Abstract: From 2014 to 2015, we investigated and compared the Hami and Wuqia populations of Gymnocarpos przewalskii in the wild, and used indices such as flowering amplitude, relative flowering intensity, synchrony index. The two populations flowered once in a year, late April to early May into flowering onset; the flowering phenology of the two populations was similar, and the plant has the characteristics of high flowering synchrony and a typical “mass flowering” pattern. At the individual level, there were significant differences both in the first flowering date and the end flowering date by population and year; the single flower longevity and mean longevities of individual flowers were similar at different years. Different populations differed: the flowering duration of a single flower of Wuqia was slightly longer than for the Hami population. We found a significant positive correlation between flower number and seed number, whereas the synchrony indices were negatively correlated with relative flowering intensity in different populations. The similarity in flowering amplitude and flowering synchrony in the two species and years of G. przewalskii may be explained by their phylogenetic relationships, whereas the differences may result from their evolutionary history in heterogeneous habitats (altitude, temperature, precipitation and light). These results can provide useful information for research on reproductive biology and conservation for the species.
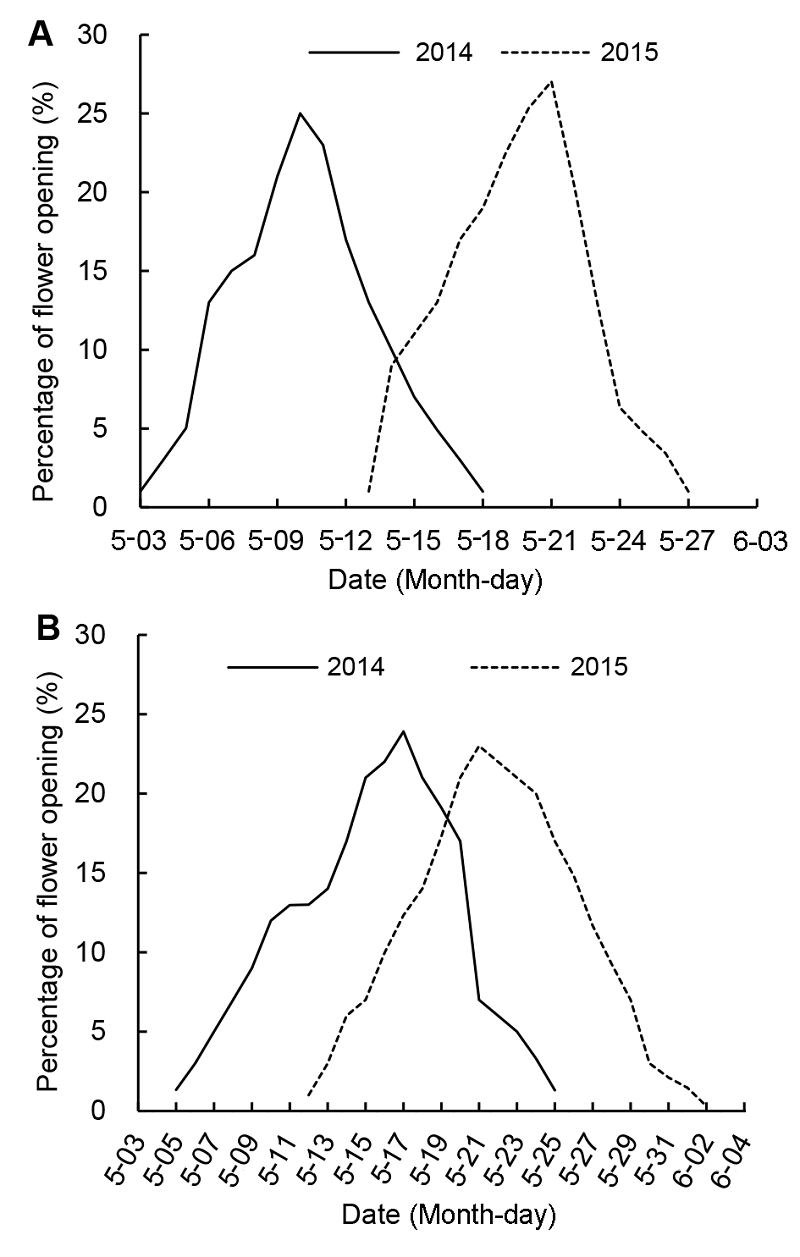
2 结果与讨论2.1 种群及个体水平的开花物候2014和2015年不同裸果木的开花物候如表2所示。在种群水平上, 2014和2015年哈密和乌恰裸果木种群每年均开1次花, 3月下旬至4月上旬开始长出新的叶片, 4月下旬到5月上旬进入初花期, 5月中旬进入盛花期, 5月下旬或6月上旬进入终花期; 哈密种群的始花期和终花期均早于乌恰种群。 在个体水平上, 哈密种群连续2年的始花期和终花期均早于乌恰种群, 但乌恰种群的开花持续时间比哈密种群长; 在不同年份间, 不同裸果木种群的开花振幅基本相同, 均呈单峰曲线, 当开花比例达到最大值后逐渐下降, 只是在2015年两种群的始花日期和开花高峰日期均比2014年略有推迟(图1)。哈密和乌恰种群个体的始花期(F=2.740, P<0.05; F=2.399, P<0.05)、盛花期(F=1.291, P<0.05; F=1.550, P< 0.05)、终花期(F=2.376, P<0.05; F=2.064, P<0.05)在年际间均存在显著差异; 而开花振幅(F=7.378, P>0.05; F=1.670, P>0.05)和开花同步指数(F=0.543, P>0.05; F=0.485, P>0.05)在年际间均差异不显著。2014和2015年, 裸果木个体始花期(F=24.347, P< 0.05; F=15.539, P<0.05)、盛花期(F=2.959, P<0.05; F=1.537, P<0.05)和终花期(F=3.110, P<0.05; F= 1.099, P<0.05)在种群间均差异显著, 而其开花同步指数(F=4.598, P>0.05; F=8.319, P>0.05)及开花振幅(F=2.983, P>0.05; F=7.559, P>0.05)在种群间不存在显著差异。哈密和乌恰裸果木种群在2014和2015年的平均开花同步性分别为0.833±0.011、0.831±0.004、0.819±0.027及0.813±0.026, 表现出较高的同步性特征, 呈现出一种“集中大量”的开花模式。由图2可知, 不同种群裸果木个体的开花强度 表2 Table 2 表2 表2 2014和2015年不同种群裸果木在种群及个体水平的开花物候 Table 2 Phenology of Gymnocarpos przewalskii at the level of the population and individual level in 2014 and 2015
Observation item
Population of Hami
Population of Wuqia
2014
2015
2014
2015
Population level
Onset
April 29
May 6
May 3
May 9
Date of peak flowering
May 9
May 17
May 23
May 30
End date
May 29
June 11
June 13
June 24
Duration (d)
30
36
41
46
Individual level
Number of flowers
303.4±63.7
317.65±55.36
232.3±33.10
234±20.66
Onset
4.55±1.82 (May 3)
12.00±0.86 (May 11)
6.95±0.19 (May 6)
13.50±1.47 (May 13)
Date of peak flowering
10.90±1.07
21.45±0.76
17.00±1.17
23.25±1.92
End date
16.70±1.13
28.10±1.17
24.95±1.76
33.60±2.04
Duration (d)
12.15±2.37
16.10±1.41
18.10±2.17
20.10±2.63
Range
3-19
11-30
6-30
19-39
RFI (%)
9.57±7.79
10.13±8.30
11.46±7.06
12.16±7.53
FSI
0.833±0.011
0.831±0.004
0.819±0.027
0.813±0.026
Data shown are means±SD, and range of values. RFI: Relative flowering intensity; FSI: Flowering synchrony index. 数据显示平均值±标准差和范围。RFI: 相对开花强度; FSI: 开花同步性指数。
表2 2014和2015年不同种群裸果木在种群及个体水平的开花物候 Table 2 Phenology of Gymnocarpos przewalskii at the level of the population and individual level in 2014 and 2015
2.3 开花物候参数及结籽数的相关性哈密和乌恰裸果木种群的开花物候参数(始花期、开花持续时间、开花同步性指数、相对开花强度、开花数)及结籽数之间的相关性分析结果显示(表4)。不同裸果木种群的开花数与结籽数均呈极显著正相关, 表明植株个体开花数越大, 结籽数越高; 开花同步性指数与相对开花强度间呈显著负相关; 哈密裸果木种群 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-487/img_2.png<b>图1</b> 不同裸果木种群在不同年份间个体的开花振幅曲线(A) 哈密种群; (B) 乌恰种群<br/><b>Figure 1</b> Flowering amplitude curves of <i>Gymnocarpos przewalskii</i> individual plant of two populations in different years(A) Hami population; (B) Wuqia population Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-487/img_2.png<b>图1</b> 不同裸果木种群在不同年份间个体的开花振幅曲线(A) 哈密种群; (B) 乌恰种群<br/><b>Figure 1</b> Flowering amplitude curves of <i>Gymnocarpos przewalskii</i> individual plant of two populations in different years(A) Hami population; (B) Wuqia population
图1 不同裸果木种群在不同年份间个体的开花振幅曲线(A) 哈密种群; (B) 乌恰种群 Figure 1 Flowering amplitude curves of Gymnocarpos przewalskii individual plant of two populations in different years(A) Hami population; (B) Wuqia population
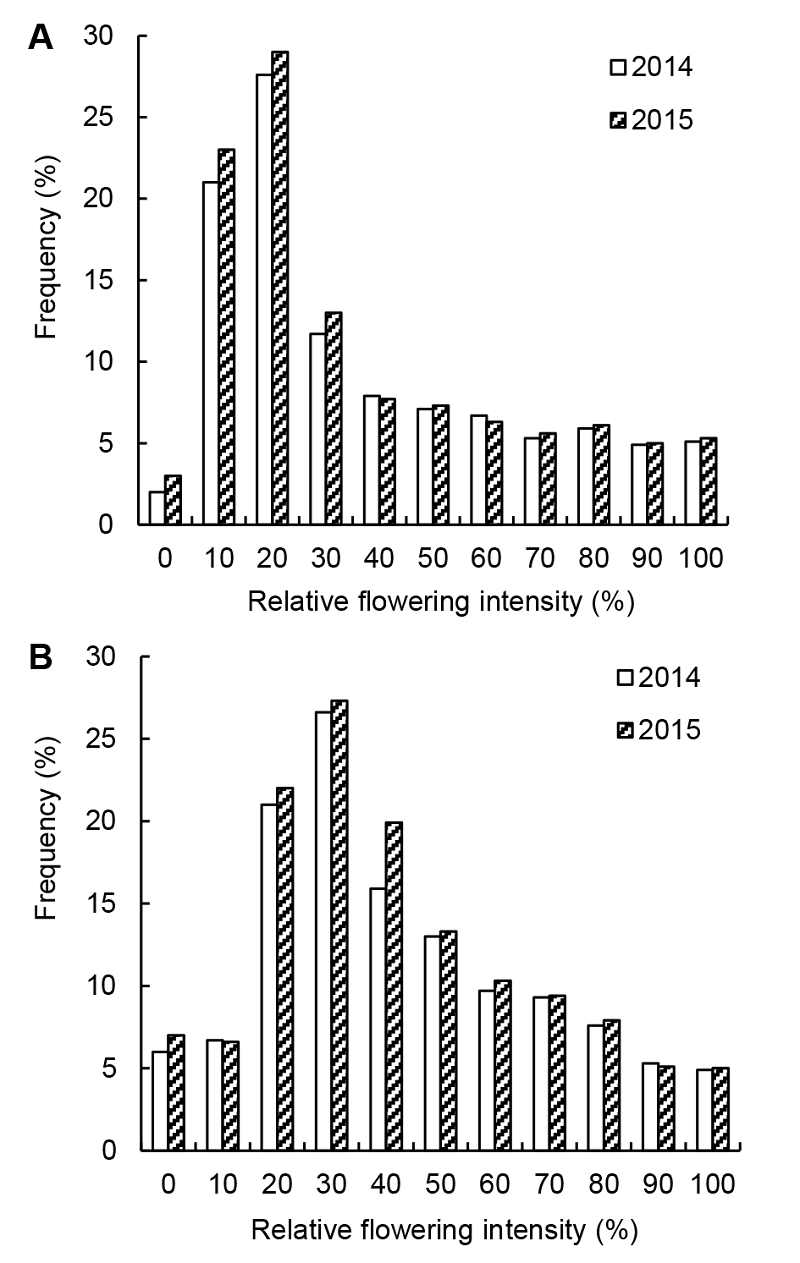
2.4 讨论2.4.1 开花物候特征及其影响因子 2014和2015年连续2年的开花物候观察表明, 裸果木个体水平的开花振幅在年际间和种群间均呈单峰曲线, 开花比例达最大值后逐渐降低, 只有1个开花高峰期表现出较高的开花同步性, 呈现出“集中开花”模式。相对开花强度影响植物花粉的转运模式(Dafni 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-487/img_3.png<b>图2</b> 不同裸果木种群在不同年份间个体的相对开花强度频率分布(A) 哈密种群; (B) 乌恰种群<br/><b>Figure 2</b> Frequency distributions of relative flowering intensity of <i>Gymnocarpos przewalskii</i> individual plants of two populations in different years(A) Hami population; (B) Wuqia population Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-487/img_3.png<b>图2</b> 不同裸果木种群在不同年份间个体的相对开花强度频率分布(A) 哈密种群; (B) 乌恰种群<br/><b>Figure 2</b> Frequency distributions of relative flowering intensity of <i>Gymnocarpos przewalskii</i> individual plants of two populations in different years(A) Hami population; (B) Wuqia population
图2 不同裸果木种群在不同年份间个体的相对开花强度频率分布(A) 哈密种群; (B) 乌恰种群 Figure 2 Frequency distributions of relative flowering intensity of Gymnocarpos przewalskii individual plants of two populations in different years(A) Hami population; (B) Wuqia population
et al., 2005), 是衡量植物花资源空间分布的指标(Buide et al., 2002)。裸果木相对开花强度较低, 但强度分布偏斜, 表明在两种群中存在相对开花强度较高的植株个体。上述特征与许多荒漠植物如蒙古沙冬青(Ammopiptanthus mongolicus)、新疆沙冬青(A. nanus)、准噶尔无叶豆(Eremosparton songoricum)、弯花黄芪(Astragalus flexus)、耳叶补血草(Limonium otolepis)、沙拐枣属(Calligonum)及银沙槐(Ammo- dendron argenteum)的研究结果相似(李新蓉等, 表3 Table 3 表3 表3 裸果木单花开花动态 Table 3 The flowering dynamic of Gymnocarpos przewalskii single flower
Stages
Calyx opening situation
Anthers changes in color
Relative position of stigma and anthers
Pre-dehiscence
Unopened
Yellow
Stamens lower than the pistil
Initial dehiscence
Calyx open
Yellow
Stamens lower than the pistil or flush
Full dehiscence
Calyx fully open and the maximum diameter
Yellow or orange
Stamens higher than the pistil or flush
Final dehiscence
After reaching the maximum diameter, the flower diameter began to decrease
Orange or fall off
Stamens higher than the pistil or flush
Withering
Flower diameter decreased and gradua- lly closed
Fall off
Stamens lower than the pistil or flush
表3 裸果木单花开花动态 Table 3 The flowering dynamic of Gymnocarpos przewalskii single flower
表4 Table 4 表4 表4 2015年不同种群裸果木开花物候参数及结籽数的相关性分析 Table 4 Pearson correlation coefficients of flowering phenology items and number of seeds of Gymnocarpos przewalskii in different populations of 2015
表4 2015年不同种群裸果木开花物候参数及结籽数的相关性分析 Table 4 Pearson correlation coefficients of flowering phenology items and number of seeds of Gymnocarpos przewalskii in different populations of 2015
2006; 李新蓉和谭敦炎, 2007; 马文宝等, 2008; 赵文忠, 2008; 黄云兰等, 2012; 康晓珊等, 2012; 李志成等, 2013)。同时, 形成这些特征的原因一方面是该物种为第三纪孑遗植物且种群间断分布, 另一方面与其种群内存在龄级差异较大的植株个体有关。植物的开花式样(指花在种群水平上表现出的特征, 即花在某一时间段开放的数量和在花序上的空间排列)在整个花期内不断变化, 种群开花早期和末期, 花的开放数量均比盛花期要少(Huang et al., 2006)。裸果木在始花期只有个别植株和花序上开了几朵花, 但盛花期则大量开花, 这可能是由于其主要生长在荒漠砾石戈壁及山前洪积扇戈壁等地区, 自然生境恶劣, 土壤贫瘠, 昼夜温差大且多风沙, 在没有充足传粉者的情况下, 植物通过大量开花的方式增加授粉成功率(Harder and Johnson, 2005)。对热带灌木植物Hybanthus prunifolius的研究表明, 增加花的开放数量能够提高对群居蜜蜂的吸引力, 从而增加传粉成功的可能性(Augspurger, 1980, 1981)。 哈密和乌恰裸果木种群个体的始花期、盛花期及终花期在年际间均存在显著差异; 个体间始花期、盛花期、终花期及开花振幅在种群间均差异显著, 而开花振幅和开花同步性指数在年际间均差异不显著, 其开花同步性指数在种群间也不存在显著差异。以上结果表明, 开花物候特征不仅受植物物种的地理发生和遗传特性的影响, 而且与其生境因子的变化(如纬度、海拔、光照、温度和降水等)也有密切的联系(Michèle et al., 2007)。分布于不同地区或同一地区不同海拔的同种植物开花物候存在一定差异, 这主要是由于生境中光照和温度等作用的结果(柴胜丰等, 2009; 任崇勇等, 2015)。由表1可知, 哈密种群的温度和日照连续2年均高于乌恰种群, 但降水量低于乌恰种群。在温度和光照相对较高的条件下, 哈密种群连续2年的始花期、盛花期、终花期及果实成熟期均比乌恰种群早10天左右。高海拔的植物比低海拔的植物有更长的花寿命(Primack, 1985)。乌恰种群海拔高2 130 m, 哈密种群海拔高1 323 m, 温度随海拔的升高而降低。乌恰种群的单花持续时间连续2年均长于哈密种群, 这可能与不同种群的海拔高度差异有关。我们通过前期的调查发现, 裸果木多生长在荒漠戈壁的干河沟或山前冲积扇上, 对土壤水分有一定要求, 乌恰种群的降水量连续2年均高于哈密种群, 其种群盖度和植株密度均大于哈密种群。2015年较低的气温使不同裸果木种群的花期均有所推迟, 其中哈密和乌恰种群始花日期2015年较2014年分别推迟7天和6天。此外, 哈密种群所在地风力较大(多为3-4级), 据我们观测, 许多花朵及花蕾在此环境条件下传粉受到影响, 进而直接影响坐果数。由此可看出, 不同种群间物候的差异主要受海拔、温度、降水、光照和风力的影响, 降水则可能是引起种群在年际间物候差异的重要原因。 表1 Table 1 表1 表1 哈密和乌恰2014和2015年1-7月主要气象因子 Table 1 Main meteorological factors at Hami and Wuqia from January to July of 2014 and 2015
Research area
Main meteorological factors
Year
Month
January
February
March
April
May
June
July
Hami
Mean temperature (°C)
2014
-12.5
-4.3
9.2
15.9
21
24.7
26.9
2015
-9.3
-4.5
5.8
14.4
19.9
24.3
27.6
Precipitation (mm)
2014
0
4.1
0
0
1.5
3.4
4.4
2015
0
0
0
0.6
3.5
26.6
4.9
Sunshine duration (h)
2014
226.1
237.6
311.3
329
366.9
329.1
378.2
2015
230.4
237.2
309.5
303.1
367.8
331.1
383.1
Wuqia
Mean temperature (°C)
2014
-5.2
-5.1
3.5
9.8
14.7
17.2
20.2
2015
-5.1
-2.2
4.9
10.5
15.3
17.5
22.8
Precipitation (mm)
2014
0.3
4
0
1.9
18.1
11.8
9.2
2015
1.4
9.5
0.5
0
2.7
13.7
1.3
Sunshine duration (h)
2014
169.6
180.8
244.2
248.7
282.3
348.7
364.9
2015
189.4
168.7
225.1
240
252.1
325.9
360.8
表1 哈密和乌恰2014和2015年1-7月主要气象因子 Table 1 Main meteorological factors at Hami and Wuqia from January to July of 2014 and 2015
AugspurgerCK (1980). Mass-flowering of a tropical shrub (Hybanthus prunifolius): influence on pollinator attraction and movement.Evolution 34, 475-488. [本文引用: 1]
[19]
AugspurgerCK (1981). Reproductive synchrony of a tropical shrub: experimental studies on effects of pollinators and seed predators on Hybanthus prunifolius (Violaceae).Eco- logy 62, 775-788. [本文引用: 1]
[20]
AugspurgerCK (1983). Phenology, flowering synchrony, and fruit set of six neotropical shrubs.Biotropical 15, 257-267. [本文引用: 1]
[21]
BawaKS, KangH, GrayumMH (2003). Relationships among time, frequency, and duration of flowering in tropical rain forest trees.Am J Bot 90, 877-887. [本文引用: 1]
[22]
BuideML, Diaz-PeromingoJA, GuitiánJ (2002). Flowering phenology and female reproductive success inSilene acutifolia Link ex Rohrb. Plant Ecol 163, 93-103. [本文引用: 1]
[23]
CarmerondF (1976). The inheritance of flowering time in Townsville Styllo (Stylosanthes humilis). Aust J Agr Res 27, 825-838. [本文引用: 1]
[24]
DafniA (1992).Pollination Ecology:a Practical Approach.Oxford: Oxford University Press. pp. 171-181. [本文引用: 1]
GuoH, YangH, MocklerTC, LinC (1998). Regulation of flowering time by Arabidopsis photoreceptors.Science 279, 1360-1363. [本文引用: 1]
[27]
HarderLD, JohnsonSD (2005). Adaptive plasticity of floral display size in animal-pollinated plants.P Roy Soc B-Biol Sci 272, 2615-2657. [本文引用: 1]
[28]
HerreraJ (1986). Flowering and fruiting phenology in the coastal shrublands of Doňana South Spain.Vegetatio 68, 91-98. [本文引用: 1]
[29]
HuangSQ, TangLL, SunJF, LuY (2006). Pollinator response to female and male floral display in a monoecious species and its implications for the evolution of floral dimorphism.New Phytol 171, 417-424. [本文引用: 1]
[30]
MaSM, ZhangML (2012). Phylogeography and conservation genetics of the relic Gymnocarpos przewalskii(Caryophyllaceae) restricted to northwestern China. Conserv Ge- net 13, 1531-1541. [本文引用: 1]
[31]
MaSM, ZhangML, SandersonSC (2012). Phylogeography of the rare Gymnocarpos przewalskii (Caryophyllaceae): indications of multiple glacial refugia in north-western China.Aust J Bot 60, 20-31. [本文引用: 1]
[32]
McIntoshME (2002). Flowering phenology and reproductive output in two sister species of Ferocactus (Cactaceae).Plant Ecol 159, 1-13.
[33]
MichèleT, GillianneB, AgnèsSL, MyriamB, AnneA (2007). Flowering phenology of Ulex europaeus: ecological consequences of variation within and among populations.Evol Ecol 21, 395-409. [本文引用: 1]
[34]
OllertonJ, LackA (1992). Flowering phenology: an example of relaxation of natural selection.Trends Ecol Evol 7, 274-276. [本文引用: 1]
[35]
PickeringCM (1995). Variation in flowering parameters within and among five species of Australian alpine Ran- unculus. Aust J Bot 43, 103-112. [本文引用: 1]
SutherlandS, DelphLF (1984). On the importance of male fitness in plants: patterns of fruit-set.Ecology 65, 1093-1104. [本文引用: 1]
[39]
WhiteLM (1995). Predicting flowering of 130 plant at 8 locations with temperature and day length. J Range Manage 48, 108-114. [本文引用: 1]
[40]
WidénB (1991). Environmental and genetic influences on phenology and plant size in a perennial herb,Senecio integrifolius. Can J Bot 69, 209-217. [本文引用: 1]

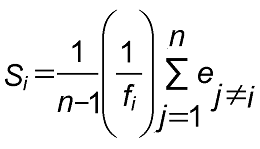

{kind=link}
{kind=link}
{kind=link}
{kind=link}