Abstract: The control of flowering time is a crucial environmental adaptation in plants; numerous CCT domain genes control flowering in plants. Bioinformatics was used for a genome-wide research of CCT domain genes in Aegilops tauschii. In this study, 26 CCT domain genes were identified in A. tauschii, distributed on seven chromosomes of the A. tauschii genome. The predicted molecular weight of this family was spread over 14.9 to 83.2 kDa, and 25 proteins contain a complete CCT conserved structure domain. Twelve pairs of A. tauschii-Triticum urartu and 9 pairs of A. tauschii-rice CCT proteins were orthologous in the phylogenetic tree. Specific expression and constitutive expression were found in the CCT gene family; nine AetCCT genes were constitutively expressed in all organisms, including AetCCT3, AetCCT4, AetCCT7 and AetCCT9; AetCCT15, AetCCT21 and AetCCT25 were specifically expressed in leaf, seed and roots of A. tauschii, respectively. Moreover, the members responded to phytohormone treatments differently, which suggested a complex function and characteristic in metabolism of this family. Light conditions affect the expression of AetCCT, and this gene family is regulated by photoperiod and vernalization. The results of this paper not only provide useful information for wheat evolution studies, but also provide theory basis for comprehensive understanding of formation and interaction characteristics of important traits.
Key words:Aegilops tauschii ; CCT gene family ; sequence analysis ; responses to phytohormones
小麦(Triticum aestivum)是世界上重要的粮食作物, 种植区域极为广泛, 目前只有少数与小麦花期和春化相关的基因被克隆。其中VRN1、VRN2与VRN3被认为是春化途径中3个关键基因(Yan et al., 2006); 而zCCT1、zCCT2、Ppd以及CO则是调控光周期的主要基因(Trevaskis et al., 2007; Dennis and Peacock, 2009)。小麦具有广泛的适应性, 能适应不同的地理和生态环境, 抽穗期不可能仅由少数几个基因控制, 因此尚有许多相关调控基因未被发现。而CCT家族基因在植物生长发育过程中起着非常重要的作用, 可能参与小麦驯化和改良过程。由于小麦基因组庞大, 导致其研究相对滞后, 与花期、产量、非生物逆境以及生长发育等方面相关的基因克隆进展缓慢。作为小麦D基因组供体, 粗山羊草(Aegilops tauschii)已经完成基因组测序, 功能基因组和单倍型图谱正在构建, 这将极大地促进小麦基因组研究, 也为小麦的驯化和传播研究提供重要思路和线索。目前, 水稻、拟南芥和大麦(Hordeum vulgare) CCT家族成员的组成已基本清楚(Cockram et al., 2012), 而小麦D基因组供体粗山羊草中CCT基因分类、数量及功能尚未见相关报道。本研究采用生物信息学方法分离了粗山羊草CCT家族基因(AetCCT), 进一步分析了该家族成员的染色体位置、序列特征、基因结构、表达、进化以及对外源激素的响应, 相关结果可为禾谷类作物CCT基因家族的深入研究与利用提供有价值的信息。
1 材料与方法1.1 数据来源粗山羊草(Aegilops tauschii Cosson.)基因组序列、蛋白序列和表达数据均来自AGDB数据库(http://dd. agrinome.org/)。粗山羊草定位序列片段和小麦(Triti- cum aestivum L.) EST标记信息来自GrainGene2.0数据库(http://wheat.pw.usda.gov/)。拟南芥(Arabi- dopsis thaliana (L.) Heynh.)、水稻 (Oryza sativa L.)和小麦CCT蛋白序列下载自NCBI (http://www.ncbi. nlm.nih.gov/)。
2 结果与讨论2.1 粗山羊草CCT家族基因的分离和染色体定位经登录PFAM网站查询, 得到CCT家族蛋白保守结构域编号为PF06203, 保守蛋白结构域序列为EAALL- RYKEKRKTRKFEKKIRYESRKAVAESRPRVKGR- FVKQA。我们利用隐马模型文件在本地蛋白数据库中进行检索及CCT保守结构域检测, 共获得26条粗山羊草CCT蛋白序列, 根据序列编号获得对应的CDS序列和基因组序列。利用GrainGene2.0数据库中的定位结果, 将得到的26条序列定位于粗山羊草的7条染色体上(图1)。根据所在染色体的位置信息, 将上述CCT基因依次命名为AetCCT1-26 (表1)。其中AetCCT5和AetCCT6位于同一条scaffold上, 是1对串联重复基因。 由定位结果可知, AetCCT各成员在7条染色体上均有分布, 在4D和7D上分布较多, 均为6个; AetCCT基因在1D、2D、3D、5D和6D上的分布数目分别为3、3、2、4和2个。在此基础上, 我们利用Y201和Y2272的F2:3家系进行抽穗期QTL和AetCCT共定位。从图1可以看出, 包含CCT基因的QTL共有7个, 除3D染色体上未定位到相关QTL, 其余染色体均有抽穗期相关QTL, 表明对应成员可能与粗山羊草抽穗相关。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_1.png<b>图1</b> 粗山羊草<i>CCT</i>家族基因的染色体定位和QTL定位<br/>左侧为遗传标尺, 黑色表示抽穗期QTL与<i>AetCCT</i>的共定位结果。<br/><b>Figure 1</b> Chromosome mapping and QTL distribution of <i>CCT</i> genes in <i>Aegilops tauschii</i> genome<br/>Genetic scale is indicated on the left side of chromosomes, the heading date QTLs are indicated on the right side of the chromosomes in black. Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_1.png<b>图1</b> 粗山羊草<i>CCT</i>家族基因的染色体定位和QTL定位<br/>左侧为遗传标尺, 黑色表示抽穗期QTL与<i>AetCCT</i>的共定位结果。<br/><b>Figure 1</b> Chromosome mapping and QTL distribution of <i>CCT</i> genes in <i>Aegilops tauschii</i> genome<br/>Genetic scale is indicated on the left side of chromosomes, the heading date QTLs are indicated on the right side of the chromosomes in black.
图1 粗山羊草CCT家族基因的染色体定位和QTL定位 左侧为遗传标尺, 黑色表示抽穗期QTL与AetCCT的共定位结果。 Figure 1 Chromosome mapping and QTL distribution of CCT genes in Aegilops tauschii genome Genetic scale is indicated on the left side of chromosomes, the heading date QTLs are indicated on the right side of the chromosomes in black.
2.2 AetCCT基因结构分析26个AetCCT基因在序列长度和内含子(外显子)数目等方面差异较大。序列最长的基因为AetCCT12, 其序列长度为4 521 bp; 而最短的是AetCCT15, 其序列长度仅为603 bp。通过聚类分析将26个AetCCT基因划分为5组(图2)。基因结构分析表明, AetCCT基因中分别包括1、6、5、4、4、3和3个成员, 分别包含0、1、2、3、4、5和7个内含子, 差异较大。从基因结构中发现, CCT各基序都是由其中较大的外显子编码。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_2.png<b>图2</b> 粗山羊草<i>AetCCT</i>基因结构<br/>棒状结构代表外显子部分, 线状代表内含子部分。<br/><b>Figure 2</b> Intron-exon structures of <i>CCT</i> genes in <i>Aegilops tauschii<br/></i>Exons are represented by bars, introns are represented by connecting lines. Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_2.png<b>图2</b> 粗山羊草<i>AetCCT</i>基因结构<br/>棒状结构代表外显子部分, 线状代表内含子部分。<br/><b>Figure 2</b> Intron-exon structures of <i>CCT</i> genes in <i>Aegilops tauschii<br/></i>Exons are represented by bars, introns are represented by connecting lines.
图2 粗山羊草AetCCT基因结构 棒状结构代表外显子部分, 线状代表内含子部分。 Figure 2 Intron-exon structures of CCT genes in Aegilops tauschii Exons are represented by bars, introns are represented by connecting lines.
2.3 AetCCT蛋白基序分析AetCCT蛋白的分子量变化范围较大, 从14.9 kDa (Aet- CCT3)至83.2 kDa (AetCCT12); 等电点范围从4.2 (AetCCT13)至10.2 (AetCCT14)。基序分析结果表明, AetCCT家族共包含4个基序(图3), 其中Motif1和Motif2为CCT保守结构域, Motif3为REC基序, Motif4为B-box基序(图4)。Logo图中显示Motif1和Motif2的保守性明显高于Motif3和Motif4。有25个(占96.2%) AetCCT蛋白具有Motif1和Motif2, 而AetCCT13包含了部分CCT结构, 只含有Motif2。另外, AetCCT4、7、9和12 (4个蛋白, 占15.4%)蛋白含有Motif3, AetCCT5、11、19、21、22和25 (6个蛋白, 占23.1%)蛋白含有Motif4。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_3.png<b>图3</b> 粗山羊草CCT蛋白基序的分布<br/><b>Figure 3</b> Motif distribution of AetCCT proteins in <i>Aegilops tauschii</i> Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_3.png<b>图3</b> 粗山羊草CCT蛋白基序的分布<br/><b>Figure 3</b> Motif distribution of AetCCT proteins in <i>Aegilops tauschii</i>
图5 CCT蛋白的系统发生 乌拉尔图小麦和粗山羊草的CCT蛋白分别用三角形和圆形标注。 Figure 5 Phylogenesis of CCT proteins CCT proteins of Triticum urartu and Aegilops tauschii were indicated by triangles and circles, respectively.
2.5 AetCCT家族基因启动子序列分析截取AetCCT基因起始密码子上游1 500 bp的基因组序列, 对启动子区参与激素响应的顺式作用组件进行分析。在启动子区发现与不同激素响应有关的9种顺式作用组件, 分别是与脱落酸响应有关的ABRE Motif、与MeJA调控相关的CGTCA Motif、与GA4调控相关的GARE Motif、P-Box和TATC-box、与IAA相关的TGA-Element和AuxRR Core Motif、与乙烯响应有关的ERE Motif及与SA调控相关的TCA-Element。从图6可以看出, 启动子区域顺式作用组件的类型在AetCCT成员间存在差异, 暗示AetCCT家族成员可通过不同的激素调控途径广泛参与粗山羊草的生长发育。 图6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_6.png<b>图6</b> 粗山羊草<i>CCT</i>基因启动子区激素响应的顺式作用组件<br/><b>Figure 6</b> <i>Cis</i>-elements in <i>CCT</i> genes promoters in response to phytohormones of <i>Aegilops tauschii</i> Figure 6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_6.png<b>图6</b> 粗山羊草<i>CCT</i>基因启动子区激素响应的顺式作用组件<br/><b>Figure 6</b> <i>Cis</i>-elements in <i>CCT</i> genes promoters in response to phytohormones of <i>Aegilops tauschii</i>
图8 粗山羊草CCT家族基因在不同激素处理下的表达分析 (A) 处理24小时; (B) 处理72小时 Figure 8 Hierarchical clustering of CCT domain genes under different phytohormones treatments in Aegilops tauschii (A) Phytohormones treatment for 24 h; (B) Phytohormones treatment for 72 h
2.8 AetCCT基因在不同光照条件下的表达分析CCT家族基因多位于植物开花调控路径的上游, 受光周期调控, 因此我们进一步分析了AetCCT成员在春化和不同光照条件下的基因表达(图9)。结果表明, 在不同光照条件下各成员间基因表达差异较大。AetCCT1、4、11、17、19和22在长日照条件下表达量较高; 而AetCCT3、5、18、21和26在短日照条件下表达量较高。春化也明显影响AetCCT基因的表达, 经春化后AetCCT1、3、17和18的表达量升高, 而AetCCT5、9、11、12、16和20表达量降低。可见, 光照和春化是影响和调控AetCCT表达的重要因素。此外, 处于同一进化分支的AetCCT基因对于光照条件往往具有相似的响应。例如, AetCCT4、9、12和17在长日照条件下表达量较高, 而AetCCT18、21和26在短日照条件下表达量较高, 暗示处于相同进化分支的AetCCT成员可能具有相似的功能。 图9https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_9.png<b>图9</b> 粗山羊草<i>CCT</i>家族基因在不同光照条件下的表达分析<br/>NV-LD: 未春化-长日照(16小时光照/8小时黑暗); NV-SD: 未春化-短日照(8小时光照/16小时黑暗); LD: 春化-长日照(16小时光照/8小时黑暗); SD: 春化-短日照(8小时光照/16小时黑暗)。<br/><b>Figure 9</b> Expression of <i>CCT</i> domain genes under different light treatments in <i>Aegilops tauschii<br/></i>NV-LD: Long-day condition without vernalization (16 h light/8 h dark); NV-SD: Short-day condition without vernalization (8 h light/16 h dark); LD: Long-day condition after vernalization (16 h light/8 h dark); SD: Short-day condition after vernalization (8 h light/16 h dark). Figure 9https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-2-188/img_9.png<b>图9</b> 粗山羊草<i>CCT</i>家族基因在不同光照条件下的表达分析<br/>NV-LD: 未春化-长日照(16小时光照/8小时黑暗); NV-SD: 未春化-短日照(8小时光照/16小时黑暗); LD: 春化-长日照(16小时光照/8小时黑暗); SD: 春化-短日照(8小时光照/16小时黑暗)。<br/><b>Figure 9</b> Expression of <i>CCT</i> domain genes under different light treatments in <i>Aegilops tauschii<br/></i>NV-LD: Long-day condition without vernalization (16 h light/8 h dark); NV-SD: Short-day condition without vernalization (8 h light/16 h dark); LD: Long-day condition after vernalization (16 h light/8 h dark); SD: Short-day condition after vernalization (8 h light/16 h dark).
图9 粗山羊草CCT家族基因在不同光照条件下的表达分析 NV-LD: 未春化-长日照(16小时光照/8小时黑暗); NV-SD: 未春化-短日照(8小时光照/16小时黑暗); LD: 春化-长日照(16小时光照/8小时黑暗); SD: 春化-短日照(8小时光照/16小时黑暗)。 Figure 9 Expression of CCT domain genes under different light treatments in Aegilops tauschii NV-LD: Long-day condition without vernalization (16 h light/8 h dark); NV-SD: Short-day condition without vernalization (8 h light/16 h dark); LD: Long-day condition after vernalization (16 h light/8 h dark); SD: Short-day condition after vernalization (8 h light/16 h dark).
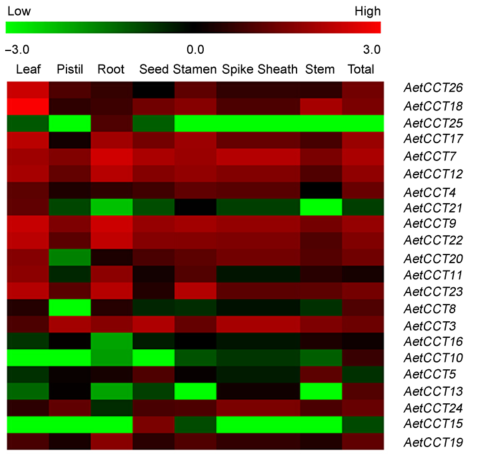
2.9 讨论开花是植物重要的生命过程, 对生育周期与产量影响较大。CCT基因在裸子植物和被子植物中广泛存在, 多数成员在花期的调控过程中发挥重要作用(Ando et al., 2013; Riboni et al., 2013)。该基因家族编码的蛋白质分子量也明显不同, 等电点分布十分广泛, 包括从酸性到碱性的各种蛋白。如水稻Os12g01080编码的蛋白质最小, 仅有147个氨基酸残基; 而Os03g- 17570编码的蛋白质最大, 含有768个氨基酸残基(陈华夏等, 2010)。此外, CCT基因含有丰富的结构域类型, 不仅基因结构差异较大, 而且编码的蛋白质理化性质差别也很大(Hsu et al., 2013)。本研究分离到的26个AetCCT在序列长度和基因结构上差异较大, 序列最长的基因AetCCT12长为4 521 bp, 而序列最短的AetCCT15仅长603 bp, 编码蛋白质远远大于水稻。Cockram等(2007)通过整合拟南芥、水稻、高粱(Sorghum bicolor)和大麦的最新测序信息对CCT基因进行了重新统计并分为4类, 而AetCCT家族也包含4个亚族。此外, 我们发现AetCCT5和AetCCT6为串联重复排列, 可能存在拷贝数变异位置效应和剂量效应。粗山羊草CCT蛋白经聚类分析可划分为9个组, 相关成员在进化树的各分支都有分布。AetCCT与小麦CCT蛋白之间亲缘关系最近, 其次为乌拉尔图小麦、水稻和拟南芥。该结果符合禾本科植物物种形成的一般规律。此外, AetCCT17独立于各分支, 具有物种特异性, 暗示其可能有新的功能。可见CCT基因在物种间具有一定的保守性, 在物种分化过程中部分成员在结构和功能上发生分化以适应相应环境。 研究表明, 植物开花受环境信号和内源激素共同调控, 主要包括自主开花途径、春化途径、光调控途径以及赤霉素信号途径(Rademacher et al., 2011)。CCT基因的功能在不同物种间发生了很大分化, 不仅参与调控开花途径, 还影响植物产量、繁殖和适应性等性状(Zhang et al., 2015)。例如, Ghd7在长日照条件下抑制水稻开花, 同时突变体的株高和单株产量明显提高; 玉米(Zea mays) Ghd7的同源基因则是通过CACTA转座子顺式组件的甲基化程度来调控对光照的敏感度, 从而影响驯化后的传播(Xue et al., 2008; Yang et al., 2015)。分析基因在特定组织的表达丰度是研究基因功能的重要方法。以往研究表明, 光周期基因往往仅在茎尖和幼穗等发育器官中表达, 从而决定相关器官的发育进程(Yan et al., 2004)。 Xiao等(2014)发现受春化诱导表达的Jacalin类凝集素蛋白VER2具有部分取代春化的作用, 可在叶片、茎 和茎尖等部位通过磷酸化与TaGRP2作用启动春化开花。AetCCT有2种表达模式。一种为组成型表达, 在各生育时期和各组织中均有表达。例如, AetCCT9、AetCCT7、AetCCT12、AetCCT17和AetCCT22在大部分生长部位表达量较高, 这种类型的AetCCT基因可能参与植株不同的生长发育过程(Grasser et al., 2005; Lolas et al., 2010)。另一类AetCCT基因表达具有较强的组织特异性, 只在生长锥、叶片和幼穗等光反应器官中表达, 暗示这类基因为光周期基因, 主要参与花期的调控。在此基础上, 本研究探讨了Aet- CCT对外源激素的响应, 进一步探讨不同成员参与调控的机制。结果表明, AetCCT不同成员对6种激素有不同的响应机制, 所有成员对GA4非常敏感, GA4可以抑制AetCCT17和AetCCT22的表达, 其余成员在GA4处理条件下表达量升高, 尤其在处理24小时后AetCCT8最为敏感, 比对照增强了12倍。由此推测, 大部分AetCCT基因是通过赤霉素信号途径参与粗山羊草的光反应代谢。IAA和NAA几乎参与植物所有的生长代谢过程(Vanneste and Friml, 2009; Strader et al., 2010)。AetCCT基因参与IAA和NAA代谢途径的成员也较多, 且各成员对2种激素具有明显的协同效应。例如, IAA和NAA对AetCCT1、AetCCT19及Aet- CCT20的表达都有促进作用, 而AetCCT12和Aet- CCT22等基因的表达则受到抑制。此外, 从组织特异表达结果来看, 对IAA和NAA敏感的成员多为组成型表达, 推测AetCCT主要是通过这2类激素调控网络参与粗山羊草的生长代谢。进一步分析发现, 不同AetCCT基因在激素响应方面还表现出结构域类型相似的特点。例如, AetCCT1、AetCCT3、AetCCT18和AetCCT26都含有Motif1, 施用IAA、NAA和GA4的响应方式为增加表达量。相同结构域成员对于激素也有不同的响应方式。例如, AetCCT5、AetCCT19、AetCCT21和AetCCT22都含有Motif1与Motif4, 但IAA和NAA对AetCCT5与AetCCT19具有促进作用, 对AetCCT21及AetCCT22则表现为抑制表达, 可能与N端功能结构域的功能歧化有关。施用激素24小时和72小时各成员对激素响应整体趋势表现一致, 72小时处理中大部分基因与施用24小时组相比表达量降低, 可能是由于植株自身代谢或是与外源激素降解有关。激素响应结果进一步验证了CCT家族基因功能在物种进化中存在较高的保守性, 而结构相似的基因呈现的表达模式也存在差异。物种形成和进化是一个漫长的过程, 而AetCCT上游调控机制存在差异, 导致部分家族成员发生功能分化, 一些成员在防御和应激等方面更广泛地参与植物的生长繁殖过程。如Yang等(2013)发现在玉米中无转座子插入ZmCCT抗病等位基因后, 玉米苗期的抗病性与光周期无关, 而抗病的ZmCCT等位基因可同时介导玉米对低氮和高盐的抗性。AetCCT8、AetCCT17和AetCCT26在MeJA和SA存在的条件下表达量升高, 而这2种激素多与应激和抗病虫有关, 进一步说明了AetCCT基因功能的多样性。 现已发现CCT家族基因多位于植物开花调控路径的上游。目前对拟南芥相关基因的研究较为深入, 相关调控路径已基本清楚, 如TIMEING OF CAB EXPRESS1、ZEITLUPE、TEJ、TIME FOR CO- FFEE、LUXARRHYTHMO和FIONA1等(Strayer et al., 2000; Panda et al., 2002; Kim et al., 2008)。通过筛选不同日照条件下控制植物生物进程的突变体, 如开花时间和下胚轴伸长等, 相继发现了EARLY FLO- WERING3、EARLY FLOWERING4、GIGANTEA和LATE ELONGATED等基因(Hicks et al., 2001; Doyle et al., 2002)。本研究通过抽穗期QTL和Aet- CCT共定位, 发现包含CCT基因的QTL共有7个, 这些成员可能与对应QTL的抽穗期相关, 在今后的研究中应重点关注。此外, 在不同光照和春化条件下, AetCCT成员基因的表达差异很大, 这与Cockram等(2012)的研究结果一致, 进一步证实了光照条件与春化作用是影响和调控AetCCT表达的重要因素。借助粗山羊草基因组测序进行CCT基因家族生物信息学和相关表达研究, 有助于系统地了解该家族基因的进化, 并为小麦遗传改良提供有价值的信息。 致谢 中国农业科学院耿帅峰博士在相关实验中给予指导和帮助, 谨致谢忱。
The authors have declared that no competing interests exist.
AndoE, OhnishiM, WangY, MatsushitaT, WatanabeA, HayashiY, FujiiM, MaJF, InoueS, KinoshitaT (2013). TWIN SISTER OF FT,GIGANTEA, and CONSTANS ha- ve a positive but indirect effect on blue light-induced stom- atal opening in Arabidopsis. Plant Physiol 162, 1529-1538. [本文引用: 1]
[3]
ChoLH, ChoiH, AnG (2010). OsCOL4 is a constitutive flowering repressor upstream of Ehd1 and downstream of OsphyB.Plant J 63, 8-30.
[4]
CockramJ, JonesH, LeighFJ, O’SullivanD, PowellW (2007). Control of flowering time in temperate cereals: genes, domestication and sustainable productivity.J Exp Bot 58, 1231-1244.
[5]
CockramJ, ThielT, SteuernagelB, SteinN, TaudienS, BaileyPC, SullivanDM (2012). Genome dynamics explain the evolution of flowering time CCT domain gene families in the Poaceae.PLoS One 7, e45307. [本文引用: 1]
GrasserKD (2005). Emerging role for transcript elongation in plant development.Trends Plant Sci 10, 484-490. [本文引用: 1]
[11]
HarmonF, ImaizumiT, GrayWM (2008). CUL1 regulates TOC1 protein stability in the Arabidopsis circadian clock.Plant J 55, 568-579.
[12]
HicksKA, AlbertsonTM, WagnerDR (2001). EARLY FLO- WERING3 encodes a novel protein that regulates circadian clock function and flowering in Arabidopsis.Plant Cell 13, 1281-1292. [本文引用: 1]
[13]
HsuCY, AdamsJP, NoK, LiangH, MeilanR, PechanovaO, BarakatA, CarlsonJE, PageGP, YuceerC (2012). Overexpression ofCONSTANS homologs CO1 and CO2 fails to alter normal reproductive onset and fall bud set in woody perennial poplar. PLoS One 7, e45448. [本文引用: 1]
[14]
KimJ, KimY, YeomM, KimJH, NamHG (2008). FIONA1 is essential for regulating period length in the Arabidopsis circadian clock. Plant Cell 20, 307-319. [本文引用: 1]
[15]
KimSK, ParkHY, JangYH, LeeJH, KimJK (2013). The sequence variation responsible for the functional difference between the CONSTANS protein, and the CONSTANS-like COL1 and COL2 proteins, resides mostly in the region encoded by their first exons. Plant Sci 199-200, 71-78.
[16]
LeeYS, JeongDH, LeeDY, YiJ, RyuCH, KimSL, JeongHJ, ChoiSC, JinP, YangJ, RademacherEH, MollerB, LokerseAS, Llavata-PerisCI, van den BergW, WeijersD (2011). A cellular expression map of the ArabidopsisAUXIN RESPONSE FACTOR gene family. Plant J 68, 597-606.
[17]
LolasIB, HimanenK, GronlundJT, LynggaardC, HoubenA, MelzerM, Van LijsebettensM, GrasserKD (2010). The transcript elongation factor FACT affects Arabidopsis vegetative and reproductive development and genetically interacts with HUB1/2.Plant J 61, 686-697. [本文引用: 1]
[18]
PandaS, PoirierGG, KaySA (2002). Tej defines a role for poly(ADP-ribosyl)ation in establishing period length of the Arabidopsis circadian oscillator.Dev Cell 3, 51-61. [本文引用: 1]
[19]
PutterillJ, RobsonF, LeeK, SimonR, CouplandG (1995). TheCONSTANS gene of Arabidopsis promotes flowering and encodes a protein showing similarities to zinc finger transcription factors. Cell 80, 847-857.
[20]
RiboniM, GalbiatiM, TonelliC, ContiL (2013). GIGANTEA enables drought escape response via abscisic acid- dependent activation of the florigens andSUPPRESSOR OF OVEREXPRESSION OF CONSTANS. Plant Physiol 162, 1706-1719. [本文引用: 1]
StrayerC, OyamaT, SchultzTF, RamanR, SomersDE, MásP, PandaS, KrepsJA, KaySA (2000). Cloning of the Arabidopsis clock geneTOC1, an autoregulatory response regulator homolog. Science 289, 768-771. [本文引用: 2]
[23]
TrevaskisB, HemmingMN, DennisES, PeacockWJ (2007). The molecular basis of vernalization-induced flo- wering in cereals.Trends Plant Sci 12, 352-357. [本文引用: 1]
[24]
VannesteS, FrimlJ (2009). Auxin: a trigger for change in plant development.Cell 136, 1005-1016. [本文引用: 1]
[25]
WuF, PriceB, HaiderW, SeufferheldG, NelsonR, HanzawaY (2014). Functional and evolutionary characterization of theCONSTANS gene family in short-day photoperiodic flowering in soybean. PLoS One 9, e85754.
[26]
WuWX, ZhengXM, LuGW, ZhongZZ, GaoH, ChenLP, WuCY, WangHJ, WangQ, ZhouKN, WangJL, WuFQ, ZhangX, GuoXP, ChengZJ, LeiCL, LinQB, JiangL, WangHY, GeS, WanJM (2013). Association of functional nucleotide polymorphisms atDTH2 with the northward expansion of rice cultivation in Asia. Proc Natl Acad Sci USA 110, 2775-2780.
[27]
XiaoJ, XuS, LiC, XuY, XingL, NiuY, HuanQ, TangY, ZhaoC, WagnerD, GaoC, ChongK (2014). O-GlcNAc- mediated interaction between ?VER2 and ?TaGRP2 elicits ?TaVRN1 mRNA accumulation during vernalization in win- ter wheat. Nat Commun 5, 4572-4578.
[28]
XueWY, XingYZ, WengXY, ZhaoY, TangWJ, WangL, ZhouHJ, YuSB, XuCG, LiXH, ZhangQF (2008). Natural variation inGhd7 is an important regulator of hea- ding date and yield potential in rice. Nat Genet 40, 761-767. [本文引用: 1]
[29]
YanL, FuDL, LiC, BlechlA, TranquilliG, BonafedeM, SanchezA, ValarikM, YasudaS, DubcovskyJ (2006). The wheat and barley vernalization geneVRN3 is an orthologue of FT. Proc Natl Acad Sci USA 103, 19581-19586. [本文引用: 1]
[30]
YanL, LoukoianovA, BlechlA, TranquilliG, Ramakri- shnaW, SanMiguelP, BennetzenJL, EcheniqueV, DubcovskyJ (2004). The wheatVRN2 gene is a flowering repressor down-regulated by vernalization. Science 303, 1640-1644. [本文引用: 1]
[31]
YangQ, LiZ, LiWQ, KuLX, WangC, YeJR, LiK, YangN, LiYP, ZhongT, LiJS, ChenYH, YanJB, YangXH, XuML (2013). CACTA-like transposable element inZmCCT attenuated photoperiod sensitivity and accelerated the postdomestication spread of maize. Proc Natl Acad Sci USA 110, 16969-16974. [本文引用: 1]
[32]
YanoM, InoueH, TanisakaT (2012). Ef7 encodes an ELF3-like protein and promotes rice flowering by negatively regulating the floral repressor geneGhd7 under both short- and long-day conditions. Plant Cell Physiol 53, 717-728.
[33]
ZhangL, LiQP, DongHJ, HeQ, LiangLW, TanC, HanZM, YaoW, LiGW, ZhaoH, XieWB, XinYZ (2015). Three CCT domain-containing genes were identified to regulate heading date by candidate gene-based association mapping and transformation in rice. Sci Rep 5, 7663. [本文引用: 1]
... 小麦(Triticum aestivum)是世界上重要的粮食作物, 种植区域极为广泛, 目前只有少数与小麦花期和春化相关的基因被克隆.其中VRN1、VRN2与VRN3被认为是春化途径中3个关键基因(Yan et al., 2006); 而zCCT1、zCCT2、Ppd以及CO则是调控光周期的主要基因(Trevaskis et al., 2007; Dennis and Peacock, 2009).小麦具有广泛的适应性, 能适应不同的地理和生态环境, 抽穗期不可能仅由少数几个基因控制, 因此尚有许多相关调控基因未被发现.而CCT家族基因在植物生长发育过程中起着非常重要的作用, 可能参与小麦驯化和改良过程.由于小麦基因组庞大, 导致其研究相对滞后, 与花期、产量、非生物逆境以及生长发育等方面相关的基因克隆进展缓慢.作为小麦D基因组供体, 粗山羊草(Aegilops tauschii)已经完成基因组测序, 功能基因组和单倍型图谱正在构建, 这将极大地促进小麦基因组研究, 也为小麦的驯化和传播研究提供重要思路和线索.目前, 水稻、拟南芥和大麦(Hordeum vulgare) CCT家族成员的组成已基本清楚(Cockram et al., 2012), 而小麦D基因组供体粗山羊草中CCT基因分类、数量及功能尚未见相关报道.本研究采用生物信息学方法分离了粗山羊草CCT家族基因(AetCCT), 进一步分析了该家族成员的染色体位置、序列特征、基因结构、表达、进化以及对外源激素的响应, 相关结果可为禾谷类作物CCT基因家族的深入研究与利用提供有价值的信息. ...
1 2009
... 小麦(Triticum aestivum)是世界上重要的粮食作物, 种植区域极为广泛, 目前只有少数与小麦花期和春化相关的基因被克隆.其中VRN1、VRN2与VRN3被认为是春化途径中3个关键基因(Yan et al., 2006); 而zCCT1、zCCT2、Ppd以及CO则是调控光周期的主要基因(Trevaskis et al., 2007; Dennis and Peacock, 2009).小麦具有广泛的适应性, 能适应不同的地理和生态环境, 抽穗期不可能仅由少数几个基因控制, 因此尚有许多相关调控基因未被发现.而CCT家族基因在植物生长发育过程中起着非常重要的作用, 可能参与小麦驯化和改良过程.由于小麦基因组庞大, 导致其研究相对滞后, 与花期、产量、非生物逆境以及生长发育等方面相关的基因克隆进展缓慢.作为小麦D基因组供体, 粗山羊草(Aegilops tauschii)已经完成基因组测序, 功能基因组和单倍型图谱正在构建, 这将极大地促进小麦基因组研究, 也为小麦的驯化和传播研究提供重要思路和线索.目前, 水稻、拟南芥和大麦(Hordeum vulgare) CCT家族成员的组成已基本清楚(Cockram et al., 2012), 而小麦D基因组供体粗山羊草中CCT基因分类、数量及功能尚未见相关报道.本研究采用生物信息学方法分离了粗山羊草CCT家族基因(AetCCT), 进一步分析了该家族成员的染色体位置、序列特征、基因结构、表达、进化以及对外源激素的响应, 相关结果可为禾谷类作物CCT基因家族的深入研究与利用提供有价值的信息. ...
2004
1 2002
... 现已发现CCT家族基因多位于植物开花调控路径的上游.目前对拟南芥相关基因的研究较为深入, 相关调控路径已基本清楚, 如TIMEING OF CAB EXPRESS1、ZEITLUPE、TEJ、TIME FOR CO- FFEE、LUXARRHYTHMO和FIONA1等(Strayer et al., 2000; Panda et al., 2002; Kim et al., 2008).通过筛选不同日照条件下控制植物生物进程的突变体, 如开花时间和下胚轴伸长等, 相继发现了EARLY FLO- WERING3、EARLY FLOWERING4、GIGANTEA和LATE ELONGATED等基因(Hicks et al., 2001; Doyle et al., 2002).本研究通过抽穗期QTL和Aet- CCT共定位, 发现包含CCT基因的QTL共有7个, 这些成员可能与对应QTL的抽穗期相关, 在今后的研究中应重点关注.此外, 在不同光照和春化条件下, AetCCT成员基因的表达差异很大, 这与Cockram等(2012)的研究结果一致, 进一步证实了光照条件与春化作用是影响和调控AetCCT表达的重要因素.借助粗山羊草基因组测序进行CCT基因家族生物信息学和相关表达研究, 有助于系统地了解该家族基因的进化, 并为小麦遗传改良提供有价值的信息. ...
2013
1 2005
... 研究表明, 植物开花受环境信号和内源激素共同调控, 主要包括自主开花途径、春化途径、光调控途径以及赤霉素信号途径(Rademacher et al., 2011).CCT基因的功能在不同物种间发生了很大分化, 不仅参与调控开花途径, 还影响植物产量、繁殖和适应性等性状(Zhang et al., 2015).例如, Ghd7在长日照条件下抑制水稻开花, 同时突变体的株高和单株产量明显提高; 玉米(Zea mays) Ghd7的同源基因则是通过CACTA转座子顺式组件的甲基化程度来调控对光照的敏感度, 从而影响驯化后的传播(Xue et al., 2008; Yang et al., 2015).分析基因在特定组织的表达丰度是研究基因功能的重要方法.以往研究表明, 光周期基因往往仅在茎尖和幼穗等发育器官中表达, 从而决定相关器官的发育进程(Yan et al., 2004). Xiao等(2014)发现受春化诱导表达的Jacalin类凝集素蛋白VER2具有部分取代春化的作用, 可在叶片、茎 和茎尖等部位通过磷酸化与TaGRP2作用启动春化开花.AetCCT有2种表达模式.一种为组成型表达, 在各生育时期和各组织中均有表达.例如, AetCCT9、AetCCT7、AetCCT12、AetCCT17和AetCCT22在大部分生长部位表达量较高, 这种类型的AetCCT基因可能参与植株不同的生长发育过程(Grasser et al., 2005; Lolas et al., 2010).另一类AetCCT基因表达具有较强的组织特异性, 只在生长锥、叶片和幼穗等光反应器官中表达, 暗示这类基因为光周期基因, 主要参与花期的调控.在此基础上, 本研究探讨了Aet- CCT对外源激素的响应, 进一步探讨不同成员参与调控的机制.结果表明, AetCCT不同成员对6种激素有不同的响应机制, 所有成员对GA4非常敏感, GA4可以抑制AetCCT17和AetCCT22的表达, 其余成员在GA4处理条件下表达量升高, 尤其在处理24小时后AetCCT8最为敏感, 比对照增强了12倍.由此推测, 大部分AetCCT基因是通过赤霉素信号途径参与粗山羊草的光反应代谢.IAA和NAA几乎参与植物所有的生长代谢过程(Vanneste and Friml, 2009; Strader et al., 2010).AetCCT基因参与IAA和NAA代谢途径的成员也较多, 且各成员对2种激素具有明显的协同效应.例如, IAA和NAA对AetCCT1、AetCCT19及Aet- CCT20的表达都有促进作用, 而AetCCT12和Aet- CCT22等基因的表达则受到抑制.此外, 从组织特异表达结果来看, 对IAA和NAA敏感的成员多为组成型表达, 推测AetCCT主要是通过这2类激素调控网络参与粗山羊草的生长代谢.进一步分析发现, 不同AetCCT基因在激素响应方面还表现出结构域类型相似的特点.例如, AetCCT1、AetCCT3、AetCCT18和AetCCT26都含有Motif1, 施用IAA、NAA和GA4的响应方式为增加表达量.相同结构域成员对于激素也有不同的响应方式.例如, AetCCT5、AetCCT19、AetCCT21和AetCCT22都含有Motif1与Motif4, 但IAA和NAA对AetCCT5与AetCCT19具有促进作用, 对AetCCT21及AetCCT22则表现为抑制表达, 可能与N端功能结构域的功能歧化有关.施用激素24小时和72小时各成员对激素响应整体趋势表现一致, 72小时处理中大部分基因与施用24小时组相比表达量降低, 可能是由于植株自身代谢或是与外源激素降解有关.激素响应结果进一步验证了CCT家族基因功能在物种进化中存在较高的保守性, 而结构相似的基因呈现的表达模式也存在差异.物种形成和进化是一个漫长的过程, 而AetCCT上游调控机制存在差异, 导致部分家族成员发生功能分化, 一些成员在防御和应激等方面更广泛地参与植物的生长繁殖过程.如Yang等(2013)发现在玉米中无转座子插入ZmCCT抗病等位基因后, 玉米苗期的抗病性与光周期无关, 而抗病的ZmCCT等位基因可同时介导玉米对低氮和高盐的抗性.AetCCT8、AetCCT17和AetCCT26在MeJA和SA存在的条件下表达量升高, 而这2种激素多与应激和抗病虫有关, 进一步说明了AetCCT基因功能的多样性. ...
2008
1 2001
... 现已发现CCT家族基因多位于植物开花调控路径的上游.目前对拟南芥相关基因的研究较为深入, 相关调控路径已基本清楚, 如TIMEING OF CAB EXPRESS1、ZEITLUPE、TEJ、TIME FOR CO- FFEE、LUXARRHYTHMO和FIONA1等(Strayer et al., 2000; Panda et al., 2002; Kim et al., 2008).通过筛选不同日照条件下控制植物生物进程的突变体, 如开花时间和下胚轴伸长等, 相继发现了EARLY FLO- WERING3、EARLY FLOWERING4、GIGANTEA和LATE ELONGATED等基因(Hicks et al., 2001; Doyle et al., 2002).本研究通过抽穗期QTL和Aet- CCT共定位, 发现包含CCT基因的QTL共有7个, 这些成员可能与对应QTL的抽穗期相关, 在今后的研究中应重点关注.此外, 在不同光照和春化条件下, AetCCT成员基因的表达差异很大, 这与Cockram等(2012)的研究结果一致, 进一步证实了光照条件与春化作用是影响和调控AetCCT表达的重要因素.借助粗山羊草基因组测序进行CCT基因家族生物信息学和相关表达研究, 有助于系统地了解该家族基因的进化, 并为小麦遗传改良提供有价值的信息. ...
... 现已发现CCT家族基因多位于植物开花调控路径的上游.目前对拟南芥相关基因的研究较为深入, 相关调控路径已基本清楚, 如TIMEING OF CAB EXPRESS1、ZEITLUPE、TEJ、TIME FOR CO- FFEE、LUXARRHYTHMO和FIONA1等(Strayer et al., 2000; Panda et al., 2002; Kim et al., 2008).通过筛选不同日照条件下控制植物生物进程的突变体, 如开花时间和下胚轴伸长等, 相继发现了EARLY FLO- WERING3、EARLY FLOWERING4、GIGANTEA和LATE ELONGATED等基因(Hicks et al., 2001; Doyle et al., 2002).本研究通过抽穗期QTL和Aet- CCT共定位, 发现包含CCT基因的QTL共有7个, 这些成员可能与对应QTL的抽穗期相关, 在今后的研究中应重点关注.此外, 在不同光照和春化条件下, AetCCT成员基因的表达差异很大, 这与Cockram等(2012)的研究结果一致, 进一步证实了光照条件与春化作用是影响和调控AetCCT表达的重要因素.借助粗山羊草基因组测序进行CCT基因家族生物信息学和相关表达研究, 有助于系统地了解该家族基因的进化, 并为小麦遗传改良提供有价值的信息. ...
2013
2011
1 2010
... 研究表明, 植物开花受环境信号和内源激素共同调控, 主要包括自主开花途径、春化途径、光调控途径以及赤霉素信号途径(Rademacher et al., 2011).CCT基因的功能在不同物种间发生了很大分化, 不仅参与调控开花途径, 还影响植物产量、繁殖和适应性等性状(Zhang et al., 2015).例如, Ghd7在长日照条件下抑制水稻开花, 同时突变体的株高和单株产量明显提高; 玉米(Zea mays) Ghd7的同源基因则是通过CACTA转座子顺式组件的甲基化程度来调控对光照的敏感度, 从而影响驯化后的传播(Xue et al., 2008; Yang et al., 2015).分析基因在特定组织的表达丰度是研究基因功能的重要方法.以往研究表明, 光周期基因往往仅在茎尖和幼穗等发育器官中表达, 从而决定相关器官的发育进程(Yan et al., 2004). Xiao等(2014)发现受春化诱导表达的Jacalin类凝集素蛋白VER2具有部分取代春化的作用, 可在叶片、茎 和茎尖等部位通过磷酸化与TaGRP2作用启动春化开花.AetCCT有2种表达模式.一种为组成型表达, 在各生育时期和各组织中均有表达.例如, AetCCT9、AetCCT7、AetCCT12、AetCCT17和AetCCT22在大部分生长部位表达量较高, 这种类型的AetCCT基因可能参与植株不同的生长发育过程(Grasser et al., 2005; Lolas et al., 2010).另一类AetCCT基因表达具有较强的组织特异性, 只在生长锥、叶片和幼穗等光反应器官中表达, 暗示这类基因为光周期基因, 主要参与花期的调控.在此基础上, 本研究探讨了Aet- CCT对外源激素的响应, 进一步探讨不同成员参与调控的机制.结果表明, AetCCT不同成员对6种激素有不同的响应机制, 所有成员对GA4非常敏感, GA4可以抑制AetCCT17和AetCCT22的表达, 其余成员在GA4处理条件下表达量升高, 尤其在处理24小时后AetCCT8最为敏感, 比对照增强了12倍.由此推测, 大部分AetCCT基因是通过赤霉素信号途径参与粗山羊草的光反应代谢.IAA和NAA几乎参与植物所有的生长代谢过程(Vanneste and Friml, 2009; Strader et al., 2010).AetCCT基因参与IAA和NAA代谢途径的成员也较多, 且各成员对2种激素具有明显的协同效应.例如, IAA和NAA对AetCCT1、AetCCT19及Aet- CCT20的表达都有促进作用, 而AetCCT12和Aet- CCT22等基因的表达则受到抑制.此外, 从组织特异表达结果来看, 对IAA和NAA敏感的成员多为组成型表达, 推测AetCCT主要是通过这2类激素调控网络参与粗山羊草的生长代谢.进一步分析发现, 不同AetCCT基因在激素响应方面还表现出结构域类型相似的特点.例如, AetCCT1、AetCCT3、AetCCT18和AetCCT26都含有Motif1, 施用IAA、NAA和GA4的响应方式为增加表达量.相同结构域成员对于激素也有不同的响应方式.例如, AetCCT5、AetCCT19、AetCCT21和AetCCT22都含有Motif1与Motif4, 但IAA和NAA对AetCCT5与AetCCT19具有促进作用, 对AetCCT21及AetCCT22则表现为抑制表达, 可能与N端功能结构域的功能歧化有关.施用激素24小时和72小时各成员对激素响应整体趋势表现一致, 72小时处理中大部分基因与施用24小时组相比表达量降低, 可能是由于植株自身代谢或是与外源激素降解有关.激素响应结果进一步验证了CCT家族基因功能在物种进化中存在较高的保守性, 而结构相似的基因呈现的表达模式也存在差异.物种形成和进化是一个漫长的过程, 而AetCCT上游调控机制存在差异, 导致部分家族成员发生功能分化, 一些成员在防御和应激等方面更广泛地参与植物的生长繁殖过程.如Yang等(2013)发现在玉米中无转座子插入ZmCCT抗病等位基因后, 玉米苗期的抗病性与光周期无关, 而抗病的ZmCCT等位基因可同时介导玉米对低氮和高盐的抗性.AetCCT8、AetCCT17和AetCCT26在MeJA和SA存在的条件下表达量升高, 而这2种激素多与应激和抗病虫有关, 进一步说明了AetCCT基因功能的多样性. ...
1 2002
... 现已发现CCT家族基因多位于植物开花调控路径的上游.目前对拟南芥相关基因的研究较为深入, 相关调控路径已基本清楚, 如TIMEING OF CAB EXPRESS1、ZEITLUPE、TEJ、TIME FOR CO- FFEE、LUXARRHYTHMO和FIONA1等(Strayer et al., 2000; Panda et al., 2002; Kim et al., 2008).通过筛选不同日照条件下控制植物生物进程的突变体, 如开花时间和下胚轴伸长等, 相继发现了EARLY FLO- WERING3、EARLY FLOWERING4、GIGANTEA和LATE ELONGATED等基因(Hicks et al., 2001; Doyle et al., 2002).本研究通过抽穗期QTL和Aet- CCT共定位, 发现包含CCT基因的QTL共有7个, 这些成员可能与对应QTL的抽穗期相关, 在今后的研究中应重点关注.此外, 在不同光照和春化条件下, AetCCT成员基因的表达差异很大, 这与Cockram等(2012)的研究结果一致, 进一步证实了光照条件与春化作用是影响和调控AetCCT表达的重要因素.借助粗山羊草基因组测序进行CCT基因家族生物信息学和相关表达研究, 有助于系统地了解该家族基因的进化, 并为小麦遗传改良提供有价值的信息. ...
 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}