张金飞1,2, 李霞2,*,, 谢寅峰1 1南京林业大学生物与环境学院, 南京 2100372江苏省农业科学院粮食作物研究所, 江苏省优质水稻工程技术研究中心, 国家水稻改良中心南京分中心, 南京 210014 ZhangJinfei1,2, LiXia2,*,, XieYinfeng1 1College of Biology and Environment, Nanjing Forestry University, Nanjing 210037, China2Nanjing Branch of China National Center for Rice Improvement, Jiangsu High Quality Rice Engineering Technology Research Center, Institute of Food Crops, Jiangsu Academy of Agricultural Sciences, Nanjing 210014, China 引用本文 张金飞, 李霞, 谢寅峰. 植物SnRKs家族在胁迫信号通路中的调节作用. , 2017, 52(3): 346-357
Abstract: SnRKs (sucrose nonfermenting-1-related protein kinases) are key protein kinases in stress responses. In response to biotic stresses, they are involved in reactive oxygen species and salicylic acid-mediated signaling transduction pathways to enhance the plant tolerance to biological stress. In response to abiotic stresses, SnRKs enhance the plant tolerance to drought, salinity and high temperatures by intracellular signaling mediated by abscisic acid (ABA), regulate cellular energy homeostasis and maintain ion balance by ABA-independent signaling. SnRKs are the main regulators in stress signaling in recent research and we give a brief outlook for future study.
1.2 植物SnRK2蛋白激酶亚家族植物SnRK2亚家族成员含有2个典型结构域: N端催化结构域和C末端调节结构域(Halford and Hardie, 1998)。C末端结构域又包括2个子结构域, 即结构域1和结构域2。其中, 结构域1被称为ABA结合框, 因其富含负电荷, 可与PP2C (phosphatase 2C)结合; 结构域2主要是SnRK2的2和3亚组成员的特有结构, 富含天冬氨酸(Yoshida et al., 2006)。对SnRK2亚家族进行全基因组分析, 发现其在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中各有10个成员, 但是染色体定位在二者之间有差异(Wang et al., 2015a)。如在拟南芥所有染色体上都有SnRK2亚家族基因, 而在水稻中其分别位于1、4、7、10和12号染色体, 在小麦(Triticum aestivum)中则定位于1A、1B及1D染色体上, 暗示其生理功能可能因物种的不同而异(Tian et al., 2013; Saha et al., 2014)。
1.3 植物SnRK3蛋白激酶亚家族植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015)。其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014)。PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011)。同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012)。例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物。而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014)。
2.2 植物SnRKs参与水杨酸介导的信号通路水杨酸(salicylic acid, SA)在植物中调控防御基因的表达及宿主细胞死亡, 从而在宿主获得系统性免疫(systemic acquired resistance, SAR)中发挥重要作用, 而NPR1 (nonexpressor of pathogenesis-related gene1)是SA介导信号通路中的主要调节因子(Vlot et al., 2009; Yan and Dong, 2014)。SA介导通路中的信号分子可以激活编码SnRK2.8蛋白激酶的基因, 上调该基因的表达(Lee et al., 2015)。同时, SnRK2.8也可磷酸化NPR1, 使其入核并调控相应的转录因子表达, 增强植株的SAR能力(Lee et al., 2015)。此外, 在拟南芥SnRK3.22基因缺失突变体中, 由于SnRK3.22磷酸化NPR1的缺失, 进入核内的NPR1减少, NPR1的靶基因WRKY38 (sequence-specific DNA-bind- ing proteins38 of the N-terminal sequence WRKY- GQK)和WRKY62的表达下调, 减弱了植株对病虫害的抗性(Xie et al., 2010)。可见, 在植株受到生物胁迫时, SnRKs可通过磷酸化NPR1参与SA介导的信号通路, 从转录水平上迅速启动防御机制, 以增强植株抵抗生物胁迫的能力。
2.3 植物SnRKs参与生物逆境响应在病毒或微生物入侵时, 植物SnRK1可以通过磷酸化帽结合蛋白、翻译起始因子或者病毒蛋白参与生物胁迫响应。拟南芥SnRK1也可以磷酸化双生病毒AL2蛋白, 抑制双生病毒的转录与翻译, 延迟白菜卷叶病毒的感染(Shen et al., 2014)。在小麦中, SnRK1与TaFROG (Triticum aestivum fusarium resistance orphan gene)编码的蛋白质相互作用, 增强小麦抵御禾谷镰刀菌的能力以及对呕吐毒素(deoxynivalenol, DON)的耐性(Perochon et al., 2015)。Kim等(2015) 发现受到真菌和细菌病原体侵染的水稻OSK35-D (水稻SnRK1b的编码基因)激活型突变体能增强基因表达的重编辑, 从而提高对病原体的抗性。此外, 当双生病毒感染拟南芥时, SnRK1磷酸化AtREM4s (Arabidopsis thaliana remorin group 4s), 进而参与26S蛋白酶体途径, 降解甜菜严重曲顶病毒, 增强抗病性(Son et al., 2014)。
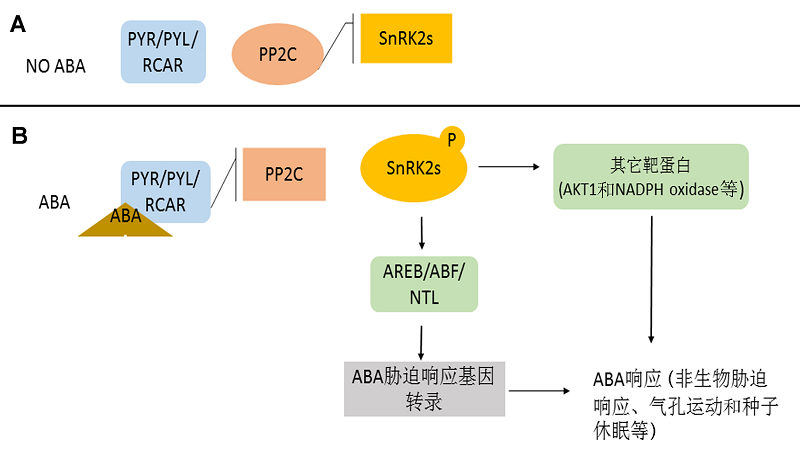
3 植物SnRKs参与非生物胁迫应答3.1 植物SnRKs参与ABA信号介导的非生物胁迫应答2009年, ABA受体的发现为研究SnRKs参与胁迫信号转导通路提供了重要的节点。其中, ABA作为胁迫的响应因子, 可分别与PYR/PYL/RCAR (pyrabactin resistant/PYR-like/regulatory component of ABA) ABA受体结合, 激活的ABA受体抑制包括ABI1 (abs- cisic acid-insenitive 1)、ABI2、HAB1 (homology to ABI1)、HAB2、AHG1 (ABA-hypersensitive germination 1)以及AHG3等在内的9个A组PP2C的活性。失去活性的A组PP2C无法抑制SnRK2s的活性, SnRK2s通过自身磷酸化激活。激活的SnRK2s磷酸化下游的靶标, 从而进一步放大胁迫信号, 增强植株对非生物胁迫的耐性(Yoshida et al., 2006; Hirayama and Umezawa, 2010; Danquah et al., 2014)。 植物SnRKs参与ABA信号介导的非生物胁迫应答大致可分为直接和间接2种途径。直接途径是SnRKs通过自身的磷酸化或被磷酸化, 直接参与ABA信号介导非生物胁迫过程, 增强植株对干旱或盐碱等非生物胁迫的耐性。例如, 在拟南芥中, SnRK- 2.2/SnRK2.3/SnRK2.6是ABA信号转导通路中的重要调节因子, 其中在保卫细胞中响应ABA的ABAR (ab- scisic acid receptor)可直接与SnRK2.6相互作用, 调控ABA信号转导, 进而调节气孔运动(Liang et al., 2015)。而在ABA信号通路中产生的NO, 还可使保卫细胞中的SnRK2.2/SnRK2.3/SnRK2.6的巯基亚硝基化, 抑制SnRK2.2/SnRK2.3/SnRK2.6的活性, 调节由ABA参与调控的种子萌发及幼苗生长(Wang et al., 2015b)。PP2Cs的3个成员(ABI1、AHG1和AHG3)与SnRK2.10强烈相互作用, 介导SnRK2.10参与ABA信号转导, 响应毛果杨(Populus trichocarpa)对干旱和盐碱胁迫的应答(Song et al., 2015)。 另外, 在ABA信号级联通路中, SnRKs可与ABA信号通路中的调节因子相互作用, 间接参与ABA信号转导途径, 调控植株对非生物胁迫的耐受性(Chen et al., 2012)。例如, ABI2可分别与SnRK3.11、SnRK- 3.13和SnRK3.15相互作用, 而ABI1能够与SnRK3.6发生强烈相互作用, 它们互作后都能通过调控相关基因的表达增强植株对ABA信号的敏感性, 从而增强对盐胁迫的耐受性(Ohta et al., 2003)。在拟南芥中, SnRK3.22可以磷酸化ABI5, 间接参与ABA响应基因的表达, 从而调节种子发芽(Zhou et al., 2015b)。处于种子萌发期以及幼苗期的拟南芥, 在它们细胞核中, SnRK2.2、SnRK2.3和SnRK2.6通过磷酸化RAV1 (related to ABI3 (ABA-insensitive3)/VP1 transcription factor, RAV)负调控编码转录因子ABI3、ABI4和ABI5的表达, 参与ABA信号级联通路(Feng et al., 2014)。SnRK2.2、SnRK2.6和SnRK2.3 (SRK2D/E/I)还可激活调节胁迫应答基因表达AREB1 (ABRE (ABA-responsive element)-binding factors1)、AREB2和AREB3, 从而调控植株响应渗透胁迫, 维持植物正常的营养生长(Yoshida et al., 2015)。最新研究发现, ABA信号通路的新成员植物U-Box类E3连接酶(PUB (plant U-box E3 ligases))、PUB12和PUB13可以通过与ABA的信号元件ABI1相互作用, 释放与ABI1结合的SnRK2.6 (也被称为OST1), 促进OST1对下游靶标的磷酸化。研究者基于此提出了依赖ABA作用的新模型(Xuan et al., 2015)。我们按照最新研究进展(Axel et al., 2016), 将SnRK2s参与的ABA途径总结为图1。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-346/img_1.png<b>图1</b> 植物SnRK2s参与的主要ABA途径(改自<xref ref-type="bibr" rid="b3-1674-3466-52-3-346">Axel et al., 2016</xref>)<br/>(A) 无ABA时, ABA受体失活, PP2C抑制SnRK2s的活性; (B) 有ABA时, ABA受体与ABA结合, 抑制PP2C活性, SnRK2s通过自身磷酸化激活, 激活的SnRK2s磷酸化下游转录因子, 磷酸化的转录因子调控ABA胁迫响应基因的表达。同时, SnRK2s也可以磷酸化其它蛋白, 如AKT1及NADPH氧化酶等。这些事件共同引起植株体内ABA响应。<br/><b>Figure 1</b> The plant SnRK2s is involved in the main ABA pathway (motified from Axel et al., 2016)<br/>(A) In the absence of ABA, the ABA receptors are inactivated and PP2C (Protein Phosphatase 2C) proteins inhibit SnRK2s; (B) In the presence of ABA, the ABA receptors PYR/PIL/RCAR proteins bind to ABA and in turn inhibit PP2C activity, allowing the activation of SnRK2s through autophosphorylation. Activated SnRK2s then phosphorylate their downstream transcription factors, which modulate ABA responding gene expression. They can also phosphorylate other proteins, such as AKT1 and NADPH oxidases. Together, these events lead to the establishment of the ABA response. Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-346/img_1.png<b>图1</b> 植物SnRK2s参与的主要ABA途径(改自<xref ref-type="bibr" rid="b3-1674-3466-52-3-346">Axel et al., 2016</xref>)<br/>(A) 无ABA时, ABA受体失活, PP2C抑制SnRK2s的活性; (B) 有ABA时, ABA受体与ABA结合, 抑制PP2C活性, SnRK2s通过自身磷酸化激活, 激活的SnRK2s磷酸化下游转录因子, 磷酸化的转录因子调控ABA胁迫响应基因的表达。同时, SnRK2s也可以磷酸化其它蛋白, 如AKT1及NADPH氧化酶等。这些事件共同引起植株体内ABA响应。<br/><b>Figure 1</b> The plant SnRK2s is involved in the main ABA pathway (motified from Axel et al., 2016)<br/>(A) In the absence of ABA, the ABA receptors are inactivated and PP2C (Protein Phosphatase 2C) proteins inhibit SnRK2s; (B) In the presence of ABA, the ABA receptors PYR/PIL/RCAR proteins bind to ABA and in turn inhibit PP2C activity, allowing the activation of SnRK2s through autophosphorylation. Activated SnRK2s then phosphorylate their downstream transcription factors, which modulate ABA responding gene expression. They can also phosphorylate other proteins, such as AKT1 and NADPH oxidases. Together, these events lead to the establishment of the ABA response.
图1 植物SnRK2s参与的主要ABA途径(改自Axel et al., 2016) (A) 无ABA时, ABA受体失活, PP2C抑制SnRK2s的活性; (B) 有ABA时, ABA受体与ABA结合, 抑制PP2C活性, SnRK2s通过自身磷酸化激活, 激活的SnRK2s磷酸化下游转录因子, 磷酸化的转录因子调控ABA胁迫响应基因的表达。同时, SnRK2s也可以磷酸化其它蛋白, 如AKT1及NADPH氧化酶等。这些事件共同引起植株体内ABA响应。 Figure 1 The plant SnRK2s is involved in the main ABA pathway (motified from Axel et al., 2016) (A) In the absence of ABA, the ABA receptors are inactivated and PP2C (Protein Phosphatase 2C) proteins inhibit SnRK2s; (B) In the presence of ABA, the ABA receptors PYR/PIL/RCAR proteins bind to ABA and in turn inhibit PP2C activity, allowing the activation of SnRK2s through autophosphorylation. Activated SnRK2s then phosphorylate their downstream transcription factors, which modulate ABA responding gene expression. They can also phosphorylate other proteins, such as AKT1 and NADPH oxidases. Together, these events lead to the establishment of the ABA response.
3.2 植物SnRKs参与不依赖ABA信号的非生物胁迫应答3.2.1 植物SnRKs参与调节能量代谢 植物SnRK1作为细胞信号网络中的关键组分, 可以分别与细胞内代表能量状态的不同形式的糖作用, 参与调节能量状态转变的其它激酶活性, 并与调控能量状态的转录因子相互作用, 调节细胞内的能量平衡, 是能量和逆境信号的交汇点(Baena-González and Sheen, 2008)。例如, SnRK1可在黑暗、缺氧和盐碱条件下响应高浓度或低浓度的葡萄糖水平, 参与胁迫下植物能量代谢的调控(Polge and Thomas, 2007)。海藻糖-6-磷酸(T6P)、葡萄糖-6-磷酸(G6P)和葡萄糖-1-磷酸(G1P)可分别抑制SnRK1的活性(Nägele and Weckwerth, 2014)。在T6P/SnRK1信号通路中, 通过磷酸化过程抑制SnRK1活性, 加强细胞代谢活性, 加速生长(Lawlor and Paul, 2014)。绿色器官中, 核糖-5-磷酸在向核糖-1-磷酸转化过程中能量的消耗, 也可以间接抑制SnRK1的活性(Nunes et al., 2013)。SnRK1可与拟南芥中催化生成59-AMP的ADK (ade- nosine kinase)相互作用, 调节细胞中AMP与ATP之间的能量平衡(Mohannath et al., 2014)。此外, 拟南芥中的SnRK1可磷酸化转录因子bZIP63, 调节下游靶基因的表达, 从而调控体内能量平衡(Mair et al., 2015)。当细胞处于低能量状态时, AKIN10 (SnRK1的催化亚基)与拟南芥中的PTL (petal loss)相互作用, 抑制细胞分裂(O'Brien et al., 2015)。同时, AKIN10也可以磷酸化花期调控转录因子IDD8 (indeterminate domain-containing transcription factor8), 参与IDD8介导的代谢过程, 调控开花时间, 使花期延迟(Jeong et al., 2015)。 3.2.2 植物SnRKs参与调节离子平衡 植物SnRK3在参与调控细胞内离子平衡尤其在抵御盐碱胁迫时发挥重要作用。一方面, SnRK3蛋白家族成员参与最重要的胁迫信号转导通路(即SOS通路), SOS3和SCaBP8与SnRK3.11 (又被称为SOS2)相互作用, 激活SnRK3.11, 并在质膜上形成复合体活化Na+/H+泵, 维持细胞内Na+/H+平衡(Lin et al., 2014)。同时, Park等(2013)在研究拟南芥的开花启动时, 发现SnRK3.11可以与光周期和生物钟调控因子GI (gigantea)相互作用。在正常条件下, GI: SnRK3.11复合体可以抑制植物体内主要的Na+/H+泵(SOS1激酶)的活性, 介导对盐碱的适应性(Park et al., 2013)。在盐胁迫下, GI:SnRK3.11复合体崩解且释放SnRK3.11, 激活SOS1, 调控胞内Na+/H+平衡; 与此同时, Sn- RK3.11可以与多种蛋白相互作用, 调控植株不同生长发育时期的耐盐碱性(Park et al., 2013)。SnRK3还可以激活离子通道蛋白参与钾离子运输。例如, Wang等(2016)发现, CBL1/9-SnRK3.23复合体和AtKC1 (Arabidopsis thaliana K+ rectifying channel 1) 可以共同调控AKT1 (Arabidopsis K+ transporter), 平衡AKT1介导的低K+响应中K+的吸收与释放。此外, SnRK3-CBL复合体也可以激活VvK1.2 (Vitis vinifera K+ channel 1.2)。激活的VvK1.2作为电压-门控, 当整流电压低于-100 mv, 且受到外界刺激时, 激活内向K+通道, 介导K+的快速运输(Cuéllar et al., 2013)。 植物SnRK3通过与多种蛋白相互作用, 还可维持细胞内的Mg2+平衡, 防御Mg2+毒害。例如, 拟南芥SnRK3.17/SnRK3.12/SnRK3.23与液泡膜上的CBL 2/3相互作用, 共同螯合液泡中Mg2+ (Tang et al., 2015)。同时, 拟南芥中的SnRK3.17/SnRK3.12/ SnRK3.23 (又称CIPK3/9/23)还可与SnRK2D/E/I相互作用, 通过感受细胞外高浓度Mg2+, 并通过与SnRK2D/E/I相互作用, 传递胁迫信号, 共同维持细胞内Mg2+的动态平衡(Mogami et al., 2015)。 植物SnRK3还可与CBL相互作用, 而CBL对于细胞内外的Ca2+浓度变化敏感。因此, SnRK3与各种底物蛋白相互作用后, 最终也会引起细胞内Ca2+的浓度变化, 参与Ca2+信号通路, 调控胞内Ca2+浓度, 维持细胞的正常生长发育, 或者参与细胞内钙依赖的盐碱干旱胁迫响应。例如, SnRK3.9的过度表达诱导细胞产生严重液泡化表型, 且SnRK3.9基因缺失突变体极性生长受损(Steinhorst et al., 2015)。这是由于液泡内的Ca2+可以直接激活CBL2/3-SnRK3.9复合体, 使得花粉管迅速生长(Steinhorst et al., 2015)。Zhou等(2015a)则发现SnRK3.5过表达可引起极性缺失, 通过尖端膨胀引起胞内Ca2+浓度升高。因此, SnRK3.5也可参与平衡Ca2+流, 避免Ca2+毒害(Zhou, 2015a)。此外, SnRK3也可间接参与平衡胞内Ca2+。如SnRK3.22一方面能够通过磷酸化调控质膜上的质子泵(ATP酶)活性, 引起质膜内外pH值变化; 另一方面, 当其与SCaBP1结合时, 质膜外高pH值可以触发增加胞浆内Ca2+浓度。由于胞浆内质子浓度与盐胁迫相关, 因此, SnRK3.22间接参与Ca2+信号通路, 这也是植物细胞迅速启动盐胁迫应答的一种重要方式(Fuglsang et al., 2007)。 3.2.3 植物SnRKs参与调控非生物胁迫下植株的生长发育 植物SnRKs可以通过磷酸化与生长发育过程中的各种调节因子相互作用, 直接参与调控非生物胁迫下植株的生长发育, 增强植株对胁迫的耐性。烟草(Nico- tiana tabacum) SnRK1可以与作为种子萌发负调控因子的HSPRO (Ortholog of sugar beet Hs1pro-1)相互作用, 调控幼苗的发育(Schuck et al., 2013)。马铃薯(Solanum tuberosum)中细胞程序性死亡抑制因子Adi3 (AvrPto-dependent Pto-interacting protein3)可以磷酸化SnRK1的β亚基, 磷酸化的SnRK1不能与底物特异性相互作用, 抑制SnRK1活性, 调节生长发育过程(Avila et al., 2012)。AKIN10可通过磷酸化IDD8延迟花期(Jeong et al., 2015)。Kim等(2012)则在研究拟南芥响应干旱胁迫时, 发现SnRK2.8磷酸化转录因子NTL6 (NAC (nam/ataf1/2/cuc2) with transmembrane motif1-like6), 延缓植株在响应干旱胁迫时的休眠进程。此外, 在拟南芥响应盐胁迫时, SnRK2.4和SnRK2.10可以与磷脂酸(phosphatidic acid, PA)结合, 参与不依赖ABA的非生物胁迫信号通路, 其中SnRK2.4有磷脂酸结合区域(PA-binding domain, PABD), 此结构域影响拟南芥的根系发育(Wang et al., 2015b)。 综上所述, 我们总结了植物不同SnRKs家族基因参与细胞内不同的生命过程(表1)。 表1 Table 1 表1 表1 拟南芥SnRKs基因家族成员的功能分类 Table 1 Different functions of SnRKs gene family of Arabidopsis
功能
途径
SnRKs亚家族基因名称
基因座位号
基因证号
参考文献
信号
糖
SnRK1.1 (KIN10)
At3G01090
821259
Polge and Thomas, 2007
SnRK1.2 (KIN11)
At3G29160
822566
Polge and Thomas, 2007
SnRK1.3
At5G39440
833940
Mair et al., 2015
钙离子
SnRK3.16 (CIPK1)
At3G17510
821016
D'Angelo et al., 2006
SnRK3.5 (CIPK19)
At5G45810
834621
Zhou et al., 2015a
硝酸盐
SnRK3.13 (CIPK8, PKS11)
At4G24400
828542
Hu et al., 2009
激素 信号
脱落酸
SnRK2.2
At3G50500
824214
Feng et al., 2014
SnRK2.3
AT5G66880
9305254
Wang et al., 2015a
SnRK2.6 (OST1)
At4G33950
829541
Yoshida et al., 2015
SnRK3.14 (CIPK6)
At4G30960
829221
Chen et al., 2013
SnRK3.17 (CIPK3, PKS12)
At2G26980
817240
Kim et al., 2003
SnRK3.22 (CIPK11, PKS5)
At2G30360
817586
Zhou et al., 2015b
生物 胁迫
水杨酸
SnRK2.8
At1G78290
844164
Lee et al., 2015
SnRK3.22 (CIPK11, PKS5)
At2G30360
817586
Xie et al., 2010
活性氧
SnRK3.26 (CIPK26)
At5G21326
832246
Drerup et al., 2013
SnRK3.14 (CIPK6)
At4G30960
829221
Chen et al., 2013
非生物胁迫
非生物胁迫
SnRK3.1 (CIPK15, PKS3)
At5G01810
830556
Kanwar et al., 2014
SnRK3.2 (CIPK2)
At5G07070
830598
SnRK3.6 (CIPK20, PKS18)
At5G45820
834622
SnRK3.7 (CIPK13)
At2G34180
817979
SnRK3.8 (CIPK10)
At5G58380
835951
寒冷
SnRK3.10 (CIPK7, PKS7)
At3G23000
821874
Huang et al., 2011
SnRK3.17 (CIPK3, PKS12)
At2G26980
817240
Kim et al., 2003
缺氧
SnRK3.25 (CIPK25)
At5G25110
832582
Meena et al., 2015
渗透胁迫
SnRK3.18 (CIPK16, PKS15)
At2G25090
817047
Amarasinghe et al., 2016
SnRK3.4 (CIPK21)
At5G57630
835868
Pandey et al., 2015
SnRK2.7 (SRK2F)
At4G40010
830162
Fujii et al., 2011
SnRK2.9
At2G23030
816833
Fujii et al., 2011
SnRK2.1 (SRK2G)
At5G08590
830760
Fujii et al., 2011
盐碱
SnRK3.9 (CIPK12)
At4G18700
827604
Steinhorst et al., 2015
SnRK3.11 (SOS2, CIPK24)
At5G35410
833502
Park et al., 2013
SnRK3.12 (CIPK9, PKS6)
At1G01140
839349
Mogami et al., 2015
SnRK3.15 (PKS24, CIPK14)
At5G01820
831765
Lin et al., 2014
SnRK3.22 (CIPK11, PKS5)
At2G30360
817586
Zhou et al., 2015b
SnRK3.18 (CIPK16, PKS15)
At2G25090
817047
Amarasinghe et al., 2016
SnRK3.23 (CIPK23, PKS17)
At1G30270
839907
Wang et al., 2016
生长 发育
根生长
SnRK2.10
At1G60940
842385
McLoughlin et al., 2012
SnRK2.4 (ASK1, SRK2A)
At1G10940
837637
McLoughlin et al., 2012
SnRK3.25 (CIPK25)
At5G25110
832582
Meena et al., 2015
种子发芽
SnRK3.22 (CIPK11, PKS5)
At2G30360
817586
Zhou et al., 2015b
幼苗生长
SnRK1.1 (KIN10)
At3G01090
821259
O'Brien et al., 2015
开花
SnRK1.1 (KIN10)
At3G01090
821259
Jeong et al., 2015
数据来源包括基因库http://www.ncbi.nlm.nih.gov/。Sources include GenBank http://www.ncbi.nlm.nih.gov/.
表1 拟南芥SnRKs基因家族成员的功能分类 Table 1 Different functions of SnRKs gene family of Arabidopsis
4 植物SnRKs参与不同胁迫信号通路途径间的可能联系4.1 植物体内SnRKs的平衡调节机制当植物受到胁迫时, SnRKs迅速感受刺激并产生应激反应。此时, SnRK1主要通过调整代谢来改变细胞内的能量胁迫状态(Halford and Hey, 2009)。但是, 当应激反应强度过大且时间持久时, 活化的SnRK1则触发自身的小型泛素化修饰(small ubiquitin-like modifier, SUMO)过程, 通过蛋白酶体途径进行降解, 进而抑制细胞的正常生理活动(Nägele and Weckwerth, 2014; Crozet et al., 2016)。此外, SnRK1的降解是严格依赖其自身活性的(Crozet et al., 2016)。例如, 无活性的SnRK1在植物体内是非常稳定的, 但是, 当SnRK1被激活时, 同时启动了SUMO化过程(Crozet et al., 2016)。此外, Rodrigues等(2013)发现, A组PP2C作为ABA介导的胁迫信号通路中的抑制因子, 可以与SnRK1的催化亚基相互作用使其去磷酸化并失活。同时, SnRK1靶基因表达分析进一步证明, 由于ABA抑制PP2C使SnRK1激活, 在能量匮乏时, SnRK1激活下游的靶标(Rodrigues et al., 2013)。可见, SnRK1的失活与激活可能通过自身的严格控制而达到平衡。
AmarasingheS, Watson-HaighNS, GillihamM, RoyS, BaumannU (2016). The evolutionary origin ofCIPK16: a gene involved in enhanced salt tolerance. Mol Phylogenet Evol 100, 135-147.
[2]
AvilaJ, GregoryOG, SuD, DeeterTA, ChenS, Silva- SanchezC, XuS, MartinGB, DevarenneTP (2012). The β-subunit of the SnRK1 complex is phosphorylated by the plant cell death suppressor Adi3.Plant Physiol 159, 1277-1290. [本文引用: 1]
[3]
AxelZ, JeanC, HeribertH (2016). The role of MAPK modu- les and ABA during abiotic stress signaling.Trends Plant Sci 21, 677-685. [本文引用: 2]
[4]
Baena-GonzálezE, SheenJ (2008). Convergent energy and stress signaling.Trends Plant Sci 13, 474-482. [本文引用: 2]
[5]
BatističO, KudlaJ (2009). Plant calcineurin B-like proteins and their interacting protein kinases.Biochim Biophys Acta 1793, 985-992. [本文引用: 1]
[6]
BatističO, WaadtR, SteinhorstL, HeldK, KudlaJ (2010). CBL-mediated targeting of CIPKs facilitates the decoding of calcium signals emanating from distinct cellular stores.Plant J 61, 211-222. [本文引用: 1]
[7]
Chaves-SanjuanA, Sanchez-BarrenaMJ, Gonzalez- RubioJM, MorenoM, RagelP, JimenezM, PardoJM, Martinez-RipollM, QuinteroFJ, AlbertA (2014). Struc- tural basis of the regulatory mechanism of the plant CIPK family of protein kinases controlling ion homeostasis and abiotic stress.Proc Natl Acad Sci USA 111, 4532-4541. [本文引用: 1]
[8]
ChenL, RenF, ZhouL, WangQQ, ZhongH, LiXB (2012). The Brassica napus calcineurin B-Like 1/CBL-interacting protein kinase 6 (CBL1/CIPK6) component is involved in the plant response to abiotic stress and ABA signaling. J Exp Bot 63, 6211-6222. [本文引用: 1]
[9]
ChenL, WangQQ, ZhouL, RenF, LiDD, LiXB (2013). Arabidopsis CBL-interacting protein kinase (CIPK6) is involved in plant response to salt/osmotic stress and ABA. Mol Biol Rep 40, 4759-4767.
[10]
CoelloP, HeySJ, HalfordNG (2011). The sucrose non- fermenting-1-related (SnRK) family of protein kinases: potential for manipulation to improve stress tolerance and increase yield.J Exp Bot 62, 883-893. [本文引用: 2]
[11]
ConfrariaA, MartinhoC, EliasA, Rubio-SomozaI, Baena-GonzálezE (2013). miRNAs mediate SnRK1- dependent energy signaling in Arabidopsis.Front Plant Sci 4, 197. [本文引用: 1]
CuéllarT, AzeemF, AndrianteranagnaM, PascaudF, VerdeilJL, SentenacH, ZimmermannS, GaillardI (2013). Potassium transport in developing fleshy fruits: the grapevine inward K+ channel VvK1.2 is activated by CIPK-CBL complexes and induced in ripening berry flesh cells.Plant J 73, 1006-1018. [本文引用: 1]
[14]
D'AngeloC, WeinlS, BatisticO, PandeyGK, CheongYH, SchültkeS (2006). Alternative complex formation of the Ca2+ regulated protein kinase CIPK1 controls abscisic acid dependent and independent stress responses in Arabidopsis.Plant J 48, 857-872.
[15]
DanquahA, de ZelicourtA, ColcombetJ, HirtH (2014). The role of ABA and MAPK signaling pathways in plant abiotic stress responses.Biotechnol Adv 32, 40-52. [本文引用: 1]
[16]
DongXF, CuiN, WangL, ZhaoXC, QuB, LiTL, ZhangGL (2012). The SnRK protein kinase family and the function of SnRK1 protein kinase.Int J Agric Biol 14, 196-200. [本文引用: 1]
[17]
DrerupMM, SchlückingK, HashimotoK, ManishankarP, SteinhorstL, KuchitsuK, KudlaJ (2013). The calcineurin B-like calcium sensors CBL1 and CBL9 together with their interacting protein kinase CIPK26 regulate the Arabidopsis NADPH oxidase RBOHF.Mol Plant 6, 559-569. [本文引用: 1]
[18]
EmanuelleS, HossainMI, MollerIE, PedersenHL, MeeneAM, DoblinMS, KoayA, OakhillJS, ScottJW, WillatsWGT, KempBE (2015). SnRK1 from Arabidopsis thaliana is an atypical AMPK. Plant J 82, 183-192. [本文引用: 6]
[19]
FengCZ, ChenY, WangC, KongYH, WuWH, ChenYF (2014). Arabidopsis RAV1 transcription factor, phosphorylated by SnRK2 kinases, regulates the expression of ABI3, ABI4, and ABI5 during seed germination and early seedling development.Plant J 80, 654-668. [本文引用: 1]
[20]
FuglsangAT, GuoY, CuinTA, QiuQ, SongC, KristiansenKA, BychK, SchulzA, ShabalaS, SchumakerKS (2007). Arabidopsis protein kinase PKS5 inhibits the plasma membrane H+-ATPase by preventing interaction with 14-3-3 protein.Plant Cell 19, 1617-1634. [本文引用: 1]
[21]
FujiiH, VersluesPE, ZhuJK (2011). Arabidopsis decuple mutant reveals the importance of SnRK2 kinases in osmotic stress responses in vivo. Proc Natl Acad Sci USA 108, 1717-1722.
[22]
GhillebertR, SwinnenE, WenJ, VandesteeneL, RamonM, NorgaK, RollandF, WinderickxJ (2011). The AMP- K/SNF1/SnRK1 fuel gauge and energy regulator: structure, function and regulation. FEBS J 278, 3978-3990. [本文引用: 1]
[23]
HalfordNG, HardieDG (1998). SNF1-related protein kinases: global regulators of carbon metabolism in plants.Plant Mol Biol 37, 735-748. [本文引用: 1]
[24]
HalfordNG, HeySJ (2009). Snf1-related protein kinases (SnRKs) act within an intricate network that links metabolic and stress signaling in plants.Biochem J 419, 247-259. [本文引用: 1]
[25]
HalfordNG, HeySJ, JhurreeaD, LaurieS, McKibbinRS, ZhangY, PaulMJ (2004). Highly conserved protein kinases involved in the regulation of carbon and amino acid metabolism.J Exp Bot 55, 35-42. [本文引用: 1]
[26]
HashimotoK, EckertC, AnschützU, ScholzM, HeldK, WaadtR, ReyerA, HipplerM, BeckerD, KudlaJ (2012). Phosphorylation of calcineurin B-like (CBL) calcium sensor proteins by their CBL-interacting protein kinases (CIPKs) is required for full activity of CBL-CIPK complexes toward their target proteins.J Biol Chem 287, 7956-7968. [本文引用: 1]
[27]
HirayamaT, UmezawaT (2010). The PP2C-SnRK2 complex: the central regulator of an abscisic acid signaling pathway.Plant Signal Behave 5, 160-163. [本文引用: 1]
[28]
HuHC, WangYY, TsayYF (2009). AtCIPK8, a CBL-inter- acting protein kinase, regulates the low-affinity phase of the primary nitrate response.Plant J 57, 264-278.
[29]
HuangC, DingS, ZhangH, DuH, AnL (2011). CIPK7 is involved in cold response by interacting with CBL1 inArabidopsis thaliana. Plant Sci 181, 57-64.
[30]
JeongEY, SeoPJ, WooJC, ParkCM (2015). AKIN10 delays flowering by inactivating IDD8 transcription factor through protein phosphorylation in Arabidopsis.BMC Plant Biol 15, 110. [本文引用: 2]
KanwarP, SanyalSK, TokasI, YadavAK, PandeyA, KapoorS, PandeyGK (2014). Comprehensive structural, interaction and expression analysis of CBL and CIPK complement during abiotic stresses and development in rice.Cell Calcium 56, 81-95.
[33]
KimCY, VoKTX, AnG, JeonJS (2015). A rice sucrose non-fermenting-1 related protein kinase 1, OSK35, plays an important role in fungal and bacterial disease resistance.J Korean Soc Appl Bi 58, 669-675.
[34]
KimKN, CheongYH, GrantJJ, PandeyGK, LuanS (2003). CIPK3, a calcium sensor-associated protein kin- ase that regulates abscisic acid and cold signal transduction in Arabidopsis.Plant Cell 15, 411-423.
[35]
KimMJ, ParkMJ, SeoPJ, SongJS, KimHJ, ParkCM (2012). Controlled nuclear import of the transcription factor NTL6 reveals a cytoplasmic role of SnRK2.8 in the drought-stress response.Biochem J 448, 353-363.
[36]
KimuraS, KawarazakiT, NiboriH, MichikawaM, ImaiA, KayaH, KuchitsuK (2013). The CBL-interacting protein kinase CIPK26 is a novel interactor of Arabidopsis NADPH oxidase AtRbohF that negatively modulates its ROS-producing activity in a heterologous expression sys- tem.J Biochem 153, 191-195.
[37]
KleistTJ, SpencleyAL, LuanS (2014). Comparative phylogenomics of the CBL-CIPK calcium-decoding network in the moss Physcomitrella, Arabidopsis, and other green lineages. Front Plant Sci 5, 187-207. [本文引用: 2]
[38]
LanWZ, LeeSC, CheYF, JiangYQ, LuanS (2011). Mechanistic analysis of AKT1 regulation by the CBL- CIPK-PP2CA interactions.Mol Plant 4, 527-536. [本文引用: 1]
[39]
LawlorDW, PaulMJ (2014). Source/sink interactions underpin crop yield: the case for trehalose 6-phosphate/ SnRK1 in improvement of wheat.Front Plant Sci 5, 418-432. [本文引用: 1]
LiangS, LuK, WuZ, JiangSC, YuYT, BiC, ZhangDP (2015). A link between magnesium-chelatase H subunit and sucrose nonfermenting 1 (SNF1)-related protein kinase SnRK2.6/OST1 in Arabidopsis guard cell signaling in response to abscisic acid.J Exp Bot 66, 6355-6369. [本文引用: 1]
[42]
LinH, DuW, YangY, SchumakerKS, GuoY (2014). A calcium-independent activation of the Arabidopsis SOS2- like protein kinase24 by its interacting SOS3-like calcium binding protein1.Plant Physiol 164, 2197-2206. [本文引用: 1]
[43]
MairA, PedrottiL, WurzingerB, AnratherD, SimeunovicA, WeisteC, ValerioC, DietrichK, KirchlerT, NägeleT (2015). SnRK1-triggered switch of bZIP63 dimerization mediates the low-energy response in plants.eLife 4, e05828. [本文引用: 1]
[44]
McLoughlinF, Galvan-AmpudiaCS, JulkowskaMM, CaarlsL, van der DoesD, LaurièreC, MunnikT, HaringMA, TesterinkC (2012). The Snf1-related protein kinases SnRK2.4 and SnRK2.10 are involved in maintenance of root system architecture during salt stress.Plant J 72, 436-449.
[45]
MeenaMK, SanjayG, VikasD, AnsumanR, DebasisC (2015). Expression of chickpea CIPK25 enhances root growth and tolerance to dehydration and salt stress in transgenic tobacco.Front Plant Sci 6, 683.
[46]
MogamiJ, FujitaY, YoshidaT, TsukioriY, NakagamiH, NomuraY, FujiwaraT, NishidaS, YanagisawaS, TakahashiF (2015). Two distinct families of protein kinases are required for plant growth under high external Mg2+ concentrations in Arabidopsis.Plant Physiol 167, 1039-1057. [本文引用: 1]
[47]
MohannathG, JackelJN, LeeYH, BuchmannRC, WangH, PatilV, AdamsAK, BisaroDM (2014). A complex containing SNF1-related kinase (SnRK1) and adenosine kinase in Arabidopsis.PLoS One 9, e87592. [本文引用: 1]
[48]
NägeleT, WeckwerthW (2014). Mathematical modeling reveals that metabolic feedback regulation of SnRK1 and hexokinase is sufficient to control sugar homeostasis from energy depletion to full recovery.Front Plant Sci 5, 365-376. [本文引用: 2]
[49]
NunesC, PrimavesiLF, PatelMK, Martinez-BarajasE, PowersSJ, SagaR, FevereiroPS, DavisBG, PaulMJ (2013). Inhibition of SnRK1 by metabolites: tissuedepen- dent effects and cooperative inhibition by glucose 1- phosphate in combination with trehalose 6-phosphate.Plant Physiol Biochem 63, 89-98. [本文引用: 1]
[50]
O’BrienM, Kaplan-LevyRN, QuonT, SapplPG, SmythDR (2015). PETAL LOSS, a trihelix transcription factor that represses growth in Arabidopsis thaliana, binds the energy-sensing SnRK1 kinase AKIN10. J Exp Bot 66, 2475-2485. [本文引用: 1]
[51]
OhtaM, GuoY, HalfterU, ZhuJK (2003). A novel domain in the protein kinase SOS2 mediates interaction with the protein phosphatase 2C ABI2.Proc Natl Acad Sci USA 100, 11771-11776. [本文引用: 1]
[52]
PandeyGK, KanwarP, SinghA, SteinhorstL, PandeyA, YadavAK, CheongYH (2015). Calcineurin B-like protein-interacting protein kinase CIPK21 regulates osmotic and salt stress responses in Arabidopsis.Plant Physiol 169, 780-792.
[53]
ParkHJ, KimWY, YunDJ (2013). A role for GIGANTEA: keeping the balance between flowering and salinity stress tolerance.Plant Signal Behav 8, e24820. [本文引用: 2]
[54]
PerochonA, JianguangJ, KahlaA, ArunachalamC, ScofieldSR, BowdenS, WallingtonE, DoohanFM (2015). TaFROG encodes a Pooideae orphan protein that interacts with SnRK1 and enhances resistance to the mycotoxigenic fungus Fusarium graminearum. Plant Phy- siol 169, 2895-2906. [本文引用: 1]
[55]
PolgeC, ThomasM (2007). SNF1/AMPK/SnRK1 kinases, global regulators at the heart of energy control.Trends Plant Sci 12, 20-28. [本文引用: 1]
[56]
RamonM, RuelensP, LiY, SheenJ, GeutenK, RollandF (2013). The hybrid Four-CBS-Domain KINβγ subunit functions as the canonical γ subunit of the plant energy sensor SnRK1. Plant J 75, 11-25. [本文引用: 1]
[57]
RodriguesA, AdamoM, CrozetP, MargalhaL, ConfrariaA, MartinhoC, EliasA, RabissiA, LumbrerasV, GonzálezGM (2013). ABI1 and PP2CA phosphatases are negative regulators of Snf1-related protein kinase1 signaling in Arabidopsis.Plant Cell 25, 3871-3884. [本文引用: 1]
[58]
SahaJ, ChatterjeeC, SenguptaA, GuptaK, GuptaB (2014). Genome-wide analysis and evolutionary study of sucrose non-fermenting 1-related protein kinase 2 (Sn- RK2) gene family members in Arabidopsis andOryza. Comput Biol Chem 49, 59-70. [本文引用: 1]
[59]
SanyalSK, PandeyA, PandeyGK (2015). The CBL-CIPK signaling module in plants: a mechanistic perspective.Physiol Plant 155, 89-108. [本文引用: 1]
[60]
SchuckS, BaldwinIT, BonaventureG (2013). HSPRO acts via SnRK1-mediated signaling in the regulation of Nicotiana attenuata seedling growth promoted by Pirifor- mospora indica. Plant Signali Behav 8, e23537. [本文引用: 1]
SonS, OhCJ, AnCS (2014). Arabidopsis thaliana remorins interact with SnRK1 and play a role in susceptibility to beet curly top virus and beet severe curly top virus.Plant Pathol J 30, 269-278. [本文引用: 1]
[63]
SongX, OhtaniM, HoriC, TakebayasiA, HiroyamaR, RejabNA, SuzukiT, DemuraT, YinT (2015). Physical interaction between SnRK2 and PP2C is conserved in Populus trichocarpa. Plant Biotech 32, 337-341. [本文引用: 1]
Thoday-KennedyEL, JacobsAK, RoySJ (2015). The role of the CBL-CIPK calcium signaling network in regulating ion transport in response to abiotic stress.Plant Growth Regul 76, 3-12. [本文引用: 1]
[67]
TianS, MaoX, ZhangH, ChenS, ZhaiC, YangS, JingR (2013). Cloning and characterization of TaSnRK2.3, a novel SnRK2 gene in common wheat. J Exp Bot 64, 2063-2080. [本文引用: 1]
[68]
TorreF, Gutiérrez-BeltránE, Pareja-JaimeY, Chakravar- thyS, MartinGB, PozoO (2013). The tomato calcium sensor Cbl10 and its interacting protein kinase Cipk6 define a signaling pathway in plant immunity.Plant Cell 25, 2748-2764. [本文引用: 1]
[69]
VlotAC, DempseyDMA, KlessigDF (2009). Salicylic acid, a multifaceted hormone to combat disease.Annu Rev Phytopathol 47, 177-206. [本文引用: 1]
[70]
WangL, HuW, SunJ, LiangX, YangX, WeiS, WangX, ZhouY, XiaoQ, YangG (2015a). Genome-wide analysis of SnRK gene family in Brachypodium distachyon and functional characterization of BdSnRK2.9. Plant Sci 237, 33-45. [本文引用: 1]
[71]
WangP, XueL, BatelliG, LeeS, HouYJ, VanOMJ, ZhangH, TaoWA, ZhuJK (2013). Quantitative phosphoproteomics identifies SnRK2 protein kinase substrates and reveals the effectors of abscisic acid action.Proc Natl Acad Sci USA 110, 11205-11210. [本文引用: 1]
[72]
WangP, ZhuJK, LangZ (2015b). Nitric oxide suppresses the inhibitory effect of abscisic acid on seed germination by S-nitrosylation of SnRK2 proteins. Plant Signal Behav 10, e1031939. [本文引用: 2]
XieC, ZhouX, DengX, GuoY (2010). PKS5, a SNF1- related kinase, interacts with and phosphorylates NPR1, and modulates expression of WRKY38 and WRKY62.J Genet Genomics 37, 359-369. [本文引用: 1]
[75]
XuanMA, XinMB, FengCX (2015). New players in ABA signaling: identification of PUB12/13 involved in degradation of ABA co-receptor ABI1.Sci China Life Sci 58, 1173-1174. [本文引用: 1]
YanS, DongX (2014). Perception of the plant immune signal salicylic acid.Curr Opin Plant Biol 20, 64-68. [本文引用: 1]
[78]
YoshidaR, UmezawaT, MizoguchiT, TakahashiS, TakahashiF, ShinozakiK (2006). The regulatory domain of SRK2E/OST1/SnRK2.6 interacts with ABI1 and integrates abscisic acid (ABA) and osmotic stress signals controlling stomatal closure in Arabidopsis.J Biol Chem 281, 5310-5318. [本文引用: 3]
[79]
YoshidaT, FujitaY, MaruyamaK, MogamiJ, TodakaD, ShinozakiK, YamaaguchiSK (2015). Four Arabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abs- cise acid signaling in response to osmotic stress.Plant Cell Environ 38, 35-49. [本文引用: 1]
[80]
YoshidaT, NishimuraN, KitahataN, KuromoriT, ItoT, AsamiT, ShinozakiK, HirayamaT (2006). ABA-Hypersensitive Germination3 encodes a protein phospha- tase 2C (AtPP2CA) that strongly regulates abscisic acid signaling during germination among Arabidopsis protein phosphatase 2Cs.Plant Physiol 140, 115-126.
[81]
ZhouL, LanW, ChenB, FangW, LuanS (2015a). A calcium sensor-regulated protein kinase, calcineurin B-like protein-interacting protein kinase19, is required for pollen tube growth and polarity.Plant Physiol 167, 1351-1360. [本文引用: 1]
[82]
ZhouX, HaoH, ZhangY, BaiY, ZhuW, QinY, YuanF, ZhaoF, WangM, HuJ (2015b). SOS2-like protein kinase5, an snf1-related protein kinase3-type protein kin- ase, is important for abscisic acid responses in Arabidopsis through phosphorylation of abscisic acid-insensitive.Plant Physiol 168, 659-676. [本文引用: 1]
[83]
ZulawskiM, BraginetsR, SchulzeWX (2013). PhosPhAt goes kinases-searchable protein kinase target information in the plant phosphorylation site database PhosPhAt.Nucleic Acids Res 41, D1176-D1184. [本文引用: 1]
2016
1 2012
... 植物SnRKs可以通过磷酸化与生长发育过程中的各种调节因子相互作用, 直接参与调控非生物胁迫下植株的生长发育, 增强植株对胁迫的耐性.烟草(Nico- tiana tabacum) SnRK1可以与作为种子萌发负调控因子的HSPRO (Ortholog of sugar beet Hs1pro-1)相互作用, 调控幼苗的发育(Schuck et al., 2013).马铃薯(Solanum tuberosum)中细胞程序性死亡抑制因子Adi3 (AvrPto-dependent Pto-interacting protein3)可以磷酸化SnRK1的β亚基, 磷酸化的SnRK1不能与底物特异性相互作用, 抑制SnRK1活性, 调节生长发育过程(Avila et al., 2012).AKIN10可通过磷酸化IDD8延迟花期(Jeong et al., 2015).Kim等(2012)则在研究拟南芥响应干旱胁迫时, 发现SnRK2.8磷酸化转录因子NTL6 (NAC (nam/ataf1/2/cuc2) with transmembrane motif1-like6), 延缓植株在响应干旱胁迫时的休眠进程.此外, 在拟南芥响应盐胁迫时, SnRK2.4和SnRK2.10可以与磷脂酸(phosphatidic acid, PA)结合, 参与不依赖ABA的非生物胁迫信号通路, 其中SnRK2.4有磷脂酸结合区域(PA-binding domain, PABD), 此结构域影响拟南芥的根系发育(Wang et al., 2015b). ...
2 2016
... 另外, 在ABA信号级联通路中, SnRKs可与ABA信号通路中的调节因子相互作用, 间接参与ABA信号转导途径, 调控植株对非生物胁迫的耐受性(Chen et al., 2012).例如, ABI2可分别与SnRK3.11、SnRK- 3.13和SnRK3.15相互作用, 而ABI1能够与SnRK3.6发生强烈相互作用, 它们互作后都能通过调控相关基因的表达增强植株对ABA信号的敏感性, 从而增强对盐胁迫的耐受性(Ohta et al., 2003).在拟南芥中, SnRK3.22可以磷酸化ABI5, 间接参与ABA响应基因的表达, 从而调节种子发芽(Zhou et al., 2015b).处于种子萌发期以及幼苗期的拟南芥, 在它们细胞核中, SnRK2.2、SnRK2.3和SnRK2.6通过磷酸化RAV1 (related to ABI3 (ABA-insensitive3)/VP1 transcription factor, RAV)负调控编码转录因子ABI3、ABI4和ABI5的表达, 参与ABA信号级联通路(Feng et al., 2014).SnRK2.2、SnRK2.6和SnRK2.3 (SRK2D/E/I)还可激活调节胁迫应答基因表达AREB1 (ABRE (ABA-responsive element)-binding factors1)、AREB2和AREB3, 从而调控植株响应渗透胁迫, 维持植物正常的营养生长(Yoshida et al., 2015).最新研究发现, ABA信号通路的新成员植物U-Box类E3连接酶(PUB (plant U-box E3 ligases))、PUB12和PUB13可以通过与ABA的信号元件ABI1相互作用, 释放与ABI1结合的SnRK2.6 (也被称为OST1), 促进OST1对下游靶标的磷酸化.研究者基于此提出了依赖ABA作用的新模型(Xuan et al., 2015).我们按照最新研究进展(Axel et al., 2016), 将SnRK2s参与的ABA途径总结为图1. ... ... 植物SnRK2s参与的主要ABA途径(改自Axel et al., 2016) (A) 无ABA时, ABA受体失活, PP2C抑制SnRK2s的活性; (B) 有ABA时, ABA受体与ABA结合, 抑制PP2C活性, SnRK2s通过自身磷酸化激活, 激活的SnRK2s磷酸化下游转录因子, 磷酸化的转录因子调控ABA胁迫响应基因的表达.同时, SnRK2s也可以磷酸化其它蛋白, 如AKT1及NADPH氧化酶等.这些事件共同引起植株体内ABA响应. ...
2 2008
... 植物在其生活史中一直面对各种生物与非生物胁迫.为了生存, 植物必须感知周围环境的变化, 并且针对这些变化产生防御机制以平衡生存, 完成生长和繁殖等生命过程.为此, 植物进化出了复杂且精密的逆境响应机制.其中, 植物蔗糖非发酵1 (SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinases, SnRKs)家族作为植物体内响应胁迫的一类关键蛋白激酶家族, 通过级联放大胁迫信号调控细胞内多条信号通路, 启动胁迫应答.SnRKs根据其不同的蛋白质三级结构可以细分为3个亚家族——SnRK1、SnRK2和SnRK3 (Coello et al., 2011).其中, SnRK1参与植物生命活动的氮、蔗糖和脂质代谢、器官发生以及衰老等过程, 且在逆境条件下通过调节能量和胁迫信号响应生物与非生物胁迫, 从而调节植物的生长发育(Baena-Gonzalez and Sheen, 2008; Confraria et al., 2013; Emanuelle et al., 2015); SnRK2亚家族成员因其C-末端具有2个独特的子结构域而与其它家族区分开(Yoshida et al., 2006).而对SnRK2的定量磷酸化蛋白质组学分析表明, SnRK2的底物蛋白有84种, 其参与生长发育, 如花期调控、RNA和DNA的结合、miRNA与表观遗传调控、信号转导、叶绿体功能以及离子运输等多种逆境响应过程(Wang et al., 2013).SnRK3蛋白激酶因有丰富的作用底物, 可以分别与CBL (calcineurin-B- like CBL calcium sensors)、SOS3 (salt overly sensitive)和ScaBP (SOS3-like calcium binding protein)相互作用(Dong et al., 2012), 因此, 又分别命名为CIPKs (CBL interacting protein kinases)、SOS2类蛋白激酶(SOS2-like kinases)或者与SOS2相关蛋白激酶PKSs (protein kinases related to SOS2), 它可与分布在质膜、液泡膜、细胞质基质以及细胞核中的10种CBL相互作用, 感受细胞内盐碱及干旱等胁迫信号, 启动细胞发生胁迫响应, 被认为位于植物能量代谢和胁迫信号连接的中心(Batistič et al., 2010; Thoday-Kennedy et al., 2015).因此, 深入了解SnRKs的作用机制将有助于植物耐性改良.虽然植物SnRKs家族发现较早, 但是由于其蛋白种类繁多、来源广泛且功能多样, 因此人们对其作用机制了解得还比较分散.本文通过总结植物SnRKs家族参与逆境信号转导的研究进展, 旨在拓展对SnRKs在 植物抗逆中作用的认识, 为植物抗逆机理的研究提供新思路. ... ... 植物SnRK1作为细胞信号网络中的关键组分, 可以分别与细胞内代表能量状态的不同形式的糖作用, 参与调节能量状态转变的其它激酶活性, 并与调控能量状态的转录因子相互作用, 调节细胞内的能量平衡, 是能量和逆境信号的交汇点(Baena-González and Sheen, 2008).例如, SnRK1可在黑暗、缺氧和盐碱条件下响应高浓度或低浓度的葡萄糖水平, 参与胁迫下植物能量代谢的调控(Polge and Thomas, 2007).海藻糖-6-磷酸(T6P)、葡萄糖-6-磷酸(G6P)和葡萄糖-1-磷酸(G1P)可分别抑制SnRK1的活性(Nägele and Weckwerth, 2014).在T6P/SnRK1信号通路中, 通过磷酸化过程抑制SnRK1活性, 加强细胞代谢活性, 加速生长(Lawlor and Paul, 2014).绿色器官中, 核糖-5-磷酸在向核糖-1-磷酸转化过程中能量的消耗, 也可以间接抑制SnRK1的活性(Nunes et al., 2013).SnRK1可与拟南芥中催化生成59-AMP的ADK (ade- nosine kinase)相互作用, 调节细胞中AMP与ATP之间的能量平衡(Mohannath et al., 2014).此外, 拟南芥中的SnRK1可磷酸化转录因子bZIP63, 调节下游靶基因的表达, 从而调控体内能量平衡(Mair et al., 2015).当细胞处于低能量状态时, AKIN10 (SnRK1的催化亚基)与拟南芥中的PTL (petal loss)相互作用, 抑制细胞分裂(O'Brien et al., 2015).同时, AKIN10也可以磷酸化花期调控转录因子IDD8 (indeterminate domain-containing transcription factor8), 参与IDD8介导的代谢过程, 调控开花时间, 使花期延迟(Jeong et al., 2015). ...
1 2009
... 植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015).其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014).PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011).同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ...
1 2010
... 植物在其生活史中一直面对各种生物与非生物胁迫.为了生存, 植物必须感知周围环境的变化, 并且针对这些变化产生防御机制以平衡生存, 完成生长和繁殖等生命过程.为此, 植物进化出了复杂且精密的逆境响应机制.其中, 植物蔗糖非发酵1 (SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinases, SnRKs)家族作为植物体内响应胁迫的一类关键蛋白激酶家族, 通过级联放大胁迫信号调控细胞内多条信号通路, 启动胁迫应答.SnRKs根据其不同的蛋白质三级结构可以细分为3个亚家族——SnRK1、SnRK2和SnRK3 (Coello et al., 2011).其中, SnRK1参与植物生命活动的氮、蔗糖和脂质代谢、器官发生以及衰老等过程, 且在逆境条件下通过调节能量和胁迫信号响应生物与非生物胁迫, 从而调节植物的生长发育(Baena-Gonzalez and Sheen, 2008; Confraria et al., 2013; Emanuelle et al., 2015); SnRK2亚家族成员因其C-末端具有2个独特的子结构域而与其它家族区分开(Yoshida et al., 2006).而对SnRK2的定量磷酸化蛋白质组学分析表明, SnRK2的底物蛋白有84种, 其参与生长发育, 如花期调控、RNA和DNA的结合、miRNA与表观遗传调控、信号转导、叶绿体功能以及离子运输等多种逆境响应过程(Wang et al., 2013).SnRK3蛋白激酶因有丰富的作用底物, 可以分别与CBL (calcineurin-B- like CBL calcium sensors)、SOS3 (salt overly sensitive)和ScaBP (SOS3-like calcium binding protein)相互作用(Dong et al., 2012), 因此, 又分别命名为CIPKs (CBL interacting protein kinases)、SOS2类蛋白激酶(SOS2-like kinases)或者与SOS2相关蛋白激酶PKSs (protein kinases related to SOS2), 它可与分布在质膜、液泡膜、细胞质基质以及细胞核中的10种CBL相互作用, 感受细胞内盐碱及干旱等胁迫信号, 启动细胞发生胁迫响应, 被认为位于植物能量代谢和胁迫信号连接的中心(Batistič et al., 2010; Thoday-Kennedy et al., 2015).因此, 深入了解SnRKs的作用机制将有助于植物耐性改良.虽然植物SnRKs家族发现较早, 但是由于其蛋白种类繁多、来源广泛且功能多样, 因此人们对其作用机制了解得还比较分散.本文通过总结植物SnRKs家族参与逆境信号转导的研究进展, 旨在拓展对SnRKs在 植物抗逆中作用的认识, 为植物抗逆机理的研究提供新思路. ...
1 2014
... 植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015).其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014).PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011).同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ...
1 2012
... 另外, 在ABA信号级联通路中, SnRKs可与ABA信号通路中的调节因子相互作用, 间接参与ABA信号转导途径, 调控植株对非生物胁迫的耐受性(Chen et al., 2012).例如, ABI2可分别与SnRK3.11、SnRK- 3.13和SnRK3.15相互作用, 而ABI1能够与SnRK3.6发生强烈相互作用, 它们互作后都能通过调控相关基因的表达增强植株对ABA信号的敏感性, 从而增强对盐胁迫的耐受性(Ohta et al., 2003).在拟南芥中, SnRK3.22可以磷酸化ABI5, 间接参与ABA响应基因的表达, 从而调节种子发芽(Zhou et al., 2015b).处于种子萌发期以及幼苗期的拟南芥, 在它们细胞核中, SnRK2.2、SnRK2.3和SnRK2.6通过磷酸化RAV1 (related to ABI3 (ABA-insensitive3)/VP1 transcription factor, RAV)负调控编码转录因子ABI3、ABI4和ABI5的表达, 参与ABA信号级联通路(Feng et al., 2014).SnRK2.2、SnRK2.6和SnRK2.3 (SRK2D/E/I)还可激活调节胁迫应答基因表达AREB1 (ABRE (ABA-responsive element)-binding factors1)、AREB2和AREB3, 从而调控植株响应渗透胁迫, 维持植物正常的营养生长(Yoshida et al., 2015).最新研究发现, ABA信号通路的新成员植物U-Box类E3连接酶(PUB (plant U-box E3 ligases))、PUB12和PUB13可以通过与ABA的信号元件ABI1相互作用, 释放与ABI1结合的SnRK2.6 (也被称为OST1), 促进OST1对下游靶标的磷酸化.研究者基于此提出了依赖ABA作用的新模型(Xuan et al., 2015).我们按照最新研究进展(Axel et al., 2016), 将SnRK2s参与的ABA途径总结为图1. ...
2013
2 2011
... 植物在其生活史中一直面对各种生物与非生物胁迫.为了生存, 植物必须感知周围环境的变化, 并且针对这些变化产生防御机制以平衡生存, 完成生长和繁殖等生命过程.为此, 植物进化出了复杂且精密的逆境响应机制.其中, 植物蔗糖非发酵1 (SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinases, SnRKs)家族作为植物体内响应胁迫的一类关键蛋白激酶家族, 通过级联放大胁迫信号调控细胞内多条信号通路, 启动胁迫应答.SnRKs根据其不同的蛋白质三级结构可以细分为3个亚家族——SnRK1、SnRK2和SnRK3 (Coello et al., 2011).其中, SnRK1参与植物生命活动的氮、蔗糖和脂质代谢、器官发生以及衰老等过程, 且在逆境条件下通过调节能量和胁迫信号响应生物与非生物胁迫, 从而调节植物的生长发育(Baena-Gonzalez and Sheen, 2008; Confraria et al., 2013; Emanuelle et al., 2015); SnRK2亚家族成员因其C-末端具有2个独特的子结构域而与其它家族区分开(Yoshida et al., 2006).而对SnRK2的定量磷酸化蛋白质组学分析表明, SnRK2的底物蛋白有84种, 其参与生长发育, 如花期调控、RNA和DNA的结合、miRNA与表观遗传调控、信号转导、叶绿体功能以及离子运输等多种逆境响应过程(Wang et al., 2013).SnRK3蛋白激酶因有丰富的作用底物, 可以分别与CBL (calcineurin-B- like CBL calcium sensors)、SOS3 (salt overly sensitive)和ScaBP (SOS3-like calcium binding protein)相互作用(Dong et al., 2012), 因此, 又分别命名为CIPKs (CBL interacting protein kinases)、SOS2类蛋白激酶(SOS2-like kinases)或者与SOS2相关蛋白激酶PKSs (protein kinases related to SOS2), 它可与分布在质膜、液泡膜、细胞质基质以及细胞核中的10种CBL相互作用, 感受细胞内盐碱及干旱等胁迫信号, 启动细胞发生胁迫响应, 被认为位于植物能量代谢和胁迫信号连接的中心(Batistič et al., 2010; Thoday-Kennedy et al., 2015).因此, 深入了解SnRKs的作用机制将有助于植物耐性改良.虽然植物SnRKs家族发现较早, 但是由于其蛋白种类繁多、来源广泛且功能多样, 因此人们对其作用机制了解得还比较分散.本文通过总结植物SnRKs家族参与逆境信号转导的研究进展, 旨在拓展对SnRKs在 植物抗逆中作用的认识, 为植物抗逆机理的研究提供新思路. ... ... 丝氨酸/苏氨酸蛋白激酶是一个相对较大的蛋白激酶家族, 其中, AMPK (AMP-activated kinase)/SnRKs是高度保守的丝氨酸/苏氨酸蛋白激酶家族的重要成员(Emanuelle et al., 2015). SnRKs可以细分成3个亚家族, 即SnRK1、SnRK2和SnRK3 (Coello et al., 2011).酵母SNF1 (sucrose nonfermenting1)、蛔虫AAK (AMP-activated kinase)、昆虫AMPK、哺乳动物的AMPK以及植物的SnRK1都属于SnRK1蛋白激酶亚家族, 而SnRK2和SnRK3则为植物特有的亚家族(Ghillebert et al., 2011; Emanuelle et al., 2015). ...
1 2013
... 植物在其生活史中一直面对各种生物与非生物胁迫.为了生存, 植物必须感知周围环境的变化, 并且针对这些变化产生防御机制以平衡生存, 完成生长和繁殖等生命过程.为此, 植物进化出了复杂且精密的逆境响应机制.其中, 植物蔗糖非发酵1 (SNF1)相关蛋白激酶(sucrose nonfermenting-1-related protein kinases, SnRKs)家族作为植物体内响应胁迫的一类关键蛋白激酶家族, 通过级联放大胁迫信号调控细胞内多条信号通路, 启动胁迫应答.SnRKs根据其不同的蛋白质三级结构可以细分为3个亚家族——SnRK1、SnRK2和SnRK3 (Coello et al., 2011).其中, SnRK1参与植物生命活动的氮、蔗糖和脂质代谢、器官发生以及衰老等过程, 且在逆境条件下通过调节能量和胁迫信号响应生物与非生物胁迫, 从而调节植物的生长发育(Baena-Gonzalez and Sheen, 2008; Confraria et al., 2013; Emanuelle et al., 2015); SnRK2亚家族成员因其C-末端具有2个独特的子结构域而与其它家族区分开(Yoshida et al., 2006).而对SnRK2的定量磷酸化蛋白质组学分析表明, SnRK2的底物蛋白有84种, 其参与生长发育, 如花期调控、RNA和DNA的结合、miRNA与表观遗传调控、信号转导、叶绿体功能以及离子运输等多种逆境响应过程(Wang et al., 2013).SnRK3蛋白激酶因有丰富的作用底物, 可以分别与CBL (calcineurin-B- like CBL calcium sensors)、SOS3 (salt overly sensitive)和ScaBP (SOS3-like calcium binding protein)相互作用(Dong et al., 2012), 因此, 又分别命名为CIPKs (CBL interacting protein kinases)、SOS2类蛋白激酶(SOS2-like kinases)或者与SOS2相关蛋白激酶PKSs (protein kinases related to SOS2), 它可与分布在质膜、液泡膜、细胞质基质以及细胞核中的10种CBL相互作用, 感受细胞内盐碱及干旱等胁迫信号, 启动细胞发生胁迫响应, 被认为位于植物能量代谢和胁迫信号连接的中心(Batistič et al., 2010; Thoday-Kennedy et al., 2015).因此, 深入了解SnRKs的作用机制将有助于植物耐性改良.虽然植物SnRKs家族发现较早, 但是由于其蛋白种类繁多、来源广泛且功能多样, 因此人们对其作用机制了解得还比较分散.本文通过总结植物SnRKs家族参与逆境信号转导的研究进展, 旨在拓展对SnRKs在 植物抗逆中作用的认识, 为植物抗逆机理的研究提供新思路. ...
3 2016
... 当植物受到胁迫时, SnRKs迅速感受刺激并产生应激反应.此时, SnRK1主要通过调整代谢来改变细胞内的能量胁迫状态(Halford and Hey, 2009).但是, 当应激反应强度过大且时间持久时, 活化的SnRK1则触发自身的小型泛素化修饰(small ubiquitin-like modifier, SUMO)过程, 通过蛋白酶体途径进行降解, 进而抑制细胞的正常生理活动(Nägele and Weckwerth, 2014; Crozet et al., 2016).此外, SnRK1的降解是严格依赖其自身活性的(Crozet et al., 2016).例如, 无活性的SnRK1在植物体内是非常稳定的, 但是, 当SnRK1被激活时, 同时启动了SUMO化过程(Crozet et al., 2016).此外, Rodrigues等(2013)发现, A组PP2C作为ABA介导的胁迫信号通路中的抑制因子, 可以与SnRK1的催化亚基相互作用使其去磷酸化并失活.同时, SnRK1靶基因表达分析进一步证明, 由于ABA抑制PP2C使SnRK1激活, 在能量匮乏时, SnRK1激活下游的靶标(Rodrigues et al., 2013).可见, SnRK1的失活与激活可能通过自身的严格控制而达到平衡. ... ... ).此外, SnRK1的降解是严格依赖其自身活性的(Crozet et al., 2016).例如, 无活性的SnRK1在植物体内是非常稳定的, 但是, 当SnRK1被激活时, 同时启动了SUMO化过程(Crozet et al., 2016).此外, Rodrigues等(2013)发现, A组PP2C作为ABA介导的胁迫信号通路中的抑制因子, 可以与SnRK1的催化亚基相互作用使其去磷酸化并失活.同时, SnRK1靶基因表达分析进一步证明, 由于ABA抑制PP2C使SnRK1激活, 在能量匮乏时, SnRK1激活下游的靶标(Rodrigues et al., 2013).可见, SnRK1的失活与激活可能通过自身的严格控制而达到平衡. ... ... ).例如, 无活性的SnRK1在植物体内是非常稳定的, 但是, 当SnRK1被激活时, 同时启动了SUMO化过程(Crozet et al., 2016).此外, Rodrigues等(2013)发现, A组PP2C作为ABA介导的胁迫信号通路中的抑制因子, 可以与SnRK1的催化亚基相互作用使其去磷酸化并失活.同时, SnRK1靶基因表达分析进一步证明, 由于ABA抑制PP2C使SnRK1激活, 在能量匮乏时, SnRK1激活下游的靶标(Rodrigues et al., 2013).可见, SnRK1的失活与激活可能通过自身的严格控制而达到平衡. ...
... 丝氨酸/苏氨酸蛋白激酶是一个相对较大的蛋白激酶家族, 其中, AMPK (AMP-activated kinase)/SnRKs是高度保守的丝氨酸/苏氨酸蛋白激酶家族的重要成员(Emanuelle et al., 2015). SnRKs可以细分成3个亚家族, 即SnRK1、SnRK2和SnRK3 (Coello et al., 2011).酵母SNF1 (sucrose nonfermenting1)、蛔虫AAK (AMP-activated kinase)、昆虫AMPK、哺乳动物的AMPK以及植物的SnRK1都属于SnRK1蛋白激酶亚家族, 而SnRK2和SnRK3则为植物特有的亚家族(Ghillebert et al., 2011; Emanuelle et al., 2015). ...
1 1998
... 植物SnRK2亚家族成员含有2个典型结构域: N端催化结构域和C末端调节结构域(Halford and Hardie, 1998).C末端结构域又包括2个子结构域, 即结构域1和结构域2.其中, 结构域1被称为ABA结合框, 因其富含负电荷, 可与PP2C (phosphatase 2C)结合; 结构域2主要是SnRK2的2和3亚组成员的特有结构, 富含天冬氨酸(Yoshida et al., 2006).对SnRK2亚家族进行全基因组分析, 发现其在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中各有10个成员, 但是染色体定位在二者之间有差异(Wang et al., 2015a).如在拟南芥所有染色体上都有SnRK2亚家族基因, 而在水稻中其分别位于1、4、7、10和12号染色体, 在小麦(Triticum aestivum)中则定位于1A、1B及1D染色体上, 暗示其生理功能可能因物种的不同而异(Tian et al., 2013; Saha et al., 2014). ...
1 2009
... 当植物受到胁迫时, SnRKs迅速感受刺激并产生应激反应.此时, SnRK1主要通过调整代谢来改变细胞内的能量胁迫状态(Halford and Hey, 2009).但是, 当应激反应强度过大且时间持久时, 活化的SnRK1则触发自身的小型泛素化修饰(small ubiquitin-like modifier, SUMO)过程, 通过蛋白酶体途径进行降解, 进而抑制细胞的正常生理活动(Nägele and Weckwerth, 2014; Crozet et al., 2016).此外, SnRK1的降解是严格依赖其自身活性的(Crozet et al., 2016).例如, 无活性的SnRK1在植物体内是非常稳定的, 但是, 当SnRK1被激活时, 同时启动了SUMO化过程(Crozet et al., 2016).此外, Rodrigues等(2013)发现, A组PP2C作为ABA介导的胁迫信号通路中的抑制因子, 可以与SnRK1的催化亚基相互作用使其去磷酸化并失活.同时, SnRK1靶基因表达分析进一步证明, 由于ABA抑制PP2C使SnRK1激活, 在能量匮乏时, SnRK1激活下游的靶标(Rodrigues et al., 2013).可见, SnRK1的失活与激活可能通过自身的严格控制而达到平衡. ...
1 2004
... 在真核生物中, AMPK/SNF1/SnRK1蛋白激酶是进化上高度保守的代谢感知因子(Emanuelle et al., 2015).SnRK1与SNF1和AMPK的氨基酸序列一致性均为48%, 其中催化域的氨基酸序列约有62%一致(Halford et al., 2004).SnRK1通常是由具催化功能的α亚基、起调节作用的β亚基以及有活化功能的γ亚基组成的多聚体蛋白激酶(Emanuelle et al., 2015).而拟南芥中SnRK1则是由α亚基、非典型的β3亚基以及植物中特有的βγ亚基组成(Ramon et al., 2013; Emanuelle et al., 2015). ...
1 2012
... 植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015).其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014).PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011).同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ...
1 2010
... 2009年, ABA受体的发现为研究SnRKs参与胁迫信号转导通路提供了重要的节点.其中, ABA作为胁迫的响应因子, 可分别与PYR/PYL/RCAR (pyrabactin resistant/PYR-like/regulatory component of ABA) ABA受体结合, 激活的ABA受体抑制包括ABI1 (abs- cisic acid-insenitive 1)、ABI2、HAB1 (homology to ABI1)、HAB2、AHG1 (ABA-hypersensitive germination 1)以及AHG3等在内的9个A组PP2C的活性.失去活性的A组PP2C无法抑制SnRK2s的活性, SnRK2s通过自身磷酸化激活.激活的SnRK2s磷酸化下游的靶标, 从而进一步放大胁迫信号, 增强植株对非生物胁迫的耐性(Yoshida et al., 2006; Hirayama and Umezawa, 2010; Danquah et al., 2014). ...
2009
2011
2 2015
... 植物SnRK1作为细胞信号网络中的关键组分, 可以分别与细胞内代表能量状态的不同形式的糖作用, 参与调节能量状态转变的其它激酶活性, 并与调控能量状态的转录因子相互作用, 调节细胞内的能量平衡, 是能量和逆境信号的交汇点(Baena-González and Sheen, 2008).例如, SnRK1可在黑暗、缺氧和盐碱条件下响应高浓度或低浓度的葡萄糖水平, 参与胁迫下植物能量代谢的调控(Polge and Thomas, 2007).海藻糖-6-磷酸(T6P)、葡萄糖-6-磷酸(G6P)和葡萄糖-1-磷酸(G1P)可分别抑制SnRK1的活性(Nägele and Weckwerth, 2014).在T6P/SnRK1信号通路中, 通过磷酸化过程抑制SnRK1活性, 加强细胞代谢活性, 加速生长(Lawlor and Paul, 2014).绿色器官中, 核糖-5-磷酸在向核糖-1-磷酸转化过程中能量的消耗, 也可以间接抑制SnRK1的活性(Nunes et al., 2013).SnRK1可与拟南芥中催化生成59-AMP的ADK (ade- nosine kinase)相互作用, 调节细胞中AMP与ATP之间的能量平衡(Mohannath et al., 2014).此外, 拟南芥中的SnRK1可磷酸化转录因子bZIP63, 调节下游靶基因的表达, 从而调控体内能量平衡(Mair et al., 2015).当细胞处于低能量状态时, AKIN10 (SnRK1的催化亚基)与拟南芥中的PTL (petal loss)相互作用, 抑制细胞分裂(O'Brien et al., 2015).同时, AKIN10也可以磷酸化花期调控转录因子IDD8 (indeterminate domain-containing transcription factor8), 参与IDD8介导的代谢过程, 调控开花时间, 使花期延迟(Jeong et al., 2015). ... ... 植物SnRKs可以通过磷酸化与生长发育过程中的各种调节因子相互作用, 直接参与调控非生物胁迫下植株的生长发育, 增强植株对胁迫的耐性.烟草(Nico- tiana tabacum) SnRK1可以与作为种子萌发负调控因子的HSPRO (Ortholog of sugar beet Hs1pro-1)相互作用, 调控幼苗的发育(Schuck et al., 2013).马铃薯(Solanum tuberosum)中细胞程序性死亡抑制因子Adi3 (AvrPto-dependent Pto-interacting protein3)可以磷酸化SnRK1的β亚基, 磷酸化的SnRK1不能与底物特异性相互作用, 抑制SnRK1活性, 调节生长发育过程(Avila et al., 2012).AKIN10可通过磷酸化IDD8延迟花期(Jeong et al., 2015).Kim等(2012)则在研究拟南芥响应干旱胁迫时, 发现SnRK2.8磷酸化转录因子NTL6 (NAC (nam/ataf1/2/cuc2) with transmembrane motif1-like6), 延缓植株在响应干旱胁迫时的休眠进程.此外, 在拟南芥响应盐胁迫时, SnRK2.4和SnRK2.10可以与磷脂酸(phosphatidic acid, PA)结合, 参与不依赖ABA的非生物胁迫信号通路, 其中SnRK2.4有磷脂酸结合区域(PA-binding domain, PABD), 此结构域影响拟南芥的根系发育(Wang et al., 2015b). ...
... 植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015).其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014).PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011).同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ... ... ).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ...
1 2011
... 植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015).其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014).PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011).同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ...
1 2014
... 植物SnRK1作为细胞信号网络中的关键组分, 可以分别与细胞内代表能量状态的不同形式的糖作用, 参与调节能量状态转变的其它激酶活性, 并与调控能量状态的转录因子相互作用, 调节细胞内的能量平衡, 是能量和逆境信号的交汇点(Baena-González and Sheen, 2008).例如, SnRK1可在黑暗、缺氧和盐碱条件下响应高浓度或低浓度的葡萄糖水平, 参与胁迫下植物能量代谢的调控(Polge and Thomas, 2007).海藻糖-6-磷酸(T6P)、葡萄糖-6-磷酸(G6P)和葡萄糖-1-磷酸(G1P)可分别抑制SnRK1的活性(Nägele and Weckwerth, 2014).在T6P/SnRK1信号通路中, 通过磷酸化过程抑制SnRK1活性, 加强细胞代谢活性, 加速生长(Lawlor and Paul, 2014).绿色器官中, 核糖-5-磷酸在向核糖-1-磷酸转化过程中能量的消耗, 也可以间接抑制SnRK1的活性(Nunes et al., 2013).SnRK1可与拟南芥中催化生成59-AMP的ADK (ade- nosine kinase)相互作用, 调节细胞中AMP与ATP之间的能量平衡(Mohannath et al., 2014).此外, 拟南芥中的SnRK1可磷酸化转录因子bZIP63, 调节下游靶基因的表达, 从而调控体内能量平衡(Mair et al., 2015).当细胞处于低能量状态时, AKIN10 (SnRK1的催化亚基)与拟南芥中的PTL (petal loss)相互作用, 抑制细胞分裂(O'Brien et al., 2015).同时, AKIN10也可以磷酸化花期调控转录因子IDD8 (indeterminate domain-containing transcription factor8), 参与IDD8介导的代谢过程, 调控开花时间, 使花期延迟(Jeong et al., 2015). ...
2 2015
... 水杨酸(salicylic acid, SA)在植物中调控防御基因的表达及宿主细胞死亡, 从而在宿主获得系统性免疫(systemic acquired resistance, SAR)中发挥重要作用, 而NPR1 (nonexpressor of pathogenesis-related gene1)是SA介导信号通路中的主要调节因子(Vlot et al., 2009; Yan and Dong, 2014).SA介导通路中的信号分子可以激活编码SnRK2.8蛋白激酶的基因, 上调该基因的表达(Lee et al., 2015).同时, SnRK2.8也可磷酸化NPR1, 使其入核并调控相应的转录因子表达, 增强植株的SAR能力(Lee et al., 2015).此外, 在拟南芥SnRK3.22基因缺失突变体中, 由于SnRK3.22磷酸化NPR1的缺失, 进入核内的NPR1减少, NPR1的靶基因WRKY38 (sequence-specific DNA-bind- ing proteins38 of the N-terminal sequence WRKY- GQK)和WRKY62的表达下调, 减弱了植株对病虫害的抗性(Xie et al., 2010).可见, 在植株受到生物胁迫时, SnRKs可通过磷酸化NPR1参与SA介导的信号通路, 从转录水平上迅速启动防御机制, 以增强植株抵抗生物胁迫的能力. ... ... ).同时, SnRK2.8也可磷酸化NPR1, 使其入核并调控相应的转录因子表达, 增强植株的SAR能力(Lee et al., 2015).此外, 在拟南芥SnRK3.22基因缺失突变体中, 由于SnRK3.22磷酸化NPR1的缺失, 进入核内的NPR1减少, NPR1的靶基因WRKY38 (sequence-specific DNA-bind- ing proteins38 of the N-terminal sequence WRKY- GQK)和WRKY62的表达下调, 减弱了植株对病虫害的抗性(Xie et al., 2010).可见, 在植株受到生物胁迫时, SnRKs可通过磷酸化NPR1参与SA介导的信号通路, 从转录水平上迅速启动防御机制, 以增强植株抵抗生物胁迫的能力. ...
1 2015
... 植物SnRKs参与ABA信号介导的非生物胁迫应答大致可分为直接和间接2种途径.直接途径是SnRKs通过自身的磷酸化或被磷酸化, 直接参与ABA信号介导非生物胁迫过程, 增强植株对干旱或盐碱等非生物胁迫的耐性.例如, 在拟南芥中, SnRK- 2.2/SnRK2.3/SnRK2.6是ABA信号转导通路中的重要调节因子, 其中在保卫细胞中响应ABA的ABAR (ab- scisic acid receptor)可直接与SnRK2.6相互作用, 调控ABA信号转导, 进而调节气孔运动(Liang et al., 2015).而在ABA信号通路中产生的NO, 还可使保卫细胞中的SnRK2.2/SnRK2.3/SnRK2.6的巯基亚硝基化, 抑制SnRK2.2/SnRK2.3/SnRK2.6的活性, 调节由ABA参与调控的种子萌发及幼苗生长(Wang et al., 2015b).PP2Cs的3个成员(ABI1、AHG1和AHG3)与SnRK2.10强烈相互作用, 介导SnRK2.10参与ABA信号转导, 响应毛果杨(Populus trichocarpa)对干旱和盐碱胁迫的应答(Song et al., 2015). ...
... 在病毒或微生物入侵时, 植物SnRK1可以通过磷酸化帽结合蛋白、翻译起始因子或者病毒蛋白参与生物胁迫响应.拟南芥SnRK1也可以磷酸化双生病毒AL2蛋白, 抑制双生病毒的转录与翻译, 延迟白菜卷叶病毒的感染(Shen et al., 2014).在小麦中, SnRK1与TaFROG (Triticum aestivum fusarium resistance orphan gene)编码的蛋白质相互作用, 增强小麦抵御禾谷镰刀菌的能力以及对呕吐毒素(deoxynivalenol, DON)的耐性(Perochon et al., 2015).Kim等(2015) 发现受到真菌和细菌病原体侵染的水稻OSK35-D (水稻SnRK1b的编码基因)激活型突变体能增强基因表达的重编辑, 从而提高对病原体的抗性.此外, 当双生病毒感染拟南芥时, SnRK1磷酸化AtREM4s (Arabidopsis thaliana remorin group 4s), 进而参与26S蛋白酶体途径, 降解甜菜严重曲顶病毒, 增强抗病性(Son et al., 2014). ...
1 2007
... 植物SnRK1作为细胞信号网络中的关键组分, 可以分别与细胞内代表能量状态的不同形式的糖作用, 参与调节能量状态转变的其它激酶活性, 并与调控能量状态的转录因子相互作用, 调节细胞内的能量平衡, 是能量和逆境信号的交汇点(Baena-González and Sheen, 2008).例如, SnRK1可在黑暗、缺氧和盐碱条件下响应高浓度或低浓度的葡萄糖水平, 参与胁迫下植物能量代谢的调控(Polge and Thomas, 2007).海藻糖-6-磷酸(T6P)、葡萄糖-6-磷酸(G6P)和葡萄糖-1-磷酸(G1P)可分别抑制SnRK1的活性(Nägele and Weckwerth, 2014).在T6P/SnRK1信号通路中, 通过磷酸化过程抑制SnRK1活性, 加强细胞代谢活性, 加速生长(Lawlor and Paul, 2014).绿色器官中, 核糖-5-磷酸在向核糖-1-磷酸转化过程中能量的消耗, 也可以间接抑制SnRK1的活性(Nunes et al., 2013).SnRK1可与拟南芥中催化生成59-AMP的ADK (ade- nosine kinase)相互作用, 调节细胞中AMP与ATP之间的能量平衡(Mohannath et al., 2014).此外, 拟南芥中的SnRK1可磷酸化转录因子bZIP63, 调节下游靶基因的表达, 从而调控体内能量平衡(Mair et al., 2015).当细胞处于低能量状态时, AKIN10 (SnRK1的催化亚基)与拟南芥中的PTL (petal loss)相互作用, 抑制细胞分裂(O'Brien et al., 2015).同时, AKIN10也可以磷酸化花期调控转录因子IDD8 (indeterminate domain-containing transcription factor8), 参与IDD8介导的代谢过程, 调控开花时间, 使花期延迟(Jeong et al., 2015). ...
1 2013
... 在真核生物中, AMPK/SNF1/SnRK1蛋白激酶是进化上高度保守的代谢感知因子(Emanuelle et al., 2015).SnRK1与SNF1和AMPK的氨基酸序列一致性均为48%, 其中催化域的氨基酸序列约有62%一致(Halford et al., 2004).SnRK1通常是由具催化功能的α亚基、起调节作用的β亚基以及有活化功能的γ亚基组成的多聚体蛋白激酶(Emanuelle et al., 2015).而拟南芥中SnRK1则是由α亚基、非典型的β3亚基以及植物中特有的βγ亚基组成(Ramon et al., 2013; Emanuelle et al., 2015). ...
1 2013
... 当植物受到胁迫时, SnRKs迅速感受刺激并产生应激反应.此时, SnRK1主要通过调整代谢来改变细胞内的能量胁迫状态(Halford and Hey, 2009).但是, 当应激反应强度过大且时间持久时, 活化的SnRK1则触发自身的小型泛素化修饰(small ubiquitin-like modifier, SUMO)过程, 通过蛋白酶体途径进行降解, 进而抑制细胞的正常生理活动(Nägele and Weckwerth, 2014; Crozet et al., 2016).此外, SnRK1的降解是严格依赖其自身活性的(Crozet et al., 2016).例如, 无活性的SnRK1在植物体内是非常稳定的, 但是, 当SnRK1被激活时, 同时启动了SUMO化过程(Crozet et al., 2016).此外, Rodrigues等(2013)发现, A组PP2C作为ABA介导的胁迫信号通路中的抑制因子, 可以与SnRK1的催化亚基相互作用使其去磷酸化并失活.同时, SnRK1靶基因表达分析进一步证明, 由于ABA抑制PP2C使SnRK1激活, 在能量匮乏时, SnRK1激活下游的靶标(Rodrigues et al., 2013).可见, SnRK1的失活与激活可能通过自身的严格控制而达到平衡. ...
1 2014
... 植物SnRK2亚家族成员含有2个典型结构域: N端催化结构域和C末端调节结构域(Halford and Hardie, 1998).C末端结构域又包括2个子结构域, 即结构域1和结构域2.其中, 结构域1被称为ABA结合框, 因其富含负电荷, 可与PP2C (phosphatase 2C)结合; 结构域2主要是SnRK2的2和3亚组成员的特有结构, 富含天冬氨酸(Yoshida et al., 2006).对SnRK2亚家族进行全基因组分析, 发现其在拟南芥(Arabidopsis thaliana)和水稻(Oryza sativa)中各有10个成员, 但是染色体定位在二者之间有差异(Wang et al., 2015a).如在拟南芥所有染色体上都有SnRK2亚家族基因, 而在水稻中其分别位于1、4、7、10和12号染色体, 在小麦(Triticum aestivum)中则定位于1A、1B及1D染色体上, 暗示其生理功能可能因物种的不同而异(Tian et al., 2013; Saha et al., 2014). ...
1 2015
... 植物SnRK3蛋白激酶具有丰富的作用底物, 使其在细胞胁迫信号通路与碳、氮代谢通路中均发挥重要作用, 它包括1个N末端激酶催化结构域、1个连接结构域和1个高度可变的C末端调节结构域(Sanyal et al., 2015).其中, 催化结构域包括1个活化环, 调节结构域还可分为NAF (designated according to the prominent amino acids N, A and F)结构域和PPI结构域(protein phosphatase interaction) (Batistič and Kudla, 2009), 推测其保守的钙依赖性NAF结构域不仅是调控SnRK3的基础, 也是上游激酶可变调节的基础(Chaves-Sanjuan et al., 2014), 介导了SnRK3与CBL之间的相互作用(Kleist et al., 2014).PPI是SnRK3与ABI相互作用时的结合位点(Lan et al., 2011).同时, SnRK3激酶的激活需要活性环从活化位点释放以及NAF结构域从核苷酸结合位点释放, 从而基于NAF自抑制结构域与CBL的相互作用体现其生物活性(Hashimoto et al., 2012).例如, SnRK3.23的活化环可折叠成有序的结构, 阻碍活性位点接触底物.而SnRK3.11的活性环能够产生活性位点, 并可进行催化反应(Kleist et al., 2014). ...
1 2013
... 植物SnRKs可以通过磷酸化与生长发育过程中的各种调节因子相互作用, 直接参与调控非生物胁迫下植株的生长发育, 增强植株对胁迫的耐性.烟草(Nico- tiana tabacum) SnRK1可以与作为种子萌发负调控因子的HSPRO (Ortholog of sugar beet Hs1pro-1)相互作用, 调控幼苗的发育(Schuck et al., 2013).马铃薯(Solanum tuberosum)中细胞程序性死亡抑制因子Adi3 (AvrPto-dependent Pto-interacting protein3)可以磷酸化SnRK1的β亚基, 磷酸化的SnRK1不能与底物特异性相互作用, 抑制SnRK1活性, 调节生长发育过程(Avila et al., 2012).AKIN10可通过磷酸化IDD8延迟花期(Jeong et al., 2015).Kim等(2012)则在研究拟南芥响应干旱胁迫时, 发现SnRK2.8磷酸化转录因子NTL6 (NAC (nam/ataf1/2/cuc2) with transmembrane motif1-like6), 延缓植株在响应干旱胁迫时的休眠进程.此外, 在拟南芥响应盐胁迫时, SnRK2.4和SnRK2.10可以与磷脂酸(phosphatidic acid, PA)结合, 参与不依赖ABA的非生物胁迫信号通路, 其中SnRK2.4有磷脂酸结合区域(PA-binding domain, PABD), 此结构域影响拟南芥的根系发育(Wang et al., 2015b). ...
1 2014
... 在病毒或微生物入侵时, 植物SnRK1可以通过磷酸化帽结合蛋白、翻译起始因子或者病毒蛋白参与生物胁迫响应.拟南芥SnRK1也可以磷酸化双生病毒AL2蛋白, 抑制双生病毒的转录与翻译, 延迟白菜卷叶病毒的感染(Shen et al., 2014).在小麦中, SnRK1与TaFROG (Triticum aestivum fusarium resistance orphan gene)编码的蛋白质相互作用, 增强小麦抵御禾谷镰刀菌的能力以及对呕吐毒素(deoxynivalenol, DON)的耐性(Perochon et al., 2015).Kim等(2015) 发现受到真菌和细菌病原体侵染的水稻OSK35-D (水稻SnRK1b的编码基因)激活型突变体能增强基因表达的重编辑, 从而提高对病原体的抗性.此外, 当双生病毒感染拟南芥时, SnRK1磷酸化AtREM4s (Arabidopsis thaliana remorin group 4s), 进而参与26S蛋白酶体途径, 降解甜菜严重曲顶病毒, 增强抗病性(Son et al., 2014). ...
1 2014
... 在病毒或微生物入侵时, 植物SnRK1可以通过磷酸化帽结合蛋白、翻译起始因子或者病毒蛋白参与生物胁迫响应.拟南芥SnRK1也可以磷酸化双生病毒AL2蛋白, 抑制双生病毒的转录与翻译, 延迟白菜卷叶病毒的感染(Shen et al., 2014).在小麦中, SnRK1与TaFROG (Triticum aestivum fusarium resistance orphan gene)编码的蛋白质相互作用, 增强小麦抵御禾谷镰刀菌的能力以及对呕吐毒素(deoxynivalenol, DON)的耐性(Perochon et al., 2015).Kim等(2015) 发现受到真菌和细菌病原体侵染的水稻OSK35-D (水稻SnRK1b的编码基因)激活型突变体能增强基因表达的重编辑, 从而提高对病原体的抗性.此外, 当双生病毒感染拟南芥时, SnRK1磷酸化AtREM4s (Arabidopsis thaliana remorin group 4s), 进而参与26S蛋白酶体途径, 降解甜菜严重曲顶病毒, 增强抗病性(Son et al., 2014). ...
1 2015
... 植物SnRKs参与ABA信号介导的非生物胁迫应答大致可分为直接和间接2种途径.直接途径是SnRKs通过自身的磷酸化或被磷酸化, 直接参与ABA信号介导非生物胁迫过程, 增强植株对干旱或盐碱等非生物胁迫的耐性.例如, 在拟南芥中, SnRK- 2.2/SnRK2.3/SnRK2.6是ABA信号转导通路中的重要调节因子, 其中在保卫细胞中响应ABA的ABAR (ab- scisic acid receptor)可直接与SnRK2.6相互作用, 调控ABA信号转导, 进而调节气孔运动(Liang et al., 2015).而在ABA信号通路中产生的NO, 还可使保卫细胞中的SnRK2.2/SnRK2.3/SnRK2.6的巯基亚硝基化, 抑制SnRK2.2/SnRK2.3/SnRK2.6的活性, 调节由ABA参与调控的种子萌发及幼苗生长(Wang et al., 2015b).PP2Cs的3个成员(ABI1、AHG1和AHG3)与SnRK2.10强烈相互作用, 介导SnRK2.10参与ABA信号转导, 响应毛果杨(Populus trichocarpa)对干旱和盐碱胁迫的应答(Song et al., 2015). ...
 ,
, 

{kind=link}
{kind=link}