导度或改变液流离子浓度来提高局部水力导度(Martins et al., 2016; Roddy et al., 2018; McCulloh et al., 2019). ...
... Cleaf的缓冲作用可以在不同层次上体现.在针叶树和蕨类植物中, 随VPD增加, Cleaf越低, 气孔关闭越快.这些植物类群一般表现为被动气孔控制(Brodribb & McAdam, 2011; Brodribb et al., 2014), 气孔直接响应于叶片的水分状况.因此, 单位蒸腾损失增加时, 低Cleaf可反映出较低的Ψleaf和高VPD诱发的气孔迅速关闭(Cardoso et al., 2019).Martins等(2016)发现, 在Cleaf较低的植物中, 气孔迅速关闭并没有阻止Ψleaf下降, 这里暗示了一种受低Cleaf的低缓冲能力所驱动的被动水力响应.而在被子植物中, gs下降50%所需的时间(T50)均高于针叶树和蕨类植物, 但T50和Cleaf之间的关系仍随其等水调节程度而不同(Fu et al., 2019).被子植物对水分亏缺的主动调节机制使其对Ψleaf下降具有高敏感性(Brodribb & McAdam, 2011, 2017).高Cleaf会提供一种保护机制以避免Ψleaf下降, 另外, 末端叶脉塌陷导致Kleaf下降, 也能防止水分进一步流失, 避免Ψleaf下降(Zhang et al., 2016; Lechthaler et al., 2019). ...
Bark water uptake promotes localized hydraulic recovery in coastal redwood crown 2 2016
Recovery performance in xylem hydraulic conductivity is correlated with cavitation resistance for temperate deciduous tree species 3 2013
... 许多水力安全阈值较窄的木本植物经常处于接近栓塞临界阈值的水势下, 当可利用水分增加时, 其水势和水分运输能力会迅速恢复(Ogasa et al., 2013), 表明很多植物具有水力恢复策略(Trifilò et al., 2015; Klein et al., 2018).有研究表明, 高水力风险的物种具有较高的水力恢复能力, 而水力对策较保守的物种可能并不依赖水力恢复对策(Ogasa et al., 2013).因此Klein等(2018)将恢复力作为一个额外的维度, 加入“安全-效率”权衡中, 形成一个新的“安全-效率-恢复”框架.了解在干旱胁迫下存活树木的水力恢复过程同样重要. ...
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
Water relations of Calycanthus flowers: hydraulic conductance, capacitance, and embolism resistance 1 2018
... 除了gs和Kleaf之外, 叶水容(Cleaf)被认为是决定Ψleaf的第3个关键特征.水容(C)是植物组织储存水分并将其释放到蒸腾流的能力, 在蒸腾速率增加后, 可以暂时缓冲水势的变化(Tyree & Ewers, 1991; Sperry et al., 2008).Cleaf可以通过压力-体积曲线来确定.C对木质部压力的短暂缓冲效应可以用时间常数(τ)来量化, 该值为水力阻力和水容的乘积, 是VPD增加引起蒸腾速率增加后达到最终恒定水势值63%所需的时间(McCulloh et al., 2019).因此, 在VPD增加时, 高水容物种的τ较高, 缓冲效应持续时间较长, 并可通过降低气孔导度或改变液流离子浓度来提高局部水力导度(Martins et al., 2016; Roddy et al., 2018; McCulloh et al., 2019). ...
Mapping xylem failure in disparate organs of whole plants reveals extreme resistance in olive roots 2 2018
... 气孔关闭后, 水仍会通过气孔渗漏, 以及表皮和树皮等其他组织流失, 使植物水势持续缓慢下降.而植物也会释放内部储存水来进一步缓冲Ψx的下降.与此同时, 植物水力导度会通过一系列生物物理和生理机制的改变而下降, 包括叶脉塌缩、植物水通道蛋白调节、细根皮层腔隙的形成等(Choat et al., 2018).当水势下降到一个临界点(P50)时, 栓塞便会在木质部中迅速增长, 这一过程会发生在包括根茎叶在内的整个植物水力系统中(Creek et al., 2018; Rodriguez-Dominguez et al., 2018).由于气孔长期关闭, 光合作用受到抑制, 而呼吸作用还在继续, 从而消耗大量储存的碳水化合物.此外, 植物需要利用碳水化合物来驱动韧皮部运输、维持膨压, 以及重新填充栓塞的木质部, 因此维持呼吸所需的碳水化合物进一步减少.而水和碳的限制又逐渐增加了木质部栓塞的比例, 限制了对冠层的水分输送, 导致分枝斑块性死亡, 冠层叶片面积显著减少(McDowell et al., 2011).随着栓塞逐渐遍布整个输水网络, 造成植物水力系统不可逆的损伤(P88), 最终导致整个植株死亡(Anderegg et al., 2012; Choat et al., 2018).事实上, 在干旱胁迫下, 植物达到死亡临界阈值的概率和时间最终取决于相关生理和形态学特征的相互作用(图2).这种相互作用的多维性是维管植物能够生长在地球上几乎所有的陆地生态系统的有效策略, 并为抗旱提供了大量可能的形态和生理解决方案. ...
... 不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同.树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低.在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008).有两种假设解释了植物的这种保护机制.一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018).而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降.两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018).此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角. ...
The role of isohydric and anisohydric species in determining ecosystem-scale response to severe drought 1 2015
... 干旱导致树木死亡的生理机制假设主要有2种: 1)水力失衡假说: 干旱诱导的栓塞导致木质部水力功能部分或完全丧失, 阻碍水分运输, 从而导致树木因水力衰竭而死亡; 2)碳饥饿假说: 由于干旱引起气孔关闭, 促使碳水化合物供需失衡导致碳饥饿, 这将无法满足渗透、代谢和防御性碳的需要(McDowell et al., 2008, 2011, 2013; McDowell, 2011; Sevanto et al., 2014; Roman et al., 2015; Adams et al., 2017).但植物不同死亡机制是一个相互包容而非独立的过程(McDowell et al., 2011).大量研究表明, 水力失衡是干旱期间植被冠层枯梢或植物死亡的主要因子(Hoffmann et al., 2011; McDowell et al., 2011; Anderegg et al., 2012,2015, 2016); 但在水力系统还未被完全破坏的情况下, 植物应对水分运输能力降低的调节对策可能会导致储备碳水化合物因呼吸作用而被大量消耗, 使植物对病虫害胁迫的脆弱性增加, 最终导致死亡(McDowell et al., 2011).因此, 植物对干旱具有复杂的多维响应机制.鉴于未来极端干旱事件的频率和强度可能随温度的升高而增加, 迫切需要更好地了解不同植物的干旱响应策略差异, 以及不同植物水力特征对死亡时间和机制的影响(Choat et al., 2018).因此, 本文阐述了植物响应干旱胁迫的两个过程: 1)干旱胁迫期间植物的生理响应过程, 重点综述木本植物叶、茎和根水力性状的协调和变异, 以及对土壤-植物-大气连续体中水分运输的影响, 并评价衡量植物干旱抗性指标——气孔-水力安全阈值, 以提升对植物在干旱胁迫下死亡风险的预测; 2)干旱胁迫后的植物水力恢复过程, 主要包括树皮和叶表面吸水以及非结构性碳水化合物对栓塞修复的作用. ...
1 2016
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
Hydraulic acclimation in a Mediterranean oak subjected to permanent throughfall exclusion results in increased stem hydraulic capacitance 1 2020
... 茎水容(Cstem)为给定水势区间内水含量变化的比率, 是由水分释放曲线的斜率确定的(Sperry et al., 2008).茎水分释放曲线理论上分为3个阶段.第一阶段植物木质部水势相对较高, 释放的水来自细胞间隙和开放的导管、管胞和纤维中, 被称为毛细储水.毛细储水量主要取决于管腔空间, 尤其是纤维腔.纤维腔越大, 木质部密度越低, 因此毛细储水量与木质部密度成反比.第二阶段中, 水主要从薄壁组织、形成层和周皮组织等弹性活细胞中释放, 被称为弹性储水.此时曲线的斜率相对平缓, 且木质组织中弹性储水的体积一般很小.第三阶段的水主要来自栓塞的导管或管胞腔内, 所释放的水量主要取决于栓塞导管或管胞的数量和大小, 被称为空化储水(Pratt & Jacobsen, 2017).Cstem的范围在20- 500 kg·m-3·MPa-1, 主要受边材密度和含水量的影响, 反映了木质部水分运输受阻时蒸腾有效水分的来源(Pratt & Jacobsen, 2017).此外, Cstem还取决于树的大小.树越大, 边材体积越大, 储水空间越大, 且大树相对更依赖于储存水来满足蒸腾需求.表明Cstem是一种与生物量分配模式相关的可塑性特征(McCulloh et al., 2014).Cstem的缓冲作用可以避免植物水势大幅度波动达到不可逆值.高Cstem物种具有更高的Ψmin, 且木质部栓塞抗性与Cstem呈反比(Meinzer et al., 2009).因此, Cstem可能是与木质部安全-效率权衡相关的重要特征(Sperry et al., 2008; McCulloh et al., 2014; Pratt & Jacobsen, 2017).有研究认为, “避旱”树种WD低、Cstem高、栓塞脆弱性高, 而“抗旱”树种WD高、Cstem低, 且在低水势下仍能维持木质部功能(Meinzer et al., 2009; McCulloh et al., 2014).然而这种模式并不完全适用于种内水平.Barnard等(2011)的研究发现, 生长在干旱地区的花旗松(Pseudotsuga menziesii)和黄松(Pinus ponderosa)具有更高的Cstem和栓塞脆弱性, 并认为在较干燥的条件下Cstem的增加一方面可能部分解释了与木质部栓塞相关结构特征的适应性的不足, 另一方面则表明生物量分配的结构调整(如降低叶与边材面积比)是优化用水的关键策略.Salomón等(2020)在对Quercus ilex进行穿透雨排除实验后, 其水分释放曲线第一阶段的Cstem比对照组高, 而在其他阶段差异不显著.表明对于Q. ilex这样的耐旱树种而言, Cstem对水分亏缺的响应以优化碳增益为重. ...
Woody tissue photosynthesis and its contribution to trunk growth and bud development in young plants 1 2010
... 当叶片C同化和韧皮部C运输受限时, 茎光合作用成为一个重要的替代C源, 对树木在干旱胁迫下的生存至关重要(Cernusak & Cheesman, 2015; Vandegehuchte et al., 2015; De Roo et al., 2020a).Ávila等(2014)将茎表皮上通过气孔或皮孔固定大气CO2的净光合作用称为茎净光合作用(SNP), 而将非光合组织呼吸释放的CO2通过茎皮层组织再同化的过程称为茎循环光合作用(SRP).SNP通常需要植物茎的表皮上存在气孔或皮孔, 以促进CO2从大气扩散到茎皮层的光合细胞中, 一般存在于一些沙漠灌木和早期演替豆科植物中, 且与叶光合作用有相似的特征, 如相似的环境响应曲线, 包括光量子通量密度、胞间CO2浓度和VPD等(Ávila et al., 2014).SRP又称内部再同化作用, 是茎光呼吸速率和茎暗呼吸速率之差.茎内CO2部分源于活细胞的局部呼吸作用, 其中包括地下根呼吸产物通过蒸腾流向上输送.树木通过光合作用将这些CO2重新固定在含有叶绿素的细胞中, 这些细胞存在于树皮、木质部薄壁组织射线甚至茎髓中.Saveyn等(2010)将这个过程称为木质组织光合作用(Pwt), 以表达CO2被整个茎吸收的概念. ...
Species-specific reversal of stem xylem embolism after a prolonged drought correlates to endpoint concentration of soluble sugars 2 2016
... 尽管人们对栓塞导管再充水的实际情况持怀疑态度, 但仍有大量研究对相关机制进行了详细探索.研究发现, 木质部栓塞修复常常伴随着淀粉降解和糖在导管中的释放(Tomasella et al., 2017, 2019b; Wang et al., 2018a), 与此同时, 一些参与NSCs代谢和运输的基因以及水通道蛋白的表达均增强, 表明干旱复水后, 栓塞再充水是基于释放在管道中的糖和离子产生的渗透梯度驱动水流入木质部、韧皮部和薄壁组织完成的(Trifilò et al., 2019).在这种情况下, 淀粉降解似乎在提供渗透梯度方面发挥了关键作用.因此, NSCs贮存量可能直接影响干旱胁迫后植物的水力恢复能力(Tomasella et al., 2019a; Trifilò et al., 2019).Savi等(2016)发现, 被子植物茎木质部可溶性糖含量与水力恢复程度相关.针叶树种中, 水力恢复会伴随着茎木质部中NSCs的耗竭(Tomasella et al., 2017).因此, 高NSCs含量可以支持干旱胁迫缓解后的水力恢复, 而较低NSCs可利用性则会限制栓塞导管修复, 导致植物水力功能退化(Savi et al., 2016).此外, 有研究表明, 边材中淀粉和可溶性糖的相互转化与栓塞的形成和修复周期有关(Yoshimura et al., 2016).但Tomasella等(2019b)对欧洲山毛榉(Fagus sylvatica)的研究发现, 尽管该物种发生了淀粉到糖的转化, 但补水后栓塞导管并未恢复水力功能.而在第二次干旱循环后, 其木质部中NSCs总量较高.表明山毛榉不能从高水平栓塞恢复, 但多次干旱可能提高茎NSCs的可用性. ...
Direct uptake of canopy rainwater causes turgor-driven growth spurts in the mangrove Avicennia marina 1 2018
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
A case-study of water transport in co-occurring ring- versus diffuse-porous trees: contrasts in water-status, conducting capacity, cavitation and vessel refilling 1 2008
Variability among species of stomatal control under fluctuating soil water status and evaporative demand: modelling isohydric and anisohydric behaviours 1 1998
... 在陆地植物中, 由于绝大多数气体交换是在气孔中进行的, 因此水分流失和CO2吸收紧密耦合(Skelton et al., 2015).当气孔打开吸收CO2时, 水分就会流失, 这可能使植物接近水力功能障碍阈值, 导致水力失衡(Pivovaroff et al., 2018).因此, 气孔调节对植物水分状况以及水碳平衡具有重要意义(Meinzer et al., 2017; Creek et al., 2020).已有研究系统地综述了植物应对干旱胁迫的气孔调节机制, 并阐述了气孔调节的演化过程, 及其与水力调节的关系(罗丹丹等, 2019).植物在面对干旱导致的水力破坏的潜在风险时, 演化出不同的气孔调节策略.之前的研究以气孔动力学为基础, 将这种调节行为划分为等水调节与非等水调节.在大气水汽压亏缺(VPD)增加或土壤水势(ΨS)减少时, 等水调节植物通过严格的气孔调节, 迅速关闭气孔使最小叶水势(Ψleaf)维持相对恒定(罗丹丹等, 2017).但由于气孔完全关闭较早不能进行光合碳同化, 随着干旱的持续, 等水调节物种将更多地依赖储存的碳水化合物满足其呼吸、渗透调节或防御方面的碳需求, 因此在长期干旱胁迫下更容易死于碳饥饿.相反, 非等水植物气孔调节对环境变化不太敏感, 在干旱胁迫时, 气孔不会立刻关闭, 而是保持气体交换, 其Ψleaf随VPD的增加或ΨS减少而降低(Tardieu & Simonneau, 1998; Klein, 2014).由于非等水植物在干旱时气孔完全关闭较晚, 以水势下降为代价换取碳摄入, 因而更易死于水力失衡. ...
Non-structural carbohydrate and hydraulic dynamics during drought and recovery in Fraxinus ornus and Ostrya carpinifolia saplings 3 2019a
... 尽管人们对栓塞导管再充水的实际情况持怀疑态度, 但仍有大量研究对相关机制进行了详细探索.研究发现, 木质部栓塞修复常常伴随着淀粉降解和糖在导管中的释放(Tomasella et al., 2017, 2019b; Wang et al., 2018a), 与此同时, 一些参与NSCs代谢和运输的基因以及水通道蛋白的表达均增强, 表明干旱复水后, 栓塞再充水是基于释放在管道中的糖和离子产生的渗透梯度驱动水流入木质部、韧皮部和薄壁组织完成的(Trifilò et al., 2019).在这种情况下, 淀粉降解似乎在提供渗透梯度方面发挥了关键作用.因此, NSCs贮存量可能直接影响干旱胁迫后植物的水力恢复能力(Tomasella et al., 2019a; Trifilò et al., 2019).Savi等(2016)发现, 被子植物茎木质部可溶性糖含量与水力恢复程度相关.针叶树种中, 水力恢复会伴随着茎木质部中NSCs的耗竭(Tomasella et al., 2017).因此, 高NSCs含量可以支持干旱胁迫缓解后的水力恢复, 而较低NSCs可利用性则会限制栓塞导管修复, 导致植物水力功能退化(Savi et al., 2016).此外, 有研究表明, 边材中淀粉和可溶性糖的相互转化与栓塞的形成和修复周期有关(Yoshimura et al., 2016).但Tomasella等(2019b)对欧洲山毛榉(Fagus sylvatica)的研究发现, 尽管该物种发生了淀粉到糖的转化, 但补水后栓塞导管并未恢复水力功能.而在第二次干旱循环后, 其木质部中NSCs总量较高.表明山毛榉不能从高水平栓塞恢复, 但多次干旱可能提高茎NSCs的可用性. ...
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
Relationships between stomatal behavior, xylem vulnerability to cavitation and leaf water relations in two cultivars of Vitis vinifera 1 2014
... 有研究发现, Kx和Kox均与gs和蒸腾速率(E)呈正相关关系, 但Kox与gs (或E)之间的相关性很弱, 茎水力导度(Kstem)与叶Kx呈正相关关系, 但与Kox不相关, 表明叶片Kox减弱了叶片水分运输和气体交换之间的协调能力以及茎和叶之间的水力联系, 提出木质部水分运输效率是gs和光合速率的重要决定因素(Liu et al., 2019b).叶柄和叶蒸发部位之间的水势梯度代表了水通过叶片输送的驱动力.一般来说, 由于叶木质部外水力阻力较高, 所以植物所需的木质部外水分运输驱动力(ΔΨox)比木质部水力运输驱动力(ΔΨx)更大.高ΔΨox能够补偿低Kox, 以达到所需的gs或E.此外, 在干旱胁迫条件下, 具有高ΔΨox的物种气孔关闭更早, 以防止Ψleaf过度下降和叶肉组织过度脱水(Liu et al., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
Stomatal closure is induced by hydraulic signals and maintained by ABA in drought- stressed grapevine 1 2015
... 植物对干旱胁迫的响应过程主要分为两个阶段(图1).第一阶段是干旱胁迫开始到气孔闭合期间, 持续时间主要取决于植物的水分获取能力; 第二阶段是气孔闭合到木质部完全栓塞期间, 持续时间主要取决于植物水分维持能力及其耐旱特征(Martin- Stpaul et al., 2017).在干旱期间, 降水减少导致土壤湿度下降, 并伴随着温度的升高和大气蒸发需求的增加, 导致植物Ψx下降.植物应对干旱胁迫的第一道防线是气孔调节(Martínez-Vilalta & Garcia-Forner, 2017).随Ψx下降, 植物叶片失去膨压, 导致气孔关闭(Ψgs88), 这明显减缓了植物脱水和Ψx的下降速度.气孔关闭通常与水力功能失调的开始相吻合(Ishida et al., 2008; Tombesi et al., 2015; Buckley, 2019).在较短时间尺度内, 气孔关闭会导致叶片温度调节能力的下降和光损伤概率的增加(Leigh et al., 2017); 在较长时间下, 则会导致非结构性碳水化合物库的耗尽, 从而干扰糖在韧皮部转运, 以及抵抗食草动物和病原体所需的防御化合物的合成(McDowell et al., 2008; Mitchell et al., 2013; Sevanto et al., 2014).尽管如此, 等水调节植物气孔关闭仍发生在空穴化产生之前, 表明避免木质部空穴化栓塞对此类植物的长期生存至关重要(Skelton et al., 2015; Martin-Stpaul et al., 2017; Choat et al., 2018). ...
Effects of prolonged drought on stem non-structural carbohydrates content and post- drought hydraulic recovery in Laurus nobilis L.: the possible link between carbon starvation and hydraulic failure 2 2017
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
... ., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
Hydraulic recovery from xylem embolism in excised branches of twelve woody species: relationships with parenchyma cells and non-structural carbohydrates 3 2019
... 尽管人们对栓塞导管再充水的实际情况持怀疑态度, 但仍有大量研究对相关机制进行了详细探索.研究发现, 木质部栓塞修复常常伴随着淀粉降解和糖在导管中的释放(Tomasella et al., 2017, 2019b; Wang et al., 2018a), 与此同时, 一些参与NSCs代谢和运输的基因以及水通道蛋白的表达均增强, 表明干旱复水后, 栓塞再充水是基于释放在管道中的糖和离子产生的渗透梯度驱动水流入木质部、韧皮部和薄壁组织完成的(Trifilò et al., 2019).在这种情况下, 淀粉降解似乎在提供渗透梯度方面发挥了关键作用.因此, NSCs贮存量可能直接影响干旱胁迫后植物的水力恢复能力(Tomasella et al., 2019a; Trifilò et al., 2019).Savi等(2016)发现, 被子植物茎木质部可溶性糖含量与水力恢复程度相关.针叶树种中, 水力恢复会伴随着茎木质部中NSCs的耗竭(Tomasella et al., 2017).因此, 高NSCs含量可以支持干旱胁迫缓解后的水力恢复, 而较低NSCs可利用性则会限制栓塞导管修复, 导致植物水力功能退化(Savi et al., 2016).此外, 有研究表明, 边材中淀粉和可溶性糖的相互转化与栓塞的形成和修复周期有关(Yoshimura et al., 2016).但Tomasella等(2019b)对欧洲山毛榉(Fagus sylvatica)的研究发现, 尽管该物种发生了淀粉到糖的转化, 但补水后栓塞导管并未恢复水力功能.而在第二次干旱循环后, 其木质部中NSCs总量较高.表明山毛榉不能从高水平栓塞恢复, 但多次干旱可能提高茎NSCs的可用性. ...
... ., 2019).Savi等(2016)发现, 被子植物茎木质部可溶性糖含量与水力恢复程度相关.针叶树种中, 水力恢复会伴随着茎木质部中NSCs的耗竭(Tomasella et al., 2017).因此, 高NSCs含量可以支持干旱胁迫缓解后的水力恢复, 而较低NSCs可利用性则会限制栓塞导管修复, 导致植物水力功能退化(Savi et al., 2016).此外, 有研究表明, 边材中淀粉和可溶性糖的相互转化与栓塞的形成和修复周期有关(Yoshimura et al., 2016).但Tomasella等(2019b)对欧洲山毛榉(Fagus sylvatica)的研究发现, 尽管该物种发生了淀粉到糖的转化, 但补水后栓塞导管并未恢复水力功能.而在第二次干旱循环后, 其木质部中NSCs总量较高.表明山毛榉不能从高水平栓塞恢复, 但多次干旱可能提高茎NSCs的可用性. ...
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
Diurnal changes in embolism rate in nine dry forest trees: relationships with species-specific xylem vulnerability, hydraulic strategy and wood traits 2 2015
... 许多水力安全阈值较窄的木本植物经常处于接近栓塞临界阈值的水势下, 当可利用水分增加时, 其水势和水分运输能力会迅速恢复(Ogasa et al., 2013), 表明很多植物具有水力恢复策略(Trifilò et al., 2015; Klein et al., 2018).有研究表明, 高水力风险的物种具有较高的水力恢复能力, 而水力对策较保守的物种可能并不依赖水力恢复对策(Ogasa et al., 2013).因此Klein等(2018)将恢复力作为一个额外的维度, 加入“安全-效率”权衡中, 形成一个新的“安全-效率-恢复”框架.了解在干旱胁迫下存活树木的水力恢复过程同样重要. ...
... 另一方面, 具有低WD和高Cstem的物种, 水力恢复能力通常较强, 而WD较高的树种在补水后往往无法完全恢复其木质部功能(Ogasa et al., 2013; Trifilò et al., 2015).WD与Cstem呈负相关关系, 且与薄壁组织占木材体积的百分比有关(Pratt & Jacobsen, 2017).Trifilò等(2019)对12种木本植物栓塞恢复能力进行研究, 证明栓塞导管修复主要靠NSCs提供的渗透势推动, 且薄壁组织比例更高的物种, 水力恢复能力更强.表明薄壁组织丰度、NSCs含量和木质部栓塞恢复能力之间相互关联.Trifilò等(2017)在不同干旱胁迫时间下月桂(Laurus nobilis)的水力恢复和NSCs研究中发现, 长期干旱条件下, 植物NSCs库耗竭, 导致其水力恢复能力下降.而NSCs耗竭主要是由于光合作用对碳的吸收与对碳底物的需求之间的不平衡(Trifilò et al., 2017).对Fraxinus ornus和Ostrya carpinifolia的研究表明, 两物种在干旱胁迫和复水期间具有不同的水分调节对策和NSCs浓度(Tomasella et al., 2019a).Ostrya carpinifolia由于其更高的气孔敏感性(等水调节), 避免木质部栓塞增长.而非等水调节的Fraxinus ornus在干旱胁迫时水力导度下降, NSCs浓度升高, 而在补水后木质部水力功能完全恢复初始水平, 且仅在植株死亡时耗尽NSCs (Tomasella et al., 2019a).表明NSCs储量对干旱胁迫的响应具有物种特异性.根据McDowell等(2008)提出的框架, 干旱胁迫期间等水调节植物更容易发生碳饥饿, 而非等水调节植物更易死于栓塞导致的水力失衡.然而, 最近的研究发现, Cstem和NSCs存储量之间存在一种权衡, 且随等水调节程度的下降(Cstem下降), NSCs储存量耗损增加(Jiang et al., 2021).更趋向于等水调节的植物在干旱期间表现出NSCs的积累, 表明等水调节植物可能并不会受到更高的碳限制.相反地, 非等水调节植物可能更依赖于淀粉转化为糖, 以通过渗透调节改变ΨTLP来应对Ψleaf的下降(Martínez-Vilalta & Garcia-Forner, 2017; Hartmann et al., 2021).这为我们对整株植物水分调节和干旱机制的理解提供了一个新视角. ...
The contribution of vascular and extra-vascular water pathways to drought-induced decline of leaf hydraulic conductance 2 2016
... 有研究发现, Kx和Kox均与gs和蒸腾速率(E)呈正相关关系, 但Kox与gs (或E)之间的相关性很弱, 茎水力导度(Kstem)与叶Kx呈正相关关系, 但与Kox不相关, 表明叶片Kox减弱了叶片水分运输和气体交换之间的协调能力以及茎和叶之间的水力联系, 提出木质部水分运输效率是gs和光合速率的重要决定因素(Liu et al., 2019b).叶柄和叶蒸发部位之间的水势梯度代表了水通过叶片输送的驱动力.一般来说, 由于叶木质部外水力阻力较高, 所以植物所需的木质部外水分运输驱动力(ΔΨox)比木质部水力运输驱动力(ΔΨx)更大.高ΔΨox能够补偿低Kox, 以达到所需的gs或E.此外, 在干旱胁迫条件下, 具有高ΔΨox的物种气孔关闭更早, 以防止Ψleaf过度下降和叶肉组织过度脱水(Liu et al., 2019b).这可能表明相对较低的Kox、较高的ΔΨox和保守的气孔调节策略可能是保护茎、叶木质部减少栓塞的有效措施.因此, 在轻至中度干旱胁迫条件下, 气孔可能是防止水力失衡的安全阀(Hochberg et al., 2017).然而, Wang等(2018b)研究显示, Kleaf下降50%是在gs和叶肉导度(gm)下降50%之前发生的, 表明在水稻(Oryza sativa)中, 气孔没有起到安全阀的作用.Trifiló等(2016)和Scoffoni等(2017)研究表明, 大多数物种在达到ΨTLP之前, 木质部外的水力脆弱性能够解释Kleaf下降的75%-100%.因此他们认为Kleaf的变化主要是由Kox的变化所致(Trifilò et al., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
... ., 2016; Scoffoni et al., 2017).Albuquerque等(2020)的研究也表明, Kleaf的下降与叶片木质部栓塞无关.对此我们认为, 上述现象可能与植物应对水胁迫的不同水分调节策略有关(Tombesi et al., 2014; Flexas et al., 2018). ...
The hydraulic architecture of trees and other woody plants 2 1991
... 除了gs和Kleaf之外, 叶水容(Cleaf)被认为是决定Ψleaf的第3个关键特征.水容(C)是植物组织储存水分并将其释放到蒸腾流的能力, 在蒸腾速率增加后, 可以暂时缓冲水势的变化(Tyree & Ewers, 1991; Sperry et al., 2008).Cleaf可以通过压力-体积曲线来确定.C对木质部压力的短暂缓冲效应可以用时间常数(τ)来量化, 该值为水力阻力和水容的乘积, 是VPD增加引起蒸腾速率增加后达到最终恒定水势值63%所需的时间(McCulloh et al., 2019).因此, 在VPD增加时, 高水容物种的τ较高, 缓冲效应持续时间较长, 并可通过降低气孔导度或改变液流离子浓度来提高局部水力导度(Martins et al., 2016; Roddy et al., 2018; McCulloh et al., 2019). ...
... 不同植物叶、茎和根性状之间的协调性差异也会导致其水力模式的不同.树木茎木质部是一项长期投资, 碳水化合物投入成本较高, 但叶和细根的成本相对较低.在干旱胁迫下, 植物放弃末端器官(叶和细根), 减少蒸腾耗水, 有助于保持茎的水力完整性(Tyree & Ewers, 1991), 以便在干旱胁迫得到缓解后促进其生理机制的恢复(Ishida et al., 2008).有两种假设解释了植物的这种保护机制.一些研究认为, 叶和茎的栓塞脆弱性是相似的, 但叶片是植物水力运输的最远端, 因此Ψleaf要比Ψstem更负, 而植物不同器官的水势梯度差异会导致叶比茎更容易发生水力功能障碍(Skelton et al., 2018).而脆弱性分割假说则认为, 叶的栓塞脆弱性比茎更高, 这样可以保证在茎木质部出现大量栓塞前, 叶片的水力失衡导致整体失水速率迅速下降.两种假说都在一些研究中得到支持(Bartlett et al., 2016; Jin et al., 2016; Rodriguez-Dominguez et al., 2018; Skelton et al., 2018).此外, Hochberg等(2017)研究发现, 葡萄(Vitis vinifera)植株的基生叶、顶端叶和茎的栓塞脆弱性均差异显著, 且基生叶比顶端叶更容易栓塞, 支持了脆弱性分割假说, 并为不同位置叶片之间的分割增加了新的视角. ...
Do woody plants operate near the point of catastrophic xylem dysfunction caused by dynamic water stress? Answers from a model 1 1988
... 在干旱条件下, 植物木质部内的张力增加, 导致空穴化, 降低植物导水率.随着干旱加剧, 木质部空穴化的程度增加最终导致灾难性的水力失调(Tyree & Sperry, 1988).因此, 拟合植物导水率随木质部水势(Ψx)变化的关系所得到的木质部栓塞脆弱性曲线, 以及水力导度损失50%或88%时对应的水势(P50和P88)已成为与植物死亡率相关的关键生理指标(Brodribb & Cochard, 2009; Choat et al., 2012).当Ψx低于P50时, 木质部栓塞加速扩散, 水分运输功能明显受损.在脆弱性曲线中, 通过P50和栓塞扩散速率(斜率)可计算出木质部几乎完全失效的水势, 该水势下的导水率约占最大导水率的88% (Choat et al., 2012).P50和P88值越负, 木质部抵抗栓塞能力越强.研究表明, 不同树种P50和P88存在巨大差异, 并与其生长环境的年降水量和干旱程度有关(Choat et al., 2012).空穴化脆弱性的差异受木质部解剖学特征(导管或管胞的长度、直径和分布等)和纹孔结构(纹孔数量、纹孔膜厚度、孔隙度和纹孔腔深度等)(Lens et al., 2011)影响, 这些特征决定了气体通过纹孔膜, 导致相邻导管(或管胞)空穴化、栓塞在木质部中扩散的临界水势.然而, 脆弱性曲线测定方法比较繁琐, 且技术上具有挑战性, 因此, 相对较容易测量的木材密度(WD)被作为表征树木耐旱性的重要指标(Liang et al., 2021).研究表明, 在热带森林中, WD较高的树种, 不仅木质部饱和渗透势和膨压损失点渗透势均较低(De Guzman et al., 2021), 其叶ΨTLP亦较低(Fu & Meinzer, 2019), 表明高WD的植物能够忍受更低的水势, 且在全球范围内, WD较高的物种在干旱期间死亡率较低(Greenwood et al., 2017).但在相似的WD下, 裸子植物的致死水势要比被子植物更低.这种差异反映了两类植物不同木质部结构的功能特征(Liang et al., 2021).目前WD对致死水势的预测力仅在少数物种中进行了检验, 且数据主要来自幼树盆栽实验, 因此需要更多的工作来验证该模式的普遍性. ...
Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees 1 2013
... 随着全球气候变化的加剧, 极端气象事件频发、降雨格局变化等将加剧区域尺度的干旱程度, 造成全球范围内森林大面积死亡, 这对生态系统的功能和碳平衡产生了深远的影响(Urli et al., 2013; Allen et al., 2015; Anderegg et al., 2015; Clark et al., 2016; Choat et al., 2018; Martinez-Vilalta et al., 2019).其中, 植物水力系统失调, 是干旱期间树木死亡的主要机制(Anderegg et al., 2016; Adams et al., 2017).土壤水分有效性和大气蒸发需求变化显著影响植物水分运输能力, 因为叶片水分蒸腾和植物体内水分运输系统(从土壤到叶片)控制水分运输的效率.因此, 植物水力学可作为整合植物和生态系统功能的中心枢纽(Sack et al., 2016).然而, 树木对干旱胁迫的响应非常复杂, 不同树种在干旱胁迫期间的调节对策和水力特征变化很大, 甚至同一地区的共存物种也存在类似现象(Johnson et al., 2018).因此, 不同树木对干旱胁迫的响应对策, 以及不同植物水力特征如何延缓植物在干旱时死亡的时间, 降低干旱致死率等的机制备受关注(Choat et al., 2018). ...
Woody tissue photosynthesis in trees: salve on the wounds of drought 1 2015
... 当叶片C同化和韧皮部C运输受限时, 茎光合作用成为一个重要的替代C源, 对树木在干旱胁迫下的生存至关重要(Cernusak & Cheesman, 2015; Vandegehuchte et al., 2015; De Roo et al., 2020a).Ávila等(2014)将茎表皮上通过气孔或皮孔固定大气CO2的净光合作用称为茎净光合作用(SNP), 而将非光合组织呼吸释放的CO2通过茎皮层组织再同化的过程称为茎循环光合作用(SRP).SNP通常需要植物茎的表皮上存在气孔或皮孔, 以促进CO2从大气扩散到茎皮层的光合细胞中, 一般存在于一些沙漠灌木和早期演替豆科植物中, 且与叶光合作用有相似的特征, 如相似的环境响应曲线, 包括光量子通量密度、胞间CO2浓度和VPD等(Ávila et al., 2014).SRP又称内部再同化作用, 是茎光呼吸速率和茎暗呼吸速率之差.茎内CO2部分源于活细胞的局部呼吸作用, 其中包括地下根呼吸产物通过蒸腾流向上输送.树木通过光合作用将这些CO2重新固定在含有叶绿素的细胞中, 这些细胞存在于树皮、木质部薄壁组织射线甚至茎髓中.Saveyn等(2010)将这个过程称为木质组织光合作用(Pwt), 以表达CO2被整个茎吸收的概念. ...
Plant xylem hydraulics: What we understand, current research, and future challenges 1 2017
... 维管植物的叶片中, 水通过木质部导管在叶柄和整个叶脉网络中运动, 之后穿过维管束鞘、叶肉和表皮细胞, 扩散到细胞间隙, 最后通过气孔扩散到叶片外(Buckley et al., 2015).叶水力导度(Kleaf)代表了叶片中水分的运输效率, 包括叶木质部水力导度(Kx)和木质部外水力导度(Kox).木质部水分运输是水在内聚力-张力机制下通过负压在木质部中移动的过程.Kx主要由叶脉密度、排列方式、形状和导管直径和长度等结构特征所决定(Venturas et al., 2017).在干旱胁迫下, 木质部张力增加, 形成空穴化栓塞, 则导致Kx下降.木质部外水分运输路径较为复杂, 是水通过质外体或共质体途径在活细胞内移动的过程.该水分运输途径涉及多种组织, 而这些组织的导水阻力在很大程度上仍是未知的. ...
Effect of leaf water potential on internal humidity and CO2 dissolution: reverse transpiration and improved water use efficiency under negative pressure 2 2017
... 叶片吸水以液相和气相两种形式出现.如果水以气相进入叶片, 则需要叶表皮内细胞间隙具有低于周围空气的蒸汽压, 此时的FWU则是由水蒸气浓度梯度所驱动(Vesala et al., 2017).由于气孔相较于其他结构而言, 水的通量相对较高, 因此在气孔开放时, 该结构是FWU的主导途径, 且将这种现象称为“逆向蒸腾” (Binks et al., 2020).当气孔关闭时, 表皮是FWU的主要途径.不同的叶表皮特性(表皮蜡质层的化学结构、气孔结构、毛状体、刺毛和内生菌等)和叶结构(叶表皮、叶肉细胞和细胞间隙的大小和形状等)均会影响FWU (Boanares et al., 2018; Berry et al., 2019a).通过FWU进入叶片的水首先到达细胞间隙或表皮细胞, 并继续沿着水势梯度移动.在细胞内, 水分移动是通过质外体或共质体途径, 其中质外体途径占主导作用(Buckley et al., 2015).对于FWU来说, 气相水分输送可能是叶片内水分运动的常规途径, 并与细胞中的液相输送同时发生.之后, 这些水可以进入叶肉并用于光合作用或补充水容(Lehmann et al., 2018), 或进入导管系统(Berry et al., 2019a), 也有可能会蒸发到大气中.Steppe等(2018)发现, FWU在细胞膨压变化的驱动下诱导植物茎的径向生长, 表明FWU水也可以补充木材储水.Cassana等(2016)通过同位素示踪分析(δ2H)发现, 在干旱胁迫后的南洋杉(Araucaria angustifolia)中, 叶片吸收的水分可以通过木质部输送到靠近根部的土壤中, 以改善植物水分状况.但Berry等(2014)对冷杉(Abies fraseri)和云杉(Picea rubens)进行了类似的研究, 并未在土壤中发现FWU的水同位素信号(δ18O和δ2H).此外, 有研究认为叶片和大气之间可能会双向交换水同位素, 所以仅靠水同位素无法确定FWU的净吸收量, 需要开发更多维度和更高分辨率的方法来验证(Goldsmith et al., 2017; Berry et al., 2019a).Vesala等(2017)对红杉(Sequoia sempervirens)的研究中, 用蒸腾速率和FWU建立叶片水分的模型, 并发现FWU通量可以用Ψleaf来解释.因为Ψleaf降低会增加叶片和大气边界层的水势梯度, 所以较低的Ψleaf会导致FWU通量较高.但这里有一个潜在的权衡: Ψleaf较低时会导致叶片传导率(gs和gm)下降, 限制FWU.Gotsch等(2015)发现叶片较厚且Cleaf较大的物种FWU较低, 而高FWU的物种能承受较低的水势, 但其储水能力较弱.此外, Cleaf也会改变FWU的持续时间.在FWU条件下, Cleaf会影响水势的增长.高Cleaf物种水势变化较慢, 水势梯度持续时间较长, 导致FWU持续时间较长, 表明叶片水分储存和叶片吸水可能是相互联系的.Eller等(2016)发现, FWU能力较低的物种, 会更依赖替代策略, 如更等水的气孔调节, 以在干旱胁迫下维持叶膨压.而非等水调节物种则显示出更高的FWU频率, 并且FWU可作为一个短暂的水源, 以降低其水力风险.但同时, 非等水调节物种对FWU的依赖性使其更容易受到干旱和气候变化的影响(Eller et al., 2016; Berry et al., 2019b; Fuenzalida et al., 2019). ...
 ,, 王传宽
,, 王传宽 ,*, 金鹰
,*, 金鹰

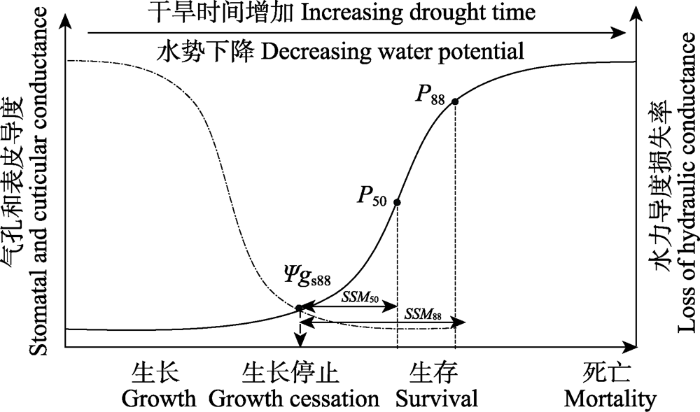 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT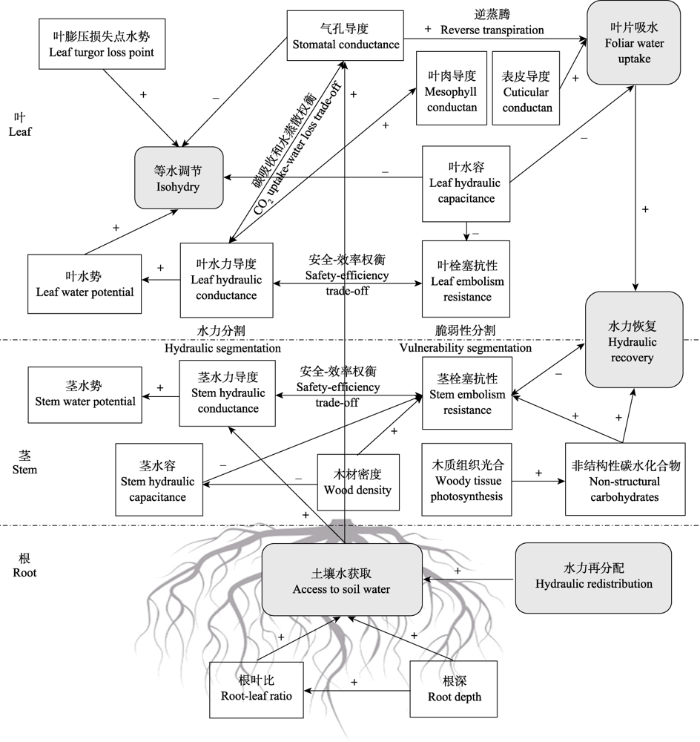 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}