Abstract: Located between the cell wall and plasma membrane as a secondary metabolite, suberin typically distributes in rhizodermis and the boundary tissue of stems. Based on glycerol, suberin is a heteropolymer composed of polya- liphatics and polyaromatics, and it could slow the outflow of water and nutrient substance, limit pathogen invasion and prevent toxic gas from diffusing to plants. Recently, with people’s focus on the storage and processing of fruits and vegetables, as well plant resistance, the research in suberin is increasing, especially in the aspects of metabolic enzymes and corresponding genes and the metabolite’s function. In this paper, we elaborate the research progress in suberin histochemistry, the biosynthesis pathway as well related enzymes and genes. We introduce recent advances in the transport of suberin components intracellularly and to the cell wall, polymer assembly, and the regulation of suberin deposition and present the research development of suberin physiological function. This research is expected to provide significant information for further research and application of suberin.
图1 木栓质在根不同发育阶段的堆积位点(改自Vishwanath et al., 2015) 图中所示为典型双子叶植物(例如拟南芥)根的横切面。(A) 内胚层发育过程中细胞壁的初生生长阶段, 木栓质片层在其内表面有所积累(黄色), 凯氏带(红色)在内皮层细胞间的结合点处积累(ML: 中间层; PW: 初生细胞壁; SL: 木栓质片层; PM: 细胞质膜; Cy: 细胞质); (B) 次生生长阶段, 木栓质堆积于木栓组织/栓化细胞(周皮)中 Figure 1 Sites of suberin deposition in different developmental stages of roots (modified from Vishwanath et al., 2015) The diagrams show the root cross-sections of typical dicot plants (e.g. Arabidopsis thaliana). (A) Primary growth stage of cell wall during endodermis development with suberin lamellae deposition in the inner face of endodermal cell walls (yellow) and deposition of Casparian bands (red) localized at endodermal cell-cell junctions (ML: Middle lamellae; PW: Primary cell wall; SL: Suberin lamellae; PM: Plasma membrane; Cy: Cytoplasm); (B) Secondary growth stage with suberin deposition in the phellem/cork cells (periderms)
木栓质片层超微结构可进一步通过透射电子显微镜观察。Schmutz等(1996)通过抑制脂肪酸链的延长, 发现木栓质浅色片层的厚度和脂肪酸的烷基链长度存在相关性。Bernards (2002)研究表明, 木栓质深色或者黑色条带和酚类物质的含量相关。酚类物质可以通过自发荧光或者样品染色后在显微镜下进行观察检测。但是, 这些检测不能区分木栓质酚类物质、木质素或者细胞壁中的其它酚类成分。此外, 木栓质形成早期或者少量时, 染色技术经常不够灵敏。例如, 对湿地植物Gyceria maxima根的横截面进行自发荧光检测, 结果发现酚类物质已经开始堆积, GC-MS分析也证实了皮下层组织已形成较多的脂肪类化合物。但是, 苏丹红7B或者荧光黄88染料并不能检测到木栓质(Soukup et al., 2007)。与此类似, 在拟南芥种子发育后期, 仅种子合点区域可以被苏丹红染色(Beis- son et al., 2007; Franke et al., 2009)。 在细胞壁内, 木栓质的化学成分及其分布排列因其生理功能或者组织的成熟阶段而异。Machado等(2013)对栓皮栎(Quercus variabilis)次生根的木栓化进行观察, 透射电镜下通过中性红染料染色, 最先在内皮层凯氏带内检测到木栓质堆积。苏丹红7B和荧光黄88染料则可以在凯氏带不被着色的情况下特异性染色片层木栓质。周皮细胞染色结果进一步证实了木栓质片层可被苏丹红7B染色。与最外层形成层细胞相比, 新形成的木栓质片层细胞具有特异的中性红信号。
2.2 木栓质单体间的相互连接为探究木栓质单体间的相互作用, 前人通过多糖水解酶降解典型的木栓化组织或富含木栓质的细胞壁, 将分离下来的低分子量化合物进行色谱和质谱鉴定。之后, 将被鉴定的物质在原位进行重组。然而, 由于不同化合物及其分子间连接的复杂性, 这种方式并不可行(Thomas et al., 2007)。也有研究者尝试利用碱水解和酯基转移作用清除脂肪类物质间的连接, 释放出脂肪酸衍生物, 让其仅和酚酸相连, 以此探究木栓质单体间的连接。但是, 这种适用于木质素的降解方法并不适用于检测木栓质单体间的作用。多酚类物质中, 醚的形成和C-C连接的可能性使其它类似处理也并未取得成功(Graça and Pereira, 2000)。对马铃薯损伤周皮进行13C固态核磁共振光谱分析(NMR), 试图揭示木栓质多聚物中羧基功能的化学转变, 然而仅仅得到了追踪信号, 并未得到详细的功能变换数据(Bern- ards et al., 1995; Yan and Stark, 2000)。 虽然对木栓质的组成仍不明晰, 但是通过其超微结构和已明确的单体物质, 可进行木栓质低聚物分析。Graça和Santos (2007)应用Ca(OH)2-甲醇分解法溶解了栓皮栎周皮大约10%的木栓质, 使用点喷射电离串联质谱对这些物质的结构进行测定, 鉴定的结构物包括ω-羟基脂肪酸-甘油二聚物、α, ω-二羧酸-甘油二聚物、ω-羟基脂肪酸-阿魏酰酯以及连接2个α, ω-二羧酸的甘油酯和ω-羟基脂肪酸-阿魏酰甘油酯, 成功呈现了木栓质中甘油-酰基和阿魏酸的连接。在马铃薯周皮中, 通过固态核磁共振方法也确定了这些组分与它们之间的相互连接, 且长链脂肪类物质、多酚类物质和细胞壁多糖彼此共价连接(Stark et al., 1994; Yan and Stark, 2000)。据Bernards (2002)研究推测, 酚类物质先与初生细胞壁连接, 再与多聚脂肪类物质连接。其中, 苯丙烷代谢途径(PPP)的中间产物阿魏酸在建立和交联芳香类物质过程中可与脂肪区域相连。结合以上鉴定到的低聚物单元, 以及它们的功能和透射电镜下的片层结构, Graça和Santos (2007)得出了以两末端羧基分别连接甘油的长链α, ω-二羧酸为支柱骨架的木栓质大分子结构新模式: 这些支柱骨架单元堆积后的分子大小与透射电镜观察到的片层结构大小相符; 多芳香类物质含量与不透明片层厚度相关, 覆盖了聚酯区域; 酯化成酰基单元的阿魏酰连接聚酯和多芳香类。 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997)。与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009)。以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009)。Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构。然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致。例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009)。以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程。此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010)。虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究。
3 木栓质合成的酶学研究目前已经确定的参与木栓质合成的酶包括细胞色素P450单加氧酶、氧化还原酶、脂肪酸延长酶和过氧化物酶。它们分别参与脂肪酸ω-羟基化、二酸形成、脂肪酸延长和芳香类化合物聚合, 其活性可在多种植物组织的聚酯形成过程中检测到(Agrawal and Kola- ttukudy, 1977; Razem and Bernards, 2003; Schrei- ber et al., 2005c)。 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加。相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012)。苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008)。块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002)。Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum)。Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性。相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高。因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证。此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003)。然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释。
4 木栓质合成的基因组学研究木栓化不仅受空间限制, 也受到环境和自身发育的调控。因此, 木栓质的生物合成与堆积在细胞和组织水平上受到严格调控。拟南芥木栓质的化学特性研究已经深入到基因层面, 如具有脂肪酸延长作用的β-酮酯酰-CoA合成酶基因(KCS)和脂肪酸羟基化作用的细胞色素P450加氧酶基因(CYP)已被鉴定(Franke et al., 2012)。 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制。然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005)。作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007)。Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定。例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因。在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达。在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性。在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性。在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010)。这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控。 4.1 CYP86亚家族细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011)。P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006)。Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化。复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程。融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008)。与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008)。与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b)。 在拟南芥根的内皮层中, CYP86B1的表达也具有单体特异性, 即在ralph突变体中, CYP86B1的缺失对木栓质成分产生显著影响(Compagnon et al., 2009)。在此突变体的根和种皮的木栓质中, 超长链ω-羟基酸和α, ω-二羧酸, 尤其链长为C22和C24的羧酸几乎完全缺失(Compagnon et al., 2009; Molina et al., 2009)。在拟南芥RNAi株系中, CYP86B1表达水平的局部降低也导致这些特殊长链羧酸含量减少, 这再次证明了CYP86B1是生成木栓质ω-羟化脂肪酸(链长≥C22)所必需的酶。虽然该酶的具体功能仍未确定, 但这些研究均表明CYP86B1编码一种超长链脂肪酸ω-羟基化酶。此外, 有研究表明超长链氧化脂肪酸的损耗可以通过增加未被氧化的C22和C24脂肪酸来补充。即在cyp86b1/ralph突变体的木栓质多聚酯中, 双官能团脂肪酸可以被单官能团脂肪酸所代替。然而, Molina等(2009)认为, 单官能团的单体很大一部分并不延伸或者交联其它脂肪族多聚物, 因此单官能团脂肪酸会使酯多聚物链终止。所以, 还未被鉴定的木栓质成分对准确描述木栓质的结构非常重要。
4.2 GPAT5基因与CYP基因相似, GPAT5是木栓化过程中首批被鉴定的基因之一, 主要在根的内皮层组织中表达(Beis- son et al., 2007)。对突变体gpat5的研究表明, 其编码的甘油-3-磷酸酰基转移酶(GPAT5)参与根和种皮木栓质中C22与C24超长链酰基单体的形成。在gpat5突变体根中, C22/C24单酰基甘油和超长链烷羟基肉桂酸酯的含量减少, 说明GPAT5在脂类聚合物的生物合成中发挥关键作用。与贮存型脂肪生物合成中GPATs的sn-1酰基化作用不同, 在木栓化过程中, GPAT5催化甘油的sn-2酰基化(Yang et al., 2010)。与相关蛋白质的作用特性相比较, 仅在木栓质合成中发现了GPATs的sn-2酰基化作用。动物、微生物和藻类也缺失具有sn-2酰基化作用的GPATs, 这可能与陆地植物的进化过程相关(Yang et al., 2010)。
4.3 KCS基因角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008)。因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE。KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008)。然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定。在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008)。此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006)。对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009)。同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程。 在拟南芥基因(KCS2)敲除突变体(daisy)的根木栓质中, C22和C24超长链单体含量显著减少, KCS2的表达位置与模式也与木栓质的堆积和功能相一致(Lee et al., 2009; Franke et al., 2009)。然而, 在DAISY/KCS2突变体的其它组织中, C22和C24或者中间链长的脂肪酸仍出现在其它脂类结构(鞘脂类和蜡质)中, 表明促进木栓化可能并不是KCS2的唯一功能。双突变分析进一步证实了KCS2与KCS20在木 栓质和蜡质生物合成中具有双重作用(Lee et al., 2009)。 StKCS6在木栓化的马铃薯块茎周皮中高表达(Serra et al., 2009a)。若下调StKCS6的表达丰度, 链长为C28和更长链脂肪酸的含量下降50%-95%, 而链长C26和更短链的化合物却发生堆积, 进一步证明StKCS6对脂肪酸链的延长有特定作用(Serra et al., 2009a)。据此推测, StKCS6参与了木栓质前体脂肪酸链的延长。
5 木栓质前体物质的输出和装配多种木栓质前体物质, 无论它们是以单体还是部分形成低聚物的形式, 都需要从内质网输出并穿过细胞质膜, 然后聚集形成木栓质。虽然木栓质中多种脂肪类物质的合成已经研究得比较透彻, 但是它们的运输和聚合机制仍不清楚。 木栓质直接前体物质的分子属性、分泌和装配过程对木栓质的形成非常重要。最近, DeBolt等(2009)发现, 拟南芥UDP-葡萄糖-甾醇糖基转移酶基因的突变体(ugt80B1)种脐区缺乏自发荧光。糖基转移酶催化植物次生代谢产物的合成, 是植物次生代谢过程中的重要部分。UDP-葡萄糖-甾醇糖基转移酶催化UDP葡萄糖的糖基转移到甾醇, 形成甾醇苷类物质。化学分析揭示了UDP-葡萄糖-甾醇糖基转移酶在木栓化过程中的作用。在ugtB1和ugtA2双突变的种子中, 除了C24 α, ω-二羧酸, 大部分聚酯单体含量显著降低。通过透射电镜观察, 发现ugtA2/ugtB1外种皮缺失高电子密度层, 而细胞质内却出现高电子密度体, 说明脂肪聚酯前体物质在细胞质内积累。由此推测, 甾醇苷类和丰富的等离子膜组分对于脂类聚酯前体输出或者运输到质外体是必需的。 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究。这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013)。质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输。例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014)。最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014)。此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010)。ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索。但是, 木栓质在形成过程中其成分是不断变化的。在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制。与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出。因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用。 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知。近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012)。与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007)。一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006)。 综合以上关于木栓质合成的酶学、基因组学及其前体物质输出方面的研究, 我们将木栓质的合成及其运输过程归纳为图2。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-358/img_2.png<b>图2</b> 木栓质合成及其运输至细胞壁的路径概述<br/>脂肪酸合成发生在细胞质内, 之后被脂肪酸-CoA连接酶(FACL)和长链酰基-CoA合成酶(LACSs)活化, 生成脂肪酰CoA类物质, 然后被一系列木栓质合成酶修饰。脂肪酸延长酶(FAE)复合体调控脂肪酰延长, 产生超长链脂肪酸(VLCFAs); 脂肪酰还原酶(FARs)还原酰基, 生成脂肪醇类和α, ω-二醇; 细胞色素P450酶类(CYPs)氧化脂肪酰, 生成ω-羟基脂肪酸(ω-OHs)和α, ω-二羧酸(DCAs); ω-OHs可被细胞色素P450s进一步氧化成DCAs; ω-OHs和DCAs被甘油-3-磷酸酰基转移酶(GPATs)酯化成<i>sn</i>-2单酰基甘油酯。ATP-结合盒(ABC)转运蛋白参与木栓质单体穿过质膜的运输。多酯合酶(PS)可能用其它木栓质单体延伸<i>sn</i>-2单酰基甘油, 使所有成分聚集, 产生高分子质量聚酯。在苯丙氨酸解氨酶(PAL)等酶作用下, 苯丙烷代谢途径生成香豆酸、咖啡酸和阿魏酸, 它们在酰基连接酶等作用下, 再生成阿魏酰-CoA。阿魏酰-CoA在脂肪木栓质阿魏酰转移酶(ASFT)作用下, 与脂肪醇、α, ω-二醇和单酰基甘油酯连接, 再生成相应的酯类物质。<br/><b>Figure 2</b> Overview of the suberin biosynthetic pathway with subsequent transport to the cell wall<br/>Fatty acids are synthesed in the cytoplasm, activated into fatty acyl-CoAs by fatty acid-CoA ligase (FACL) and long chain acyl-CoA synthetases (LACSs), and then modified by series suberin biosynthetic enzymes. Fatty acyl elongation is controlled via the fatty acid elongation (FAE) complex producing very long chain fatty acids (VLCFAs); acyl reduction by fatty acyl reductases (FARs) producing fatty alcohol and α, ω-diols; fatty acyl oxidation by cytochrome P450 enzymes (CYPs) producing ω-hydroxy fatty acids (ω-OHs) and α, ω-dicarboxylic acids (DCAs); ω-OHs could be also further oxidized to DCAs by P450s; and esterification of ω-OHs and DCAs by glycerol 3-phosphate acyltransferases (GPATs) producing <i>sn</i>-2 monoacylglycerols. ATP-binding- cassette (ABC) transporters are involved in transport of suberin monomers across the plasma membrane. Polyester synthase(s) (PS) may extend <i>sn</i>-2 monoacylglycerols with other suberin monomers to eventually gather into high molecular weight polyesters. With the catalytic action of phenylalnine ammonialyase (PAL) and other related enzymes, phenylalanine metabolic pathways provide coumaric, caffeic, and ferulic acids, which then are transformed to feruloyl-CoA by acyltransferase. Moreover, feruloyl-CoA is linked with fatty alcohols, α, ω-diols and monoacylglycerols through aliphatic suberin feruloyl transferase (ASFT), to finally produce esters. Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-358/img_2.png<b>图2</b> 木栓质合成及其运输至细胞壁的路径概述<br/>脂肪酸合成发生在细胞质内, 之后被脂肪酸-CoA连接酶(FACL)和长链酰基-CoA合成酶(LACSs)活化, 生成脂肪酰CoA类物质, 然后被一系列木栓质合成酶修饰。脂肪酸延长酶(FAE)复合体调控脂肪酰延长, 产生超长链脂肪酸(VLCFAs); 脂肪酰还原酶(FARs)还原酰基, 生成脂肪醇类和α, ω-二醇; 细胞色素P450酶类(CYPs)氧化脂肪酰, 生成ω-羟基脂肪酸(ω-OHs)和α, ω-二羧酸(DCAs); ω-OHs可被细胞色素P450s进一步氧化成DCAs; ω-OHs和DCAs被甘油-3-磷酸酰基转移酶(GPATs)酯化成<i>sn</i>-2单酰基甘油酯。ATP-结合盒(ABC)转运蛋白参与木栓质单体穿过质膜的运输。多酯合酶(PS)可能用其它木栓质单体延伸<i>sn</i>-2单酰基甘油, 使所有成分聚集, 产生高分子质量聚酯。在苯丙氨酸解氨酶(PAL)等酶作用下, 苯丙烷代谢途径生成香豆酸、咖啡酸和阿魏酸, 它们在酰基连接酶等作用下, 再生成阿魏酰-CoA。阿魏酰-CoA在脂肪木栓质阿魏酰转移酶(ASFT)作用下, 与脂肪醇、α, ω-二醇和单酰基甘油酯连接, 再生成相应的酯类物质。<br/><b>Figure 2</b> Overview of the suberin biosynthetic pathway with subsequent transport to the cell wall<br/>Fatty acids are synthesed in the cytoplasm, activated into fatty acyl-CoAs by fatty acid-CoA ligase (FACL) and long chain acyl-CoA synthetases (LACSs), and then modified by series suberin biosynthetic enzymes. Fatty acyl elongation is controlled via the fatty acid elongation (FAE) complex producing very long chain fatty acids (VLCFAs); acyl reduction by fatty acyl reductases (FARs) producing fatty alcohol and α, ω-diols; fatty acyl oxidation by cytochrome P450 enzymes (CYPs) producing ω-hydroxy fatty acids (ω-OHs) and α, ω-dicarboxylic acids (DCAs); ω-OHs could be also further oxidized to DCAs by P450s; and esterification of ω-OHs and DCAs by glycerol 3-phosphate acyltransferases (GPATs) producing <i>sn</i>-2 monoacylglycerols. ATP-binding- cassette (ABC) transporters are involved in transport of suberin monomers across the plasma membrane. Polyester synthase(s) (PS) may extend <i>sn</i>-2 monoacylglycerols with other suberin monomers to eventually gather into high molecular weight polyesters. With the catalytic action of phenylalnine ammonialyase (PAL) and other related enzymes, phenylalanine metabolic pathways provide coumaric, caffeic, and ferulic acids, which then are transformed to feruloyl-CoA by acyltransferase. Moreover, feruloyl-CoA is linked with fatty alcohols, α, ω-diols and monoacylglycerols through aliphatic suberin feruloyl transferase (ASFT), to finally produce esters.
图2 木栓质合成及其运输至细胞壁的路径概述 脂肪酸合成发生在细胞质内, 之后被脂肪酸-CoA连接酶(FACL)和长链酰基-CoA合成酶(LACSs)活化, 生成脂肪酰CoA类物质, 然后被一系列木栓质合成酶修饰。脂肪酸延长酶(FAE)复合体调控脂肪酰延长, 产生超长链脂肪酸(VLCFAs); 脂肪酰还原酶(FARs)还原酰基, 生成脂肪醇类和α, ω-二醇; 细胞色素P450酶类(CYPs)氧化脂肪酰, 生成ω-羟基脂肪酸(ω-OHs)和α, ω-二羧酸(DCAs); ω-OHs可被细胞色素P450s进一步氧化成DCAs; ω-OHs和DCAs被甘油-3-磷酸酰基转移酶(GPATs)酯化成sn-2单酰基甘油酯。ATP-结合盒(ABC)转运蛋白参与木栓质单体穿过质膜的运输。多酯合酶(PS)可能用其它木栓质单体延伸sn-2单酰基甘油, 使所有成分聚集, 产生高分子质量聚酯。在苯丙氨酸解氨酶(PAL)等酶作用下, 苯丙烷代谢途径生成香豆酸、咖啡酸和阿魏酸, 它们在酰基连接酶等作用下, 再生成阿魏酰-CoA。阿魏酰-CoA在脂肪木栓质阿魏酰转移酶(ASFT)作用下, 与脂肪醇、α, ω-二醇和单酰基甘油酯连接, 再生成相应的酯类物质。 Figure 2 Overview of the suberin biosynthetic pathway with subsequent transport to the cell wall Fatty acids are synthesed in the cytoplasm, activated into fatty acyl-CoAs by fatty acid-CoA ligase (FACL) and long chain acyl-CoA synthetases (LACSs), and then modified by series suberin biosynthetic enzymes. Fatty acyl elongation is controlled via the fatty acid elongation (FAE) complex producing very long chain fatty acids (VLCFAs); acyl reduction by fatty acyl reductases (FARs) producing fatty alcohol and α, ω-diols; fatty acyl oxidation by cytochrome P450 enzymes (CYPs) producing ω-hydroxy fatty acids (ω-OHs) and α, ω-dicarboxylic acids (DCAs); ω-OHs could be also further oxidized to DCAs by P450s; and esterification of ω-OHs and DCAs by glycerol 3-phosphate acyltransferases (GPATs) producing sn-2 monoacylglycerols. ATP-binding- cassette (ABC) transporters are involved in transport of suberin monomers across the plasma membrane. Polyester synthase(s) (PS) may extend sn-2 monoacylglycerols with other suberin monomers to eventually gather into high molecular weight polyesters. With the catalytic action of phenylalnine ammonialyase (PAL) and other related enzymes, phenylalanine metabolic pathways provide coumaric, caffeic, and ferulic acids, which then are transformed to feruloyl-CoA by acyltransferase. Moreover, feruloyl-CoA is linked with fatty alcohols, α, ω-diols and monoacylglycerols through aliphatic suberin feruloyl transferase (ASFT), to finally produce esters.
6 脱落酸对木栓质合成的影响20世纪90年代, 研究者普遍采用显微观察技术研究木栓化组织的结构及其复杂成分(Schreiber et al., 1999)。之后, 结合GC-MS等技术, 不同物种、不同组织和不同环境下的特异木栓质可以被定量和定性描述(Zimmermann et al., 2000; Schreiber et al., 2005b)。随着分析技术的成熟, 影响木栓质形成和堆积的因素逐渐清晰, 包括各种环境胁迫、机械损伤和激素处理。在环境胁迫如盐胁迫(Krishnamurthy et al., 2009)或缺氧(Kotula et al., 2009)条件下, 水稻(Oryza sativa)根外皮层的凯氏带和片层木栓质的堆积量显著增加。目前, 激素对木栓质形成和堆积的影响, 尤其是脱落酸(ABA)的作用, 逐渐成为研究热点。 马铃薯块茎在采收、处理和种切过程中会发生损伤, 伤口的快速愈合对于避免病原菌侵染等非常重要。Lulai和Suttle (2004)证明, 虽然乙烯参与了块茎的损伤反应, 但是乙烯并不直接影响损伤诱导木栓质的合成。ABA调控植物对干旱和盐胁迫的应答(Himm- elbach et al., 2003), 且ABA与水通道蛋白基因的表达相关, 因此推测其具有防止细胞干燥等保护块茎的作用(Efetova et al., 2007)。 Lulai等(2008)测定了ABA在损伤诱导的木栓化调控和块茎水分损失中的作用。液质联用色谱分析显示, 整个块茎中ABA总含量基本不变, 但是在损伤周皮附近含量较高, 在中心部位最低。块茎损伤诱导了ABA分布发生变化。损伤后, 在愈合组织块中ABA含量先降低, 24小时达到最低, 损伤后第3-7天ABA含量逐渐增加。这表明内源ABA参与损伤诱导木栓化的形成, 避免由于脱水而致细胞死亡。 Kumar等(2010)研究表明, 马铃薯块茎的损伤愈合能力受年龄和ABA调控。马铃薯块茎损伤诱导木栓化近封闭层和损伤周皮形成, 可抗干燥和微生物的侵染。在损伤应答过程中, ABA和苯丙氨酸解氨酶的含量及活性均增加, 是木栓化近封闭层和损伤周皮形成的关键过程。随着块茎年龄的增加, 损伤愈合能力下降。未损伤的且年龄较高的块茎中, ABA含量比贮藏期短的新块茎低86%。在伤后24小时之内, 新块茎的PAL转录活性提高, 但在较老块茎中则延后5天。对老块茎进行ABA处理, 可提高其PAL活性, 强化损伤处对水分蒸发的阻碍能力。在贮藏时间较长的老块茎中, 损伤诱导的木栓质酚类和脂肪类物质含量均较低。而ABA处理可以通过增加木栓质酚类物质(SPP)的积累, 部分恢复老块茎的愈合能力。 ABA也调控番茄果实茎疤组织中木栓质的形成(Leide et al., 2012)。新采收的樱桃番茄果实在茎疤处存在损伤, 3天内迅速愈合。对新合成的茎疤组织进行化学成分分析, 发现芳香类和脂肪类物质在此处聚合, 形成木栓质聚酯。茎疤区域的ABA含量分析结果显示, 在损伤愈合期间, 茎疤组织中ABA的含量明显增加。然而, ABA缺陷突变体中茎疤损伤处的木栓化过程被推迟或者木栓质含量较低。Tao等(2016)也证明了外源ABA参与以木栓质为基础的番茄损伤愈合过程。
BaxterI, HosmaniPS, RusA, LahnerB, BorevitzJO, MuthukumarB, MickelbartMV, SchreiberL, FrankeRB, SaltDE (2009). Root suberin forms an extracellular barrier that affects water relations and mineral nutrition in Arabidopsis.PLoS Genet 5, e1000492. [本文引用: 1]
[6]
BeissonF, LiY, BonaventureG, PollardM, OhlroggeJB (2007). The acyltransferase GPAT5 is required for the synthesis of suberin in seed coat and root of Arabidopsis.Plant Cell 19, 351-368. [本文引用: 4]
[7]
BeissonF, Li-BeissonY, PollardM (2012). Solving the puzzles of cutin and suberin polymer biosynthesis.Curr Opin Plant Biol 15, 329-337. [本文引用: 1]
[8]
BenvenisteI, TijetN, AdasF, PhilippsG, SalaünJP, DurstF (1998). CYP86A1 from Arabidopsis thaliana encodes a cytochrome P450-dependent fatty acid omega- hydroxylase. Biochem Biophys Res Commun 243, 688-693. [本文引用: 1]
BrundrettMC, EnstoneDE, PetersonCA (1988). A berberine-aniline blue fluorescent staining procedure for suberin, lignin, and callose in plant tissue.Protoplasma 146, 133-142. [本文引用: 1]
[17]
BrundrettMC, KendrickB, PetersonCA (1991). Efficient lipid staining in plant material with Sudan Red 7B or Fluo- ral Yellow 088 in polyethylene glycol-glycerol.Biotech Hi- stochem 66, 111-116. [本文引用: 1]
[18]
ChoiH, JinJY, ChoiS, HwangJU, KimYY, SuhMC, LeeY (2011). An ABCG/WBC-type ABC transporter is ess- ential for transport of sporopollenin precursors for exine formation in developing pollen.Plant J 65, 181-193. [本文引用: 1]
[19]
ClarksonD, RobardsA, StephensJ, StarkM (1987). Suberin lamellae in the hypodermis of maize (Zea mays) roots, development and factors affecting the permeability of hypodermal layers. Plant Cell Environ 10, 83-93. [本文引用: 1]
[20]
CompagnonV, DiehlP, BenvenisteI, MeyerD, SchallerH, SchreiberL, FrankeR, PinotF (2009). CYP86B1 is required for very long chain ω-hydroxyacid and α, ω- dicarboxylic acid synthesis in root and seed suberin polyester.Plant Physiol 150, 1831-1843. [本文引用: 4]
[21]
D’AuriaJC (2006). Acyltransferases in plants: a good time to be BAHD.Curr Opin Plant Biol 9, 331-340. [本文引用: 1]
[22]
De SimoneO, HaaseK, MüllerE, JunkWJ, HartmannK, SchreiberL, SchmidtW (2003). Apoplasmic barriers and oxygen transport properties of hypodermal cell walls in roots from four Amazonian tree species.Plant Physiol 132, 206-217. [本文引用: 1]
[23]
DeBoltS, ScheibleWR, SchrickK, AuerM, BeissonF, BischoffV, Bouvier-NavéP, CarrollA, HematyK, LiY (2009). Mutations in UDP-glucose: sterol glucosyltrans- ferase in Arabidopsis cause transparent testa phenotype and suberization defect in seeds.Plant Physiol 151, 78-87. [本文引用: 1]
[24]
DomergueF, VishwanathSJ, JoubèsJ, OnoJ, LeeJA, BourdonM, AlhattabR, LoweC, PascalS, LessireR (2010). Three Arabidopsis fatty acyl-coenzyme A reduc- tases, FAR1, FAR4, and FAR5, generate primary fatty al- cohols associated with suberin deposition.Plant Physiol 153, 1539-1554. [本文引用: 2]
[25]
EfetovaM, ZeierJ, RiedererM, LeeCW, StinglN, MuellerM, HartungW, HedrichR, DeekenR (2007). A central role of abscisic acid in drought stress protection of Agro- bacterium-induced tumors on Arabidopsis. Plant Physiol 145, 853-862. [本文引用: 1]
[26]
EnstoneDE, PetersonCA, MaF (2003). Root endodermis and exodermis: structure, function, and responses to the environment.J Plant Growth Regul 21, 335-351. [本文引用: 2]
[27]
FrankeR, BriesenI, WojciechowskiT, FaustA, Yephr- emovA, NawrathC, SchreiberL (2005). Apoplastic polyesters in Arabidopsis surface tissues—a typical sub- erin and a particular cutin.Phytochemistry 66, 2643-2658. [本文引用: 4]
[28]
FrankeR, HöferR, BriesenI, EmsermannM, EfremovaN, YephremovA, SchreiberL (2009). The DAISY gene from Arabidopsis encodes a fatty acid elongase con- densing enzyme involved in the biosynthesis of aliphatic suberin in roots and the chalaza-micropyle region of seeds. Plant J 57, 80-95. [本文引用: 5]
FrankeRB, DombrinkI, SchreiberL (2012). Suberin goes genomics: use of a short living plant to investigate a long lasting polymer.Front Plant Sci 3, 1-8. [本文引用: 3]
[31]
GandiniA (2008). Polymers from renewable resources: a challenge for the future of macromolecular materials.Ma- cromolecules 41, 9491-9504. [本文引用: 1]
[32]
GarthwaiteAJ, ArmstrongW, ColmerTD (2008). Assess- ment of O2 diffusivity across the barrier to radial O2 loss in adventitious roots of Hordeum marinum. New Phytol 179, 405-416. [本文引用: 1]
[33]
GirardAL, MounetF, Lemaire-ChamleyM, GaillardC, ElmorjaniK, VivancosJ, RunavotJL, QuemenerB, PetitJ, GermainV (2012). Tomato GDSL1 is required for cutin deposition in the fruit cuticle.Plant Cell 24, 3119-3134. [本文引用: 1]
[34]
GouJY, YuXH, LiuCJ (2009). A hydroxycinnamoyltransfe- rase responsible for synthesizing suberin aromatics in Arabidopsis.Proc Natl Acad Sci USA 106, 18855-18860. [本文引用: 1]
KatoM, HayakawaY, HyodoH, IkomaY, YanoM (2000). Wound-induced ethylene synthesis and expression and formation of 1-aminocyclopropane-1-carboxylate (ACC) synthase, ACC oxidase, phenylalanine ammonia-lyase, and peroxidase in wounded mesocarp tissue of Cucurbita maxima. Plant Cell Physiol 41, 440-447. [本文引用: 1]
[45]
KesanakurtiD, KolattukudyPE, KirtiPB (2012). Fruit- specific overexpression of wound-induced tap1 under E8 promoter in tomato confers resistance to fungal patho- gens at ripening stage.Physiol Plant 146, 136-148.
[46]
KilianJ, WhiteheadD, HorakJ, WankeD, WeinlS, BatisticO, D’AngeloC, Bornberg-BauerE, KudlaJ, HarterK (2007). The AtGenExpress global stress expre- ssion data set: protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses.Plant J 50, 347-363. [本文引用: 2]
[47]
KolattukudyP (1971). Enzymatic synthesis of fatty alcohols in Brassica oleracea. Arch Biochem Biophys 142, 701-709. [本文引用: 1]
[48]
KolattukudyPE (1981). Structure, biosynthesis, and biode- gradation of cutin and suberin.Annu Rev Plant Physiol 32, 539-567.
[49]
KolattukudyPE (2001). Polyesters in higher plants. In: Biopolyesters. Berlin Heidelberg: Springer Press. pp. 1-49. [本文引用: 6]
[50]
KotulaL, RanathungeK, SchreiberL, SteudleE (2009). Functional and chemical comparison of apoplastic barr- iers to radial oxygen loss in roots of rice (Oryza sativa L.) grown in aerated or deoxygenated solution. J Exp Bot 60, 2155-2167. [本文引用: 1]
[51]
KrishnamurthyP, RanathungeK, FrankeR, PrakashH, SchreiberL, MathewM (2009). The role of root apo- plastic transport barriers in salt tolerance of rice (Oryza sativa L.). Planta 230, 119-134. [本文引用: 2]
[52]
KrishnamurthyP, RanathungeK, NayakS, SchreiberL, MathewM (2011). Root apoplastic barriers block Na+ transport to shoots in rice (Oryza sativa L.). J Exp Bot 62, 4215-4228.
[53]
KumarG, KnowlesNR (2003). Wound-induced superoxide production and PAL activity decline with potato tuber age and wound healing ability.Physiol Plant 117, 108-117. [本文引用: 2]
[54]
KumarGM, IyerS, KnowlesNR (2007). Strboh A homolo- gue of NADPH oxidase regulates wound-induced oxida- tive burst and facilitates wound-healing in potato tubers.Planta 227, 25-36. [本文引用: 1]
[55]
KumarGM, LulaiEC, SuttleJC, KnowlesNR (2010). Age-induced loss of wound-healing ability in potato tubers is partly regulated by ABA.Planta 232, 1433-1445. [本文引用: 1]
[56]
KurdyukovS, FaustA, NawrathC, BärS, VoisinD, EfremovaN, FrankeR, SchreiberL, SaedlerH, MétrauxJP (2006). The epidermis-specific extracellular BODYGUARD controls cuticle development and morpho- genesis in Arabidopsis.Plant Cell 18, 321-339. [本文引用: 1]
[57]
LandgrafR, SmolkaU, AltmannS, Eschen-LippoldL, SenningM, SonnewaldS, WeigelB, FrolovaN, Stre- hmelN, HauseG (2014). The ABC transporter ABCG1 is required for suberin formation in potato tuber periderm.Plant Cell 26, 3403-3415. [本文引用: 1]
[58]
LasserreE, JobetE, LlauroC, DelsenyM (2008). AtERF38 (At2g35700), an AP2/ERF family transcription factor gene from Arabidopsis thaliana, is expressed in sp- ecific cell types of roots, stems and seeds that undergo suberization. Plant Physiol Biochem 46, 1051-1061. [本文引用: 1]
[59]
LeeSB, JungSJ, GoYS, KimHU, KimJK, ChoHJ, ParkOK, SuhMC (2009). Two Arabidopsis 3-ketoacyl CoA synthase genes,KCS20 and KCS2/DAISY, are func- tionally redundant in cuticular wax and root suberin bio- synthesis, but differentially controlled by osmotic stress. Plant J 60, 462-475. [本文引用: 5]
[60]
LeideJ, HildebrandtU, HartungW, RiedererM, VoggG (2012). Abscisic acid mediates the formation of a suber- ized stem scar tissue in tomato fruits.New Phytol 194, 402-415. [本文引用: 1]
[61]
LiY, BeissonF, KooAJ, MolinaI, PollardM, OhlroggeJ (2007). Identification of acyltransferases required for cutin biosynthesis and production of cutin with suberin-like mo- nomers.Proc Natl Acad Sci USA 104, 18339-18344. [本文引用: 2]
LotfyS, NegrelJ, JavelleF (1994). Formation of ω-feruloy- loxypalmitic acid by an enzyme from wound-healing pota- to tuber discs.Phytochemistry 35, 1419-1424. [本文引用: 1]
[64]
LüS, SongT, KosmaDK, ParsonsEP, RowlandO, JenksMA (2009). Arabidopsis CER8 encodes LONG-CHAIN ACYL-COA SYNTHETASE 1 (LACS1) that has overlap- ping functions with LACS2 in plant wax and cutin synthe- sis. Plant J 59, 553-564. [本文引用: 2]
[65]
LulaiEC, CorsiniDL (1998). Differential deposition of suberin phenolic and aliphatic domains and their roles in resistance to infection during potato tuber (Solanum tube- rosum L.) wound-healing. Physiol Mol Plant Pathol 53, 209-222. [本文引用: 2]
[66]
LulaiEC, SuttleJC (2004). The involvement of ethylene in wound-induced suberization of potato tuber (Solanum tu- berosum L.): a critical assessment. Postharvest Biol Te- chnol 34, 105-112.
[67]
LulaiEC, SuttleJC, PedersonSM (2008). Regulatory involvement of abscisic acid in potato tuber wound-hea- ling.J Exp Bot 59, 1175-1186. [本文引用: 3]
[68]
LuxA, ŠottníkováA, OpatrnaJ, GregerM (2004). Differ- ences in structure of adventitious roots in Salix clones with contrasting characteristics of cadmium accumulation and sensitivity. Physiol Plant 120, 537-545. [本文引用: 1]
[69]
MachadoA, PereiraH, TeixeiraRT (2013). Anatomy and development of the endodermis and phellem of Quercus suber L. roots. Microsc Microanal 19, 525-534. [本文引用: 1]
[70]
MatsudaF, MorinoK, MiyashitaM, MiyagawaH (2003). Metabolic flux analysis of the phenylpropanoid pathway in wound-healing potato tuber tissue using stable isotope- labeled tracer and LC-MS spectroscopy.Plant Cell Phy- siol 44, 510-517. [本文引用: 1]
[71]
MetzJG, PollardMR, AndersonL, HayesTR, LassnerMW (2000). Purification of a jojoba embryo fatty acyl- coenzyme A reductase and expression of its cDNA in high erucic acid rapeseed.Plant Physiol 122, 635-644. [本文引用: 1]
[72]
MeyerCJ, SeagoJL, PetersonCA (2009). Environmental effects on the maturation of the endodermis and multi- seriate exodermis of Iris germanica roots. Ann Bot 103, 687-702. [本文引用: 1]
[73]
MitsudaN, SekiM, ShinozakiK, Ohme-TakagiM (2005). The NAC transcription factors NST1 and NST2 of Arabi- dopsis regulate secondary wall thickenings and are required for anther dehiscence.Plant Cell 17, 2993-3006. [本文引用: 1]
[74]
MolinaI, BonaventureG, OhlroggeJ, PollardM (2006). The lipid polyester composition of Arabidopsis thaliana and Brassica napus seeds. Phytochemistry 67, 2597-2610. [本文引用: 1]
[75]
MolinaI, Li-BeissonY, BeissonF, OhlroggeJB, PollardM (2009). Identification of an Arabidopsis feruloyl-coen- zyme A transferase required for suberin synthesis.Plant Physiol 151, 1317-1328. [本文引用: 5]
[76]
ObayashiT, HayashiS, SaekiM, OhtaH, KinoshitaK (2009). ATTED-II provides coexpressed gene networks for Arabidopsis.Nucleic Acids Res 37, D987-D991. [本文引用: 1]
[77]
ObayashiT, KinoshitaK, NakaiK, ShibaokaM, HayashiS, SaekiM, ShibataD, SaitoK, OhtaH (2007). AT- TED-II: a database of co-expressed genes and cis elements for identifying co-regulated gene groups in Arabidopsis. Nucleic Acids Res 35, D863-D869. [本文引用: 1]
[78]
OlsonDA, ShearesVV (2006). Preparation of unsaturated linear aliphatic polyesters using condensation polymeri- zation.Macromolecules 39, 2808-2814. [本文引用: 1]
[79]
OlssonA, LindströmM, IversenT (2007). Lipase-cataly- zed synthesis of an epoxy-functionalized polyester from the suberin monomer cis-9, 10-epoxy-18-hydroxyoctade- canoic acid. Biomacromolecules 8, 757-760. [本文引用: 1]
[80]
PanikashviliD, Savaldi-GoldsteinS, MandelT, YifharT, FrankeRB, HöferR, SchreiberL, ChoryJ, AharoniA (2007). The Arabidopsis DESPERADO/AtWBC11 trans- porter is required for cutin and wax secretion.Plant Phy- siol 145, 1345-1360. [本文引用: 1]
[81]
PanikashviliD, ShiJX, BocobzaS, FrankeRB, SchreiberL, AharoniA (2010). The Arabidopsis DSO/ABCG11 transporter affects cutin metabolism in reproductive or- gans and suberin in roots.Mol Plant 3, 563-575. [本文引用: 1]
[82]
PaulS, GableK, BeaudoinF, CahoonE, JaworskiJ, NapierJA, DunnTM (2006). Members of the Arabidopsis FAE1-like 3-ketoacyl-CoA synthase gene family substitute for the Elop proteins of Saccharomyces cerevisiae. J Biol Chem 281, 9018-9029. [本文引用: 1]
RaffaeleS, VailleauF, LégerA, JoubèsJ, MierschO, HuardC, BléeE, MongrandS, DomergueF, RobyD (2008). A MYB transcription factor regulates very-long- chain fatty acid biosynthesis for activation of the hyper- sensitive cell death response in Arabidopsis.Plant Cell 20, 752-767. [本文引用: 1]
[89]
RanathungeK, SchreiberL (2011). Water and solute per- meabilities of Arabidopsis roots in relation to the amount and composition of aliphatic suberin.J Exp Bot 62, 1961-1974. [本文引用: 1]
[90]
RanathungeK, SchreiberL, FrankeR (2011). Suberin research in the genomics era—new interest for an old po- lymer.Plant Sci 180, 399-413. [本文引用: 1]
[91]
RanathungeK, SteudleE, LafitteR (2005). Blockage of apoplastic bypass-flow of water in rice roots by insoluble salt precipitates analogous to a Pfeffer cell.Plant Cell En- viron 28, 121-133. [本文引用: 2]
[92]
RazemFA, BernardsMA (2003). Reactive oxygen species production in association with suberization: evidence for an NADPH-dependent oxidase.J Exp Bot 54, 935-941. [本文引用: 2]
[93]
RowlandO, DomergueF (2012). Plant fatty acyl reduc- tases: enzymes generating fatty alcohols for protective layers with potential for industrial applications.Plant Sci 193, 28-38. [本文引用: 2]
[94]
RowlandO, ZhengH, HepworthSR, LamP, JetterR, KunstL (2006). CER4 encodes an alcohol-forming fatty acyl-coenzyme A reductase involved in cuticular wax production in Arabidopsis.Plant Physiol 142, 866-877. [本文引用: 1]
[95]
SchmutzA, BuchalaAJ, RyserU (1996). Changing the dimensions of suberin lamellae of green cotton fibers with a specific inhibitor of the endoplasmic reticulum-asso- ciated fatty acid elongases.Plant Physiol 110, 403-411. [本文引用: 1]
[96]
SchnurrJ, ShockeyJ (2004). The acyl-CoA synthetase encoded by LACS2 is essential for normal cuticle deve- lopment in Arabidopsis. Plant Cell 16, 629-642. [本文引用: 1]
[97]
SchnurrJA, ShockeyJM, de BoerGJ (2002). Fatty acid export from the chloroplast. Molecular characterization of a major plastidial acyl-coenzyme A synthetase from Arab- idopsis.Plant Physiol 129, 1700-1709. [本文引用: 1]
[98]
SchreiberL (2010). Transport barriers made of cutin, suberin and associated waxes.Trends Plant Sci 15, 546-553. [本文引用: 1]
[99]
SchreiberL, FrankeR, HartmannK (2005a). Wax and suberin development of native and wound periderm of potato (Solanum tuberosum L.) and its relation to peri- dermal transpiration. Planta 220, 520-530. [本文引用: 2]
[100]
SchreiberL, FrankeR, HartmannKD, RanathungeK, SteudleE (2005b). The chemical composition of suberin in apoplastic barriers affects radial hydraulic conductivity differently in the roots of rice (Oryza sativa L. cv. IR64) and corn(Zea mays L. cv. Helix). J Exp Bot 56, 1427-1436. [本文引用: 2]
[101]
SchreiberL, FrankeR, LessireR (2005c). Biochemical characterization of elongase activity in corn (Zea mays L.) roots. Phytochemistry 66, 131-138. [本文引用: 1]
[102]
SchreiberL, HartmannK, SkrabsM, ZeierJ (1999). Apoplastic barriers in roots: chemical composition of end- odermal and hypodermal cell walls.J Exp Bot 50, 1267-1280. [本文引用: 1]
[103]
SchulerMA, DuanH, BilginM, AliS (2006). Arabidopsis cytochrome P450s through the looking glass: a window on plant biochemistry.Phytochem Rev 5, 205-237. [本文引用: 1]
[104]
SerraO, HohnC, FrankeR, PratS, MolinasM, FiguerasM (2010). A feruloyl transferase involved in the bio- synthesis of suberin and suberin-associated wax is re- quired for maturation and sealing properties of potato periderm.Plant J 62, 277-290. [本文引用: 2]
[105]
SerraO, SolerM, HohnC, FrankeR, SchreiberL, PratS, MolinasM, FiguerasM (2009a). Silencing of StKCS6 in potato periderm leads to reduced chain lengths of suberin and wax compounds and increased peridermal transpir- ation. J Exp Bot 60, 697-707. [本文引用: 2]
[106]
SerraO, SolerM, HohnC, SauveplaneV, PinotF, FrankeR, SchreiberL, PratS, MolinasM, FiguerasM (2009b).CYP86A33-targeted gene silencing in potato tuber alters suberin composition, distorts suberin lamellae, and imp- airs the periderm’s water barrier function. Plant Physiol 149, 1050-1060. [本文引用: 2]
[107]
ShockeyJM, FuldaMS (2003). Arabidopsis contains a large superfamily of acyl-activating enzymes. Phylogen- etic and biochemical analysis reveals a new class of acyl-coenzyme A synthetases.Plant Physiol 132, 1065-1076. [本文引用: 1]
[108]
SolerM, SerraO, MolinasM, García-BerthouE, CaritatA, FiguerasM (2008). Seasonal variation in transcript abun- dance in cork tissue analyzed by real time RT-PCR.Tree Physiol 28, 743-751.
[109]
SolerM, SerraO, MolinasM, HuguetG, FluchS, Figu- erasM (2007). A genomic approach to suberin biosynthe- sis and cork differentiation.Plant Physiol 144, 419-431. [本文引用: 2]
[110]
SoukupA, ArmstrongW, SchreiberL, FrankeR, VotrubováO (2007). Apoplastic barriers to radial oxygen loss and solute penetration: a chemical and functional comparison of the exodermis of two wetland species, Phr- agmites australis and Glyceria maxima. New Phytol 173, 264-278. [本文引用: 2]
[111]
SoukupA, VotrubováO, ČížkováH (2002). Development of anatomical structure of roots of Phragmites australis. New Phytol 153, 277-287. [本文引用: 1]
[112]
StarkRE, SohnW, Pacchiano JrRA, Al-BashirM, GarbowJR (1994). Following suberization in potato wound periderm by histochemical and solid-state 13C nuclear magnetic resonance methods.Plant Physiol 104, 527-533. [本文引用: 1]
ThomasR, FangX, RanathungeK, AndersonTR, PetersonCA, BernardsMA (2007). Soybean root su- berin: anatomical distribution, chemical composition, and relationship to partial resistance to Phytophthora sojae. Plant Physiol 144, 299-311. [本文引用: 1]
[117]
TrenkampS, MartinW, TietjenK (2004). Specific and differential inhibition of very-long-chain fatty acid elong- ases from Arabidopsis thaliana by different herbicides. Proc Natl Acad Sci USA 101, 11903-11908. [本文引用: 1]
[118]
TyermanSD, BohnertH, MaurelC, SteudleE, SmithJ (1999). Plant aquaporins: their molecular biology, bio- physics and significance for plant water relations.J Exp Bot 50, 1055-1071. [本文引用: 1]
[119]
VioqueJ, KolattukudyP (1997). Resolution and purification of an aldehyde-generating and an alcohol-generating fatty acyl-CoA reductase from pea leaves (Pisum sativum L.). Arch Biochem Biophys 340, 64-72. [本文引用: 1]
[120]
VishwanathSJ, DeludeC, DomergueF (2015). Suberin: biosynthesis, regulation, and polymer assembly of a pro- tective extracellular barrier.Plant Cell Rep 34, 573-586. [本文引用: 2]
[121]
VisserE, ColmerT, BlomC, VoesenekL (2000). Changes in growth, porosity, and radial oxygen loss from adven- titious roots of selected mono- and dicotyledonous wetl- and species with contrasting types of aerenchyma.Plant Cell Environ 23, 1237-1245. [本文引用: 1]
[122]
WaduwaraCI, WalcottSE, PetersonCA (2008). Suberin lamellae of the onion root endodermis: their pattern of development and continuity.Botany 86, 623-632. [本文引用: 1]
[123]
WeiH, PerssonS, MehtaT, SrinivasasainagendraV, ChenL, PageGP, SomervilleC, LoraineA (2006). Transcriptional coordination of the metabolic network in Arabidopsis.Plant Physiol 142, 762-774. [本文引用: 1]
[124]
WengJK, ChappleC (2010). The origin and evolution of lignin biosynthesis.New Phytol 187, 273-285. [本文引用: 1]
[125]
WhitePJ (2001). The pathways of calcium movement to the xylem.J Exp Bot 52, 891-899. [本文引用: 1]
[126]
YadavV, MolinaI, RanathungeK, CastilloIQ, RothsteinSJ, ReedJW (2014). ABCG transporters are required for suberin and pollen wall extracellular barriers in Arabidop- sis.Plant Cell 26, 3569-3588. [本文引用: 1]
[127]
YanB, StarkRE (2000). Biosynthesis, molecular structure, and domain architecture of potato suberin: a 13C NMR study using isotopically labeled precursors.J Agric Food Chem 48, 3298-3304. [本文引用: 2]
[128]
YangW, PollardM, Li-BeissonY, BeissonF, FeigM, OhlroggeJ (2010). A distinct type of glycerol-3-phos- phate acyltransferase with sn-2 preference and phos- phatase activity producing 2-monoacylglycerol. Proc Natl Acad Sci USA 107, 12040-12045. [本文引用: 2]
[129]
YeatsTH, MartinLB, ViartHM, IsaacsonT, HeY, ZhaoL, MatasAJ, BudaGJ, DomozychDS, ClausenMH (2012). The identification of cutin synthase: formation of the plant polyester cutin.Nat Chem Biol 8, 609-611. [本文引用: 1]
[130]
ZhongR, DemuraT, YeZH (2006). SND1, a NAC domain transcription factor, is a key regulator of secondary wall synthesis in fibers of Arabidopsis.Plant Cell 18, 3158-3170. [本文引用: 1]
[131]
ZhongR, LeeC, YeZH (2010). Functional characterization of poplar wood-associated NAC domain transcription fac- tors.Plant Physiol 152, 1044-1055. [本文引用: 1]
[132]
ZhouJ, LeeC, ZhongR, YeZH (2009). MYB58 and MYB63 are transcriptional activators of the lignin biosynthetic pathway during secondary cell wall formation in Arabi- dopsis.Plant Cell 21, 248-266. [本文引用: 1]
[133]
ZimmermannHM, HartmannK, SchreiberL, SteudleE (2000). Chemical composition of apoplastic transport bar- riers in relation to radial hydraulic conductivity of corn roots (Zea mays L.). Planta 210, 302-311. [本文引用: 1]
1 1977
... 目前已经确定的参与木栓质合成的酶包括细胞色素P450单加氧酶、氧化还原酶、脂肪酸延长酶和过氧化物酶.它们分别参与脂肪酸ω-羟基化、二酸形成、脂肪酸延长和芳香类化合物聚合, 其活性可在多种植物组织的聚酯形成过程中检测到(Agrawal and Kola- ttukudy, 1977; Razem and Bernards, 2003; Schrei- ber et al., 2005c). ...
1 2007
... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
1 2005
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
1 2000
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
1 2009
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
4 2007
... 木栓质片层超微结构可进一步通过透射电子显微镜观察.Schmutz等(1996)通过抑制脂肪酸链的延长, 发现木栓质浅色片层的厚度和脂肪酸的烷基链长度存在相关性.Bernards (2002)研究表明, 木栓质深色或者黑色条带和酚类物质的含量相关.酚类物质可以通过自发荧光或者样品染色后在显微镜下进行观察检测.但是, 这些检测不能区分木栓质酚类物质、木质素或者细胞壁中的其它酚类成分.此外, 木栓质形成早期或者少量时, 染色技术经常不够灵敏.例如, 对湿地植物Gyceria maxima根的横截面进行自发荧光检测, 结果发现酚类物质已经开始堆积, GC-MS分析也证实了皮下层组织已形成较多的脂肪类化合物.但是, 苏丹红7B或者荧光黄88染料并不能检测到木栓质(Soukup et al., 2007).与此类似, 在拟南芥种子发育后期, 仅种子合点区域可以被苏丹红染色(Beis- son et al., 2007; Franke et al., 2009). ... ... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ... ... 与CYP基因相似, GPAT5是木栓化过程中首批被鉴定的基因之一, 主要在根的内皮层组织中表达(Beis- son et al., 2007).对突变体gpat5的研究表明, 其编码的甘油-3-磷酸酰基转移酶(GPAT5)参与根和种皮木栓质中C22与C24超长链酰基单体的形成.在gpat5突变体根中, C22/C24单酰基甘油和超长链烷羟基肉桂酸酯的含量减少, 说明GPAT5在脂类聚合物的生物合成中发挥关键作用.与贮存型脂肪生物合成中GPATs的sn-1酰基化作用不同, 在木栓化过程中, GPAT5催化甘油的sn-2酰基化(Yang et al., 2010).与相关蛋白质的作用特性相比较, 仅在木栓质合成中发现了GPATs的sn-2酰基化作用.动物、微生物和藻类也缺失具有sn-2酰基化作用的GPATs, 这可能与陆地植物的进化过程相关(Yang et al., 2010). ... ... 在CYP86A1基因突变和CYP86A33基因沉默的植物中, 超长链ω-羟基酸和α, ω-二羧酸(>C20)并没有减少(Li et al., 2007; Höfer et al., 2008; Serra et al., 2009b), 表明有其它P450s参与木栓质单体合成.敲除或者沉默GPAT5和KCS2基因仅导致单体的部分损失, 影响部分脂肪酸链的延长, 说明还有1种或多种其它的GPATs和KCS参与此过程(Beisson et al., 2007; Serra et al., 2009a).转录水平研究证明, 渗透胁迫与盐胁迫可诱导KCS2和KCS20共同表达(Lee et al., 2009; Franke et al., 2009). ...
1 2012
... 借助基因工程技术和相关分析方法, 已在马铃薯(Solanum tuberosum)和拟南芥(Arabidopsis thali- ana)的木栓质组成、结构、分布模式和功能的研究 上取得了较大进展(Kolattukudy, 2001; Bernards, 2002; Ranathunge et al., 2011; Beisson et al., 2012; Franke et al., 2012).本文回顾和总结了近年来木栓质的相关研究成果, 并对未来的研究方向进行了展望. ...
1 1998
... 细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011).P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006).Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化.复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程.融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008).与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ...
4 2002
... 借助基因工程技术和相关分析方法, 已在马铃薯(Solanum tuberosum)和拟南芥(Arabidopsis thali- ana)的木栓质组成、结构、分布模式和功能的研究 上取得了较大进展(Kolattukudy, 2001; Bernards, 2002; Ranathunge et al., 2011; Beisson et al., 2012; Franke et al., 2012).本文回顾和总结了近年来木栓质的相关研究成果, 并对未来的研究方向进行了展望. ... ... 除此之外, 木栓质也广泛存在于损伤愈合和受环境胁迫的组织中, 如因非生物胁迫和微生物侵染等造成损伤的部位(Lulai and Corsini, 1998; Kolattukudy, 2001; Enstone et al., 2003).因表皮损伤而重建的马铃薯块茎周皮已经作为研究木栓质的模式材料(Kol- attukudy, 2001; Bernards, 2002).当植物需要防护性屏障时, 就会合成和积累木栓质(Kolattukudy, 2001; Franke and Schreiber, 2007).因此, 当植物需要将其自身或者其组织与环境相隔开, 木栓质就会在任何位置进行堆积, 形成屏障(Kolattukudy, 2001).但是, 木栓质的性能因植物种类、木栓质单体多样性及其堆积程度而异. ... ... 组织学研究证实, 木栓质位于细胞壁和质膜之间(Bernards, 2002; Lulai et al., 2008).木栓化的细胞壁主要成分是芳香类(以酚类物质为主)和脂肪类化合物.在荧光显微镜下观察非染色样品, 可以检测到木栓质的多酚类物质.用特定染料如小檗碱、甲苯胺蓝、苏丹红或者荧光黄88对样品进行染色, 显微镜下可以检测到木栓质的脂肪类和其它组分(Brundrett et al., 1988, 1991).通过这些技术, 我们可以在植物的周皮、块茎和根部观察到木栓质(图1). ... ... 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加.相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012).苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008).块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002).Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ...
1 1995
... 为探究木栓质单体间的相互作用, 前人通过多糖水解酶降解典型的木栓化组织或富含木栓质的细胞壁, 将分离下来的低分子量化合物进行色谱和质谱鉴定.之后, 将被鉴定的物质在原位进行重组.然而, 由于不同化合物及其分子间连接的复杂性, 这种方式并不可行(Thomas et al., 2007).也有研究者尝试利用碱水解和酯基转移作用清除脂肪类物质间的连接, 释放出脂肪酸衍生物, 让其仅和酚酸相连, 以此探究木栓质单体间的连接.但是, 这种适用于木质素的降解方法并不适用于检测木栓质单体间的作用.多酚类物质中, 醚的形成和C-C连接的可能性使其它类似处理也并未取得成功(Graça and Pereira, 2000).对马铃薯损伤周皮进行13C固态核磁共振光谱分析(NMR), 试图揭示木栓质多聚物中羧基功能的化学转变, 然而仅仅得到了追踪信号, 并未得到详细的功能变换数据(Bern- ards et al., 1995; Yan and Stark, 2000). ...
1 2008
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 2003
... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ...
1 2006
... 角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008).因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE.KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008).然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定.在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008).此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006).对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009).同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程. ...
1 2008
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2007
... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ...
1 1988
... 组织学研究证实, 木栓质位于细胞壁和质膜之间(Bernards, 2002; Lulai et al., 2008).木栓化的细胞壁主要成分是芳香类(以酚类物质为主)和脂肪类化合物.在荧光显微镜下观察非染色样品, 可以检测到木栓质的多酚类物质.用特定染料如小檗碱、甲苯胺蓝、苏丹红或者荧光黄88对样品进行染色, 显微镜下可以检测到木栓质的脂肪类和其它组分(Brundrett et al., 1988, 1991).通过这些技术, 我们可以在植物的周皮、块茎和根部观察到木栓质(图1). ...
1 1991
... 组织学研究证实, 木栓质位于细胞壁和质膜之间(Bernards, 2002; Lulai et al., 2008).木栓化的细胞壁主要成分是芳香类(以酚类物质为主)和脂肪类化合物.在荧光显微镜下观察非染色样品, 可以检测到木栓质的多酚类物质.用特定染料如小檗碱、甲苯胺蓝、苏丹红或者荧光黄88对样品进行染色, 显微镜下可以检测到木栓质的脂肪类和其它组分(Brundrett et al., 1988, 1991).通过这些技术, 我们可以在植物的周皮、块茎和根部观察到木栓质(图1). ...
1 2011
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 1987
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
4 2009
... 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997).与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009).以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009).Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构.然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致.例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ... ... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ... ... 在拟南芥根的内皮层中, CYP86B1的表达也具有单体特异性, 即在ralph突变体中, CYP86B1的缺失对木栓质成分产生显著影响(Compagnon et al., 2009).在此突变体的根和种皮的木栓质中, 超长链ω-羟基酸和α, ω-二羧酸, 尤其链长为C22和C24的羧酸几乎完全缺失(Compagnon et al., 2009; Molina et al., 2009).在拟南芥RNAi株系中, CYP86B1表达水平的局部降低也导致这些特殊长链羧酸含量减少, 这再次证明了CYP86B1是生成木栓质ω-羟化脂肪酸(链长≥C22)所必需的酶.虽然该酶的具体功能仍未确定, 但这些研究均表明CYP86B1编码一种超长链脂肪酸ω-羟基化酶.此外, 有研究表明超长链氧化脂肪酸的损耗可以通过增加未被氧化的C22和C24脂肪酸来补充.即在cyp86b1/ralph突变体的木栓质多聚酯中, 双官能团脂肪酸可以被单官能团脂肪酸所代替.然而, Molina等(2009)认为, 单官能团的单体很大一部分并不延伸或者交联其它脂肪族多聚物, 因此单官能团脂肪酸会使酯多聚物链终止.所以, 还未被鉴定的木栓质成分对准确描述木栓质的结构非常重要. ... ... ).在此突变体的根和种皮的木栓质中, 超长链ω-羟基酸和α, ω-二羧酸, 尤其链长为C22和C24的羧酸几乎完全缺失(Compagnon et al., 2009; Molina et al., 2009).在拟南芥RNAi株系中, CYP86B1表达水平的局部降低也导致这些特殊长链羧酸含量减少, 这再次证明了CYP86B1是生成木栓质ω-羟化脂肪酸(链长≥C22)所必需的酶.虽然该酶的具体功能仍未确定, 但这些研究均表明CYP86B1编码一种超长链脂肪酸ω-羟基化酶.此外, 有研究表明超长链氧化脂肪酸的损耗可以通过增加未被氧化的C22和C24脂肪酸来补充.即在cyp86b1/ralph突变体的木栓质多聚酯中, 双官能团脂肪酸可以被单官能团脂肪酸所代替.然而, Molina等(2009)认为, 单官能团的单体很大一部分并不延伸或者交联其它脂肪族多聚物, 因此单官能团脂肪酸会使酯多聚物链终止.所以, 还未被鉴定的木栓质成分对准确描述木栓质的结构非常重要. ...
1 2006
... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
1 2003
... 湿地植物的外皮层或周皮组织, 普遍存在木栓质和木质素等非原生质体生化聚合物, 它们形成了一种渗氧(ROL)屏障, 阻碍氧气扩散到根际土壤中.同时, 这些木栓质屏障也阻碍了土壤和微生物释放的有毒气体渗透到根中(Visser et al., 2000; De Simone et al., 2003; Soukup et al., 2007; Garthwaite et al., 2008). ...
... 湿地植物的外皮层或周皮组织, 普遍存在木栓质和木质素等非原生质体生化聚合物, 它们形成了一种渗氧(ROL)屏障, 阻碍氧气扩散到根际土壤中.同时, 这些木栓质屏障也阻碍了土壤和微生物释放的有毒气体渗透到根中(Visser et al., 2000; De Simone et al., 2003; Soukup et al., 2007; Garthwaite et al., 2008). ...
1 2012
... 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知.近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012).与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007).一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006). ...
1 2009
... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
... 木栓质是一种非原生质体的甘油酚-脂类生化多聚物, 其堆积具有组织特异性(如植物周皮、地上表皮和地下根等组织).研究最早的木栓质组织是欧洲栓皮栎(Quercus suber)的树皮周皮, 其木栓质含量超过周皮干重的50% (Graça and Pereira, 1997).作为植物的最外层防护, 这些边界组织可以控制水、矿物质和主要气体的流通, 防御微生物的进攻和有毒化合物的渗入. ... ... 木栓质是一种以甘油为基础且化学成分复杂得多聚物, 包括与细胞壁相关的多芳香类、脂肪类物质和部分蜡质(Franke and Schreiber, 2007; Pollard et al., 2008).其脂肪类物质包括饱和或不饱和的长链与超长链羟基二羧酸, 它们或者环氧化或者在链中部被二醇基取代.另外, 也包括中链氧化脂肪酸和脂肪醇类物质, 它们一般通过与阿魏酰辅酶A形成酯而与酚类区域连接(Graça and Pereira, 1997; Kolattukudy, 2001; Pollard et al., 2008).木栓质中甘油被酯化为ω-羟基甘油酯和α, ω-二羧酸甘油酯等(Graça and Santos, 2007).组成木栓质的单体与角质单体类似, 但是在木栓质中, 其脂肪酸链更长, α, ω-二羧酸所占的比例更大.通过酯基转移反应, 将拟南芥木栓质解聚, 检测到木栓质的单体包括链长为C16-C24的ω-羟基脂肪酸、α, ω-二羧酸、未被氧化的长链脂肪酸(>C18)、链长为C18-C22的脂肪醇和阿魏酸等酚类物质(Franke et al., 2005; Molina et al., 2006). ... ... 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997).与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009).以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009).Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构.然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致.例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ...
1 2000
... 为探究木栓质单体间的相互作用, 前人通过多糖水解酶降解典型的木栓化组织或富含木栓质的细胞壁, 将分离下来的低分子量化合物进行色谱和质谱鉴定.之后, 将被鉴定的物质在原位进行重组.然而, 由于不同化合物及其分子间连接的复杂性, 这种方式并不可行(Thomas et al., 2007).也有研究者尝试利用碱水解和酯基转移作用清除脂肪类物质间的连接, 释放出脂肪酸衍生物, 让其仅和酚酸相连, 以此探究木栓质单体间的连接.但是, 这种适用于木质素的降解方法并不适用于检测木栓质单体间的作用.多酚类物质中, 醚的形成和C-C连接的可能性使其它类似处理也并未取得成功(Graça and Pereira, 2000).对马铃薯损伤周皮进行13C固态核磁共振光谱分析(NMR), 试图揭示木栓质多聚物中羧基功能的化学转变, 然而仅仅得到了追踪信号, 并未得到详细的功能变换数据(Bern- ards et al., 1995; Yan and Stark, 2000). ...
1 2007
... 木栓质是一种以甘油为基础且化学成分复杂得多聚物, 包括与细胞壁相关的多芳香类、脂肪类物质和部分蜡质(Franke and Schreiber, 2007; Pollard et al., 2008).其脂肪类物质包括饱和或不饱和的长链与超长链羟基二羧酸, 它们或者环氧化或者在链中部被二醇基取代.另外, 也包括中链氧化脂肪酸和脂肪醇类物质, 它们一般通过与阿魏酰辅酶A形成酯而与酚类区域连接(Graça and Pereira, 1997; Kolattukudy, 2001; Pollard et al., 2008).木栓质中甘油被酯化为ω-羟基甘油酯和α, ω-二羧酸甘油酯等(Graça and Santos, 2007).组成木栓质的单体与角质单体类似, 但是在木栓质中, 其脂肪酸链更长, α, ω-二羧酸所占的比例更大.通过酯基转移反应, 将拟南芥木栓质解聚, 检测到木栓质的单体包括链长为C16-C24的ω-羟基脂肪酸、α, ω-二羧酸、未被氧化的长链脂肪酸(>C18)、链长为C18-C22的脂肪醇和阿魏酸等酚类物质(Franke et al., 2005; Molina et al., 2006). ...
1 2003
... 马铃薯块茎在采收、处理和种切过程中会发生损伤, 伤口的快速愈合对于避免病原菌侵染等非常重要.Lulai和Suttle (2004)证明, 虽然乙烯参与了块茎的损伤反应, 但是乙烯并不直接影响损伤诱导木栓质的合成.ABA调控植物对干旱和盐胁迫的应答(Himm- elbach et al., 2003), 且ABA与水通道蛋白基因的表达相关, 因此推测其具有防止细胞干燥等保护块茎的作用(Efetova et al., 2007). ...
4 2008
... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ... ... 细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011).P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006).Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化.复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程.融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008).与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ... ... ; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ... ... 在CYP86A1基因突变和CYP86A33基因沉默的植物中, 超长链ω-羟基酸和α, ω-二羧酸(>C20)并没有减少(Li et al., 2007; Höfer et al., 2008; Serra et al., 2009b), 表明有其它P450s参与木栓质单体合成.敲除或者沉默GPAT5和KCS2基因仅导致单体的部分损失, 影响部分脂肪酸链的延长, 说明还有1种或多种其它的GPATs和KCS参与此过程(Beisson et al., 2007; Serra et al., 2009a).转录水平研究证明, 渗透胁迫与盐胁迫可诱导KCS2和KCS20共同表达(Lee et al., 2009; Franke et al., 2009). ...
2 2011
... 脂肪酸合成以后, 一般会被长链酰基-CoA合成酶(LACSs)转化成酰基-CoA酯.最近的研究表明, 至少3个长链酰基-CoA合成异构酶参与拟南芥蜡质的合成, 它们催化游离脂肪酸活化为脂肪酰CoA硫酯(Lü et al., 2009; Weng and Chapple, 2010; Jessen et al., 2011).进一步研究显示, 拟南芥中包括9个属于酰基活化酶超家族的LACSs, 它们也参与脂肪酸的运输过程(Shockey and Fulda, 2003).LACS1-3是芽殖酵母(Saccharomyces cerevisiae)中脂类运输所必需的酶(Pulsifer et al., 2012).LACS1和2也参与了角质单体的合成, 激活脂肪酸生成酰基-CoA酯(Schnurr and Shockey, 2004).LACS的活性位点是在叶绿体被膜的内表面还是外表面仍不清楚. ... ... 在lacs1突变体中, 茎中的蜡质总含量减少了40%, 游离的超长链脂肪酸在蜡质中堆积, 所有烷烃形成路径中的化合物含量均降低.在lacs2/lacs3双突变体中, 茎中蜡质含量减少了5% (Lü et al., 2009).关于lacs1和lacs4双敲除突变体的研究, 证明了LACS1和LACS4都是含油花粉外被皮中脂类形成所必需的酶(Jessen et al., 2011).然而, 对lacs9突变体的研究显示LACS9对脂类的合成没有影响, 表明这个蛋白质家族的功能可能具有冗余性(Schnurr et al., 2002). ...
1 2008
... 角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008).因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE.KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008).然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定.在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008).此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006).对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009).同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程. ...
2006
1 2000
... 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加.相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012).苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008).块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002).Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ...
2012
2 2007
... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ... ... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 1971
... 饱和长链(C18、C20和C22)醇类物质是木栓质的普遍成分(Franke et al., 2005).对荷荷巴(Simmondsia chinensis)种子木栓质合成相关酶的研究表明, 以超长链脂肪酸为底物生成醇类的反应过程由脂肪酰还原酶(FAR)催化完成(Kolattukudy, 1971; Vioque and Kolattukudy, 1997; Metz et al., 2000).通过对酵母中超长链(C24和C26)醇类物质的产生过程进行研究, 确定了FAR的活性(Rowland and Domergue, 2012).脂肪酰还原酶(FARs)催化活化的脂肪酸还原为醇类物质.对FAR编码基因的研究显示, 拟南芥基因组中包括8个FAR基因序列, 分别编码FAR1-8, 其中1个序列被鉴定为CER4 (Rowland et al., 2006).在CER4突变体木栓质中, 链长C24-C28的主要醇类几乎完全缺失, 但是仍有部分C30醇类, 说明其它FAR或者相关的功能酶可能对C30醇的形成发挥作用(Rowl- and and Domergue, 2012).在研究FAR基因家族的表达模式时, 发现FAR1、FAR4和FAR5在拟南芥根内皮和其它含有木栓质的组织中出现, 相应的三突变体导致C18、C20和C22链长的木栓质醇类物质含量减少(Domergue et al., 2010), 表明FAR参与了根和种皮中木栓质长链(≥C18)醇类的合成. ...
1981
6 2001
... 借助基因工程技术和相关分析方法, 已在马铃薯(Solanum tuberosum)和拟南芥(Arabidopsis thali- ana)的木栓质组成、结构、分布模式和功能的研究 上取得了较大进展(Kolattukudy, 2001; Bernards, 2002; Ranathunge et al., 2011; Beisson et al., 2012; Franke et al., 2012).本文回顾和总结了近年来木栓质的相关研究成果, 并对未来的研究方向进行了展望. ... ... 除此之外, 木栓质也广泛存在于损伤愈合和受环境胁迫的组织中, 如因非生物胁迫和微生物侵染等造成损伤的部位(Lulai and Corsini, 1998; Kolattukudy, 2001; Enstone et al., 2003).因表皮损伤而重建的马铃薯块茎周皮已经作为研究木栓质的模式材料(Kol- attukudy, 2001; Bernards, 2002).当植物需要防护性屏障时, 就会合成和积累木栓质(Kolattukudy, 2001; Franke and Schreiber, 2007).因此, 当植物需要将其自身或者其组织与环境相隔开, 木栓质就会在任何位置进行堆积, 形成屏障(Kolattukudy, 2001).但是, 木栓质的性能因植物种类、木栓质单体多样性及其堆积程度而异. ... ... ).因表皮损伤而重建的马铃薯块茎周皮已经作为研究木栓质的模式材料(Kol- attukudy, 2001; Bernards, 2002).当植物需要防护性屏障时, 就会合成和积累木栓质(Kolattukudy, 2001; Franke and Schreiber, 2007).因此, 当植物需要将其自身或者其组织与环境相隔开, 木栓质就会在任何位置进行堆积, 形成屏障(Kolattukudy, 2001).但是, 木栓质的性能因植物种类、木栓质单体多样性及其堆积程度而异. ... ... ).当植物需要防护性屏障时, 就会合成和积累木栓质(Kolattukudy, 2001; Franke and Schreiber, 2007).因此, 当植物需要将其自身或者其组织与环境相隔开, 木栓质就会在任何位置进行堆积, 形成屏障(Kolattukudy, 2001).但是, 木栓质的性能因植物种类、木栓质单体多样性及其堆积程度而异. ... ... ).因此, 当植物需要将其自身或者其组织与环境相隔开, 木栓质就会在任何位置进行堆积, 形成屏障(Kolattukudy, 2001).但是, 木栓质的性能因植物种类、木栓质单体多样性及其堆积程度而异. ... ... 木栓质是一种以甘油为基础且化学成分复杂得多聚物, 包括与细胞壁相关的多芳香类、脂肪类物质和部分蜡质(Franke and Schreiber, 2007; Pollard et al., 2008).其脂肪类物质包括饱和或不饱和的长链与超长链羟基二羧酸, 它们或者环氧化或者在链中部被二醇基取代.另外, 也包括中链氧化脂肪酸和脂肪醇类物质, 它们一般通过与阿魏酰辅酶A形成酯而与酚类区域连接(Graça and Pereira, 1997; Kolattukudy, 2001; Pollard et al., 2008).木栓质中甘油被酯化为ω-羟基甘油酯和α, ω-二羧酸甘油酯等(Graça and Santos, 2007).组成木栓质的单体与角质单体类似, 但是在木栓质中, 其脂肪酸链更长, α, ω-二羧酸所占的比例更大.通过酯基转移反应, 将拟南芥木栓质解聚, 检测到木栓质的单体包括链长为C16-C24的ω-羟基脂肪酸、α, ω-二羧酸、未被氧化的长链脂肪酸(>C18)、链长为C18-C22的脂肪醇和阿魏酸等酚类物质(Franke et al., 2005; Molina et al., 2006). ...
1 2009
... 20世纪90年代, 研究者普遍采用显微观察技术研究木栓化组织的结构及其复杂成分(Schreiber et al., 1999).之后, 结合GC-MS等技术, 不同物种、不同组织和不同环境下的特异木栓质可以被定量和定性描述(Zimmermann et al., 2000; Schreiber et al., 2005b).随着分析技术的成熟, 影响木栓质形成和堆积的因素逐渐清晰, 包括各种环境胁迫、机械损伤和激素处理.在环境胁迫如盐胁迫(Krishnamurthy et al., 2009)或缺氧(Kotula et al., 2009)条件下, 水稻(Oryza sativa)根外皮层的凯氏带和片层木栓质的堆积量显著增加.目前, 激素对木栓质形成和堆积的影响, 尤其是脱落酸(ABA)的作用, 逐渐成为研究热点. ...
2 2009
... 20世纪90年代, 研究者普遍采用显微观察技术研究木栓化组织的结构及其复杂成分(Schreiber et al., 1999).之后, 结合GC-MS等技术, 不同物种、不同组织和不同环境下的特异木栓质可以被定量和定性描述(Zimmermann et al., 2000; Schreiber et al., 2005b).随着分析技术的成熟, 影响木栓质形成和堆积的因素逐渐清晰, 包括各种环境胁迫、机械损伤和激素处理.在环境胁迫如盐胁迫(Krishnamurthy et al., 2009)或缺氧(Kotula et al., 2009)条件下, 水稻(Oryza sativa)根外皮层的凯氏带和片层木栓质的堆积量显著增加.目前, 激素对木栓质形成和堆积的影响, 尤其是脱落酸(ABA)的作用, 逐渐成为研究热点. ... ... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
2011
2 2003
... 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加.相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012).苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008).块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002).Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ... ... ).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ...
1 2007
... 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加.相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012).苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008).块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002).Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ...
... 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知.近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012).与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007).一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006). ...
1 2014
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 2008
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
5 2009
... 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997).与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009).以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009).Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构.然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致.例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ... ... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ... ... 角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008).因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE.KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008).然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定.在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008).此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006).对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009).同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程. ... ... 在拟南芥基因(KCS2)敲除突变体(daisy)的根木栓质中, C22和C24超长链单体含量显著减少, KCS2的表达位置与模式也与木栓质的堆积和功能相一致(Lee et al., 2009; Franke et al., 2009).然而, 在DAISY/KCS2突变体的其它组织中, C22和C24或者中间链长的脂肪酸仍出现在其它脂类结构(鞘脂类和蜡质)中, 表明促进木栓化可能并不是KCS2的唯一功能.双突变分析进一步证实了KCS2与KCS20在木 栓质和蜡质生物合成中具有双重作用(Lee et al., 2009). ... ... 在CYP86A1基因突变和CYP86A33基因沉默的植物中, 超长链ω-羟基酸和α, ω-二羧酸(>C20)并没有减少(Li et al., 2007; Höfer et al., 2008; Serra et al., 2009b), 表明有其它P450s参与木栓质单体合成.敲除或者沉默GPAT5和KCS2基因仅导致单体的部分损失, 影响部分脂肪酸链的延长, 说明还有1种或多种其它的GPATs和KCS参与此过程(Beisson et al., 2007; Serra et al., 2009a).转录水平研究证明, 渗透胁迫与盐胁迫可诱导KCS2和KCS20共同表达(Lee et al., 2009; Franke et al., 2009). ...
... 细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011).P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006).Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化.复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程.融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008).与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ... ... 在CYP86A1基因突变和CYP86A33基因沉默的植物中, 超长链ω-羟基酸和α, ω-二羧酸(>C20)并没有减少(Li et al., 2007; Höfer et al., 2008; Serra et al., 2009b), 表明有其它P450s参与木栓质单体合成.敲除或者沉默GPAT5和KCS2基因仅导致单体的部分损失, 影响部分脂肪酸链的延长, 说明还有1种或多种其它的GPATs和KCS参与此过程(Beisson et al., 2007; Serra et al., 2009a).转录水平研究证明, 渗透胁迫与盐胁迫可诱导KCS2和KCS20共同表达(Lee et al., 2009; Franke et al., 2009). ...
3 2013
... 目前, 还没有关于LACS基因参与木栓质合成的相关报道.但是LACS2基因功能缺失突变体的分析显示, LACS2对木栓质的形成具有一定作用(Li-Beisson et al., 2013).LACS也可能在脂肪酸酯化前发挥修饰脂肪酸的作用(如ω-羟基化或羧基化). ... ... 木栓质生物合成还涉及脂肪醇类物质的合成、环氧化作用、单体输出到质外体和多聚物的聚合装配等过程, 而这些过程亦受多种酶调控, 也可能有更大的未知代谢网络参与调控木栓质的合成(Franke and Schreiber, 2007; Li-Beisson et al., 2013). ... ... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 1994
... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
2 2009
... 脂肪酸合成以后, 一般会被长链酰基-CoA合成酶(LACSs)转化成酰基-CoA酯.最近的研究表明, 至少3个长链酰基-CoA合成异构酶参与拟南芥蜡质的合成, 它们催化游离脂肪酸活化为脂肪酰CoA硫酯(Lü et al., 2009; Weng and Chapple, 2010; Jessen et al., 2011).进一步研究显示, 拟南芥中包括9个属于酰基活化酶超家族的LACSs, 它们也参与脂肪酸的运输过程(Shockey and Fulda, 2003).LACS1-3是芽殖酵母(Saccharomyces cerevisiae)中脂类运输所必需的酶(Pulsifer et al., 2012).LACS1和2也参与了角质单体的合成, 激活脂肪酸生成酰基-CoA酯(Schnurr and Shockey, 2004).LACS的活性位点是在叶绿体被膜的内表面还是外表面仍不清楚. ... ... 在lacs1突变体中, 茎中的蜡质总含量减少了40%, 游离的超长链脂肪酸在蜡质中堆积, 所有烷烃形成路径中的化合物含量均降低.在lacs2/lacs3双突变体中, 茎中蜡质含量减少了5% (Lü et al., 2009).关于lacs1和lacs4双敲除突变体的研究, 证明了LACS1和LACS4都是含油花粉外被皮中脂类形成所必需的酶(Jessen et al., 2011).然而, 对lacs9突变体的研究显示LACS9对脂类的合成没有影响, 表明这个蛋白质家族的功能可能具有冗余性(Schnurr et al., 2002). ...
2 1998
... 除此之外, 木栓质也广泛存在于损伤愈合和受环境胁迫的组织中, 如因非生物胁迫和微生物侵染等造成损伤的部位(Lulai and Corsini, 1998; Kolattukudy, 2001; Enstone et al., 2003).因表皮损伤而重建的马铃薯块茎周皮已经作为研究木栓质的模式材料(Kol- attukudy, 2001; Bernards, 2002).当植物需要防护性屏障时, 就会合成和积累木栓质(Kolattukudy, 2001; Franke and Schreiber, 2007).因此, 当植物需要将其自身或者其组织与环境相隔开, 木栓质就会在任何位置进行堆积, 形成屏障(Kolattukudy, 2001).但是, 木栓质的性能因植物种类、木栓质单体多样性及其堆积程度而异. ... ... 木栓质坚固的结构及其疏水多聚物的化学特性在植物体内具有重要的生理作用.木栓质建立了组织屏障, 可以防止水分以及离子流失, 也可抵抗病原物的侵染(Machado et al., 2013).通过对环境胁迫下的植物组织化学特性进行研究, 已证明环境胁迫可以增强木栓化水平(Enstone et al., 2003).马铃薯块茎的损伤愈合也是通过形成木栓化的封闭层和损伤周皮发展而成.木栓化的细胞和相关疏水物质共同防卫其邻近的组织免受真菌以及微生物病原的侵染(Lulai and Corsini, 1998; Matsuda et al., 2003; Schreiber et al., 2005b). ...
2004
3 2008
... 组织学研究证实, 木栓质位于细胞壁和质膜之间(Bernards, 2002; Lulai et al., 2008).木栓化的细胞壁主要成分是芳香类(以酚类物质为主)和脂肪类化合物.在荧光显微镜下观察非染色样品, 可以检测到木栓质的多酚类物质.用特定染料如小檗碱、甲苯胺蓝、苏丹红或者荧光黄88对样品进行染色, 显微镜下可以检测到木栓质的脂肪类和其它组分(Brundrett et al., 1988, 1991).通过这些技术, 我们可以在植物的周皮、块茎和根部观察到木栓质(图1). ... ... 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加.相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012).苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008).块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002).Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ... ... Lulai等(2008)测定了ABA在损伤诱导的木栓化调控和块茎水分损失中的作用.液质联用色谱分析显示, 整个块茎中ABA总含量基本不变, 但是在损伤周皮附近含量较高, 在中心部位最低.块茎损伤诱导了ABA分布发生变化.损伤后, 在愈合组织块中ABA含量先降低, 24小时达到最低, 损伤后第3-7天ABA含量逐渐增加.这表明内源ABA参与损伤诱导木栓化的形成, 避免由于脱水而致细胞死亡. ...
1 2004
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
1 2013
... 木栓质坚固的结构及其疏水多聚物的化学特性在植物体内具有重要的生理作用.木栓质建立了组织屏障, 可以防止水分以及离子流失, 也可抵抗病原物的侵染(Machado et al., 2013).通过对环境胁迫下的植物组织化学特性进行研究, 已证明环境胁迫可以增强木栓化水平(Enstone et al., 2003).马铃薯块茎的损伤愈合也是通过形成木栓化的封闭层和损伤周皮发展而成.木栓化的细胞和相关疏水物质共同防卫其邻近的组织免受真菌以及微生物病原的侵染(Lulai and Corsini, 1998; Matsuda et al., 2003; Schreiber et al., 2005b). ...
1 2003
... 木栓质坚固的结构及其疏水多聚物的化学特性在植物体内具有重要的生理作用.木栓质建立了组织屏障, 可以防止水分以及离子流失, 也可抵抗病原物的侵染(Machado et al., 2013).通过对环境胁迫下的植物组织化学特性进行研究, 已证明环境胁迫可以增强木栓化水平(Enstone et al., 2003).马铃薯块茎的损伤愈合也是通过形成木栓化的封闭层和损伤周皮发展而成.木栓化的细胞和相关疏水物质共同防卫其邻近的组织免受真菌以及微生物病原的侵染(Lulai and Corsini, 1998; Matsuda et al., 2003; Schreiber et al., 2005b). ...
1 2000
... 饱和长链(C18、C20和C22)醇类物质是木栓质的普遍成分(Franke et al., 2005).对荷荷巴(Simmondsia chinensis)种子木栓质合成相关酶的研究表明, 以超长链脂肪酸为底物生成醇类的反应过程由脂肪酰还原酶(FAR)催化完成(Kolattukudy, 1971; Vioque and Kolattukudy, 1997; Metz et al., 2000).通过对酵母中超长链(C24和C26)醇类物质的产生过程进行研究, 确定了FAR的活性(Rowland and Domergue, 2012).脂肪酰还原酶(FARs)催化活化的脂肪酸还原为醇类物质.对FAR编码基因的研究显示, 拟南芥基因组中包括8个FAR基因序列, 分别编码FAR1-8, 其中1个序列被鉴定为CER4 (Rowland et al., 2006).在CER4突变体木栓质中, 链长C24-C28的主要醇类几乎完全缺失, 但是仍有部分C30醇类, 说明其它FAR或者相关的功能酶可能对C30醇的形成发挥作用(Rowl- and and Domergue, 2012).在研究FAR基因家族的表达模式时, 发现FAR1、FAR4和FAR5在拟南芥根内皮和其它含有木栓质的组织中出现, 相应的三突变体导致C18、C20和C22链长的木栓质醇类物质含量减少(Domergue et al., 2010), 表明FAR参与了根和种皮中木栓质长链(≥C18)醇类的合成. ...
1 2009
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
1 2005
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2006
... 木栓质是一种以甘油为基础且化学成分复杂得多聚物, 包括与细胞壁相关的多芳香类、脂肪类物质和部分蜡质(Franke and Schreiber, 2007; Pollard et al., 2008).其脂肪类物质包括饱和或不饱和的长链与超长链羟基二羧酸, 它们或者环氧化或者在链中部被二醇基取代.另外, 也包括中链氧化脂肪酸和脂肪醇类物质, 它们一般通过与阿魏酰辅酶A形成酯而与酚类区域连接(Graça and Pereira, 1997; Kolattukudy, 2001; Pollard et al., 2008).木栓质中甘油被酯化为ω-羟基甘油酯和α, ω-二羧酸甘油酯等(Graça and Santos, 2007).组成木栓质的单体与角质单体类似, 但是在木栓质中, 其脂肪酸链更长, α, ω-二羧酸所占的比例更大.通过酯基转移反应, 将拟南芥木栓质解聚, 检测到木栓质的单体包括链长为C16-C24的ω-羟基脂肪酸、α, ω-二羧酸、未被氧化的长链脂肪酸(>C18)、链长为C18-C22的脂肪醇和阿魏酸等酚类物质(Franke et al., 2005; Molina et al., 2006). ...
5 2009
... 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997).与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009).以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009).Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构.然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致.例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ... ... ; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ... ... 突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ... ... 在拟南芥根的内皮层中, CYP86B1的表达也具有单体特异性, 即在ralph突变体中, CYP86B1的缺失对木栓质成分产生显著影响(Compagnon et al., 2009).在此突变体的根和种皮的木栓质中, 超长链ω-羟基酸和α, ω-二羧酸, 尤其链长为C22和C24的羧酸几乎完全缺失(Compagnon et al., 2009; Molina et al., 2009).在拟南芥RNAi株系中, CYP86B1表达水平的局部降低也导致这些特殊长链羧酸含量减少, 这再次证明了CYP86B1是生成木栓质ω-羟化脂肪酸(链长≥C22)所必需的酶.虽然该酶的具体功能仍未确定, 但这些研究均表明CYP86B1编码一种超长链脂肪酸ω-羟基化酶.此外, 有研究表明超长链氧化脂肪酸的损耗可以通过增加未被氧化的C22和C24脂肪酸来补充.即在cyp86b1/ralph突变体的木栓质多聚酯中, 双官能团脂肪酸可以被单官能团脂肪酸所代替.然而, Molina等(2009)认为, 单官能团的单体很大一部分并不延伸或者交联其它脂肪族多聚物, 因此单官能团脂肪酸会使酯多聚物链终止.所以, 还未被鉴定的木栓质成分对准确描述木栓质的结构非常重要. ... ... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
1 2009
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2007
... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
1 2006
... 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知.近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012).与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007).一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006). ...
1 2007
... 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知.近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012).与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007).一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006). ...
1 2007
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 2010
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 2006
... 角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008).因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE.KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008).然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定.在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008).此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006).对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009).同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程. ...
1 2004
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
1 2011
... 细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011).P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006).Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化.复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程.融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008).与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ...
... 木栓质是一种以甘油为基础且化学成分复杂得多聚物, 包括与细胞壁相关的多芳香类、脂肪类物质和部分蜡质(Franke and Schreiber, 2007; Pollard et al., 2008).其脂肪类物质包括饱和或不饱和的长链与超长链羟基二羧酸, 它们或者环氧化或者在链中部被二醇基取代.另外, 也包括中链氧化脂肪酸和脂肪醇类物质, 它们一般通过与阿魏酰辅酶A形成酯而与酚类区域连接(Graça and Pereira, 1997; Kolattukudy, 2001; Pollard et al., 2008).木栓质中甘油被酯化为ω-羟基甘油酯和α, ω-二羧酸甘油酯等(Graça and Santos, 2007).组成木栓质的单体与角质单体类似, 但是在木栓质中, 其脂肪酸链更长, α, ω-二羧酸所占的比例更大.通过酯基转移反应, 将拟南芥木栓质解聚, 检测到木栓质的单体包括链长为C16-C24的ω-羟基脂肪酸、α, ω-二羧酸、未被氧化的长链脂肪酸(>C18)、链长为C18-C22的脂肪醇和阿魏酸等酚类物质(Franke et al., 2005; Molina et al., 2006). ... ... ; Pollard et al., 2008).木栓质中甘油被酯化为ω-羟基甘油酯和α, ω-二羧酸甘油酯等(Graça and Santos, 2007).组成木栓质的单体与角质单体类似, 但是在木栓质中, 其脂肪酸链更长, α, ω-二羧酸所占的比例更大.通过酯基转移反应, 将拟南芥木栓质解聚, 检测到木栓质的单体包括链长为C16-C24的ω-羟基脂肪酸、α, ω-二羧酸、未被氧化的长链脂肪酸(>C18)、链长为C18-C22的脂肪醇和阿魏酸等酚类物质(Franke et al., 2005; Molina et al., 2006). ... ... 之前使用的木栓质模式材料(栓皮栎和马铃薯), 由于生长周期较长、基因突变和种间基因组差异, 分子方法在木栓质的合成研究中受到严重限制.然而, 当在短寿命周期植物中发现木栓质的分布和检测到其相关化学特性后, 具有基因资源优势的拟南芥在木栓质的合成研究中被充分利用(Franke et al., 2005).作为模式植物, 拟南芥可以产生公共的基因转录组位点, 并具有木栓质组织特异性, 大大促进了可能涉及多类膜家族蛋白参与的木栓质生物合成的基因筛选(Birnbaum et al., 2003; Brady et al., 2007; Kilian et al., 2007).Birnbaum等(2003)提出了一种基于基因转录丰度的研究方法, 即基于木栓质组织的基因转录丰度, 参照主要代谢过程如脂肪酸氧化、延长和结合作用等, 列出候选基因清单, 在候选基因的数字表达谱中, 对参与木栓质形成的基因进行鉴定.例如, 将报告基因与基因启动子融合, 探究拟南芥木栓质合成的调控基因.在根内皮中, 甘油-3-磷酸酰基转移酶基因(GPAT5、CYP86A1、CYP86B1、阿魏酰转移酶基因(ASFT/AtHHT)、脂肪酰还原酶基因(FAR1、 FAR4和FAR5)、KCS20和DAISY/KCS2)优先表达.在根周皮和种皮中, 可以检测到GPAT5和ASFT的启动子活性.在种脐区域, 可以检测到GPAT5、DAISY/ KCS2、ASFT、FAR1和FAR4的启动子活性.在损伤后的组织中, 可检测到DAISY/KCS2、FAR1、FAR4和FAR5的启动子活性(Beisson et al., 2007; Höfer et al., 2008; Pollard et al., 2008; Compagnon et al., 2009; Lee et al., 2009; Domergue et al., 2010).这些研究证明, 木栓质的生物合成同时受到多种基因的协同调控. ... ... 角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008).因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE.KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008).然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定.在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008).此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006).对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009).同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程. ...
1 2012
... 脂肪酸合成以后, 一般会被长链酰基-CoA合成酶(LACSs)转化成酰基-CoA酯.最近的研究表明, 至少3个长链酰基-CoA合成异构酶参与拟南芥蜡质的合成, 它们催化游离脂肪酸活化为脂肪酰CoA硫酯(Lü et al., 2009; Weng and Chapple, 2010; Jessen et al., 2011).进一步研究显示, 拟南芥中包括9个属于酰基活化酶超家族的LACSs, 它们也参与脂肪酸的运输过程(Shockey and Fulda, 2003).LACS1-3是芽殖酵母(Saccharomyces cerevisiae)中脂类运输所必需的酶(Pulsifer et al., 2012).LACS1和2也参与了角质单体的合成, 激活脂肪酸生成酰基-CoA酯(Schnurr and Shockey, 2004).LACS的活性位点是在叶绿体被膜的内表面还是外表面仍不清楚. ...
1 2008
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2011
... 借助基因工程技术和相关分析方法, 已在马铃薯(Solanum tuberosum)和拟南芥(Arabidopsis thali- ana)的木栓质组成、结构、分布模式和功能的研究 上取得了较大进展(Kolattukudy, 2001; Bernards, 2002; Ranathunge et al., 2011; Beisson et al., 2012; Franke et al., 2012).本文回顾和总结了近年来木栓质的相关研究成果, 并对未来的研究方向进行了展望. ...
1 2011
... 根压探头技术和压力势等的测定技术已经广泛应用于推断切根和整根系统对水及溶质的渗透性, 根对水以及溶质的透性随着皮层木栓质的堆积而变化(Ranathunge and Schreiber, 2011).然而, 也有研究表明, 具有完整木栓质片层的成熟水稻根区的导水率与没有片层或部分片层的根尖区域的导水率并无显著差异(Ranathunge et al., 2005).因此, 木栓质片层可能并不显著影响细胞壁的透水性.这种现象的可能原因有: 细胞膜中的水通道蛋白可以显著改变膜的渗透性(Tyerman et al., 1999); 片层中的穿孔可以作为水自由进出细胞的位点(Waduwara et al., 2008); 非原生质体屏障中木栓质的堆积和多聚物的排列顺序均会对其透性产生影响(Schreiber et al., 2005a). ...
2 2005
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ... ... 根压探头技术和压力势等的测定技术已经广泛应用于推断切根和整根系统对水及溶质的渗透性, 根对水以及溶质的透性随着皮层木栓质的堆积而变化(Ranathunge and Schreiber, 2011).然而, 也有研究表明, 具有完整木栓质片层的成熟水稻根区的导水率与没有片层或部分片层的根尖区域的导水率并无显著差异(Ranathunge et al., 2005).因此, 木栓质片层可能并不显著影响细胞壁的透水性.这种现象的可能原因有: 细胞膜中的水通道蛋白可以显著改变膜的渗透性(Tyerman et al., 1999); 片层中的穿孔可以作为水自由进出细胞的位点(Waduwara et al., 2008); 非原生质体屏障中木栓质的堆积和多聚物的排列顺序均会对其透性产生影响(Schreiber et al., 2005a). ...
2 2003
... 目前已经确定的参与木栓质合成的酶包括细胞色素P450单加氧酶、氧化还原酶、脂肪酸延长酶和过氧化物酶.它们分别参与脂肪酸ω-羟基化、二酸形成、脂肪酸延长和芳香类化合物聚合, 其活性可在多种植物组织的聚酯形成过程中检测到(Agrawal and Kola- ttukudy, 1977; Razem and Bernards, 2003; Schrei- ber et al., 2005c). ... ... 在拟南芥突变体cyp86a1中, CYP活性降低, 导致表皮中羟基脂肪酸及木栓质的总含量严重减少, 进一步导致根对水分和溶质的渗透性增加.相反, 在CYP过表达的突变体中, 其根的木栓质含量增加了2倍, 提高了根中水的利用效率, 也有效控制了根对矿物质的吸收和运输(Franke et al., 2012).苯丙氨酸解氨酶(PAL)是催化苯丙氨酸代谢途径第1步反应过程中的酶, 也是该过程的限速酶(Kato et al., 2000), 在马铃薯块茎损伤愈合过程中发挥重要作用(Kumar and Knowles, 2003; Kumar et al., 2007; Lulai et al., 2008).块茎应答损伤时, PAL转录水平增加, 催化苯丙烷代谢产生肉桂酸和对-香豆酸单体, 最终聚合成木栓质多酚类物质(Bernards, 2002).Kesanakurti等(2012)培育了过表达过氧化物酶的转基因番茄(Lyc- opersicon esculentum).Northern杂交实验表明, 在应答损伤时, 转基因果实表皮中具有很高的过氧化物酶活性, 而对照组番茄表皮中则未检测到此酶活性.相应的荧光共聚焦显微观察结果显示, 在转基因果实的损伤位点有大量的酚类物质积累, 果实对真菌等的抗性也相应提高.因此推测酚类物质的积累与损伤木栓质的形成有关, 但此推测尚待进一步验证.此外, 也有损伤实验表明, 损伤诱导的超氧游离基可能参与受过氧化物酶调控的酚类单体的氧化聚集过程(Kum- ar and Knowles, 2003; Razem and Bernards, 2003).然而, 由于木栓质分布的组织特异性, 以及多酶复合体和膜关联酶的定位问题, 木栓质通路中的主要反应及参与酶尚未得到明确阐释. ...
2 2012
... 饱和长链(C18、C20和C22)醇类物质是木栓质的普遍成分(Franke et al., 2005).对荷荷巴(Simmondsia chinensis)种子木栓质合成相关酶的研究表明, 以超长链脂肪酸为底物生成醇类的反应过程由脂肪酰还原酶(FAR)催化完成(Kolattukudy, 1971; Vioque and Kolattukudy, 1997; Metz et al., 2000).通过对酵母中超长链(C24和C26)醇类物质的产生过程进行研究, 确定了FAR的活性(Rowland and Domergue, 2012).脂肪酰还原酶(FARs)催化活化的脂肪酸还原为醇类物质.对FAR编码基因的研究显示, 拟南芥基因组中包括8个FAR基因序列, 分别编码FAR1-8, 其中1个序列被鉴定为CER4 (Rowland et al., 2006).在CER4突变体木栓质中, 链长C24-C28的主要醇类几乎完全缺失, 但是仍有部分C30醇类, 说明其它FAR或者相关的功能酶可能对C30醇的形成发挥作用(Rowl- and and Domergue, 2012).在研究FAR基因家族的表达模式时, 发现FAR1、FAR4和FAR5在拟南芥根内皮和其它含有木栓质的组织中出现, 相应的三突变体导致C18、C20和C22链长的木栓质醇类物质含量减少(Domergue et al., 2010), 表明FAR参与了根和种皮中木栓质长链(≥C18)醇类的合成. ... ... ).在CER4突变体木栓质中, 链长C24-C28的主要醇类几乎完全缺失, 但是仍有部分C30醇类, 说明其它FAR或者相关的功能酶可能对C30醇的形成发挥作用(Rowl- and and Domergue, 2012).在研究FAR基因家族的表达模式时, 发现FAR1、FAR4和FAR5在拟南芥根内皮和其它含有木栓质的组织中出现, 相应的三突变体导致C18、C20和C22链长的木栓质醇类物质含量减少(Domergue et al., 2010), 表明FAR参与了根和种皮中木栓质长链(≥C18)醇类的合成. ...
1 2006
... 饱和长链(C18、C20和C22)醇类物质是木栓质的普遍成分(Franke et al., 2005).对荷荷巴(Simmondsia chinensis)种子木栓质合成相关酶的研究表明, 以超长链脂肪酸为底物生成醇类的反应过程由脂肪酰还原酶(FAR)催化完成(Kolattukudy, 1971; Vioque and Kolattukudy, 1997; Metz et al., 2000).通过对酵母中超长链(C24和C26)醇类物质的产生过程进行研究, 确定了FAR的活性(Rowland and Domergue, 2012).脂肪酰还原酶(FARs)催化活化的脂肪酸还原为醇类物质.对FAR编码基因的研究显示, 拟南芥基因组中包括8个FAR基因序列, 分别编码FAR1-8, 其中1个序列被鉴定为CER4 (Rowland et al., 2006).在CER4突变体木栓质中, 链长C24-C28的主要醇类几乎完全缺失, 但是仍有部分C30醇类, 说明其它FAR或者相关的功能酶可能对C30醇的形成发挥作用(Rowl- and and Domergue, 2012).在研究FAR基因家族的表达模式时, 发现FAR1、FAR4和FAR5在拟南芥根内皮和其它含有木栓质的组织中出现, 相应的三突变体导致C18、C20和C22链长的木栓质醇类物质含量减少(Domergue et al., 2010), 表明FAR参与了根和种皮中木栓质长链(≥C18)醇类的合成. ...
1 1996
... 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997).与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009).以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009).Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构.然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致.例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ...
1 2004
... 脂肪酸合成以后, 一般会被长链酰基-CoA合成酶(LACSs)转化成酰基-CoA酯.最近的研究表明, 至少3个长链酰基-CoA合成异构酶参与拟南芥蜡质的合成, 它们催化游离脂肪酸活化为脂肪酰CoA硫酯(Lü et al., 2009; Weng and Chapple, 2010; Jessen et al., 2011).进一步研究显示, 拟南芥中包括9个属于酰基活化酶超家族的LACSs, 它们也参与脂肪酸的运输过程(Shockey and Fulda, 2003).LACS1-3是芽殖酵母(Saccharomyces cerevisiae)中脂类运输所必需的酶(Pulsifer et al., 2012).LACS1和2也参与了角质单体的合成, 激活脂肪酸生成酰基-CoA酯(Schnurr and Shockey, 2004).LACS的活性位点是在叶绿体被膜的内表面还是外表面仍不清楚. ...
1 2002
... 在lacs1突变体中, 茎中的蜡质总含量减少了40%, 游离的超长链脂肪酸在蜡质中堆积, 所有烷烃形成路径中的化合物含量均降低.在lacs2/lacs3双突变体中, 茎中蜡质含量减少了5% (Lü et al., 2009).关于lacs1和lacs4双敲除突变体的研究, 证明了LACS1和LACS4都是含油花粉外被皮中脂类形成所必需的酶(Jessen et al., 2011).然而, 对lacs9突变体的研究显示LACS9对脂类的合成没有影响, 表明这个蛋白质家族的功能可能具有冗余性(Schnurr et al., 2002). ...
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ... ... 根压探头技术和压力势等的测定技术已经广泛应用于推断切根和整根系统对水及溶质的渗透性, 根对水以及溶质的透性随着皮层木栓质的堆积而变化(Ranathunge and Schreiber, 2011).然而, 也有研究表明, 具有完整木栓质片层的成熟水稻根区的导水率与没有片层或部分片层的根尖区域的导水率并无显著差异(Ranathunge et al., 2005).因此, 木栓质片层可能并不显著影响细胞壁的透水性.这种现象的可能原因有: 细胞膜中的水通道蛋白可以显著改变膜的渗透性(Tyerman et al., 1999); 片层中的穿孔可以作为水自由进出细胞的位点(Waduwara et al., 2008); 非原生质体屏障中木栓质的堆积和多聚物的排列顺序均会对其透性产生影响(Schreiber et al., 2005a). ...
2 2005
... 20世纪90年代, 研究者普遍采用显微观察技术研究木栓化组织的结构及其复杂成分(Schreiber et al., 1999).之后, 结合GC-MS等技术, 不同物种、不同组织和不同环境下的特异木栓质可以被定量和定性描述(Zimmermann et al., 2000; Schreiber et al., 2005b).随着分析技术的成熟, 影响木栓质形成和堆积的因素逐渐清晰, 包括各种环境胁迫、机械损伤和激素处理.在环境胁迫如盐胁迫(Krishnamurthy et al., 2009)或缺氧(Kotula et al., 2009)条件下, 水稻(Oryza sativa)根外皮层的凯氏带和片层木栓质的堆积量显著增加.目前, 激素对木栓质形成和堆积的影响, 尤其是脱落酸(ABA)的作用, 逐渐成为研究热点. ... ... 木栓质坚固的结构及其疏水多聚物的化学特性在植物体内具有重要的生理作用.木栓质建立了组织屏障, 可以防止水分以及离子流失, 也可抵抗病原物的侵染(Machado et al., 2013).通过对环境胁迫下的植物组织化学特性进行研究, 已证明环境胁迫可以增强木栓化水平(Enstone et al., 2003).马铃薯块茎的损伤愈合也是通过形成木栓化的封闭层和损伤周皮发展而成.木栓化的细胞和相关疏水物质共同防卫其邻近的组织免受真菌以及微生物病原的侵染(Lulai and Corsini, 1998; Matsuda et al., 2003; Schreiber et al., 2005b). ...
1 2005
... 目前已经确定的参与木栓质合成的酶包括细胞色素P450单加氧酶、氧化还原酶、脂肪酸延长酶和过氧化物酶.它们分别参与脂肪酸ω-羟基化、二酸形成、脂肪酸延长和芳香类化合物聚合, 其活性可在多种植物组织的聚酯形成过程中检测到(Agrawal and Kola- ttukudy, 1977; Razem and Bernards, 2003; Schrei- ber et al., 2005c). ...
1 1999
... 20世纪90年代, 研究者普遍采用显微观察技术研究木栓化组织的结构及其复杂成分(Schreiber et al., 1999).之后, 结合GC-MS等技术, 不同物种、不同组织和不同环境下的特异木栓质可以被定量和定性描述(Zimmermann et al., 2000; Schreiber et al., 2005b).随着分析技术的成熟, 影响木栓质形成和堆积的因素逐渐清晰, 包括各种环境胁迫、机械损伤和激素处理.在环境胁迫如盐胁迫(Krishnamurthy et al., 2009)或缺氧(Kotula et al., 2009)条件下, 水稻(Oryza sativa)根外皮层的凯氏带和片层木栓质的堆积量显著增加.目前, 激素对木栓质形成和堆积的影响, 尤其是脱落酸(ABA)的作用, 逐渐成为研究热点. ...
1 2006
... 细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011).P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006).Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化.复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程.融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008).与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ...
2 2010
... 将马铃薯细胞色素P450单加氧酶(CYPs)基因CYP86A33沉默, 其周皮木栓质中将缺失ω-羟基脂肪酸和α, ω-二羧酸成分, 脂肪酸和脂肪醇类含量降低了50%, 甘油含量也降低了60%, 导致大分子结构紊乱(Graça and Pereira, 1997).与此类似, 在cyp- 86a1突变体中, ω-羟基脂肪酸和二羧酸含量明显减少, 导致根部周皮木栓质片层结构被破坏(Molina et al., 2009).以上结果表明, 木栓质组成成分或者含量的变化将显著影响木栓质片层结构(Schmutz et al., 1996; Lee et al., 2009).Kolattukudy (1981)和Bern- ards (2002)也都证实了长链双官能团成分的含量显著影响木栓质结构.然而, 对一些突变体木栓质成分的研究结果与这一模式并不一致.例如, cyp86b2突变体的种皮聚酯中, 单官能团脂肪类物质(羧酸类和脂肪醇类)的数量超过了双官能团(ω-羟基脂肪酸和α, ω-二羧酸)单体的含量(Compagnon et al., 2009; Molina et al., 2009).以上结果表明拟南芥木栓质的生物合成可以从羧酸类和脂肪醇类中产生聚酯, 并不一定需要氧化过程.此外, 在酰基转移酶基因ASFT/ AtHHT和FHT突变体的木栓质中, 酯连接的阿魏酸含量减少既未影响木栓质的不溶性, 也未影响透射电镜下的片层结构(Molina et al., 2009; Serra et al., 2010).虽然已经有较多的关于木栓质低聚物及其连接方面的研究, 但是, 对于多聚芳香类及多聚脂肪类物质聚合成大分子的结论性证据仍需进一步验证和研究. ... ... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
2 2009
... StKCS6在木栓化的马铃薯块茎周皮中高表达(Serra et al., 2009a).若下调StKCS6的表达丰度, 链长为C28和更长链脂肪酸的含量下降50%-95%, 而链长C26和更短链的化合物却发生堆积, 进一步证明StKCS6对脂肪酸链的延长有特定作用(Serra et al., 2009a).据此推测, StKCS6参与了木栓质前体脂肪酸链的延长. ... ... 在CYP86A1基因突变和CYP86A33基因沉默的植物中, 超长链ω-羟基酸和α, ω-二羧酸(>C20)并没有减少(Li et al., 2007; Höfer et al., 2008; Serra et al., 2009b), 表明有其它P450s参与木栓质单体合成.敲除或者沉默GPAT5和KCS2基因仅导致单体的部分损失, 影响部分脂肪酸链的延长, 说明还有1种或多种其它的GPATs和KCS参与此过程(Beisson et al., 2007; Serra et al., 2009a).转录水平研究证明, 渗透胁迫与盐胁迫可诱导KCS2和KCS20共同表达(Lee et al., 2009; Franke et al., 2009). ...
2 2009
... 细胞色素P450单加氧酶(P450s)催化脂肪酸ω-羟基化, 该反应是形成木栓质单体必不可少的步骤(Pinot and Beisson, 2011).P450s在拟南芥中为1个由272个基因编码的蛋白质家族(Schuler et al., 2006).Kandel等(2006)对拟南芥根的基因表达数据进行筛选, 在表达丰度排名前50的P450s基因中, 鉴定出属于CYP86亚家族的成员, 已经证实此亚家族可以催化脂肪酸ω-羟基化.复合体CYP86A1是植物中首个被鉴定出的C16和C18脂肪酸ω-羟基化酶(Benven- iste et al., 1998), 其相应的horst突变体研究进一步揭示了CYP86A1参与的生化过程.融合报告基因实验结果显示, CYP86A1特异性地在拟南芥根(尤其在根的内皮层)中表达(Höfer et al., 2008).与复合体CYP86A1的催化功能一致, 在cyp86a1/horst突变体的木栓质中, 羟基化C16和C18脂肪酸含量显著降低, 导致木栓质含量降低60% (Li et al., 2007; Höfer et al., 2008).与拟南芥cyp86a1突变体表型类似, 在马铃薯块茎中, 对CYP86A33基因进行RNA干扰导致C16与C18 ω-羟基酸和α, ω-二羧酸(DCAs)含量下降约70%-90% (Serra et al., 2009b). ... ... 在CYP86A1基因突变和CYP86A33基因沉默的植物中, 超长链ω-羟基酸和α, ω-二羧酸(>C20)并没有减少(Li et al., 2007; Höfer et al., 2008; Serra et al., 2009b), 表明有其它P450s参与木栓质单体合成.敲除或者沉默GPAT5和KCS2基因仅导致单体的部分损失, 影响部分脂肪酸链的延长, 说明还有1种或多种其它的GPATs和KCS参与此过程(Beisson et al., 2007; Serra et al., 2009a).转录水平研究证明, 渗透胁迫与盐胁迫可诱导KCS2和KCS20共同表达(Lee et al., 2009; Franke et al., 2009). ...
1 2003
... 脂肪酸合成以后, 一般会被长链酰基-CoA合成酶(LACSs)转化成酰基-CoA酯.最近的研究表明, 至少3个长链酰基-CoA合成异构酶参与拟南芥蜡质的合成, 它们催化游离脂肪酸活化为脂肪酰CoA硫酯(Lü et al., 2009; Weng and Chapple, 2010; Jessen et al., 2011).进一步研究显示, 拟南芥中包括9个属于酰基活化酶超家族的LACSs, 它们也参与脂肪酸的运输过程(Shockey and Fulda, 2003).LACS1-3是芽殖酵母(Saccharomyces cerevisiae)中脂类运输所必需的酶(Pulsifer et al., 2012).LACS1和2也参与了角质单体的合成, 激活脂肪酸生成酰基-CoA酯(Schnurr and Shockey, 2004).LACS的活性位点是在叶绿体被膜的内表面还是外表面仍不清楚. ...
2008
2 2007
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ... ... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
2 2007
... 木栓质片层超微结构可进一步通过透射电子显微镜观察.Schmutz等(1996)通过抑制脂肪酸链的延长, 发现木栓质浅色片层的厚度和脂肪酸的烷基链长度存在相关性.Bernards (2002)研究表明, 木栓质深色或者黑色条带和酚类物质的含量相关.酚类物质可以通过自发荧光或者样品染色后在显微镜下进行观察检测.但是, 这些检测不能区分木栓质酚类物质、木质素或者细胞壁中的其它酚类成分.此外, 木栓质形成早期或者少量时, 染色技术经常不够灵敏.例如, 对湿地植物Gyceria maxima根的横截面进行自发荧光检测, 结果发现酚类物质已经开始堆积, GC-MS分析也证实了皮下层组织已形成较多的脂肪类化合物.但是, 苏丹红7B或者荧光黄88染料并不能检测到木栓质(Soukup et al., 2007).与此类似, 在拟南芥种子发育后期, 仅种子合点区域可以被苏丹红染色(Beis- son et al., 2007; Franke et al., 2009). ... ... 湿地植物的外皮层或周皮组织, 普遍存在木栓质和木质素等非原生质体生化聚合物, 它们形成了一种渗氧(ROL)屏障, 阻碍氧气扩散到根际土壤中.同时, 这些木栓质屏障也阻碍了土壤和微生物释放的有毒气体渗透到根中(Visser et al., 2000; De Simone et al., 2003; Soukup et al., 2007; Garthwaite et al., 2008). ...
1 2002
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
... 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知.近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012).与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007).一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006). ...
1 1998
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
2016
1 2007
... 为探究木栓质单体间的相互作用, 前人通过多糖水解酶降解典型的木栓化组织或富含木栓质的细胞壁, 将分离下来的低分子量化合物进行色谱和质谱鉴定.之后, 将被鉴定的物质在原位进行重组.然而, 由于不同化合物及其分子间连接的复杂性, 这种方式并不可行(Thomas et al., 2007).也有研究者尝试利用碱水解和酯基转移作用清除脂肪类物质间的连接, 释放出脂肪酸衍生物, 让其仅和酚酸相连, 以此探究木栓质单体间的连接.但是, 这种适用于木质素的降解方法并不适用于检测木栓质单体间的作用.多酚类物质中, 醚的形成和C-C连接的可能性使其它类似处理也并未取得成功(Graça and Pereira, 2000).对马铃薯损伤周皮进行13C固态核磁共振光谱分析(NMR), 试图揭示木栓质多聚物中羧基功能的化学转变, 然而仅仅得到了追踪信号, 并未得到详细的功能变换数据(Bern- ards et al., 1995; Yan and Stark, 2000). ...
1 2004
... 角质和木栓质在脂肪聚酯的化学成分上有所不同, 角质以C16和C18脂肪聚酯为主, 而木栓质含有超长链脂肪单体及其聚酯(Franke et al., 2005; Pollard et al., 2008).因此, 木栓质合成的另一分子目标是脂肪酸延长酶FAE.KCS基因编码的β-酮酯CoA合成酶(KCS)是FAE复合体的组成部分, 是脂肪酸延长过程中的关键酶(Joubès et al., 2008).然而, 由于多数酶在功能和表达模式上存在广泛的冗余, 减慢了对KCS候选基因的鉴定.在拟南芥根中已发现至少7个高表达的KCS基因, 但是仅有3个参与长链C24木栓质单体的C骨架延长(Joubès et al., 2008).此外, 作为拟南芥根木栓质中的主要超长链单体C22, 至多有5个拟南芥KCS基因参与C22的延长(Trenkamp et al., 2004; Blacklock and Jaworski, 2006; Paul et al., 2006).对KCS突变体的木栓质研究也暗示, KCS可能参与其它需要超长链脂肪酸的过程(如膜脂形成), 进而影响木栓质单体生物合成的脂肪酸含量(Franke et al., 2009; Lee et al., 2009).同时, 这些酶或者基因的功能冗余现象减慢了相关候选基因的鉴定过程. ...
1 1999
... 根压探头技术和压力势等的测定技术已经广泛应用于推断切根和整根系统对水及溶质的渗透性, 根对水以及溶质的透性随着皮层木栓质的堆积而变化(Ranathunge and Schreiber, 2011).然而, 也有研究表明, 具有完整木栓质片层的成熟水稻根区的导水率与没有片层或部分片层的根尖区域的导水率并无显著差异(Ranathunge et al., 2005).因此, 木栓质片层可能并不显著影响细胞壁的透水性.这种现象的可能原因有: 细胞膜中的水通道蛋白可以显著改变膜的渗透性(Tyerman et al., 1999); 片层中的穿孔可以作为水自由进出细胞的位点(Waduwara et al., 2008); 非原生质体屏障中木栓质的堆积和多聚物的排列顺序均会对其透性产生影响(Schreiber et al., 2005a). ...
1 1997
... 饱和长链(C18、C20和C22)醇类物质是木栓质的普遍成分(Franke et al., 2005).对荷荷巴(Simmondsia chinensis)种子木栓质合成相关酶的研究表明, 以超长链脂肪酸为底物生成醇类的反应过程由脂肪酰还原酶(FAR)催化完成(Kolattukudy, 1971; Vioque and Kolattukudy, 1997; Metz et al., 2000).通过对酵母中超长链(C24和C26)醇类物质的产生过程进行研究, 确定了FAR的活性(Rowland and Domergue, 2012).脂肪酰还原酶(FARs)催化活化的脂肪酸还原为醇类物质.对FAR编码基因的研究显示, 拟南芥基因组中包括8个FAR基因序列, 分别编码FAR1-8, 其中1个序列被鉴定为CER4 (Rowland et al., 2006).在CER4突变体木栓质中, 链长C24-C28的主要醇类几乎完全缺失, 但是仍有部分C30醇类, 说明其它FAR或者相关的功能酶可能对C30醇的形成发挥作用(Rowl- and and Domergue, 2012).在研究FAR基因家族的表达模式时, 发现FAR1、FAR4和FAR5在拟南芥根内皮和其它含有木栓质的组织中出现, 相应的三突变体导致C18、C20和C22链长的木栓质醇类物质含量减少(Domergue et al., 2010), 表明FAR参与了根和种皮中木栓质长链(≥C18)醇类的合成. ...
2 2015
... 木栓质在根不同发育阶段的堆积位点(改自Vishwanath et al., 2015) 图中所示为典型双子叶植物(例如拟南芥)根的横切面.(A) 内胚层发育过程中细胞壁的初生生长阶段, 木栓质片层在其内表面有所积累(黄色), 凯氏带(红色)在内皮层细胞间的结合点处积累(ML: 中间层; PW: 初生细胞壁; SL: 木栓质片层; PM: 细胞质膜; Cy: 细胞质); (B) 次生生长阶段, 木栓质堆积于木栓组织/栓化细胞(周皮)中 ... ... Sites of suberin deposition in different developmental stages of roots (modified from Vishwanath et al., 2015) The diagrams show the root cross-sections of typical dicot plants (e.g. Arabidopsis thaliana). (A) Primary growth stage of cell wall during endodermis development with suberin lamellae deposition in the inner face of endodermal cell walls (yellow) and deposition of Casparian bands (red) localized at endodermal cell-cell junctions (ML: Middle lamellae; PW: Primary cell wall; SL: Suberin lamellae; PM: Plasma membrane; Cy: Cytoplasm); (B) Secondary growth stage with suberin deposition in the phellem/cork cells (periderms) ...
1 2000
... 湿地植物的外皮层或周皮组织, 普遍存在木栓质和木质素等非原生质体生化聚合物, 它们形成了一种渗氧(ROL)屏障, 阻碍氧气扩散到根际土壤中.同时, 这些木栓质屏障也阻碍了土壤和微生物释放的有毒气体渗透到根中(Visser et al., 2000; De Simone et al., 2003; Soukup et al., 2007; Garthwaite et al., 2008). ...
1 2008
... 根压探头技术和压力势等的测定技术已经广泛应用于推断切根和整根系统对水及溶质的渗透性, 根对水以及溶质的透性随着皮层木栓质的堆积而变化(Ranathunge and Schreiber, 2011).然而, 也有研究表明, 具有完整木栓质片层的成熟水稻根区的导水率与没有片层或部分片层的根尖区域的导水率并无显著差异(Ranathunge et al., 2005).因此, 木栓质片层可能并不显著影响细胞壁的透水性.这种现象的可能原因有: 细胞膜中的水通道蛋白可以显著改变膜的渗透性(Tyerman et al., 1999); 片层中的穿孔可以作为水自由进出细胞的位点(Waduwara et al., 2008); 非原生质体屏障中木栓质的堆积和多聚物的排列顺序均会对其透性产生影响(Schreiber et al., 2005a). ...
1 2006
... 通过将木栓质组织特异的转录数据与数据库候选基因序列进行比对, 已成功找到脂肪木栓质阿魏酰转移酶(ASFT)基因(Wei et al., 2006; Aoki et al., 2007; Obayashi et al., 2007).ASFT和马铃薯脂肪ω-羟基酸/脂肪醇羟基肉桂酰转移酶(FHT)属于酰基转移酶BAHD家族.BAHD蛋白以羟基肉桂酰-CoA酯作为酰基供体, 催化脂肪的阿魏酰化(D’Auria, 2006).拟南芥CYP86A1和GPAT5的共表达分析表明, BAHD家族基因ASFT也被显著共调控.因此, ASFT可作为木栓质酰基转移酶的候选基因(Molina et al., 2009).异源表达的ASFT蛋白特性描述进一步确定了ASFT催化酰基从阿魏酰-CoA转移到ω-羟基脂肪酸和脂肪醇上(Gou et al., 2009).FHT的活性也已经通过马铃薯切片实验得到证实, FHT-RNAi沉默的马铃薯周皮中, 酯连接的阿魏酸含量明显降低, 再次证明了FHT在马铃薯木栓化中发挥重要作用(Lotfy et al., 1994; Serra et al., 2010).此外, Soler等(2007, 2008)在栓皮栎中发现了另1种与ASFT高度相似的BAHD家族基因, 其在木栓组织和栓皮生长过程中高度上调表达, 但其基因功能还需要进一步确定. ...
1 2010
... 脂肪酸合成以后, 一般会被长链酰基-CoA合成酶(LACSs)转化成酰基-CoA酯.最近的研究表明, 至少3个长链酰基-CoA合成异构酶参与拟南芥蜡质的合成, 它们催化游离脂肪酸活化为脂肪酰CoA硫酯(Lü et al., 2009; Weng and Chapple, 2010; Jessen et al., 2011).进一步研究显示, 拟南芥中包括9个属于酰基活化酶超家族的LACSs, 它们也参与脂肪酸的运输过程(Shockey and Fulda, 2003).LACS1-3是芽殖酵母(Saccharomyces cerevisiae)中脂类运输所必需的酶(Pulsifer et al., 2012).LACS1和2也参与了角质单体的合成, 激活脂肪酸生成酰基-CoA酯(Schnurr and Shockey, 2004).LACS的活性位点是在叶绿体被膜的内表面还是外表面仍不清楚. ...
1 2001
... 有研究表明, 细胞壁并不是一种完美的离子屏障(Lux et al., 2004; Armstrong and Armstrong, 2005; Ranathunge et al., 2005).大部分研究表明, 木栓质具有较强的离子不透性(Barrowclough et al., 2000).在拟南芥根中, 木栓质与Ca2+通量间的密切关系已得到证实(White, 2001).由于木栓质的阻碍作用, 导致根对Ca2+的运输能力下降, 不恰当的木栓质障碍也导致离子沿非原生质体侧流, 使茎中Ca2+含量降低.拟南芥突变体(esb1)的茎中Ca2+含量与根中木栓质含量呈负相关, 其根中木栓质含量是对照组的2倍, 相应的茎中Ca2+含量则降低了50% (Baxter et al., 2009), 这进一步验证了以上结论.芦苇(Phragmites australis)和鸢尾(Iris tectorum)木栓化的根外皮阻止了非原生质体内Fe2+的流动(Soukup et al., 2002; Meyer et al., 2009).在玉米(Zea mays)根中, 皮层组织的木栓化降低了Rb+的透性(Clarkson et al., 1987).耐盐水稻品种(Pokkali)的根木栓化程度很高, Na+外流量很低.然而, 盐敏感品种(IR20)的根木栓化程度很低, 对应的Na+吸收量很高, 说明木栓化组织阻碍离子流入根部(Krishnamurthy et al., 2009).Krishnamurthy等(2011)再次证明, 水稻根的木栓质含量与流进茎中的Na+积累量呈负相关.在蓖麻(Ricinus communis) (Schreiber et al., 2005a)和拟南芥(Franke et al., 2009)根中, 也进一步验证了木栓质阻隔离子的作用. ...
1 2014
... 酰基单体、酰基甘油、阿魏酰偶联物或者其它预形成的二聚物或低聚物如何被引导并输出到质外体, 仍需进一步探究.这些物质是通过高尔基体的囊泡还是直接通过质膜泵或者借助ATP结合盒(ABC)转运蛋白被运输仍然未知(Li-Beisson et al., 2013).质膜上属于ABC转运蛋白超家族的G亚家族蛋白, 有助于表皮中孢粉素前体的输出(Pighin et al., 2004; Choi et al., 2011), 所以推测其可能也参与了木栓质的运输.例如, 在栓皮栎木栓质组织中, 编码G亚家族WBC型转运蛋白的基因, 其表达水平显著上调(Soler et al., 2007); 在马铃薯块茎的周皮内, 也发现ABCG1是木栓质形成所必需的蛋白(Landgraf et al., 2014).最近的研究显示, ABCG2、ABCG6和ABCG20参与拟南芥根内皮和种皮中木栓质片层的形成, 其相应的三突变体abcg2/abcg6/abcg20根部和种皮的木栓质结构、组成及性能均发生了变化(Yadav et al., 2014).此外, 参与角质聚酯形成的ABC转运蛋白ABCG11 (Pani- kashvili et al., 2007; Bird, 2008), 也直接或间接影响根木栓质含量(Panikashvili et al., 2010).ABC转运蛋白的详细生化特性研究可为聚酯前体的鉴定提供线索.但是, 木栓质在形成过程中其成分是不断变化的.在拟南芥根木栓质形成过程中, 由于前体物质的结构和大小不断变化, 膜转运蛋白的功能会受到限制.与此形成对比, 胞外运输机制或者其它膜融合过程能够使细胞内结构多样的低聚物和已经结合到细胞壁上的糖类大量输出.因此, 木栓质前体物质的输出可能涉及多种运输机制的协同作用. ...
2 2000
... 为探究木栓质单体间的相互作用, 前人通过多糖水解酶降解典型的木栓化组织或富含木栓质的细胞壁, 将分离下来的低分子量化合物进行色谱和质谱鉴定.之后, 将被鉴定的物质在原位进行重组.然而, 由于不同化合物及其分子间连接的复杂性, 这种方式并不可行(Thomas et al., 2007).也有研究者尝试利用碱水解和酯基转移作用清除脂肪类物质间的连接, 释放出脂肪酸衍生物, 让其仅和酚酸相连, 以此探究木栓质单体间的连接.但是, 这种适用于木质素的降解方法并不适用于检测木栓质单体间的作用.多酚类物质中, 醚的形成和C-C连接的可能性使其它类似处理也并未取得成功(Graça and Pereira, 2000).对马铃薯损伤周皮进行13C固态核磁共振光谱分析(NMR), 试图揭示木栓质多聚物中羧基功能的化学转变, 然而仅仅得到了追踪信号, 并未得到详细的功能变换数据(Bern- ards et al., 1995; Yan and Stark, 2000). ... ... 虽然对木栓质的组成仍不明晰, 但是通过其超微结构和已明确的单体物质, 可进行木栓质低聚物分析.Graça和Santos (2007)应用Ca(OH)2-甲醇分解法溶解了栓皮栎周皮大约10%的木栓质, 使用点喷射电离串联质谱对这些物质的结构进行测定, 鉴定的结构物包括ω-羟基脂肪酸-甘油二聚物、α, ω-二羧酸-甘油二聚物、ω-羟基脂肪酸-阿魏酰酯以及连接2个α, ω-二羧酸的甘油酯和ω-羟基脂肪酸-阿魏酰甘油酯, 成功呈现了木栓质中甘油-酰基和阿魏酸的连接.在马铃薯周皮中, 通过固态核磁共振方法也确定了这些组分与它们之间的相互连接, 且长链脂肪类物质、多酚类物质和细胞壁多糖彼此共价连接(Stark et al., 1994; Yan and Stark, 2000).据Bernards (2002)研究推测, 酚类物质先与初生细胞壁连接, 再与多聚脂肪类物质连接.其中, 苯丙烷代谢途径(PPP)的中间产物阿魏酸在建立和交联芳香类物质过程中可与脂肪区域相连.结合以上鉴定到的低聚物单元, 以及它们的功能和透射电镜下的片层结构, Graça和Santos (2007)得出了以两末端羧基分别连接甘油的长链α, ω-二羧酸为支柱骨架的木栓质大分子结构新模式: 这些支柱骨架单元堆积后的分子大小与透射电镜观察到的片层结构大小相符; 多芳香类物质含量与不透明片层厚度相关, 覆盖了聚酯区域; 酯化成酰基单元的阿魏酰连接聚酯和多芳香类. ...
2 2010
... 与CYP基因相似, GPAT5是木栓化过程中首批被鉴定的基因之一, 主要在根的内皮层组织中表达(Beis- son et al., 2007).对突变体gpat5的研究表明, 其编码的甘油-3-磷酸酰基转移酶(GPAT5)参与根和种皮木栓质中C22与C24超长链酰基单体的形成.在gpat5突变体根中, C22/C24单酰基甘油和超长链烷羟基肉桂酸酯的含量减少, 说明GPAT5在脂类聚合物的生物合成中发挥关键作用.与贮存型脂肪生物合成中GPATs的sn-1酰基化作用不同, 在木栓化过程中, GPAT5催化甘油的sn-2酰基化(Yang et al., 2010).与相关蛋白质的作用特性相比较, 仅在木栓质合成中发现了GPATs的sn-2酰基化作用.动物、微生物和藻类也缺失具有sn-2酰基化作用的GPATs, 这可能与陆地植物的进化过程相关(Yang et al., 2010). ... ... -2酰基化作用的GPATs, 这可能与陆地植物的进化过程相关(Yang et al., 2010). ...
1 2012
... 木栓质具有规律的片层结构, 说明聚合物装配是1个受调控的过程, 但是在自由聚合过程中, 是受到空间位阻还是蛋白调控或者其它因素的调控仍不得而知.近年来, 通过对番茄第一角质合酶(CD1)的鉴定, 已经可以较好地阐释表面脂类聚合物的堆积机理(Girard et al., 2012; Yeats et al., 2012).与此类似, 对于木栓质聚酯框架, 根据推断出的酯形成酶, 通过一种可以聚合脂肪的模型底物和脂肪酶, 已经在体外演示了聚酯形成过程(Olson and Sheares, 2006; Olsson et al., 2007).一些其它候选蛋白质在酯类聚合物形成中也可能发挥重要作用, 如GDSL-motif羧酸酯酶和BDG (BODYGUARD)家族的类脂酶蛋白(Suh et al., 2005; Kurdyukov et al., 2006). ...
1 2006
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2010
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2009
... 根据木栓质的发生和基因表达的空间分布关系, 推测木栓质合成的可能调控因子位于WRKY-、NAC-和MYB-特异转录因子区域.转录组数据显示, 这些转录因子家族成员在木栓化组织中优先表达(Kilian et al., 2007), 它们是与木栓质基因共表达的调控蛋白(Obayashi et al., 2009).这些调控因子在栓皮栎木栓组织中上调表达, 参与木栓化的相关过程(Soler et al., 2007).MYBs参与苯丙烷类物质的生物合成(Tamagnone et al., 1998; Bomal et al., 2008; Zhou et al., 2009)以及脂肪酸链的延长(Raffaele et al., 2008).NAC家族成员和乙烯响应(AP2/ERF)转录因子(AtERF38)参与次生细胞壁修饰(Mitsuda et al., 2005; Zhong et al., 2006, 2010; Lasserre et al., 2008).然而, 这些转录因子参与木栓化过程的直接证据还不充分, 尚需深入研究. ...
1 2000
... 20世纪90年代, 研究者普遍采用显微观察技术研究木栓化组织的结构及其复杂成分(Schreiber et al., 1999).之后, 结合GC-MS等技术, 不同物种、不同组织和不同环境下的特异木栓质可以被定量和定性描述(Zimmermann et al., 2000; Schreiber et al., 2005b).随着分析技术的成熟, 影响木栓质形成和堆积的因素逐渐清晰, 包括各种环境胁迫、机械损伤和激素处理.在环境胁迫如盐胁迫(Krishnamurthy et al., 2009)或缺氧(Kotula et al., 2009)条件下, 水稻(Oryza sativa)根外皮层的凯氏带和片层木栓质的堆积量显著增加.目前, 激素对木栓质形成和堆积的影响, 尤其是脱落酸(ABA)的作用, 逐渐成为研究热点. ...

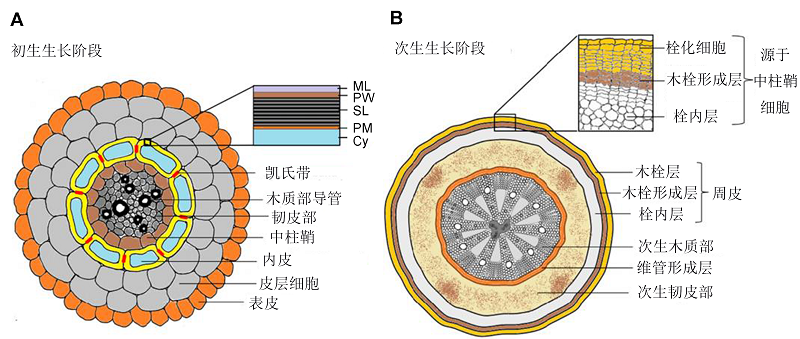
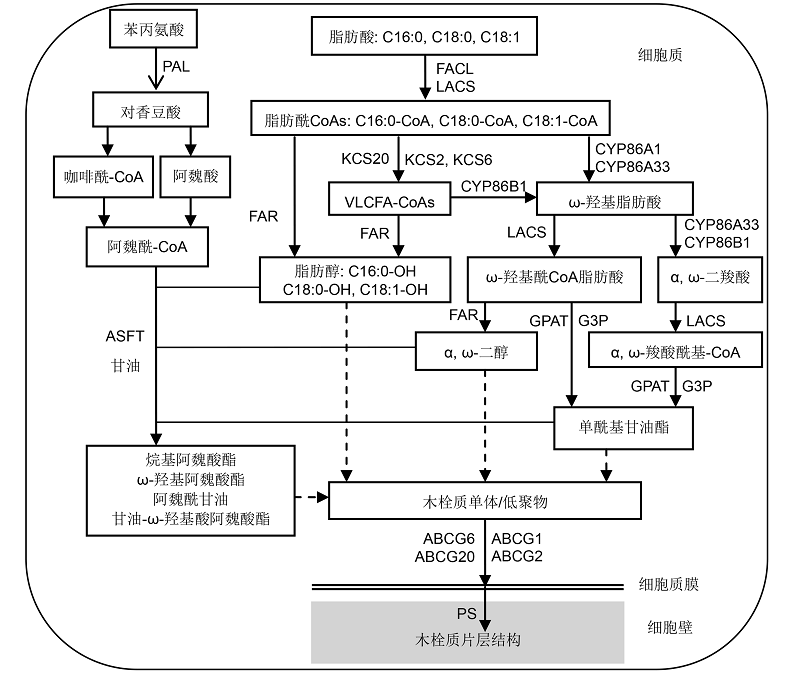

{kind=link}
{kind=link}
{kind=link}
{kind=link}