Effects of non-structural carbohydrate and nitrogen allocation on the ability of Populus deltoides and P. cathayana to resist soil salinity stress
Xia-Zhen LIN1, Lin LIU2, Ting-Ting DONG2, Qi-Bo FANG2, Qing-Xue GUO,2,*1Teaching Center, Zhejiang Open University, Hangzhou 310012, China 2College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 311121, China
Abstract Aims The increasing level of soil salinization has been one of the most important factors to limit the development of forestry. The fast-growing Populusspp. are widely used for tree plantations and afforestation around the world and play crucial role in economic and ecological functions. Linking carbon and nitrogen metabolism with the resistance to soil salinity stress, will help to well develop the Populus plantations in salinization area. Methods The present study used P. deltoidesand P. cathayana for materials, while two salt (NaCl) concentrations and two defoliation treatments were applied. The carbon supply ability and allocation, nitrogen metabolism and allocation of the two poplar species were mainly investigated in different treatments. Important findings We found that the P. deltoides had higher total biomass and photosynthetic rate than P. cathayana under salinity stress. The chlorophyll concentration and the PSII maximum photochemical efficiency of P. deltoides were significantly higher than those of P. cathayana especially under defoliation with salinity stress, which demonstrated stronger damage on P. cathayana. The defoliation treatment aggravated the damage of NaCl on P. cathayana. The Na+ concentration in leaf and stem of P. deltoides was significantly lower than that of P. cathayana under salinity stress, demonstrating that the P. deltoides strongly restricted Na+ up-transport from root. Stem and root of P. deltoides had higher concentrations of starch, soluble sugars and sucrose than P. cathayana under salinity stress. The higher adenosine diphosphate glucose pyrophosphorylase activity facilitated the production of starch in P. deltoides than in P. cathayana. The defoliation greatly reduced the resistant ability of P. cathayana to salinity because of lower supply of non-structural carbohydrate to osmoregulation function. The allocation of nitrogen to sodium dodecyl sulfate-soluble protein of P. cathayana was significantly reduced by increasing salt, whereas NH4+ concentration, glutamate dehydrogenase activity and proline concentration were significantly higher than those of P. deltoides. Our results demonstrated the crucial role of non-structural carbohydrate of plant species in resisting soil salinity stress. Keywords:non-structural carbohydrate;osmoregulation;nitrogen allocation;Populus deltoides; Populus cathayana
PDF (4107KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文 引用本文 林夏珍, 刘林, 董婷婷, 方琦博, 郭庆学. 非结构性碳水化合物与氮分配对美洲黑杨和青杨耐盐能力的影响. 植物生态学报, 2021, 45(9): 961-971. DOI: 10.17521/cjpe.2021.0240 LIN Xia-Zhen, LIU Lin, DONG Ting-Ting, FANG Qi-Bo, GUO Qing-Xue. Effects of non-structural carbohydrate and nitrogen allocation on the ability of Populus deltoides and P. cathayana to resist soil salinity stress. Chinese Journal of Plant Ecology, 2021, 45(9): 961-971. DOI: 10.17521/cjpe.2021.0240
土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020)。土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a)。针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b)。然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮。
非结构性碳水化合物(淀粉与可溶性糖的总和)是植物赖以生长、生存和繁殖的能量物质, 可溶性糖为植物生长、呼吸、防御以及渗透调节等生理活动提供可直接利用的底物和能量(Hüve et al., 2012; Hartmann & Trumbore, 2016), 而淀粉则作为短暂或长期的贮藏碳源, 可水解为可溶性糖, 供给植物所需(MacNeill et al., 2017)。不同生长环境下, 植物光合固碳能力以及耗碳过程(如呼吸和分泌)共同决定了植物各器官中非结构性碳水化合物的浓度(Guo et al., 2020)。处于环境胁迫中的植物, 光合固碳能力下降, 但渗透调节能力以及抗氧化能力提高(陈少良等, 2001; Ottow et al., 2005; Guo et al., 2020)。许多研究发现植物主动提高体内非结构性碳水化合物的浓度, 以提高植物抗环境胁迫的能力。例如, 淀粉浓度的升高增强了木本植物的抗旱能力(MacNeill et al., 2017), 而可溶性糖浓度升高则提高了植物在盐胁迫条件下的渗透调节能力(陈少良等, 2001; Liu et al., 2021a)。
去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021)。植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018)。植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020)。作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011)。在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力。叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配。研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018)。植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020)。植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害。
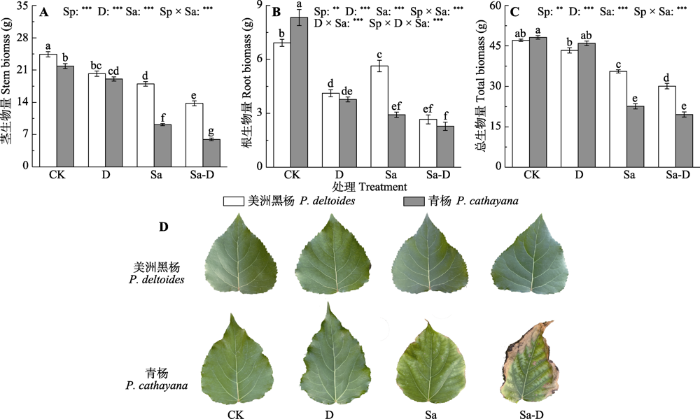
新窗口打开|下载原图ZIP|生成PPT 图1不同处理下美洲黑杨和青杨茎生物量(A)、根生物量(B)、总生物量(C)和叶片特征(D)(平均值±标准误)。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶; Sp, 物种。不同小写字母代表差异显著(p < 0.05)。**, 0.001 < p ≤0.01; ***, p ≤ 0.001。
Fig. 1Stem biomass (A), root biomass (B), total biomass (C) and leaf performance (D) of Populus deltoides and P. cathayana under different treatments (mean ± SE). CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). **, 0.001 < p ≤0.01; ***, p ≤ 0.001.
新窗口打开|下载原图ZIP|生成PPT 图2不同处理下美洲黑杨和青杨叶、茎和根中Na+与Cl-浓度(平均值±标准误)。由于青杨盐胁迫与去叶组根系生物量不足, 所以未测定根系Cl-浓度。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶; Sp, 物种。不同小写字母代表差异显著(p < 0.05)。*, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001。
Fig. 2Na+ and Cl- concentrations of leaf, stem and root in both Populus deltoides and P. cathayana under different treatments (mean ± SE). Root Cl- concentration was not tested because the root biomass of P. cathayana in the salt stress and defoliation treatment was not sufficient. CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
新窗口打开|下载原图ZIP|生成PPT 图3不同处理下美洲黑杨和青杨叶片光合与荧光特征(平均值±标准误)。Fv/Fm, 最大光量子效率; gs, 气孔导度; Pn, 净光合速率; YII, 实际光量子效率。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶; Sp, 物种。不同小写字母代表差异显著(p < 0.05)。*, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001。
Fig. 3The leaf photosynthetic and chlorophyll fluorescence traits of Populus deltoides and P. cathayana under different treatments (mean ± SE). Fv/Fm, maximum yield of primary photochemistry; gs, stomatal conductance; Pn, net photosynthetic rate; YII, effective quantum yield of PSII. CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
新窗口打开|下载原图ZIP|生成PPT 图4不同处理下美洲黑杨和青杨叶、茎和根淀粉浓度(平均值±标准误)。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶。不同小写字母代表差异显著(p < 0.05)。*, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001。
Fig. 4Leaf, stem and root starch concentration of Populus deltoides and P. cathayana under different treatments (mean ± SE). CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
Table 1 表1 表1不同处理条件下美洲黑杨与青杨叶、茎和根中可溶性糖与蔗糖浓度(平均值±标准误) Table 1Soluble sugars and sucrose concentration of leaf, stem and root in both Populus deltoides and P. cathayana under different treatments (mean ± SE)
处理 Treatment
可溶性糖 Soluble sugars (mg·g-1)
蔗糖 Sucrose (mg·g-1)
叶 Leaf
茎 Stem
根 Root
叶 Leaf
茎 Stem
根 Root
美洲黑杨P. deltoides
对照 Control (CK)
134.1 ± 4.4a
71.2 ± 2.2a
51.6 ± 1.3b
75.6 ± 3.9a
27.6 ± 1.9ab
27.6 ± 1.4b
去叶 Defoliation (D)
143.1 ± 4.6a
58.4 ± 1.9bc
33.0 ± 1.3e
73.8 ± 4.3a
26.5 ± 2.6abc
19.3 ± 1.4c
盐胁迫 Salt stress (Sa)
144.9 ± 6.0a
67.2 ± 3.0ab
60.3 ± 1.8a
79.4 ± 5.8a
35.0 ± 4.4a
37.7 ± 3.2a
盐胁迫与去叶 Salt stress and Defoliation (Sa-D)
149.0 ± 2.3a
54.1 ± 1.6c
45.1 ± 1.1bc
75.4 ± 3.9a
25.3 ± 3.2abc
22.0 ± 1.4bc
青杨P. cathayana
CK
144.6 ± 3.4a
59.7 ± 1.8bc
39.4 ± 1.5cd
61.0 ± 2.4ab
27.0 ± 3.3abc
19.4 ± 0.6c
D
141.6 ± 1.1a
44.4 ± 1.9d
38.7 ± 2.0cd
47.2 ± 2.8b
17.8 ± 2.0bc
21.2 ± 1.8bc
Sa
113.6 ± 5.5b
34.6 ± 1.4e
29.9 ± 3.4e
49.0 ± 8.1b
15.2 ± 1.6bc
14.5 ± 1.2cd
Sa-D
113.0 ± 4.2b
31.0 ± 1.1e
18.5 ± 1.2f
44.5 ± 3.4b
14.6 ± 1.2c
11.0 ± 1.4d
p
物种 Species (Sp)
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
D
0.432
<0.001
<0.001
0.076
0.012
<0.001
Sa
0.001
<0.001
0.094
0.483
0.264
0.636
Sp × D
0.168
0.216
<0.001
0.352
0.894
<0.001
Sp × Sa
<0.001
<0.001
<0.001
0.138
0.010
<0.001
D × Sa
0.833
0.044
0.174
0.598
1.000
0.012
Sp × D × Sa
0.549
0.035
0.010
0.388
0.034
0.676
若因子交互作用有显著性差异(p < 0.05), 则用Turkey分析进行多重比较, 同列中的不同小写字母表示显著性差异。 D, defoliation; Sa, salt stress; Sp, species. Different lowercase letters indicate significant differences among treatments according to Tukey's HSD test at a significance level of p < 0.05.
新窗口打开|下载原图ZIP|生成PPT 图5不同处理下美洲黑杨和青杨叶、根中腺苷二磷酸葡萄糖(ADPG)焦磷酸化酶活性(平均值±标准误)。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶; Sp, 物种。不同小写字母代表差异显著(p < 0.05)。*, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001。
Fig. 5Adenosine diphosphate glucose (ADPG) pyrophosphorylase activity in the leaf and root of Populus deltoides and P. cathayana under different treatments (mean ± SE). CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
新窗口打开|下载原图ZIP|生成PPT 图6不同处理下美洲黑杨与青杨叶氮稳定同位素比值(δ15N)(平均值±标准误)。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶; Sp, 物种。不同小写字母代表差异显著(p < 0.05)。*, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001。
Fig. 6Nitrogen stable isotope ratio (δ15N) in leaves of Populus deltoides and P. cathayana under different treatments (mean ± SE). CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
图7
新窗口打开|下载原图ZIP|生成PPT 图7不同处理下美洲黑杨和青杨叶与根中NH4+、NO3-和脯氨酸浓度(平均值±标准误)。CK, 对照; D, 去叶; Sa, 盐胁迫; Sa-D, 盐胁迫与去叶; Sp, 物种。不同小写字母代表差异显著(p < 0.05)。*, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001。
Fig. 7NH4+, NO3- and proline concentrations in the leaf and root of Populus deltoides and P. cathayana under different treatments (mean ± SE). CK, control; D, defoliation; Sa, salt stress; Sa-D, salt stress and defoliation; Sp, species. Different lowercase letters indicate significant differences among treatments (p < 0.05). *, 0.01 < p ≤ 0.05; **, 0.001 < p ≤ 0.01; ***, p ≤ 0.001.
Table 2 表2 表2不同处理条件下美洲黑杨与青杨叶和根谷氨酸脱氢酶与谷氨酰胺合成酶活性(平均值±标准误) Table 2Glutamate dehydrogenase (GDH) and glutamine synthase (GS) activity of leaf and root in both Populus deltoides and P. cathayana under different treatments (mean ± SE)
处理 Treatment
谷氨酸脱氢酶 GDH (U)
谷氨酰胺合成酶 GS (U)
叶 Leaf
根 Root
叶 Leaf
根 Root
美洲黑杨P. deltoides
对照 Control (CK)
202.1 ± 25.3cd
63.7 ± 7.7b
7.5 ± 1.7a
5.3 ± 0.5
去叶 Defoliation (D)
111.6 ± 26.2de
82.6 ± 4.9b
9.5 ± 1.6a
4.6 ± 0.4
盐胁迫 Salt stress (Sa)
411.9 ± 40.6b
129.3 ± 9.3a
11.3 ± 1.8a
4.6 ± 0.4
盐胁迫与去叶 Salt stress and Defoliation (Sa-D)
685.1 ± 22.6a
49.2 ± 9.4b
13.7 ± 1.5a
3.8 ± 0.2
青杨P. cathayana
CK
114.6 ± 15.2de
73.5 ± 7.1b
7.2 ± 0.9a
3.5 ± 0.3
D
58.3 ± 12.6e
85.7 ± 9.1b
7.9 ± 0.9a
3.7 ± 0.9
Sa
248.8 ± 36.7c
78.4 ± 11.4b
4.1 ± 0.5a
3.5 ± 0.2
Sa-D
248.1 ± 28.1c
78.2 ± 11.2b
5.8 ± 0.9a
3.8 ± 0.2
p
物种 Spices (Sp)
<0.001
0.726
<0.001
0.004
D
0.114
0.063
0.078
0.491
Sa
<0.001
0.253
0.454
0.242
Sp × D
0.004
0.007
0.582
0.136
Sp × Sa
<0.001
0.181
0.001
0.216
D × Sa
<0.001
<0.001
0.736
0.920
Sp × D × Sa
<0.001
0.002
0.880
0.888
若因子交互作用有显著性差异(p < 0.05), 则用Turkey分析进行多重比较, 同列中的不同小写字母表示显著性差异。 Different lowercase letters indicate significant differences among treatments according to Tukey's HSD test at a significance level of p < 0.05.
Supplement I Superoxide dismutase (SOD) and peroxidase (POD) activity in the leaf and root of Populus deltoides and P. cathayana under different treatments
AhmadP, HashemA, Abd-AllahEF, AlqarawiAA, JohnR, EgamberdievaD, GucelS (2015). Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system Frontiers in Plant Science, 6, 868. DOI: 10.3389/fpls.2015.00868. [本文引用: 1]
BaiAX, LuXY (2020). Effects of calcium and calcium effectors on antioxidant system and osmotic adjustment substances content of sour jujube (Ziziphus jujuba var. spinosa) seedlings under NaCl stress Plant Physiology Journal, 56, 1910-1920. [本文引用: 1]
BegumN, QinC, AhangerMA, RazaS, KhanMI, AshrafM, AhmedN, ZhangLX (2019). Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance Frontiers in Plant Science, 10, 1068. DOI: 10.3389/fpls.2019.01068. URL [本文引用: 1]
BehdadA, MohsenzadehS, AziziM (2021). Growth, leaf gas exchange and physiological parameters of two Glycyrrhiza glabra L. populations subjected to salt stress condition Rhizosphere, 17, 100319. DOI: 10.1016/j.rhisph.2021.100319. URL [本文引用: 1]
ChenJ, DongTF, DuanBL, KorpelainenH, NiinemetsÜ, LiCY (2015). Sexual competition and N supply interactively affect the dimorphism and competiveness of opposite sexes in Populus cathayana Plant, Cell & Environment, 38, 1285-1298. [本文引用: 2]
ChenSL, LiJK, BiWF, WangSS (2001). Genotypic variation in accumulation of salt ions betaine and sugars in poplar under conditions of salt stress Chinese Bulletin of Botany, 18, 587-596. [本文引用: 2]
EvansJR, SeemannJR (1989). The allocation of protein nitrogen in the photosynthetic apparatus: costs, consequences and control Plant Biology, 183-205. [本文引用: 1]
FaseelaP, SinishaAK, BrestičM, PuthurJT (2019). Chlorophyll a fluorescence parameters as indicators of a particular abiotic stress in rice Photosynthetica, 58, 293-3005. DOI:10.32615/ps.2019.147URL [本文引用: 1]
HüveK, BicheleI, IvanovaH, KeerbergO, ParnikT, RasulovB, TobiasM, NiinemetsÜ (2012). Temperature responses of dark respiration in relation to leaf sugar concentration Physiologia Plantarum, 144, 320-334. DOI:10.1111/ppl.2012.144.issue-4URL [本文引用: 1]
LinSJ, SunM (2017). Analysis of physiological response and salt tolerance mechanism of Crossostephium chinense and four species of Chrysanthemum under salt stress Acta Botanica Boreali-Occidentalia Sinica, 37, 1137-1144. [本文引用: 1]
LiuM, KorpelainenH, LiCY (2021a). Sexual differences and sex ratios of dioecious plants under stressful environments Journal of Plant Ecology, 14, 920-933. DOI:10.1093/jpe/rtab038URL [本文引用: 2]
LiuM, LiuXC, DuXH, KorpelainenH, NiinemetsÜ, LiCY (2021b). Anatomical variation of mesophyll conductance due to salt stress in Populus cathayana females and males growing under different inorganic nitrogen sources Tree Physiology, 41, 1462-1478. DOI:10.1093/treephys/tpab017URL [本文引用: 2]
LuoJ, ZhouJ, LiH, ShiWG, PolleA, LuMZ, SunXM, LuoZB (2015). Global poplar root and leaf transcriptomes reveal links between growth and stress responses under nitrogen starvation and excess Tree Physiology, 35, 1283-1302. DOI:10.1093/treephys/tpv091URL [本文引用: 1]
MacneillGJ, MehrpouyanS, MinowMAA, PattersonJA, TetlowIJ, EmesMJ (2017). Starch as a source, starch as a sink: the bifunctional role of starch in carbon allocation Journal of Experimental Botany, 68, 4433-4453. DOI:10.1093/jxb/erx291URL [本文引用: 4]
MeuriotF, Morvan-BertrandA, Noiraud-RomyN, DecauML, Escobar-GutiérrezAJ, GastalF, Prud'hommeMP (2018). Short-term effects of defoliation intensity on sugar remobilization and N fluxes in ryegrass Journal of Experimental Botany, 69, 3975-3986. DOI:10.1093/jxb/ery211URL [本文引用: 3]
MunnsR, TesterM (2008). Mechanisms of salinity tolerance Annual Review of Plant Biology, 59, 651-681. DOI:10.1146/annurev.arplant.59.032607.092911PMID:18444910 [本文引用: 1] The physiological and molecular mechanisms of tolerance to osmotic and ionic components of salinity stress are reviewed at the cellular, organ, and whole-plant level. Plant growth responds to salinity in two phases: a rapid, osmotic phase that inhibits growth of young leaves, and a slower, ionic phase that accelerates senescence of mature leaves. Plant adaptations to salinity are of three distinct types: osmotic stress tolerance, Na(+) or Cl() exclusion, and the tolerance of tissue to accumulated Na(+) or Cl(). Our understanding of the role of the HKT gene family in Na(+) exclusion from leaves is increasing, as is the understanding of the molecular bases for many other transport processes at the cellular level. However, we have a limited molecular understanding of the overall control of Na(+) accumulation and of osmotic stress tolerance at the whole-plant level. Molecular genetics and functional genomics provide a new opportunity to synthesize molecular and physiological knowledge to improve the salinity tolerance of plants relevant to food production and environmental sustainability.
OttowEA, BrinkerM, TeichmannT, FritzE, KaiserW, Brosche?M, Kangasja?rviJ, JiangXN, PolleA (2005). Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress Plant Physiology, 139, 1762-1772. PMID:16299175 [本文引用: 2] Populus euphratica Olivier is known to exist in saline and arid environments. In this study we investigated the physiological mechanisms enabling this species to cope with stress caused by salinity. Acclimation to increasing Na+ concentrations required adjustments of the osmotic pressure of leaves, which were achieved by accumulation of Na+ and compensatory decreases in calcium and soluble carbohydrates. The counterbalance of Na+/Ca2+ was also observed in mature leaves from field-grown P. euphratica trees exposed to an environmental gradient of increasing salinity. X-ray microanalysis showed that a primary strategy to protect the cytosol against sodium toxicity was apoplastic but not vacuolar salt accumulation. The ability to cope with salinity also included maintenance of cytosolic potassium concentrations and development of leaf succulence due to an increase in cell number and cell volume leading to sodium dilution. Decreases in apoplastic and vacuolar Ca2+ combined with suppression of calcineurin B-like protein transcripts suggest that Na+ adaptation required suppression of calcium-related signaling pathways. Significant increases in galactinol synthase and alternative oxidase after salt shock and salt adaptation point to shifts in carbohydrate metabolism and suppression of reactive oxygen species in mitochondria under salt stress.
SatheeL, JhaSK, RajputOS, SinghD, KumarS, KumarA (2021). Expression dynamics of genes encoding nitrate and ammonium assimilation enzymes in rice genotypes exposed to reproductive stage salinity stress Plant Physiology and Biochemistry, 165, 161-172. DOI:10.1016/j.plaphy.2021.05.013URL [本文引用: 2]
SecchiF, ZwienieckiMA (2011). Sensing embolism in xylem vessels: the role of sucrose as a trigger for refilling Plant, Cell & Environment, 34, 514-524. [本文引用: 1]
TakashimaT, HikosakaK, HiroseT (2004). Photosynthesis or persistence: nitrogen allocation in leaves of evergreen and deciduous Quercus species Plant, Cell & Environment, 27, 1047-1054. [本文引用: 2]
WangZQ, YuanYZ, OuJQ, LinQH, ZhangCF (2007). Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat (Triticum aestivum) seedlings exposed to different salinity Journal of Plant Physiology, 164, 695-701. DOI:10.1016/j.jplph.2006.05.001URL [本文引用: 1]
WenXM, XianT, LiuJY, XuX, DongTF (2021). Effects of defoliation on growth and non-structural carbohydrates in female and male Populus cathayana. Chinese Journal of Ecology, 40, 41-48. [本文引用: 1]
WuXY, LiuJT, MengQQ, FangSY, KangJY, GuoQX (2021). Differences in carbon and nitrogen metabolism between male and female Populus cathayana in response to deficient nitrogen Tree Physiology, 41, 119-133. DOI:10.1093/treephys/tpaa108URL [本文引用: 4]
XuXH, DiaoHJ, QinCY, HaoJ, ShenY, DongKH, WangCH (2021). Response of soil net nitrogen mineralization to different levels of nitrogen addition in a saline-alkaline grassland of northern China Chinese Journal of Plant Ecology, 45, 85-95. DOI:10.17521/cjpe.2020.0153URL [本文引用: 1]
ZhaoR, ChenSL (2020). The salt-stress signaling network involved in the regulation of ionic and ROS homeostasis in poplar Scientia Sinica (Vitae), 50, 167-175. [本文引用: 3]
ZhouHH, LiWH (2015). Responses and adaptation of xylem hydraulic conductivity to salt stress in Populus euphratica Chinese Journal of Plant Ecology, 39, 81-91. DOI:10.17521/cjpe.2015.0009URL [本文引用: 2]
[ 周洪华, 李卫红 (2015). 胡杨木质部水分传导对盐胁迫的响应与适应 植物生态学报, 39, 81-91.] DOI:10.17521/cjpe.2015.0009 [本文引用: 2] 解析植物木质部导水率对逆境的响应和适应对促进植物抗逆性机理研究和受损植被恢复具有重要意义。该文以荒漠河岸林建群种胡杨(Populus euphratica)为研究对象, 系统分析了胡杨幼株根、茎、叶水分传输通道对不同浓度盐胁迫的响应和适应。结果表明: (1)胡杨幼株根系对盐胁迫的敏感性高于茎和叶, 盐胁迫下根系生长和根尖数显著受到抑制, 根木质部易于发生栓塞, 导水率明显降低。(2)胡杨幼株茎木质部导水率对盐胁迫的响应依盐浓度而定, 轻度(0.05 mol·L<sup>-1</sup> NaCl)和中度(0.15 mol·L<sup>-1</sup> NaCl)盐胁迫下, 胡杨可以通过协调导管输水的有效性和安全性来调节木质部的导水率, 维持植物正常生长; 重度(0.30 mol·L<sup>-1</sup> NaCl)盐胁迫下, 胡杨茎木质部导管输水有效性和安全性均明显降低, 木质部导水率显著下降, 并伴随叶片气孔导度的显著降低, 从而严重抑制了胡杨的光合和生长。 Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system 1 2015
... 土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020).土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a).针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b).然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮. ...
钙和钙效应剂对NaCl胁迫下酸枣幼苗抗氧化系统及渗透调节物质含量的影响 1 2020
... 去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021).植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018).植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020).作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011).在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力.叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配.研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018).植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020).植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害. ...
钙和钙效应剂对NaCl胁迫下酸枣幼苗抗氧化系统及渗透调节物质含量的影响 1 2020
... 去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021).植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018).植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020).作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011).在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力.叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配.研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018).植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020).植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害. ...
Role of arbuscular mycorrhizal fungi in plant growth regulation: implications in abiotic stress tolerance 1 2019
... 土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020).土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a).针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b).然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮. ...
Growth, leaf gas exchange and physiological parameters of two Glycyrrhiza glabra L. populations subjected to salt stress condition 1 2021
Photosynthesis, water use efficiency and stable carbon isotope composition are associated with anatomical properties of leaf and xylem in six poplar species 1 2012
Stronger intra- specific competition aggravates negative effects of drought on the growth of Cunninghamia lanceolata 6 2020
... 非结构性碳水化合物(淀粉与可溶性糖的总和)是植物赖以生长、生存和繁殖的能量物质, 可溶性糖为植物生长、呼吸、防御以及渗透调节等生理活动提供可直接利用的底物和能量(Hüve et al., 2012; Hartmann & Trumbore, 2016), 而淀粉则作为短暂或长期的贮藏碳源, 可水解为可溶性糖, 供给植物所需(MacNeill et al., 2017).不同生长环境下, 植物光合固碳能力以及耗碳过程(如呼吸和分泌)共同决定了植物各器官中非结构性碳水化合物的浓度(Guo et al., 2020).处于环境胁迫中的植物, 光合固碳能力下降, 但渗透调节能力以及抗氧化能力提高(陈少良等, 2001; Ottow et al., 2005; Guo et al., 2020).许多研究发现植物主动提高体内非结构性碳水化合物的浓度, 以提高植物抗环境胁迫的能力.例如, 淀粉浓度的升高增强了木本植物的抗旱能力(MacNeill et al., 2017), 而可溶性糖浓度升高则提高了植物在盐胁迫条件下的渗透调节能力(陈少良等, 2001; Liu et al., 2021a). ...
... ., 2020).许多研究发现植物主动提高体内非结构性碳水化合物的浓度, 以提高植物抗环境胁迫的能力.例如, 淀粉浓度的升高增强了木本植物的抗旱能力(MacNeill et al., 2017), 而可溶性糖浓度升高则提高了植物在盐胁迫条件下的渗透调节能力(陈少良等, 2001; Liu et al., 2021a). ...
... 去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021).植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018).植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020).作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011).在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力.叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配.研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018).植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020).植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害. ...
Temperature responses of dark respiration in relation to leaf sugar concentration 1 2012
... 非结构性碳水化合物(淀粉与可溶性糖的总和)是植物赖以生长、生存和繁殖的能量物质, 可溶性糖为植物生长、呼吸、防御以及渗透调节等生理活动提供可直接利用的底物和能量(Hüve et al., 2012; Hartmann & Trumbore, 2016), 而淀粉则作为短暂或长期的贮藏碳源, 可水解为可溶性糖, 供给植物所需(MacNeill et al., 2017).不同生长环境下, 植物光合固碳能力以及耗碳过程(如呼吸和分泌)共同决定了植物各器官中非结构性碳水化合物的浓度(Guo et al., 2020).处于环境胁迫中的植物, 光合固碳能力下降, 但渗透调节能力以及抗氧化能力提高(陈少良等, 2001; Ottow et al., 2005; Guo et al., 2020).许多研究发现植物主动提高体内非结构性碳水化合物的浓度, 以提高植物抗环境胁迫的能力.例如, 淀粉浓度的升高增强了木本植物的抗旱能力(MacNeill et al., 2017), 而可溶性糖浓度升高则提高了植物在盐胁迫条件下的渗透调节能力(陈少良等, 2001; Liu et al., 2021a). ...
盐胁迫下芙蓉菊与4种菊属植物生理响应特征及其耐盐机理分析 1 2017
... 土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020).土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a).针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b).然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮. ...
盐胁迫下芙蓉菊与4种菊属植物生理响应特征及其耐盐机理分析 1 2017
... 土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020).土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a).针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b).然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮. ...
Sexual differences and sex ratios of dioecious plants under stressful environments 2 2021a
... 土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020).土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a).针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b).然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮. ...
... 非结构性碳水化合物(淀粉与可溶性糖的总和)是植物赖以生长、生存和繁殖的能量物质, 可溶性糖为植物生长、呼吸、防御以及渗透调节等生理活动提供可直接利用的底物和能量(Hüve et al., 2012; Hartmann & Trumbore, 2016), 而淀粉则作为短暂或长期的贮藏碳源, 可水解为可溶性糖, 供给植物所需(MacNeill et al., 2017).不同生长环境下, 植物光合固碳能力以及耗碳过程(如呼吸和分泌)共同决定了植物各器官中非结构性碳水化合物的浓度(Guo et al., 2020).处于环境胁迫中的植物, 光合固碳能力下降, 但渗透调节能力以及抗氧化能力提高(陈少良等, 2001; Ottow et al., 2005; Guo et al., 2020).许多研究发现植物主动提高体内非结构性碳水化合物的浓度, 以提高植物抗环境胁迫的能力.例如, 淀粉浓度的升高增强了木本植物的抗旱能力(MacNeill et al., 2017), 而可溶性糖浓度升高则提高了植物在盐胁迫条件下的渗透调节能力(陈少良等, 2001; Liu et al., 2021a). ...
Anatomical variation of mesophyll conductance due to salt stress in Populus cathayana females and males growing under different inorganic nitrogen sources 2 2021b
... 土壤盐渍化严重阻碍了我国林业的可持续发展, 气候变化以及不科学的管理方式导致盐渍化土壤面积不断扩大, 成为困扰我国的主要生态问题(林双冀和孙明, 2017; 赵瑞和陈少良, 2020).土壤盐分升高降低了土壤水势和孔隙度, 限制植物根系生长以及对水分和养分的吸收(Ahmad et al., 2015; 徐小惠等, 2021), 同时过量积累的Na+或Cl-破坏植物膜系统, 导致光合色素降解, 光合作用能力下降(Liu et al., 2021a).针对土壤盐胁迫, 植物可以通过提高抗氧化水平降低活性氧物质的毒害, 也可以通过渗透调节、排Na+或者将Na+区隔化到液泡里等方式适应盐胁迫或者提高耐盐能力(Begum et al. 2019; 赵瑞和陈少良, 2020; Liu et al. 2021b).然而, 植物适应盐胁迫或者提高耐盐能力的生理活动依赖于植物自身可提供的能量与底物, 比如碳和氮. ...
Populus euphratica displays apoplastic sodium accumulation, osmotic adjustment by decreases in calcium and soluble carbohydrates, and develops leaf succulence under salt stress 2 2005
... 非结构性碳水化合物(淀粉与可溶性糖的总和)是植物赖以生长、生存和繁殖的能量物质, 可溶性糖为植物生长、呼吸、防御以及渗透调节等生理活动提供可直接利用的底物和能量(Hüve et al., 2012; Hartmann & Trumbore, 2016), 而淀粉则作为短暂或长期的贮藏碳源, 可水解为可溶性糖, 供给植物所需(MacNeill et al., 2017).不同生长环境下, 植物光合固碳能力以及耗碳过程(如呼吸和分泌)共同决定了植物各器官中非结构性碳水化合物的浓度(Guo et al., 2020).处于环境胁迫中的植物, 光合固碳能力下降, 但渗透调节能力以及抗氧化能力提高(陈少良等, 2001; Ottow et al., 2005; Guo et al., 2020).许多研究发现植物主动提高体内非结构性碳水化合物的浓度, 以提高植物抗环境胁迫的能力.例如, 淀粉浓度的升高增强了木本植物的抗旱能力(MacNeill et al., 2017), 而可溶性糖浓度升高则提高了植物在盐胁迫条件下的渗透调节能力(陈少良等, 2001; Liu et al., 2021a). ...
... 非结构性碳水化合物为合成渗透物质提供底物和能量(MacNeill et al., 2017), 同时影响植物的氮吸收能力(Guo et al., 2017; Wu et al. 2021).研究发现, 非结构性碳水化合物浓度降低后植物吸收氮的能力也明显降低(Guo et al., 2017).盐胁迫和去叶导致青杨非结构性碳水化合物浓度严重下降, 限制N的吸收, 青杨更依赖贮藏的氮源, 如NH4+、NO3-、蛋白质和氨基酸, 因此会有较高的δ15N.叶中的氮可简单地分为脂不溶性蛋白质氮(主要分布在细胞壁)、脂溶性蛋白质氮(主要分布在膜系统、基粒和基质中)和水溶性蛋白质氮(如Rubisco酶)(Evans & Seemann, 1989; Takashima et al., 2004).在盐胁迫下青杨叶绿素浓度以及PSII反应中心Fv/Fm降低, 表明植物的膜系统蛋白质结构受到了破坏或水解, 导致脂溶性蛋白质氮浓度降低.蛋白质和氨基酸也是植物体内重要的渗透调节物质(Sathee et al., 2021).盐胁迫下青杨显著提高了脯氨酸的浓度, 而膜系统相关蛋白的水解可能为脯氨酸的合成提供了氮源.研究发现对盐敏感或耐盐性较差的植物, 高的谷氨酸脱氢酶(GDH)活性与脯氨酸浓度成正相关关系(Sathee et al., 2021).NH4+过高可引起植物细胞基质内pH失衡以及破坏光和磷酸化过程.NH4+的同化依赖于谷氨酰胺合成酶途径, 即在谷氨酰胺合成酶(GS)作用下生产谷氨酰胺, 或依赖于谷氨酸脱氢酶途径, 在GDH的作用下以NAD(P)H为氢供体, 形成谷氨酸.Wang等(2007)发现盐胁迫下NH4+的同化依赖于GDH-NAD(P)H途径, 而谷氨酸是脯氨酸合成的前体, 因为高GDH活性使得青杨中积累较多的脯氨酸.美洲黑杨叶和根中的脯氨酸浓度并没有随着GDH活性的提高而增加, 这可能是因为美洲黑杨可溶性糖(包括蔗糖)浓度较高, 满足了其渗透调节功能, 而氮则用于积累生物量而不是增加脯氨酸的合成. ...
Glutamine synthetase and glutamate dehydrogenase contribute differentially to proline accumulation in leaves of wheat (Triticum aestivum) seedlings exposed to different salinity 1 2007
... 非结构性碳水化合物为合成渗透物质提供底物和能量(MacNeill et al., 2017), 同时影响植物的氮吸收能力(Guo et al., 2017; Wu et al. 2021).研究发现, 非结构性碳水化合物浓度降低后植物吸收氮的能力也明显降低(Guo et al., 2017).盐胁迫和去叶导致青杨非结构性碳水化合物浓度严重下降, 限制N的吸收, 青杨更依赖贮藏的氮源, 如NH4+、NO3-、蛋白质和氨基酸, 因此会有较高的δ15N.叶中的氮可简单地分为脂不溶性蛋白质氮(主要分布在细胞壁)、脂溶性蛋白质氮(主要分布在膜系统、基粒和基质中)和水溶性蛋白质氮(如Rubisco酶)(Evans & Seemann, 1989; Takashima et al., 2004).在盐胁迫下青杨叶绿素浓度以及PSII反应中心Fv/Fm降低, 表明植物的膜系统蛋白质结构受到了破坏或水解, 导致脂溶性蛋白质氮浓度降低.蛋白质和氨基酸也是植物体内重要的渗透调节物质(Sathee et al., 2021).盐胁迫下青杨显著提高了脯氨酸的浓度, 而膜系统相关蛋白的水解可能为脯氨酸的合成提供了氮源.研究发现对盐敏感或耐盐性较差的植物, 高的谷氨酸脱氢酶(GDH)活性与脯氨酸浓度成正相关关系(Sathee et al., 2021).NH4+过高可引起植物细胞基质内pH失衡以及破坏光和磷酸化过程.NH4+的同化依赖于谷氨酰胺合成酶途径, 即在谷氨酰胺合成酶(GS)作用下生产谷氨酰胺, 或依赖于谷氨酸脱氢酶途径, 在GDH的作用下以NAD(P)H为氢供体, 形成谷氨酸.Wang等(2007)发现盐胁迫下NH4+的同化依赖于GDH-NAD(P)H途径, 而谷氨酸是脯氨酸合成的前体, 因为高GDH活性使得青杨中积累较多的脯氨酸.美洲黑杨叶和根中的脯氨酸浓度并没有随着GDH活性的提高而增加, 这可能是因为美洲黑杨可溶性糖(包括蔗糖)浓度较高, 满足了其渗透调节功能, 而氮则用于积累生物量而不是增加脯氨酸的合成. ...
去叶对青杨雌雄植株生长和非结构性碳水化合物的影响 1 2021
... 去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021).植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018).植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020).作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011).在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力.叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配.研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018).植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020).植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害. ...
去叶对青杨雌雄植株生长和非结构性碳水化合物的影响 1 2021
... 去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021).植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018).植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020).作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011).在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力.叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配.研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018).植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020).植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害. ...
Differences in carbon and nitrogen metabolism between male and female Populus cathayana in response to deficient nitrogen 4 2021
... 去叶是研究植物碳代谢特征以及模拟食叶害虫对植物碳平衡影响的重要措施(Meuriot et al., 2018; 文小梅等, 2021; Wu et al., 2021).植物叶片遭受较为严重的破坏或损失后, 短时间内光合固碳能力急剧下降甚至停滞(Meuriot et al., 2018).植物呼吸、渗透调节和防御等则依赖于体内贮藏的非结构性碳水化合物(MacNeill et al., 2017; Guo et al., 2020).作为植物体内光合碳运输的主要形式, 蔗糖是植物感知环境胁迫的重要信号, 也是修复植物损伤的关键物质(Secchi & Zwieniecki, 2011).在土壤盐胁迫下, 植物叶片遭受严重损失, 非结构性碳水化合物浓度高的植物可能具有维持较高水平的渗透调节和抗氧化能力.叶片损失不仅影响植物光合碳固定以及输出, 也影响了植物对氮的吸收和分配.研究发现植物叶片损失后, 根系吸收氮的速率明显下降(Meuriot et al., 2018).植物需要主动积累氨基酸和蛋白质等物质, 以此来适应盐胁迫或增强耐盐能力(白爱兴和鲁晓燕, 2020).植物叶片损失不仅导致植物氮储量减少, 同时还降低根系的氮吸收速率, 可能使得植物积累渗透物质所需的氮源不足, 最终导致耐盐能力弱的植物遭受更严重的盐毒害. ...
 ,2,*
,2,*

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT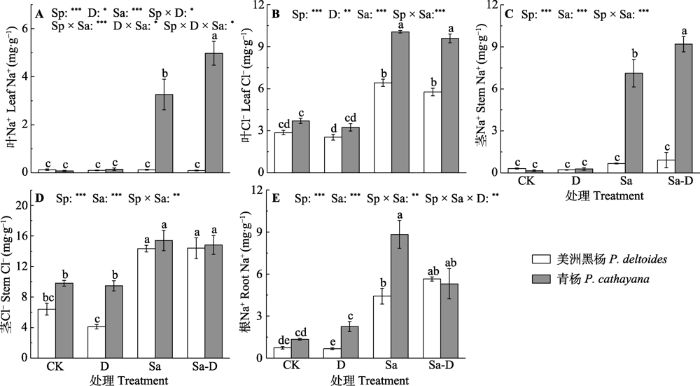 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT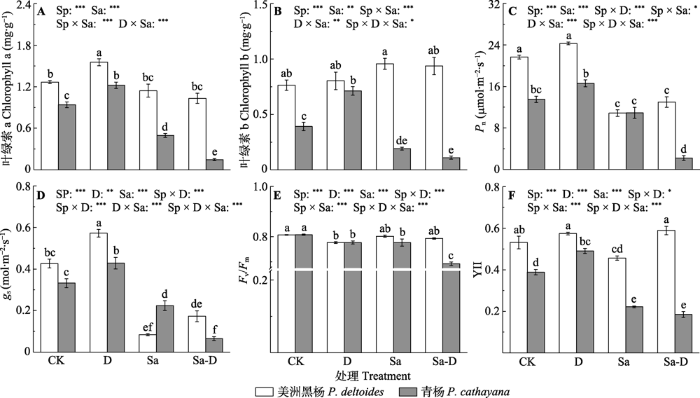 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT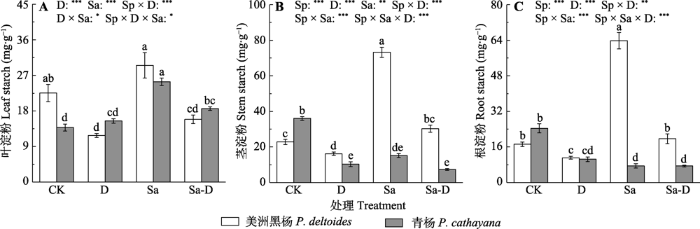 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT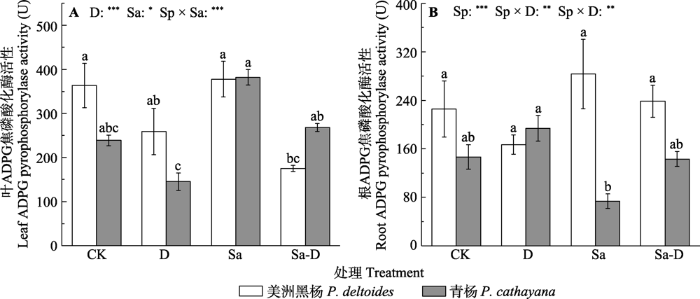 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT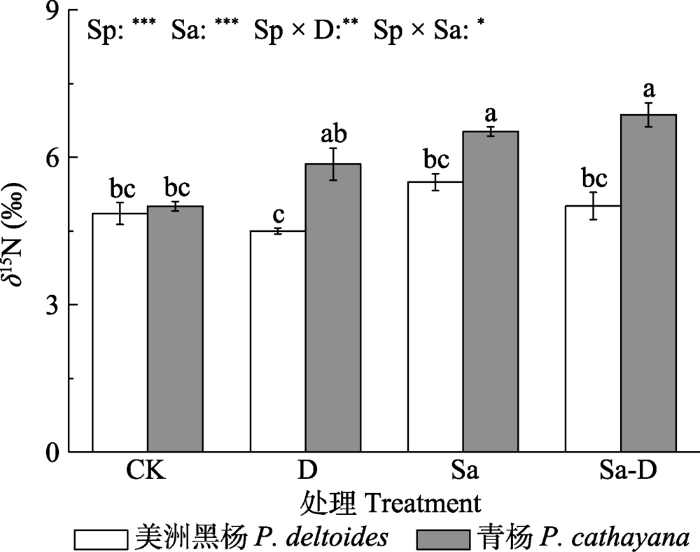 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT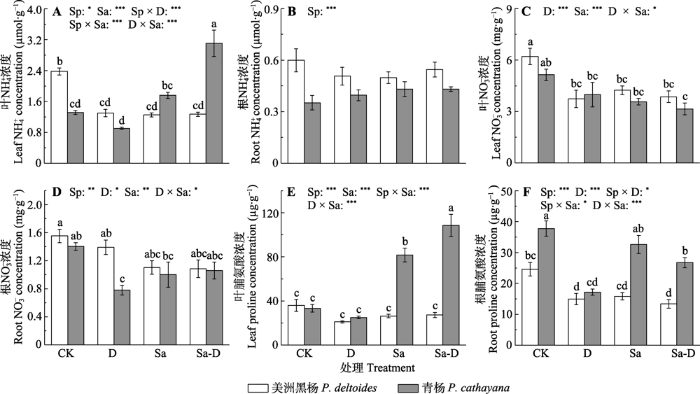 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}