Abstract: As pores surrounded by two guard cells on the surface of plants, stomata act as important pathways exchanging water and gas between plants and the external atmosphere. Stomata can also adapt to changes in the environment by affecting the photosynthesis, transpiration and subsequent biological processes in plants. Auxin, the first discovered plant hormone, regulates various developmental processes in plants. Recent studies revealed that auxin is involved in stomatal development by transcription regulation of STOMAGEN via the auxin carrier protein-TIR1/AFB receptor-AUXIN/IAA-ARFs signaling pathway. Eventually, stomatal development is initiated by the STOMAGEN-LRR receptor-like protein kinase ERf-MAPK cascade protein kinase-SPCH signaling pathway. EPF1, EPF2 and the LRR receptor-like protein kinase TMM appear not key components in auxin-regulated stomatal development. Light signaling affects this regulatory process by the E3 ubiquitin ligase COP1 acting upstream of MAPK protein kinases.
Key words:auxin ; stomatal development ; signaling pathway
气孔是由两个保卫细胞围成的特化的植物茎叶等器官表皮结构, 通过控制植物与外界气体交换来影响植物的生长发育等生物学过程。植物激素是植物体内合成的有机小分子代谢物, 较低浓度即可显著调节植物的生长和发育(潘瑞炽, 2008)。目前研究较多的植物激素有生长素(auxin, IAA)、赤霉素(gibberellin, GA)、细胞分裂素(cytokinin, CTK)、脱落酸(abscisic acid, ABA)、乙烯(ethylene, ETH)、油菜素内酯(brassinolide, BR)、茉莉酸(jasmonic acid, JA)、水杨酸(salicylic acid, SA)和独脚金内酯(strigolactone, SL)。随着模式植物拟南芥(Arabidopsis thaliana)气孔发育重要信号分子及相关基因的鉴定, 人们对植物激素在气孔发育中的作用也有了更加深入的认识。Saibo等(2003)的研究发现, 拟南芥下胚轴的生长及气孔发育由GA和其它激素协同调控。ABA能够限制气孔发育过程的启动, 并影响气孔的大小(Matthias et al., 2009; Tanaka et al., 2013)。BR通过磷酸化MAPKKK蛋白激酶(YODA)和SPCH转录因子来调控气孔的发育(Gudesblat et al., 2012; Kim et al., 2012)。SL应答突变体max2的叶片气孔密度显著增加(Ha et al., 2014)。JA和SA均参与气孔运动的调控(Poór and Tari, 2012; Khokon et al., 2015), 但二者参与气孔发育的研究目前报道尚少。生长素是具极性运输特性的植物激素(Ljung et al., 2001; Peer et al., 2011), 被发现与气孔运动有关(Snaith and Mansfield, 1984), 且参与气孔发育的调控(Saibo et al., 2003)。本文主要综述了生长素参与调控气孔发育的信号通路, 及光信号对这一生物学过程的影响。
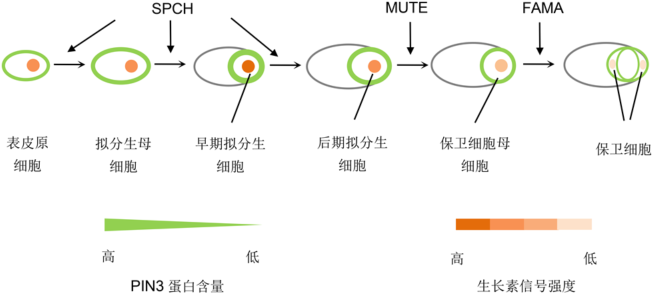
1 生长素载体蛋白和生长素运输对气孔发育的影响在模式植物拟南芥中, 气孔发育由表皮原细胞(pro- todermal cell)开始, 经历拟分生母细胞(mother of meristemoids cells, MMC)、拟分生细胞(meriste- moids cell, MC)、保卫细胞母细胞(guard mother cell, GMC)和保卫细胞(guard cell, GC) 4个主要阶段, 分别受SPCH (SPEECHLESS)、MUTE以及FAMA (Mac- Alister et al., 2007; Pillitteri et al., 2007; Lau et al., 2014) 3个bHLH转录因子(basic Helix Loop Helix)调控(Bergmann and Sack, 2007) (图1)。生长素作为植物生长发育的重要激素之一, 参与气孔发育的调节(Saibo et al., 2003)。生长素在调控过程中存在极 性运输(polar auxin transport, PAT), 此过程由生 长素输入载体蛋白AUX/LAX和输出载体蛋白PIN保持(Paponov et al., 2005; Revalska et al., 2015) (图2)。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-235/img_1.png<b>图1</b> 生长素载体蛋白PIN3及生长素调控气孔发育<br/>图示SPCH、MUTE和FAMA的相应调控位置, 气孔发育过程伴随着生长素输出蛋白PIN3含量以及生长素活性的变化。<br/><b>Figure 1</b> Auxin carrier protein PIN3 and the regulation of auxin on stomatal development<br/>Diagram shows relevant regulatory positions of SPCH, MUTE and FAMA and stomatal development following the content changes of PIN3 and activity changes of auxin. Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-235/img_1.png<b>图1</b> 生长素载体蛋白PIN3及生长素调控气孔发育<br/>图示SPCH、MUTE和FAMA的相应调控位置, 气孔发育过程伴随着生长素输出蛋白PIN3含量以及生长素活性的变化。<br/><b>Figure 1</b> Auxin carrier protein PIN3 and the regulation of auxin on stomatal development<br/>Diagram shows relevant regulatory positions of SPCH, MUTE and FAMA and stomatal development following the content changes of PIN3 and activity changes of auxin.
图1 生长素载体蛋白PIN3及生长素调控气孔发育 图示SPCH、MUTE和FAMA的相应调控位置, 气孔发育过程伴随着生长素输出蛋白PIN3含量以及生长素活性的变化。 Figure 1 Auxin carrier protein PIN3 and the regulation of auxin on stomatal development Diagram shows relevant regulatory positions of SPCH, MUTE and FAMA and stomatal development following the content changes of PIN3 and activity changes of auxin.
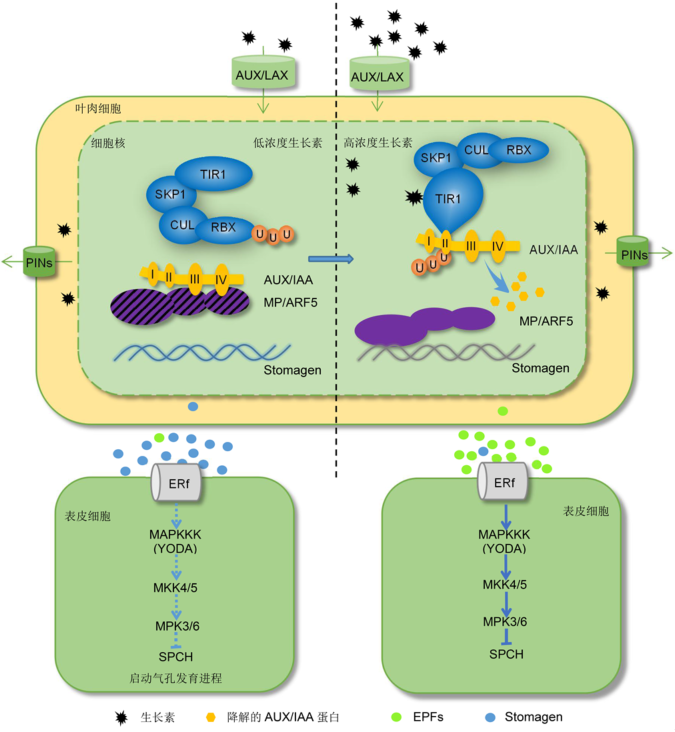
2 生长素调控气孔发育的分子机制生长素输出载体蛋白PIN使生长素在细胞中的含量降低, 细胞趋于由GMC向保卫细胞转化, 此过程会促进气孔的发育; 反之, 如果生长素含量较高, 则会抑制气孔的发育。近年的研究对这一过程的分子机制进行了解析(图2)。当生长素含量增加时, 会结合TIR1/ AFB (TRANSPORT INHIBITOR RESPONSE1/ AUXIN-BINDING F-BOX)受体, 形成以核蛋白TIR1/ AFB为F-box的E3泛素连接酶SCF复合体, TIR1/AFB特异识别生长素响应因子AUX/IAA核蛋白, 通过泛素化将其降解, 使得AUX/IAA与生长素响应转录因子ARFs (auxin response factors)形成的二聚体解离, 释放出ARFs (Gray et al., 2001)。ARFs解除抑制状态后, 与靶基因的启动子结合, 启动或抑制生长素响应基因的转录(Ulmasov et al., 1997; Tiwari et al., 2004; 司马晓娇和郑炳松, 2015) (图2)。 Sugano等(2009)和Zhang等(2014)的研究表明, 在叶肉细胞中, 处于活性状态的ARFs家族成员MP/ARF5会与气孔发育正调控基因STOMAGEN/ EPFL9的启动子结合, 抑制其转录, 使其表达产物减少, 转运到表皮的Stomagen蛋白量也随之减少。通常情况下, 与Stomagen属同一EPF (epidermal patter- ning factor)家族成员的EPF1和EPF2会竞争性结合在表皮细胞膜上的TMM (too many mouths)及ERf (ERECTA)类受体蛋白上(Hara et al., 2007, 2009; Hunt and Gray, 2009), 从而激发MAPKKK (mitogen-activated protein kinase kinase kinase/YODA)到MKK4/5 (MAPK kinases, MAPKKs)再到MPK3/6 (MAPKs) (Widmann et al., 1999; 肖文娟等, 2004; Wang et al., 2007)的MAPK级联反应。MPK3/6磷酸化SPCH转录因子, 抑制其转录活性(Lampard et al., 2008) (图2), 从而抑制气孔的发育过程(图1)。但是在生长素参与的气孔发育调控中, ER、ERL1和ERL2是必需元件, EPF1和EPF2小分子肽及TMM类受体蛋白为非必需元件(Balcerowicz et al., 2014)。Stoma- gen特异结合ERf类受体蛋白(Lee et al., 2015), 其它EPF类因子可能与其竞争结合ERf。当生长素含量降低时, Stomagen的量增加, 从而促进气孔的发育(图2)。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-235/img_2.png<b>图2</b> 生长素信号参与气孔建成的信号通路示意图<br/>生长素极性运输由输入载体蛋白AUX/LAX和输出载体蛋白PIN保持。生长素浓度低时, AUX/IAA蛋白的结构域III、IV与ARF转录因子结合, 促进<i>STOMAGEN</i>基因正常表达。Stomagen由叶肉细胞运转到表皮细胞, 结合并抑制类LRR受体蛋白ERf, 导致MAPKs信号级联反应被抑制, SPCH转录因子启动气孔的发育。生长素浓度高时, SCF复合体的F-box蛋白TIR1识别结构域II, 最终AUX/IAA蛋白被降解, 释放ARF转录因子。ARF转录因子与<i>STOMAGEN</i>启动子结合, <i>STOMAGEN</i>的表达被抑制。与Stomagen竞争的EPFs, 会结合类LRR受体蛋白ERf, 激发MAPKs信号级联反应, 抑制SPCH及其调控的气孔发育。图中箭头表示正调控作用, T表示负调控作用, 虚线表示该过程被抑制。<br/><b>Figure 2</b> Signaling pathway of auxin regulated stomatal establishment<br/>The polar transport of auxin is maintained by auxin influx carrier AUX/LAX and auxin efflux carrier PIN. When auxin concentration is low in mesophyll cells, domain III and IV of AUX/IAA combine with ARF transcription factor, thus promoting <i>STOMAGEN </i>gene expression<i>. </i>Stomagen will move to epidermal cells from mesophyll cell, combine with LRR-like receptor ERf. Then MAPKs signaling cascade will mitigate the repression of SPCH transcription factor which then initiates the stomatal development entry. When auxin level increases in mesophyll cells, TIR1 F-box protein of SCF complex will recognize the domain II of AUX/IAA, leading to the degradation of AUX/IAA. Then ARF will be released to repress the expression of <i>STOMAGEN</i>. EPFs competing with Stomagen will combine LRR-like receptor ERf to phosphorylate kinases of MAPKs signaling cascade. Eventually transcription activity of SPCH and stomatal development are inhibited. Arrows and T represent positive and negative regulations, respectively. The dashed lines represent the inhibited pathway. Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-235/img_2.png<b>图2</b> 生长素信号参与气孔建成的信号通路示意图<br/>生长素极性运输由输入载体蛋白AUX/LAX和输出载体蛋白PIN保持。生长素浓度低时, AUX/IAA蛋白的结构域III、IV与ARF转录因子结合, 促进<i>STOMAGEN</i>基因正常表达。Stomagen由叶肉细胞运转到表皮细胞, 结合并抑制类LRR受体蛋白ERf, 导致MAPKs信号级联反应被抑制, SPCH转录因子启动气孔的发育。生长素浓度高时, SCF复合体的F-box蛋白TIR1识别结构域II, 最终AUX/IAA蛋白被降解, 释放ARF转录因子。ARF转录因子与<i>STOMAGEN</i>启动子结合, <i>STOMAGEN</i>的表达被抑制。与Stomagen竞争的EPFs, 会结合类LRR受体蛋白ERf, 激发MAPKs信号级联反应, 抑制SPCH及其调控的气孔发育。图中箭头表示正调控作用, T表示负调控作用, 虚线表示该过程被抑制。<br/><b>Figure 2</b> Signaling pathway of auxin regulated stomatal establishment<br/>The polar transport of auxin is maintained by auxin influx carrier AUX/LAX and auxin efflux carrier PIN. When auxin concentration is low in mesophyll cells, domain III and IV of AUX/IAA combine with ARF transcription factor, thus promoting <i>STOMAGEN </i>gene expression<i>. </i>Stomagen will move to epidermal cells from mesophyll cell, combine with LRR-like receptor ERf. Then MAPKs signaling cascade will mitigate the repression of SPCH transcription factor which then initiates the stomatal development entry. When auxin level increases in mesophyll cells, TIR1 F-box protein of SCF complex will recognize the domain II of AUX/IAA, leading to the degradation of AUX/IAA. Then ARF will be released to repress the expression of <i>STOMAGEN</i>. EPFs competing with Stomagen will combine LRR-like receptor ERf to phosphorylate kinases of MAPKs signaling cascade. Eventually transcription activity of SPCH and stomatal development are inhibited. Arrows and T represent positive and negative regulations, respectively. The dashed lines represent the inhibited pathway.
图2 生长素信号参与气孔建成的信号通路示意图 生长素极性运输由输入载体蛋白AUX/LAX和输出载体蛋白PIN保持。生长素浓度低时, AUX/IAA蛋白的结构域III、IV与ARF转录因子结合, 促进STOMAGEN基因正常表达。Stomagen由叶肉细胞运转到表皮细胞, 结合并抑制类LRR受体蛋白ERf, 导致MAPKs信号级联反应被抑制, SPCH转录因子启动气孔的发育。生长素浓度高时, SCF复合体的F-box蛋白TIR1识别结构域II, 最终AUX/IAA蛋白被降解, 释放ARF转录因子。ARF转录因子与STOMAGEN启动子结合, STOMAGEN的表达被抑制。与Stomagen竞争的EPFs, 会结合类LRR受体蛋白ERf, 激发MAPKs信号级联反应, 抑制SPCH及其调控的气孔发育。图中箭头表示正调控作用, T表示负调控作用, 虚线表示该过程被抑制。 Figure 2 Signaling pathway of auxin regulated stomatal establishment The polar transport of auxin is maintained by auxin influx carrier AUX/LAX and auxin efflux carrier PIN. When auxin concentration is low in mesophyll cells, domain III and IV of AUX/IAA combine with ARF transcription factor, thus promoting STOMAGEN gene expression. Stomagen will move to epidermal cells from mesophyll cell, combine with LRR-like receptor ERf. Then MAPKs signaling cascade will mitigate the repression of SPCH transcription factor which then initiates the stomatal development entry. When auxin level increases in mesophyll cells, TIR1 F-box protein of SCF complex will recognize the domain II of AUX/IAA, leading to the degradation of AUX/IAA. Then ARF will be released to repress the expression of STOMAGEN. EPFs competing with Stomagen will combine LRR-like receptor ERf to phosphorylate kinases of MAPKs signaling cascade. Eventually transcription activity of SPCH and stomatal development are inhibited. Arrows and T represent positive and negative regulations, respectively. The dashed lines represent the inhibited pathway.
3 生长素调控的气孔发育受光信号影响生长素调控气孔发育受到光信号的影响。在光照条件下, 隐花色素(cryptochrome, CRY)蓝光受体和光敏色素(phytochrome, PHY)红光受体被活化, 二者与光形态建成负调控因子COP1 (constitutively photomorphogenic 1) E3泛素连接酶结合, 活性受到抑制, 进而促进植物的光形态建成(Luo et al., 2014)。有研究表明, 光信号可能通过COP1参与到气孔的发育过程, 其在信号通路中位于YODA等MAPK级联反应元件及SPCH、MUTE和FAMA转录因子的上游, 与TMM类受体蛋白不在同一信号通路上(Kang et al., 2009) (图2)。 生长素在黑暗条件下也会抑制气孔的发育(Balcerowicz et al., 2014), 目前已鉴定出信号通路中的一些信号组分(图2)。有研究表明, 黑暗条件下, 在生长素合成突变体、生长素受体突变体和yoda突变体, 以及经生长素运输抑制剂NPA处理过的幼苗上都观察到了大量的气孔簇, 与光照条件下的表型一致(Balcerowicz et al., 2014)。遗传学分析结果表明, AXR3作用在bHLH转录因子SPCH、 MUTE、 FAMA以及YODA的上游, 与COP1和类受体蛋白TMM处于不同的信号通路; ERf则部分参与AXR3的信号通路, 位于其下游(Balcerowicz et al., 2014), 与光照条件下的信号通路相近。
GudesblatGE, Schneider-PizońJ, BettiC, MayerhoferJ, VanhoutteI, van DongenW, BoerenS, ZhiponovaM, de VriesS, JonakC, RussinovaE (2012). Speechless integrates brassinosteroid and stomata signaling pathways.Nat Cell Biol 14, 548-554. [本文引用: 1]
[8]
HaCV, Leyva-GonzalezMA, OsakabeY, TranUT, NishiyamaR, WatanabeY, TanakaM, SekiM, YamaguchiS, DongNV, Yamaguchi-ShinozakiK, ShinozakiK, Herrera- EstrellaL, TranLS (2014). Positive regulatory role of strigolactone in plant responses to drought and salt stress.Proc Natl Acad Sci USA 111, 851-856. [本文引用: 1]
[9]
HaraK, KajitaR, ToriiKU, BergmannDC, KakimotoT (2007). The secretory peptide geneEPF1 enforces the stomatal one-cell-spacing rule. Genes Dev 21, 1720-1725. [本文引用: 1]
[10]
HaraK, YokooT, KajitaR, OnishiT, YahataS, PetersonKM, ToriiKU, KakimotoT (2009). Epidermal cell density is autoregulated via a secretory peptide, EPIDERMAL PATTERNING FACTOR 2 in Arabidopsis leaves.Plant Cell Physiol 50, 1019-1031. [本文引用: 1]
[11]
HuntL, GrayJE (2009). The signaling peptide EPF2 controls asymmetric cell divisions during stomatal development.Curr Biol 19, 864-869. [本文引用: 1]
[12]
KangCY, LianHL, WangFF, HuangJR, YangHQ (2009). Cryptochromes, Phytochromes, and COP1 regulate light- controlled stomatal development in Arabidopsis.Plant Cell 21, 2624-2641. [本文引用: 1]
[13]
KhokonMA, SalamMA, JammesF, YeW, HossainMA, UrajiM, NakamuraY, MoriIC, KwakJM, MurataY (2015). Two guard cell mitogen-activated protein kinases, MPK9 and MPK12, function in methyl jasmonate-induced stomatal closure in Arabidopsis thaliana. Plant Biol 17, 946-952. [本文引用: 1]
[14]
KimTW, MichniewiczM, BergmannDC, WangZY (2012). Brassinosteroid regulates stomatal development by GSK3- mediated inhibition of a MAPK pathway.Nature 482, 419-422. [本文引用: 1]
[15]
LampardGR, MacalisterCA, BergmannDC (2008). Arabidopsis stomatal initiation is controlled by MAPK- mediated regulation of the bHLH SPEECHLESS.Science 322, 1113-1116. [本文引用: 1]
[16]
LauOS, DaviesKA, ChangJ, AdrianJ, RoweMH, BallengerCE, BergmannDC (2014). Direct roles of SPEECHLESS in the specification of stomatal self-renewing cells.Science 345, 1605-1609. [本文引用: 1]
[17]
LeJ, LiuXG, YangKZ, ChenXL, ZouJJ, WangHZ, WangM, VannesteS, MoritaM, TasakaM, DingZJ, FrimlJ, BeeckmanT, SackF (2014). Auxin transport and activity regulate stomatal patterning and development.Nat Com- mun 5, 3090. [本文引用: 1]
LivanosP, GiannoutsouE, ApostolakosP, GalatisB (2015). Auxin as an inducer of asymmetrical division generating the subsidiary cells in stomatal complexes ofZea mays. Plant Signal Behav 10, e984531. [本文引用: 1]
[20]
LjungK, OstinA, LioussanneL, SandbergG (2001). Developmental regulation of indole-3-acetic acid turnover in Scots pine seedlings.Plant Physiol 125, 464-475. [本文引用: 1]
[21]
LuoQ, LianHL, HeSB, LiL, JiaKP, YangHQ (2014). Cop1 and phyB physically interact with PIL1 to regulate its stability and photomorphogenic development in Arabidopsis.Plant Cell 26, 2441-2456. [本文引用: 1]
[22]
MacAlisterCA, Ohashi-ItoK, BergmannDC (2007). Transcription factor control of asymmetric cell divisions that establish the stomatal lineage.Nature 445, 537-540. [本文引用: 1]
[23]
MatthiasA, Jorg-PeterS, BarbaraE, RobertH, TheoL, HeinzR, AxelH, ErwinG, JorgF (2009). Expression of the Arabidopsis mutant abi1 gene alters abscisic acid sensitivity, stomatal development, and growth morphology in gray poplars. Plant Physiol 151, 2110-2119. [本文引用: 1]
PeerWA, BlakesleeJJ, YangHB, MurphyAS (2011). Seven things we think we know about auxin transport.Mol Plant 4, 487-504. [本文引用: 1]
[26]
PillitteriLJ, SloanDB, BogenschutzNL, ToriiKU (2007). Termination of asymmetric cell division and differentiation of stomata.Nature 445, 501-505. [本文引用: 1]
[27]
PoórP, TariI (2012). Regulation of stomatal movement and photosynthetic activity in guard cells of tomato abaxial epidermal peels by salicylic acid.Funct Plant Biol 39, 1028-1037. [本文引用: 1]
[28]
RevalskaM, VassilevaV, ZechirovG, LantchevaA (2015). Is the auxin influx carrier LAX3 essential for plant growth and development in the model plantsMedicago truncatula, Lotus japonicus and Arabidopsis thaliana? Biotechnol Biotec Eq 29, 786-797. [本文引用: 1]
[29]
SaiboNJM, VriezenWH, BeemsterGTS, StraetenDVD (2003). Growth and stomata development of Arabidopsis, hypocotyls are controlled by gibberellins and modulated by ethylene and auxins.Plant J 33, 989-1000. [本文引用: 2]
[30]
SnaithPJ, MansfieldTA (1984). Studies of the inhibition of stomatal opening by naphth-1-ylacetic acid and abscisic acid.J Exp Bot 35, 1410-1418. [本文引用: 1]
[31]
SuganoSS, ShimadaT, ImaiY, OkawaK, TamaiA, MoriM, Hara-NishimuraI (2009). Stomagen positively regulates stomatal density in Arabidopsis.Nature 463, 241-244. [本文引用: 1]
[32]
TanakaY, NoseT, JikumaruY, KamiyaY (2013). ABA inhibits entry into stomatal-lineage development in Arabi- dopsis leaves.Plant J 74, 448-457. [本文引用: 1]
UlmasovT, MurfettJ, HagenG, GuilfoyleTJ (1997). Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements.Plant Cell 9, 1963-1971. [本文引用: 1]
[35]
WangHC, NgwenyamaN, LiuYD, WalkerJC, ZhangSQ (2007). Stomatal development and patterning are regulated by environmentally responsive mitogen-activated protein kinases in Arabidopsis.Plant Cell 19, 63-73. [本文引用: 1]
[36]
WidmannC, GibsonS, JarpeB, JohnsonGL (1999). Mitogen-activated protein kinase: conservation of a three- kinase module from yeast to human.Phys Rev 79, 143-180. [本文引用: 1]
[37]
ZhangJY, HeSB, LiL, YangHQ (2014). Auxin inhibits stomatal development through monopteros repression of a mobile peptide gene stomagen in mesophyll.Proc Natl Acad Sci USA 111, 3015-3023.
1 2008
... 气孔是由两个保卫细胞围成的特化的植物茎叶等器官表皮结构, 通过控制植物与外界气体交换来影响植物的生长发育等生物学过程.植物激素是植物体内合成的有机小分子代谢物, 较低浓度即可显著调节植物的生长和发育(潘瑞炽, 2008).目前研究较多的植物激素有生长素(auxin, IAA)、赤霉素(gibberellin, GA)、细胞分裂素(cytokinin, CTK)、脱落酸(abscisic acid, ABA)、乙烯(ethylene, ETH)、油菜素内酯(brassinolide, BR)、茉莉酸(jasmonic acid, JA)、水杨酸(salicylic acid, SA)和独脚金内酯(strigolactone, SL).随着模式植物拟南芥(Arabidopsis thaliana)气孔发育重要信号分子及相关基因的鉴定, 人们对植物激素在气孔发育中的作用也有了更加深入的认识.Saibo等(2003)的研究发现, 拟南芥下胚轴的生长及气孔发育由GA和其它激素协同调控.ABA能够限制气孔发育过程的启动, 并影响气孔的大小(Matthias et al., 2009; Tanaka et al., 2013).BR通过磷酸化MAPKKK蛋白激酶(YODA)和SPCH转录因子来调控气孔的发育(Gudesblat et al., 2012; Kim et al., 2012).SL应答突变体max2的叶片气孔密度显著增加(Ha et al., 2014).JA和SA均参与气孔运动的调控(Poór and Tari, 2012; Khokon et al., 2015), 但二者参与气孔发育的研究目前报道尚少.生长素是具极性运输特性的植物激素(Ljung et al., 2001; Peer et al., 2011), 被发现与气孔运动有关(Snaith and Mansfield, 1984), 且参与气孔发育的调控(Saibo et al., 2003).本文主要综述了生长素参与调控气孔发育的信号通路, 及光信号对这一生物学过程的影响. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}