王春成
1 , 张云玲
2 , 马松梅
, , 3 , * , 黄刚
1 , 张丹
1 , 闫涵
3 1 石河子大学生命科学学院, 绿洲城镇与山盆系统生态兵团重点实验室, 新疆石河子 8320002 新疆维吾尔自治区草原总站, 乌鲁木齐 8300493 石河子大学理学院, 绿洲城镇与山盆系统生态兵团重点实验室, 新疆石河子 832000Phylogeny and species differentiation of four wild almond species of subgen. Amygdalus in China Chun-Cheng WANG
1 , Yun-Ling ZHANG
2 , Song-Mei MA
, , 3 , * , Gang HUANG
1 , Dan ZHANG
1 , Han YAN
3 1 Key Laboratory of Ecological Corps for Oasis City and Mountain Basin System, College of Life Sciences, Shihezi University, Shihezi, Xinjiang 832000, China2 General Grassland Station of Xinjiang, Ürümqi 830049, China3 Key Laboratory of Ecological Corps for Oasis City and Mountain Basin System, College of Science, Shihezi University, Shihezi, Xinjiang 832000, China通讯作者: *马松梅: ORCID:0000-0002-3107-2256,(
shzmsm@126.com )
编委: 李镇清
责任编辑: 李敏
收稿日期: 2020-11-9
接受日期: 2021-06-2
基金资助: 国家自然科学基金 (41261011 )国家自然科学基金 (41561007 )新疆维吾尔自治区草原生态修复治理补助项目 (XJCYZZ202007 )
Corresponding authors: *(
shzmsm@126.com )
Received: 2020-11-9
Accepted: 2021-06-2
Fund supported: National Natural Science Foundation of China (41261011 )National Natural Science Foundation of China (41561007 )Grassland Ecological Restoration and Management Grant Project of Xinjiang Uygur Autonomous Region (XJCYZZ202007 )
摘要 基于核基因ITS序列研究中国4种野生扁桃: 新疆野扁桃(
Amygdalus ledebouriana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )和西康扁桃(
A. tangutica )的系统发育关系和物种分化, 为4种植物的遗传与演化研究提供数据支撑。利用单倍型网络和主坐标分析揭示单倍型的聚类关系; 利用最大似然树和贝叶斯系统树分析单倍型的系统发育关系; 利用R语言“ecospat”包分析4种扁桃的生态位分化及其环境驱动因子。4种扁桃ITS1-ITS4片段总长为634 bp, 鉴别出27个核苷酸变异位点, 共定义了28个单倍型。4种扁桃种间最小遗传距离均大于种内最大遗传距离, 种间存在显著的遗传分化。4种扁桃的单倍型聚为两支: 新疆野扁桃、蒙古扁桃和西康扁桃聚为一支, 长柄扁桃为另一支; 单倍型网络和主坐标分析揭示的单倍型聚类关系与系统树一致。西康扁桃与蒙古扁桃、与长柄扁桃之间均表现出了显著的生态位分化, 最暖月最高气温、年平均气温、最冷月最低气温和最暖季降水量是驱动物种生态位分化的关键因子。
关键词: 野生扁桃 ;
ITS序列 ;
系统发育 ;
物种分化 ;
生态位分化 Abstract Aims Based on the ITS sequences, we aimed to analyze the spatial genetic structure, genealogy relationship, and species differentiation of the
Amygdalus ledebouriana, A. mongolica, A. pedunculata, A. tangutica in China, and provide data for the future studies on the four species' genetics and evolution.
Methods The median-joining network and principal coordinate analysis (PCoA) were used to reveal haplotype clustering. The maximum likelihood method and Bayesian method were used to analyze the phylogenetic relationships of haplotypes. The “ecospat” package in R 4.0.2 was used to analyze the ecological niche divergence of four almond species and their environmental drivers.
Important findings The total length of the ITS1-ITS4 fragment after corrected alignment was 634 bp, 27 nucleotide variants detected, and a total of 28 haplotypes were identified. The minimum genetic distance among the four almond species is greater than the maximum genetic distance within species, and there are significant genetic differentiations among species. The haplotypes of the four almond species clustered into two branches:
A. ledebouriana, A. mongolica and
A. tangutica for one clade, and
A. pedunculata for the other. The revealed dendrogram relationship of haplotype network and PCoA analysis is consistent with the phylogenetic tree. The significant niche divergence was observed between
A. tangutica and
A. mongolica, as well as between
A. tangutica and
A. pedunculata, with annual mean temperature, max temperature of warmest month, min temperature of coldest month and precipitation of warmest quarter as key drivers of niche divergence.
Keywords: wild almond species ;
ITS sequences ;
phylogeny ;
species differentiation ;
niche divergence PDF (25618KB) 元数据 多维度评价 相关文章 导出 EndNote |
Ris |
Bibtex 收藏本文 引用本文 王春成, 张云玲, 马松梅, 黄刚, 张丹, 闫涵. 中国扁桃亚属四种野生扁桃的系统发育与物种分化. 植物生态学报, 2021, 45(9): 987-995. DOI:
10.17521/cjpe.2020.0366 WANG Chun-Cheng, ZHANG Yun-Ling, MA Song-Mei, HUANG Gang, ZHANG Dan, YAN Han.
Phylogeny and species differentiation of four wild almond species of subgen. Amygdalus in China .
Chinese Journal of Plant Ecology , 2021, 45(9): 987-995. DOI:
10.17521/cjpe.2020.0366 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 )。李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 )。扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 )。在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 )。具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 )。4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值。
对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 )。上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树。我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈。
核糖体DNA内转录间隔区(ITS)序列为双亲遗传, 具有长度保守性和高度变异性, 能有效地反映种间杂交或基因渐渗, 被称为研究属、种和亚种间遗传分化和系统发育关系的通用工具(于华会等,
2010 )。但要注意ITS序列在使用中易出现多倍体杂合位点问题。生态位相似性分析, 分析并可视化物种间的生态位宽度和生态位关系, 有助于识别驱动物种分化与进化的生态驱动力(Slatyer
et al .,
2013 )。若异域分布的姐妹种在环境空间上完全分离, 那么生态因素很可能在物种形成过程中扮演了主要作用, 而如果姐妹种种间具有相同或相似的环境空间, 生态因素则可能只是地理隔离导致的附带分歧(Peterson
et al .,
1999 )。本研究基于核基因ITS序列, 利用群体遗传、系统发育及生态位相似性分析方法, 研究我国4种野生扁桃的种间系统关系和物种分化。旨在解决以下科学目标: 1)基于ITS序列变异的4种扁桃的单倍型系统发育关系与物种分化; 2) 4种扁桃物种间的生态位分化及其环境驱动力; 3)生态因素对4种扁桃物种间的谱系分化与进化所起的作用。
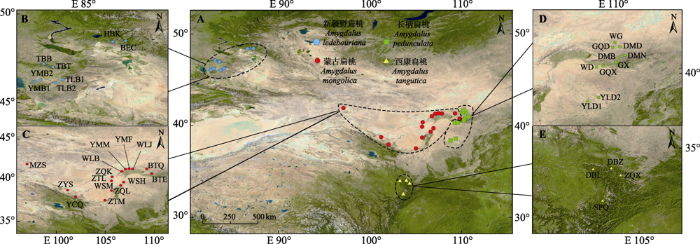
1 材料和方法 1.1 野外调查与采样 本研究共采集扁桃亚属4种野生植物的37个种群, 每个种群采集8-12份样品。采样的扁桃株间至少相距10 m, 均采集新鲜、幼嫩、无病斑的叶片, 用硅胶干燥保存于自封袋, 带回实验室置于-20 ℃冷藏。利用Excel随机抽取公式随机挑选每个种群的4个个体, 共148份样品开展4种扁桃植物的系统发育和物种分化研究(
图1 )。
图1 新窗口打开 |
下载原图ZIP |
生成PPT 图14种扁桃亚属野生植物的野外采样点。BEC, 布尔津县; BTE, 大青山2; BTQ, 大青山1; DBL, 迭部县1; DBZ, 迭部县2; DMB, 达茂旗3; DMD, 达茂旗1; DMN, 达茂旗2; GQD, 固阳1; GQX, 固阳3; GX, 固阳2; HBK, 哈巴河县; MZS, 马鬃山; SPQ, 松潘县; TBB, 塔城2; TBT, 塔城1; TLB1, 托里县1; TLB2, 托里县2; WD, 乌拉山; WG, 乌盖苏木乡; WLB, 大坝口; WLJ, 乌加禾镇; WSH, 苏海图; WSM, 大麦力沟; YCQ, 龙首山; YLD1, 榆林1; YLD2, 榆林2; YMB1, 裕民县1; YMB2, 裕民县2; YMF, 阴山2; YMM, 阴山1; ZQK, 图克木嘎查; ZQL, 贺兰山; ZQX, 舟曲县; ZTL, 特莫乌拉嘎查; ZTM, 苏力图嘎查; ZYS, 祁连山。 Fig. 1Field sampling points of four subgen. Amygdalus wild plants. BEC, Buerjin County; BTE, Group 2 in Daqing Mountain; BTQ, Group 1 in Daqing Mountain; DBL, Group 1 in Têwo County; DBZ, Group 2 in Têwo County; DMB, Group 3 in Damao Banner; DMD, Group 1 in Damao Banner; DMN, Group 2 in Damao Banner; GQD, Group 1 in Guyang County; GQX, Group 3 in Guyang County; GX, Group 2 in Guyang County; HBK, Habahe County; MZS, Mazong Mountain; SPQ, Songpan County; TBB, Group 2 Tacheng City; TBT, Group 1 Tacheng City; TLB1, Group 1 in Toli County; TLB2, Group 2 in Toli County; WD, Wula Mountain; WG, Ugai Sumu Township; WLB, Dam Mouth; WLJ, Wujiahe Town; WSH, Suhaitu; WSM, Damaili Furrow; YCQ, Longshou Mountain; YLD1, Group 1 in Yulin City; YLD2, Group 2 in Yulin City; YMB1, Group 1 in Yumin County; YMB2, Group 2 in Yumin County; YMF, Group 2 in Yinshan Mountains; YMM, Group 1 in Yinshan Mountains; ZQK, Tukemu Gazha; ZQL, Helan Mountain; ZQX, Zhugqu County; ZTL, Temowula Gazha; ZTM, Su Litu Gazha; ZYS, Qilianshan Mountain. 1.2 研究方法 总基因组DNA利用天根公司新型植物基因组提取试剂盒(DP320)进行提取。利用Thermo核酸检测仪(Nanodrop 2000/2000c, 北京博恒科创生物科技有限公司, 北京)测定DNA的浓度。利用核DNA序列通用引物ITS1-ITS4进行目的片段的PCR扩增(Hall,
1999 )。PCR扩增反应体系为25 μL, 包括10 × Easy Pfu Buffer 2 μL, 0.25 mmol·mol
-1 dNTPs 2 μL, 2.5 U Easy Pfu DNA polymerase 0.5 μL, 上下游引物各1 μL, DNA模板1 μL, 灭菌双蒸馏水17.5 μL。反应程序为94 ℃预变性5 min; 94 ℃变性30 s, 退火30 s, 退火温度为55 ℃, 72 ℃延伸1 min, 35个循环; 72 ℃延伸10 min。获得的PCR产物利用1%的琼脂糖进行凝胶电泳, EB染色观察。将扩增产物和测序引物送上海生物工程技术有限公司进行正反向测序, 测序引物同PCR扩增引物。
1.3 数据处理 测序数据利用BioEdit 7.0进行多重比对、人工校对及序列编辑与拼接。对测序后存在的多倍体杂合位点, 利用PHASE 2.1的贝叶斯方法(Stephens & Donnelly,
2003 ), 在DnaSP 6.0中使用默认参数进行矫正, 并利用MEGA-X进行核查(Librado & Rozas,
2009 )。利用DnaSP 6.0对ITS1-ITS4进行序列分析: 统计插入或缺失碱基的个数, 鉴别不同的单倍型序列。序列中的插入或缺失都作为一个进化事件处理(Simmons & Ochoterena,
2000 ), 用于后续的聚类和建树分析。利用R 4.0.2的“vegan”包(Peakall & Smouse,
2006 ), 基于成对遗传距离矩阵, 进行种群水平的主坐标分析(PCoA), 并利用“scatter plot 3d”包对聚类结果进行3D可视化。
选择我国分布的稠李(
Padus racemosa )和山桃(
Amygdalus davidiana )(GenBank登录号分别为MH 711214.1和MH711207.1)作为外类群(邱蓉等,
2012 )。利用软件Network 10.0基于中接法(Median-joining)分析单倍型的网络关系。利用MEGA-X (Tamura
et al .,
2007 ), 基于Kimura-2参数模型计算4种扁桃种间种内的遗传距离, 并利用最大似然法构建单倍型的系统发育树。利用BEAST 1.7.4 (Ronquist
et al .,
2012 ) 构建单倍型的贝叶斯系统发育树, 具体方法参照文献(Ma
et al .,
2019 )。
1.4 生态位相似性分析 基于4种扁桃的自然分布点和环境因子数据, 利用R 3.6.2的“ecospat”包分析4种植物的生态位分化, 分析环境变量对种间生态位分化的相对贡献, 利用统计学检验检测种间生态位分化的显著性。首先, 整合本研究种群采样点和已发表文献采样点(段义忠等,
2020 ), 共获取4种野生扁桃自然分布点239个; 基于“biomod2”包和“raster”包剔除重复和模糊分布点, 最终获得102个分布点。从世界气候网站(Worldclim)获取基准时段(1970-2000年)的19个生物气候变量, 利用“Hmisc”包进行Pearson相关性分析, 剔除相关性较高(>80%)的变量, 最终获得7个环境因子: 年平均气温(Bio1)、温度季节性变化标准差(Bio4)、最暖月最高气温(Bio5)、最冷月最低气温(Bio6)、降水量的季节性变异系数(Bio15)、最干季降水量(Bio17)和最暖季降水量(Bio18)。基于筛选后的分布点和环境因子, 利用“ecospat”包可视化4种扁桃的生态位空间, 并计算生态位重叠分数
D (观测值), 其范围从0到1, 表示生态位从没有叠加到完全叠加(Warren
et al .,
2008 )。利用“factoextra”包的主成分分析(PCA)计算各驱动因子对种间生态位分化的相对贡献。
2 结果和分析 2.1 4种扁桃的遗传结构和物种分化 4种扁桃的核序列ITS1-ITS4多重比对后的总长为634 bp, 包括27个核苷酸变异位点, 共定义了28个单倍型(
图2 )。单倍型网络表明, 28个单倍型聚为4支, 分别对应新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃, 种间无共享单倍型(
图2 )。4种植物的遗传距离分析显示: 种内和种间的遗传分化距离分别为0-0.009和0.014-0.066, 种内和种间的距离间隙为0.005, 表明4个物种种间存在显著的遗传分化(
图3 )。4种扁桃的PCoA显示, 新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃总体上各聚为一组, 其中新疆野扁桃和蒙古扁桃及西康扁桃之间的遗传距离相对较小, 而三者与长柄扁桃的遗传距离相对较大(
图4 )。
图2 新窗口打开 |
下载原图ZIP |
生成PPT 图2扁桃亚属4种野生植物28个单倍型(R1-R28)的地理分布及其单倍型网络。图中种群采样点编码与图1一致, 饼图表示各种群的单倍型频率。A, 单倍型网络, 图中圆圈大小与单倍型频率成正比, 节点间的分支长度大致与单倍型的突变数成正比, 相应分支附近附有步长; 稠李和山桃作为外类群。 Fig. 2Geographical distribution and the haplotype network of 28 haplotypes (R1-R28) of four subgen. Amygdalus wild plants. The population field sampling point codes in the figure are consistent with the ones in Fig. 1. Pie graphs indicate the frequency of each haplotype of these populations. A, In the median-joining haplotypes network, the sizes of the circles in the network are proportional to the haplotype frequencies. Branch lengths are roughly proportional to the number of mutation steps between haplotypes and nodes; the true number of steps is shown near the corresponding branch sections. Padus racemosa and Amygdalus davidiana was used as outgroup. 图3 新窗口打开 |
下载原图ZIP |
生成PPT 图3基于ITS序列计算的扁桃亚属4种野生植物种间和种内的遗传距离。 Fig. 3Genetic distance among four subgen. Amygdalus wild plants based on ITS sequences. 图4 新窗口打开 |
下载原图ZIP |
生成PPT 图4扁桃亚属4种野生植物基于种群水平的前3个坐标的主坐标分析(PCoA)。 Fig. 4Plots of the first three coordinates of the principal coordinates analysis (PCoA) at the population level for four subgen. Amygdalus wild plants. 2.2 4种扁桃单倍型的系统发育分析 基于ITS序列构建的4种扁桃的最大似然树和贝叶斯树均表明28个单倍型聚为两支和4个亚支, 其中, 新疆野扁桃、蒙古扁桃和西康扁桃聚为一支, 长柄扁桃为另一支; 单倍型的4个亚支分别对应新疆野扁桃(R1-R6)、蒙古扁桃(R7-R17)、长柄扁桃(R18-R25)和西康扁桃(R26-R28)的单倍型(
图5 )。
图5 新窗口打开 |
下载原图ZIP |
生成PPT 图5扁桃亚属4种野生植物基于ITS序列的单倍型系统发育树。A, 最大似然(ML)树。分支点上方的数字为大于等于70的自展支持率。B, 贝叶斯树。分支节点右侧的数字表示大于0.70的后验概率值, 两系统发育树右侧的中括号表示为4个物种相应的亚支。 Fig. 5Haplotype phylogenetic trees of four subgen. Amygdalus wild plants based on ITS sequences. A, Maximum likelihood (ML) tree. Bootstrap values equal to or greater than 70 are shown above the corresponding branching points. B, Bayesian tree. The values on the right of the branching points represent the posterior probability greater than 0.70; the brackets on the right of the two phylogenetic trees indicate the corresponding subclades of the four species. 2.3 4种扁桃的生态位相似性及其驱动因子 生态位相似性分析表明, 新疆野扁桃与其他3种扁桃, 蒙古扁桃与长柄扁桃之间生态位分化均不明显,
D 的范围为0.047 4-0.405 4, 统计检验均不显著(
p > 0.05), 可视化的生态位空间均显示存在一定范围的生态位重叠。西康扁桃与蒙古扁桃, 与长柄扁桃均存在显著的生态位分化(
p < 0.05),
D 均为0,可视化生态位均无重叠(
图5 )。基于筛选后的7个环境因子的PCA表明: 前两个PC轴解释了研究区环境因子方差的70.84% (PC1: 39.62%; PC2: 31.22%); 最暖月最高气温(Bio5)、年平均气温(Bio1)、最冷月最低气温(Bio6)和最暖季降水量(Bio18)主要影响了西康扁桃与蒙古扁桃, 与长柄扁桃之间的生态位分化(
图6 )。
图6 新窗口打开 |
下载原图ZIP |
生成PPT 图6扁桃亚属4种野生植物生态位相似性与驱动因子贡献率。A-F, 生态位可视化。绿色表示第一个种的生态位空间, 红色表示第二个种的生态位空间, 蓝色表示重叠空间。D为生态位重叠分数。G, 驱动因子主成分分析。箭头表示相关的方向(同一方向表示高度相关)。Bio1, 年平均气温; Bio4, 温度季节性变化标准差; Bio5, 最暖月最高气温; Bio6, 最冷月最低气温; Bio15, 降水量的季节性变异系数; Bio17, 最干季降水量; Bio18, 最暖季降水量。环境因子后括号里的数字为贡献率排名。 Fig. 6Ecological niche differentiation and the contribution of driving factors of four subgen. Amygdalus plants. A-F, Visualization of ecological niches. The green colour depicts the niche space of the first species, red of the second species and the overlapping range is shown in blue. D is the ecological niche overlap score. G, Driver factors principal component analysis. The arrow depicts the direction of correlation (same direction indicates a high correlation). Bio1, annual mean temperature; Bio4, temperature seasonality (standard deviation × 100); Bio5, max temperature of warmest month; Bio6, min temperature of coldest month; Bio15, precipitation seasonality (coefficient of variation); Bio17, precipitation of driest quarter; Bio18, precipitation of warmest quarter. The numbers in brackets after the environmental factors are the contribution rankings. 3 讨论 3.1 4种扁桃的系统发育关系 目前, 对于李亚科的分属存在两种划分观点, 一类为广义李属(Benthan & Hooker,
1865 ), 即: 桃、李、梅、杏等核果统归于李属, 再下设7个组; 另一类为狭义李属(俞德浚, 1979), 采用小属的划分方法, 共7个属: 桃属、杏属(
Armeniaca )、樱属(
Cerasus )、桂樱属(
Laurocerasus )、稠李属(
Padus )和李属(
Prunus ), 每个属下又分亚属(如桃属分扁桃亚属和桃亚属(subgen.
Persica )), 该方法同时也是《中国植物志》的分类方法。李亚科内所有植物均为单心皮核果, 关于其属下亚属或组, 以及种的划分, 几百年来均无统一结果。本研究基于ITS序列, 最大似然树、贝叶斯树、单倍型网络及PCoA均表明4种扁桃的单倍型呈4个亚支, 种间无共享单倍型(
图2 ,
图4 ,
图5 )。形态上, 4种扁桃的枝、叶、果实和种子等也均存在明显差异(赵一之,
1995 ; 毛祖梅等,
1996 )。
前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 )。本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 )。4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 )。4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 )。基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 )。从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 )。根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 )。
3.2 4种扁桃谱系分化与生态位分化 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关。第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 )。同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 )。4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 )。
4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用。新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木。西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 )。种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 )。西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 )。年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 )。3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 )。另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠。
[1] Bentham G Hooker JD 1865 ). Genera Plantarum. [本文引用: 1] [2] Delplancke M Yazbek M Arrigo N Espíndola A Joly H Alvarez N 2016 ). Combining conservative and variable markers to infer the evolutionary history of Prunus subgen. Amygdalus s.l. under domesticationGenetic Resources & Crop Evolution 221 -234 . [本文引用: 1] [3] Dong SP 2015 ). Phylogeny Research of Prunus triloba and Related Species Based on Chromosome Kayotype and Single-copy Nuclear Gene DNA SequencesMaster degree dissertation, Beijing Forestry University, Beijing [本文引用: 2] [ 董山平 (2015 ). 基于染色体核型和单拷贝核基因DNA序列的榆叶梅及其近缘种系统学研究硕士学位论文, 北京林业大学, 北京 .] [本文引用: 2] [4] Duan YZ Wang JH Wang C Wang HT Du ZY 2020 ). Analysis on the potential suitable areas of four species of subgen. Amygdalus in arid Northwest China under future climate changeChinese Journal of Ecology 2193 -2204 . [本文引用: 1] [ 段义忠 , 王佳豪 , 王驰 , 王海涛 , 杜忠毓 (2020 ). 未来气候变化下西北干旱区4种扁桃亚属植物潜在适生区分析生态学杂志 , 39, 2193 -2204 .] [本文引用: 1] [5] Hall TA 1999 ). BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NTNucleic Acids Symposium Series 95 -98 . [本文引用: 1] [6] Jing ZB Cheng JM Guo CH Wang XP 2013 ). Seed traits, nutrient elements and assessment of genetic diversity for almond (Amygdalus spp.) endangered to China as revealed using SRAP markersBiochemical Systematics and Ecology 51 -57 . DOI:
10.1016/j.bse.2013.03.015 URL [本文引用: 1] [7] Li YX 2001 ). Evolution of plant cell karyotypeBulletin of Biology 2 ), 16 -17 . [本文引用: 1] [ 李雅轩 (2001 ). 植物细胞核型的进化生物学通报 , 36(2 ), 16 -17 .] [本文引用: 1] [8] Librado P Rozas J 2009 ). DnaSP v5: a software for comprehensive analysis of DNA polymorphism dataBioinformatics 1451 -1452 . DOI:
10.1093/bioinformatics/btp187 PMID:
19346325 [本文引用: 1] DnaSP is a software package for a comprehensive analysis of DNA polymorphism data. Version 5 implements a number of new features and analytical methods allowing extensive DNA polymorphism analyses on large datasets. Among other features, the newly implemented methods allow for: (i) analyses on multiple data files; (ii) haplotype phasing; (iii) analyses on insertion/deletion polymorphism data; (iv) visualizing sliding window results integrated with available genome annotations in the UCSC browser.Freely available to academic users from: (http://www.ub.edu/dnasp).
[9] Liu N Huang Y 2010 ). Advances in molecular systematics of polyneopteraEntomotaxonomia 304 -312 . [本文引用: 1] [ 刘念 , 黄原 (2010 ). 多新翅类昆虫分子系统学的研究现状昆虫分类学报 , 32, 304 -312 .] [本文引用: 1] [10] Ma SM Nie YB Jiang XL Xu Z Ji WQ 2019 ). Genetic structure of the endangered, relict shrub Amygdalus mongolica (Rosaceae) in arid northwest ChinaAustralian Journal of Botany 128 -139 . DOI:
10.1071/BT18188 URL [本文引用: 2] [11] Mao ZM Li XY Yang CY 1996 ). Flora of Xinjiang. 355 . [本文引用: 2] [ 毛祖美 , 李学禹 , 杨昌友 (1996 ). 新疆植物志新疆科技卫生出版社, 乌鲁木齐 . 355 .] [本文引用: 2] [12] Mei LX Liu WQ Wei Y Jiang B 2014 ). Evaluation of the resources and development potential of Amygdalus spp. in ChinaJournal of Northwest Forestry University 1 ), 69 -72 . [本文引用: 1] [ 梅立新 , 刘文倩 , 魏钰 , 蒋宝 (2014 ). 中国扁桃资源与利用价值分析西北林学院学报 , 29(1 ), 69 -72 .] [本文引用: 1] [13] Peakall R Smouse PE 2006 ). Genalex 6: genetic analysis in Excel. Population genetic software for teaching and researchMolecular Ecology Notes 288 -295 . DOI:
10.1111/men.2006.6.issue-1 URL [本文引用: 1] [14] Peterson AT 1999 ). Conservatism of ecological niches in evolutionary timeScience 1265 -1267 . PMID:
10455053 Theory predicts low niche differentiation between species over evolutionary time scales, but little empirical evidence is available. Reciprocal geographic predictions based on ecological niche models of sister taxon pairs of birds, mammals, and butterflies in southern Mexico indicate niche conservatism over several million years of independent evolution (between putative sister taxon pairs) but little conservatism at the level of families. Niche conservatism over such time scales indicates that speciation takes place in geographic, not ecological, dimensions and that ecological differences evolve later.
[15] Peterson AT Soberón J Sánchez CV 1999 ). Conservatism of ecological niches in evolutionary timeScience 1265 -1267 . PMID:
10455053 [本文引用: 1] Theory predicts low niche differentiation between species over evolutionary time scales, but little empirical evidence is available. Reciprocal geographic predictions based on ecological niche models of sister taxon pairs of birds, mammals, and butterflies in southern Mexico indicate niche conservatism over several million years of independent evolution (between putative sister taxon pairs) but little conservatism at the level of families. Niche conservatism over such time scales indicates that speciation takes place in geographic, not ecological, dimensions and that ecological differences evolve later.
[16] Qiu R Cheng ZP Wang ZL 2012 ). Studies on genetic relationship and evolutionary path of subgenus Amygdalu s in ChinaActa Horticulturae Sinica 205 -214 . [本文引用: 2] [ 邱蓉 , 程中平 , 王章利 (2012 ). 中国扁桃亚属植物亲缘关系及其演化途径研究园艺学报 , 39, 205 -214 .] [本文引用: 2] [17] Ronquist F Teslenko M van der Mark P Ayres DL Darling A Höhna S Larget B Liu L Suchard MA Huelsenbeck JP 2012 ). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model spaceSystematic Biology 539 -542 . DOI:
10.1093/sysbio/sys029 PMID:
22357727 [本文引用: 1] Since its introduction in 2001, MrBayes has grown in popularity as a software package for Bayesian phylogenetic inference using Markov chain Monte Carlo (MCMC) methods. With this note, we announce the release of version 3.2, a major upgrade to the latest official release presented in 2003. The new version provides convergence diagnostics and allows multiple analyses to be run in parallel with convergence progress monitored on the fly. The introduction of new proposals and automatic optimization of tuning parameters has improved convergence for many problems. The new version also sports significantly faster likelihood calculations through streaming single-instruction-multiple-data extensions (SSE) and support of the BEAGLE library, allowing likelihood calculations to be delegated to graphics processing units (GPUs) on compatible hardware. Speedup factors range from around 2 with SSE code to more than 50 with BEAGLE for codon problems. Checkpointing across all models allows long runs to be completed even when an analysis is prematurely terminated. New models include relaxed clocks, dating, model averaging across time-reversible substitution models, and support for hard, negative, and partial (backbone) tree constraints. Inference of species trees from gene trees is supported by full incorporation of the Bayesian estimation of species trees (BEST) algorithms. Marginal model likelihoods for Bayes factor tests can be estimated accurately across the entire model space using the stepping stone method. The new version provides more output options than previously, including samples of ancestral states, site rates, site d(N)/d(S) rations, branch rates, and node dates. A wide range of statistics on tree parameters can also be output for visualization in FigTree and compatible software.
[18] Shi WT 2018 ). Classification of Prunus L. sensu lato Based on Fruit MorphologyMaster degree dissertation, South China Agricultural University, Guangdong [本文引用: 2] [ 石文婷 (2018 ). 基于果实形态的广义李属植物系统分类学研究硕士学位论文, 华南农业大学, 广东 .] [本文引用: 2] [19] Simmons MP Ochoterena H 2000 ). Gaps as characters in sequence-based phylogenetic analysesSystematic Biology 369 -381 . PMID:
12118412 [本文引用: 1] [20] Slatyer RA Hirst M Sexton JP 2013 ). Niche breadth predicts geographical range size: a general ecological patternEcology Letters 1104 -1114 . DOI:
10.1111/ele.12140 PMID:
23773417 [本文引用: 1] The range of resources that a species uses (i.e. its niche breadth) might determine the geographical area it can occupy, but consensus on whether a niche breadth-range size relationship generally exists among species has been slow to emerge. The validity of this hypothesis is a key question in ecology in that it proposes a mechanism for commonness and rarity, and if true, may help predict species' vulnerability to extinction. We identified 64 studies that measured niche breadth and range size, and we used a meta-analytic approach to test for the presence of a niche breadth-range size relationship. We found a significant positive relationship between range size and environmental tolerance breadth (z = 0.49), habitat breadth (z = 0.45), and diet breadth (z = 0.28). The overall positive effect persisted even when incorporating sampling effects. Despite significant variability in the strength of the relationship among studies, the general positive relationship suggests that specialist species might be disproportionately vulnerable to habitat loss and climate change due to synergistic effects of a narrow niche and small range size. An understanding of the ecological and evolutionary mechanisms that drive and cause deviations from this niche breadth-range size pattern is an important future research goal.© 2013 John Wiley & Sons Ltd/CNRS.
[21] Stephens M Donnelly P 2003 ). A comparison of Bayesian methods for haplotype reconstruction from population genotype dataThe American Journal of Human Genetics 1162 -1169 . DOI:
10.1086/379378 URL [本文引用: 1] [22] Tamura K Dudley J Nei M Kumar S 2007 ). MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0Molecular Biology and Evolution 1596 - 1599 . DOI:
10.1093/molbev/msm092 URL [本文引用: 1] [23] Tian JB He Y Cheng EM 2008 ). Chinese AlmondChina Agriculture Press, Beijing 54 -56 . [本文引用: 3] [ 田建保 , 何勇 , 程恩明 (2008 ). 中国扁桃中国农业出版社, 北京 54 -56 .] [本文引用: 3] [24] Warren DL Glor RE Turelli M 2008 ). Environmental niche equivalency versus conservatism: quantitative approaches to niche evolutionEvolution 2868 -2883 . DOI:
10.1111/j.1558-5646.2008.00482.x PMID:
18752605 [本文引用: 2] Environmental niche models, which are generated by combining species occurrence data with environmental GIS data layers, are increasingly used to answer fundamental questions about niche evolution, speciation, and the accumulation of ecological diversity within clades. The question of whether environmental niches are conserved over evolutionary time scales has attracted considerable attention, but often produced conflicting conclusions. This conflict, however, may result from differences in how niche similarity is measured and the specific null hypothesis being tested. We develop new methods for quantifying niche overlap that rely on a traditional ecological measure and a metric from mathematical statistics. We reexamine a classic study of niche conservatism between sister species in several groups of Mexican animals, and, for the first time, address alternative definitions of "niche conservatism" within a single framework using consistent methods. As expected, we find that environmental niches of sister species are more similar than expected under three distinct null hypotheses, but that they are rarely identical. We demonstrate how our measures can be used in phylogenetic comparative analyses by reexamining niche divergence in an adaptive radiation of Cuban anoles. Our results show that environmental niche overlap is closely tied to geographic overlap, but not to phylogenetic distances, suggesting that niche conservatism has not constrained local communities in this group to consist of closely related species. We suggest various randomization tests that may prove useful in other areas of ecology and evolutionary biology.
[25] Wei Y Guo CH Zhang GQ Meng QC Guo GG 2012 ). Studies on cold-resistance of several Chinese almond speciesJournal of Northwest A&F University Natural Science Edition ), 40(6 ), 99 -106 . [本文引用: 1] [ 魏钰 , 郭春会 , 张国庆 , 孟庆超 , 郭改改 (2012 ). 我国几个扁桃种抗寒性的研究西北农林科技大学学报(自然科学版) , 40(6 ), 99 -106 .] [本文引用: 1] [26] Yang B Meng QY Zhang K Duan YZ 2020 ). Chloroplast genome characterization and identification of genetic relationship of relict endangered plant Amygdalus nana. Bulletin of Botanical Research 686 -695 . [本文引用: 1] [ 杨斌 , 孟庆瑶 , 张凯 , 段义忠 (2020 ). 孑遗濒危植物矮扁桃叶绿体全基因组特征分析及亲缘关系鉴定植物研究 , 40, 686 - 695 .] [本文引用: 1] [27] Yazbek M Oh SH 2013 ). Peaches and almonds: phylogeny of Prunus subg. Amygdalus (Rosaceae) based on DNA sequences and morphologyPlant Systematics and Evolution 1403 -1418 . DOI:
10.1007/s00606-013-0802-1 URL [本文引用: 3] [28] Yu DJ 1986 ). Flora Reipublicae Popularis SinicaeScience Press, Beijing 11 -17 . [本文引用: 2] [ 俞德浚 (1986 ). 中国植物志科学出版社, 北京 11 -17 .] [本文引用: 2] [29] Yu HH Yang ZL Yang X Tang ZF Shu X Liu RN 2010 ). Advances in studies on ITS sequence of medicinal plants germplasm resourcesChinese Traditional & Herbal Drugs 491 -496 . [本文引用: 1] [ 于华会 , 杨志玲 , 杨旭 , 谭梓峰 , 舒枭 , 刘若楠 (2010 ). 药用植物种质资源ITS序列研究进展中草药 , 41, 419 -496 .] [本文引用: 1] [30] Zeng B 2008 ). A Study on Propagate Biological Characteristics and Genetic Diversity of Germplasm Resources of Amygdalus ledebouriana SchlechtPhD dissertation, Xinjiang Agricultural University, Ürümqi [本文引用: 1] [ 曾斌 (2008 ). 新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究博士学位论文, 新疆农业大学, 乌鲁木齐 .] [本文引用: 1] [31] Zhang HX Wang Q Jia SW 2020 ). Genomic phylogeography of Gymnocarpos przewalskii (Caryophyllaceae): insights into habitat fragmentation in arid northwestern ChinaDiversity 335 . DOI:
10.3390/d12090335 URL [本文引用: 1] [32] Zhang YJ Li DZ 2011 ). Advances in phylogenomics based on complete chloroplast genomesPlant Diversity and Resources 365 -375 . [本文引用: 1] [ 张韵洁 , 李德铢 (2011 ). 叶绿体系统发育基因组学的研究进展植物分类与资源学报 , 33, 365 -375 .] [本文引用: 1] [33] Zhao YZ 1995 ). Study on floristic geographical distribution of Amygdalus mongolica Acta Scientiarum Naturalium Universitatis Neimongol 713 -715 . [本文引用: 3] [ 赵一之 (1995 ). 蒙古扁桃的植物区系地理分布研究内蒙古大学学报(自然科学版) , 26, 713 -715 .] [本文引用: 3] 1
1865
... 目前, 对于李亚科的分属存在两种划分观点, 一类为广义李属(Benthan & Hooker,
1865 ), 即: 桃、李、梅、杏等核果统归于李属, 再下设7个组; 另一类为狭义李属(俞德浚, 1979), 采用小属的划分方法, 共7个属: 桃属、杏属(
Armeniaca )、樱属(
Cerasus )、桂樱属(
Laurocerasus )、稠李属(
Padus )和李属(
Prunus ), 每个属下又分亚属(如桃属分扁桃亚属和桃亚属(subgen.
Persica )), 该方法同时也是《中国植物志》的分类方法.李亚科内所有植物均为单心皮核果, 关于其属下亚属或组, 以及种的划分, 几百年来均无统一结果.本研究基于ITS序列, 最大似然树、贝叶斯树、单倍型网络及PCoA均表明4种扁桃的单倍型呈4个亚支, 种间无共享单倍型(
图2 ,
图4 ,
图5 ).形态上, 4种扁桃的枝、叶、果实和种子等也均存在明显差异(赵一之,
1995 ; 毛祖梅等,
1996 ). ...
Combining conservative and variable markers to infer the evolutionary history of
Prunus subgen.
Amygdalus s.l. under domestication
1
2016
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
基于染色体核型和单拷贝核基因DNA序列的榆叶梅及其近缘种系统学研究
2
2015
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
... ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
基于染色体核型和单拷贝核基因DNA序列的榆叶梅及其近缘种系统学研究
2
2015
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
... ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
未来气候变化下西北干旱区4种扁桃亚属植物潜在适生区分析
1
2020
... 基于4种扁桃的自然分布点和环境因子数据, 利用R 3.6.2的“ecospat”包分析4种植物的生态位分化, 分析环境变量对种间生态位分化的相对贡献, 利用统计学检验检测种间生态位分化的显著性.首先, 整合本研究种群采样点和已发表文献采样点(段义忠等,
2020 ), 共获取4种野生扁桃自然分布点239个; 基于“biomod2”包和“raster”包剔除重复和模糊分布点, 最终获得102个分布点.从世界气候网站(Worldclim)获取基准时段(1970-2000年)的19个生物气候变量, 利用“Hmisc”包进行Pearson相关性分析, 剔除相关性较高(>80%)的变量, 最终获得7个环境因子: 年平均气温(Bio1)、温度季节性变化标准差(Bio4)、最暖月最高气温(Bio5)、最冷月最低气温(Bio6)、降水量的季节性变异系数(Bio15)、最干季降水量(Bio17)和最暖季降水量(Bio18).基于筛选后的分布点和环境因子, 利用“ecospat”包可视化4种扁桃的生态位空间, 并计算生态位重叠分数
D (观测值), 其范围从0到1, 表示生态位从没有叠加到完全叠加(Warren
et al .,
2008 ).利用“factoextra”包的主成分分析(PCA)计算各驱动因子对种间生态位分化的相对贡献. ...
未来气候变化下西北干旱区4种扁桃亚属植物潜在适生区分析
1
2020
... 基于4种扁桃的自然分布点和环境因子数据, 利用R 3.6.2的“ecospat”包分析4种植物的生态位分化, 分析环境变量对种间生态位分化的相对贡献, 利用统计学检验检测种间生态位分化的显著性.首先, 整合本研究种群采样点和已发表文献采样点(段义忠等,
2020 ), 共获取4种野生扁桃自然分布点239个; 基于“biomod2”包和“raster”包剔除重复和模糊分布点, 最终获得102个分布点.从世界气候网站(Worldclim)获取基准时段(1970-2000年)的19个生物气候变量, 利用“Hmisc”包进行Pearson相关性分析, 剔除相关性较高(>80%)的变量, 最终获得7个环境因子: 年平均气温(Bio1)、温度季节性变化标准差(Bio4)、最暖月最高气温(Bio5)、最冷月最低气温(Bio6)、降水量的季节性变异系数(Bio15)、最干季降水量(Bio17)和最暖季降水量(Bio18).基于筛选后的分布点和环境因子, 利用“ecospat”包可视化4种扁桃的生态位空间, 并计算生态位重叠分数
D (观测值), 其范围从0到1, 表示生态位从没有叠加到完全叠加(Warren
et al .,
2008 ).利用“factoextra”包的主成分分析(PCA)计算各驱动因子对种间生态位分化的相对贡献. ...
BioEdit:a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT
1
1999
... 总基因组DNA利用天根公司新型植物基因组提取试剂盒(DP320)进行提取.利用Thermo核酸检测仪(Nanodrop 2000/2000c, 北京博恒科创生物科技有限公司, 北京)测定DNA的浓度.利用核DNA序列通用引物ITS1-ITS4进行目的片段的PCR扩增(Hall,
1999 ).PCR扩增反应体系为25 μL, 包括10 × Easy Pfu Buffer 2 μL, 0.25 mmol·mol
-1 dNTPs 2 μL, 2.5 U Easy Pfu DNA polymerase 0.5 μL, 上下游引物各1 μL, DNA模板1 μL, 灭菌双蒸馏水17.5 μL.反应程序为94 ℃预变性5 min; 94 ℃变性30 s, 退火30 s, 退火温度为55 ℃, 72 ℃延伸1 min, 35个循环; 72 ℃延伸10 min.获得的PCR产物利用1%的琼脂糖进行凝胶电泳, EB染色观察.将扩增产物和测序引物送上海生物工程技术有限公司进行正反向测序, 测序引物同PCR扩增引物. ...
Seed traits, nutrient elements and assessment of genetic diversity for almond (
Amygdalus spp.) endangered to China as revealed using SRAP markers
1
2013
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
植物细胞核型的进化
1
2001
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
植物细胞核型的进化
1
2001
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
DnaSP v5: a software for comprehensive analysis of DNA polymorphism data
1
2009
... 测序数据利用BioEdit 7.0进行多重比对、人工校对及序列编辑与拼接.对测序后存在的多倍体杂合位点, 利用PHASE 2.1的贝叶斯方法(Stephens & Donnelly,
2003 ), 在DnaSP 6.0中使用默认参数进行矫正, 并利用MEGA-X进行核查(Librado & Rozas,
2009 ).利用DnaSP 6.0对ITS1-ITS4进行序列分析: 统计插入或缺失碱基的个数, 鉴别不同的单倍型序列.序列中的插入或缺失都作为一个进化事件处理(Simmons & Ochoterena,
2000 ), 用于后续的聚类和建树分析.利用R 4.0.2的“vegan”包(Peakall & Smouse,
2006 ), 基于成对遗传距离矩阵, 进行种群水平的主坐标分析(PCoA), 并利用“scatter plot 3d”包对聚类结果进行3D可视化. ...
多新翅类昆虫分子系统学的研究现状
1
2010
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
多新翅类昆虫分子系统学的研究现状
1
2010
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
Genetic structure of the endangered, relict shrub
Amygdalus mongolica (Rosaceae) in arid northwest China
2
2019
... 选择我国分布的稠李(
Padus racemosa )和山桃(
Amygdalus davidiana )(GenBank登录号分别为MH 711214.1和MH711207.1)作为外类群(邱蓉等,
2012 ).利用软件Network 10.0基于中接法(Median-joining)分析单倍型的网络关系.利用MEGA-X (Tamura
et al .,
2007 ), 基于Kimura-2参数模型计算4种扁桃种间种内的遗传距离, 并利用最大似然法构建单倍型的系统发育树.利用BEAST 1.7.4 (Ronquist
et al .,
2012 ) 构建单倍型的贝叶斯系统发育树, 具体方法参照文献(Ma
et al .,
2019 ). ...
... 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关.第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 ).同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 ).4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 ). ...
新疆植物志
2
1996
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 目前, 对于李亚科的分属存在两种划分观点, 一类为广义李属(Benthan & Hooker,
1865 ), 即: 桃、李、梅、杏等核果统归于李属, 再下设7个组; 另一类为狭义李属(俞德浚, 1979), 采用小属的划分方法, 共7个属: 桃属、杏属(
Armeniaca )、樱属(
Cerasus )、桂樱属(
Laurocerasus )、稠李属(
Padus )和李属(
Prunus ), 每个属下又分亚属(如桃属分扁桃亚属和桃亚属(subgen.
Persica )), 该方法同时也是《中国植物志》的分类方法.李亚科内所有植物均为单心皮核果, 关于其属下亚属或组, 以及种的划分, 几百年来均无统一结果.本研究基于ITS序列, 最大似然树、贝叶斯树、单倍型网络及PCoA均表明4种扁桃的单倍型呈4个亚支, 种间无共享单倍型(
图2 ,
图4 ,
图5 ).形态上, 4种扁桃的枝、叶、果实和种子等也均存在明显差异(赵一之,
1995 ; 毛祖梅等,
1996 ). ...
新疆植物志
2
1996
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 目前, 对于李亚科的分属存在两种划分观点, 一类为广义李属(Benthan & Hooker,
1865 ), 即: 桃、李、梅、杏等核果统归于李属, 再下设7个组; 另一类为狭义李属(俞德浚, 1979), 采用小属的划分方法, 共7个属: 桃属、杏属(
Armeniaca )、樱属(
Cerasus )、桂樱属(
Laurocerasus )、稠李属(
Padus )和李属(
Prunus ), 每个属下又分亚属(如桃属分扁桃亚属和桃亚属(subgen.
Persica )), 该方法同时也是《中国植物志》的分类方法.李亚科内所有植物均为单心皮核果, 关于其属下亚属或组, 以及种的划分, 几百年来均无统一结果.本研究基于ITS序列, 最大似然树、贝叶斯树、单倍型网络及PCoA均表明4种扁桃的单倍型呈4个亚支, 种间无共享单倍型(
图2 ,
图4 ,
图5 ).形态上, 4种扁桃的枝、叶、果实和种子等也均存在明显差异(赵一之,
1995 ; 毛祖梅等,
1996 ). ...
中国扁桃资源与利用价值分析
1
2014
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
中国扁桃资源与利用价值分析
1
2014
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
Genalex 6: genetic analysis in Excel. Population genetic software for teaching and research
1
2006
... 测序数据利用BioEdit 7.0进行多重比对、人工校对及序列编辑与拼接.对测序后存在的多倍体杂合位点, 利用PHASE 2.1的贝叶斯方法(Stephens & Donnelly,
2003 ), 在DnaSP 6.0中使用默认参数进行矫正, 并利用MEGA-X进行核查(Librado & Rozas,
2009 ).利用DnaSP 6.0对ITS1-ITS4进行序列分析: 统计插入或缺失碱基的个数, 鉴别不同的单倍型序列.序列中的插入或缺失都作为一个进化事件处理(Simmons & Ochoterena,
2000 ), 用于后续的聚类和建树分析.利用R 4.0.2的“vegan”包(Peakall & Smouse,
2006 ), 基于成对遗传距离矩阵, 进行种群水平的主坐标分析(PCoA), 并利用“scatter plot 3d”包对聚类结果进行3D可视化. ...
Conservatism of ecological niches in evolutionary time
0
1999
Conservatism of ecological niches in evolutionary time
1
1999
... 核糖体DNA内转录间隔区(ITS)序列为双亲遗传, 具有长度保守性和高度变异性, 能有效地反映种间杂交或基因渐渗, 被称为研究属、种和亚种间遗传分化和系统发育关系的通用工具(于华会等,
2010 ).但要注意ITS序列在使用中易出现多倍体杂合位点问题.生态位相似性分析, 分析并可视化物种间的生态位宽度和生态位关系, 有助于识别驱动物种分化与进化的生态驱动力(Slatyer
et al .,
2013 ).若异域分布的姐妹种在环境空间上完全分离, 那么生态因素很可能在物种形成过程中扮演了主要作用, 而如果姐妹种种间具有相同或相似的环境空间, 生态因素则可能只是地理隔离导致的附带分歧(Peterson
et al .,
1999 ).本研究基于核基因ITS序列, 利用群体遗传、系统发育及生态位相似性分析方法, 研究我国4种野生扁桃的种间系统关系和物种分化.旨在解决以下科学目标: 1)基于ITS序列变异的4种扁桃的单倍型系统发育关系与物种分化; 2) 4种扁桃物种间的生态位分化及其环境驱动力; 3)生态因素对4种扁桃物种间的谱系分化与进化所起的作用. ...
中国扁桃亚属植物亲缘关系及其演化途径研究
2
2012
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
... 选择我国分布的稠李(
Padus racemosa )和山桃(
Amygdalus davidiana )(GenBank登录号分别为MH 711214.1和MH711207.1)作为外类群(邱蓉等,
2012 ).利用软件Network 10.0基于中接法(Median-joining)分析单倍型的网络关系.利用MEGA-X (Tamura
et al .,
2007 ), 基于Kimura-2参数模型计算4种扁桃种间种内的遗传距离, 并利用最大似然法构建单倍型的系统发育树.利用BEAST 1.7.4 (Ronquist
et al .,
2012 ) 构建单倍型的贝叶斯系统发育树, 具体方法参照文献(Ma
et al .,
2019 ). ...
中国扁桃亚属植物亲缘关系及其演化途径研究
2
2012
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
... 选择我国分布的稠李(
Padus racemosa )和山桃(
Amygdalus davidiana )(GenBank登录号分别为MH 711214.1和MH711207.1)作为外类群(邱蓉等,
2012 ).利用软件Network 10.0基于中接法(Median-joining)分析单倍型的网络关系.利用MEGA-X (Tamura
et al .,
2007 ), 基于Kimura-2参数模型计算4种扁桃种间种内的遗传距离, 并利用最大似然法构建单倍型的系统发育树.利用BEAST 1.7.4 (Ronquist
et al .,
2012 ) 构建单倍型的贝叶斯系统发育树, 具体方法参照文献(Ma
et al .,
2019 ). ...
MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space
1
2012
... 选择我国分布的稠李(
Padus racemosa )和山桃(
Amygdalus davidiana )(GenBank登录号分别为MH 711214.1和MH711207.1)作为外类群(邱蓉等,
2012 ).利用软件Network 10.0基于中接法(Median-joining)分析单倍型的网络关系.利用MEGA-X (Tamura
et al .,
2007 ), 基于Kimura-2参数模型计算4种扁桃种间种内的遗传距离, 并利用最大似然法构建单倍型的系统发育树.利用BEAST 1.7.4 (Ronquist
et al .,
2012 ) 构建单倍型的贝叶斯系统发育树, 具体方法参照文献(Ma
et al .,
2019 ). ...
基于果实形态的广义李属植物系统分类学研究
2
2018
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
基于果实形态的广义李属植物系统分类学研究
2
2018
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
Gaps as characters in sequence-based phylogenetic analyses
1
2000
... 测序数据利用BioEdit 7.0进行多重比对、人工校对及序列编辑与拼接.对测序后存在的多倍体杂合位点, 利用PHASE 2.1的贝叶斯方法(Stephens & Donnelly,
2003 ), 在DnaSP 6.0中使用默认参数进行矫正, 并利用MEGA-X进行核查(Librado & Rozas,
2009 ).利用DnaSP 6.0对ITS1-ITS4进行序列分析: 统计插入或缺失碱基的个数, 鉴别不同的单倍型序列.序列中的插入或缺失都作为一个进化事件处理(Simmons & Ochoterena,
2000 ), 用于后续的聚类和建树分析.利用R 4.0.2的“vegan”包(Peakall & Smouse,
2006 ), 基于成对遗传距离矩阵, 进行种群水平的主坐标分析(PCoA), 并利用“scatter plot 3d”包对聚类结果进行3D可视化. ...
Niche breadth predicts geographical range size: a general ecological pattern
1
2013
... 核糖体DNA内转录间隔区(ITS)序列为双亲遗传, 具有长度保守性和高度变异性, 能有效地反映种间杂交或基因渐渗, 被称为研究属、种和亚种间遗传分化和系统发育关系的通用工具(于华会等,
2010 ).但要注意ITS序列在使用中易出现多倍体杂合位点问题.生态位相似性分析, 分析并可视化物种间的生态位宽度和生态位关系, 有助于识别驱动物种分化与进化的生态驱动力(Slatyer
et al .,
2013 ).若异域分布的姐妹种在环境空间上完全分离, 那么生态因素很可能在物种形成过程中扮演了主要作用, 而如果姐妹种种间具有相同或相似的环境空间, 生态因素则可能只是地理隔离导致的附带分歧(Peterson
et al .,
1999 ).本研究基于核基因ITS序列, 利用群体遗传、系统发育及生态位相似性分析方法, 研究我国4种野生扁桃的种间系统关系和物种分化.旨在解决以下科学目标: 1)基于ITS序列变异的4种扁桃的单倍型系统发育关系与物种分化; 2) 4种扁桃物种间的生态位分化及其环境驱动力; 3)生态因素对4种扁桃物种间的谱系分化与进化所起的作用. ...
A comparison of Bayesian methods for haplotype reconstruction from population genotype data
1
2003
... 测序数据利用BioEdit 7.0进行多重比对、人工校对及序列编辑与拼接.对测序后存在的多倍体杂合位点, 利用PHASE 2.1的贝叶斯方法(Stephens & Donnelly,
2003 ), 在DnaSP 6.0中使用默认参数进行矫正, 并利用MEGA-X进行核查(Librado & Rozas,
2009 ).利用DnaSP 6.0对ITS1-ITS4进行序列分析: 统计插入或缺失碱基的个数, 鉴别不同的单倍型序列.序列中的插入或缺失都作为一个进化事件处理(Simmons & Ochoterena,
2000 ), 用于后续的聚类和建树分析.利用R 4.0.2的“vegan”包(Peakall & Smouse,
2006 ), 基于成对遗传距离矩阵, 进行种群水平的主坐标分析(PCoA), 并利用“scatter plot 3d”包对聚类结果进行3D可视化. ...
MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0
1
2007
... 选择我国分布的稠李(
Padus racemosa )和山桃(
Amygdalus davidiana )(GenBank登录号分别为MH 711214.1和MH711207.1)作为外类群(邱蓉等,
2012 ).利用软件Network 10.0基于中接法(Median-joining)分析单倍型的网络关系.利用MEGA-X (Tamura
et al .,
2007 ), 基于Kimura-2参数模型计算4种扁桃种间种内的遗传距离, 并利用最大似然法构建单倍型的系统发育树.利用BEAST 1.7.4 (Ronquist
et al .,
2012 ) 构建单倍型的贝叶斯系统发育树, 具体方法参照文献(Ma
et al .,
2019 ). ...
中国扁桃
3
2008
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关.第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 ).同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 ).4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 ). ...
中国扁桃
3
2008
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关.第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 ).同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 ).4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 ). ...
Environmental niche equivalency versus conservatism: quantitative approaches to niche evolution
2
2008
... 基于4种扁桃的自然分布点和环境因子数据, 利用R 3.6.2的“ecospat”包分析4种植物的生态位分化, 分析环境变量对种间生态位分化的相对贡献, 利用统计学检验检测种间生态位分化的显著性.首先, 整合本研究种群采样点和已发表文献采样点(段义忠等,
2020 ), 共获取4种野生扁桃自然分布点239个; 基于“biomod2”包和“raster”包剔除重复和模糊分布点, 最终获得102个分布点.从世界气候网站(Worldclim)获取基准时段(1970-2000年)的19个生物气候变量, 利用“Hmisc”包进行Pearson相关性分析, 剔除相关性较高(>80%)的变量, 最终获得7个环境因子: 年平均气温(Bio1)、温度季节性变化标准差(Bio4)、最暖月最高气温(Bio5)、最冷月最低气温(Bio6)、降水量的季节性变异系数(Bio15)、最干季降水量(Bio17)和最暖季降水量(Bio18).基于筛选后的分布点和环境因子, 利用“ecospat”包可视化4种扁桃的生态位空间, 并计算生态位重叠分数
D (观测值), 其范围从0到1, 表示生态位从没有叠加到完全叠加(Warren
et al .,
2008 ).利用“factoextra”包的主成分分析(PCA)计算各驱动因子对种间生态位分化的相对贡献. ...
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
我国几个扁桃种抗寒性的研究
1
2012
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
我国几个扁桃种抗寒性的研究
1
2012
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
孑遗濒危植物矮扁桃叶绿体全基因组特征分析及亲缘关系鉴定
1
2020
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
孑遗濒危植物矮扁桃叶绿体全基因组特征分析及亲缘关系鉴定
1
2020
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
Peaches and almonds: phylogeny of
Prunus subg.
Amygdalus (Rosaceae) based on DNA sequences and morphology
3
2013
... 对扁桃亚属植物亲缘关系的研究主要体现在: 基于核标记、叶绿体全基因组结合植物学性状开展的系统发育分析, 核标记分析表明蒙古扁桃与西康扁桃, 长柄扁桃与榆叶梅, 新疆野扁桃与扁桃, 具有相对较近的亲缘关系(邱蓉等,
2012 ; Jing
et al .,
2013 ; Yazbek & Oh,
2013 ); 叶绿体全基因组分析表明, 蒙古扁桃和新疆野扁桃, 西康扁桃、长柄扁桃和榆叶梅亲缘关系相对较近(石文婷,
2018 ; 杨斌等,
2020 ).上述研究的研究材料缺乏采样范围的明确设置, 主要采用的是当地标本库的材料或美国国家生物技术信息中心(NCBI)的测序序列建树.我国不同地区分布的4种野生扁桃的种间系统发育关系, 物种分化及其环境驱动力等问题尚不清楚, 成为深入理解4种野生扁桃分布与进化的瓶颈. ...
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
... ; Yazbek & Oh,
2013 ). ...
中国植物志
2
1986
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
中国植物志
2
1986
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 前人利用叶绿体序列和单拷贝核基因序列建树得出, 新疆野扁桃、长柄扁桃与欧洲李、黑刺李(
Prunus spinosa )和稠李(
Padus avium )等李属、稠李属植物, 蒙古扁桃和西康扁桃与桃(
Prunus persica )、紫叶桃(
Prunus persica ‘Zi Ye Tao')、山桃等桃属植物分别具有较近的亲缘关系(Yazbek & Oh, 2010,
2013 ; 董山平,
2015 ).本研究表明4种野生扁桃与两个外类群山桃和稠李间的亲缘及聚类关系均较远(
图1 ,
图3 ,
图4 ).4种扁桃果实成熟后开裂无汁液与山桃和稠李果实成熟后不开裂多汁也具有明显的形态差异(俞德浚,
1986 ).4种扁桃的PCoA和系统发育分析均表明, 新疆野扁桃、蒙古扁桃、西康扁桃之间亲缘关系相对较近, 而与长柄扁桃的亲缘关系均相对较远(
图4 ,
图5 ).基于核标记和形态特征对扁桃亚属系统发育的研究也揭示了长柄扁桃和其他3种扁桃的亲缘关系相对较远(Delplancke
et al .,
2016 ).从染色体核型上, 新疆野扁桃、蒙古扁桃、西康扁桃均为二倍体16条染色体, 长柄扁桃为十二倍体96条染色体(董山平,
2015 ).根据植物由二倍体向多倍体演化的规律, 分布区为4种植物分布最东界的长柄扁桃很有可能为4种扁桃中最年轻的物种(李雅轩,
2001 ; Yazbek & Oh,
2013 ). ...
药用植物种质资源ITS序列研究进展
1
2010
... 核糖体DNA内转录间隔区(ITS)序列为双亲遗传, 具有长度保守性和高度变异性, 能有效地反映种间杂交或基因渐渗, 被称为研究属、种和亚种间遗传分化和系统发育关系的通用工具(于华会等,
2010 ).但要注意ITS序列在使用中易出现多倍体杂合位点问题.生态位相似性分析, 分析并可视化物种间的生态位宽度和生态位关系, 有助于识别驱动物种分化与进化的生态驱动力(Slatyer
et al .,
2013 ).若异域分布的姐妹种在环境空间上完全分离, 那么生态因素很可能在物种形成过程中扮演了主要作用, 而如果姐妹种种间具有相同或相似的环境空间, 生态因素则可能只是地理隔离导致的附带分歧(Peterson
et al .,
1999 ).本研究基于核基因ITS序列, 利用群体遗传、系统发育及生态位相似性分析方法, 研究我国4种野生扁桃的种间系统关系和物种分化.旨在解决以下科学目标: 1)基于ITS序列变异的4种扁桃的单倍型系统发育关系与物种分化; 2) 4种扁桃物种间的生态位分化及其环境驱动力; 3)生态因素对4种扁桃物种间的谱系分化与进化所起的作用. ...
药用植物种质资源ITS序列研究进展
1
2010
... 核糖体DNA内转录间隔区(ITS)序列为双亲遗传, 具有长度保守性和高度变异性, 能有效地反映种间杂交或基因渐渗, 被称为研究属、种和亚种间遗传分化和系统发育关系的通用工具(于华会等,
2010 ).但要注意ITS序列在使用中易出现多倍体杂合位点问题.生态位相似性分析, 分析并可视化物种间的生态位宽度和生态位关系, 有助于识别驱动物种分化与进化的生态驱动力(Slatyer
et al .,
2013 ).若异域分布的姐妹种在环境空间上完全分离, 那么生态因素很可能在物种形成过程中扮演了主要作用, 而如果姐妹种种间具有相同或相似的环境空间, 生态因素则可能只是地理隔离导致的附带分歧(Peterson
et al .,
1999 ).本研究基于核基因ITS序列, 利用群体遗传、系统发育及生态位相似性分析方法, 研究我国4种野生扁桃的种间系统关系和物种分化.旨在解决以下科学目标: 1)基于ITS序列变异的4种扁桃的单倍型系统发育关系与物种分化; 2) 4种扁桃物种间的生态位分化及其环境驱动力; 3)生态因素对4种扁桃物种间的谱系分化与进化所起的作用. ...
新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究
1
2008
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
新疆野扁桃繁殖生物学特性及种质资源遗传多样性研究
1
2008
... 4种扁桃种间均存在一定的生态位分化(
图6 ), 表明生态因素在物种分化的过程中起到了一定的作用.新疆野扁桃、蒙古扁桃、长柄扁桃和西康扁桃分布区的气候差异明显, 分别为中温带大陆性、温带大陆性、寒温带大陆性和温带干旱半干旱气候区, 在年平均气温、年降水量、无霜期、日照时间及土壤类型等方面均存在一定差异(梅立新等,
2014 ), 分布区生态地理环境的显著差异利于4种扁桃物种的分化, 4种植物也被细分为: 小灌木、落叶旱生灌木、落叶小灌木和密生小灌木.西康扁桃与蒙古扁桃, 以及与长柄扁桃之间生态位分化较显著(
图6 ).种间生态位的分化通常由于两物种实际生态位和背景环境的分化(Warren
et al .,
2008 ).西康扁桃是我国唯一分布于南方的扁桃种, 其生长与分布生境均与蒙古扁桃和长柄扁桃存在明显差异(石文婷,
2018 ).年平均气温, 以及反映极端温度与降水的因子主要驱动了西康扁桃与蒙古扁桃、长柄扁桃的生态位分化(
图5 ).3种扁桃分布区的昼夜温差及季节温度变幅均较大, 西康扁桃分布区的年平均气温范围和降水量范围与蒙古扁桃(5-9 ℃; 小于170 mm)和长柄扁桃(7-11 ℃; 330-450 mm)相比, 均相对较大(7-12 ℃; 500-800 mm; 魏钰等,
2012 ).另外, 新疆野扁桃分布区内存在2个生态型: 气候较干旱的阿勒泰生态型和气候温和湿润的塔城生态型(曾斌,
2008 ), 这可能分别与蒙古扁桃和长柄扁桃偏旱生的生态位, 以及西康扁桃偏中生的生态位存在一定重叠. ...
Genomic phylogeography of
Gymnocarpos przewalskii (Caryophyllaceae): insights into habitat fragmentation in arid northwestern China
1
2020
... 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关.第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 ).同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 ).4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 ). ...
叶绿体系统发育基因组学的研究进展
1
2011
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
叶绿体系统发育基因组学的研究进展
1
2011
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
蒙古扁桃的植物区系地理分布研究
3
1995
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 目前, 对于李亚科的分属存在两种划分观点, 一类为广义李属(Benthan & Hooker,
1865 ), 即: 桃、李、梅、杏等核果统归于李属, 再下设7个组; 另一类为狭义李属(俞德浚, 1979), 采用小属的划分方法, 共7个属: 桃属、杏属(
Armeniaca )、樱属(
Cerasus )、桂樱属(
Laurocerasus )、稠李属(
Padus )和李属(
Prunus ), 每个属下又分亚属(如桃属分扁桃亚属和桃亚属(subgen.
Persica )), 该方法同时也是《中国植物志》的分类方法.李亚科内所有植物均为单心皮核果, 关于其属下亚属或组, 以及种的划分, 几百年来均无统一结果.本研究基于ITS序列, 最大似然树、贝叶斯树、单倍型网络及PCoA均表明4种扁桃的单倍型呈4个亚支, 种间无共享单倍型(
图2 ,
图4 ,
图5 ).形态上, 4种扁桃的枝、叶、果实和种子等也均存在明显差异(赵一之,
1995 ; 毛祖梅等,
1996 ). ...
... 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关.第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 ).同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 ).4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 ). ...
蒙古扁桃的植物区系地理分布研究
3
1995
... 系统发育研究根据生物表型特征与分子序列, 以树状结构反映生物类群间的演化关系, 研究生物类群的系统发育利于深入理解生物的起源、亲缘关系及其演化(刘念和黄原,
2010 ).李亚科植物是重要的核果类果树资源, 由于该亚科所有植物的形态特征高度趋同或平行进化, 同源多倍体及种间杂交较常见, 导致该亚科植物属、亚属甚至种间的系统关系至今仍存在较大争议(张韵洁和李德铢,
2011 ).扁桃亚属(subgen.
Amygdalus )隶属于蔷薇科李亚科桃属(
Amygdalus ), 共有6个种: 扁桃(
Amygdalus communis )、新疆野扁桃(
A. ledebouriana 和或矮扁桃
A. nana )、蒙古扁桃(
A. mongolica )、长柄扁桃(
A. pedunculata )、西康扁桃(
A. tangutica )和榆叶梅(
A. triloba )(俞德浚,
1986 ; 毛祖梅等,
1996 ; 田建保等,
2008 ).在我国, 新疆野扁桃与蒙古扁桃, 长柄扁桃和西康扁桃呈间断分布, 后3种扁桃随生态环境条件由北向南呈替代性地理分布(赵一之,
1995 ).具体说, 新疆野扁桃主要残存分布于新疆北部塔城、阿尔泰山干旱坡地、草原等; 蒙古扁桃集中分布于甘肃最北部马鬃山、河西走廊、内蒙古阿拉善左旗及其北部和中部地区的荒漠草原、丘陵坡麓、石质山坡等; 长柄扁桃分布于内蒙古北部和中部及陕西北部的浅山带、荒漠及荒漠草原等; 西康扁桃分布于甘肃南部和四川西北部的山坡向阳处或溪流边(田建保等,
2008 ).4种野生扁桃均是重要的药用经济树种, 具有速生、抗旱、耐寒、适应性强等生物学特性, 也具有重要的生态及科研价值. ...
... 目前, 对于李亚科的分属存在两种划分观点, 一类为广义李属(Benthan & Hooker,
1865 ), 即: 桃、李、梅、杏等核果统归于李属, 再下设7个组; 另一类为狭义李属(俞德浚, 1979), 采用小属的划分方法, 共7个属: 桃属、杏属(
Armeniaca )、樱属(
Cerasus )、桂樱属(
Laurocerasus )、稠李属(
Padus )和李属(
Prunus ), 每个属下又分亚属(如桃属分扁桃亚属和桃亚属(subgen.
Persica )), 该方法同时也是《中国植物志》的分类方法.李亚科内所有植物均为单心皮核果, 关于其属下亚属或组, 以及种的划分, 几百年来均无统一结果.本研究基于ITS序列, 最大似然树、贝叶斯树、单倍型网络及PCoA均表明4种扁桃的单倍型呈4个亚支, 种间无共享单倍型(
图2 ,
图4 ,
图5 ).形态上, 4种扁桃的枝、叶、果实和种子等也均存在明显差异(赵一之,
1995 ; 毛祖梅等,
1996 ). ...
... 4种植物种间存在显著的遗传分化(
图3 ), 这可能与分布区的地理隔离和环境异质性有关.第三纪以来, 4种扁桃分布区内发生了显著的地质变化, 如天山、马鬃山、阴山、秦岭等山体的若干次隆升, 古尔班通古特沙漠、巴丹吉林沙漠、腾格里沙漠及乌兰布和沙漠等的形成与扩张, 很可能造成了4种植物不同群体间的地理隔离与物种分化(Ma
et al .,
2019 ).同时, 环境异质性可引起不同群体所处的微环境显著不同, 进而导致局部遗传变异的差异和局部环境适应性分化(Zhang
et al .,
2020 ).4种扁桃在长期进化过程中, 由于分布区水热条件的差异, 逐渐适应形成了旱生、中旱生和旱中生的特性, 而且相应地在枝、叶、花和果等形态上也形成了明显不同的形态特征(赵一之,
1995 ; 田建保等,
2008 ). ...
 ,
, ,3,*, 黄刚1, 张丹1, 闫涵3
,3,*, 黄刚1, 张丹1, 闫涵3

 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT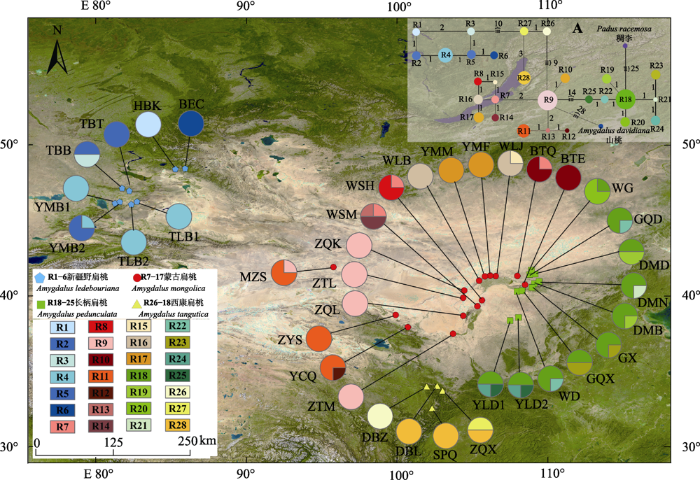 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT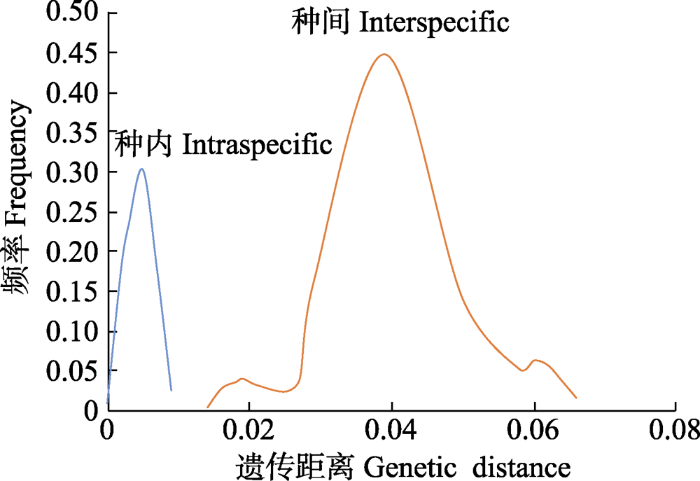 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT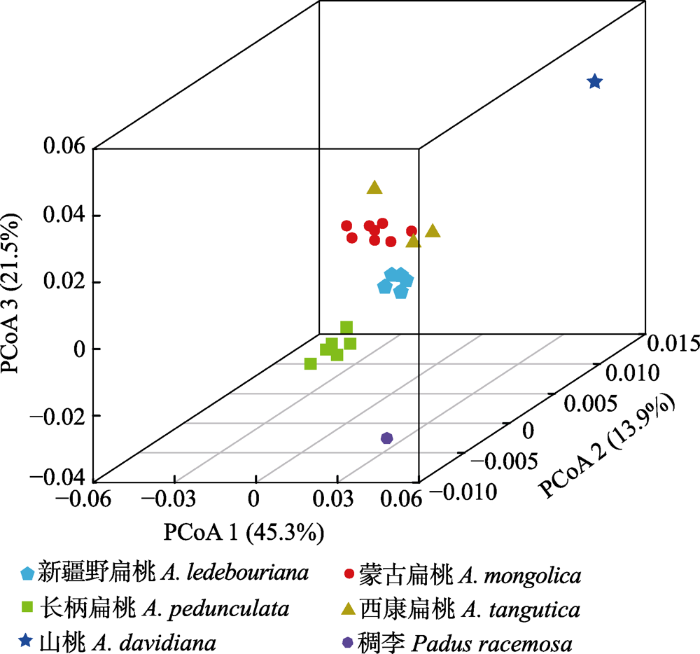 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT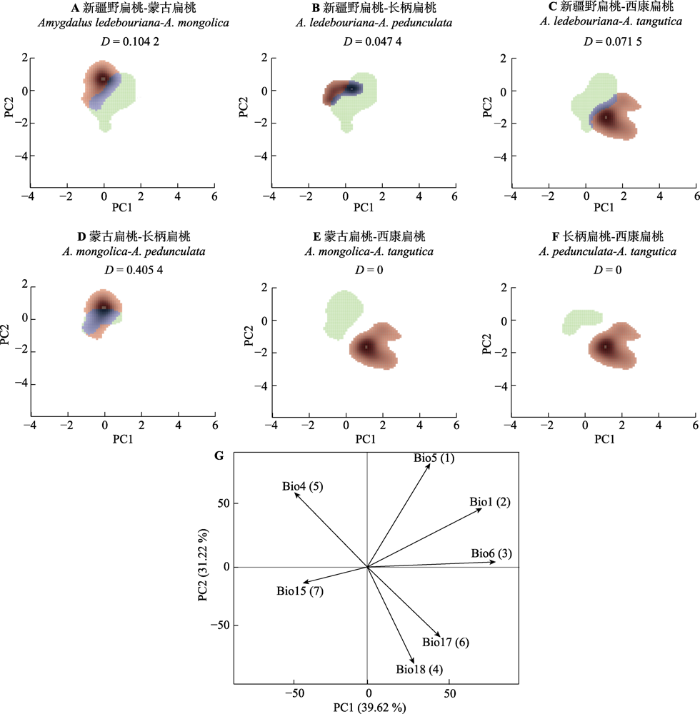 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}