Abstract: Sporangia are propagative organs of ferns and their morphology has great significance for fern taxonomy and phylogeny. In this study, we used sodium hypochlorite solution to observe fern sporangia. By this process, we could obtain sporangia photos under light microscopy. We studied the sporangia morphology of 13 species belonging to four genera of the fern family Lindsaeaceae and found that the shape of the capsule is ellipsoidal, with a vertical annulus; the pedicel is composed of three rows of cells. The cell number annulus is reduced in Odontosoria, Osmolindsaea, Tapeinidium and Lindsaea, whereas the volume of capsule and cell number of stomium and capsule are reduced in Odontosoria, Tapeinidium, Osmolindsaea and Lindsaea. As well, the same genus shows more differences between species, such as Odontosoria biflora and O. chinensis as well as Osmolindsaea odorata and Os. japonica, but less difference between the genera of Tapeinidium and Lindsaea. Studies of sporangia morphology will be useful for further research in other groups of ferns.
Lindsaea Dryand. ex Sm. Odontosoria Fée Ormoloma Maxon Sphenomeris Maxon Tapeinidium (C.Presl) C.Chr. Schizoloma Gaud. Stenoloma Fée Xyropteris Kramer Cystodium J. Sm. Lonchitis L.
Lindsaea Dryand. ex Sm. (Synonym: Ormoloma; Schizoloma) Odontosoria Fée Sphenomeris Maxon (Synonym: Stenoloma) Tapeinidium (C.Presl) C.Chr. Osmolindsaea (K. U. Kramer) Lehtonen & Christenh. Nesolindsaea Lehtonen & Christenh.
Xyropteris Kramer Cystodium J. Sm. Lonchitis L.
Presl, 1836; Christensen, 1938; Holttum, 1947; Tryon and Tryon, 1982; Kramer and Green, 1990; Lehtonen et al., 2010
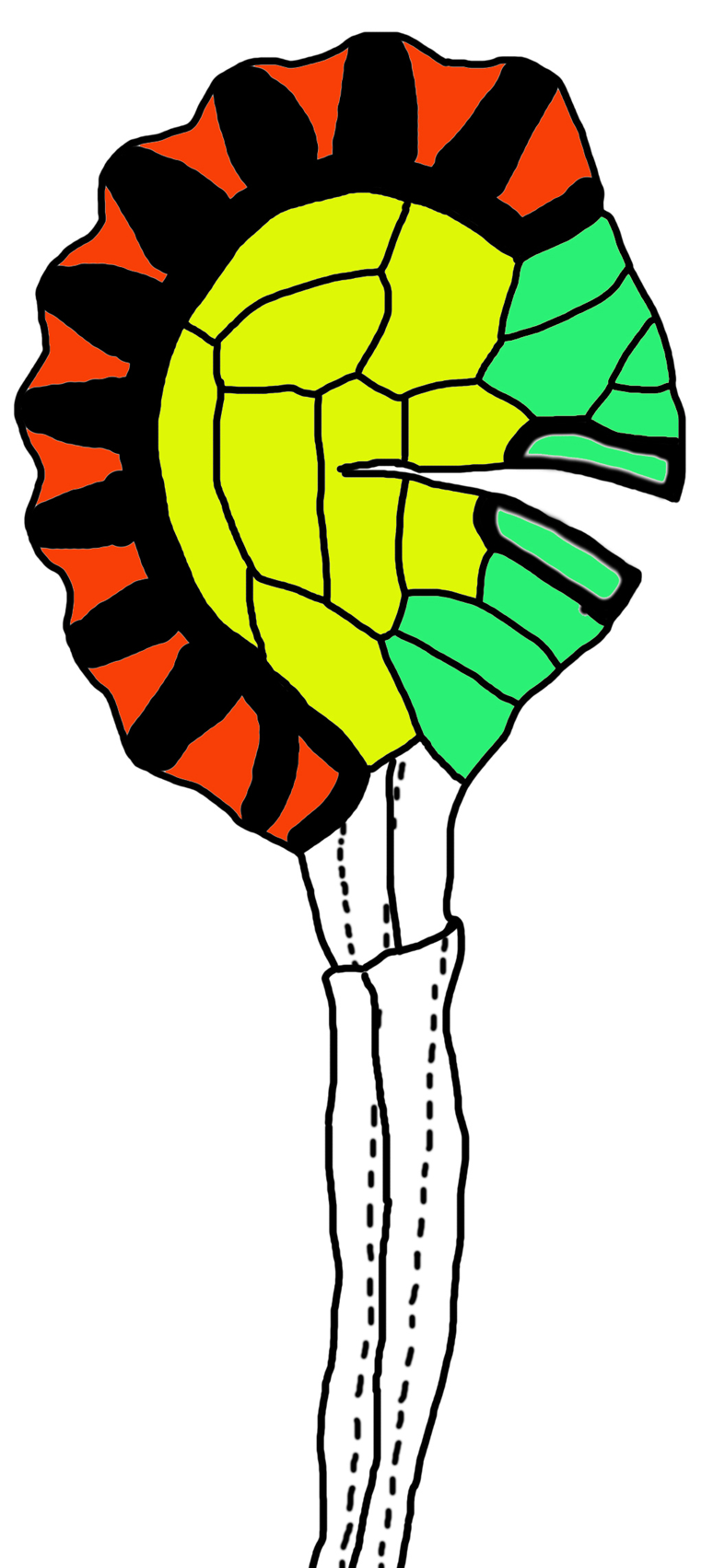
1.2 方法所取材料为当年已成熟的孢子囊, 且里面的孢子已散落。将孢子囊从叶片上攫取出来制成水装片(不盖盖玻片), 向中央滴加2滴次氯酸钠, 将材料完全浸没。由于次氯酸钠有氧化性, 因此可看到材料的颜色逐渐变深, 然后又逐渐变浅, 此时可在体视镜下持续观察。在孢子囊环带颜色褪去之前向材料滴加2滴蒸馏水稀释次氯酸钠, 以免材料过度褪色。然后用滤纸吸掉玻片上的次氯酸钠, 滴加2滴蒸馏水, 再用滤纸吸掉, 然后再次滴加2滴蒸馏水, 盖上盖玻片观察并拍照。在用滤纸吸收时, 注意不要将玻片上的溶液完全吸干, 要保证孢子囊一直有溶液浸润其中, 以免有气泡进入。上述褪色过程大概持续10-20分钟, 制备好的孢子囊环带颜色呈黄棕色, 囊壁略呈无色, 且囊壁上看不到明显杂质。 将上述制备的装片盖上盖玻片, 置于光学显微镜下观察, 选取形态特征清晰完整的孢子囊拍照并保存。由于孢子囊呈椭球形, 所用显微镜拍照软件需要有景深合成功能, 或者用第三方景深合成软件。根据照片记录每个孢子囊的环带细胞数、囊壁细胞数、唇细胞数和唇细胞加厚细胞数, 测量囊柄长度以及孢子囊蒴的长度和宽度, 再根据囊蒴的长和宽计算囊蒴的近似体积, 即囊蒴体积=(4/3)π[(囊蒴长+囊蒴宽)/4]3。记录环带细胞个数时每个种随机选取30个完整的孢子囊, 测量其它数据时选取10个正常发育的成熟孢子囊。孢子囊形态特征如图2所示。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_2.png<b>图2</b> 已开裂的具纵行环带的孢子囊示意图<br/>下面未着色的为孢子囊柄, 上面着色部分为孢子囊蒴。棕色部分为环带。粗黑线为加厚的细胞壁。绿色部分为唇细胞, 唇细胞中间位置壁较厚的为唇细胞加厚细胞。 黄色部分为孢子囊壁细胞。<br/><b>Figure 2</b> Sporangium with vertical annulus<br/>The pedicel is uncolored, the colored part is capsule. The annulus in brown color. Bold black line shows thick-wall. The stomium in green color, and with thick-walled stomium in centre. The wall of capsule in yellow color. Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_2.png<b>图2</b> 已开裂的具纵行环带的孢子囊示意图<br/>下面未着色的为孢子囊柄, 上面着色部分为孢子囊蒴。棕色部分为环带。粗黑线为加厚的细胞壁。绿色部分为唇细胞, 唇细胞中间位置壁较厚的为唇细胞加厚细胞。 黄色部分为孢子囊壁细胞。<br/><b>Figure 2</b> Sporangium with vertical annulus<br/>The pedicel is uncolored, the colored part is capsule. The annulus in brown color. Bold black line shows thick-wall. The stomium in green color, and with thick-walled stomium in centre. The wall of capsule in yellow color.
图2 已开裂的具纵行环带的孢子囊示意图 下面未着色的为孢子囊柄, 上面着色部分为孢子囊蒴。棕色部分为环带。粗黑线为加厚的细胞壁。绿色部分为唇细胞, 唇细胞中间位置壁较厚的为唇细胞加厚细胞。 黄色部分为孢子囊壁细胞。 Figure 2 Sporangium with vertical annulus The pedicel is uncolored, the colored part is capsule. The annulus in brown color. Bold black line shows thick-wall. The stomium in green color, and with thick-walled stomium in centre. The wall of capsule in yellow color.
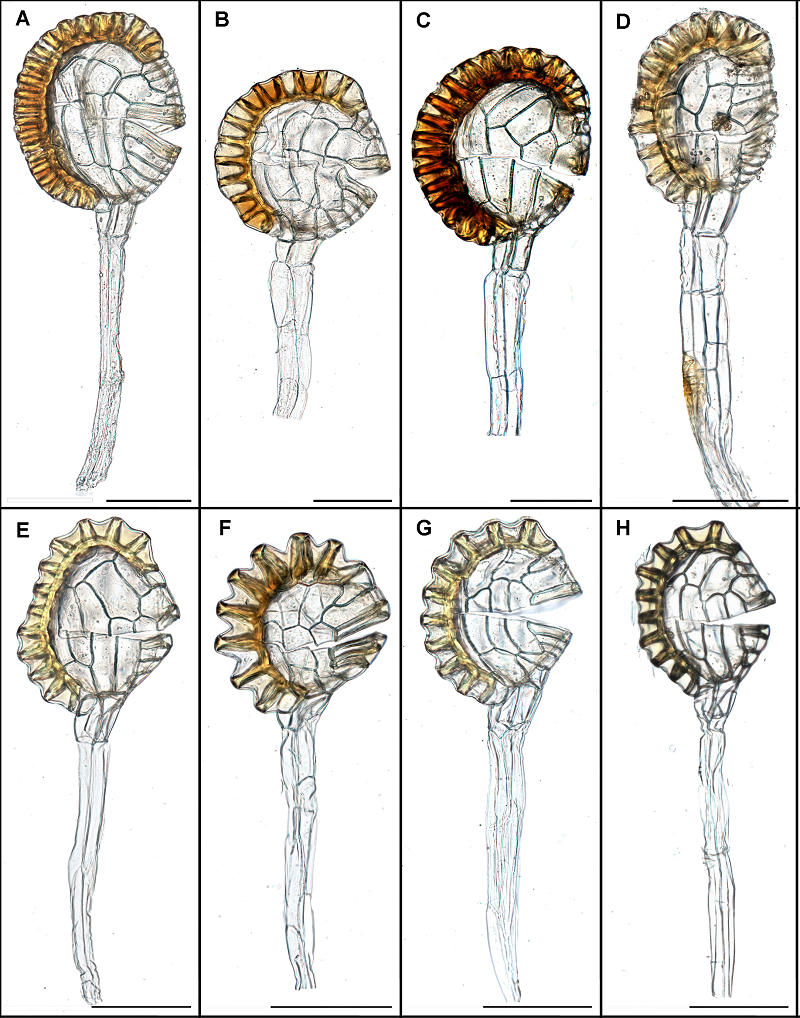
图3 鳞始蕨科孢子囊形态 (A) 阔片乌蕨; (B) 乌蕨; (C) 香鳞始蕨; (D) 日本鳞始蕨; (E) 线片鳞始蕨; (F) 攀缘鳞始蕨; (G) 爪哇鳞始蕨; (H) 团叶鳞始蕨。Bar=100 μm。图中囊柄有断裂情况, 故其长度不做比较。 Figure 3 Sporangia morphology of Lindsaeaceae (A) Odontosoria biflora; (B) O. chinensis; (C) Osmolindsaea odorata; (D) Os. japonica; (E) Lindsaea eberhardtii; (F) L. merrillii subsp. yaeyamensis; (G) L. javanensis; (H) L. orbiculata. Bar=100 μm. The length of pedicel can not be used for comparing since some of it was broken.
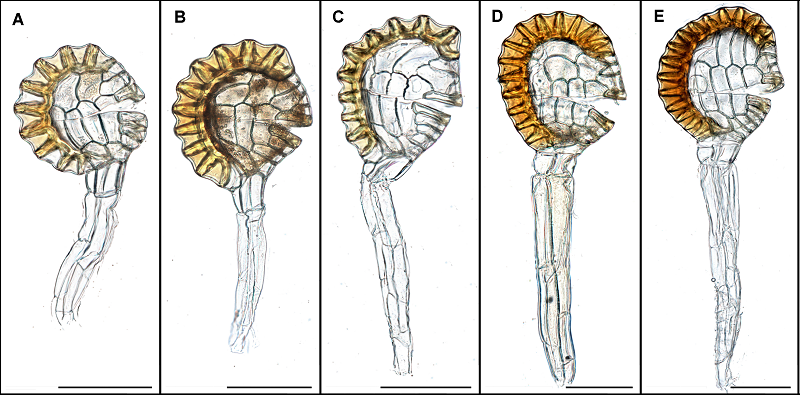
图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_4.png<b>图4</b> 鳞始蕨科孢子囊形态<br/>(A) 网脉鳞始蕨; (B) 异叶鳞始蕨; (C) 剑叶鳞始蕨; (D) 达边蕨; (E) 二羽达边蕨。Bar=100 μm。图中囊柄有断裂情况, 故其长度不做比较。<br/><b>Figure 4</b> Sporangia morphology of Lindsaeaceae<br/>(A) <i>Lindsaea cultrata</i>; (B) <i>L. heterophylla</i>; (C) <i>L. ensifolia</i>; (D) <i>Tapeinidium pinnatum</i>; (E) <i>T. pinnatum</i> var. <i>biserratum</i>. Bar=100 μm. The length of pedicel can not be used for comparing since some of it was broken. Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_4.png<b>图4</b> 鳞始蕨科孢子囊形态<br/>(A) 网脉鳞始蕨; (B) 异叶鳞始蕨; (C) 剑叶鳞始蕨; (D) 达边蕨; (E) 二羽达边蕨。Bar=100 μm。图中囊柄有断裂情况, 故其长度不做比较。<br/><b>Figure 4</b> Sporangia morphology of Lindsaeaceae<br/>(A) <i>Lindsaea cultrata</i>; (B) <i>L. heterophylla</i>; (C) <i>L. ensifolia</i>; (D) <i>Tapeinidium pinnatum</i>; (E) <i>T. pinnatum</i> var. <i>biserratum</i>. Bar=100 μm. The length of pedicel can not be used for comparing since some of it was broken.
图4 鳞始蕨科孢子囊形态 (A) 网脉鳞始蕨; (B) 异叶鳞始蕨; (C) 剑叶鳞始蕨; (D) 达边蕨; (E) 二羽达边蕨。Bar=100 μm。图中囊柄有断裂情况, 故其长度不做比较。 Figure 4 Sporangia morphology of Lindsaeaceae (A) Lindsaea cultrata; (B) L. heterophylla; (C) L. ensifolia; (D) Tapeinidium pinnatum; (E) T. pinnatum var. biserratum. Bar=100 μm. The length of pedicel can not be used for comparing since some of it was broken.
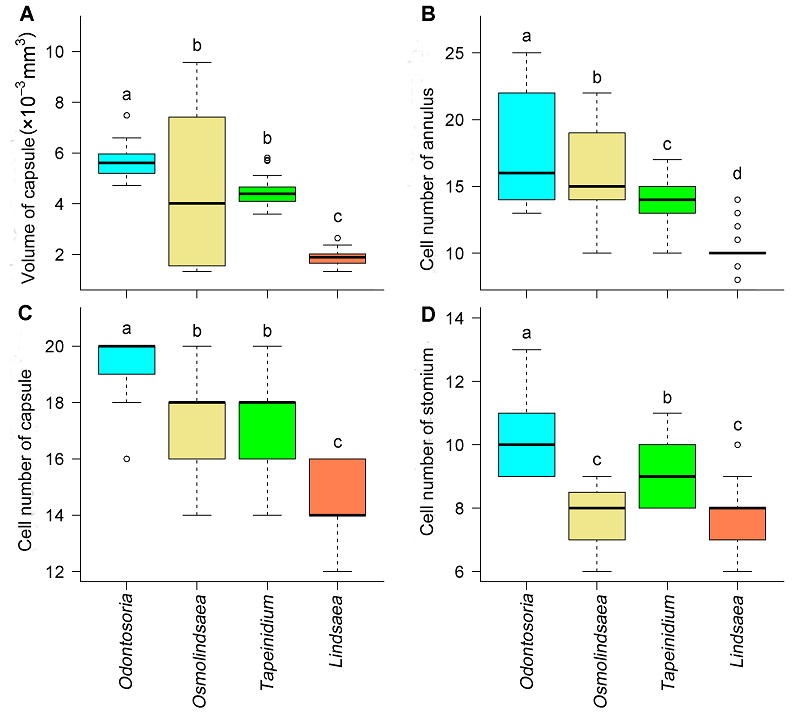
2.6 形态数据的比较根据表3的形态数据, 利用方差分析与Turkey’s多重比较法, 比较孢子囊囊蒴体积、环带细胞数、囊壁细胞数和唇细胞数4组形态数据在属间的差异, 所得结果如图5。 图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_5.png<b>图5</b> 鳞始蕨科4属孢子囊形态数据比较<br/>(A) 囊蒴体积; (B) 环带细胞数; (C) 囊壁细胞数; (D) 唇细胞数。无共同字母表示属间差异显著(Turkey’s多重比较, <i>P</i><0.05)。在每个箱线图中, 箱体显示25%、中位数和75%的四分位数。箱体外的虚线延伸至距箱体1.5倍四分位距(箱体长度)范围内的极值, 超过此限的极值以散点表示。<br/><b>Figure 5</b> Comparisons of sporangia morphology among four genera in Lindsaeaceae<br/>(A) Volume of capsule; (B) Cell number of annulus; (C) Cell number of capsule; (D) Cell number of stomium. Genera with no letters in common are significantly different (multiple comparisons using Turkey’s method, <i>P</i><0.05). For each box-and-whisker plot, the box shows 25%, median and 75% quantile of the given values. The whiskers extend to the most extreme data points that are not more than 1.5 times the interquartile range (length of the box) from the box. Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_5.png<b>图5</b> 鳞始蕨科4属孢子囊形态数据比较<br/>(A) 囊蒴体积; (B) 环带细胞数; (C) 囊壁细胞数; (D) 唇细胞数。无共同字母表示属间差异显著(Turkey’s多重比较, <i>P</i><0.05)。在每个箱线图中, 箱体显示25%、中位数和75%的四分位数。箱体外的虚线延伸至距箱体1.5倍四分位距(箱体长度)范围内的极值, 超过此限的极值以散点表示。<br/><b>Figure 5</b> Comparisons of sporangia morphology among four genera in Lindsaeaceae<br/>(A) Volume of capsule; (B) Cell number of annulus; (C) Cell number of capsule; (D) Cell number of stomium. Genera with no letters in common are significantly different (multiple comparisons using Turkey’s method, <i>P</i><0.05). For each box-and-whisker plot, the box shows 25%, median and 75% quantile of the given values. The whiskers extend to the most extreme data points that are not more than 1.5 times the interquartile range (length of the box) from the box.
图5 鳞始蕨科4属孢子囊形态数据比较 (A) 囊蒴体积; (B) 环带细胞数; (C) 囊壁细胞数; (D) 唇细胞数。无共同字母表示属间差异显著(Turkey’s多重比较, P<0.05)。在每个箱线图中, 箱体显示25%、中位数和75%的四分位数。箱体外的虚线延伸至距箱体1.5倍四分位距(箱体长度)范围内的极值, 超过此限的极值以散点表示。 Figure 5 Comparisons of sporangia morphology among four genera in Lindsaeaceae (A) Volume of capsule; (B) Cell number of annulus; (C) Cell number of capsule; (D) Cell number of stomium. Genera with no letters in common are significantly different (multiple comparisons using Turkey’s method, P<0.05). For each box-and-whisker plot, the box shows 25%, median and 75% quantile of the given values. The whiskers extend to the most extreme data points that are not more than 1.5 times the interquartile range (length of the box) from the box.
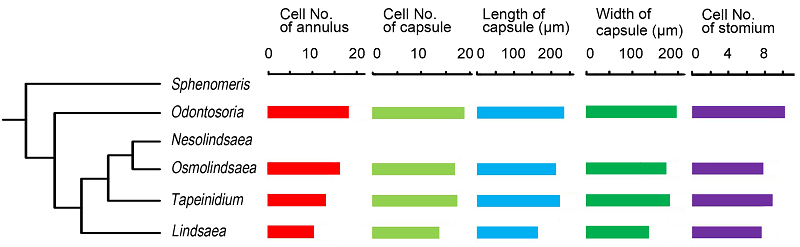
2.7 讨论2.7.1 中国鳞始蕨科各属之间孢子囊的形态变化 鳞始蕨科全世界已确定有6属, 约200种, 中国有4属18种, 分别是乌蕨属、香鳞始蕨属、达边蕨属和鳞始蕨属, 中国不产的2属为Sphenomeris和Nesolind- saea。各属之间的系统关系如图6所示(Lehtonen et al., 2010)。由表3可知, 乌蕨属、香鳞始蕨属、达边蕨属和鳞始蕨属的孢子囊形态有一定的规律性, 如环带细胞数逐渐减少, 分别为17.9、16.5、13.6和10.2个; 囊蒴体积、唇细胞数和囊壁细胞数具有相同的变化规律, 即从大(多)到小(少)依次为乌蕨属、达边蕨属、香鳞始蕨属和鳞始蕨属(图6)。图6所示的5种形态数据中, 各项数值最大的都是乌蕨属。Spheno- meris和Nesolindsaea由于中国不产, 未获得其孢子囊材料, 仅从文献中得知Sphenomeris clavata的孢子囊囊蒴大小为200-250 μm, 环带细胞15-23个(Lehtonen et al., 2010)。而乌蕨属的囊蒴长度为228.7-252.6 μm, 环带细胞14.2-21.6个。因此从这2项形态指标来看Sphenomeris clavata更接近乌蕨属。 图6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_6.png<b>图6</b> 鳞始蕨科属间系统关系(<xref ref-type="bibr" rid="b14-1674-3466-52-3-322">Lehtonen et al., 2010</xref>)<br/><b>Figure 6</b> Phylogenetic relationships within Lindsaeaceae (<xref ref-type="bibr" rid="b14-1674-3466-52-3-322">Lehtonen et al., 2010</xref>) Figure 6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-3-322/img_6.png<b>图6</b> 鳞始蕨科属间系统关系(<xref ref-type="bibr" rid="b14-1674-3466-52-3-322">Lehtonen et al., 2010</xref>)<br/><b>Figure 6</b> Phylogenetic relationships within Lindsaeaceae (<xref ref-type="bibr" rid="b14-1674-3466-52-3-322">Lehtonen et al., 2010</xref>)
BierhorstDW (1971). Morphology of Vascular Plants. New York: The Macmillan Company. pp. 291-326.
[6]
BowerFO (1923). The Ferns (Filicales) I. London: Cambridge University Press. pp. 250-255.
[7]
BowerFO (1926). The Ferns (Filicales) II. London: Cambridge University Press.
[8]
BowerFO (1928). The Ferns (Filicales) III. London: Cambridge University Press. pp. 33-34; 67-68.
[9]
ChristenhuszMJ, ChaseMW (2014). Trends and concepts in fern classification.Ann Bot 113, 571-594. [本文引用: 1]
[10]
ChristensenC (1938). Filicinae. In: Verdoorn F, ed. Manual of Pteridology. The Hague: Martinus Nijhoff. pp. 522-550. [本文引用: 1]
[11]
HolttumRE (1947). A revised classification of leptosporangiate ferns.Bot J Linn Soc 53, 123-159. [本文引用: 1]
[12]
KramerKU (1957). A revision of the genus Lindsaea in the New World with notes on allied genera. Acta Bot Neerland 6, 97-281. [本文引用: 1]
[13]
KramerKU, GreenPS (1990). Pteridophytes and gymnosperms. In: Kubitzki K, ed. The Families and Genera of Vascular Plants. Berlin: Springer-Verlag. pp. 89-93. [本文引用: 2]
[14]
LehtonenS, TuomistoH, RouhanG, ChristenhuszMJ (2010). Phylogenetics and classification of the pantropical fern family Lindsaeaceae.Bot J Linn Soc 163, 305-359. [本文引用: 8]
[15]
LellingerDB (1985). A Field Manual of the Ferns and Fern-allies of the United States and Canada, Vol.3. Wash- ington: Smithsonian Institution Press. [本文引用: 1]
[16]
NoblinX, RojasNO, WestbrookJ, LlorensC, ArgentinaM, DumaisJ (2012). The fern sporangium: a unique ca- tapult.Science 335, 1322. [本文引用: 1]
[17]
PoppingaS, HaushahnT, WarnkeM, MasselterT, SpeckT (2015). Sporangium exposure and spore release in the peruvian maidenhair fern (Adiantum peruvianum, Pteridaceae). PLoS One 10, e0138495. [本文引用: 1]
[18]
PreslCB (1836). Tentamen pteridographiae, seu, genera filicacearum praesetim juxta venarum decursum et distributionem exposita. Prague: A. Haase. [本文引用: 1]
[19]
PryerKM, SchuettpelzE, WolfPG, SchneiderH, SmithAR, CranfillR (2004). Phylogeny and evolution of ferns (monilophytes) with a focus on the early leptosporangiate divergences.Am J Bot 91, 1582-1598. [本文引用: 2]
[20]
R CoreTeam (2015). R: a Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. URL . URL [本文引用: 1]
[21]
SmithAR, PryerKM, SchuettpelzE, KorallP, SchneiderH, WolfPG (2006). A classification for extant ferns.Taxon 55, 705-731. [本文引用: 1]
[22]
TryonR, TryonA (1982). Ferns and Allied Plants with Special Reference to Tropical America. New York: Springer- Verlag. [本文引用: 1]
[23]
WuZY, RavenP, HongDY (2013). Flora of China, Vol.2-3(Pteridophytes). Beijing and St. Louis: Science Press & Missouri Botanical Garden Press. pp. 139-146. [本文引用: 6]
石松类和蕨类植物研究进展: 兼论国产类群的科级分类系统 1 2008
... 鳞始蕨科(Lindsaeaceae)为水龙骨类(polypods)的基部类群(Pryer et al., 2004; Smith et al., 2006; 刘红梅等, 2008; Christenhusz and Chase, 2014), 起源于1.33亿年前(Pryer et al., 2004), 全世界约有200种, 主要分布在泛热带地区(Kramer, 1957; Wu et al., 2013).分子系统学兴起之前, 鳞始蕨科是一个庞杂而富有争议的类群.该科部分类群的系统位置早期被处理为骨碎补科(Davalliaceae)的成员(Presl, 1836; Christensen, 1938), 后来又被处理为碗蕨科(Dennstaedtiaceae)的种类(Holttum, 1947; Tryon and Tryon, 1982; Kramer and Green, 1990).在属的划分上, 鳞始蕨科曾被划分为多个属.最新研究结果表明, 该科有6属, 另有3属的系统关系还未得到确认(表1).其中中国产4属: 乌蕨属(Odontosoria)、香鳞始蕨属(Osmolindsaea)、达边蕨属(Tapeinidium)和鳞始蕨属(Lindsaea) (Wu et al., 2013).中国不产的2个属, Sphenomeris主要分布在新热带地区, Nesolindsaea主要分布在印度洋北部地区(塞舌尔和斯里兰卡) (Lehtonen et al., 2010).基于分子证据的鳞始蕨科的系统发育关系相对稳定, 但缺乏孢子囊形态的支持. ...


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}