Phenotypic plasticity and genetic differentiation of quantitative traits in genotypes of Leymus chinensis
YANGXue, SHENJun-Fang, ZHAONian-Xi*,, GAOYu-Bao College of Life Sciences, NanKai University, Tianjin 300071, China 通讯作者:* 通信作者Author for correspondence (E-mail:zhaonianxi@nankai.edu.cn) 版权声明:2017植物生态学报编辑部本文是遵循CCAL协议的开放存取期刊,引用请务必标明出处。 基金资助:国家自然科学基金(31570427)
关键词:羊草;基因型;适应;遗传分化;表型可塑性;反应规范 Abstract Aims Adaptation mechanisms of plants to environment can be classified as genetic differentiation and phenotypic plasticity (environmental modification). The strategy and mechanism of plant adaptation is a hot topic in the field of evolutionary ecology. Leymus chinensis is one of constructive species in the Nei Mongol grassland. Particularly, Leymus chinensis is a rhizomatous and clonally reproductive grass, a genotype that can play an important role in the community. In this study, we aimed to (1) investigate the phenotypic plasticity of L. chinensis under different conditions, and (2) test the genetic differentiation and reaction norms (the relationship between the environment and the phenotype of an individual or a group of individuals) under four environmental conditions among different genotypes of L. chinensis. Methods Ten genotypes of L. chinensis were randomly selected. Under the control condition, we studied the effects of genotype, defoliation, drought and their interactions on 11 quantitative traits of growth (8 traits including photochemical efficiency of photosystem II, maximum net photosynthetic rate, transpiration rate, specific leaf area, relative growth rate, the number of tillers increased, aboveground and underground biomass growth), defense (total phenol concentration of leaf) and tolerance (non-structural carbohydrate content of root, root/shoot ratio) of L. chinensis. We studied the phenotypic plasticity, genetic differentiation and reaction norms mainly through tested the effect of environment and genotype on these traits. Important findings First, all 11 traits showed obvious phenotypic plasticity (i.e., significant effect of drought, defoliation and their interactions). The expression of 10 genotypes of L. chinensis was divergent under different environmental conditions. Interactions of genotype and environment significantly affected the maximum net photosynthetic rate, transpiration rate, specific leaf area, relative growth rate, total phenolic concentration of leaf, and total non-structural carbohydrate content of root. This indicated that the phenotypic plasticity of these five traits exhibited genetic differentiation. Second, the increase of number of tillers, belowground biomass and non-structural carbohydrate content of root did not show genetic differentiation under the same condition. The other eight traits showed significantly genetic differentiation, and the heritabilities (H2) of six traits related to growth were higher than 0.5. The leaf total phenol concentration and root/shoot ratio showed genetically differentiation only under the drought and defoliation condition, with the heritabilities being 0.145 and 0.201, respectively. These results explained why L. chinensis widely distributed in the Nei Mongol grassland, and provided genetic and environmental basis for related application and species conservation in this grassland ecosystem.
Keywords:Leymus chinensis;genotype;adaptation;genetic differentiation;phenotypic plasticity;reaction norm -->0 PDF (2815KB)元数据多维度评价相关文章收藏文章 本文引用格式导出EndNoteRisBibtex收藏本文--> 杨雪, 申俊芳, 赵念席, 高玉葆. 不同基因型羊草数量性状的可塑性及遗传分化. 植物生态学报, 2017, 41(3): 359-368 https://doi.org/10.17521/cjpe.2015.0257 YANGXue, SHENJun-Fang, ZHAONian-Xi, GAOYu-Bao. Phenotypic plasticity and genetic differentiation of quantitative traits in genotypes of Leymus chinensis. Chinese Journal of Plant Ecology, 2017, 41(3): 359-368 https://doi.org/10.17521/cjpe.2015.0257 生物在生长发育过程中不可避免地经历外部环境的变化, 生物对环境的适应机制主要有两种: 遗传分化(genetic differentiation)和表型可塑性(phenotypic plasticity) (Liefting et al., 2009)。前者指通过遗传的改变来产生可稳定遗传的表型变异(Hedrick, 2005); 后者则通过环境饰变来产生依赖于环境的表型变异(Sultan, 2000)。同一基因型在不同环境下产生的一系列表型被称为反应规范(reaction norm) (Miner et al., 2005), 反应规范在同一物种内不同基因型间也会存在显著差异(环境因素×基因型作用显著), 即反应规范的遗传分化或表型可塑性的遗传分化(Mallitt et al., 2010)。表型可塑性与遗传分化是生物适应环境变化的两种不同但并不互相排斥的策略, 同一物种的适应可以既有表型可塑性又有遗传分化, 两者对环境变化适应的相对重要性与具体性状、生态因子等有关(Sultan, 1995)。对物种适应机制的深入研究不仅能够了解该物种对环境的适应策略和维持机制, 而且能够对未来全球变化下物种的进化潜力进行正确预测, 因此, 该 方面的研究一直以来都是种群生态学及种群遗传学研究的热点问题(Lande, 2009; Draghi & Whitlock, 2012)。 随着种群遗传学研究的不断深入, 人们发现种内基因型间的性状变异不仅对种群的适应性进化具有重要作用, 而且能发挥物种水平的作用, 对群落乃至生态系统进程也具有重要影响(Crutsinger et al., 2009; Cook-Patton et al., 2011)。如不同基因型的大叶藻(Zostera marina)不仅在生物量方面存在差异, 而且在铵态氮和硝态氮的利用方面也存在差异, 不同基因型表现出养分利用的分化(Hughes et al., 2009); 不同基因型的美洲山杨(Populus tremuloides)对食草动物的防御和耐受存在差异, 且这种差异是由遗传因素决定的(Stevens et al., 2007)。因此, 关于重要物种基因型间差异的研究不仅能反映植物适应策略, 而且对该物种在群落中的重要作用的正确评估也有一定的积极意义(Johnson & Agrawal, 2005; Kanaga et al., 2008; Hughes et al., 2009; Le Corre & Kremer, 2012; Strasburg et al., 2012)。 近半个世纪, 特别是20世纪80年代以来, 全球气候变化和人类活动加剧引起生态系统退化加重。草原作为重要的生态系统, 退化尤为严重, 草原群落的恢复和重建成为亟须解决的问题(Joshi et al., 2006)。而这一问题的解决主要集中在草原建群种和优势种对环境变化的适应机制和适应策略的研究上, 因为优势物种对生态系统有重要的指示和调节作用(Seliskar et al., 2002)。羊草(Leymus chinensis)作为内蒙古草原的建群种, 具有生态幅广、适应性强等特点, 如早期的研究发现, 羊草种群间遗传分化显著(胡宝忠等, 2001; 刘惠芬等, 2004a, 2004b), 数量性状(生长和生理性状)存在显著的差异(Wang & Gao, 2001; 李红等, 2004; 李海燕等, 2011); 但针对这一重要物种适应机制的探讨并没有深入展开。另外, 羊草作为根茎型禾草, 通过克隆繁殖, 某些特定的基因型在群落中也能起到很重要的作用。最近有研究发现, 羊草不同基因型间具有类似于物种间的生态位互补效应(申俊芳等, 2015), 但这种互补效应是否有遗传学基础并没有得到证实, 且不同基因型间性状的差异是否能够反映适应策略的不同也未见报道。因此, 基于内蒙古草原区一直经历严重的放牧压力和水分分布不均匀的特点, 本研究选择不同基因型羊草为研究对象, 并进行刈割、干旱处理, 探讨: (1)随环境条件变化, 同一基因型羊草的数量性状的表型可塑性及对应的适应策略; (2)性状的反应规范在基因型间是否存在显著差异; (3)同一环境条件下, 羊草数量性状是否受基因型影响显著, 并重点关注胁迫环境下与防御和耐受相关的性状。所得结果不仅能够了解羊草对环境变化的适应机制及策略, 为合理预测未来气候变化对该物种的影响提供理论基础, 而且对羊草种群及其所在群落的合理利用、保护和资源开发也具有重要的意义。
1 材料和方法
1.1 实验材料
本实验所用羊草为2010年于内蒙古锡林浩特市阿巴嘎旗典型草原西界(43.90° N, 115.34° E)采集的羊草基株, 利用ISSR分子标记(AG)7T和(CA)6A确定基因型(王丹, 2004), 并进行编号。在同样的条件下培养以去除母体效应, 通过根茎无性繁殖得到同一基因型的大量分蘖。从已有的基因型中随机选择10个基因型(Y7、Y8、Y18、Y23、Y24、Y25、3/0、3/1、3/25、4/6)进行实验。
基因型、干旱、刈割及其交互作用对不同指标的影响并不完全相同(表1), 图1比较直观地给出了10种基因型羊草在4种处理下各个指标的表型可塑性和反应规范, 结果发现: 没有哪个基因型在所有观测的指标中都表现出最佳值; 刈割、干旱及其交互作用对多数观测指标影响显著(表1)(p < 0.05), 即所有性状均具有表型可塑性, 如刈割处理提高RGR以及叶总酚浓度(图1E、1I), 干旱处理提高植物根冠比(图1K)。而基因型与环境因子之间交互作用只对某些指标影响显著, 如基因型×刈割对Pnmax、Tr、RGR、叶总酚浓度和根TNC总量, 基因型×干旱对Tr、SLA、RGR、叶总酚浓度和根TNC总量, 以及基因型×刈割×干旱仅对RGR具有显著影响(p < 0.05), 表明这些性状的表型可塑性具有遗传学基础。具体来分析, 相对于对照(De-Dr-), 刈割干旱处理(De+Dr+)下基因型3/25中叶总酚浓度(防御相关)显著升高, 而Y24中叶总酚浓度降低(图1I); 相对于对照(De-Dr-), 刈割处理(De+Dr-)使基因型3/25中根TNC总量(耐受相关)显著升高, 而Y8、Y18无明显变化(图1J)。 Table 1 表1 表1不同基因型羊草表型变异的多因素方差分析及交互作用 Table 1Multivariate analysis of variance for genotype and phenotypic variations and interactions of genotype and treatments in Leymus chinensis
性状 Traits
基因型 Genotype (G)
刈割 Defoliation (De)
干旱 Drought (Dr)
De × Dr
基因型×刈割 G × De
基因型×干旱 G × Dr
基因型× 刈割×干旱 G × De × Dr
F
p
F
p
F
p
F
p
F
p
F
p
F
p
光系统II光化学效率 Photochemical efficiency of photosystem II
19.47
<0.001
2.649
0.105
17.19
<0.001
0.476
0.491
1.064
0.391
0.912
0.516
1.824
0.066
最大净光合速率 Maximum net photosynthetic rate
41.08
<0.001
315.7
<0.001
7.305
0.007
0.023
0.879
9.143
<0.001
1.385
0.197
1.353
0.212
蒸腾速率 Transpiration rate
4.878
<0.001
4.979
0.027
2.226
0.137
1.764
0.186
3.817
<0.001
2.805
0.004
0.881
0.543
比叶面积 Specific leaf area
3.781
<0.001
34.3
<0.001
2.916
0.089
11.62
0.001
1.231
0.278
2.207
0.023
1.843
0.063
相对生长速率 Relative growth rate
2.829
0.006
1 765
<0.001
212.8
<0.001
115.1
<0.001
6.55
<0.001
3.071
0.003
4.847
<0.001
分蘖增长数 The number of tillers increased
1.842
0.073
4.488
0.037
30.15
<0.001
24.72
<0.001
1.061
0.401
1.5
0.162
0.848
0.574
地上生物量 Aboveground biomass
1.454
0.167
0.007
0.933
69.06
<0.001
28.66
0.001
1.116
0.353
1.848
0.062
0.635
0.767
地下生物量 Belowground biomass
2.249
0.021
1.221
0.27
21.93
<0.001
11.07
0.001
0.527
0.854
1.644
0.105
0.664
0.741
叶总酚浓度 Total phenolic concentration of leaf
1.624
0.111
30.62
<0.001
0.002
0.962
48.21
<0.001
1.978
0.044
2.321
0.017
0.957
0.477
根非结构性碳水化合物总量 Total non-structural carbohydrates content of root
1.480
0.157
11.09
0.001
47.13
<0.001
16.21
<0.001
1.938
0.049
2.462
0.011
1.76
0.078
根冠比 Root/shoot ratio
0.976
0.461
0.099
0.753
6.005
0.015
0.479
0.49
1.099
0.365
0.855
0.567
1.093
0.369
新窗口打开 显示原图|下载原图ZIP|生成PPT 图1四种处理条件下10个基因型羊草的反应规范。De-Dr-, 对照; De+Dr-, 非干旱刈割; De-Dr+, 非刈割干旱; De+Dr+, 刈割干旱。 -->Fig. 1Reaction norms of traits of 10 Leymus chinensis genotypes under four treatments. De-Dr-, control; De+Dr-, non-arid defoliation; De-Dr+, non-defoliation but drought; De+Dr+, defoliation and drought. -->
2.2 单因素方差分析及广义遗传力
分别对4种处理条件下的性状平均值进行基因型间的差异分析, 结果显示, 8个性状在基因型间存在显著差异(p < 0.05), 而分蘖增长数、地下部生物量和TNC总量在4种环境条件下均未检测到基因型间的差异(p > 0.05)(表2)。在对照(De-Dr-)条件下, 6个性状表现出基因型间的显著差异(p < 0.05), 其中Pnmax和Tr的H2高于0.5; 在刈割(De+Dr-)条件下, 只有3个性状表现出基因型间的显著差异, 其中Pnmax和RGR的遗传力(H2)高于0.5; 而干旱(De-Dr+)条件下, 5个性状表型表现出基因型间的显著差异(p < 0.05), 其中Fv/Fm、Pnmax和RGR的H2高于0.5; 刈割干旱(De+Dr+)处理条件下, 6个性状表现出基因型间的显著差异(p < 0.05), 其中Fv/Fm、Pnmax和RGR的H2高于0.5。叶总酚浓度和根冠比仅在刈割干旱(De+Dr+)条件下表现出基因型间的显著差异, H2分别为0.145和0.202。 Table 2 表2 表2四种处理条件下羊草表型变异的单因素方差分析与广义遗传力(H2)分析 Table 2ANOVA analysis and the broad-sense heritability (H2) estimates for 10 traits of phenotypic variations under four treatments in Leymus chinensis
性状 Trait
对照 Control
刈割 Defoliation
干旱 Drought
刈割干旱 Defoliation and drought
H2
p
H2
p
H2
p
H2
p
光系统II光化学效率 Photochemical efficiency of photosystem II
0.000
<0.001
0.000
<0.001
1.000
<0.001
1.000
<0.001
最大净光合速率 Maximum net photosynthetic rate
0.847
<0.001
0.679
<0.001
0.585
<0.001
0.660
<0.001
蒸腾速率 Transpiration rate
0.538
<0.001
-
0.948
0.380
<0.001
0.168
0.029
比叶面积 Specific leaf area
0.221
0.022
-
0.127
0.171
0.022
-
0.061
相对生长速率 Relative growth rate
0.400
0.002
0.727
<0.001
0.625
0.001
0.515
0.006
分蘖增长数 The number of tillers increased
-
0.314
-
0.286
-
0.410
-
0.056
地上生物量 Aboveground biomass
0.269
0.007
-
0.787
-
0.752
-
0.075
地下生物量 Belowground biomass
-
0.151
-
0.145
-
0.523
-
0.553
叶总酚浓度 Total phenolic concentration of leaf
-
0.170
-
0.201
-
0.616
0.145
0.004
根非结构性碳水化合物总量 Total non-structural carbohydrates content of root
-
0.201
-
0.062
-
0.550
-
0.951
根冠比 Root/shoot ratio
-
0.936
-
0.061
-
0.068
0.202
0.017
-, the broad-sense heritability (H2) was not calculated because the p-value is bigger than 0.05.-, 基因型间差异不显著未进行广义遗传力(H2)计算。 新窗口打开
2.3 判别分析
4种处理的CDA表明, 两个函数就能解释总变异的59%以上, 但每种处理下, 不同基因型羊草的总体表型存在显著差异,并且对前两个函数贡献较大的指标也不完全相同; 另外, 我们还可以发现, 无论是在哪种处理条件下, 同一个基因型的几个重复的点在图中相距很近, 但并不能单独聚在一起, 表明环境和遗传因素均可以对羊草的表型变异产生影响(表3; 图2)。 Table 3 表3 表3四种处理条件下羊草性状的典则判别函数系数 Table 3Function coefficients of canonical discriminant traits of Leymus chinensis for four treatments
性状 Traits
对照 Control
刈割 Defoliation
干旱 Drought
刈割干旱 Defoliation and drought
函数1 Function 1
函数2 Function 2
函数1 Function 1
函数2 Function 2
函数1 Function 1
函数2 Function 2
函数1 Function 1
函数2 Function 2
光系统II光化学效率 Photochemical efficiency of photosystem II
0.891
0.966
-0.510
-0.191
1.585
0.717
-0.193
0.312
最大净光合速率 Maximum net photosynthetic rate
1.285
-0.462
-0.905
0.865
1.231
-1.609
0.783
-0.672
蒸腾速率 Transpiration rate
-0.561
0.125
0.491
-0.213
0.576
-0.827
1.319
0.098
比叶面积 Specific leaf area
-0.692
-0.533
0.542
0.219
1.341
-0.835
-0.042
1.452
相对生长速率 Relative growth rate
-0.588
0.815
0.962
0.953
1.235
0.236
1.101
0.267
分蘖增长数 The number of tillers increased
-0.416
0.347
-0.424
-1.030
-1.365
0.603
-0.079
0.521
地上生物量 Aboveground biomass
1.114
0.571
1.919
2.834
-0.579
3.541
1.323
2.44
地下生物量 Belowground biomass
-1.81
0.104
-1.614
-2.049
1.594
-3.385
0.055
-2.852
叶总酚浓度 Total phenolic concentration of leaf
0.126
0.905
-0.714
-0.021
-0.115
0.486
-1.226
-1.07
根非结构性碳水化合物总量 Total non-structural carbohydrates content of root
0.598
-1.076
-0.010
-0.446
-1.058
0.564
-0.129
-0.498
根冠比 Root/shoot ratio
1.174
0.781
1.201
1.319
-0.141
1.039
0.838
2.452
解释的方差 Explained variance (%)
49.8
31.9
44.5
30.2
59.2
20.4
37.3
22.0
新窗口打开 显示原图|下载原图ZIP|生成PPT 图2四种处理下10个基因型羊草的典则判别函数散点图。A, 对照。B, 刈割。C, 干旱。D, 刈割干旱。 -->Fig. 2Scatterplot of canonical discriminant of 10 genotypes in Leymus chinensis for four treatments. A, Control. B, Non-arid defoliation. C, Non-defoliation but drought. D, Defoliation and drought. -->
3 讨论
自然种群由表型多样的个体组成, 环境和遗传均可以产生表型变异并驱动种群进化(Bolnick et al., 2011)。在本研究中, 所观测指标在很大程度上均受到环境条件的显著影响, 这不仅表现在不同环境处理间, 也表现在同一环境处理条件下相同基因型间(表1; 图1, 图2), 表明表型可塑性在羊草适应环境中具有重要的作用。羊草表现出的较大的可塑性是其能够耐受水分胁迫和放牧等外界环境、在不同生境中广泛分布、大面积存在于欧亚草原的原因。可塑性能够调节自然选择对植物的影响, 为植物面对快速的气候变化提供一个缓冲, 同时又能使其快速适应新环境(Nicotra et al., 2010)。刈割处理提高根部TNC含量(图1J), 即植株将光合产物更多地转移到根部, 为植株地上部分迅速再生提供物质基础, 表现出耐受性的再生; 在干旱及干旱刈割处理下, 植株根冠比增加(图1K), 以增加根部对水源的获取和减少地上部对水分的蒸腾。然而, 与水分充足时的放牧相比, 在干旱时期放牧, 不同基因型羊草对环境的防御性和耐受性都有不同程度的降低, 即干旱降低了羊草对放牧的耐受性(Gao et al., 2008)和防御性, 对种群的维持和功能的发挥产生不利影响。类似报道, 如干旱胁迫导致苹果(Malus domestica)树生长和产生的次级防御化合物浓度的下降(Gutbrodt et al., 2012)。 除了受环境影响外, 多数生长指标受羊草基因型的影响显著, 如Pnmax、SLA、RGR。羊草相对生长速率和光合能力的种群内部差异(表1)说明不同基因型羊草的资源获取能力不同(Yang et al., 2009), 从而导致种群内基因型间生态位的差异, 使得不同基因型羊草能够以类似于物种间生态位互补的方式共存, 维持自然种群中的多样性(Chang & Smith, 2014)。这些结果合理地解释了申俊芳等(2015)研究发现的多基因型组合羊草种群在提高种群生产力以及抗干扰方面的正效应, 并深入分析得出互补效应在其中起主导作用的结果。其他人的研究结果也支持种内遗传多样性能够以与物种多样性相似的方式提高群落生产力并维护其稳定性(Hughes et al., 2009; Kotowska et al., 2010)。 此外, 基因型×刈割, 基因型×干旱, 以及基因型×刈割×干旱对若干性状产生显著影响, 表明环境条件会对植物的适应机制及策略产生影响, 这会进一步影响基因型多样性功能的发挥, 且这种影响具有遗传学基础。早有研究报道, 基因型间的分化更倾向于在“胁迫”条件下而不是在“最佳”条件下表达(Pigliucci et al., 1995)——随着胁迫水平的增加, 性状的遗传力逐渐增加(Aslam et al., 2006); 但也有人发现在干旱条件下性状遗传力会降低(Eid, 2009)。另外, 性状的遗传力的大小也随环境条件变化且没有一致的规律, 但可以发现, 仅在严重胁迫环境条件(刈割干旱处理)下叶总酚浓度和根TNC总量表现出基因型间的显著差异(表1, 表2), 说明基因型的生态适应策略发生了显著分化, 如基因型3/25用于防御的能量增加, 表现为叶总酚浓度显著升高, 而Y24中总酚浓度却非常低, 两者相差5倍左右。不同基因型对环境变化响应的差异是普遍的, 如:不同基因型的Boechera stricta耐旱性差异显著(Haugen et al., 2008); 生长在不同地点的小麦(Triticum aestivum)的26个基因型的酚酸含量有很大差异 (Fernandez-Orozco et al., 2010); 不同基因型番茄(Lycopersicon esculentum)在不同程度盐胁迫条件下, 根的生长, Na、K含量以及相关蛋白组的表达不同(Manaa et al., 2011)。Mallitt等(2010)研究发现, 在不同水分和光照强度变化条件下, 不同基因型的Lepidium bonariense表现不同, 受基因型×水分×光照交互作用影响显著。显著的基因型×环境交互作用表明: 环境变化越复杂, 基因型越不可能以统一的方式回应, 使得基因型多样性的存在能够维持种群在变异的环境中生存(Ackerly et al., 2000), 对提高群落稳定性或可恢复性十分重要。 羊草的表型差异不仅有广泛的可塑性基础, 而且不同基因型羊草多数性状差异及性状可塑性变异模式受遗传控制, 是其分布范围广、耐受性强的重要原因, 也为其应对外界环境条件变化提供了保障。结合建群种基因型多样性可以发挥类似物种水平多样性的功能已有报道(Kotowska et al., 2010; Cook-Patton et al., 2011), 对羊草草原的保护不仅要保护物种多样性, 也要保护羊草基因型多样性, 这对草原生态系统的维持及退化草原的恢复和重建具有重要的指导意义。 The authors have declared that no competing interests exist.
AckerlyDD, DudleySA, SultanSE, SchmittJ, ColemanJS, LinderCR, SandquistDR, GeberMA, EvansAS, DawsonTE (2000). The evolution of plant ecophysiological traits: Recent advances and future directions new research addresses natural selection, genetic constraints, and the adaptive evolution of plant ecophysiological traits. Bioscience, 50, 979-995. [本文引用: 1]
[2]
AslamM, KhanIA, SaleemM, AliZ (2006). Assessment of water stress tolerance in different maize accessions at germination and early growth stage. Pakistan Journal of Botany, 38, 1571-1579. [本文引用: 1]
[3]
BolnickDI, AmarasekareP, AraújoMS, BürgerR, LevineJM, NovakM, RudolfVH, SchreiberSJ, UrbanMC, VasseurDA (2011). Why intraspecific trait variation matters in community ecology. Trends in Ecology & Evolution, 26, 183-192. [本文引用: 1]
[4]
ChangCC, SmithMD (2014). Direct and indirect relationships between genetic diversity of a dominant grass, community diversity and above-ground productivity in tallgrass prairie. Journal of Vegetation Science, 25, 470-480. [本文引用: 1]
[5]
Cook-PattonSC, McArtSH, ParachnowitschAL, ThalerJS, AgrawalAA (2011). A direct comparison of the consequences of plant genotypic and species diversity on communities and ecosystem function. Ecology, 92, 915-923. [本文引用: 2]
[6]
CrutsingerGM, SandersNJ, ClassenAT (2009). Comparing intra-and inter-specific effects on litter decomposition in an old-field ecosystem. Basic and Applied Ecology, 10, 535-543. [本文引用: 1]
[7]
Da SilveiraAJ, Feitosa TelesF, StullJW (1978). A rapid technique for total nonstructural carbohydrate determination of plant tissue. Journal of Agricultural and Food Chemistry, 26, 770-772. [本文引用: 1]
[8]
DraghiJA, WhitlockMC (2012). Phenotypic plasticity facilitates mutational variance, genetic variance, and evolvability along the major axis of environmental variation. Evolution, 66, 2891-2902. [本文引用: 1]
[9]
EidMH (2009). Estimation of heritability and genetic advance of yield traits in wheat (Triticum aestivum L.) under drought condition. International Journal of Genetics and Molecular Biology, 1, 115-120. [本文引用: 1]
[10]
Fernandez-OrozcoR, LiL, HarflettC, ShewryPR, WardJL (2010). Effects of environment and genotype on phenolic acids in wheat in the health grain diversity screen. Journal of Agricultural and Food Chemistry, 58, 9341-9352. [本文引用: 1]
[11]
GaoY, WangD, BaL, BaiY, LiuB (2008). Interactions between herbivory and resource availability on grazing tolerance of Leymus chinensis. Environmental and Experimental Botany, 63, 113-122. [本文引用: 1]
GutbrodtB, DornS, ModyK (2012). Drought stress affects constitutive but not induced herbivore resistance in apple plants. Arthropod-Plant Interactions, 6, 171-179.
[14]
HaugenR, SteffesL, WolfJ, BrownP, MatznerS, SiemensDH (2008). Evolution of drought tolerance and defense: Dependence of tradeoffs on mechanism, environment and defense switching. Oikos, 117, 231-244. [本文引用: 1]
HuBZ, LiuD, HuGF, JiangSJ, ZhangAY (2001). Morphological variation and genetic diversity in Aneurolepidium chinensis. Acta Phytoecologica Sinica, 25, 83-89. (in Chinese with English abstract)[胡宝忠, 刘娣, 胡国富, 姜述君, 张阿英 (2001). 羊草遗传多样性的研究 . 植物生态学报, 25, 83-89.] [本文引用: 1]
[17]
HughesAR, StachowiczJJ, WilliamsSL (2009). Morphological and physiological variation among seagrass (Zostera marina) genotypes. Oecologia, 159, 725-733.
[18]
JohnsonMT, AgrawalAA (2005). Plant genotype and environment interact to shape a diverse arthropod community on evening primrose (Oenothera biennis). Ecology, 86, 874-885. [本文引用: 1]
KanagaMK, RyelRJ, MockKE, PfrenderME (2008). Quantitative-genetic variation in morphological and physiological traits within a quaking aspen (Populus tremuloides) population. Canadian Journal of Forest Research, 38, 1690-1694. [本文引用: 1]
LandeR (2009). Adaptation to an extraordinary environment by evolution of phenotypic plasticity and genetic assimilation. Journal of Evolutionary Biology, 22, 1435-1446. [本文引用: 2]
[23]
Le CorreV, KremerA (2012). The genetic differentiation at quantitative trait loci under local adaptation. Molecular Ecology, 21, 1548-1566. [本文引用: 2]
[24]
LiH, YangYF, LuXS (2004). Quantitative analysis of reproductive tiller characteristics of Leymus chinensis populations on the Songnen Plain of China. Acta Prataculturae Sinica, 13(4), 50-56. (in Chinese with English abstract)[李红, 杨允菲, 卢欣石 (2004). 松嫩平原羊草种群生殖分蘖株的数量特征及其定量分析 . 草业学报, 13(4), 50-56.] [本文引用: 1]
[25]
LiHY, LiJD, XuZG, ZhouJY, ZhangJF (2011). Vegetative reproduction characteristics of Leymus chinensis populations in Tumuji National Nature Reserve, Inner Mongolia. Acta Prataculturae Sinica, 20(5), 19-25. (in Chinese with English abstract)[李海燕, 李建东, 徐振国, 周景英, 张建峰 (2011). 内蒙古图牧吉自然保护区羊草种群营养繁殖特性的比较 . 草业学报 20(5), 19-25.] [本文引用: 1]
[26]
LieftingM, HoffmannAA, EllersJ (2009). Plasticity versus environmental canalization: Population differences in thermal responses along a latitudinal gradient in Drosophila serrata. Evolution, 63, 1954-1963.
[27]
LiuHF, GaoYB, RuanWB, ChenL, LiCL, ZhaoNX, WangD (2004a). Genetic differentiation within and between Leymus chinensis populations from different zones of mid-eastern Inner Mongolia steppe. Acta Ecologica Sinica, 24, 2157-2164. (in Chinese with English abstract)[刘惠芬, 高玉葆, 阮维斌, 陈磊, 李长林, 赵念席, 王丹 (2004a). 内蒙古中东部不同草原地带羊草种群遗传分化 . 生态学报, 24, 2157-2164.] [本文引用: 2]
[28]
LiuHF, GaoYB, WangD, RenAZ, RuanWB, ChenL, ZhaoNX (2004b). Genetic differentiation in eight populations of Leymus chinensis in Inner Mongolia steppe. Acta Ecologica Sinica, 24, 423-431. (in Chinese with English abstract)[刘惠芬, 高玉葆, 王丹, 任安芝, 阮维斌, 陈磊, 赵念席 (2004b). 内蒙古典型草原羊草种群遗传分化的RAPD分析 . 生态学报, 24, 423-431.] [本文引用: 1]
[29]
MalinowskiDP, AlloushGA, BeleskyDP (1998). Evidence for chemical changes on the root surface of tall fescue in response to infection with the fungal endophyte Neotyphodium coenophialum. Plant and Soil, 205, 1-12. [本文引用: 1]
[30]
MallittKL, BonserSP, HuntJ (2010). The plasticity of phenotypic integration in response to light and water availability in the pepper grass, Lepidium bonariense. Evolutionary Ecology, 24, 1321-1337. [本文引用: 1]
[31]
ManaaA, AhmedHB, ValotB, BouchetJP, Aschi-SmitiS, CausseM, FaurobertM (2011). Salt and genotype impact on plant physiology and root proteome variations in tomato. Journal of Experimental Botany, 17, 1-17.
[32]
MinerBG, SultanSE, MorganSG, PadillaDK, RelyeaRA (2005). Ecological consequences of phenotypic plasticity. Trends in Ecology & Evolution, 20, 685-692.
[33]
NicotraAB, AtkinOK, BonserSP, DavidsonAM, FinneganE, MathesiusU, PootP, PuruggananMD, RichardsC, ValladaresF (2010). Plant phenotypic plasticity in a changing climate. Trends in Plant Science, 15, 684-692.
[34]
PigliucciM, SchlichtingC, WhittonJ (1995). Reaction norms of Arabidopsis. II. Response to stress and unordered environmental variation. Functional Ecology, 9, 537-547.
[35]
RichardsJH (1993). Physiology of Plants Recovering from Defoliation. SIR Publishing, Wellington, New Zealand. 85-94. [本文引用: 2]
[36]
SeliskarDM, GallagherJL, BurdickDM, MutzLA (2002). The regulation of ecosystem functions by ecotypic variation in the dominant plant: A Spartina alterniflora salt-marsh case study. Journal of Ecology, 90, 1-11. [本文引用: 1]
[37]
ShenJF, RenHQ, XinXJ, XuB, GaoYB, ZhaoNX (2015). Leymus chinensis genotypic diversity increases the response of population to disturbance. Acta Ecologica Sinica, 35, 7682-7689. (in Chinese with English abstract)[申俊芳, 任慧琴, 辛晓静, 徐冰, 高玉葆, 赵念席 (2015). 羊草基因型多样性能增强种群对干扰的响应 . 生态学报, 35, 7682-7689.] [本文引用: 1]
[38]
StevensMT, WallerDM, LindrothRL (2007). Resistance and tolerance in Populus tremuloides: Genetic variation, costs, and environmental dependency. Evolutionary Ecology, 21, 829-847. [本文引用: 1]
[39]
StrasburgJL, ShermanNA, WrightKM, MoyleLC, WillisJH, RiesebergLH (2012). What can patterns of differentiation across plant genomes tell us about adaptation and speciation? Philosophical Transactions of the Royal Society of London B: Biological Sciences, 367, 364-373. [本文引用: 1]
SultanSE (2000). Phenotypic plasticity for plant development, function and life history. Trends in Plant Science, 5, 537-542.
[42]
TokerC (2004). Estimates of broad-sense heritability for seed yield and yield criteria in faba bean (Vicia faba L.). Hereditas, 140, 222-225. [本文引用: 1]
[43]
WangD (2004). The Variance and Differentiation of Leymus chinensis in Mid-Eastern Inner Mongolia Grassland. Master degree dissertation, Nankai University, Tianjin. 35-45. (in Chinese with English abstract)[王丹 (2004). 内蒙古中东部草原羊草种内变异与分化研究 . 硕士学位论文, 南开大学, 天津. 35-45.]
[44]
WangR, GaoQ (2001). Photosynthesis, transpiration, and water use efficiency in two divergent Leymus chinensis populations from Northeast China. Photosynthetica, 39, 123-126. [本文引用: 1]
[45]
YangY, FangJ, JiC, HanW (2009). Above- and belowground biomass allocation in Tibetan grasslands. Journal of Vegetation Science, 20, 177-184. [本文引用: 1]
The evolution of plant ecophysiological traits: Recent advances and future directions new research addresses natural selection, genetic constraints, and the adaptive evolution of plant ecophysiological traits. 1 2000
... 此外, 基因型×刈割, 基因型×干旱, 以及基因型×刈割×干旱对若干性状产生显著影响, 表明环境条件会对植物的适应机制及策略产生影响, 这会进一步影响基因型多样性功能的发挥, 且这种影响具有遗传学基础.早有研究报道, 基因型间的分化更倾向于在“胁迫”条件下而不是在“最佳”条件下表达(Pigliucci et al., 1995)——随着胁迫水平的增加, 性状的遗传力逐渐增加(Aslam et al., 2006); 但也有人发现在干旱条件下性状遗传力会降低(Eid, 2009).另外, 性状的遗传力的大小也随环境条件变化且没有一致的规律, 但可以发现, 仅在严重胁迫环境条件(刈割干旱处理)下叶总酚浓度和根TNC总量表现出基因型间的显著差异(表1, 表2), 说明基因型的生态适应策略发生了显著分化, 如基因型3/25用于防御的能量增加, 表现为叶总酚浓度显著升高, 而Y24中总酚浓度却非常低, 两者相差5倍左右.不同基因型对环境变化响应的差异是普遍的, 如:不同基因型的Boechera stricta耐旱性差异显著(Haugen et al., 2008); 生长在不同地点的小麦(Triticum aestivum)的26个基因型的酚酸含量有很大差异 (Fernandez-Orozco et al., 2010); 不同基因型番茄(Lycopersicon esculentum)在不同程度盐胁迫条件下, 根的生长, Na、K含量以及相关蛋白组的表达不同(Manaa et al., 2011).Mallitt等(2010)研究发现, 在不同水分和光照强度变化条件下, 不同基因型的Lepidium bonariense表现不同, 受基因型×水分×光照交互作用影响显著.显著的基因型×环境交互作用表明: 环境变化越复杂, 基因型越不可能以统一的方式回应, 使得基因型多样性的存在能够维持种群在变异的环境中生存(Ackerly et al., 2000), 对提高群落稳定性或可恢复性十分重要. ...
Assessment of water stress tolerance in different maize accessions at germination and early growth stage. 1 2006
... 此外, 基因型×刈割, 基因型×干旱, 以及基因型×刈割×干旱对若干性状产生显著影响, 表明环境条件会对植物的适应机制及策略产生影响, 这会进一步影响基因型多样性功能的发挥, 且这种影响具有遗传学基础.早有研究报道, 基因型间的分化更倾向于在“胁迫”条件下而不是在“最佳”条件下表达(Pigliucci et al., 1995)——随着胁迫水平的增加, 性状的遗传力逐渐增加(Aslam et al., 2006); 但也有人发现在干旱条件下性状遗传力会降低(Eid, 2009).另外, 性状的遗传力的大小也随环境条件变化且没有一致的规律, 但可以发现, 仅在严重胁迫环境条件(刈割干旱处理)下叶总酚浓度和根TNC总量表现出基因型间的显著差异(表1, 表2), 说明基因型的生态适应策略发生了显著分化, 如基因型3/25用于防御的能量增加, 表现为叶总酚浓度显著升高, 而Y24中总酚浓度却非常低, 两者相差5倍左右.不同基因型对环境变化响应的差异是普遍的, 如:不同基因型的Boechera stricta耐旱性差异显著(Haugen et al., 2008); 生长在不同地点的小麦(Triticum aestivum)的26个基因型的酚酸含量有很大差异 (Fernandez-Orozco et al., 2010); 不同基因型番茄(Lycopersicon esculentum)在不同程度盐胁迫条件下, 根的生长, Na、K含量以及相关蛋白组的表达不同(Manaa et al., 2011).Mallitt等(2010)研究发现, 在不同水分和光照强度变化条件下, 不同基因型的Lepidium bonariense表现不同, 受基因型×水分×光照交互作用影响显著.显著的基因型×环境交互作用表明: 环境变化越复杂, 基因型越不可能以统一的方式回应, 使得基因型多样性的存在能够维持种群在变异的环境中生存(Ackerly et al., 2000), 对提高群落稳定性或可恢复性十分重要. ...
Why intraspecific trait variation matters in community ecology. 1 2011
... 自然种群由表型多样的个体组成, 环境和遗传均可以产生表型变异并驱动种群进化(Bolnick et al., 2011).在本研究中, 所观测指标在很大程度上均受到环境条件的显著影响, 这不仅表现在不同环境处理间, 也表现在同一环境处理条件下相同基因型间(表1; 图1, 图2), 表明表型可塑性在羊草适应环境中具有重要的作用.羊草表现出的较大的可塑性是其能够耐受水分胁迫和放牧等外界环境、在不同生境中广泛分布、大面积存在于欧亚草原的原因.可塑性能够调节自然选择对植物的影响, 为植物面对快速的气候变化提供一个缓冲, 同时又能使其快速适应新环境(Nicotra et al., 2010).刈割处理提高根部TNC含量(图1J), 即植株将光合产物更多地转移到根部, 为植株地上部分迅速再生提供物质基础, 表现出耐受性的再生; 在干旱及干旱刈割处理下, 植株根冠比增加(图1K), 以增加根部对水源的获取和减少地上部对水分的蒸腾.然而, 与水分充足时的放牧相比, 在干旱时期放牧, 不同基因型羊草对环境的防御性和耐受性都有不同程度的降低, 即干旱降低了羊草对放牧的耐受性(Gao et al., 2008)和防御性, 对种群的维持和功能的发挥产生不利影响.类似报道, 如干旱胁迫导致苹果(Malus domestica)树生长和产生的次级防御化合物浓度的下降(Gutbrodt et al., 2012). ...
Direct and indirect relationships between genetic diversity of a dominant grass, community diversity and above-ground productivity in tallgrass prairie. 1 2014
... 除了受环境影响外, 多数生长指标受羊草基因型的影响显著, 如Pnmax、SLA、RGR.羊草相对生长速率和光合能力的种群内部差异(表1)说明不同基因型羊草的资源获取能力不同(Yang et al., 2009), 从而导致种群内基因型间生态位的差异, 使得不同基因型羊草能够以类似于物种间生态位互补的方式共存, 维持自然种群中的多样性(Chang & Smith, 2014).这些结果合理地解释了申俊芳等(2015)研究发现的多基因型组合羊草种群在提高种群生产力以及抗干扰方面的正效应, 并深入分析得出互补效应在其中起主导作用的结果.其他人的研究结果也支持种内遗传多样性能够以与物种多样性相似的方式提高群落生产力并维护其稳定性(Hughes et al., 2009; Kotowska et al., 2010). ...
A direct comparison of the consequences of plant genotypic and species diversity on communities and ecosystem function. 2 2011
... 随着种群遗传学研究的不断深入, 人们发现种内基因型间的性状变异不仅对种群的适应性进化具有重要作用, 而且能发挥物种水平的作用, 对群落乃至生态系统进程也具有重要影响(Crutsinger et al., 2009; Cook-Patton et al., 2011).如不同基因型的大叶藻(Zostera marina)不仅在生物量方面存在差异, 而且在铵态氮和硝态氮的利用方面也存在差异, 不同基因型表现出养分利用的分化(Hughes et al., ...
... 羊草的表型差异不仅有广泛的可塑性基础, 而且不同基因型羊草多数性状差异及性状可塑性变异模式受遗传控制, 是其分布范围广、耐受性强的重要原因, 也为其应对外界环境条件变化提供了保障.结合建群种基因型多样性可以发挥类似物种水平多样性的功能已有报道(Kotowska et al., 2010; Cook-Patton et al., 2011), 对羊草草原的保护不仅要保护物种多样性, 也要保护羊草基因型多样性, 这对草原生态系统的维持及退化草原的恢复和重建具有重要的指导意义. ...
Comparing intra-and inter-specific effects on litter decomposition in an old-field ecosystem. 1 2009
... 随着种群遗传学研究的不断深入, 人们发现种内基因型间的性状变异不仅对种群的适应性进化具有重要作用, 而且能发挥物种水平的作用, 对群落乃至生态系统进程也具有重要影响(Crutsinger et al., 2009; Cook-Patton et al., 2011).如不同基因型的大叶藻(Zostera marina)不仅在生物量方面存在差异, 而且在铵态氮和硝态氮的利用方面也存在差异, 不同基因型表现出养分利用的分化(Hughes et al., ...
A rapid technique for total nonstructural carbohydrate determination of plant tissue. 1 1978
... 叶总酚浓度测定采用苯酚比色法(Malinowski et al., 1998); TNC总量测定采用3, 5-二硝基水杨酸比色法(Da Silveira et al., 1978), 并根据相应的地下生物量计算TNC总量. ...
Phenotypic plasticity facilitates mutational variance, genetic variance, and evolvability along the major axis of environmental variation. 1 2012
 , 高玉葆
, 高玉葆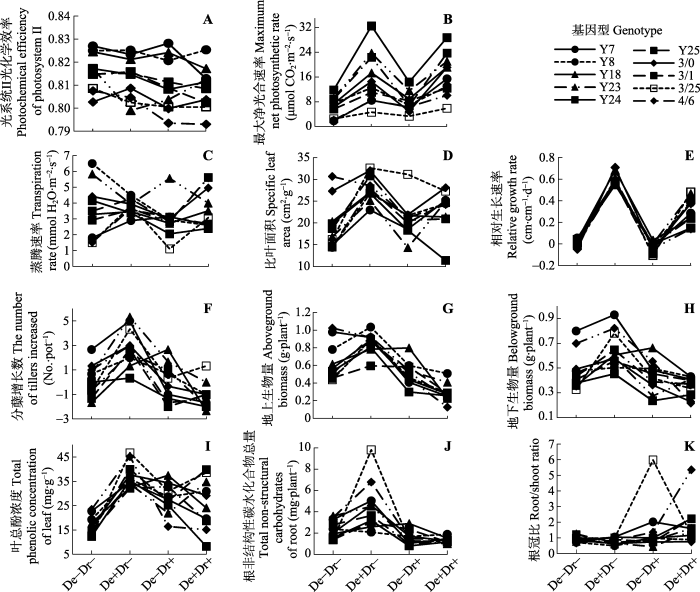 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT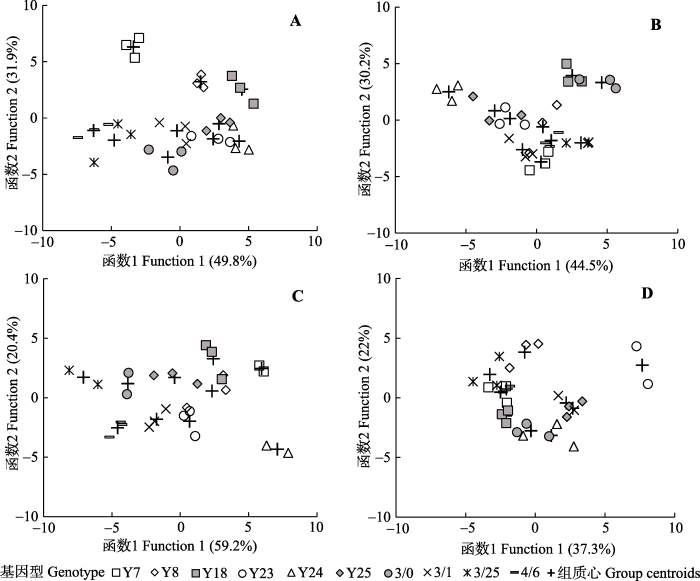 显示原图|下载原图ZIP|生成PPT
显示原图|下载原图ZIP|生成PPT

{kind=link}
{kind=link}
{kind=link}
{kind=link}