 ,
, ,*
,*Responses of leaf hydraulic traits to water conditions in eight tree species and the driving factors
Jin-Pei REN, Jun-Peng LI, Wei-Feng WANG, Yong-Xin DAI, Lin WANG,,*通讯作者: *王林: ORCID:0000-0002-1658-4720(lwanger@163.com)
编委: 罗天祥
责任编辑: 李敏
收稿日期:2021-04-14接受日期:2021-08-9
| 基金资助: |
Corresponding authors: *(lwanger@163.com)
Received:2021-04-14Accepted:2021-08-9
| Fund supported: |

摘要
树木叶片的水力效率和安全性会对水分条件的改变做出一定的响应, 进而影响树木的生长和分布, 然而叶导水率(Kleaf)和叶水力脆弱性(P50)对不同水分条件的响应模式及其影响因素尚不清楚。该研究选取了晋西北关帝山和黑茶山两种水分条件下的8种树种, 测量其水力性状、叶片导管和形态性状, 比较两地不同树种的Kleaf和P50的变化, 分析叶片水力效率和安全性之间的权衡关系, 并探讨叶片水力性状在不同树种及水分条件下的响应模式及其驱动因素。结果表明: 对同一树种而言, 湿润的关帝山叶最大导水率(Kmax)和P50均高于干旱的黑茶山; 对同一地区而言, 从在高水分条件下生长的树种到在易干旱环境生长的树种, Kmax和P50均逐渐下降。Kmax、P50、膨压丧失点水势(TLP)之间均存在显著相关关系。两地叶片P50与导管密度、导管塌陷预测值((t/b)3)、叶片厚度、比叶质量显著正相关, 与导管直径、叶面积显著负相关, 不同树种的Kleaf和P50与叶导管性状的关系大于叶形态性状。同一树种的关帝山到黑茶山P50变化量(δP50)与比叶质量和叶干物质含量在两地的变化量显著正相关, 同一树种δP50与叶形态性状变化量的关系大于与叶导管性状的。以上结果表明: 随着水分条件变差, 叶片水力效率降低, 水力安全性提高, 不同树种叶片水力效率与安全性之间存在一定的权衡关系, 不同树种叶水力性状的差别受叶导管性状影响的程度大于受叶形态性状的影响, 同一树种叶水力安全性对水分条件变化的响应主要依靠叶形态性状的驱动, 树木在提高自身叶水力安全的同时增加了叶构建的碳投资。
关键词:
Abstract
Aims The hydraulic efficiency and safety of tree leaves can respond to changes in water conditions, hence affecting the growth and distribution of trees. This study was conducted to determine the patterns of responses in leaf hydraulic conductivity (Kleaf) and leaf hydraulic vulnerability (P50) in trees to varying water conditions and the influencing factors. Methods In this study, eight tree species were selected at the study sites of Guandi Mountain and Heicha Mountain in northwestern Shanxi, and their hydraulic traits, leaf vessel and morphological traits were measured. Changes of Kleaf and P50 in those eight tree species were compared between the two locations. The trade-off relationship between leaf hydraulic efficiency and safety was analyzed. Important findings Within the same tree species, the maximum hydraulic conductivity (Kmax) and P50 were higher at the moist Guandi Mountain sites than at the dry Heicha Mountain sites; within the same study areas, Kmax and P50 were higher in tree species occurring under high water availability than those in drought-prone environment. There were significant correlations among Kmax, P50 and water potential at turgor loss points (TLP). Leaf P50 in trees in the two study areas was significantly and positively correlated with the number of vessels per unit area, the predicted value of vessel collapse ((t/b)3), leaf thickness, and leaf mass per unit area, and negatively with vessel diameter and leaf area. Kleaf and P50 in different tree species were better related with leaf vessel traits than with leaf morphological traits. The changes in P50(δP50) from Guandi Mountain to Heicha Mountain within the same tree species were significantly and positively correlated with changes in leaf mass per unit area and leaf dry mass content, and δP50 was more closely related with the leaf morphological traits than with the leaf vessel traits within the same tree species. The above results indicate that, with deterioration of water conditions, leaf hydraulic efficiency decreases while the hydraulic safety increases. There is a certain trade-off between leaf hydraulic efficiency and safety across different tree species. The differences in leaf hydraulic traits among tree species are more affected by leaf vessel traits than leaf morphological traits. The responses of leaf hydraulic safety to changes in water conditions are mainly driven by leaf morphological traits. An improvement in leaf hydraulic safety occurs at the expenses of structural carbon investment.
Keywords:
PDF (1305KB)元数据多维度评价相关文章导出EndNote|Ris|Bibtex收藏本文
引用本文
任金培, 李俊鹏, 王卫锋, 代永欣, 王林. 八个树种叶水力性状对水分条件的响应及其驱动因素. 植物生态学报, 2021, 45(9): 942-951. DOI: 10.17521/cjpe.2021.0140
REN Jin-Pei, LI Jun-Peng, WANG Wei-Feng, DAI Yong-Xin, WANG Lin.

水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al., 2016; McDowell et al., 2019)。树木水分运输系统主要由根、茎、叶3个部分组成(Tyree & Ewers, 1991), 叶片处于整株水分传输系统的终端部分, 尽管叶片水分运输路径占整个植物体的比例较小, 但叶片水分运输阻力占整株的30%-60% (甚至80%), 叶片被认为是在水分限制条件下整株植物水力系统的瓶颈(Yan et al., 2020), 叶片水力学结构与树木的光合速率密切相关, 进而影响树木的生长速率和森林生产力。叶片水力结构的“熔断机制”是树木在干旱条件下减少水分散失的有效手段(Villagra et al., 2013; McCulloh et al., 2014)。
叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio, 2014), 叶导水率高的树种倾向于在资源充足条件下以更强的水分供应能力保证较快速度的碳同化, 进而促进叶片的光合作用和植物生长(Sack & Holbrook, 2006), 叶水力脆弱性对于逆境条件下叶水力结构的维持至关重要, 能在低水势条件下维持水力结构的稳定(Nardini & Luglio, 2014)。干旱对根和茎木质部导管水分输导能力的主要影响是发生气穴栓塞, 限制水分输导能力(Johnson et al., 2009), 也有报道表明干旱还容易导致叶木质部导管壁的破裂或导管向内塌陷, 从而影响其水分输导(Brodribb & Holbrook, 2005)。叶片的水分输导除受到木质部导管水力结构的影响外, 还受到导管外的细胞间水分输导影响。因此叶片的水分输导效率和安全性影响因素较茎更为复杂。前人的研究表明: 植物的叶Kleaf和P50的变化与木质部导管直径、导管密度、导管壁特性等有关(Nardini & Salleo, 2005), 也与叶面积、比叶质量、叶片厚度、叶脉密度等因素有关。对叶片水力性状和经济性状的研究结果也普遍表明叶片水分输导安全性增加的代价是增加了碳投入(金鹰和王传宽, 2016), 并牺牲了部分在充足资源环境下的竞争能力(Nardini et al., 2012), 但目前对叶片应对干旱叶导管和导管外的响应了解不足, 这进一步限制了对不同树种叶水力特性差别及同一树种在不同水分条件下叶水力特性的响应模式及其驱动因素的了解。
前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al., 2016), 但对叶的导水率和栓塞脆弱性之间的相关关系没有一致的结论, 一些研究表明在叶水平上不存在水力效率和水力安全的权衡(Scoffoni et al., 2012; Yao et al., 2021)。但也有研究表明P50与基于叶质量的导水率(Kmass)存在相关关系(Bucci et al., 2019; Yan et al., 2020), 但与基于叶面积的导水率(Karea)关系不大(Nardini & Luglio, 2014)。原因可能是叶片的Kleaf和P50同时受到木质部导管和木质部导管外传输的影响, 从而模糊了这种关系(Yan et al., 2020)。因此, 叶的水分输导效率和安全性之间的关系需要有更多的试验证据。
晋西北森林草原过渡区降水偏少、生态脆弱, 因此这一地区易受到干旱胁迫和气候变化的影响, 研究这一地区树木叶水力结构及其影响因素有利于全面了解温带水分限制区域树木的水力结构及其对水分条件的响应模式, 同时在实践中可以为这一地区造林树种的选择和针对性抚育管理措施的制定提供理论依据。本研究选取晋西北水分条件有一定差别的关帝山和黑茶山8个树种作为研究对象(受地形影响, 关帝山降水较为充足, 黑茶山降水较少, 干旱现象较为普遍), 分析其叶水力性状、叶导管和形态性状, 比较不同水分条件及树种之间叶水力性状上的差别, 并探究叶导管和形态性状与叶水力性状的关系, 以期了解: (1)在晋西北森林草原过渡区的树种叶水分输导效率和安全性之间是否存在权衡关系; (2)降水量不同的两个地区(关帝山和黑茶山)及同一地区不同树种的水分输导效率和安全性的响应模式; (3)不同树种及不同水分条件下的同一树种叶水力性状变化的驱动因素。
1 材料和方法
1.1 研究地概况和试验设计
研究地分别选择在关帝山(G)和黑茶山(H)。关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等, 1999; 刘敏等, 2020)。黑茶山地处吕梁山脉中部北段, 地理位置37.88°-38.73° N, 111.00°-111.77° E, 属温带大陆性气候, 四季分明, 平均海拔1 702 m, 年平均气温3-5 ℃, 年降水量450 mm, 无霜期105-135天, 土壤主要为棕壤和褐土, 主要树种有油松、山杨、山杏(Armeniaca sibirica)、落叶松、云杉等(郜慧萍, 2019)。本研究分别于关帝山和黑茶山中选择8种自然更新的树种, 其中, 阔叶树种为: 青杨(Populus cathayana)、蒙古栎、榆树(Ulmus pumila)、山杏、山杨, 针叶树种为: 落叶松、油松、云杉。青杨和蒙古栎多生长在湿润的山谷地带, 而榆树、山杏和3种针叶树种多生长在干旱的山坡上, 山杨则常生长在山脊上。分别在关帝山和黑茶山选取海拔(1 800 m)、坡度(25°)、地形相似的两个地点作为取样点, 于2020年6月中旬树木旺盛生长期在每个树种最佳生境中随机选取6株生长良好的健康成熟植株作为样木, 采取树冠上部光照条件好、无病虫害的成熟叶片作为研究样品。
1.2 叶水力性状的测定
叶水力脆弱性曲线采用复水法测定(Brodribb & Holbrook, 2003)。清晨在每株样木上随机采集若干带叶枝条, 用黑色塑料袋套住并迅速带回实验室。然后, 若干个小枝条在自然状态下失水不同时间以形成不同的叶水势梯度, 达到不同水势的小枝条分别进行黑暗处理至少0.5 h (使枝条上的所有叶片水势相同)。每个枝条上选取叶片用压力室测定初始叶水势(Ψ0, MPa), 然后在水下切断其相邻叶, 并使之复水一定时间(t, 30-300 s), 复水完成后立即取出擦干, 测定水化后叶水势(Ψf, MPa)。由于复水过程类似于单个电容充电的过程, 因此Kleaf的计算如下:式中, Cleaf为基于叶质量的叶水容(mmol·g-1·MPa-1)。Cleaf用叶质量标准化, 可得到基于叶质量的叶导水率(Kmass)。
叶水力脆弱性曲线是Kleaf与Ψ0之间关系的曲线, 可用一个三参数的Sigmoid函数进行拟合。当叶水势为0时, 即可求得叶最大导水率Kmax。当叶导水率为Kmax的50%时所对应的叶水势即为P50, 代表叶水力脆弱性。
Cleaf通过压力-容积(PV)曲线测定。利用自然风干法制作PV曲线(Tyree & Hammel, 1972), 求得膨压丧失点水势(TLP, MPa)。以相对含水量(RWC, %)和叶水势(Ψl, MPa)作图, 求得膨压丧失点之前线性部分的斜率ΔRWC/ΔΨ1, 用以计算膨压丧失点之前的Cleaf, 计算公式如下:
式中, DW为叶干质量(g); WW为饱和时叶的含水量(g); M为水的摩尔质量(g·mol-1)。
1.3 叶导管和形态性状的测定
清晨在不同样木上随机采集健康的成熟叶片用FAA固定液固定, 带回实验室进行石蜡制片。参照贺冰等(2014)的植物材料制片方法, 取固定好的叶片, 用梯度酒精脱水、透明、浸蜡、包埋、切片、脱蜡、染色、封片。用光学显微镜观测, 每个叶片选取6个视野拍照, 记录叶片的横切面特征, 并用ImageJ软件测量叶片厚度(LT)、主脉的导管密度(N)和主脉导管直径(D), 主脉导管塌陷预测值(t/b)3是导管壁厚度与导管直径比值的3次方。黎明前采集10个健康叶并使其复水至饱和, 首先用1/10 000天平称量叶饱和质量, 然后用扫描仪扫描叶面积(LA), 最后在85 ℃下烘干测定叶干质量。比叶质量(LMA)是叶干质量与叶面积的比值, 叶干物质含量(LDMC)是叶干质量与叶饱和质量的比值。1.4 数据分析
采用Microsoft Office Excel 2019对数据进行预处理。不同树种之间TLP的比较采用单因素方差分析(ANOVA), 并用最小显著差异法(LSD)进行多重比较, 以p < 0.05为显著性评价标准。采用线性回归分析建立水力性状之间的关系。叶片导管性状、形态性状和水力性状之间的关系采用Pearson相关分析, p < 0.05表示显著相关, p < 0.01表示极显著相关。导管性状和形态性状对P50和Kmax的影响程度比较先采用主成分分析, 再进行多元线性回归分析。所有分析均在SPSS 25.0中完成, 用SigmaPlot 14.0进行绘图。2 结果
2.1 叶水力性状
两种立地条件下各树种的叶水力脆弱性曲线均可用“S”形曲线模拟。对同一树种而言, 湿润的关帝山的Kmax均高于干旱的黑茶山, 关帝山的青杨、蒙古栎、榆树、山杏、山杨、落叶松、油松、云杉的Kmax分别比黑茶山的相应树种高63.1%、29.2%、80.7%、59.8%、49%、31.4%、18.6%和17.4%。对同一地区而言, 从水分条件较好的山谷到易受干旱影响的山脊, Kmax逐渐下降, 关帝山的8个树种的变化范围为326.53 (青杨) - 51.56 (云杉) mmol·kg-1·MPa-1·s-1, 其中, 青杨的Kmax是山杨的3倍, 是针叶树种云杉的6.3倍。黑茶山的8个树种变化范围分别为200.22 (青杨) - 43.93 (云杉) mmol·kg-1·MPa-1·s-1, 其中, 青杨的Kmax是山杨的2.8倍, 是针叶树种云杉的4.6倍(图1)。图1
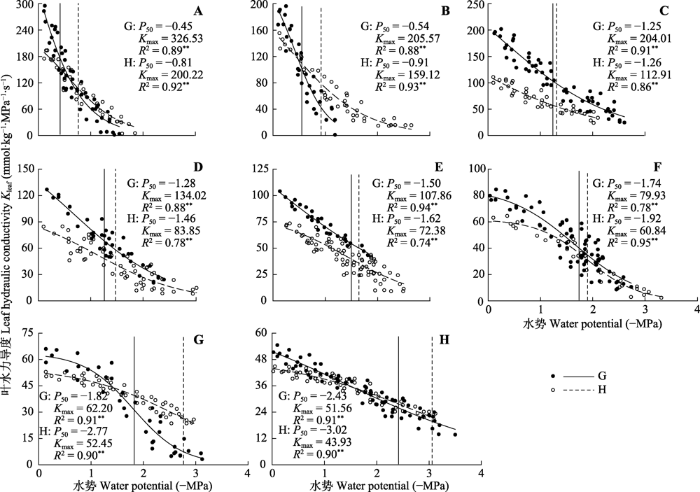 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图1关帝山和黑茶山8个树种的叶水力脆弱性曲线。A, 青杨。B, 蒙古栎。C, 榆树。D, 山杏。E, 山杨。F, 落叶松。G, 油松。H, 云杉。图中竖线为水分输导速率为最大导水率(Kmax)的50%时的水势(P50), G表示关帝山树种, H表示黑茶山树种。
Fig. 1Leaf hydraulic vulnerability curves for eight tree species in Guandi Mountain and Heicha Mountain. A, Populus cathayana. B, Quercus mongolica. C, Ulmus pumila. D, Armeniaca sibirica. E, Populus davidiana. F, Larix gmelinii. G, Pinus tabuliformis. H, Picea asperata. The vertical lines in each graph indicate the water potential (P50) when the water transport rate is at 50% of the maximum hydraulic conductivity (Kmax). G, trees in Guandi Mountain; H, trees in Heicha Mountain.
对同一树种而言, 水分条件较好的关帝山的P50均高于水分条件较差的黑茶山, 关帝山的青杨、蒙古栎、榆树、山杏、山杨、落叶松、油松、云杉的P50分别比黑茶山的相应树种高0.36、0.37、0.01、0.18、0.12、0.18、0.95和0.59 MPa。阔叶树种中, 蒙古栎的P50在关帝山和黑茶山间的降低程度最大, 为0.37 MPa, 榆树的变化程度最小为0.01 MPa, 针叶树种中油松的降低程度最大, 为0.95 MPa, 落叶松的变化程度最小, 为0.18 MPa。在同一地区从山谷到山脊, P50逐渐下降, 关帝山8个树种的变化范围为-0.45 (青杨) - -2.43 (云杉) MPa。黑茶山的8个树种变化范围分别为-0.81 (青杨) - -3.02 (云杉) MPa (图1)。
对同一树种在关帝山和黑茶山的比较, 关帝山的青杨和榆树叶片的TLP显著高于黑茶山(p < 0.05), 关帝山其他树种的TLP也略高于黑茶山相同树种, 但没有显著差异。同一地区不同树种, 从在山谷生长的物种到山坡山脊生长的物种, TLP逐渐下降, 关帝山的8个树种的TLP变化范围为-0.94 (青杨) - -2.54 (云杉) MPa, 其中, 最低的云杉是最高的青杨的2.7倍。黑茶山的8个树种的TLP变化范围为-1.37 (青杨) - -2.77 (云杉) MPa, 其中, 最低的云杉是最高的青杨的2倍(图2)。
图2
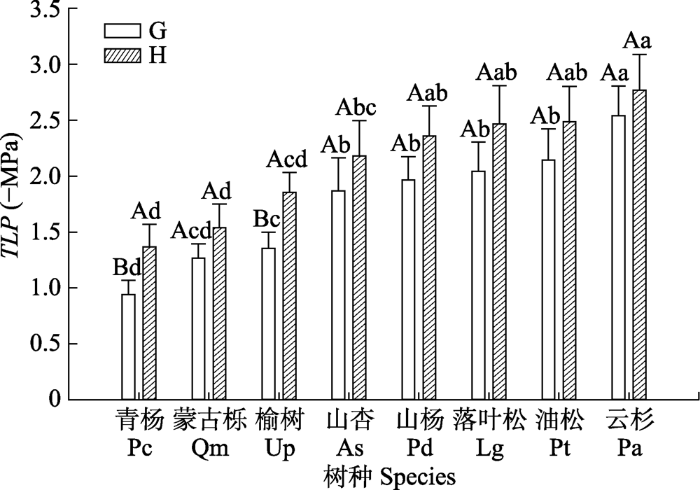 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图2关帝山和黑茶山青杨、蒙古栎、榆树、山杏、山杨、落叶松、油松和云杉叶膨压丧失点水势(TLP)的比较(平均值±标准误)。G表示关帝山树种, H表示黑茶山树种。不同大写字母代表同一树种在不同地区的差异显著, 不同小写字母代表不同树种在同一地区的差异显著(p < 0.05)。
Fig. 2Comparisons of water potential at turgor loss points (TLP) among Populus cathayana (Pc), Quercus mongolica (Qm), Ulmus pumila (Up), Armeniaca sibirica (As), Populus davidiana (Pd), Larix gmelinii (Lg), Pinus tabuliformis (Pt) and Picea asperata (Pa) in Guandi Mountain and Heicha Mountain (mean ± SE). G, trees in Guandi Mountain; H, trees in Heicha Mountain. Different uppercase letters indicate significant differences between the study areas within the same tree species; different lowercase letters indicate significant differences among tree species within the same area (p < 0.05).
阔叶树种和针叶树种相比较, 关帝山阔叶树种和针叶树种的Kmax均高于黑茶山, 阔叶树种的变化程度更大(69.90 mmol·kg-1·MPa-1·s-1)。关帝山阔叶树种和针叶树种的P50均低于黑茶山, 针叶树种在两地间的P50变化幅度更大(0.58 MPa)。关帝山阔叶树种和针叶树种的TLP均略低于黑茶山, 但差异均不显著。对同一地区而言, 阔叶树种的Kmax均高于针叶树种, P50均低于针叶树种, TLP均显著低于针叶树种(p < 0.05)(图3)。
图3
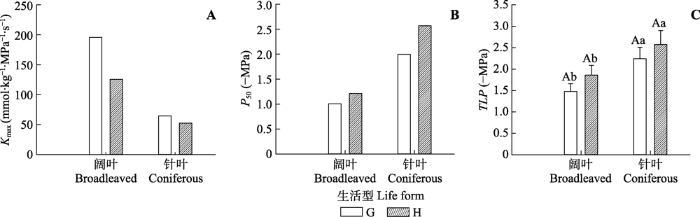 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图3关帝山和黑茶山阔叶和针叶树种叶最大导水率(Kmax)、水力脆弱性(P50)和膨压丧失点水势(TLP)的比较(平均值±标准误)。G表示关帝山树种, H表示黑茶山树种。不同大写字母代表同一树种在不同地区的差异显著, 不同小写字母代表不同树种在同一地区的差异显著(p < 0.05)。
Fig. 3Comparisons of maximum hydraulic conductivity (Kmax), hydraulic vulnerability (P50) and water potential at turgor loss points (TLP) between broadleaved and coniferous species and between Guandi Mountain and Heicha Mountain (mean ± SE). G, trees in Guandi Mountain; H, trees in Heicha Mountain. Different uppercase letters indicate significant differences between study areas within the same tree group; lowercase letters indicate significant differences between tree groups within the same study area (p < 0.05).
2.2 叶水力性状与叶导管和形态性状的相关性
关帝山和黑茶山树种的Kmax与P50之间均呈线性负相关关系, 但在不同地区两者之间表现出的关系略有差异, 随着P50的增加, 关帝山树种的Kmax下降速度略高于黑茶山。关帝山和黑茶山树种的Kmax与TLP均呈线性负相关关系, 关帝山和黑茶山树种的P50与TLP均呈线性正相关关系, 但这两种相关关系在两地之间也存在一定的差异(图4)。图4
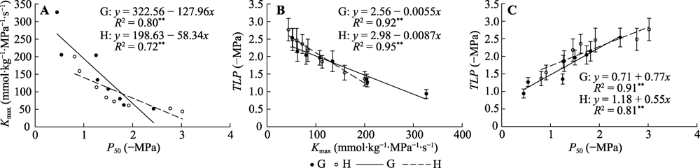 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图4关帝山和黑茶山树种叶最大导水率(Kmax)、水力脆弱性(P50)和膨压丧失点水势(TLP)的关系(平均值±标准误)。G表示关帝山树种, H表示黑茶山树种。**, p < 0.01。
Fig. 4Relationships among maximum hydraulic conductivity (Kmax), hydraulic vulnerability (P50) and water potential at turgor loss points (TLP) in Guandi Mountain and Heicha Mountain (mean ± SE). G, trees in Guandi Mountain; H, trees in Heicha Mountain. **, p < 0.01.
关帝山的P50与导管密度极显著正相关, 与(t/b)3、叶片厚度、比叶质量显著正相关, 但与导管直径和叶面积极显著负相关。关帝山树种的Kmax与导管直径极显著正相关, 与叶面积显著正相关, 与导管密度显著负相关。黑茶山树种P50与导管密度、叶片厚度和比叶质量极显著正相关, 与(t/b)3和干物质含量显著正相关, 但与导管直径极显著负相关, 与叶面积显著负相关。Kmax与导管直径和叶面积极显著正相关, 但与导管密度显著负相关(表1)。同一树种关帝山与黑茶山P50变化量(δP50)与比叶质量变化量(δLMA)和干物质含量变化量(δLDMC)均呈线性正相关关系(图5)。
Table 1
表1
表1关帝山(G)和黑茶山(H)叶水力性状与叶导管形态性状的关系
Table 1
| N (No.·mm-2) | D (μm) | (t/b)3 | LT (μm) | LA (cm2) | LMA (g·m-2) | LDMC (g·g-1) | ||
|---|---|---|---|---|---|---|---|---|
| G | P50 | 0.866** | -0.945** | 0.714* | 0.775* | -0.921** | 0.776* | 0.453 |
| Kmax | -0.749* | 0.895** | -0.563 | -0.617 | 0.813* | -0.613 | -0.392 | |
| H | P50 | 0.927** | -0.898** | 0.734* | 0.920** | -0.810* | 0.915** | 0.711* |
| Kmax | -0.724* | 0.942** | -0.597 | -0.594 | 0.900** | -0.597 | -0.327 |
新窗口打开|下载CSV
图5
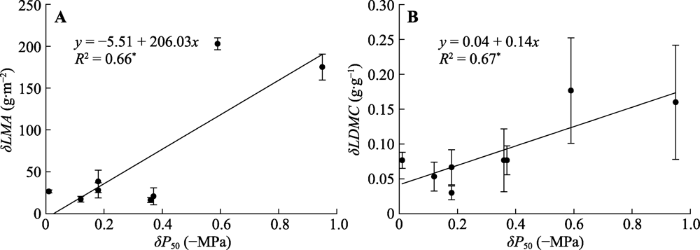 新窗口打开|下载原图ZIP|生成PPT
新窗口打开|下载原图ZIP|生成PPT图5关帝山和黑茶山两地间比叶质量变化量(δLMA)、叶干物质含量变化量(δLDMC)与水力脆弱性变化量(δP50)的关系(平均值±标准误)。*, p < 0.05。
Fig. 5Relationships of the changes in leaf mass per unit area (δLMA) and the changes in leaf dry mass content (δLDMC) with changes in hydraulic vulnerability (δP50) between Guandi Mountain and Heicha Mountain (mean ± SE). *, p < 0.05.
导管性状(VT)对P50和Kmax的影响程度均大于形态性状(MT)(表2)。两地间导管性状变化量(δVT)对P50变化量(δP50)的影响程度小于叶形态性状变化量(δMT), 两地间δVT对δKmax的影响程度大于δMT (表3)。
Table 2
表2
表2关帝山和黑茶山导管性状(VT)和形态性状(MT)对水力脆弱性(P50)和最大导水率(Kmax)的影响
Table 2
| 因子 Parameter | P50 | Kmax | ||||
|---|---|---|---|---|---|---|
| 系数 Coefficient | t | p | 系数 Coefficient | t | p | |
| VT | 0.680 | 3.875 | 0.002 | -1.034 | -3.003 | 0.010 |
| MT | 0.299 | 1.704 | 0.112 | 0.263 | 0.763 | 0.459 |
新窗口打开|下载CSV
Table 3
表3
表3关帝山和黑茶山两地间导管性状变化量(δVT)和形态性状变化量(δMT)对水力脆弱性变化量(δP50)和最大导水率变化量(δKmax)的影响
Table 3
| 因子 Parameter | δP50 | δKmax | ||||
|---|---|---|---|---|---|---|
| 系数 Coefficient | t | p | 系数 Coefficient | t | p | |
| δVT | -0.981 | -2.314 | 0.069 | -0.650 | -1.019 | 0.355 |
| δMT | 1.594 | 3.761 | 0.013 | -0.069 | -0.108 | 0.918 |
新窗口打开|下载CSV
3 讨论
3.1 叶导水率、水力脆弱性的权衡关系
以往研究通常用叶面积来标准化叶导水率(Karea), 但近10年来, 用叶质量来标准化叶导水率(Kmass)的报告逐渐增多(Nardini et al., 2012; Yan et al., 2020)。Karea侧重于叶片-大气界面上的表面生理过程, 忽略了叶肉组织中的水分运输, 只能衡量叶水力系统给叶片蒸发表面提供水分的效率。但如果叶片中的水流从叶脉组织出来后并不是直接到达表皮, 而是要在叶肉组织中流经较长的距离时(Zwieniecki et al., 2007), 用Kmass才能充分地反映出叶水力系统给整个叶组织提供水分的效率。Nardini和Luglio (2014)综合分析了全球130个木本被子植物叶片水力数据发现: P50与Karea没有相关性, 但与Kmass存在弱相关关系。因此, 在本研究中选择用叶质量来标准化叶导水率。本研究结果表明关帝山和黑茶山树种的叶P50均随Kmass增大而显著下降(图4), 这表明树种的叶水力效率与水力安全存在一定程度的权衡关系, 对叶片结构和水力结构关系的研究可以解释这种权衡关系, 如叶片水力效率和安全性与木质部特征和木质部外导水特征紧密相关, 比如水力性状可能与叶脉密度、导管直径、叶肉细胞壁体积和厚度等指标相关(Yan et al., 2020), 本研究同时也发现叶片的水力学安全性的增强基于一系列形态解剖性状的变化。系统发育分析表明叶导水效率和安全性之间存在进化关系, 但近期对茎的研究表明效率和安全性之间没有显著的进化权衡关系(Sanchez-Martinez et al., 2020), 可能是因为茎的导水主要依赖于解剖特性, 植物不同部位水力学效率和安全性之间的关系还需要更多的研究。在本研究中, Kmax和P50均与TLP存在显著相关关系(图4), 与前人的研究(Scoffoni et al., 2012; Villagra et al., 2013)一致。TLP是影响叶片对干旱胁迫耐性的重要因子(Maréchaux et al., 2015)。TLP低的树种, 其叶导水率低, 但干旱诱导的质外体抗栓塞阻力高。相反, 水分运输效率高的物种, 能够保持叶片良好的水分状况, 阻止了叶片水势的下降, 从而保持气孔开放和碳同化的时间。P50与TLP的协调关系不仅解释了叶片木质部外导水率下降的原因, 还表明了叶片内部导管导水阻力和导管外导水阻力之间的协调关系(Vilagrosaa et al., 2010; Nardini & Luglio, 2014)。在树木面临水分胁迫时, 膨压损失引起的导管外水力特性变化与导管内栓塞抵抗力的协调作用旨在诱导气孔关闭, 以响应叶片中的早期导管气穴栓塞, 从而在完全膨压损失发生之前降低蒸腾速率(Johnson et al., 2012)。从这个意义上来说, 叶的“熔断机制”可以保护活细胞不接近临界水状态。
3.2 树木叶导水率和水力安全性对水分条件的响应
叶Kleaf和P50可以在一定程度上反映树木对不同生境的适应能力。一般来说, 生长在水分条件较好的立地上的树木叶Kleaf和P50较高, 而生长在水分条件较差的立地上的树木叶Kleaf和P50较低(Nardini et al., 2012)。在本研究中, 无论是在关帝山还是黑茶山, 青杨的叶Kmax和P50在8个树种中均较高(图1), 这可能与青杨多生长在水分条件较好的山谷有关; 相反, 多生长在山坡和山脊上的树种山杏和山杨的叶Kmax和P50都相对较低(图1)。而落叶松、油松和云杉的叶Kmax和P50普遍低于阔叶树种(图1), 这一方面与其生长环境有关, 另一方面主要是因为其针叶的水分运输结构由单一叶脉所组成, 且输水单元为直径较窄的管胞, 而阔叶树种的叶片水分运输单元为直径较大的导管(Sperry et al., 2006), 因此针叶树种的输水能力较阔叶树种低。由此可见, 在同一区域范围内, 叶片导水率和水力脆弱性可以影响植物的分布, 高的叶Kleaf被认为是在水分充足生境中适应性的重要方面, 在水分较为充足的环境中, 对光和养分的竞争成为物种间竞争的主要部分, 高光合速率和生长速率树种能在水分充足的环境中占优势。相反, 叶Kleaf低的树种, 常有较高的栓塞阻力, 但栓塞脆弱性明显降低, 这使得此类物种在水资源减少的情况下保持正常成长, 但降低了这些树种在高水分资源生境中的竞争力(Nardini et al., 2012)。对于不同立地条件下的同一树种, 从水分条件较好的关帝山到水分条件较差的黑茶山, Kleaf和P50均降低, 这种改变, 损失了部分水分输导效率, 进而损失了在资源充沛条件下的生长速度和竞争能力, 但增加了在水分匮乏条件下的水力结构维持能力(Nardini et al., 2012), 这是树木对水分条件变化在水力结构上的响应, 有利于树木在水分匮乏条件下的水力结构稳定性维持。
3.3 叶水力性状与叶导管和形态性状之间的关系
叶片通过一系列形态和解剖结构调整, 提高植物的抗旱能力(Simonin et al., 2012)。有研究表明叶片水力脆弱性与主脉木质部导管直径和导管密度密切相关, 但与导管壁厚度无关(Nardini et al., 2012)。近年来使用核磁成像技术的研究表明叶片的气穴栓塞前期主要发生在主脉部位, 从而形成“熔断机制”保护叶柄以下枝条的水分供应(Scoffoni et al., 2017)。对干旱引起的功能障碍最常见的解释是木质部导管的气穴栓塞导致导水率降低, 近年来的研究表明大部分叶片的木质部同样容易受到塌陷诱导的功能障碍的影响。本研究结果表明, 关帝山和黑茶山树种的叶P50与叶主脉导管直径显著负相关, 同时与导管塌陷预测值(t/b)3显著正相关(表1), 这表明叶水力学安全性与叶片木质部导管关系密切, 也表明叶片导管内水力结构维持需要导管抗栓塞和抗塌陷能力的协同。在本研究中也发现两地叶片的导水率与导管直径极显著正相关, 而与导管密度显著负相关, P50与导管密度极显著正相关(表1), 这表明植物在降低叶木质部导管直径的同时增加了导管密度, 这可以在一定程度上弥补导管直径减小造成的导水率下降。叶导水率和水力脆弱性除受到导管特征的影响外, 也受到导管外细胞间水分输导组织的影响。本研究结果显示, P50除了与叶脉导管有密切关系外, 还与比叶质量和叶片厚度显著正相关(表1), 这表明P50的降低也依靠比叶质量和叶片厚度的增加来实现, 叶自身水力安全提高的同时, 也增加了叶片组织构建上的碳投资, 提高了叶片水分利用的成本, 对枝条水分输导安全性的研究也得到相似的结果(Hoffmann et al., 2011)。同时, 本研究也发现Kmax与叶面积显著正相关, P50与叶面积显著负相关(表1), 但普适性的水力性状与叶片大小的关系也还需要更多的试验证据。
叶片的水力学特性由导管内和导管外水力特性两部分组成, 叶片导管性状和叶片形态性状分别表征叶片导管内和导管外的水力学特性, 本研究在对导管性状和形态性状进行主成分分析的基础上, 对不同树种叶导水率和水力脆弱性影响指标的多元线性回归分析结果表明: Kmax和P50受到叶片导管性状和形态性状两方面的影响, 且导管性状对Kmax和P50的影响程度均大于叶片形态性状(表2), 这表明不同树种的叶片导水率和水力脆弱性的差别受到导管性状和叶片形态性状两方面的影响, 且导管性状对Kmax和P50的影响程度更大。本研究还比较了同一树种在关帝山和黑茶山的叶导水率和水力脆弱性变化量与叶片导管和形态变化量的关系, 结果表明同一树种在不同地区的P50变化量与比叶质量变化量、叶片干物质变化量呈显著正相关关系(图5), 且δP50与叶形态特征变化量的关系更大(表3), 这表明在水分匮乏条件下, 本研究中树种P50降低的主要模式是改变叶片厚度和干物质含量, 这表明本研究选择树种P50对水分条件变差的响应主要来自导管外叶片形态特征的响应。这种响应模式增加了叶片的碳投资, 但有利于水力结构的维持。对不同树种Kmax和P50变化及同一树种在不同水分条件下水力特性变化的研究结果表明: 树木叶导水率和水力脆弱性在树种间和树种内的差别对水分条件的响应模式是不同的, 不同树种的差别主要依靠叶导管性状的差别实现, 同一树种在水分条件变化时, 叶水力脆弱性的适应性改变主要依靠改变比叶质量等叶片形态性状来实现。但这一规律的普适性, 需要有更多的试验证据, 这是我们之后进行深入研究的重要内容。
在本研究中, 在对Kmax和P50进行多元线性回归分析之前, 采用主成分分析的方法来提取叶导管性状和叶形态性状的主成分信息, 虽然主成分的贡献率均达到或接近80%, 也会造成部分原始数据信息量的损失。
综上所述, 在晋西北森林草原过渡区, 不同树种叶片水力效率与安全性之间存在一定的权衡关系, 在水分匮乏条件下, 各个树种均有相对一致的响应, 即降低叶片导水率和水力脆弱性, 这种响应有利于树木叶片水力结构的维持。不同树种叶水力性状差别受到叶导管性状和叶片结构性状的影响, 受叶导管性状影响程度更大, 同一树种叶水力脆弱性对水分条件变化的响应主要依靠导管外叶片形态性状的驱动。树木在提高自身水力安全的同时增加了叶片构建上的碳投资。
参考文献 原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
PMID:12913171 [本文引用: 1]

The question as to what triggers stomatal closure during leaf desiccation remains controversial. This paper examines characteristics of the vascular and photosynthetic functions of the leaf to determine which responds most similarly to stomata during desiccation. Leaf hydraulic conductance (K(leaf)) was measured from the relaxation kinetics of leaf water potential (Psi(l)), and a novel application of this technique allowed the response of K(leaf) to Psi(l) to be determined. These "vulnerability curves" show that K(leaf) is highly sensitive to Psi(l) and that the response of stomatal conductance to Psi(l) is closely correlated with the response of K(leaf) to Psi(l). The turgor loss point of leaves was also correlated with K(leaf) and stomatal closure, whereas the decline in PSII quantum yield during leaf drying occurred at a lower Psi(l) than stomatal closure. These results indicate that stomatal closure is primarily coordinated with K(leaf). However, the close proximity of Psi(l) at initial stomatal closure and initial loss of K(leaf) suggest that partial loss of K(leaf) might occur regularly, presumably necessitating repair of embolisms.
PMID:15734905 [本文引用: 1]
Just as a soggy paper straw is prone to yielding under the applied suction of a thirsty drinker, the xylem tracheids in leaves seem prone to collapse as water potential declines, impeding their function. Here we describe the collapse, under tension, of lignified cells peripheral to the leaf vein of a broad-leaved rainforest conifer, Podocarpus grayi de Laub. Leaves of Podocarpus are characterized by an array of cylindrical tracheids aligned perpendicular to the leaf vein, apparently involved in the distribution of water radially through the mesophyll. During leaf desiccation the majority of these tracheids collapsed from circular to flat over the water potential range -1.5 to -2.8 MPa. An increase in the percentage of tracheids collapsed during imposed water stress was mirrored by declining leaf hydraulic conductivity (K(leaf)), implying a direct effect on water transport efficiency. Stomata responded to water stress by closing at -2.0 MPa when 45% of cells were collapsed and K(leaf) had declined by 25%. This was still substantially before the initial indications of cavitation-induced loss of hydraulic conductance in the leaf vein, at -3 MPa. Plants droughted until 49% of tracheids had collapsed were found to fully recover tracheid shape and leaf function 1 week after rewatering. A simple mechanical model of tracheid collapse, derived from the theoretical buckling pressure for pipes, accurately predicted the collapse dynamics observed in P. grayi, substantiating estimates of cell wall elasticity and measured leaf water potential. The possible adaptive advantages of collapsible vascular tissue are discussed.
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/nph.13646PMID:26378984 [本文引用: 1]
The evolution of lignified xylem allowed for the efficient transport of water under tension, but also exposed the vascular network to the risk of gas emboli and the spread of gas between xylem conduits, thus impeding sap transport to the leaves. A well-known hypothesis proposes that the safety of xylem (its ability to resist embolism formation and spread) should trade off against xylem efficiency (its capacity to transport water). We tested this safety-efficiency hypothesis in branch xylem across 335 angiosperm and 89 gymnosperm species. Safety was considered at three levels: the xylem water potentials where 12%, 50% and 88% of maximal conductivity are lost. Although correlations between safety and efficiency were weak (r(2) < 0.086), no species had high efficiency and high safety, supporting the idea for a safety-efficiency tradeoff. However, many species had low efficiency and low safety. Species with low efficiency and low safety were weakly associated (r(2) < 0.02 in most cases) with higher wood density, lower leaf- to sapwood-area and shorter stature. There appears to be no persuasive explanation for the considerable number of species with both low efficiency and low safety. These species represent a real challenge for understanding the evolution of xylem. No claim to US government works. New Phytologist © 2015 New Phytologist Trust.
[本文引用: 1]
[本文引用: 1]
DOI:10.3724/SP.J.1259.2014.00203URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/j.1365-2486.2011.02401.xURL [本文引用: 1]
DOI:10.17521/cjpe.2016.0064URL [本文引用: 1]
DOI:10.17521/cjpe.2016.0064 [本文引用: 1]
不同材性树种的解剖、叶脉分布等结构性状差异会影响树木的水分运输效率和水分利用策略, 进而限制树木的生存、生长和分布。然而, 材性对叶导水率、水力脆弱性及其潜在的权衡关系的影响尚不清楚。该研究选择东北温带森林中不同材性的9种树种(散孔材: 山杨(Populus davidiana)、紫椴(Tilia amurensis)、白桦(Betula platyphylla); 环孔材: 蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、胡桃楸(Juglans mandshurica); 无孔材: 红皮云杉(Picea koraiensis)、樟子松(Pinus sylvestris var. mongolica)、红松(Pinus koraiensis), 测量其基于叶面积和叶质量的叶导水率(K<sub>area</sub>和K<sub>mass</sub>)、水力脆弱性(P<sub>50</sub>)、膨压丧失点水势(TLP)及叶结构性状, 以比较不同材性树种叶水力性状的差异, 并探索叶水力效率与安全的权衡关系。结果表明: 3种材性树种的K<sub>area</sub>、K<sub>mass</sub>和P<sub>50</sub>均差异显著(p < 0.05)。无孔材树种的K<sub>area</sub>和K<sub>mass</sub>最低, 而散孔材和环孔材树种差异不显著; 环孔材树种P<sub>50</sub>最高, 而散孔材和无孔材树种差异不显著。K<sub>area</sub>和K<sub>mass</sub>均与P<sub>50</sub>显著负相关(p < 0.05), 但散孔材、环孔材和无孔材树种的相关关系分别呈线性、幂函数和指数函数关系。这表明叶水力效率与安全之间存在一定的权衡关系, 但该关系受树木材性的影响。K<sub>mass</sub>与TLP显著负相关(p < 0.01), 其中散孔材和环孔材树种呈线性负相关, 无孔材树种呈负指数函数关系; P<sub>50</sub>随TLP的增加而增加, 这表明树木在面临水分胁迫时, 其质外体和共质体抗旱阻力共同协调保护叶片活细胞, 防止其水分状况到达临界阈值。K<sub>mass</sub>与叶干物质含量、叶密度、比叶重均显著负相关, 而P<sub>50</sub>与之显著正相关(p < 0.01, P<sub>50</sub>与比叶重的关系除外), 表明树木叶水力特性的变化受相同叶结构特性驱动, 树木增加对水力失调的容忍需要在叶水力系统构建上增加碳投资。
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
[本文引用: 1]
DOI:10.1111/fec.2015.29.issue-10URL [本文引用: 1]
[本文引用: 1]
DOI:10.1111/nph.16151PMID:31545889 [本文引用: 1]
DOI:10.1111/fec.2014.28.issue-4URL [本文引用: 5]
DOI:10.1111/j.1469-8137.2012.04294.xPMID:22978628 [本文引用: 6]
Leaf hydraulic conductance (K(leaf) ) and vulnerability constrain plant productivity, but no clear trade-off between these fundamental functional traits has emerged in previous studies. We measured K(leaf) on a leaf area (K(leaf_area)) and mass basis (K(leaf_mass)) in six woody angiosperms, and compared these values with species' distribution and leaf tolerance to dehydration in terms of P(50), that is, the leaf water potential inducing 50% loss of K(leaf). We also measured several morphological and anatomical traits associated with carbon investment in leaf construction and water transport efficiency. Clear relationships emerged between K(leaf_mass), P(50), and leaf mass per unit area (LMA), suggesting that increased tolerance to hydraulic dysfunction implies increased carbon costs for leaf construction and water use. Low P(50) values were associated with narrower and denser vein conduits, increased thickness of conduit walls, and increased vein density. This, in turn, was associated with reduced leaf surface area. Leaf P(50) was closely associated with plants' distribution over a narrow geographical range, suggesting that this parameter contributes to shaping vegetation features. Our data also highlight the carbon costs likely to be associated with increased leaf tolerance to hydraulic dysfunction, which confers on some species the ability to thrive under reduced water availability but decreases their competitiveness in high-resource habitats.© 2012 The Authors. New Phytologist © 2012 New Phytologist Trust.
PMID:16246857 [本文引用: 1]
The hydraulic architecture, water relationships, and gas exchange of leaves of sunflower plants, grown under different levels of water stress, were measured. Plants were either irrigated with tap water (controls) or with PEG600 solutions with osmotic potential of -0.4 and -0.8 MPa (PEG04 and PEG08 plants, respectively). Mature leaves were measured for hydraulic resistance (R(leaf)) before and after making several cuts across minor veins, thus getting the hydraulic resistance of the venation system (R(venation)). R(leaf) was nearly the same in controls and PEG04 plants but it was reduced by about 30% in PEG08 plants. On the contrary, R(venation) was lowest in controls and increased in PEG04 and PEG08 plants as a likely result of reduction in the diameter of the veins' conduits. As a consequence, the contribution of R(venation) to the overall R(leaf) markedly increased from controls to PEG08 plants. Leaf conductance to water vapour (g(L)) was highest in controls and significantly lower in PEG04 and PEG08 plants. Moreover, g(L) was correlated to R(venation) and to leaf water potential (psi(leaf)) with highly significant linear relationships. It is concluded that water stress has an important effect on the hydraulic construction of leaves. This, in turn, might prove to be a crucial factor in plant-water relationships and gas exchange under water stress conditions.
[本文引用: 1]
DOI:10.1146/arplant.2006.57.issue-1URL [本文引用: 1]
DOI:10.1111/ele.v23.11URL [本文引用: 1]
DOI:10.1111/nph.2017.213.issue-3URL [本文引用: 1]
DOI:10.1093/jxb/err270URL [本文引用: 2]
[本文引用: 1]
DOI:10.3732/ajb.93.10.1490PMID:21642096 [本文引用: 1]
The wide size range of conifer tracheids and angiosperm vessels has important consequences for function. In both conduit types, bigger is better for conducting efficiency. The gain in efficiency with size is maximized by the control of conduit shape, which balances end-wall and lumen resistances. Although vessels are an order of magnitude longer than tracheids of the same diameter, they are not necessarily more efficient because they lack the low end-wall resistance of tracheids with torus-margo pits. Instead, vessels gain conducting efficiency over tracheids by achieving wider maximum diameters. End-walls contributed 56-64% to total xylem resistance in both conduit types, indicating that length limits conducting efficiency. Tracheid dimensions may be more limited by unicellularity and the need to supply strength to homoxylous wood than by the need to protect against cavitation. In contrast, the greater size of the multicellular vessel is facilitated by fibers that strengthen heteroxylous wood. Vessel dimensions may be most limited by the need to restrict intervessel pitting and cavitation by air-seeding. Stressful habitats that promote narrow vessels should favor coexistence of conifers and angiosperms. The evolution of vessels in angiosperm wood may have required early angiosperms to survive a phase of mechanic and hydraulic instability.
DOI:10.1111/nph.1991.119.issue-3URL [本文引用: 1]
DOI:10.1093/jxb/23.1.267URL [本文引用: 1]
[本文引用: 1]
DOI:10.1093/treephys/tpt098URLPMID:24284866 [本文引用: 2]
Leaves can be both a hydraulic bottleneck and a safety valve against hydraulic catastrophic dysfunctions, and thus changes in traits related to water movement in leaves and associated costs may be critical for the success of plant growth. A 4-year fertilization experiment with nitrogen (N) and phosphorus (P) addition was done in a semideciduous Atlantic forest in northeastern Argentina. Saplings of five dominant canopy species were grown in similar gaps inside the forests (five control and five N?+?P addition plots). Leaf lifespan (LL), leaf mass per unit area (LMA), leaf and stem vulnerability to cavitation, leaf hydraulic conductance (K(leaf_area) and K(leaf_mass)) and leaf turgor loss point (TLP) were measured in the five species and in both treatments. Leaf lifespan tended to decrease with the addition of fertilizers, and LMA was significantly higher in plants with nutrient addition compared with individuals in control plots. The vulnerability to cavitation of leaves (P50(leaf)) either increased or decreased with the nutrient treatment depending on the species, but the average P50(leaf) did not change with nutrient addition. The P50(leaf) decreased linearly with increasing LMA and LL across species and treatments. These trade-offs have an important functional significance because more expensive (higher LMA) and less vulnerable leaves (lower P50(leaf)) are retained for a longer period of time. Osmotic potentials at TLP and at full turgor became more negative with decreasing P50(leaf) regardless of nutrient treatment. The K(leaf) on a mass basis was negatively correlated with LMA and LL, indicating that there is a carbon cost associated with increased water transport that is compensated by a longer LL. The vulnerability to cavitation of stems and leaves were similar, particularly in fertilized plants. Leaves in the species studied may not function as safety valves at low water potentials to protect the hydraulic pathway from water stress-induced cavitation. The lack of rainfall seasonality in the subtropical forest studied probably does not act as a selective pressure to enhance hydraulic segmentation between leaves and stems.
URL [本文引用: 5]
DOI:10.1111/nph.v229.1URL [本文引用: 1]
[本文引用: 1]
Stomatal closure during leaf dehydration, correlation with other leaf physiological traits
1
2003
... 叶水力脆弱性曲线采用复水法测定(Brodribb & Holbrook,
Water stress deforms tracheids peripheral to the leaf vein of a tropical conifer
1
2005
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
Functional relationships between hydraulic traits and the timing of diurnal depression of photosynthesis
1
2019
... 前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al.,
黑茶林局主要阔叶树造林现状及对策建议
1
2019
... 研究地分别选择在关帝山(G)和黑茶山(H).关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等,
黑茶林局主要阔叶树造林现状及对策建议
1
2019
... 研究地分别选择在关帝山(G)和黑茶山(H).关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等,
Weak tradeoff between xylem safety and xylem-specific hydraulic efficiency across the world's woody plant species
1
2016
... 前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al.,
关帝山森林景观异质性及其动态的研究
1
1999
... 研究地分别选择在关帝山(G)和黑茶山(H).关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等,
关帝山森林景观异质性及其动态的研究
1
1999
... 研究地分别选择在关帝山(G)和黑茶山(H).关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等,
植物材料快速石蜡制片方法
1
2014
... 清晨在不同样木上随机采集健康的成熟叶片用FAA固定液固定, 带回实验室进行石蜡制片.参照贺冰等(
植物材料快速石蜡制片方法
1
2014
... 清晨在不同样木上随机采集健康的成熟叶片用FAA固定液固定, 带回实验室进行石蜡制片.参照贺冰等(
Hydraulic failure and tree dieback are associated with high wood density in a temperate forest under extreme drought
1
2011
... 叶导水率和水力脆弱性除受到导管特征的影响外, 也受到导管外细胞间水分输导组织的影响.本研究结果显示, P50除了与叶脉导管有密切关系外, 还与比叶质量和叶片厚度显著正相关(
九种不同材性的温带树种叶水力性状及其权衡关系
1
2016
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
九种不同材性的温带树种叶水力性状及其权衡关系
1
2016
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
Evidence for xylem embolism as a primary factor in dehydration-induced declines in leaf hydraulic conductance
1
2012
... 在本研究中, Kmax和P50均与TLP存在显著相关关系(
Leaf xylem embolism, detected acoustically and by cryo-SEM, corresponds to decreases in leaf hydraulic conductance in four evergreen species
1
2009
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
黄土高原林下草地对模拟增温的短期响应
1
2020
... 研究地分别选择在关帝山(G)和黑茶山(H).关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等,
黄土高原林下草地对模拟增温的短期响应
1
2020
... 研究地分别选择在关帝山(G)和黑茶山(H).关帝山地处吕梁山脉中部, 地理位置37.33°-38.33° N, 110.30°-111.30° E, 属温带大陆性季风气候, 平均海拔1 800 m, 年平均气温4 ℃, 年降水量831 mm, 无霜期100-125天, 土壤类型为山地棕壤, 平均土层厚度为75 cm, 全区主要是破碎陡峭的山地地貌, 岩石以花岗岩和片麻岩为主, 主要树种有油松(Pinus tabuliformis)、蒙古栎(Quercus mongolica)、山杨(Populus davidiana)、落叶松(Larix gmelinii)、云杉(Picea asperata)等(郭晋平等,
Drought tolerance as predicted by leaf water potential at turgor loss point varies strongly across species within an Amazonian forest
1
2015
... 在本研究中, Kmax和P50均与TLP存在显著相关关系(
The dynamic pipeline: hydraulic capacitance and xylem hydraulic safety in four tall conifer species
1
2014
... 水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al.,
Hydraulics in the 21st century
1
2019
... 水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al.,
Leaf hydraulic capacity and drought vulnerability: possible trade-offs and correlations with climate across three major biomes
5
2014
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
... ), 叶水力脆弱性对于逆境条件下叶水力结构的维持至关重要, 能在低水势条件下维持水力结构的稳定(Nardini & Luglio,
... 前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al.,
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近10年来, 用叶质量来标准化叶导水率(Kmass)的报告逐渐增多(Nardini et al.,
... 在本研究中, Kmax和P50均与TLP存在显著相关关系(
Trade-offs between leaf hydraulic capacity and drought vulnerability: morpho- anatomical bases, carbon costs and ecological consequences
6
2012
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近10年来, 用叶质量来标准化叶导水率(Kmass)的报告逐渐增多(Nardini et al.,
... 叶Kleaf和P50可以在一定程度上反映树木对不同生境的适应能力.一般来说, 生长在水分条件较好的立地上的树木叶Kleaf和P50较高, 而生长在水分条件较差的立地上的树木叶Kleaf和P50较低(Nardini et al.,
... .,
... 对于不同立地条件下的同一树种, 从水分条件较好的关帝山到水分条件较差的黑茶山, Kleaf和P50均降低, 这种改变, 损失了部分水分输导效率, 进而损失了在资源充沛条件下的生长速度和竞争能力, 但增加了在水分匮乏条件下的水力结构维持能力(Nardini et al.,
... 叶片通过一系列形态和解剖结构调整, 提高植物的抗旱能力(Simonin et al.,
Water stress-induced modifications of leaf hydraulic architecture in sunflower: co-ordination with gas exchange
1
2005
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
1
2016
... 水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al.,
Leaf hydraulics
1
2006
... 叶导水率(Kleaf)和叶水力脆弱性(常用水分输导速率为最大导水率的50%时的水势(P50)表征)常用于反映叶片的水分输导效率和安全性(Nardini & Luglio,
Adaptation and coordinated evolution of plant hydraulic traits
1
2020
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近10年来, 用叶质量来标准化叶导水率(Kmass)的报告逐渐增多(Nardini et al.,
Leaf vein xylem conduit diameter influences susceptibility to embolism and hydraulic decline
1
2017
... 叶片通过一系列形态和解剖结构调整, 提高植物的抗旱能力(Simonin et al.,
Dynamics of leaf hydraulic conductance with water status: quantification and analysis of species differences under steady state
2
2012
... 前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al.,
... 在本研究中, Kmax和P50均与TLP存在显著相关关系(
Hydraulic conductance of leaves correlates with leaf lifespan: implications for lifetime carbon gain
1
2012
... 叶片通过一系列形态和解剖结构调整, 提高植物的抗旱能力(Simonin et al.,
Size and function in conifer tracheids and angiosperm vessels
1
2006
... 叶Kleaf和P50可以在一定程度上反映树木对不同生境的适应能力.一般来说, 生长在水分条件较好的立地上的树木叶Kleaf和P50较高, 而生长在水分条件较差的立地上的树木叶Kleaf和P50较低(Nardini et al.,
The hydraulic architecture of trees and other woody plants
1
1991
... 水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al.,
The measurement of the turgor pressure and the water relations of plants by the pressure- bomb technique
1
1972
... Cleaf通过压力-容积(PV)曲线测定.利用自然风干法制作PV曲线(Tyree & Hammel,
Are symplast tolerance to intense drought conditions and xylem vulnerability to cavitation coordinated? An integrated analysis of photosynthetic, hydraulic and leaf level processes in two Mediterranean drought-resistant species
1
2010
... 在本研究中, Kmax和P50均与TLP存在显著相关关系(
Functional relationships between leaf hydraulics and leaf economic traits in response to nutrient addition in subtropical tree species
2
2013
... 水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al.,
... 在本研究中, Kmax和P50均与TLP存在显著相关关系(
Leaf hydraulic safety margin and safety-efficiency trade-off across angiosperm woody species
5
2020
... 水力结构在树木存活、生长、竞争和分布上起着关键作用(Sack et al.,
... 前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al.,
... .,
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近10年来, 用叶质量来标准化叶导水率(Kmass)的报告逐渐增多(Nardini et al.,
... .,
Combined high leaf hydraulic safety and efficiency provides drought tolerance in Caragana species adapted to low mean annual precipitation
1
2021
... 前人大量报道了枝条导水率和栓塞脆弱性之间的相关关系(Gleason et al.,
Hydraulic design of leaves: insights from rehydration kinetics
1
2007
... 以往研究通常用叶面积来标准化叶导水率(Karea), 但近10年来, 用叶质量来标准化叶导水率(Kmass)的报告逐渐增多(Nardini et al.,

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}