Abstract: The plasma membrane (PM) is a crucial barrier between the protoplast and environment. In addition to maintaining normal cellular homeostasis and plant nutrient status, the PM perceives and responds to various environmental stress. In recent years, plant PM proteomic investigations provide valuable information for insights into the molecular mechanisms of the plant in response to different biotic and abiotic stimuli. Plant PM proteomics have revealed diverse PM protein expression patterns in 10 plant species including Arabidopsis thaliana and Oryza sativa in response to biotic stress (e.g., Xanthomonas oryzae pv. oryzae infection) and abiotic stress (e.g., cold, salt, flooding, osmosis, high pH, Fe, nitrogen, abscisic acid, chitosan, and chitooligosaccharide). Studies have revealed the important role of the plant plasma membrane in response to stress by integrative analysis of plant PM proteomic information from recent publications, including regulation of intracellular and extracellular signal transmission; material exchange and transport by transporter, channel protein and vesicle transport-related proteins; and perception and transmission of the stress signal by signal transduction pathways, such as membrane-associated G protein, Ca2+ signaling transduction, the phosphoinositide signaling pathway, the brassinosteroid signal pathway and reversible phosphorylation of proteins, adapting the plant to stress. The research provides new clues at the protein level for understanding the molecular regulation mechanism of the PM stress response.
AlexanderssonE, SaalbachG, LarssonC, KjellbomP (2004). Arabidopsis plasma membrane proteomics identifies components of transport, signal transduction and membrane trafficking.Plant Cell Physiol 45, 1543-1556.
[5]
BaisakhN, RamanaRaoMV, RajasekaranK, SubudhiP, JandaJ, GalbraithD, VanierC, PereiraA (2012). Enhanced salt stress tolerance of rice plants expressing a vacuolar H+-ATPase subunit c1 (SaVHAc1) gene from the halophyte grass Spartina alterniflora Löisel.Plant Biotechnol J 10, 453-464. [本文引用: 1]
[6]
BariolaPA, RetelskaD, StasiakA, KammererRA, FlemingA, HijriM, FrankS, FarmerEE (2004). Remorins form a novel family of coiled coil-forming oligomeric and filamentous proteins associated with apical, vascular and embryonic tissues in plants.Plant Mol Biol 55, 579-594. [本文引用: 1]
[7]
BloomAJ, SukrapannaSS, WarnerRL (1992). Root respiration associated with ammonium and nitrate absorption and assimilation by barley.Plant Physiol 99, 1294-1301. [本文引用: 1]
BunneyTD, van den WijngaardPW, de BoerAH (2002). 14-3-3 protein regulation of proton pumps and ion channels.Plant Mol Biol 50, 1041-1051. [本文引用: 1]
[10]
CameronKD, TeeceMA, SmartLB (2006). Increased accumulation of cuticular wax and expression of lipid transfer protein in response to periodic drying events in leaves of tree tobacco.Plant Physiol 140, 176-183. [本文引用: 2]
[11]
CharronJBF, BretonG, BadawiM, SarhanF (2002). Molecular and structural analyses of a novel temperature stress-induced lipocalin from wheat and Arabidopsis.FEBS Lett 517, 129-132. [本文引用: 3]
[12]
CharronJBF, OuelletF, PelletierM, DanylukJ, ChauveC, SarhanF (2005). Identification, expression, and evolutionary analyses of plant lipocalins.Plant Physiol 139, 2017-2028. [本文引用: 1]
[13]
CheckerVG, KhuranaP (2013). Molecular and functional characterization of mulberry EST encoding remorin (Mi- REM) involved in abiotic stress.Plant Cell Rep 32, 1729-1741. [本文引用: 1]
[14]
ChenF, LiQ, HeZ (2007a). Proteomic analysis of rice plasma membrane-associated proteins in response to chitooligosaccharide elicitors.J Integr Plant Biol 49, 863-870. [本文引用: 2]
[15]
ChenF, YuanY, LiQ, HeZ (2007b). Proteomic analysis of rice plasma membrane reveals proteins involved in early defense response to bacterial blight.Proteomics 7, 1529-1539. [本文引用: 5]
[16]
ChengY, QiY, ZhuQ, ChenX, WangN, ZhaoX, ChenH, CuiX, XuL, ZhangW (2009). New changes in the plasma-membrane-associated proteome of rice roots un- der salt stress.Proteomics 9, 3100-3114. [本文引用: 11]
[17]
ChuB, SnustadDP, CarterJV (1993). Alteration of β-tubulin gene expression during low-temperature exposure in leaves of Arabidopsis thaliana.Plant Physiol 103, 371-377. [本文引用: 1]
[18]
ClouseSD (2011). Brassinosteroid signal transduction: from receptor kinase activation to transcriptional networks regu- lating plant development.Plant Cell 23, 1219-1230. [本文引用: 3]
[19]
De LorenzoL, MerchanF, LaporteP, ThompsonR, ClarkeJ, SousaC, CrespiM (2009). A novel plant leucine-rich repeat receptor kinase regulates the response of Medicago truncatula roots to salt stress.Plant Cell 21, 668-680. [本文引用: 2]
[20]
DeFalcoTA, BenderKW, SneddenWA (2010). Breaking the code: Ca2+ sensors in plant signaling.Biochem J 425, 27-40. [本文引用: 1]
[21]
DrøbakBK, Franklin-TongVE, StaigerCJ (2004). The role of the actin cytoskeleton in plant cell signaling.New Phytol 163, 13-30. [本文引用: 1]
[22]
Ellard-IveyM, HopkinsRB, WhiteTJ, LomaxTL (1999). Cloning, expression and N-terminal myristoylation of Cp- CPK1, a calcium-dependent protein kinase from zucchini (Cucurbita pepo L.).Plant Mol Biol 39, 199-208. [本文引用: 1]
[23]
EllisC, KarafyllidisI, WasternackC, TurnerJG (2002). The Arabidopsis mutant cev1 links cell wall signaling to jasmonate and ethylene responses.Plant Cell 14, 1557-1566. [本文引用: 1]
[24]
FisherM, GokhmanI, PickU, ZamirA (1997). A structu- rally novel transferrin-like protein accumulates in the plasma membrane of the unicellular green alga Dunaliella salina grown in high salinities.J Biol Chem 272, 1565-1570. [本文引用: 1]
[25]
FukudaA, ChibaK, MaedaM, NakamuraA, MaeshimaM, TanakaY (2004). Effect of salt and osmotic stresses on the expression of genes for the vacuolar H+-pyropho- sphatase, H+-ATPase subunit A, and Na+/H+ antiporter from barley.J Exp Bot 55, 585-594. [本文引用: 1]
[26]
FulgosiH, SollJ, de Faria MaraschinS, KorthoutHA, WangM, TesterinkC (2002). 14-3-3 proteins and plant development.Plant Mol Biol 50, 1019-1029. [本文引用: 1]
[27]
GalenC, RabenoldJJ, LiscumE (2007). Functional eco- logy of a blue light photoreceptor: effects of phototropin-1 on root growth enhance drought tolerance in Arabidopsis thaliana.New Phytol 173, 91-99.
GazzarriniS, LejayL, GojonA, NinnemannO, FrommerWB, von WirénN (1999). Three functional transporters for constitutive, diurnally regulated, and starvation-induc- ed uptake of ammonium into Arabidopsis roots.Plant Cell 11, 937-947. [本文引用: 1]
[30]
GensJS, FujikiM, PickardBG (2000). Arabinogalactan- protein and wall-associated kinase in a plasmalemmal reticulum with specialized vertices.Protoplasma 212, 115-134. [本文引用: 1]
[31]
GerkeV, MossSE (2002). Annexins: from structure to function.Physiol Rev 82, 331-371. [本文引用: 1]
[32]
GómezJM, JiménezA, OlmosE, SevillaF (2004). Loca- tion and effects of long-term NaCl stress on superoxide dismutase and ascorbate peroxidase isoenzymes of pea (Pisum sativum cv. ‘Puget’) chloroplasts.J Exp Bot 55, 119-130. [本文引用: 1]
[33]
HanB, ChenS, DaiS, YangN, WangT (2010). Isobaric tags for relative and absolute quantification-based comparative proteomics reveals the features of plasma membrane-associated proteomes of pollen grains and pollen tubes from Lilium davidii.J Integr Plant Biol 52, 1043-1058. [本文引用: 1]
[34]
HarrisTJ, SiuCH (2002). Reciprocal raft-receptor inter- actions and the assembly of adhesion complexes.Bioessays 24, 996-1003. [本文引用: 1]
[35]
HashimotoM, ToorchiM, MatsushitaK, IwasakiY, KomatsuS (2009). Proteome analysis of rice root plasma membrane and detection of cold stress responsive proteins.Protein Pept Lett 16, 685-697. [本文引用: 4]
[36]
HinshawJE (2000). Dynamin and its role in membrane fission.Annu Rev Cell Dev Biol 16, 483-519. [本文引用: 1]
[37]
HoeflichKP, IkuraM (2002). Calmodulin in action: diversity in target recognition and activation mechanisms.Cell 108, 739-742. [本文引用: 1]
[38]
HolwegC, NickP (2004). Arabidopsis myosin XI mutant is defective in organelle movement and polar auxin transport.Proc Natl Acad Sci USA 101, 10488-10493. [本文引用: 2]
[39]
HongW (2005). SNAREs and traffic.BBA-Mol Cell Res 1744, 120-144. [本文引用: 1]
[40]
HongZ, Geisler-LeeCJ, ZhangZ, VermaDPS (2003). Phragmoplastin dynamics: multiple forms, microtubule association and their roles in cell plate formation in plants.Plant Mol Biol 53, 297-312. [本文引用: 1]
[41]
HopffD, WienkoopS, LüthjeS (2013). The plasma membrane proteome of maize roots grown under low and high iron conditions.J Proteomics 91, 605-618. [本文引用: 13]
[42]
HowittSM, UdvardiMK (2000). Structure, function and regulation of ammonium transporters in plants.BBA-Bio- membranes 1465, 152-170. [本文引用: 2]
[43]
HuangF, FuldaS, HagemannM, NorlingB (2006). Proteomic screening of salt-stress-induced changes in plas- ma membranes of Synechocystis sp. strain PCC 6803.Proteomics 6, 910-920. [本文引用: 4]
[44]
HuntL, OtterhagL, LeeJC, LasheenT, HuntJ, SekiM, ShinozakiK, SommarinM, GilmourDJ, PicalC, GrayJE (2004). Gene-specific expression and calcium activation of Arabidopsis thaliana phospholipase C isoforms.New Phytol 162, 643-654. [本文引用: 1]
[45]
HwangI, SzeH, HarperJF (2000). A calcium-dependent protein kinase can inhibit a calmodulin-stimulated Ca2+ pump (ACA2) located in the endoplasmic reticulum of Arabidopsis.Proc Natl Acad Sci USA 97, 6224-6229. [本文引用: 1]
[46]
JametE, CanutH, BoudartG, Pont-LezicaRF (2006). Cell wall proteins: a new insight through proteomics.Trends Plant Sci 11, 33-39. [本文引用: 1]
[47]
JangJY, KimDG, KimYO, KimJS, KangH (2004). An expression analysis of a gene family encoding plasma membrane aquaporins in response to abiotic stresses in Arabidopsis thaliana.Plant Mol Biol 54, 713-725. [本文引用: 2]
[48]
JohnsonKL, JonesBJ, BacicA, SchultzCJ (2003). The fasciclin-like arabinogalactan proteins of Arabidopsis. A multigene family of putative cell adhesion molecules.Pl- ant Physiol 133, 1911-1925. [本文引用: 4]
[49]
JonesAM, AssmannSM (2004). Plants: the latest model system for G-protein research.EMBO Rep 5, 572-578. [本文引用: 1]
KalampanayilBD, WimmersLE (2001). Identification and characterization of a salt-stress-induced plasma membrane H+-ATPase in tomato.Plant Cell Environ 24, 999-1005. [本文引用: 1]
[52]
KarpinskaB, KarlssonM, SchinkelH, StrellerS, SüssKH, MelzerM, WingsleG (2001). A novel superoxide dismutase with a high isoelectric point in higher plants. Expression, regulation, and protein localization.Plant Phy- siol 126, 1668-1677. [本文引用: 1]
[53]
KatzA, WaridelP, ShevchenkoA, PickU (2007). Salt- induced changes in the plasma membrane proteome of the halotolerant alga Dunaliella salina as revealed by blue native gel electrophoresis and nano-LC-MS/MS analysis.Mol Cell Proteomics 6, 1459-1472. [本文引用: 4]
[54]
KawamuraY, UemuraM (2003). Mass spectrometric approach for identifying putative plasma membrane proteins of Arabidopsis leaves associated with cold acclimation.Plant J 36, 141-154. [本文引用: 6]
[55]
KomatsuS, WadaT, AbaléaY, NouriMZ, NanjoY, NakayamaN, ShimamuraS, YamamotoR, NakamuraT, FurukawaK (2009). Analysis of plasma membrane proteome in soybean and application to flooding stress response.J Proteome Res 8, 4487-4499. [本文引用: 5]
[56]
Leborgne-CastelN, LherminierJ, DerC, FromentinJ, HouotV, Simon-PlasF (2008). The plant defense elicitor cryptogein stimulates clathrin-mediated endocytosis correlated with reactive oxygen species production in bright yellow-2 tobacco cells.Plant Physiol 146, 1255-1266. [本文引用: 1]
[57]
LeeJ, RuddJJ (2002). Calcium-dependent protein kinases: versatile plant signaling components necessary for patho- gen defence.Trends Plant Sci 7, 97-98. [本文引用: 1]
[58]
LeustekT, MartinMN, BickJA, DaviesJP (2000). Pathways and regulation of sulfur metabolism revealed throu- gh molecular and genetic studies.Annu Rev Plant Physiol 51, 141-165. [本文引用: 1]
[59]
LeustekT, SaitoK (1999). Sulfate transport and assimila- tion in plants.Plant Physiol 120, 637-644. [本文引用: 2]
[60]
LiB, TakahashiD, KawamuraY, UemuraM (2012a). Comparison of plasma membrane proteomic changes of Arabidopsis suspension-cultured cells (T87 Line) after cold and ABA treatment in association with freezing tole- rance development.Plant Cell Physiol 53, 543-554. [本文引用: 44]
[61]
LiJ, WenJ, LeaseKA, DokeJT, TaxFE, WalkerJC (2002). BAK1, an Arabidopsis LRR receptor-like protein kinase, interacts with BRI1 and modulates brassinosteroid signaling.Cell 110, 213-222. [本文引用: 1]
[62]
LiW, LiM, ZhangW, WeltiR, WangX (2004). The plasma membrane-bound phospholipase Dδ enhances freezing tolerance in Arabidopsis thaliana.Nat Biotechnol 22, 427-433.
[63]
LiZY, XuZS, HeGY, YangGX, ChenM, LiLC, MaYZ (2012b). A mutation in Arabidopsis BSK5 encoding a bras- sinosteroid-signaling kinase protein affects responses to salinity and abscisic acid.Biochem Biophs Res Commun 426, 522-527.
[64]
LinW (1981). Inhibition of anion transport in corn root protoplasts.Plant Physiol 68, 435-438. [本文引用: 1]
[65]
LiuX, BushDR (2006). Expression and transcriptional regulation of amino acid transporters in plants.Amino Acids 30, 113-120. [本文引用: 2]
MagnottaSM, GogartenJP (2002). Multi site polyadenyla- tion and transcriptional response to stress of a vacuolar type H+-ATPase subunit A gene in Arabidopsis thaliana.BMC Plant Biol 2, 1-13. [本文引用: 1]
[68]
MalakshahSN, RezaeiMH, HeidariM, SalekdehGH (2007). Proteomics reveals new salt responsive proteins associated with rice plasma membrane.Biosci Biotechnol Biochem 71, 2144-2154. [本文引用: 4]
[69]
MartínML, BusconiL (2000). Membrane localization of a rice calcium-dependent protein kinase (CDPK) is mediated by myristoylation and palmitoylation.Plant J 24, 429-435. [本文引用: 1]
[70]
MartínML, BusconiL (2001). A rice membrane-bound calcium-dependent protein kinase is activated in response to low temperature.Plant Physiol 125, 1442-1449. [本文引用: 1]
[71]
Maruyama-NakashitaA, InoueE, Watanabe-TakahashiA, YamayaT, TakahashiH (2003). Transcriptome profiling of sulfur-responsive genes in Arabidopsis reveals global effects of sulfur nutrition on multiple metabolic pa- thways.Plant Physiol 132, 597-605. [本文引用: 1]
[72]
MeisrimlerCN, PlanchonS, RenautJ, SergeantK, LüthjeS (2011). Alteration of plasma membrane-bound redox systems of iron deficient pea roots by chitosan.J Proteomics 74, 1437-1449. [本文引用: 6]
[73]
MicheletB, BoutryM (1995). The plasma membrane H+-ATPase (a highly regulated enzyme with multiple phy- siological functions).Plant Physiol 108, 1-6. [本文引用: 1]
[74]
MikaA, LüthjeS (2003). Properties of guaiacol peroxidase activities isolated from corn root plasma membranes.Pl- ant Physiol 132, 1489-1498. [本文引用: 1]
[75]
MillsLN, HuntL, LeckieCP, AitkenFL, WentworthM, McAinshMR, GrayJE, HetheringtonAM (2004). The effects of manipulating phospholipase C on guard cell ABA-signaling.J Exp Bot 55, 199-204. [本文引用: 1]
[76]
MinamiA, FujiwaraM, FurutoA, FukaoY, YamashitaT, KamoM, KawamuraY, UemuraM (2009). Alterations in detergent-resistant plasma membrane microdomains in Arabidopsis thaliana during cold acclimation.Plant Cell Physiol 50, 341-359. [本文引用: 8]
[77]
MishraNS, TutejaR, TutejaN (2006). Signaling through MAP kinase networks in plants.Arch Biochem Biophys 452, 55-68. [本文引用: 1]
[78]
MitsukawaN, OkumuraS, ShiranoY, SatoS, KatoT, HarashimaS, ShibataD (1997). Overexpression of an Arabidopsis thaliana high-affinity phosphate transporter gene in tobacco cultured cells enhances cell growth under phosphate-limited conditions.Proc Natl Acad Sci USA 94, 7098-7102. [本文引用: 1]
MorganA, DimalineR, BurgoyneRD (1994). The ATPase activity of N-ethylmaleimide-sensitive fusion protein (NSF) is regulated by soluble NSF attachment proteins. J Biol Chem 269, 29347-29350. [本文引用: 1]
[81]
MousleyCJ, TyeryarKR, Vincent-PopeP, BankaitisVA (2007). The Sec14-superfamily and the regulatory inter- face between phospholipid metabolism and membrane trafficking.Biochim Biophys Acta 1771, 727-736. [本文引用: 1]
[82]
MoustafaK, AbuQamarS, JarrarM, Al-RajabAJ, Trémouillaux-GuillerJ (2014). MAPK cascades and major abiotic stresses.Plant Cell Rep 33, 1217-1225. [本文引用: 2]
[83]
MunnikT (2001). Phosphatidic acid: an emerging plant lipid second messenger.Trends Plant Sci 6, 227-233. [本文引用: 1]
NieL, FengJ, FanP, ChenX, GuoJ, LvS, BaoH, JiaW, TaiF, JiangP, WangJ, LiY (2015). Comparative pro- teomics of root plasma membrane proteins reveals the involvement of calcium signaling in NaCl-facilitated nitrate uptake in Salicornia europaea.J Exp Bot 66, 4497-4510. [本文引用: 14]
[86]
NoEG, LoopstraCA (2000). Hormonal and developmental regulation of two arabinogalactan-proteins in xylem of loblolly pine (Pinus taeda).Physiol Plantarum 110, 524-529.
[87]
NouriMZ, KomatsuS (2010). Comparative analysis of soybean plasma membrane proteins under osmotic stress using gel-based and LC MS/MS-based proteomics approaches.Proteomics 10, 1930-1945. [本文引用: 22]
[88]
ÖrvarBL, SangwanV, OmannF, DhindsaRS (2000). Early steps in cold sensing by plant cells: the role of actin cytoskeleton and membrane fluidity.Plant J 23, 785-794.
PokornáJ, SchwarzerováK, ZelenkováS, PetrášekJ, JanotováI, ČapkováV, OpatrnýZ (2004). Sites of actin filament initiation and reorganization in cold-treated tobacco cells.Plant Cell Environ 27, 641-653.
[93]
PopovaOV, DietzKJ, GolldackD (2003). Salt-dependent expression of a nitrate transporter and two amino acid transporter genes in Mesembryanthemum crystallinum.Plant Mol Biol 52, 569-578. [本文引用: 2]
[94]
QiuQS, ZhangN (2000). Water stress inhibits p-nitrophenyl phosphate hydrolysis activity of the plasma membrane H+-ATPase from soybean hypocotyls.Aust J Plant Phy- siol 27, 717-721. [本文引用: 1]
[95]
RaghothamaKG (2000). Phosphate transport and signaling.Curr Opin Plant Biol 3, 182-187. [本文引用: 1]
[96]
RandallSK, SzeH (1986). Properties of the partially purified tonoplast H+-pumping ATPase from oat roots.J Biol Chem 261, 1364-1371. [本文引用: 1]
[97]
RentschD, HirnerB, SchmelzerE, FrommerWB (1996). Salt stress-induced proline transporters and salt stress- repressed broad specificity amino acid permeases identified by suppression of a yeast amino acid permease- targeting mutant.Plant Cell 8, 1437-1446. [本文引用: 2]
[98]
RobertsMR (2003). 14-3-3 proteins find new partners in plant cell signaling.Trends Plant Sci 8, 218-223. [本文引用: 1]
SaijoY, HataS, KyozukaJ, ShimamotoK, IzuiK (2000). Over-expression of a single Ca2+-dependent protein kin- ase confers both cold and salt/drought tolerance on rice plants.Plant J 23, 319-327. [本文引用: 2]
SchwarzM, ZamirA, PickU (2003). Iron-binding properties of TTf, a salt-induced transferrin from the alga Dunaliella salina.J Plant Nutr 26, 2081-2091. [本文引用: 1]
[104]
SeoHM, JungY, SongS, KimY, KwonT, KimDH, JeungSJ, YiYB, YiG, NamMH, NamJ (2008). Increased expression of OsPT1, a high-affinity phosphate transporter, enhances phosphate acquisition in rice.Biotechnol Lett 30, 1833-1838. [本文引用: 1]
[105]
ShibagakiN, RoseA, McDermottJP, FujiwaraT, HayashiH, YoneyamaT, DaviesJP (2002). Selenate-resistant mutants of Arabidopsis thaliana identify Sultr1; 2, a sulfate transporter required for efficient transport of sulfate into roots.Plant J 29, 475-486.
[106]
ShibaokaH (1994). Plant hormone-induced changes in the orientation of cortical microtubules: alterations in the cross- linking between microtubules and the plasma membrane.Annu Rev Plant Physiol 45, 527-544. [本文引用: 1]
[107]
ShitamichiN, MatsuokaD, SasayamaD, FuruyaT, NanmoriT (2013). Over-expression of MAP3Kδ4, an ABA-inducible Raf-like MAP3K that confers salt tolerance in Arabidopsis.Plant Biotechnol 30, 111-118. [本文引用: 1]
SongW, SteinerHY, ZhangL, NaiderF, StaceyG, BeckerJM (1996). Cloning of a second Arabidopsis peptide tran- sport gene.Plant Physiol 110, 171-178. [本文引用: 1]
[110]
StaceyG, KohS, GrangerC, BeckerJM (2002). Peptide transport in plants.Trends Plant Sci 7, 257-263. [本文引用: 1]
[111]
SurowyTK, BoyerJS (1991). Low water potentials affect expression of genes encoding vegetative storage proteins and plasma membrane proton ATPase in soybean.Plant Mol Biol 16, 251-262. [本文引用: 1]
[112]
SuttonRB, FasshauerD, JahnR, BrungerAT (1998). Crystal structure of a SNARE complex involved in synaptic exocytosis at 2.4 Å resolution.Nature 395, 347-353. [本文引用: 1]
[113]
TakahashiD, KawamuraY, UemuraM (2013). Changes of detergent-resistant plasma membrane proteins in oat and rye during cold acclimation: association with differential freezing tolerance.J Proteome Res 12, 4998-5011. [本文引用: 34]
[114]
TangD, AdeJ, FryeCA, InnesRW (2006). A mutation in the GTP hydrolysis site of Arabidopsis dynamin-related protein 1E confers enhanced cell death in response to powdery mildew infection.Plant J 47, 75-84. [本文引用: 1]
[115]
TesterinkC, MunnikT (2005). Phosphatidic acid: a multifunctional stress signaling lipid in plants.Trends Plant Sci 10, 368-375. [本文引用: 2]
[116]
TutejaN, SoporySK (2008). Chemical signaling under abiotic stress environment in plants.Plant Signal Behav 3, 525-536. [本文引用: 1]
UenoK, KinoshitaT, InoueSI, EmiT, ShimazakiKI (2005). Biochemical characterization of plasma memb- rane H+-ATPase activation in guard cell protoplasts of Ara- bidopsis thaliana in response to blue light.Plant Cell Phy- siol 46, 955-963.
[119]
VergnolleC, VaultierMN, TaconnatL, RenouJP, KaderJC, ZachowskiA, RuellandE (2005). The cold-induced early activation of phospholipase C and D pathways determines the response of two distinct clusters of genes in Arabidopsis cell suspensions.Plant Physiol 139, 1217-1233. [本文引用: 1]
[120]
VogelJP, RaabTK, SomervilleCR, SomervilleSC (2004). Mutations in PMR5 result in powdery mildew resistance and altered cell wall composition.Plant J 40, 968-978. [本文引用: 1]
[121]
Von WittgensteinNJ, LeCH, HawkinsBJ, EhltingJ (2014). Evolutionary classification of ammonium, nitrate, and peptide transporters in land plants.BMC Evol Biol 14, 11. [本文引用: 2]
[122]
WangX, LiX, MeisenhelderJ, HunterT, YoshidaS, AsamiT, ChoryJ (2005). Autoregulation and homodi- merization are involved in the activation of the plant ste- roid receptor BRI1.Dev Cell 8, 855-865. [本文引用: 1]
[123]
WasteneysGO, YangZ (2004). New views on the plant cytoskeleton.Plant Physiol 136, 3884-3891. [本文引用: 1]
[124]
WhitePJ, BowenHC, DemidchikV, NicholsC, DaviesJM (2002). Genes for calcium-permeable channels in the plasma membrane of plant root cells.BBA-Biomembranes 1564, 299-309. [本文引用: 1]
[125]
XiongL, SchumakerKS, ZhuJK (2002). Cell signaling during cold, drought, and salt stress.Plant Cell 14, S165-S183. [本文引用: 3]
[126]
XiongL, ZhuJK (2001). Abiotic stress signal transduction in plants: molecular and genetic perspectives.Physiol Plan- tarum 112, 152-166. [本文引用: 1]
[127]
YadetaKA, ElmoreJM, CoakerG (2013). Advancements in the analysis of the Arabidopsis plasma membrane pro- teome.Front Plant Sci 4, 1-10. [本文引用: 1]
[128]
YangYL, GuoJK, ZhangF, ZhaoLQ, ZhangLX (2004). NaCl induced changes of the H+-ATPase in root plasma membrane of two wheat cultivars.Plant Sci 166, 913-918. [本文引用: 1]
[129]
YangZ (2002). Small GTPases: versatile signaling switches in plants. Plant Cell 14, S375-S388. [本文引用: 1]
[130]
YazakiK (2006). ABC transporters involved in the transport of plant secondary metabolites.FEBS Lett 580, 1183-1191. [本文引用: 1]
[131]
YoshimotoN, TakahashiH, SmithFW, YamayaT, SaitoK (2002). Two distinct high-affinity sulfate transporters with different inducibilities mediate uptake of sulfate in Arabidopsis roots.Plant J 29, 465-473. [本文引用: 1]
[132]
ZhangH, HanB, WangT, ChenS, LiH, ZhangY, DaiS (2012). Mechanisms of plant salt response: insights from proteomics.J Proteome Res 11, 49-67. [本文引用: 1]
[133]
ZhangJ, MaH, FengJ, ZengL, WangZ, ChenS (2008). Grape berry plasma membrane proteome analysis and its differential expression during ripening.J Exp Bot 59, 2979-2990. [本文引用: 1]
[134]
ZhangLF, YangHM, CuiSX, HuJ, WangJ, KuangTY, NorlingB, HuangF (2009). Proteomic analysis of plasma membranes of cyanobacterium Synechocystis sp. strain PCC 6803 in response to high pH stress.J Proteome Res 8, 2892-2902. [本文引用: 4]
[135]
ZwiewkaM, NodzyńskiT, RobertS, VannesteS, FrimlJ (2015). Osmotic stress modulates the balance between exocytosis and clathrin-mediated endocytosis in Arabidopsis thaliana.Mol Plant 8, 1175-1187. [本文引用: 1]
NaHCO3胁迫下星星草根中Ca2+与Ca2+-ATPase的超微细胞化学定位 1 2010
... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ...
活性氧调控植物生长发育的研究进展 1 2012
... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
植物细胞质外体多肽与蛋白研究 1 2006
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
2004
1 2012
... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ...
1 2004
... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ...
1 1992
... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ...
1 1995
... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ...
1 2002
... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ...
2 2006
... 脂质转移蛋白(lipid transfer protein, LTP)可以促进脂质在植物细胞外基质间的转移, 参与胞外角质蜡质的形成与沉积, 以利于抵御病原体的侵害以及逆境(如水分和低温)胁迫应答(Kader, 1996; Cameron et al., 2006).有研究表明, 烟草叶片中NgLTP基因在脱水胁迫条件下表达量增加了6倍(Cameron et al., 2006).Nouri和Komatsu (2010)利用蛋白质组学相关技术对大豆进行了研究, 发现在渗透胁迫下大豆根与下胚轴中LTP丰度减少(图1A; 附表3), 这将降低角质层蜡质的沉积, 从而削弱植物的耐旱性. ... ... 基因在脱水胁迫条件下表达量增加了6倍(Cameron et al., 2006).Nouri和Komatsu (2010)利用蛋白质组学相关技术对大豆进行了研究, 发现在渗透胁迫下大豆根与下胚轴中LTP丰度减少(图1A; 附表3), 这将降低角质层蜡质的沉积, 从而削弱植物的耐旱性. ...
3 2002
... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... ).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... 基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ...
1 2005
... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ...
1 2013
... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ...
2 2007
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
5 2007
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
11 2009
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... ; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
1 1993
... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ...
1 2010
... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ...
1 2004
... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
1 1999
... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
1 2002
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
1 1997
... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ...
1 2004
... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ...
1 2002
... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ...
2007
1 2007
... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ...
1 1999
... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ...
1 2000
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
1 2002
... 膜联蛋白(annexin, ANX)是一种钙依赖的膜结合蛋白, 在膜运输、离子通道活性调节和磷脂代谢中均发挥作用(Gerke and Moss, 2002).蛋白质组学研究表明, 在冷及ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)和氮胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度减少; 在冷胁迫的黑麦叶片(Takahashi et al., 2013)和NaCl胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度增加(图1A; 附表3).植物ANX能在PM上形成钙离子通道, ANX丰度的变化可能改变钙通道活性, 引起钙离子跨膜运输变化, 进而调节钙离子的稳态和信号转导过程(White et al., 2002). ...
1 2004
... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
4 2009
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ...
1 2000
... 网格蛋白介导的内吞作用(clathrin-mediated endocytosis, CME)是网格蛋白作为受体介导内吞作用的主要方式, 是植物细胞内向膜泡运输(vesicle transport)的重要途径.除了基本内吞作用, 此过程也受到防御诱导子的调节(Leborgne-Castel et al., 2008).高渗胁迫会导致CME增强、胞吐作用减弱; 低渗胁迫的作用则相反(Zwiewka et al., 2015).Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根和下胚轴中网格蛋白的丰度降低(图1A; 附表3), 这将减弱网格蛋白被招募到PM形成网格蛋白包被小窝(clathrin coated pits, CCPs)的能力, 进而削弱内吞作用, 不利于大豆抵御渗透胁迫.在内吞作用过程中, 动力蛋白(dy- namin)作为GTPase参与调节CCPs从膜上脱离并形成网格蛋白包被小泡的过程(Hinshaw, 2000).拟南芥中, 动力蛋白参与抗病、线粒体分裂及内吞作用中网格蛋白包被小泡的细胞板形成和分裂(Hong et al., 2003; Tang et al., 2006).蛋白质组学研究表明, 冷胁迫下拟南芥幼苗中动力相关蛋白(dynamin-related protein)的丰度增加(Minami et al., 2009) (图1A; 附表4), 这将促进网格蛋白包被小泡的释放, 利于运输货物蛋白. ...
1 2002
... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ...
2 2004
... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
1 2005
... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ...
1 2003
... 网格蛋白介导的内吞作用(clathrin-mediated endocytosis, CME)是网格蛋白作为受体介导内吞作用的主要方式, 是植物细胞内向膜泡运输(vesicle transport)的重要途径.除了基本内吞作用, 此过程也受到防御诱导子的调节(Leborgne-Castel et al., 2008).高渗胁迫会导致CME增强、胞吐作用减弱; 低渗胁迫的作用则相反(Zwiewka et al., 2015).Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根和下胚轴中网格蛋白的丰度降低(图1A; 附表3), 这将减弱网格蛋白被招募到PM形成网格蛋白包被小窝(clathrin coated pits, CCPs)的能力, 进而削弱内吞作用, 不利于大豆抵御渗透胁迫.在内吞作用过程中, 动力蛋白(dy- namin)作为GTPase参与调节CCPs从膜上脱离并形成网格蛋白包被小泡的过程(Hinshaw, 2000).拟南芥中, 动力蛋白参与抗病、线粒体分裂及内吞作用中网格蛋白包被小泡的细胞板形成和分裂(Hong et al., 2003; Tang et al., 2006).蛋白质组学研究表明, 冷胁迫下拟南芥幼苗中动力相关蛋白(dynamin-related protein)的丰度增加(Minami et al., 2009) (图1A; 附表4), 这将促进网格蛋白包被小泡的释放, 利于运输货物蛋白. ...
13 2013
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... 水通道蛋白(aquaporin, AQP)属于膜整合蛋白家族, 能促进水分的跨膜运输.在已发现的35种拟南芥水通道蛋白中, 有13种PM整合蛋白(plasma membrane intrinsic protein, PIPs) (Jang et al., 2004).这些PIPs基因在应对各种胁迫(如干旱、冷、高盐和ABA)的过程中表达模式发生变化, 对维持植物的水分平衡起关键作用(Jang et al., 2004).蛋白质组学研究表明, 在燕麦叶片应对冷胁迫(Takahashi et al., 2013)及玉米根应对铁缺乏和过量(Hopff et al., 2013)时AQP丰度减少; 在拟南芥悬浮细胞应对ABA胁迫时AQP丰度增加(Li et al., 2012a) (图1A; 附表3), 进而调整胁迫下植物的水分与渗透平衡. ... ... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 油菜素内酯(brassinosteroids, BRs)是一类促生长的类固醇激素, 参与调节植物的生长发育过程(Clouse, 2011).可逆磷酸化反应是BR信号转导的主要调控方式.油菜素受体BRI1 (brassinosteroid insensitive 1)在细胞表面感知并结合BR, 发生自磷酸化(Wang et al., 2005).BRI1类受体激酶(brassinos- teroid insensitive 1-associated receptor kinase 1, BAK1)作为BRI1的共受体, 二者相互作用对BR信号转导尤为重要, BAK1与BRI1彼此磷酸化激活BRI1, 为下游调控因子提供作用位点, 将信号向下传递(Nam and Li, 2002; Clouse, 2011).过表达BAK1导致拟南芥器官伸长, 而BAK1缺失会引起植株半矮化并降低对BR的敏感性(Li et al., 2002).蛋白质组学研究表明, 冷胁迫下拟南芥悬浮细胞中, BRI1丰度上升或下降, 而ABA胁迫下的拟南芥悬浮细胞中, BRI1丰度增加(Li et al., 2012a); 铁缺乏和过量条件下, 玉米根中BAK1丰度减少(Hopff et al., 2013) (图1B; 附表3), 这些变化使BRI1、BAK1与BR的互作受到影响, 进而影响BR的信号传递.此外, Li等(2012b)研究发现, 拟南芥BSK5转录物在非生物胁迫(包括盐、干旱、植物激素BR和ABA)下上调表达, 且拟南芥bsk5功能缺失突变体对盐和ABA的敏感性增强.另外, 蛋白质组学研究还表明, 在冷和ABA胁迫的拟南芥悬浮细胞中, BR信号的正调节蛋白——油菜素内酯信号激酶(BR-signaling kinase, BSK)的丰度增加(Li et al., 2012a) (图1B; 附表4), 这不仅有助于BSKs被BRI1磷酸化激活, 而且可以促进BR的信号传递(Clouse, 2011)及BR靶基因的表达. ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
2 2000
... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... ; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ...
4 2006
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ... ... 依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ... ... ).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ...
1 2004
... 磷酸肌醇信号途径在植物生长、发育及应对逆境过程中具有重要作用.细胞膜受体接收胞外信号, 激活磷酸肌醇特异的磷脂酶C (phosphatidylinositol-specific phospholipase C, PI-PLC), 将磷脂酰肌醇-4,5-二磷酸水解生成1,4,5-肌醇三磷酸(IP3)和二酰甘油(DAG) (Testerink and Munnik, 2005).PLC通过调节2个重要的信使(IP3和DAG)参与植物各种信号途径.IP3刺激细胞外存储的Ca2+进入细胞质基质, 使胞内Ca2+浓度升高, Ca2+进一步调节信号转导途径(Hunt et al., 2004).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ...
1 2000
... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
1 2006
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
2 2004
... 水通道蛋白(aquaporin, AQP)属于膜整合蛋白家族, 能促进水分的跨膜运输.在已发现的35种拟南芥水通道蛋白中, 有13种PM整合蛋白(plasma membrane intrinsic protein, PIPs) (Jang et al., 2004).这些PIPs基因在应对各种胁迫(如干旱、冷、高盐和ABA)的过程中表达模式发生变化, 对维持植物的水分平衡起关键作用(Jang et al., 2004).蛋白质组学研究表明, 在燕麦叶片应对冷胁迫(Takahashi et al., 2013)及玉米根应对铁缺乏和过量(Hopff et al., 2013)时AQP丰度减少; 在拟南芥悬浮细胞应对ABA胁迫时AQP丰度增加(Li et al., 2012a) (图1A; 附表3), 进而调整胁迫下植物的水分与渗透平衡. ... ... ).这些PIPs基因在应对各种胁迫(如干旱、冷、高盐和ABA)的过程中表达模式发生变化, 对维持植物的水分平衡起关键作用(Jang et al., 2004).蛋白质组学研究表明, 在燕麦叶片应对冷胁迫(Takahashi et al., 2013)及玉米根应对铁缺乏和过量(Hopff et al., 2013)时AQP丰度减少; 在拟南芥悬浮细胞应对ABA胁迫时AQP丰度增加(Li et al., 2012a) (图1A; 附表3), 进而调整胁迫下植物的水分与渗透平衡. ...
4 2003
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ... ... ; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ... ... ; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ... ... ).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
1 2004
... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ...
1 1996
... 脂质转移蛋白(lipid transfer protein, LTP)可以促进脂质在植物细胞外基质间的转移, 参与胞外角质蜡质的形成与沉积, 以利于抵御病原体的侵害以及逆境(如水分和低温)胁迫应答(Kader, 1996; Cameron et al., 2006).有研究表明, 烟草叶片中NgLTP基因在脱水胁迫条件下表达量增加了6倍(Cameron et al., 2006).Nouri和Komatsu (2010)利用蛋白质组学相关技术对大豆进行了研究, 发现在渗透胁迫下大豆根与下胚轴中LTP丰度减少(图1A; 附表3), 这将降低角质层蜡质的沉积, 从而削弱植物的耐旱性. ...
1 2001
... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ...
1 2001
... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
4 2007
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ...
6 2003
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... 磷脂酶D (phospholipase D, PLD)可以水解磷脂生成PA (Xiong et al., 2002), PA也可作为第二信使参与胁迫信号转导, 调控下游基因表达等事件(Munnik, 2001).蛋白质组学研究表明, 拟南芥悬浮细胞应对冷胁迫时PLD受到影响(Li et al., 2012a); 拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及拟南芥叶片应对冷胁迫(Kawamura and Uemura, 2003)时, PLDδ丰度均增加(图1B; 附表4).之前, Li等(2004)的研究表明, 拟南芥PLDδ突变体对冷害更敏感, 而PLDδ过表达体的抗冻性增强, 表明PLD参与的信号转导途径在细胞低温应答中具重要作用. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
5 2009
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 细胞从膜受体接受逆境信号后, 利用磷酸化蛋白质对信号进行级联转换和放大, 蛋白质可逆磷酸化在植物细胞信号转导过程中具重要作用(Xiong and Zhu, 2001).蛋白质可逆磷酸化通过蛋白激酶和磷酸酶完成, 主要发生在丝氨酸、苏氨酸和酪氨酸等氨基酸位点.丝氨酸/苏氨酸蛋白激酶能使靶蛋白的丝氨酸和苏氨酸残基磷酸化.蛋白质组学研究表明, 在黑麦和燕麦叶片应对冷胁迫(Takahashi et al., 2013)以及拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a)时, 丝氨酸/苏氨酸蛋白激酶丰度发生不同水平的改变; 在大豆根与下胚轴应对水淹胁迫时, 蛋白激酶丰度增加(Komatsu et al., 2009).此外, 蛋白质组学研究还表明, 冷胁迫下的拟南芥悬浮细胞中组氨酸激酶(histidine kinase)丰度减少(Li et al., 2012a), 从而调控相应靶蛋白的磷酸化水平, 应答环境胁迫(图1B; 附表3). ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
1 2008
... 网格蛋白介导的内吞作用(clathrin-mediated endocytosis, CME)是网格蛋白作为受体介导内吞作用的主要方式, 是植物细胞内向膜泡运输(vesicle transport)的重要途径.除了基本内吞作用, 此过程也受到防御诱导子的调节(Leborgne-Castel et al., 2008).高渗胁迫会导致CME增强、胞吐作用减弱; 低渗胁迫的作用则相反(Zwiewka et al., 2015).Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根和下胚轴中网格蛋白的丰度降低(图1A; 附表3), 这将减弱网格蛋白被招募到PM形成网格蛋白包被小窝(clathrin coated pits, CCPs)的能力, 进而削弱内吞作用, 不利于大豆抵御渗透胁迫.在内吞作用过程中, 动力蛋白(dy- namin)作为GTPase参与调节CCPs从膜上脱离并形成网格蛋白包被小泡的过程(Hinshaw, 2000).拟南芥中, 动力蛋白参与抗病、线粒体分裂及内吞作用中网格蛋白包被小泡的细胞板形成和分裂(Hong et al., 2003; Tang et al., 2006).蛋白质组学研究表明, 冷胁迫下拟南芥幼苗中动力相关蛋白(dynamin-related protein)的丰度增加(Minami et al., 2009) (图1A; 附表4), 这将促进网格蛋白包被小泡的释放, 利于运输货物蛋白. ...
1 2002
... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
1 2000
... 硫是植物生长发育必需的大量营养元素之一, 植物根主要以硫酸盐(SO42-)的形式从土壤中吸收硫元素(Leustek and Saito, 1999).硫酸盐转运蛋白(sulphate transporter, SULTR)是一类参与SO42-主动运输的载体蛋白, 与硫元素的吸收、同化和长距离运输有关(Leustek and Saito, 1999; Leustek et al., 2000).在高水平硫条件下, Sultr1;2呈现高表达; 在硫缺乏条件下, 拟南芥Sultr1;1与Sultr1;2均上调表达, 但Sultr1;2对硫缺乏的响应能力比Sultr1;1低(Yoshimoto et al., 2002).Shibagaki等(2002)对Sultr1;2突变体进行了分析, 发现该突变体对硫酸盐的吸收活性明显降低, Sultr1;2转运体对拟南芥根部吸收硫酸盐起重要作用.Sultr1;2敲除突变体中, Sultr- 1;1 mRNA水平上调, 但是不能恢复硫酸盐的吸收能力(Maruyama-Nakashita et al., 2003), 表明Sultr1;1和Sultr1;2均参与硫酸盐的吸收, 但二者对硫酸盐的吸收能力不同.蛋白质组学研究表明, 冷胁迫下的黑麦叶片中SULTR丰度下降(Takahashi et al., 2013) (图1A; 附表3), 这可能引起硫素的吸收和运输受限, 影响植物的生长. ...
2 1999
... 硫是植物生长发育必需的大量营养元素之一, 植物根主要以硫酸盐(SO42-)的形式从土壤中吸收硫元素(Leustek and Saito, 1999).硫酸盐转运蛋白(sulphate transporter, SULTR)是一类参与SO42-主动运输的载体蛋白, 与硫元素的吸收、同化和长距离运输有关(Leustek and Saito, 1999; Leustek et al., 2000).在高水平硫条件下, Sultr1;2呈现高表达; 在硫缺乏条件下, 拟南芥Sultr1;1与Sultr1;2均上调表达, 但Sultr1;2对硫缺乏的响应能力比Sultr1;1低(Yoshimoto et al., 2002).Shibagaki等(2002)对Sultr1;2突变体进行了分析, 发现该突变体对硫酸盐的吸收活性明显降低, Sultr1;2转运体对拟南芥根部吸收硫酸盐起重要作用.Sultr1;2敲除突变体中, Sultr- 1;1 mRNA水平上调, 但是不能恢复硫酸盐的吸收能力(Maruyama-Nakashita et al., 2003), 表明Sultr1;1和Sultr1;2均参与硫酸盐的吸收, 但二者对硫酸盐的吸收能力不同.蛋白质组学研究表明, 冷胁迫下的黑麦叶片中SULTR丰度下降(Takahashi et al., 2013) (图1A; 附表3), 这可能引起硫素的吸收和运输受限, 影响植物的生长. ... ... 主动运输的载体蛋白, 与硫元素的吸收、同化和长距离运输有关(Leustek and Saito, 1999; Leustek et al., 2000).在高水平硫条件下, Sultr1;2呈现高表达; 在硫缺乏条件下, 拟南芥Sultr1;1与Sultr1;2均上调表达, 但Sultr1;2对硫缺乏的响应能力比Sultr1;1低(Yoshimoto et al., 2002).Shibagaki等(2002)对Sultr1;2突变体进行了分析, 发现该突变体对硫酸盐的吸收活性明显降低, Sultr1;2转运体对拟南芥根部吸收硫酸盐起重要作用.Sultr1;2敲除突变体中, Sultr- 1;1 mRNA水平上调, 但是不能恢复硫酸盐的吸收能力(Maruyama-Nakashita et al., 2003), 表明Sultr1;1和Sultr1;2均参与硫酸盐的吸收, 但二者对硫酸盐的吸收能力不同.蛋白质组学研究表明, 冷胁迫下的黑麦叶片中SULTR丰度下降(Takahashi et al., 2013) (图1A; 附表3), 这可能引起硫素的吸收和运输受限, 影响植物的生长. ...
44 2012
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ... ... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... ); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... ; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... 水通道蛋白(aquaporin, AQP)属于膜整合蛋白家族, 能促进水分的跨膜运输.在已发现的35种拟南芥水通道蛋白中, 有13种PM整合蛋白(plasma membrane intrinsic protein, PIPs) (Jang et al., 2004).这些PIPs基因在应对各种胁迫(如干旱、冷、高盐和ABA)的过程中表达模式发生变化, 对维持植物的水分平衡起关键作用(Jang et al., 2004).蛋白质组学研究表明, 在燕麦叶片应对冷胁迫(Takahashi et al., 2013)及玉米根应对铁缺乏和过量(Hopff et al., 2013)时AQP丰度减少; 在拟南芥悬浮细胞应对ABA胁迫时AQP丰度增加(Li et al., 2012a) (图1A; 附表3), 进而调整胁迫下植物的水分与渗透平衡. ... ... 膜联蛋白(annexin, ANX)是一种钙依赖的膜结合蛋白, 在膜运输、离子通道活性调节和磷脂代谢中均发挥作用(Gerke and Moss, 2002).蛋白质组学研究表明, 在冷及ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)和氮胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度减少; 在冷胁迫的黑麦叶片(Takahashi et al., 2013)和NaCl胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度增加(图1A; 附表3).植物ANX能在PM上形成钙离子通道, ANX丰度的变化可能改变钙通道活性, 引起钙离子跨膜运输变化, 进而调节钙离子的稳态和信号转导过程(White et al., 2002). ... ... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... )时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... ); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 此外, PATL (Patellin)是一种载体蛋白, 含有磷脂酰肌醇转移蛋白(phosphatidylinositol transfer protein, Sec14)样脂质结合结构域(Sec14p-like lipid-binding domain).Sec14可转移磷脂质并调控脂质信号分子产物(Mousley et al., 2007).PATL参与膜运输、细胞板扩大和成熟, 在细胞板生物合成过程中可结合疏水分子(如磷酸肌醇)并促进其在不同细胞位点间转移(Zhang et al., 2008).在拟南芥幼苗(Minami et al., 2009)和燕麦叶片(Takahashi et al., 2013)应对冷胁迫及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, PATL丰度降低; 而在拟南芥悬浮细胞应对ABA胁迫时, PATL丰度增加(Li et al., 2012a) (图1A; 附表4), 这可能对胁迫条件下膜运输及细胞分裂过程有一定的影响. ... ... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... )的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ... ... 油菜素内酯(brassinosteroids, BRs)是一类促生长的类固醇激素, 参与调节植物的生长发育过程(Clouse, 2011).可逆磷酸化反应是BR信号转导的主要调控方式.油菜素受体BRI1 (brassinosteroid insensitive 1)在细胞表面感知并结合BR, 发生自磷酸化(Wang et al., 2005).BRI1类受体激酶(brassinos- teroid insensitive 1-associated receptor kinase 1, BAK1)作为BRI1的共受体, 二者相互作用对BR信号转导尤为重要, BAK1与BRI1彼此磷酸化激活BRI1, 为下游调控因子提供作用位点, 将信号向下传递(Nam and Li, 2002; Clouse, 2011).过表达BAK1导致拟南芥器官伸长, 而BAK1缺失会引起植株半矮化并降低对BR的敏感性(Li et al., 2002).蛋白质组学研究表明, 冷胁迫下拟南芥悬浮细胞中, BRI1丰度上升或下降, 而ABA胁迫下的拟南芥悬浮细胞中, BRI1丰度增加(Li et al., 2012a); 铁缺乏和过量条件下, 玉米根中BAK1丰度减少(Hopff et al., 2013) (图1B; 附表3), 这些变化使BRI1、BAK1与BR的互作受到影响, 进而影响BR的信号传递.此外, Li等(2012b)研究发现, 拟南芥BSK5转录物在非生物胁迫(包括盐、干旱、植物激素BR和ABA)下上调表达, 且拟南芥bsk5功能缺失突变体对盐和ABA的敏感性增强.另外, 蛋白质组学研究还表明, 在冷和ABA胁迫的拟南芥悬浮细胞中, BR信号的正调节蛋白——油菜素内酯信号激酶(BR-signaling kinase, BSK)的丰度增加(Li et al., 2012a) (图1B; 附表4), 这不仅有助于BSKs被BRI1磷酸化激活, 而且可以促进BR的信号传递(Clouse, 2011)及BR靶基因的表达. ... ... 功能缺失突变体对盐和ABA的敏感性增强.另外, 蛋白质组学研究还表明, 在冷和ABA胁迫的拟南芥悬浮细胞中, BR信号的正调节蛋白——油菜素内酯信号激酶(BR-signaling kinase, BSK)的丰度增加(Li et al., 2012a) (图1B; 附表4), 这不仅有助于BSKs被BRI1磷酸化激活, 而且可以促进BR的信号传递(Clouse, 2011)及BR靶基因的表达. ... ... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ... ... ).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ... ... 磷酸肌醇信号途径在植物生长、发育及应对逆境过程中具有重要作用.细胞膜受体接收胞外信号, 激活磷酸肌醇特异的磷脂酶C (phosphatidylinositol-specific phospholipase C, PI-PLC), 将磷脂酰肌醇-4,5-二磷酸水解生成1,4,5-肌醇三磷酸(IP3)和二酰甘油(DAG) (Testerink and Munnik, 2005).PLC通过调节2个重要的信使(IP3和DAG)参与植物各种信号途径.IP3刺激细胞外存储的Ca2+进入细胞质基质, 使胞内Ca2+浓度升高, Ca2+进一步调节信号转导途径(Hunt et al., 2004).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ... ... 磷脂酶D (phospholipase D, PLD)可以水解磷脂生成PA (Xiong et al., 2002), PA也可作为第二信使参与胁迫信号转导, 调控下游基因表达等事件(Munnik, 2001).蛋白质组学研究表明, 拟南芥悬浮细胞应对冷胁迫时PLD受到影响(Li et al., 2012a); 拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及拟南芥叶片应对冷胁迫(Kawamura and Uemura, 2003)时, PLDδ丰度均增加(图1B; 附表4).之前, Li等(2004)的研究表明, 拟南芥PLDδ突变体对冷害更敏感, 而PLDδ过表达体的抗冻性增强, 表明PLD参与的信号转导途径在细胞低温应答中具重要作用. ... ... ); 拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及拟南芥叶片应对冷胁迫(Kawamura and Uemura, 2003)时, PLDδ丰度均增加(图1B; 附表4).之前, Li等(2004)的研究表明, 拟南芥PLDδ突变体对冷害更敏感, 而PLDδ过表达体的抗冻性增强, 表明PLD参与的信号转导途径在细胞低温应答中具重要作用. ... ... 促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)是丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase)的一个家族(Mishra et al., 2006).植物中的MAPK级联途径由MAPK激酶激酶(mitogen activated protein kinase kinase kinase, MAP3K)、MAPK激酶(mitogen activated protein kinase kinase, MAPKK)和MAPK/MPK三种蛋白激酶组成, MAP3K磷酸化激活MAPKK, MAPKK磷酸化激活MAPK, 将外来信号级联放大并且传递下去(Mou- stafa et al., 2014).MAPK级联途径参与植物的生长发育、激素信号转导及应答生物和非生物胁迫(如干旱、盐和温度) (Moustafa et al., 2014).过表达MAP3Kδ4的拟南芥植株生长旺盛(Sasayama et al., 2011).在高盐、渗透、干旱、冷和ABA胁迫下MAP3Kδ4表达上调, 且ABA处理使MAP3Kδ4活性增强.过表达MAP3Kδ4的拟南芥种子萌发受ABA抑制减弱, 表明MAP3Kδ4在ABA信号通路中发挥重要作用(Shitamichi et al., 2013).蛋白质组学研究表明, 在冷和ABA胁迫下, 拟南芥悬浮细胞中MAP3K丰度增加(Li et al., 2012a) (图1B; 附表4), 这将有利于促使MAPK级联信号传递, 诱导相关基因的表达, 增强植物抗胁迫能力. ... ... 细胞从膜受体接受逆境信号后, 利用磷酸化蛋白质对信号进行级联转换和放大, 蛋白质可逆磷酸化在植物细胞信号转导过程中具重要作用(Xiong and Zhu, 2001).蛋白质可逆磷酸化通过蛋白激酶和磷酸酶完成, 主要发生在丝氨酸、苏氨酸和酪氨酸等氨基酸位点.丝氨酸/苏氨酸蛋白激酶能使靶蛋白的丝氨酸和苏氨酸残基磷酸化.蛋白质组学研究表明, 在黑麦和燕麦叶片应对冷胁迫(Takahashi et al., 2013)以及拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a)时, 丝氨酸/苏氨酸蛋白激酶丰度发生不同水平的改变; 在大豆根与下胚轴应对水淹胁迫时, 蛋白激酶丰度增加(Komatsu et al., 2009).此外, 蛋白质组学研究还表明, 冷胁迫下的拟南芥悬浮细胞中组氨酸激酶(histidine kinase)丰度减少(Li et al., 2012a), 从而调控相应靶蛋白的磷酸化水平, 应答环境胁迫(图1B; 附表3). ... ... ).此外, 蛋白质组学研究还表明, 冷胁迫下的拟南芥悬浮细胞中组氨酸激酶(histidine kinase)丰度减少(Li et al., 2012a), 从而调控相应靶蛋白的磷酸化水平, 应答环境胁迫(图1B; 附表3). ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... )、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... )、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... )、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... ), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... ), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... ; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... )和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ... ... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ...
2 2006
... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... ).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
2015
1 2002
... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ...
4 2007
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... ).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ...
1 2000
... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
1 2001
... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
1 2003
... 硫是植物生长发育必需的大量营养元素之一, 植物根主要以硫酸盐(SO42-)的形式从土壤中吸收硫元素(Leustek and Saito, 1999).硫酸盐转运蛋白(sulphate transporter, SULTR)是一类参与SO42-主动运输的载体蛋白, 与硫元素的吸收、同化和长距离运输有关(Leustek and Saito, 1999; Leustek et al., 2000).在高水平硫条件下, Sultr1;2呈现高表达; 在硫缺乏条件下, 拟南芥Sultr1;1与Sultr1;2均上调表达, 但Sultr1;2对硫缺乏的响应能力比Sultr1;1低(Yoshimoto et al., 2002).Shibagaki等(2002)对Sultr1;2突变体进行了分析, 发现该突变体对硫酸盐的吸收活性明显降低, Sultr1;2转运体对拟南芥根部吸收硫酸盐起重要作用.Sultr1;2敲除突变体中, Sultr- 1;1 mRNA水平上调, 但是不能恢复硫酸盐的吸收能力(Maruyama-Nakashita et al., 2003), 表明Sultr1;1和Sultr1;2均参与硫酸盐的吸收, 但二者对硫酸盐的吸收能力不同.蛋白质组学研究表明, 冷胁迫下的黑麦叶片中SULTR丰度下降(Takahashi et al., 2013) (图1A; 附表3), 这可能引起硫素的吸收和运输受限, 影响植物的生长. ...
6 2011
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... )中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... ; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
1 1995
... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ...
1 2003
... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
1 2004
... 磷酸肌醇信号途径在植物生长、发育及应对逆境过程中具有重要作用.细胞膜受体接收胞外信号, 激活磷酸肌醇特异的磷脂酶C (phosphatidylinositol-specific phospholipase C, PI-PLC), 将磷脂酰肌醇-4,5-二磷酸水解生成1,4,5-肌醇三磷酸(IP3)和二酰甘油(DAG) (Testerink and Munnik, 2005).PLC通过调节2个重要的信使(IP3和DAG)参与植物各种信号途径.IP3刺激细胞外存储的Ca2+进入细胞质基质, 使胞内Ca2+浓度升高, Ca2+进一步调节信号转导途径(Hunt et al., 2004).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ...
8 2009
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 网格蛋白介导的内吞作用(clathrin-mediated endocytosis, CME)是网格蛋白作为受体介导内吞作用的主要方式, 是植物细胞内向膜泡运输(vesicle transport)的重要途径.除了基本内吞作用, 此过程也受到防御诱导子的调节(Leborgne-Castel et al., 2008).高渗胁迫会导致CME增强、胞吐作用减弱; 低渗胁迫的作用则相反(Zwiewka et al., 2015).Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根和下胚轴中网格蛋白的丰度降低(图1A; 附表3), 这将减弱网格蛋白被招募到PM形成网格蛋白包被小窝(clathrin coated pits, CCPs)的能力, 进而削弱内吞作用, 不利于大豆抵御渗透胁迫.在内吞作用过程中, 动力蛋白(dy- namin)作为GTPase参与调节CCPs从膜上脱离并形成网格蛋白包被小泡的过程(Hinshaw, 2000).拟南芥中, 动力蛋白参与抗病、线粒体分裂及内吞作用中网格蛋白包被小泡的细胞板形成和分裂(Hong et al., 2003; Tang et al., 2006).蛋白质组学研究表明, 冷胁迫下拟南芥幼苗中动力相关蛋白(dynamin-related protein)的丰度增加(Minami et al., 2009) (图1A; 附表4), 这将促进网格蛋白包被小泡的释放, 利于运输货物蛋白. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 此外, PATL (Patellin)是一种载体蛋白, 含有磷脂酰肌醇转移蛋白(phosphatidylinositol transfer protein, Sec14)样脂质结合结构域(Sec14p-like lipid-binding domain).Sec14可转移磷脂质并调控脂质信号分子产物(Mousley et al., 2007).PATL参与膜运输、细胞板扩大和成熟, 在细胞板生物合成过程中可结合疏水分子(如磷酸肌醇)并促进其在不同细胞位点间转移(Zhang et al., 2008).在拟南芥幼苗(Minami et al., 2009)和燕麦叶片(Takahashi et al., 2013)应对冷胁迫及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, PATL丰度降低; 而在拟南芥悬浮细胞应对ABA胁迫时, PATL丰度增加(Li et al., 2012a) (图1A; 附表4), 这可能对胁迫条件下膜运输及细胞分裂过程有一定的影响. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... )以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
1 2006
... 促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)是丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase)的一个家族(Mishra et al., 2006).植物中的MAPK级联途径由MAPK激酶激酶(mitogen activated protein kinase kinase kinase, MAP3K)、MAPK激酶(mitogen activated protein kinase kinase, MAPKK)和MAPK/MPK三种蛋白激酶组成, MAP3K磷酸化激活MAPKK, MAPKK磷酸化激活MAPK, 将外来信号级联放大并且传递下去(Mou- stafa et al., 2014).MAPK级联途径参与植物的生长发育、激素信号转导及应答生物和非生物胁迫(如干旱、盐和温度) (Moustafa et al., 2014).过表达MAP3Kδ4的拟南芥植株生长旺盛(Sasayama et al., 2011).在高盐、渗透、干旱、冷和ABA胁迫下MAP3Kδ4表达上调, 且ABA处理使MAP3Kδ4活性增强.过表达MAP3Kδ4的拟南芥种子萌发受ABA抑制减弱, 表明MAP3Kδ4在ABA信号通路中发挥重要作用(Shitamichi et al., 2013).蛋白质组学研究表明, 在冷和ABA胁迫下, 拟南芥悬浮细胞中MAP3K丰度增加(Li et al., 2012a) (图1B; 附表4), 这将有利于促使MAPK级联信号传递, 诱导相关基因的表达, 增强植物抗胁迫能力. ...
1 1997
... 磷是植物生长发育的关键营养元素之一.植物通过磷转运蛋白(phosphate transporter, PT)吸收以及转运磷素(Raghothama, 2000).在烟草(Nicotiana glauca)细胞中过表达拟南芥磷转运蛋白基因PHT1增加了磷的吸收效率, 促进了磷缺乏条件下的细胞生长(Mitsukawa et al., 1997).在水稻中过表达OsPT1可增加植物对磷素的积累(Seo et al., 2008).Li等(2012a)的研究表明, 冷与ABA胁迫下的拟南芥悬浮细胞中PT丰度增加, 这将有利于磷的吸收和转运, 改善植物磷素的营养状态(图1A; 附表3). ...
1 2010
... 质膜(plasma membrane, PM)是细胞原生质体与外界进行物质交换和信息交流的基础, 对完成细胞间的各种代谢活动及以细胞分裂与分化为基础的生长发育过程均具重要作用.对植物细胞而言, 物质跨PM运输是细胞壁生长与代谢的基础, PM与细胞壁一起成为细胞应答外界环境的屏障(Mongrand et al., 2010).质膜与细胞内膜系统和各种细胞器膜构成了细胞内外信号分子、离子、激素以及其它代谢物等进行交换的媒介, 对细胞生长以及环境应答至关重要(Yadeta et al., 2013).在PM磷脂双分子层上, 以各种形式分布的蛋白质(整合蛋白、外周蛋白和膜锚定蛋白等)是PM行使功能的基础.它们在构成PM基本结构、感知与转导信号、细胞内外物质运输以及防御胁迫等方面都发挥重要作用.因此, 深入解析植物PM蛋白质的组成、分布与功能对认识细胞信号与代谢活动的分子机制具有重要意义. ...
1 1994
... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ...
1 2007
... 此外, PATL (Patellin)是一种载体蛋白, 含有磷脂酰肌醇转移蛋白(phosphatidylinositol transfer protein, Sec14)样脂质结合结构域(Sec14p-like lipid-binding domain).Sec14可转移磷脂质并调控脂质信号分子产物(Mousley et al., 2007).PATL参与膜运输、细胞板扩大和成熟, 在细胞板生物合成过程中可结合疏水分子(如磷酸肌醇)并促进其在不同细胞位点间转移(Zhang et al., 2008).在拟南芥幼苗(Minami et al., 2009)和燕麦叶片(Takahashi et al., 2013)应对冷胁迫及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, PATL丰度降低; 而在拟南芥悬浮细胞应对ABA胁迫时, PATL丰度增加(Li et al., 2012a) (图1A; 附表4), 这可能对胁迫条件下膜运输及细胞分裂过程有一定的影响. ...
2 2014
... 促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)是丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase)的一个家族(Mishra et al., 2006).植物中的MAPK级联途径由MAPK激酶激酶(mitogen activated protein kinase kinase kinase, MAP3K)、MAPK激酶(mitogen activated protein kinase kinase, MAPKK)和MAPK/MPK三种蛋白激酶组成, MAP3K磷酸化激活MAPKK, MAPKK磷酸化激活MAPK, 将外来信号级联放大并且传递下去(Mou- stafa et al., 2014).MAPK级联途径参与植物的生长发育、激素信号转导及应答生物和非生物胁迫(如干旱、盐和温度) (Moustafa et al., 2014).过表达MAP3Kδ4的拟南芥植株生长旺盛(Sasayama et al., 2011).在高盐、渗透、干旱、冷和ABA胁迫下MAP3Kδ4表达上调, 且ABA处理使MAP3Kδ4活性增强.过表达MAP3Kδ4的拟南芥种子萌发受ABA抑制减弱, 表明MAP3Kδ4在ABA信号通路中发挥重要作用(Shitamichi et al., 2013).蛋白质组学研究表明, 在冷和ABA胁迫下, 拟南芥悬浮细胞中MAP3K丰度增加(Li et al., 2012a) (图1B; 附表4), 这将有利于促使MAPK级联信号传递, 诱导相关基因的表达, 增强植物抗胁迫能力. ... ... ).MAPK级联途径参与植物的生长发育、激素信号转导及应答生物和非生物胁迫(如干旱、盐和温度) (Moustafa et al., 2014).过表达MAP3Kδ4的拟南芥植株生长旺盛(Sasayama et al., 2011).在高盐、渗透、干旱、冷和ABA胁迫下MAP3Kδ4表达上调, 且ABA处理使MAP3Kδ4活性增强.过表达MAP3Kδ4的拟南芥种子萌发受ABA抑制减弱, 表明MAP3Kδ4在ABA信号通路中发挥重要作用(Shitamichi et al., 2013).蛋白质组学研究表明, 在冷和ABA胁迫下, 拟南芥悬浮细胞中MAP3K丰度增加(Li et al., 2012a) (图1B; 附表4), 这将有利于促使MAPK级联信号传递, 诱导相关基因的表达, 增强植物抗胁迫能力. ...
1 2001
... 磷脂酶D (phospholipase D, PLD)可以水解磷脂生成PA (Xiong et al., 2002), PA也可作为第二信使参与胁迫信号转导, 调控下游基因表达等事件(Munnik, 2001).蛋白质组学研究表明, 拟南芥悬浮细胞应对冷胁迫时PLD受到影响(Li et al., 2012a); 拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及拟南芥叶片应对冷胁迫(Kawamura and Uemura, 2003)时, PLDδ丰度均增加(图1B; 附表4).之前, Li等(2004)的研究表明, 拟南芥PLDδ突变体对冷害更敏感, 而PLDδ过表达体的抗冻性增强, 表明PLD参与的信号转导途径在细胞低温应答中具重要作用. ...
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 膜联蛋白(annexin, ANX)是一种钙依赖的膜结合蛋白, 在膜运输、离子通道活性调节和磷脂代谢中均发挥作用(Gerke and Moss, 2002).蛋白质组学研究表明, 在冷及ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)和氮胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度减少; 在冷胁迫的黑麦叶片(Takahashi et al., 2013)和NaCl胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度增加(图1A; 附表3).植物ANX能在PM上形成钙离子通道, ANX丰度的变化可能改变钙通道活性, 引起钙离子跨膜运输变化, 进而调节钙离子的稳态和信号转导过程(White et al., 2002). ... ... )和NaCl胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度增加(图1A; 附表3).植物ANX能在PM上形成钙离子通道, ANX丰度的变化可能改变钙通道活性, 引起钙离子跨膜运输变化, 进而调节钙离子的稳态和信号转导过程(White et al., 2002). ... ... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... ), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... )、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
2000
22 2010
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 此外, PATL (Patellin)是一种载体蛋白, 含有磷脂酰肌醇转移蛋白(phosphatidylinositol transfer protein, Sec14)样脂质结合结构域(Sec14p-like lipid-binding domain).Sec14可转移磷脂质并调控脂质信号分子产物(Mousley et al., 2007).PATL参与膜运输、细胞板扩大和成熟, 在细胞板生物合成过程中可结合疏水分子(如磷酸肌醇)并促进其在不同细胞位点间转移(Zhang et al., 2008).在拟南芥幼苗(Minami et al., 2009)和燕麦叶片(Takahashi et al., 2013)应对冷胁迫及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, PATL丰度降低; 而在拟南芥悬浮细胞应对ABA胁迫时, PATL丰度增加(Li et al., 2012a) (图1A; 附表4), 这可能对胁迫条件下膜运输及细胞分裂过程有一定的影响. ... ... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... ), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... ), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 向光素(phototropin, Phot)是与PM相关的光感受器, 也是P(H+)-ATPase的调节器, 具有丝氨酸/苏氨酸激酶的催化结构域, 在蓝光下会发生自磷酸化(No- uri and Komatsu, 2010).Ueno等(2005)研究发现, 向光素参与蓝光依赖的P(H+)-ATPase激活.Galen等(2007)的研究表明, 拟南芥向光素蛋白增强了近土表层侧根系的生长效率, 提升了植物的耐旱性.另外, 蛋白质组学研究表明, 在冷胁迫下的黑麦叶片中, Phot丰度上升或下降(Takahashi et al., 2013); 在冷胁迫下的燕麦叶片(Takahashi et al., 2013)以及渗透胁迫下的大豆根与下胚轴(Nouri and Komatsu, 2010)中, Phot丰度增加(图1B; 附表4).推测向光素蛋白在胁迫条件下可能参与调控根的形态改变(Nouri and Komatsu, 2010). ... ... )以及渗透胁迫下的大豆根与下胚轴(Nouri and Komatsu, 2010)中, Phot丰度增加(图1B; 附表4).推测向光素蛋白在胁迫条件下可能参与调控根的形态改变(Nouri and Komatsu, 2010). ... ... ; 附表4).推测向光素蛋白在胁迫条件下可能参与调控根的形态改变(Nouri and Komatsu, 2010). ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... )时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... ), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... ); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ... ... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
2000
1 2001
... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ...
1 2004
... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ...
2000
2004
2 2003
... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
1 2000
... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ...
1 2000
... 磷是植物生长发育的关键营养元素之一.植物通过磷转运蛋白(phosphate transporter, PT)吸收以及转运磷素(Raghothama, 2000).在烟草(Nicotiana glauca)细胞中过表达拟南芥磷转运蛋白基因PHT1增加了磷的吸收效率, 促进了磷缺乏条件下的细胞生长(Mitsukawa et al., 1997).在水稻中过表达OsPT1可增加植物对磷素的积累(Seo et al., 2008).Li等(2012a)的研究表明, 冷与ABA胁迫下的拟南芥悬浮细胞中PT丰度增加, 这将有利于磷的吸收和转运, 改善植物磷素的营养状态(图1A; 附表3). ...
1 1986
... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ...
2 1996
... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... 受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
1 2003
... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ...
1 2000
... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
2 2000
... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ...
1 2011
... 促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)是丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase)的一个家族(Mishra et al., 2006).植物中的MAPK级联途径由MAPK激酶激酶(mitogen activated protein kinase kinase kinase, MAP3K)、MAPK激酶(mitogen activated protein kinase kinase, MAPKK)和MAPK/MPK三种蛋白激酶组成, MAP3K磷酸化激活MAPKK, MAPKK磷酸化激活MAPK, 将外来信号级联放大并且传递下去(Mou- stafa et al., 2014).MAPK级联途径参与植物的生长发育、激素信号转导及应答生物和非生物胁迫(如干旱、盐和温度) (Moustafa et al., 2014).过表达MAP3Kδ4的拟南芥植株生长旺盛(Sasayama et al., 2011).在高盐、渗透、干旱、冷和ABA胁迫下MAP3Kδ4表达上调, 且ABA处理使MAP3Kδ4活性增强.过表达MAP3Kδ4的拟南芥种子萌发受ABA抑制减弱, 表明MAP3Kδ4在ABA信号通路中发挥重要作用(Shitamichi et al., 2013).蛋白质组学研究表明, 在冷和ABA胁迫下, 拟南芥悬浮细胞中MAP3K丰度增加(Li et al., 2012a) (图1B; 附表4), 这将有利于促使MAPK级联信号传递, 诱导相关基因的表达, 增强植物抗胁迫能力. ...
1 2007
... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
1 2003
... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ...
1 2008
... 磷是植物生长发育的关键营养元素之一.植物通过磷转运蛋白(phosphate transporter, PT)吸收以及转运磷素(Raghothama, 2000).在烟草(Nicotiana glauca)细胞中过表达拟南芥磷转运蛋白基因PHT1增加了磷的吸收效率, 促进了磷缺乏条件下的细胞生长(Mitsukawa et al., 1997).在水稻中过表达OsPT1可增加植物对磷素的积累(Seo et al., 2008).Li等(2012a)的研究表明, 冷与ABA胁迫下的拟南芥悬浮细胞中PT丰度增加, 这将有利于磷的吸收和转运, 改善植物磷素的营养状态(图1A; 附表3). ...
2002
1 1994
... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
1 2013
... 促分裂原活化蛋白激酶(mitogen-activated protein kinase, MAPK)是丝氨酸/苏氨酸蛋白激酶(serine/ threonine protein kinase)的一个家族(Mishra et al., 2006).植物中的MAPK级联途径由MAPK激酶激酶(mitogen activated protein kinase kinase kinase, MAP3K)、MAPK激酶(mitogen activated protein kinase kinase, MAPKK)和MAPK/MPK三种蛋白激酶组成, MAP3K磷酸化激活MAPKK, MAPKK磷酸化激活MAPK, 将外来信号级联放大并且传递下去(Mou- stafa et al., 2014).MAPK级联途径参与植物的生长发育、激素信号转导及应答生物和非生物胁迫(如干旱、盐和温度) (Moustafa et al., 2014).过表达MAP3Kδ4的拟南芥植株生长旺盛(Sasayama et al., 2011).在高盐、渗透、干旱、冷和ABA胁迫下MAP3Kδ4表达上调, 且ABA处理使MAP3Kδ4活性增强.过表达MAP3Kδ4的拟南芥种子萌发受ABA抑制减弱, 表明MAP3Kδ4在ABA信号通路中发挥重要作用(Shitamichi et al., 2013).蛋白质组学研究表明, 在冷和ABA胁迫下, 拟南芥悬浮细胞中MAP3K丰度增加(Li et al., 2012a) (图1B; 附表4), 这将有利于促使MAPK级联信号传递, 诱导相关基因的表达, 增强植物抗胁迫能力. ...
1 1993
... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ...
1 1996
... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
1 2002
... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
1 1991
... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ...
1 1998
... 膜融合是囊泡运输的重要过程之一, 植物膜融合主要由N-乙基马来酰亚胺敏感因子附着蛋白受体(soluble N-ethylmaleimide sensitive factor attachment protein receptors, SNAREs)蛋白介导.SNA- REs是一组具有卷曲螺旋域的膜相关蛋白, 可形成极为稳定的复合体(Sutton et al., 1998).SNAREs家族成员分为Qa-SNAREs、Qb-SNAREs、Qc-SNAREs和R-SNAREs四类.SNARE核心复合体包括含有R-SNARE结构域的小突触囊泡蛋白(vesicle-associ- ated membrane protein, VAMP)、含有Qa-SNARE结构域的突触融合蛋白(syntaxin protein)和含有Qb-及Qc-SNARE结构域的25 kDa突触相关蛋白(syn- aptosomal-associated protein of 25 kDa, SNAP-25)等成员.融合开始阶段, 突触小泡通过VAMP、syntaxin1和SNAP-25的特定配对在PM处连接, 形成反式SNARE复合体, 促进膜融合发生; 当两膜融合以后, 释放囊泡内物质, 反式SNARE复合体变为顺式SNARE复合体; 膜融合完成之后, 可溶的N-乙基马来酰亚胺敏感因子附着蛋白α (alpha-soluble N-ethylmaleimide sensitive factor attachment protein, α-SNAP)结合顺式SNARE复合体并招募N-乙基马来酰亚胺敏感因子(N-ethylmaleimide sensitive factor, NSF)与顺式SNARE复合体形成瞬时的20S复合体, NSF ATP水解作用分解SNARE复合体, 促使复合体解聚, 释放的SNARE蛋白循环进入下一个膜融合过程(Hong, 2005).PM蛋白质组学研究表明, 胁迫诱导一些参与膜融合过程的蛋白质丰度发生变化.例如, 玉米根应对铁缺乏和过量(Hopff et al., 2013), 及拟南芥悬浮细胞应对冷胁迫(Li et al., 2012a)时, SNARE核心复合体成员突触融合蛋白的丰度增加; 拟南芥悬浮细胞中小突触囊泡蛋白(synaptobrevin)在冷胁迫下的丰度也增加(Li et al., 2012a) (图1A; 附表3), 这将有利于囊泡内物质释放到PM或细胞外.Nouri和Komatsu (2010)的研究还发现, 大豆根与下胚轴中NSF和α-SNAP应对渗透胁迫, 及海蓬子根中α-SNAP应对盐胁迫时丰度减少(Nie et al., 2015); 而拟南芥悬浮细胞中NSF和α-SNAP应对冷胁迫(Li et al., 2012a), 海蓬子根中α-SNAP应对氮胁迫(Nie et al., 2015), 以及水稻根尖中α-SNAP应对盐胁迫(Cheng et al., 2009)时丰度增加(图1A; 附表4).α-SNAP是一些融合活动所必需的, SNAPs作为一种接头蛋白, 可与核心膜绑定蛋白SNAREs和ATPase NSF互作(Söllner et al., 1993), 使NSF锚定到膜上, 并可促进NSF两个ATPase位点中的1个ATP水解(Morgan et al., 1994).环境胁迫下, NSF和SNAP丰度的变化将影响SNARE蛋白介导的膜融合, 不利于胁迫下植物的囊泡运输. ...
34 2013
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... ) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ... ... V(H+)-ATPase跨越液泡膜形成并维持质子电化学梯度, 为离子与代谢物的主动运输提供驱动力(Gaxiola et al., 2007).尽管V(H+)-ATPase定位于液泡膜, 其在PM蛋白质组分中也经常被鉴定到.蛋白质组学研究表明, 在拟南芥幼苗、叶片和悬浮细胞(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a), 燕麦、黑麦叶片(Takahashi et al., 2013)和水稻根(Hashimoto et al., 2009)应对冷胁迫, 玉米根应对Fe缺乏(Hopff et al., 2013), 以及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, V(H+)-ATPase的丰度均降低.然而, 在白叶枯病菌处理的水稻悬浮细胞(Chen et al., 2007b)和NaCl胁迫下的水稻根尖(Cheng et al., 2009)中, V(H+)- ATPase的丰度增加(图1A; 附表3).此外, 在缺Fe和壳聚糖处理的豌豆根(Meisrimler et al., 2011)及盐和氮胁迫的海蓬子根(Nie et al., 2015)中, V(H+)- ATPase丰度不同程度地升高或降低(图1A; 附表3).另有报道, V(H+)-ATPase A亚基的编码基因在大麦(Hordeum vulgare)耐盐品种的根(Fukuda et al., 2004)和拟南芥幼苗(Magnotta and Gogarten, 2002)中, 受盐胁迫诱导上调表达.在水稻中表达盐生植物互花米草(Spartina alterniflora) V(H+)-ATPase c1基因(SaVHAc1), 可有效驱动某些阳离子的运输和ABA信号通路相关基因的表达(Baisakh et al., 2012).然而, 蛋白质组学研究也显示, 拟南芥幼苗与叶片、黑麦与燕麦叶片, 以及水稻根中的V(H+)-ATPase在应对冷胁迫时丰度降低(图1A; 附表3), 表明在应答盐胁迫时, 植物通过调节V(H+)-ATPase的丰度变化模式来实现离子区室化, 从而维持胞内离子的平衡, 而低温会导致膜蛋白丰度降低, 细胞代谢减缓. ... ... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ... ... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... )及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... )及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... ).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... )时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ... ... 硫是植物生长发育必需的大量营养元素之一, 植物根主要以硫酸盐(SO42-)的形式从土壤中吸收硫元素(Leustek and Saito, 1999).硫酸盐转运蛋白(sulphate transporter, SULTR)是一类参与SO42-主动运输的载体蛋白, 与硫元素的吸收、同化和长距离运输有关(Leustek and Saito, 1999; Leustek et al., 2000).在高水平硫条件下, Sultr1;2呈现高表达; 在硫缺乏条件下, 拟南芥Sultr1;1与Sultr1;2均上调表达, 但Sultr1;2对硫缺乏的响应能力比Sultr1;1低(Yoshimoto et al., 2002).Shibagaki等(2002)对Sultr1;2突变体进行了分析, 发现该突变体对硫酸盐的吸收活性明显降低, Sultr1;2转运体对拟南芥根部吸收硫酸盐起重要作用.Sultr1;2敲除突变体中, Sultr- 1;1 mRNA水平上调, 但是不能恢复硫酸盐的吸收能力(Maruyama-Nakashita et al., 2003), 表明Sultr1;1和Sultr1;2均参与硫酸盐的吸收, 但二者对硫酸盐的吸收能力不同.蛋白质组学研究表明, 冷胁迫下的黑麦叶片中SULTR丰度下降(Takahashi et al., 2013) (图1A; 附表3), 这可能引起硫素的吸收和运输受限, 影响植物的生长. ... ... 脂质运载蛋白(lipocalin)参与类固醇和脂质等疏水小分子的运输, 从而影响PM的流动性(Charron et al., 2002).温度诱导的脂质运载蛋白(temperature- induced lipocalin, TIL)参与膜的生物发生与修复, 维持PM的稳定性和渗透平衡, 在温度胁迫应答过程中发挥作用(Charron et al., 2002, 2005).蛋白质组学研究表明, 玉米根应对铁缺乏和过量(Hopff et al., 2013)及大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, TIL的丰度降低; 而冷胁迫下水稻根(Hashimoto et al., 2009)、拟南芥悬浮细胞(Li et al., 2012a)和黑麦叶片(Takahashi et al., 2013)中TIL丰度增加(图1A; 附表4).此前, 人们也发现拟南芥中Attil基因和小麦中Tatil基因分别在4°C冷胁迫和45°C热休克处理时上调表达(Charron et al., 2002).与之相似, 冷胁迫下拟南芥叶片中外膜脂蛋白样蛋白(outer membrane lipoprotein-like)的丰度增加(Kaw- amura and Uemura, 2003) (附表4).低温胁迫下, 脂质运载蛋白和脂质运载蛋白样蛋白的丰度上调对于维持膜稳定性和减少膜损伤十分必要. ... ... 水通道蛋白(aquaporin, AQP)属于膜整合蛋白家族, 能促进水分的跨膜运输.在已发现的35种拟南芥水通道蛋白中, 有13种PM整合蛋白(plasma membrane intrinsic protein, PIPs) (Jang et al., 2004).这些PIPs基因在应对各种胁迫(如干旱、冷、高盐和ABA)的过程中表达模式发生变化, 对维持植物的水分平衡起关键作用(Jang et al., 2004).蛋白质组学研究表明, 在燕麦叶片应对冷胁迫(Takahashi et al., 2013)及玉米根应对铁缺乏和过量(Hopff et al., 2013)时AQP丰度减少; 在拟南芥悬浮细胞应对ABA胁迫时AQP丰度增加(Li et al., 2012a) (图1A; 附表3), 进而调整胁迫下植物的水分与渗透平衡. ... ... 膜联蛋白(annexin, ANX)是一种钙依赖的膜结合蛋白, 在膜运输、离子通道活性调节和磷脂代谢中均发挥作用(Gerke and Moss, 2002).蛋白质组学研究表明, 在冷及ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)和氮胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度减少; 在冷胁迫的黑麦叶片(Takahashi et al., 2013)和NaCl胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度增加(图1A; 附表3).植物ANX能在PM上形成钙离子通道, ANX丰度的变化可能改变钙通道活性, 引起钙离子跨膜运输变化, 进而调节钙离子的稳态和信号转导过程(White et al., 2002). ... ... Remorin是植物特有的PM脂质筏相关蛋白, 在细胞骨架与膜骨架形成中发挥作用(Bariola et al., 2004).蛋白质组学研究表明, 大豆根与下胚轴应对水淹胁迫(Komatsu et al., 2009), 拟南芥幼苗(Minami et al., 2009)和悬浮细胞(Li et al., 2012a)应对冷胁迫, 水稻根尖(Cheng et al., 2009)、水稻根(Malakshah et al., 2007)和海蓬子根(Nie et al., 2015)应对盐胁迫时, Remorin的丰度均增加; 而铁缺乏胁迫的玉米根 (Hopff et al., 2013)、渗透胁迫的大豆根及下胚轴(Nouri and Komatsu, 2010)、冷胁迫的黑麦叶片(Takahashi et al., 2013)中, Remorin的丰度减少(图1A; 附表4).之前的研究也发现, 桑树(Morus indica)中Remorin基因(MiREM)在脱水和盐胁迫下上调表达; 拟南芥中过表达group1 remorin可增强植物在萌发和幼苗期对脱水和盐分的耐受性(Checker and Khurana, 2013).故Remorin可能参与胁迫信号途径的调控, 对感应环境刺激及稳定胁迫造成的PM损伤具重要作用(Malakshah et al., 2007; Cheng et al., 2009).此外, 转铁蛋白(transferrin, Tf)能介导铁的吸收(Schwarz et al., 2003), 在长期NaCl (3 mol·L-1)胁迫下的杜氏盐藻细胞中, Tf丰度增加(Katz et al., 2007) (图1A; 附表4), 这将有助于高盐胁迫下有限的离子利用, 增强离子的吸收(Fisher et al., 1997). ... ... 此外, PATL (Patellin)是一种载体蛋白, 含有磷脂酰肌醇转移蛋白(phosphatidylinositol transfer protein, Sec14)样脂质结合结构域(Sec14p-like lipid-binding domain).Sec14可转移磷脂质并调控脂质信号分子产物(Mousley et al., 2007).PATL参与膜运输、细胞板扩大和成熟, 在细胞板生物合成过程中可结合疏水分子(如磷酸肌醇)并促进其在不同细胞位点间转移(Zhang et al., 2008).在拟南芥幼苗(Minami et al., 2009)和燕麦叶片(Takahashi et al., 2013)应对冷胁迫及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, PATL丰度降低; 而在拟南芥悬浮细胞应对ABA胁迫时, PATL丰度增加(Li et al., 2012a) (图1A; 附表4), 这可能对胁迫条件下膜运输及细胞分裂过程有一定的影响. ... ... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... ); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ... ... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ... ... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ... ... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ... ... 磷酸肌醇信号途径在植物生长、发育及应对逆境过程中具有重要作用.细胞膜受体接收胞外信号, 激活磷酸肌醇特异的磷脂酶C (phosphatidylinositol-specific phospholipase C, PI-PLC), 将磷脂酰肌醇-4,5-二磷酸水解生成1,4,5-肌醇三磷酸(IP3)和二酰甘油(DAG) (Testerink and Munnik, 2005).PLC通过调节2个重要的信使(IP3和DAG)参与植物各种信号途径.IP3刺激细胞外存储的Ca2+进入细胞质基质, 使胞内Ca2+浓度升高, Ca2+进一步调节信号转导途径(Hunt et al., 2004).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ... ... 向光素(phototropin, Phot)是与PM相关的光感受器, 也是P(H+)-ATPase的调节器, 具有丝氨酸/苏氨酸激酶的催化结构域, 在蓝光下会发生自磷酸化(No- uri and Komatsu, 2010).Ueno等(2005)研究发现, 向光素参与蓝光依赖的P(H+)-ATPase激活.Galen等(2007)的研究表明, 拟南芥向光素蛋白增强了近土表层侧根系的生长效率, 提升了植物的耐旱性.另外, 蛋白质组学研究表明, 在冷胁迫下的黑麦叶片中, Phot丰度上升或下降(Takahashi et al., 2013); 在冷胁迫下的燕麦叶片(Takahashi et al., 2013)以及渗透胁迫下的大豆根与下胚轴(Nouri and Komatsu, 2010)中, Phot丰度增加(图1B; 附表4).推测向光素蛋白在胁迫条件下可能参与调控根的形态改变(Nouri and Komatsu, 2010). ... ... ); 在冷胁迫下的燕麦叶片(Takahashi et al., 2013)以及渗透胁迫下的大豆根与下胚轴(Nouri and Komatsu, 2010)中, Phot丰度增加(图1B; 附表4).推测向光素蛋白在胁迫条件下可能参与调控根的形态改变(Nouri and Komatsu, 2010). ... ... 细胞从膜受体接受逆境信号后, 利用磷酸化蛋白质对信号进行级联转换和放大, 蛋白质可逆磷酸化在植物细胞信号转导过程中具重要作用(Xiong and Zhu, 2001).蛋白质可逆磷酸化通过蛋白激酶和磷酸酶完成, 主要发生在丝氨酸、苏氨酸和酪氨酸等氨基酸位点.丝氨酸/苏氨酸蛋白激酶能使靶蛋白的丝氨酸和苏氨酸残基磷酸化.蛋白质组学研究表明, 在黑麦和燕麦叶片应对冷胁迫(Takahashi et al., 2013)以及拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a)时, 丝氨酸/苏氨酸蛋白激酶丰度发生不同水平的改变; 在大豆根与下胚轴应对水淹胁迫时, 蛋白激酶丰度增加(Komatsu et al., 2009).此外, 蛋白质组学研究还表明, 冷胁迫下的拟南芥悬浮细胞中组氨酸激酶(histidine kinase)丰度减少(Li et al., 2012a), 从而调控相应靶蛋白的磷酸化水平, 应答环境胁迫(图1B; 附表3). ... ... P(H+)-ATPase活性受到可逆磷酸化的调节, 被磷酸化的P(H+)-ATPase活性受到抑制.蛋白质组学研究表明, 在应对渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和Fe缺乏及过量胁迫(Hopff et al., 2013)时, 大豆根与下胚轴、燕麦叶片、拟南芥悬浮细胞及玉米根中蛋白激酶(protein kinase, PK)的丰度受到影响; 在渗透(Nouri and Komatsu, 2010)、冷(Li et al., 2012a; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... ; Takahashi et al., 2013)、ABA (Li et al., 2012a)和白叶枯病(Chen et al., 2007b)胁迫下, 大豆根与下胚轴, 燕麦和黑麦叶片, 拟南芥悬浮细胞, Xa21转基因水稻悬浮细胞中的蛋白磷酸酶(protein phosphatase, PP)丰度发生变化.其中, 渗透胁迫下大豆根与下胚轴中的PK丰度减少, PP丰度增加(No- uri and Komatsu, 2010); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... ); 冷胁迫下燕麦叶片中的PK和PP, 黑麦叶片中的PP (Takahashi et al., 2013)以及白叶枯病Xoo PXO99A和DY89031感染的Xa21转基因水稻悬浮细胞中的PP丰度均增加(Chen et al., 2007b) (图1B; 附表3).可见, P(H+)-ATPase可逆磷酸化在应答渗透、冷和病菌胁迫时具重要作用.同时, P(H+)-ATPase活性也受到14-3-3蛋白质的调节.14-3-3蛋白通过与P(H+)-ATPase C端互作, 正向调节PM H+-ATPase活性, 从而控制跨PM的电化学梯度, 调控离子运输和细胞质pH值(Palmgren, 2001).与之相似, PM离子通道活性也受到14-3-3蛋白的调节, 从而维持离子的平衡(Bunney et al., 2002).此外, 植物14-3-3蛋白不仅可通过调节不同靶蛋白的活性调控多种代谢过程, 如离子运输、信号转导、转录激活和防御等(Roberts, 2003), 而且可作为转录因子复合体组件参与ABA诱导的基因表达调控(Fulgosi et al., 2002).蛋白质组学研究表明, 14-3-3蛋白在多种胁迫应答中丰度发生变化, 如氮胁迫下海蓬子根中的14-3-3蛋白(Nie et al., 2015), 盐胁迫下水稻根尖(Cheng et al., 2009), 渗透胁迫下大豆根与下胚轴中的14-3-3类蛋白A和D (Nouri and Komatsu, 2010), 以及冷胁迫下拟南芥悬浮细胞中的14-3-3类蛋白(GF14 PHI) (Li et al., 2012a)丰度均减少; 而水淹胁迫下大豆根与下胚轴中的14-3-3蛋白(Komatsu et al., 2009), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... ), 冷胁迫下黑麦叶片(Takahashi et al., 2013)及盐胁迫下水稻根中的GF14a和GF14b (Malakshah et al., 2007), ABA胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的GF14丰度增加(图1B; 附表4).上述研究表明, 14-3-3蛋白家族不同成员参与多种胁迫(如盐、渗透、水淹和冷等)应答的信号与代谢调节过程. ... ... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... ).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ... ... 逆境引起的植物体内活性氧(reactive oxygen species, ROS)自由基过量会损伤一些大分子(如DNA、蛋白质和叶绿素)以及细胞膜的结构(Gómez et al., 2004).植物通过启动多种抗氧化途径来清除过量的ROS (Roxas et al., 2000; 林植芳和刘楠, 2012).在大豆根与下胚轴、海蓬子根、豌豆根、水稻根尖和悬浮细胞、拟南芥叶片和悬浮细胞以及黑麦叶片的PM组分中, 分别鉴定到了响应渗透、水淹、冷、NaCl、ABA、Fe缺乏与过量、壳聚糖、白叶枯病菌侵染和氮素等胁迫时丰度发生变化的超氧化物歧化酶(super- oxide dismutase, SOD)、过氧化氢酶(catalase)、抗坏血酸过氧化物酶(ascorbate peroxidase)、单脱氢抗坏血酸还原酶(monodehydroascorbate reductase)、谷胱甘肽硫转移酶(glutathione S-transferase)和过氧化物酶(peroxidase, POD)等抗氧化酶(Kawamura and Uemura, 2003; Chen et al., 2007b; Cheng et al., 2009; Komatsu et al., 2009; Nouri and Komatsu, 2010; Meisrimler et al., 2011; Li et al., 2012a; Takahashi et al., 2013; Nie et al., 2015) (图1D; 附表3, 附表4).然而, 其中只有Cu/Zn SOD和POD曾被报道与PM相关联(Karpinska et al., 2001; Mika and Lüthje, 2003).Karpinska等(2001)利用免疫胶体金电镜技术, 发现欧洲赤松(Pinus sylvestris)子叶筛胞PM中存在Cu/Zn SOD.Mika和Lüthje (2003)通过阴离子交换层析, 在玉米根部PM中发现了PM结合的POD (pmPOX1和pmPOX2).其它参与ROS清除的酶类存在于细胞质与细胞器中.虽然有很多报道表明, 它们在逆境应答过程中对清除过量的ROS具重要作用, 但是其是否真正与PM相关联还有待进一步研究. ...
1 2006
... 网格蛋白介导的内吞作用(clathrin-mediated endocytosis, CME)是网格蛋白作为受体介导内吞作用的主要方式, 是植物细胞内向膜泡运输(vesicle transport)的重要途径.除了基本内吞作用, 此过程也受到防御诱导子的调节(Leborgne-Castel et al., 2008).高渗胁迫会导致CME增强、胞吐作用减弱; 低渗胁迫的作用则相反(Zwiewka et al., 2015).Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根和下胚轴中网格蛋白的丰度降低(图1A; 附表3), 这将减弱网格蛋白被招募到PM形成网格蛋白包被小窝(clathrin coated pits, CCPs)的能力, 进而削弱内吞作用, 不利于大豆抵御渗透胁迫.在内吞作用过程中, 动力蛋白(dy- namin)作为GTPase参与调节CCPs从膜上脱离并形成网格蛋白包被小泡的过程(Hinshaw, 2000).拟南芥中, 动力蛋白参与抗病、线粒体分裂及内吞作用中网格蛋白包被小泡的细胞板形成和分裂(Hong et al., 2003; Tang et al., 2006).蛋白质组学研究表明, 冷胁迫下拟南芥幼苗中动力相关蛋白(dynamin-related protein)的丰度增加(Minami et al., 2009) (图1A; 附表4), 这将促进网格蛋白包被小泡的释放, 利于运输货物蛋白. ...
2 2005
... 磷酸肌醇信号途径在植物生长、发育及应对逆境过程中具有重要作用.细胞膜受体接收胞外信号, 激活磷酸肌醇特异的磷脂酶C (phosphatidylinositol-specific phospholipase C, PI-PLC), 将磷脂酰肌醇-4,5-二磷酸水解生成1,4,5-肌醇三磷酸(IP3)和二酰甘油(DAG) (Testerink and Munnik, 2005).PLC通过调节2个重要的信使(IP3和DAG)参与植物各种信号途径.IP3刺激细胞外存储的Ca2+进入细胞质基质, 使胞内Ca2+浓度升高, Ca2+进一步调节信号转导途径(Hunt et al., 2004).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ... ... ).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ...
1 2008
... 在G蛋白质介导的信号通路中, Ca2+作为第二信使, 与各种Ca2+传感蛋白保守的EF-手型结构(EF-hand motif)结合(DeFalco et al., 2010)参与信号转导, 调控胁迫响应基因的表达(Saijo et al., 2000; 范玲玲等, 2010; Meisrimler et al., 2011).蛋白质组学研究表明, 有多种Ca2+信号途径相关蛋白参与胁迫应答过程中的Ca2+平衡调节, 如钙调素蛋白(calmodulin, CaM)和钙网蛋白(calreticulin, CRT)等.渗透胁迫下的大豆根和下胚轴(Nouri and Komatsu, 2010)及盐胁迫下的水稻根尖(Cheng et al., 2009)中, CaM的丰度减少; 盐胁迫的海蓬子根(Nie et al., 2015)及冷胁迫的拟南芥悬浮细胞(Li et al., 2012a)中, 类钙调素蛋白(calmodulin like protein, CML)的丰度增加; 而缺Fe和壳聚糖诱导的豌豆根中CaM和CML的丰度分别升高或降低(Meisrimler et al., 2011) (图1B; 附表4).CaM作为一种钙结合和钙调节蛋白, 在真核生物中高度保守(Meisrimler et al., 2011).CaM本身无酶活性, 当植物受到胁迫刺激后, 细胞内Ca2+水平升高, CaM与Ca2+结合, 形成Ca2+-CaM复合体激活下游的各种靶蛋白(钙调素依赖的蛋白质) (Hoeflich and Ikura, 2002), 从而实现对细胞周期及代谢过程的调控, 增强植物的耐受性.此外, 在大豆根与下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)及玉米根应对铁缺乏和过量(Hopff et al., 2013)胁迫时, CRT蛋白的丰度减少; 而在NaCl (3 mol·L-1)长期胁迫的杜氏盐藻细胞(Katz et al., 2007)、盐和氮胁迫的海蓬子根(Nie et al., 2015)以及冷胁迫的燕麦叶片(Takahashi et al., 2013)中, CRT丰度增加(图1B; 附表3).同时, 钙联接蛋白(calnexin)作为CRT的分子伴侣(Tuteja and Sopory, 2008), 在渗透胁迫的大豆根和下胚轴中的丰度增加(Nouri and Komatsu, 2010) (图1B; 附表3).这些Ca2+结合蛋白传递Ca2+信号, 进而调节下游特定蛋白激酶的活性, 应对胁迫环境. ...
1 2001
... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
2005
1 2005
... 磷酸肌醇信号途径在植物生长、发育及应对逆境过程中具有重要作用.细胞膜受体接收胞外信号, 激活磷酸肌醇特异的磷脂酶C (phosphatidylinositol-specific phospholipase C, PI-PLC), 将磷脂酰肌醇-4,5-二磷酸水解生成1,4,5-肌醇三磷酸(IP3)和二酰甘油(DAG) (Testerink and Munnik, 2005).PLC通过调节2个重要的信使(IP3和DAG)参与植物各种信号途径.IP3刺激细胞外存储的Ca2+进入细胞质基质, 使胞内Ca2+浓度升高, Ca2+进一步调节信号转导途径(Hunt et al., 2004).DAG被二酰甘油激酶迅速磷酸化为磷脂酸(PA), PA作为次级信使发挥作用(Testerink and Munnik, 2005).冷胁迫下, PLC被激活, PLC信号途径参与冷胁迫应答(Vergnolle et al., 2005).此外, 植物PI-PLC也参与ABA信号途径(Mills et al., 2004).蛋白质组学研究表明, 拟南芥悬浮细胞在应对冷和ABA胁迫(Li et al., 2012a), 以及黑麦叶片在应对冷胁迫(Takahashi et al., 2013)时, PI-PLC1和PI-PLC2的丰度均增加(图1B; 附表4), 表明植物PI-PLC信号途径被激活以应对外源ABA和冷胁迫. ...
1 2004
... 植物细胞壁重塑在细胞生长(扩增、分裂和分化)与胁迫应答过程中具重要作用(Ellis et al., 2002; Vogel et al., 2004).高等植物细胞壁主要由高分子量的多糖、蛋白质和木质素等成分组成(Jamet et al., 2006), 这些成分主要通过跨膜运输从细胞内分泌到细胞外(孙颖和孙大业, 2006).其中, 细胞壁蛋白质对于细胞壁结构的形成、细胞内外信号转导以及细胞壁代谢活动具重要作用.许多细胞壁蛋白质与细胞膜在结构和功能上有密切的联系.蛋白质组学研究表明, 一些参与细胞壁重塑的蛋白质存在于PM组分中, 如类成束蛋白阿拉伯半乳聚糖蛋白(fasciclin-like arabinogalactan protein, FLA)、成束蛋白(fasciclin)、葡糖苷酶(glucosidase)、果胶酯酶(pectinesterase)和糖基水解酶(glycosyl hydrolase), 它们可能在植物应对逆境胁迫过程中发挥作用.其中, 在渗透胁迫的大豆根与下胚轴(Nouri and Komatsu, 2010)及缺铁胁迫的玉米根(Hopff et al., 2013)中, FLA丰度降低; 冷和ABA胁迫的拟南芥悬浮细胞中, FLA丰度增加(Li et al., 2012a) (图1E; 附表3).FLA是阿拉伯半乳聚糖蛋白(arabinogalactan protein, AGP)家族成员, 是高度糖基化的蛋白质, 参与调控细胞扩增与分化(Johnson et al., 2003).FLA有成束蛋白结构域, 具细胞黏附功能.一些FLAs通过糖基磷脂酰肌醇(glycosylphospha- tidylinositol, GPI)锚定于PM, 它们可能与受体类蛋白激酶(如细胞壁相关激酶)互作, 参与PM与细胞壁的信号转导过程(Gens et al., 2000; Johnson et al., 2003).拟南芥基因组中至少有21种FLAs, 其中14种FLAs的C端可能由GPI锚定(PM形成膜筏)取代(Harris and Siu, 2002; Johnson et al., 2003).GPI锚定形式的FLAs在细胞扩增过程中参与保持PM的完整性, 非GPI锚定的FLAs被分泌到初生细胞壁中调控细胞扩增(Johnson et al., 2003).No和Loopstra (2000)研究发现, 火炬松(Pinus taeda)下胚轴与上胚轴中的AGP (PtX14A9和PtX3H6)参与木质部的发生过程, 干旱胁迫2天后其mRNA表达均下调.可见, 干旱胁迫导致FLA的丰度降低, 从而使植物细胞的黏附性下降, 细胞膜与初生细胞壁间的黏连被破坏, 进而影响细胞的生长与正常代谢活动. ...
2 2014
... 氮素是植物生长发育过程中必需的大量元素之一, 无机氮吸收、运输和氨基酸分配是植物生长发育所必需的(Popova et al., 2003), 铵盐和硝酸盐是植物吸收的主要氮源(Howitt and Udvardi, 2000).铵态氮的吸收主要通过铵转运蛋白(ammonium transporter, AMT), 而NO3-的吸收主要通过硝酸盐转运蛋白(nitrate transporter, NRT)介导(Von Wittgenstein et al., 2014).铵同化作用需要的能量比硝酸盐少(Bloom et al., 1992), 在铵盐和硝酸盐浓度相同的情况下, 植物优先吸收铵盐(Gazzarrini et al., 1999; Howitt and Udvardi, 2000).蛋白质组学研究表明, 冷胁迫2天及ABA胁迫24小时的拟南芥悬浮细胞中AMT丰度增加(Li et al., 2012a), 而冷胁迫4周的黑麦叶片中AMT丰度降低(Takahashi et al., 2013); NRT的丰度在拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及燕麦叶片应对冷胁迫(Takahashi et al., 2013)时均降低, 表明植物可能优先通过铵转运蛋白吸收无机氮素营养(图1A; 附表3). ... ... 另外, 蛋白质组学研究也表明, 燕麦叶片中肽/硝酸盐转运蛋白(peptide/nitrate transporter)在冷胁迫下丰度降低(Takahashi et al., 2013), 而肽转运蛋白(peptide transporter, PTR)在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)及冷胁迫的黑麦叶片(Takahashi et al., 2013)中丰度增加(图1A; 附表3).PTR又称NRT1/PTR家族, 与低亲和NRT1属于同一基因家族(Von Wittgenstein et al., 2014).寡肽运输是真核生物和原核生物中常见的生理现象, 细胞可通过耗能的方式跨膜运输寡肽, 细胞内的寡肽在肽酶作用下被迅速水解为氨基酸, 用于蛋白质合成或作为碳源和氮源(Song et al., 1996; Stacey et al., 2002).此外, 植物中存在大量的氨基酸转运蛋白(amino acid transporter).其底物专一性、细胞和组织特异性、生长发育及环境条件不同, 表达模式与活性各异.氨基酸转运蛋白包括2个超家族, 即氨基酸-多胺-胆碱家族APC和氨基酸转运蛋白家族ATF.拟南芥中APC转运蛋白主要包括阳离子氨基酸转运蛋白(AtCATs)和γ-氨基丁酸(GABA)转运蛋白; 而ATF家族包括氨基酸通透酶(AAPs)、赖氨酸/组氨酸转运蛋白(LHTs)、脯氨酸转运蛋白(ProTs)、生长素转运蛋白(AUXs)以及芳香族和中性氨基酸转运蛋白(AtANTs) (Liu and Bush, 2006).蛋白质组学研究还表明, 氨基酸转运蛋白在冷胁迫的黑麦叶片中丰度增加(Takahashi et al., 2013), 在渗透胁迫的大豆根和下胚轴中丰度降低(Nouri and Komatsu, 2010) (图1A; 附表3).而在拟南芥悬浮细胞中, 氨基酸通透酶(amino acid permease, AAP)应对ABA胁迫(Li et al., 2012a)时其丰度增加; 在黑麦叶片脯氨酸转运蛋白(proline transporter, ProT)应对冷胁迫时其丰度增加(Takahashi et al., 2013) (图1A; 附表3).已有研究表明, ProT (如拟南芥AtProT2和大麦HvProT)编码基因受水分和盐胁迫诱导(Rentsch et al., 1996; Ueda et al., 2001), 而拟南芥AtAAP4和AtAAP6受水分和盐胁迫抑制(Rent- sch et al., 1996).盐胁迫下, 冰叶日中花(Mesembry- anthemum crystallinum) ProT家族的McAAT1在叶片中上调表达, 而LHT家族的McAAT2在根中下调表达(Popova et al., 2003).这些氨基酸转运蛋白家族成员协同作用来维持胁迫下氨基酸的吸收和分配, 对植物适应逆境非常重要(Liu and Bush, 2006). ...
... 植物细胞PM的特定区域经常与细胞骨架相连(Shibaoka, 1994).质膜与肌动蛋白丝和微管共同调节胞内运输, 对植物形态建成和逆境应答过程中的细胞形态与极性调节非常重要(Schmidt and Panstr- uga, 2007).蛋白质组学研究表明, 在各种胁迫条件下, 肌动蛋白(actin)、微管蛋白(tubulin)、肌球蛋白重链蛋白(myosin heavy chain)、驱动蛋白(kinesin)、类肌动蛋白解聚因子蛋白(actin depolymerizing factor-like protein, ADFP)和前纤维蛋白(profilin)等肌动蛋白结合蛋白(actin-binding proteins, ABPs)的丰度均发生改变(图1C; 附表4).其中, 玉米根应对铁过量时肌动蛋白的丰度增加(Hopff et al., 2013); 海蓬子根应对氮素胁迫(Nie et al., 2015), 拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a), 以及拟南芥幼苗应对冷胁迫(Minami et al., 2009)时肌动蛋白的丰度减少(图1C; 附表4).Pokorná等(2004)的研究表明, 烟草细胞肌动蛋白丝和微管在0°C下立即解聚, 恢复到25°C又重新聚合.Örvar等(2000)的研究表明, 冷胁迫下膜硬化(membrane rigidification)导致肌动蛋白微丝重组, 进而调控苜蓿(M. sativa)适应冷胁迫过程, 说明冷胁迫引起了植物肌动蛋白微丝结构变化.ABPs参与肌动蛋白细胞骨架的重塑, 并调控肌动蛋白微丝的装配和动态(Wasteneys and Yang, 2004).ADFP可改变肌动蛋白动态, 并调节肌动蛋白的解聚合, 而Profilin可结合单体肌动蛋白, 引起肌动蛋白丝解聚合或聚合(Drøbak et al., 2004).蛋白质组学研究表明, 渗透胁迫下大豆根与下胚轴中的ABPs (包括ADFP和Profilin)丰度减少(Nouri and Komatsu, 2010); 冷胁迫下黑麦叶片中的ADFP丰度减少(Taka- hashi et al., 2013), 进而影响肌动蛋白的动态重塑(图1C; 附表4).此外, 蛋白质组学研究还表明, ABA胁迫下拟南芥悬浮细胞中微管蛋白β的丰度增加(Li et al., 2012a); 渗透胁迫下大豆根与下胚轴(Nouri and Komatsu, 2010)以及冷胁迫下拟南芥幼苗(Minami et al., 2009)中, 微管蛋白α和β的丰度减少, 冷胁迫1天拟南芥叶片微管蛋白β的丰度增加, 而冷胁迫3天则减少(Kawamura and Uemura, 2003) (图1C; 附表4).之前的研究表明, 4°C下, 拟南芥叶片中微管蛋白家族的不同成员的转录水平发生变化.例如, TUB2、TUB3、TUB6和TUB8转录水平下调, TUB4、TUB5和TUB7不变, 而TUB9上调(Chu et al., 1993).另外, 蛋白质组学研究还表明, 冷胁迫导致黑麦叶片驱动蛋白的丰度增加(Takahashi et al., 2013).同时, 盐胁迫下水稻根尖(Cheng et al., 2009)和冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)中的肌球蛋白重链蛋白的丰度增加(图1C; 附表4).肌球蛋白作为分子马达蛋白, 以需能的方式沿肌动蛋白丝移动(Holweg and Nick, 2004).拟南芥肌球蛋白XI MYA2突变导致植株矮化, 细胞器运动和生长素极性运输减弱(Holweg and Nick, 2004). ...
1 2002
... 膜联蛋白(annexin, ANX)是一种钙依赖的膜结合蛋白, 在膜运输、离子通道活性调节和磷脂代谢中均发挥作用(Gerke and Moss, 2002).蛋白质组学研究表明, 在冷及ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a)和氮胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度减少; 在冷胁迫的黑麦叶片(Takahashi et al., 2013)和NaCl胁迫的海蓬子根(Nie et al., 2015)中, ANX丰度增加(图1A; 附表3).植物ANX能在PM上形成钙离子通道, ANX丰度的变化可能改变钙通道活性, 引起钙离子跨膜运输变化, 进而调节钙离子的稳态和信号转导过程(White et al., 2002). ...
3 2002
... 钙依赖蛋白激酶(calcium-dependent protein kinase, CDPK)是受Ca2+调节的蛋白质之一(Lee and Rudd, 2002).蛋白质组学研究表明, 水稻Xa21转基因悬浮细胞受壳寡糖诱导(Chen et al., 2007a)及黑麦和燕麦叶片受冷胁迫(Takahashi et al., 2013)时, CDPK的丰度增加; 而冷胁迫下拟南芥悬浮细胞中, CDPK的丰度呈现不同水平的改变(Li et al., 2012a) (图1B; 附表3).CDPK具有丝氨酸/苏氨酸蛋白激酶结构域和C端钙调蛋白样结构域, 4个EF-手型结构可以直接结合Ca2+, 调控下游信号事件(Xiong et al., 2002).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ... ... ).CDPK通过含有的N端十四酰化结构与膜结合(Xiong et al., 2002).水稻OsCPK2与小西葫芦(Cucurbita pepo) CpCPK1可以被十四酰化和棕榈酰化, 并且靶向到PM (Ellard-Ivey et al., 1999; Martín and Busconi, 2000).由此推测CDPKs可能参与非生物胁迫信号转导过程(Hwang et al., 2000).例如, 冷处理下水稻中的CDPK被激活(Martín and Busconi, 2001), 过表达OsCDPK7可增强水稻对冷和渗透胁迫的耐受能力(Saijo et al., 2000). ... ... 磷脂酶D (phospholipase D, PLD)可以水解磷脂生成PA (Xiong et al., 2002), PA也可作为第二信使参与胁迫信号转导, 调控下游基因表达等事件(Munnik, 2001).蛋白质组学研究表明, 拟南芥悬浮细胞应对冷胁迫时PLD受到影响(Li et al., 2012a); 拟南芥悬浮细胞应对ABA胁迫(Li et al., 2012a)及拟南芥叶片应对冷胁迫(Kawamura and Uemura, 2003)时, PLDδ丰度均增加(图1B; 附表4).之前, Li等(2004)的研究表明, 拟南芥PLDδ突变体对冷害更敏感, 而PLDδ过表达体的抗冻性增强, 表明PLD参与的信号转导途径在细胞低温应答中具重要作用. ...
1 2001
... 细胞从膜受体接受逆境信号后, 利用磷酸化蛋白质对信号进行级联转换和放大, 蛋白质可逆磷酸化在植物细胞信号转导过程中具重要作用(Xiong and Zhu, 2001).蛋白质可逆磷酸化通过蛋白激酶和磷酸酶完成, 主要发生在丝氨酸、苏氨酸和酪氨酸等氨基酸位点.丝氨酸/苏氨酸蛋白激酶能使靶蛋白的丝氨酸和苏氨酸残基磷酸化.蛋白质组学研究表明, 在黑麦和燕麦叶片应对冷胁迫(Takahashi et al., 2013)以及拟南芥悬浮细胞应对冷和ABA胁迫(Li et al., 2012a)时, 丝氨酸/苏氨酸蛋白激酶丰度发生不同水平的改变; 在大豆根与下胚轴应对水淹胁迫时, 蛋白激酶丰度增加(Komatsu et al., 2009).此外, 蛋白质组学研究还表明, 冷胁迫下的拟南芥悬浮细胞中组氨酸激酶(histidine kinase)丰度减少(Li et al., 2012a), 从而调控相应靶蛋白的磷酸化水平, 应答环境胁迫(图1B; 附表3). ...
1 2013
... 质膜(plasma membrane, PM)是细胞原生质体与外界进行物质交换和信息交流的基础, 对完成细胞间的各种代谢活动及以细胞分裂与分化为基础的生长发育过程均具重要作用.对植物细胞而言, 物质跨PM运输是细胞壁生长与代谢的基础, PM与细胞壁一起成为细胞应答外界环境的屏障(Mongrand et al., 2010).质膜与细胞内膜系统和各种细胞器膜构成了细胞内外信号分子、离子、激素以及其它代谢物等进行交换的媒介, 对细胞生长以及环境应答至关重要(Yadeta et al., 2013).在PM磷脂双分子层上, 以各种形式分布的蛋白质(整合蛋白、外周蛋白和膜锚定蛋白等)是PM行使功能的基础.它们在构成PM基本结构、感知与转导信号、细胞内外物质运输以及防御胁迫等方面都发挥重要作用.因此, 深入解析植物PM蛋白质的组成、分布与功能对认识细胞信号与代谢活动的分子机制具有重要意义. ...
1 2004
... H+-ATPase是ATP依赖的跨膜质子泵, 根据功能和位置的不同, 可分为PM H+-ATPase (P(H+)-ATPase)、液泡膜H+-ATPase (V(H+)-ATPase)以及线粒体H+- ATPase (F(H+)-ATPase) 3种.它们对于调节细胞质pH值、膜电位及细胞膨压均具重要作用(Michelet and Boutry, 1995; Minami et al., 2009).P(H+)-ATPase包含磷酸酶结构域、转导结构域和蛋白激酶结构域, 参与调节细胞内外质子的平衡.蛋白质组学研究表明, P(H+)-ATPase参与了多种胁迫应答过程.Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根与下胚轴中P(H+)-ATPase的丰度增加(图1A; 附表3).与之相似, 在水分亏缺处理55和96小时的大豆根中P(H+)-ATPase mRNA的丰度明显增加(Surowy and Boyer, 1991), 其增加很可能是为了补偿聚乙二醇(polyethylene glycol, PEG)对P(H+)-ATPase磷酸酶结构域催化活性抑制所致的P(H+)-ATPase ATP水解酶活性降低, 进而保证植物在低水势下生长(Qiu and Zhang, 2000).此外, 蛋白质组学研究还表明, P(H+)-ATPase在冷和ABA胁迫的拟南芥悬浮细胞中的丰度受到影响(Li et al., 2012a); 在冷胁迫的黑麦叶片(Takahashi et al., 2013), 铁缺乏和过量的玉米根(Hopff et al., 2013), 以及长期NaCl (3 mol·L-1)处理的杜氏盐藻细胞(Katz et al., 2007)中丰度增加(图1A; 附表3).另有报道, P(H+)-ATPase编码基因LHA8在番茄(Lycopersicon esculentum)叶片与根应答盐胁迫时上调表达, 在应答与其等渗的PEG胁迫时则没有变化, 表明LHA8可能主要参与Na+平衡的调节(Kalampanayil and Wimmers, 2001).与之相似, 盐胁迫下, P(H+)-ATPase在小麦(Triticum aestivum)品种Longchun20根中的丰度与活性都上升(Yang et al., 2004).这表明渗透胁迫以及盐胁迫下, P(H+)- ATPase的丰度与活性增加可能有利于离子的外流, 从而维持膜内外离子的平衡. ...
1 2002
... 在植物中, G蛋白(GTP binding protein)与小G蛋白(small GTP binding protein)参与应答多种胁迫(激素、干旱、臭氧和病原菌)信号通路, 是重要的分子开关(Yang, 2002; Perfus-Barbeoch et al., 2004).G蛋白通过与G蛋白耦联受体作用感知并转换细胞外信号, 激活细胞内的效应器酶(Jones and Assmann, 2004).蛋白质组学研究表明, 在盐、冷和ABA胁迫下, G蛋白和小G蛋白的丰度发生变化.例如, 在冷和/或ABA胁迫下, 拟南芥悬浮细胞中G蛋白、小G蛋白Rab和Ran2丰度增加, 小G蛋白Ras类GTP结合蛋白丰度减少(Li et al., 2012a); 冷胁迫下黑麦叶片中小G蛋白Rho丰度减少(Takahashi et al., 2013); 盐胁迫下海蓬子根中Rab11C丰度增加, Rab7b丰度减少(Nie et al., 2015).同时, 冷胁迫下拟南芥悬浮细胞(Li et al., 2012a)和水稻根(Hashimoto et al., 2009)中, ADP核糖基化因子(ADP-ribosylation factor, ARF)丰度均增加(图1B; 附表4).ARF属于酰化的小GTP结合蛋白(约21 kDa), 其与Ras-GTPase超家族和异源三聚体G蛋白α亚基成员的结构相似(Boman and Kahn, 1995).此外, Ma等(2015)研究发现, 水稻COLD1 (CHILLING TOLERANCE DIVERGENCE 1)可增强水稻G蛋白α1活性, 引起Ca2+内流进入细胞质, 引发下游冷胁迫应答基因的表达, 提高水稻的耐寒性.上述研究结果表明, G蛋白可与活化的G蛋白偶联受体作用, 感知并转换冷胁迫信号以应对冷胁迫. ...
1 2006
... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ...
1 2002
... 硫是植物生长发育必需的大量营养元素之一, 植物根主要以硫酸盐(SO42-)的形式从土壤中吸收硫元素(Leustek and Saito, 1999).硫酸盐转运蛋白(sulphate transporter, SULTR)是一类参与SO42-主动运输的载体蛋白, 与硫元素的吸收、同化和长距离运输有关(Leustek and Saito, 1999; Leustek et al., 2000).在高水平硫条件下, Sultr1;2呈现高表达; 在硫缺乏条件下, 拟南芥Sultr1;1与Sultr1;2均上调表达, 但Sultr1;2对硫缺乏的响应能力比Sultr1;1低(Yoshimoto et al., 2002).Shibagaki等(2002)对Sultr1;2突变体进行了分析, 发现该突变体对硫酸盐的吸收活性明显降低, Sultr1;2转运体对拟南芥根部吸收硫酸盐起重要作用.Sultr1;2敲除突变体中, Sultr- 1;1 mRNA水平上调, 但是不能恢复硫酸盐的吸收能力(Maruyama-Nakashita et al., 2003), 表明Sultr1;1和Sultr1;2均参与硫酸盐的吸收, 但二者对硫酸盐的吸收能力不同.蛋白质组学研究表明, 冷胁迫下的黑麦叶片中SULTR丰度下降(Takahashi et al., 2013) (图1A; 附表3), 这可能引起硫素的吸收和运输受限, 影响植物的生长. ...
1 2012
... 植物细胞表面的受体蛋白激酶(receptor protein kinase, RPK)在感知外部环境信号中发挥重要作用(De Lorenzo et al., 2009; Zhang et al., 2012).定位于PM的RPK包含胞外配体结合域、跨膜结构域和细胞质激酶结构域, 其胞外结构域能选择性地响应各种信号(De Lorenzo et al., 2009), 并通过蛋白激酶结构域转换信号.蛋白质组学研究表明, 水稻根尖、大豆根与下胚轴、黑麦和燕麦叶片及拟南芥悬浮细胞的RPK应对盐、渗透、冷和ABA胁迫(Cheng et al., 2009; Nouri and Komatsu, 2010; Li et al., 2012a; Takahashi et al., 2013), 拟南芥悬浮细胞中富含亮氨酸重复序列的蛋白激酶(leucine-rich repeat protein kinase)应对冷和ABA胁迫(Li et al., 2012a), 海蓬子根与拟南芥悬浮细胞中受体丝氨酸/苏氨酸蛋白激酶(receptor serine/threonine kinase, RSTK)应对氮素、盐、冷和ABA (Li et al., 2012a; Nie et al., 2015)等逆境胁迫时丰度发生变化(图1B; 附表3).其中, 冷胁迫的高度耐寒黑麦叶片中, RPK丰度增加; 而耐旱性低的燕麦叶片中, RPK丰度减少(Takahashi et al., 2013).同时, 盐胁迫的水稻根尖中, RPK丰度增加(Cheng et al., 2009); 渗透胁迫的大豆根和下胚轴中RPK丰度减少(Nouri and Komatsu, 2010).De Lorenzo等(2009)研究发现, 蒺藜苜蓿(Medicago truncatula)受150 mmol·L-1 NaCl胁迫1和6小时, 其根中富含亮氨酸重复序列的受体类蛋白激酶(LRR- RLK)基因Srlk受到诱导; 并且NaCl (150 mmol·L-1)胁迫6小时后, Srlk-RNAi (RNA interference)的根和Srlk突变体中早期盐调节基因(钙依赖性蛋白激酶CDPK3和转录因子Zpt2-1)表达下调, 表明Srlk参与根部盐胁迫应答过程中受体激酶调控的信号转导过程.此外, 蛋白质组学研究表明, 在铁缺乏和过量时玉米根中赤霉素受体(Gibberellin receptor GID1L2) (Hopff et al., 2013) (图1B; 附表4), 在冷和ABA胁迫时拟南芥悬浮细胞类受体蛋白(receptor like protein) (Li et al., 2012a)的丰度均增加; 在冷和ABA胁迫的拟南芥悬浮细胞中, 富含亮氨酸重复序列蛋白(Leucine- rich repeat protein)的丰度也增加或降低(Li et al., 2012a); 而在冷胁迫的黑麦和燕麦叶片中该受体蛋白的丰度增加(Takahashi et al., 2013) (图1B; 附表3).可见, 这些受体蛋白参与介导植物感知冷、ABA和Fe信号, 进而启动特定的信号转导通路. ...
1 2008
... 此外, PATL (Patellin)是一种载体蛋白, 含有磷脂酰肌醇转移蛋白(phosphatidylinositol transfer protein, Sec14)样脂质结合结构域(Sec14p-like lipid-binding domain).Sec14可转移磷脂质并调控脂质信号分子产物(Mousley et al., 2007).PATL参与膜运输、细胞板扩大和成熟, 在细胞板生物合成过程中可结合疏水分子(如磷酸肌醇)并促进其在不同细胞位点间转移(Zhang et al., 2008).在拟南芥幼苗(Minami et al., 2009)和燕麦叶片(Takahashi et al., 2013)应对冷胁迫及大豆根和下胚轴应对渗透胁迫(Nouri and Komatsu, 2010)时, PATL丰度降低; 而在拟南芥悬浮细胞应对ABA胁迫时, PATL丰度增加(Li et al., 2012a) (图1A; 附表4), 这可能对胁迫条件下膜运输及细胞分裂过程有一定的影响. ...
4 2009
... 近年来, 关于植物PM逆境(生物胁迫(如白叶枯病菌(Xanthomonas oryzae pv. oryzae, Xoo)感染)以及非生物胁迫(冷、盐、水淹、渗透、高pH值、Fe缺乏及过量、脱落酸、壳聚糖和壳寡糖))应答蛋白质组学研究已有大量报道.拟南芥(Kawamura and Uemura, 2003; Minami et al., 2009; Li et al., 2012a)、水稻(Oryza sativa) (Chen et al., 2007a, 2007b; Ma- lakshah et al., 2007; Hashimoto et al., 2009; Cheng et al., 2009)、大豆(Glycine max) (Komatsu et al., 2009; Nouri and Komatsu, 2010)、玉米(Zea mays) (Hopff et al., 2013)、豌豆(Pisum sativum) (Meisrim- ler et al., 2011)、黑麦(Secale cereale) (Takahashi et al., 2013)、燕麦(Avena sativa) (Takahashi et al., 2013)、海蓬子(Salicornia europaea) (Nie et al., 2015)、杜氏盐藻(Dunaliella salina) (Katz et al., 2007)及集胞藻属(Synechocystis)植物(Huang et al., 2006; Zhang et al., 2009)幼苗、叶片、根和下胚轴等器官, 以及单细胞PM在应答上述各种胁迫过程中的丰度差异蛋白质见表1.我们整合分析了上述10种植物中参与各种逆境应答过程的723种PM蛋白质, 利用PSI以及PHI-BLAST分析(http://www.ncbi.nlm.nih. gov/BLAST/), 对其中89种推测的蛋白质(Predicted/ hypothetical/putative proteins)进行了注释; 同时, 我们根据TMHMM2.0服务器(http://www.cbs.dtu.dk/ services/TMHMM)跨膜结构域预测的结果, 将这些蛋白质分为具有跨膜结构域的整合蛋白(486种) (附表1)和膜相关蛋白(包括外周蛋白、膜锚定蛋白以及其它未检测到跨膜结构域的蛋白质) (237种)两类(附表2).在此基础上, 通过分析不同物种中蛋白质的同源性与功能, 将这些蛋白质整理为非冗余的整合蛋白(100种) (附表3)和膜相关蛋白(70种) (附表4).这些蛋白质丰度的动态变化为认识逆境应答过程中PM参与的跨膜转运、信号转导、细胞骨架动态、细胞壁重塑及胁迫防御等生物学事件的分子调控机制提供了新线索. ... ... 植物ABC (ATP-binding cassette)转运体参与多种物质的跨膜运输, 尤其是胁迫相关的内源性次生代谢物(如生物碱、萜类化合物、酚类和蜡质)的跨膜运输, 具有底物专一性(Yazaki, 2006).蛋白质组学研究表明, ABC转运蛋白在冷胁迫的燕麦和黑麦叶片中丰度增加(Takahashi et al., 2013); 同时, 在冷和ABA胁迫的拟南芥悬浮细胞(Li et al., 2012a), 以及盐和高pH胁迫的集胞藻细胞(Huang et al., 2006; Zhang et al., 2009) PM中, 多种ABC转运蛋白的丰度发生变化(图1A; 附表3), 这些蛋白质主要参与转运硝酸/亚硝酸、寡肽、磷酸和葡糖基甘油等.其中, 参与转运硝酸/磺酸/碳酸盐、磷酸盐和离子的ABC转运蛋白丰度增加(如高pH条件下的Na+依赖的碳酸盐转运蛋白SbtB和磷酸盐转运蛋白PstB), 而参与尿素(如UrtA)和碳酸氢盐转运(如CmpA)的ABC转运蛋白的丰度降低(Huang et al., 2006; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ... ... ; Zhang et al., 2009).这些ABC转运蛋白丰度的改变有助于调节相应底物的转运效率, 从而克服高盐或高pH造成的营养缺乏(Huang et al., 2006; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ... ... ; Zhang et al., 2009).此外, Zhang等(2009)研究发现, 集胞藻细胞中参与阴离子转运的ABC转运蛋白的丰度受到高pH胁迫的抑制.这与之前发现的高pH会导致ATPase活性下降(Randall and Sze, 1986)和阴离子吸收比率降低(Lin, 1981)等相一致. ...
1 2015
... 网格蛋白介导的内吞作用(clathrin-mediated endocytosis, CME)是网格蛋白作为受体介导内吞作用的主要方式, 是植物细胞内向膜泡运输(vesicle transport)的重要途径.除了基本内吞作用, 此过程也受到防御诱导子的调节(Leborgne-Castel et al., 2008).高渗胁迫会导致CME增强、胞吐作用减弱; 低渗胁迫的作用则相反(Zwiewka et al., 2015).Nouri和Komatsu (2010)研究发现, 渗透胁迫下大豆根和下胚轴中网格蛋白的丰度降低(图1A; 附表3), 这将减弱网格蛋白被招募到PM形成网格蛋白包被小窝(clathrin coated pits, CCPs)的能力, 进而削弱内吞作用, 不利于大豆抵御渗透胁迫.在内吞作用过程中, 动力蛋白(dy- namin)作为GTPase参与调节CCPs从膜上脱离并形成网格蛋白包被小泡的过程(Hinshaw, 2000).拟南芥中, 动力蛋白参与抗病、线粒体分裂及内吞作用中网格蛋白包被小泡的细胞板形成和分裂(Hong et al., 2003; Tang et al., 2006).蛋白质组学研究表明, 冷胁迫下拟南芥幼苗中动力相关蛋白(dynamin-related protein)的丰度增加(Minami et al., 2009) (图1A; 附表4), 这将促进网格蛋白包被小泡的释放, 利于运输货物蛋白. ...
 ,
, 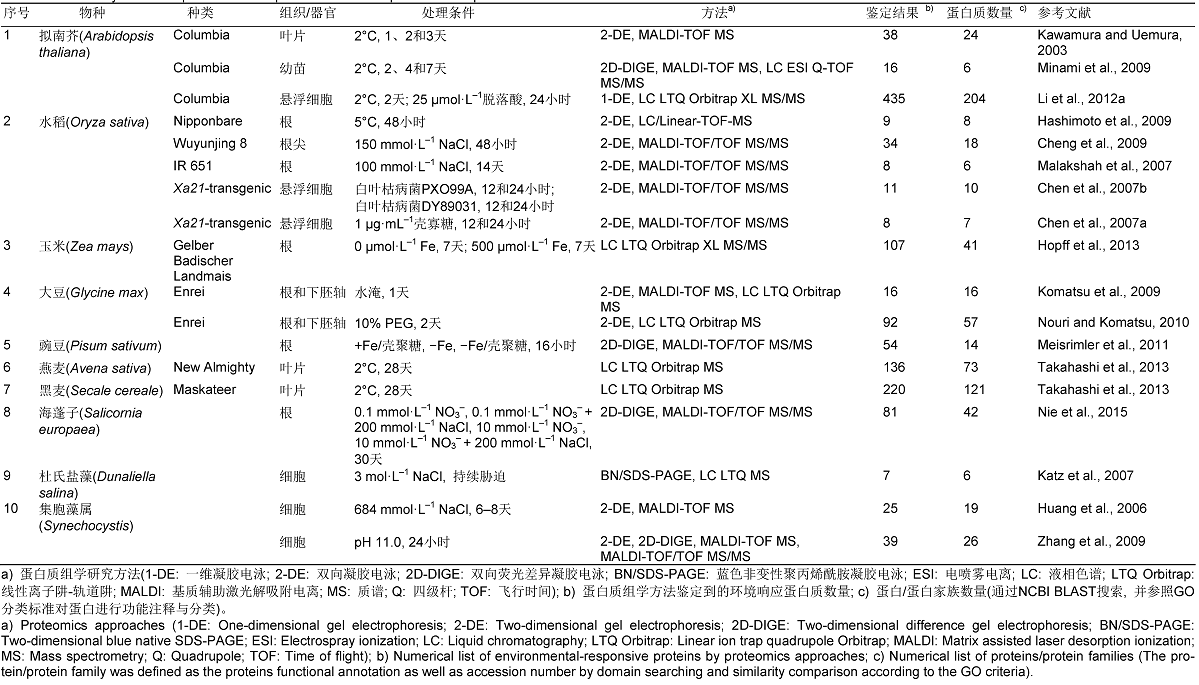
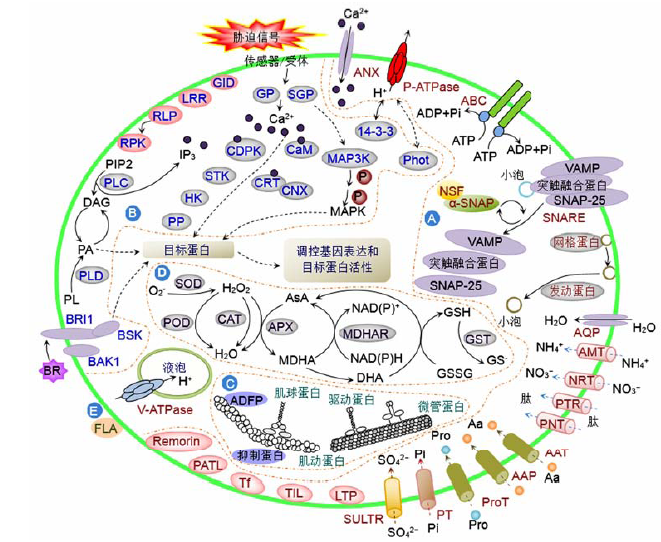

{kind=link}
{kind=link}