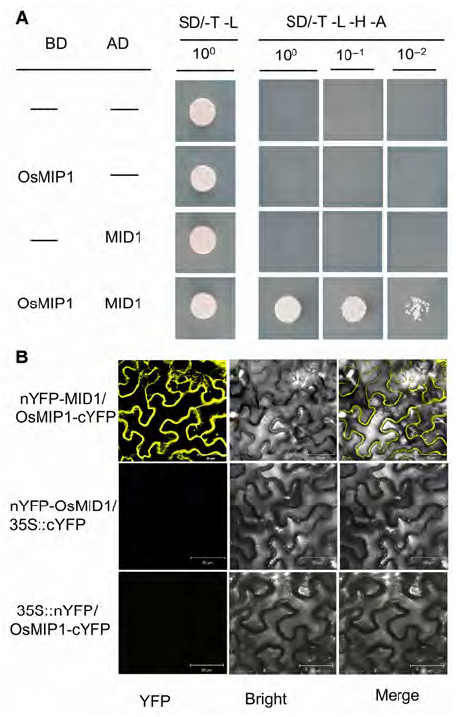
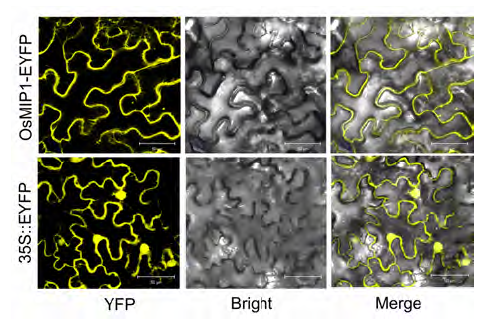
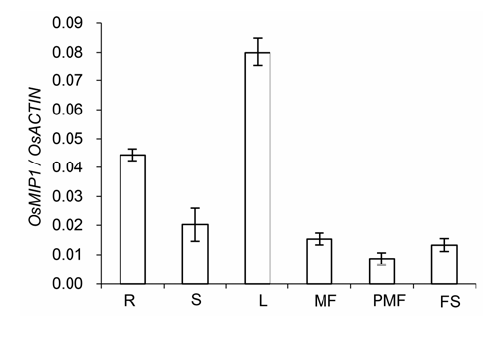
Abstract: The gene MID1 (MYB IMPORTANT FOR DROUGHT RESPONSE1) encodes a putative R-R type MYB transcription factor; is induced by abiotic stresses, especially drought in reproductive stage; and can improve pollen fertility and rice production. To understand the role of MID1 in abiotic stress responses, we used the yeast two-hybrid system to find an interacting protein, OsMIP1 (Oryza sativa MID1 interaction protein 1). The interaction was further confirmed by BiFC (bimolecular fluorescence complementation) analysis in tobacco leaf cells. OsMIP1 encodes a putative transmembrane protein with an ENTH/ANTH/VHS domain. It is expressed in the root, stem, leaf, panicle and endosperm. Under drought stress, its expression is upregulated in leaf and reproductive organs, especially in post-meiotic flowers. OsMIP1 may play a role in response to drought stress during reproductive development. OsMIP1 expression during vegetative development can be induced by other abiotic stress, including NaCl and mannitol, which suggests that OsMIP1 can respond to other abiotic stresses. There is little analysis of the evolution of genes encoding proteins with the ENTH/ ANTH/VHS domain, so we analyzed the molecular evolution of MIP1 homologs in flowering plants. The evolution analysis of the MIP1 family in angiosperms showed that MIP1 homologs can be divided into 6 types, which originated from at least 6 copies of MIP1 homologous genes in the ancestor of extent angiosperms. After gene-duplication and -loss events, MIP1 family members widely distributed in the angiosperms and might have various functions, possibly in stress responses.
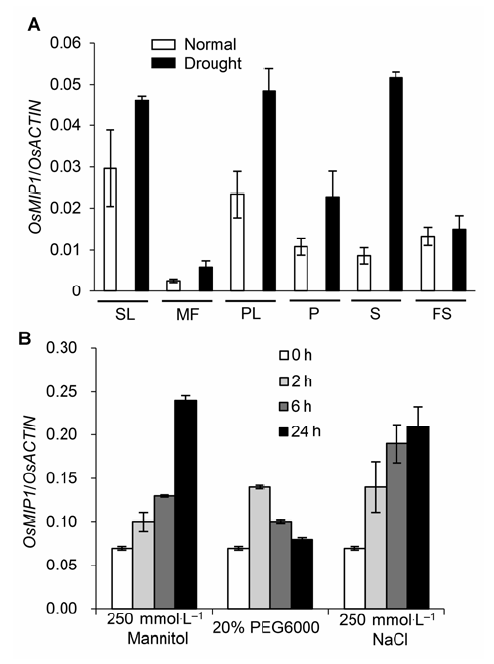
图5 OsMIP1在不同非生物胁迫下的表达模式 (A) OsMIP1在干旱胁迫下不同组织中的表达模式; (B) OsMIP1在200 mmol·L-1甘露醇、20% PEG和200 mmol·L-1 NaCl胁迫下的表达模式。SL: 苗期叶片; MF: 减数分裂时期的小花; PL: 内稃和外稃; P: 雌蕊; S: 雄蕊; DS: 灌浆5天的种子 Figure 5 Expression patterns of OsMIP1 in different abiotic stress (A) Expression patterns of OsMIP1 in different tissues under drought; (B) Expression patterns of OsMIP1 in 200 mmol·L-1 mannitol, 20% PEG and 200 mmol·L-1 NaCl stress. SL: Seedling leaf; MF: Meiotic floret; PL: Palea and lemma; P: Pistil; S: Stamen; FS: 5-day filling seeds
2.5 物种的选择与序列比对为了进一步揭示OsMIP1蛋白亚家族在被子植物中的进化关系, 我们从现在已经完成全基因组测序的植物 中选取具有代表性且进化位置较为明确的物种(表1); 同时, 从实验室完成转录组测序的植物中选取数据较好且进化位置较为明确的物种(表2), 通过同源基因搜索获取OsMIP1家族蛋白序列, 构建OsMIP1蛋白亚家族在被子植物中的进化树, 分析各同源基因的进化关系。在低等植物中也能搜索到OsMIP1蛋白亚家族的成员。 表1 Table 1 表1 表1 基因组数据中选择的物种及信息 Table 1 Species from genome database and their information
Class
Order
Species
Numbers
Eudicots
Cucurbitales
Cucumis sativus
5
Fabales
Glycine max
16
Malvales
Theobroma cacao
6
Myrtales
Eucalyptus robusta
8
Brassicales
Carica papaya
3
Ranunculales
Aquilegia viridiflora
7
Vitales
Vitis vinifera
7
Brassicales
Arabidopsis thaliana
9
Malpighiales
Populus trichocarpa
12
Lamiales
Mimulus luteus
9
Solanales
Solanum tuberosum
1
Monocots
Poales
Zea mays
14
Poales
Brachypodium distachyon
10
Poales
Oryza sativa
10
Poales
Panicum virgatum
16
Poales
Setaria italica
9
Poales
Sorghum bicolor
10
ANA grade
Amborellales
Amborella trichopoda
7
表1 基因组数据中选择的物种及信息 Table 1 Species from genome database and their information
表2 Table 2 表2 表2 转录组数据中选择的物种及信息 Table 2 Species from transcriptome database and their in- formation
Class
Order
Species
Numbers
Eudicots
Apiales
Hedera nepalensis
9
Aquifoliales
Ilex chinensis
8
Asterales
Lactuca sativa
7
Celastrales
Euonymus carnosus
9
Cornales
Swida wilsoniana
9
Dipsacales
Lonicera japonica
7
Ericales
Camellia japonica
11
Fagales
Cyclobalanopsis glauca
6
Cornales
Aucuba japonica
8
Gentianales
Vinca major
8
Proteales
Platanus occidentalis
5
Oxalidales
Oxalis corniculata
7
Proteales
Meliosma arviflora
12
Sapindales
Buxus sinica
6
Monocots
Alismatales
Acorus calamus
6
Asparagales
Asparagus officinalis
4
Asparagales
Phalaenopsis amabilis
6
Liliales
Dioscorea opposita
8
Liliales
Yucca smalliana
6
Pandanales
Pandanus tectorius
5
Zingiberales
Canna indica
8
表2 转录组数据中选择的物种及信息 Table 2 Species from transcriptome database and their in- formation
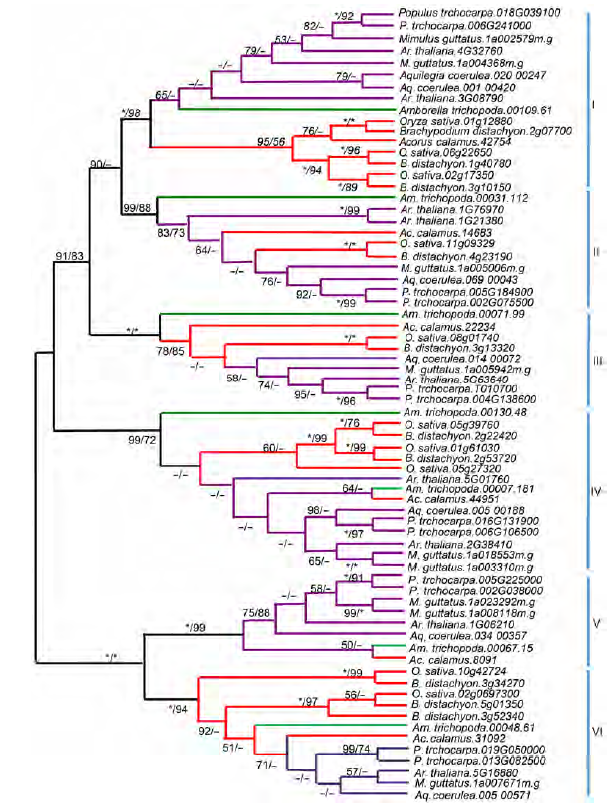
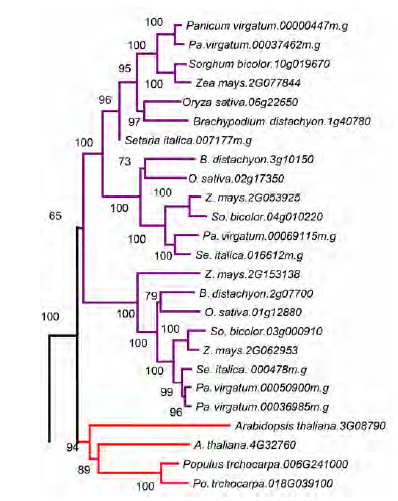
2.6 基因家族系统发育重建将同源序列比对, 并进行局部调整后, 用于构建被子植物中MIP1基因家族的NJ和ML树。根据基因树的拓扑结构, 被子植物的MIP1同源基因可分为6大支(图6, I-VI类), 每大支的NJ树支持率均至少为99%, 而且每一大支中都包含了真双子叶植物、单子叶植物和无油樟等的基因, 说明MIP1蛋白在被子植物的祖先中就已经通过基因重复过程拥有至少6个拷贝。后期的演化过程中, 多数MIP1同源基因在大部分植物中被保留下来, 并且在被子植物的分化过程中, 有些MIP1同源基因又经历了不同的进化模式, 或者拷贝数增加, 或者拷贝丢失。以第I类中的单子叶和真双子叶植物为例, 在经过简化的I类进化树(图7)中(只使用了部分基因组的序列, 包括所有禾本科植物、拟南芥和杨树), 第1个禾本科植物的小分支内的基因大部分属于直系同源基因(除了近期复制而有2个拷贝的柳枝稷外), 而禾本科植物的3个小分支之间属于旁系同源基因。在来源于转录组数据的单子叶其它物种中, 美人蕉和薯蓣等也有2-3个拷贝, 丝兰和芦笋则只有1个拷贝, 推测第I类的MIP1同源基因在单子叶植物祖先进行分化前, 就可能通过基因重复事件拥有3个重复拷贝, 并分别被保留到了各个单子叶植物中。之后, 不同单子叶植物中的拷贝命运并不相同, 有些又经历了重复事件(如柳枝稷); 而有些可能经历了拷贝丢失事件(如丝兰等)。另一种解释则为, 部分物种的转录组数据并未能检测到所有MIP1的同源基因。马铃薯仅有的1个拷贝属于第V类, 可能其在较为近期的进化过程中丢失了绝大部分MIP1大分支的拷贝, 只剩下1个, 如果排除基因组测序可能存在的深度问题, 那么这唯一的拷贝的功能会显得尤为重要。当然, 这一结论尚需分析更多的茄科植物MIP1家族的拷贝数目与进化历史才能得到有效验证。 图6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-43/img_6.png<b>图6</b> 被子植物<i>MIP1</i>基因家族邻接/最大似然(NJ/ML)进化树<br/>被子植物中, <i>MIP1</i>基因可分为6大类(亚家族); 红色: 单子叶植物; 紫色: 真双子叶植物; 绿色: 无油樟。分支上的数字分别表示邻接(neighboring joining, NJ)和最大似然(maximum likelihood, ML)树自展分析的支持率。星号代表自展支持率为100%; 减号表示支持率低于50%。枝长无意义。<br/><b>Figure 6</b> The NJ/ML (neighbor joining/maximum likelihood) tree of <i>MIP1</i> genes family in angiosperms<br/>The <i>MIP1</i> genes can be divided into six types (subfamilies) in angiosperm. Red: Monocotyledons; Purple: Eudicotyledons; Green: <i>Amborella trichopoda</i>. The numbers on each branch are bootstrap support values of NJ/ML (neighbor joining/maximum likelihood) trees. Asterisks stand for bootstrap of 100% and minus stand for bootstrap below 50%. The length of each branch is meaningless. Figure 6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-43/img_6.png<b>图6</b> 被子植物<i>MIP1</i>基因家族邻接/最大似然(NJ/ML)进化树<br/>被子植物中, <i>MIP1</i>基因可分为6大类(亚家族); 红色: 单子叶植物; 紫色: 真双子叶植物; 绿色: 无油樟。分支上的数字分别表示邻接(neighboring joining, NJ)和最大似然(maximum likelihood, ML)树自展分析的支持率。星号代表自展支持率为100%; 减号表示支持率低于50%。枝长无意义。<br/><b>Figure 6</b> The NJ/ML (neighbor joining/maximum likelihood) tree of <i>MIP1</i> genes family in angiosperms<br/>The <i>MIP1</i> genes can be divided into six types (subfamilies) in angiosperm. Red: Monocotyledons; Purple: Eudicotyledons; Green: <i>Amborella trichopoda</i>. The numbers on each branch are bootstrap support values of NJ/ML (neighbor joining/maximum likelihood) trees. Asterisks stand for bootstrap of 100% and minus stand for bootstrap below 50%. The length of each branch is meaningless.
图6 被子植物MIP1基因家族邻接/最大似然(NJ/ML)进化树 被子植物中, MIP1基因可分为6大类(亚家族); 红色: 单子叶植物; 紫色: 真双子叶植物; 绿色: 无油樟。分支上的数字分别表示邻接(neighboring joining, NJ)和最大似然(maximum likelihood, ML)树自展分析的支持率。星号代表自展支持率为100%; 减号表示支持率低于50%。枝长无意义。 Figure 6 The NJ/ML (neighbor joining/maximum likelihood) tree of MIP1 genes family in angiosperms The MIP1 genes can be divided into six types (subfamilies) in angiosperm. Red: Monocotyledons; Purple: Eudicotyledons; Green: Amborella trichopoda. The numbers on each branch are bootstrap support values of NJ/ML (neighbor joining/maximum likelihood) trees. Asterisks stand for bootstrap of 100% and minus stand for bootstrap below 50%. The length of each branch is meaningless.
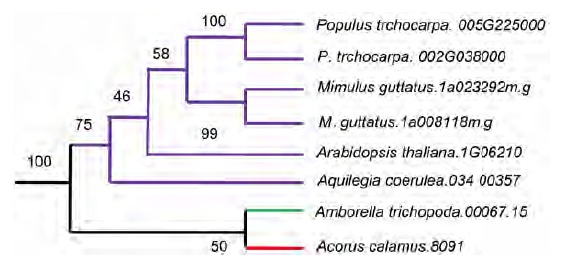
图7https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-43/img_7.png<b>图7</b> 简化的I类<i>MIP1</i>基因进化树<br/>红色: 双子叶植物; 紫色: 单子叶植物<br/><b>Figure 7</b> Simplified gene tree of type I <i>MIP1</i> homologs<br/>Red: Dicotyledons; Purple: Monocotyledons Figure 7https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-1-43/img_7.png<b>图7</b> 简化的I类<i>MIP1</i>基因进化树<br/>红色: 双子叶植物; 紫色: 单子叶植物<br/><b>Figure 7</b> Simplified gene tree of type I <i>MIP1</i> homologs<br/>Red: Dicotyledons; Purple: Monocotyledons
BaisaGA, MayersJR, BednarekSY (2013). Budding and braking news about clathrin-mediated endocytosis.Curr Opin Plant Biol 16, 718-725. [本文引用: 1]
[2]
BarnabasB, JagerK, FeherA (2008). The effect of drought and heat stress on reproductive processes in cereals.Plant Cell Environ 31, 11-38. [本文引用: 1]
[3]
CantrellRP, ReevesTG (2002). The rice genome. The cereal of the world's poor takes center stage.Science 296, 53. [本文引用: 1]
[4]
ChenYH, YangXY, HeK, LiuMH, LiJG, GaoZF, LinZQ, ZhangYF, WangXX, QiuXX, ShenYP, ZhangLi, DengXH, LuoJC, DengXW, ChenZL, GuHY, QuLJ (2006). The MYB transcription factor superfamily of Arabidopsis: expression analysis and phylogenetic comparison with the rice MYB family.Plant Mol Biol 60, 107-124.
[5]
FangYJ, XieKB, HouX, HuHH, XiongLH (2010). Systematic analysis of GT factor family of rice reveals a novel subfamily involved in stress responses.Mol Genet Genomics 283, 157-169. [本文引用: 1]
[6]
GuoCK, YouCJ, WangSS, CuiJ, GeXC, MaH (2016). MID1 plays an important role in response to drought stress during reproductive development. Plant J 88, 280-293. [本文引用: 1]
[7]
HadiartoT, TranLS (2011). Progress studies of drought- responsive genes in rice. Plant Cell Rep 30, 297-310.
[8]
JinY, YangH, WeiZ, MaH, GeX (2013). Rice male deve- lopment under drought stress: phenotypic changes and stage-dependent transcriptomic reprogramming.Mol Plant 6, 1630-1645.
[9]
LeeHK, ChoSK, SonO, XuZY, HwangI, KimWT (2009). Drought stress-induced Rma1H1, a RING membrane- anchor E3 ubiquitin ligase homolog, regulates aquaporin levels via ubiquitination in transgenic Arabidopsis plants.Plant Cell 21, 622-641. [本文引用: 1]
[10]
LiH, YuanZ, Vizcay-BarrenaG, YangC, LiangW, ZongJ, WilsonZA, ZhangDB (2011). PERSISTENT TAPET- AL CELL1 encodes a PHD-finger protein that is required for tapetal cell death and pollen development in rice.Plant Physiol 156, 615-630. [本文引用: 1]
ShinozakiK, Yamaguchi-ShinozakiK, SekiM (2003). Regulatory network of gene expression in the drought and cold stress responses.Curr Opin Plant Biol 6, 410-417. [本文引用: 2]
[13]
SuZ, MaX, GuoH, SukiranNL, GuoB, AssmannSM, MaH (2013). Flower development under drought stress: morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis.Plant Cell 25, 3785-3807. [本文引用: 1]
XiongJ, ZhangL, FuGF, YangYJ, ZhuC, TaoL (2012). Drought-induced proline accumulation is uninvolved with increased nitric oxide, which alleviates drought stress by decreasing transpiration in rice.J Plant Res 125, 155-164. [本文引用: 1]
[16]
ZengL, ZhangQ, SunR, KongH, ZhangN, MaH (2014). Resolving deep angiosperm phylogenetic relationships using conserved nuclear genes.Nat Commun 5, 4956. [本文引用: 1]
[17]
ZhuQH, RammK, ShivakkumarR, DennisES, Upad- hyayaNM (2004). The ANTHER INDEHISCENCE1 gene encoding a single MYB domain protein is involved in anther development in rice.Plant Physiol 135, 1514-1525. [本文引用: 1]
[18]
ZouharJ, SauerM (2014). Helping hands for budding prospects: ENTH/ANTH/VHS accessory proteins in endocytosis, vacuolar transport, and secretion.Plant Cell 26, 4232-4244. [本文引用: 1]
1 2013
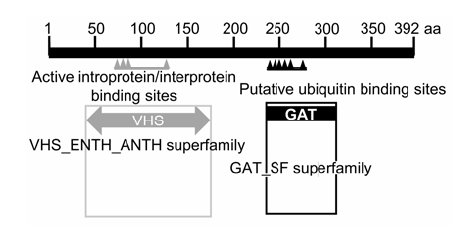
... 非生物胁迫条件下, 细胞膜蛋白对于稳定细胞结构、调节细胞渗透压和细胞内外蛋白物质的运输有重要作用(Lee et al., 2009).细胞膜系统在细胞生命活动中不可或缺, 它不仅使细胞具有一个相对稳定的内环境, 而且有利于细胞与环境之间的物质运输、能量交换和信息传递.细胞内的生物膜将细胞分隔成多个小室(如细胞器), 各个细胞器蛋白的正确运输和定位对于其执行相应的生物学功能至关重要(Baisa et al., 2013).动物中含有ENTH/ANTH/VHS结构域的一类蛋白可以通过肌醇和磷脂类蛋白包裹成一个超螺旋结构, 该结构域蛋白能介导生成网络蛋白包裹的小泡, 从而增加膜的通透性(Marchler-Bauer et al., 2007).目前, 植物中关于ENTH/ANTH/VHS结构域蛋白的研究较少, 其具体的生物学功能尚不清楚(Zouhar and Sauer, 2014). ...
1 2008
... 非生物胁迫, 如干旱、盐、渗透、高温和低温胁迫是植物生长发育的主要限制因素, 它们是导致水稻(Oryza sativa)、大豆(Glycine max)和玉米(Zea mays)等主要作物产量下降的重要原因(Shinozaki et al., 2003; Barnabas et al., 2008).植物体感受外界环境刺激信号后, 通过调控相关基因的转录表达进而合成不同的蛋白, 以实现其生理生化活动的调控, 并最终引发自身复杂的生理反应来抵御逆境(Li et al., 2011). ...
1 2002
... 水稻是世界上重要的粮食作物之一, 同时也是分子生物学研究的模式植物(Cantrell and Reeves, 2002).目前, 对水稻抗旱方面的研究主要集中在营养生长阶段的抗旱能力(Xiong et al., 2012), 关于生殖时期对干旱抵抗能力的分子水平的研究很少.Jin等(2013)观察并分析了水稻生殖发育时期干旱胁迫下花发育的表型及转录组变化, 发现干旱胁迫会引起大量基因的表达改变, 其中包括编码转录因子、酶类和分子伴侣等的基因. ...
2006
1 2010
... 转录因子在非生物胁迫信号转导途径中起重要的调节作用, 它通过与下游基因的启动子结合来调节基因的表达从而响应逆境胁迫(Wang et al., 2011).AP2、bZIP、Zinc-finger、MYB和NAC等转录因子在抗逆途径中起调节干旱响应的作用(Shinozaki et al., 2003; Zhu et al., 2004; Fang et al., 2010).基因MID1 (MYB Important for Drought Response 1)编码R-R类型的MYB-like转录因子, 蛋白定位于细胞核中, 能改善水稻营养期和生殖期的抗旱能力, 当生殖期遭遇干旱, 过表达MID1的转基因水稻能保持花粉育性和水稻产量稳定(Guo et al., 2016). ...
1 2016
... 转录因子在非生物胁迫信号转导途径中起重要的调节作用, 它通过与下游基因的启动子结合来调节基因的表达从而响应逆境胁迫(Wang et al., 2011).AP2、bZIP、Zinc-finger、MYB和NAC等转录因子在抗逆途径中起调节干旱响应的作用(Shinozaki et al., 2003; Zhu et al., 2004; Fang et al., 2010).基因MID1 (MYB Important for Drought Response 1)编码R-R类型的MYB-like转录因子, 蛋白定位于细胞核中, 能改善水稻营养期和生殖期的抗旱能力, 当生殖期遭遇干旱, 过表达MID1的转基因水稻能保持花粉育性和水稻产量稳定(Guo et al., 2016). ...
2011
2013
1 2009
... 非生物胁迫条件下, 细胞膜蛋白对于稳定细胞结构、调节细胞渗透压和细胞内外蛋白物质的运输有重要作用(Lee et al., 2009).细胞膜系统在细胞生命活动中不可或缺, 它不仅使细胞具有一个相对稳定的内环境, 而且有利于细胞与环境之间的物质运输、能量交换和信息传递.细胞内的生物膜将细胞分隔成多个小室(如细胞器), 各个细胞器蛋白的正确运输和定位对于其执行相应的生物学功能至关重要(Baisa et al., 2013).动物中含有ENTH/ANTH/VHS结构域的一类蛋白可以通过肌醇和磷脂类蛋白包裹成一个超螺旋结构, 该结构域蛋白能介导生成网络蛋白包裹的小泡, 从而增加膜的通透性(Marchler-Bauer et al., 2007).目前, 植物中关于ENTH/ANTH/VHS结构域蛋白的研究较少, 其具体的生物学功能尚不清楚(Zouhar and Sauer, 2014). ...
1 2011
... 非生物胁迫, 如干旱、盐、渗透、高温和低温胁迫是植物生长发育的主要限制因素, 它们是导致水稻(Oryza sativa)、大豆(Glycine max)和玉米(Zea mays)等主要作物产量下降的重要原因(Shinozaki et al., 2003; Barnabas et al., 2008).植物体感受外界环境刺激信号后, 通过调控相关基因的转录表达进而合成不同的蛋白, 以实现其生理生化活动的调控, 并最终引发自身复杂的生理反应来抵御逆境(Li et al., 2011). ...
1 2007
... 非生物胁迫条件下, 细胞膜蛋白对于稳定细胞结构、调节细胞渗透压和细胞内外蛋白物质的运输有重要作用(Lee et al., 2009).细胞膜系统在细胞生命活动中不可或缺, 它不仅使细胞具有一个相对稳定的内环境, 而且有利于细胞与环境之间的物质运输、能量交换和信息传递.细胞内的生物膜将细胞分隔成多个小室(如细胞器), 各个细胞器蛋白的正确运输和定位对于其执行相应的生物学功能至关重要(Baisa et al., 2013).动物中含有ENTH/ANTH/VHS结构域的一类蛋白可以通过肌醇和磷脂类蛋白包裹成一个超螺旋结构, 该结构域蛋白能介导生成网络蛋白包裹的小泡, 从而增加膜的通透性(Marchler-Bauer et al., 2007).目前, 植物中关于ENTH/ANTH/VHS结构域蛋白的研究较少, 其具体的生物学功能尚不清楚(Zouhar and Sauer, 2014). ...
2 2003
... 非生物胁迫, 如干旱、盐、渗透、高温和低温胁迫是植物生长发育的主要限制因素, 它们是导致水稻(Oryza sativa)、大豆(Glycine max)和玉米(Zea mays)等主要作物产量下降的重要原因(Shinozaki et al., 2003; Barnabas et al., 2008).植物体感受外界环境刺激信号后, 通过调控相关基因的转录表达进而合成不同的蛋白, 以实现其生理生化活动的调控, 并最终引发自身复杂的生理反应来抵御逆境(Li et al., 2011). ... ... 转录因子在非生物胁迫信号转导途径中起重要的调节作用, 它通过与下游基因的启动子结合来调节基因的表达从而响应逆境胁迫(Wang et al., 2011).AP2、bZIP、Zinc-finger、MYB和NAC等转录因子在抗逆途径中起调节干旱响应的作用(Shinozaki et al., 2003; Zhu et al., 2004; Fang et al., 2010).基因MID1 (MYB Important for Drought Response 1)编码R-R类型的MYB-like转录因子, 蛋白定位于细胞核中, 能改善水稻营养期和生殖期的抗旱能力, 当生殖期遭遇干旱, 过表达MID1的转基因水稻能保持花粉育性和水稻产量稳定(Guo et al., 2016). ...
... 转录因子在非生物胁迫信号转导途径中起重要的调节作用, 它通过与下游基因的启动子结合来调节基因的表达从而响应逆境胁迫(Wang et al., 2011).AP2、bZIP、Zinc-finger、MYB和NAC等转录因子在抗逆途径中起调节干旱响应的作用(Shinozaki et al., 2003; Zhu et al., 2004; Fang et al., 2010).基因MID1 (MYB Important for Drought Response 1)编码R-R类型的MYB-like转录因子, 蛋白定位于细胞核中, 能改善水稻营养期和生殖期的抗旱能力, 当生殖期遭遇干旱, 过表达MID1的转基因水稻能保持花粉育性和水稻产量稳定(Guo et al., 2016). ...
1 2012
... 水稻是世界上重要的粮食作物之一, 同时也是分子生物学研究的模式植物(Cantrell and Reeves, 2002).目前, 对水稻抗旱方面的研究主要集中在营养生长阶段的抗旱能力(Xiong et al., 2012), 关于生殖时期对干旱抵抗能力的分子水平的研究很少.Jin等(2013)观察并分析了水稻生殖发育时期干旱胁迫下花发育的表型及转录组变化, 发现干旱胁迫会引起大量基因的表达改变, 其中包括编码转录因子、酶类和分子伴侣等的基因. ...
... 转录因子在非生物胁迫信号转导途径中起重要的调节作用, 它通过与下游基因的启动子结合来调节基因的表达从而响应逆境胁迫(Wang et al., 2011).AP2、bZIP、Zinc-finger、MYB和NAC等转录因子在抗逆途径中起调节干旱响应的作用(Shinozaki et al., 2003; Zhu et al., 2004; Fang et al., 2010).基因MID1 (MYB Important for Drought Response 1)编码R-R类型的MYB-like转录因子, 蛋白定位于细胞核中, 能改善水稻营养期和生殖期的抗旱能力, 当生殖期遭遇干旱, 过表达MID1的转基因水稻能保持花粉育性和水稻产量稳定(Guo et al., 2016). ...
1 2014
... 非生物胁迫条件下, 细胞膜蛋白对于稳定细胞结构、调节细胞渗透压和细胞内外蛋白物质的运输有重要作用(Lee et al., 2009).细胞膜系统在细胞生命活动中不可或缺, 它不仅使细胞具有一个相对稳定的内环境, 而且有利于细胞与环境之间的物质运输、能量交换和信息传递.细胞内的生物膜将细胞分隔成多个小室(如细胞器), 各个细胞器蛋白的正确运输和定位对于其执行相应的生物学功能至关重要(Baisa et al., 2013).动物中含有ENTH/ANTH/VHS结构域的一类蛋白可以通过肌醇和磷脂类蛋白包裹成一个超螺旋结构, 该结构域蛋白能介导生成网络蛋白包裹的小泡, 从而增加膜的通透性(Marchler-Bauer et al., 2007).目前, 植物中关于ENTH/ANTH/VHS结构域蛋白的研究较少, 其具体的生物学功能尚不清楚(Zouhar and Sauer, 2014). ...
 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}