叶诺楠1, 沈娜娉2, 商天其1, 高洪娣3, 管杰然1, 伊力塔1,*, 1浙江农林大学林业与生物技术学院, 临安 3113002国家林业局华东林业调查规划设计院, 杭州 310019 3浙江省林业生态工程管理中心, 杭州 310020 YeNuonan1, ShenNaping2, ShangTianqi1, GaoHongdi3, GuanJieran1, YiLita1,*, 1School of Forestry and Biotechnology, Zhejiang Agriculture and Forestry University, Lin’an 311300, China;2The East China Institute of Forest Inventory and Planning of the State Forestry Administration, Hangzhou 310019, China3Ecological Management Center, Forestry Department of Zhejiang Province, Hangzhou 310020, China 引用本文 叶诺楠, 沈娜娉, 商天其, 高洪娣, 管杰然, 伊力塔. 浙江瑞安公益林群落结构及其与环境的相关性. , 2017, 52(4): 496-510
Abstract: With the construction of an ecological service forest (ESF) in Zhejiang province, the ESF has provided many ecological and social benefits to humans; thus, the number of studies about ESF has also increased. This study describes how to analyze the vegetation structure and the internal relationship between distribution patterns of vegetation and environment in 92 permanent sample plots of the ESF in Rui’an city, Zhejiang province. The vegetation structure and the internal relationship between the distribution patterns of vegetation and environment were studied by using canonical correspondence analysis (CCA), two-way indicator species analysis (TWINSPAN), biodiversity and interspecies association. (1) The elevation, slop aspect and slop position were main environment factors in the community distribution. (2) Combined with the results of CCA, the 92 plots could be divided into 13 groups by TWINSPAN, and the associations of Pinus massoniana, Cunninghamia lanceolata and Cryptomeria fortunei were the most constructive species. (3) The diversity of vegetation was high, and the groups with P. massonian, C. lanceolata and C. fortunei as dominant species had higher diversity. The species diversities of different layers in the community were in the order of shrub layer>tree layer>herb layer. (4) The stability of the community was not high (less than 2), but the tree layer was relatively stable because the lower ratios of positive and negative association theoretically implied the instable vegetation structure. Therefore, the internal relationship between the distribution patterns of vegetation and environment and the dominant species association could be explained by a combination of all methods; these methods could provide a scientific foundation for the classification management of a regional ESF.
Key words:ecological service forest ; canonical correspondence analysis ; two-way indicator species analysis ; biodiversity ; interspecific association
3 结果与讨论3.1 物种重要值依据重要值公式, 计算92个样地的物种群落重要值, 得到重要值大于0.1的物种114种, 其中乔木层35种, 灌木层59种, 草本层20种。结果如表1所示。 表1 Table 1 表1 表1 浙江瑞安公益林92个样地中主要乔木、灌木和草本植物的重要值 Table 1 Importance values of the main tree, shrub and herb on 92 plots in ecological service forest of Rui’an city in Zhejiang province
Layer
Number
Species
Importance value
Layer
Number
Species
Importance value
Tree
1
Pinus massoniana
30.1251
48
Syzygium buxifolium
2.6616
2
Cunninghamia lanceolata
19.1471
49
Camellia oleifera
2.6044
3
Cryptomeria fortunei
11.7255
50
Pinus massoniana
2.4924
4
Phyllostachys heterocycla
10.4433
51
Camellia sinensis
2.3143
5
Schima superba
6.4891
52
Pleioblastus amarus
2.1880
6
Lithocarpus glaber
2.7277
53
Smilax china
2.1092
7
Liquidambar formosana
2.5786
54
Smilax sieboldii
1.9226
8
Cyclobalanopsis glauca
2.1817
55
Mallotus japonicus
1.8279
9
Castanopsis sclerophylla
2.0855
56
Liquidambar formosana
1.7230
10
Castanopsis eyrei
1.5263
57
Litsea cubeba
1.6368
11
Symplocos sumuntia
1.4795
58
Schima superba
1.4864
12
Myrica rubra
1.0891
59
Rubus phoenicolasius
1.1355
13
Eurya japonica
0.8298
60
Cyclobalanopsis glauca
0.9140
14
Pinus taiwanensis
0.8262
61
Symplocos stellaris
0.8884
15
Machilus thunbergii
0.7395
62
Rubus buergeri
0.7625
16
Ilex chinensis
0.6225
63
Camellia japonica
0.7053
17
Toxicodendron succedaneum
0.4550
64
Castanopsis sclerophylla
0.6913
18
Cinnamomum camphora
0.4451
65
Toxicodendron succedaneum
0.6529
19
Litsea cubeba
0.3609
66
Trachelospermum jasminoides
0.5940
20
Vernicia montana
0.3327
67
Adinandra millettii
0.5748
21
Ulmus pumila
0.3149
68
Dendrocalamopsis oldhami
0.5642
22
Camellia japonica
0.2716
69
Cryptomeria fortunei
0.5083
23
Elaeocarpus decipiens
0.2526
70
Lindera aggregata
0.4966
24
Loropetalum chinense
0.2059
71
Rubus xanthocarpus
0.4675
25
Ficus microcarpa
0.2039
72
Quercus fabri
0.4185
26
Mallotus japonicus
0.1838
73
Myrica rubra
0.4185
27
Cyclobalanopsis myrsinifolia
0.1728
74
Lespedeza bicolor
0.4092
28
Eucalyptus camaldulensis
0.1678
75
Pinus taiwanensis
0.4092
29
Diospyros kaki
0.1445
76
Rosa laevigata
0.4092
30
Rhus chinensis
0.1296
77
Rhus chinensis
0.3742
31
Albizia kalkora
0.1260
78
Neocinnamomum chekiangense
0.3602
32
Paulownia tomentosa
0.1199
79
Osmanthus fragrans
0.3509
33
Machilus leptophylla
0.1132
80
Glochidion puberum
0.3101
34
Adinandra millettii
0.1109
81
Ulmus pumila
0.3019
35
Quercus fabri
0.1099
82
Gardenia jasminoides
0.2985
Shrub
36
Eurya japonica
13.5139
83
Vernicia fordii
0.2845
37
Rhododendron simsii
7.6406
84
Toxicodendron sylvestre
0.2553
38
Loropetalum chinense
5.6483
85
Eucalyptus camaldulensis
0.2437
39
Phyllostachys heteroclada
5.5905
86
Diospyros kaki
0.2437
40
Cunninghamia lanceolata
4.3673
87
Dalbergia hupeana
0.2145
41
Symplocos sumuntia
4.3183
88
Photinia serratifolia
0.2029
42
Brachystachyum densiflorum
4.3177
89
Melia azedarach
0.1655
43
Castanopsis eyrei
3.6525
90
Phyllostachys edulis
0.1655
44
Vaccinium bracteatum
3.3764
91
Pinus elliottii
0.1655
45
Lithocarpus glaber
3.1092
92
Machilus leptophylla
0.1364
46
Ilex chinensis
3.0220
93
Lindera glauca
0.1073
47
Smilax corbularia
2.7980
94
Phoebe sheareri
0.1073
Herb
95
Osmunda japonica
60.1798
105
Boehmeria nivea
0.6881
96
Dryopteris fuscipes
6.8201
106
Dicranopteris pedata
0.5281
97
Miscanthus floridulus
6.7660
107
Juncus setchuensis
0.3454
98
Cyperus difformis
6.5565
108
Onychium japonicum
0.2944
99
Imperata cylindrica
4.9332
109
Pteris dispar
0.2816
100
Miscanthus sinensis
4.0732
110
Carex brunnea
0.1684
101
Polytrichum sp.
3.7467
111
Woodwardia japonica
0.1472
102
Pueraria montana
1.7609
112
Boehmeria spicata
0.1472
103
Diplopterygium glaucum
1.4828
113
Yucca gloriosa
0.1387
104
Chrysanthemum indicum
0.8028
114
Lygodium japonicum
0.1387
表1 浙江瑞安公益林92个样地中主要乔木、灌木和草本植物的重要值 Table 1 Importance values of the main tree, shrub and herb on 92 plots in ecological service forest of Rui’an city in Zhejiang province
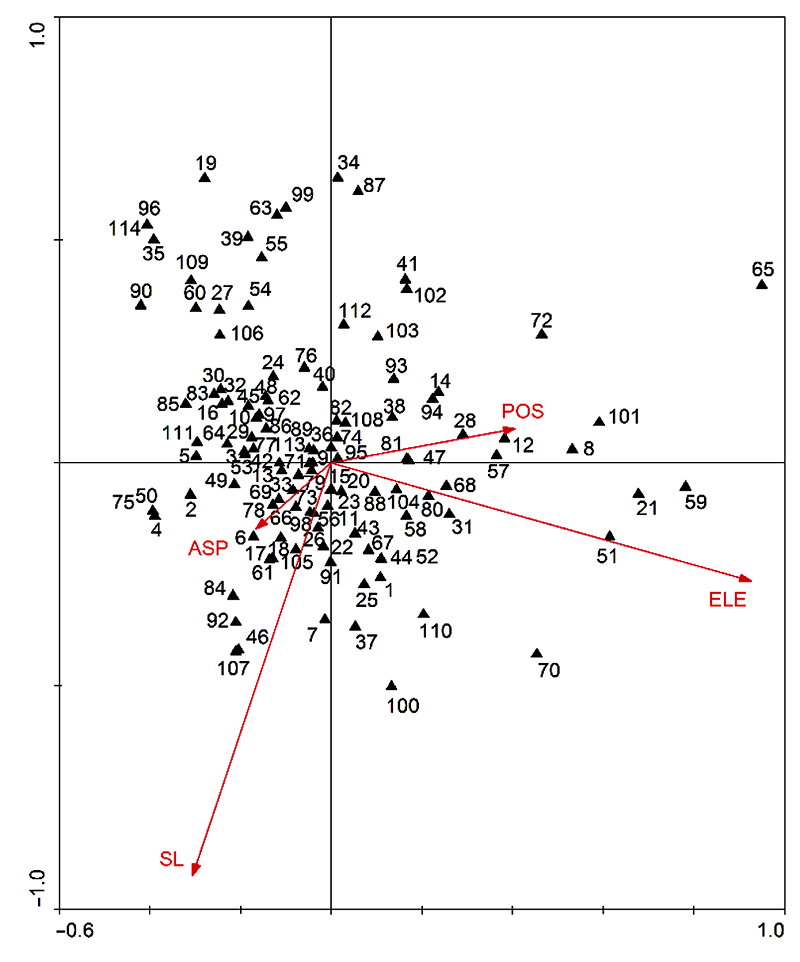
3.2 种-环境的CCA排序运用Canoco 5.0软件对研究区的114个优势物种进行典范对应分析, 得到排序轴与地形因子的关系以及各排序轴的特征值(表2)。其中第1轴和第2轴与地形因子呈显著相关(P<0.001), 特征值分别为0.290 0和0.198 0, 物种-环境相关性分别为0.820 0和0.741 0, 包含了69.1%的生态信息。故本研究采用第1轴和第2轴的信息对研究样地的环境及样地中物种与生长环境之间的关系分别进行分析。 表2 Table 2 表2 表2 浙江瑞安公益林92个样地典范对应分析(CCA)排序前4轴系数与环境因子的关系 Table 2 Coefficients between the first 4 axes in canonical correspondence analysis (CCA) ordination and environment factors of 92 plots in ecological service forest of Rui’an city in Zhejiang province
Environmental factors
Axis 1
Axis 2
Axis 3
Axis 4
Elevation
0.7595*
-0.1964
0.1744
0.0626
Slope
-0.2506
-0.6853*
-0.1253
0.0902
Aspect
-0.1357
-0.1096
0.6827*
0.0054
Position
0.3322
0.0569
0.0666
0.5886*
Eigenvalues
0.2900
0.1980
0.1110
0.1070
Species-environment correlations
0.8200
0.7410
0.7000
0.6500
Cumulative percentage variance of species-environment relation (%)
41.00
69.10
84.40
100.00
* P<0.001
表2 浙江瑞安公益林92个样地典范对应分析(CCA)排序前4轴系数与环境因子的关系 Table 2 Coefficients between the first 4 axes in canonical correspondence analysis (CCA) ordination and environment factors of 92 plots in ecological service forest of Rui’an city in Zhejiang province
在排序图(图1)中, 每个箭头都指示一种地形因子, 箭头与原点连线长度代表该地形因子对物种分布的影响大小, 连线越长, 影响程度越大; 箭头与排序轴所成的夹角大小则显示出该地形因子与排序轴之间的相关性, 角度越小, 相关性越大(郭晋平和张芸香, 2003; 郭逍宇等, 2004; 贾晓妮等, 2007)。综合表2和图1可知, 海拔是与第1轴相关性最高的地形因子, 呈正相关, 相关系数为0.759 5, 其次是坡位和坡度; 与第2轴相关性最高的地形因子是坡度, 呈负相关, 相关系数为-0.685 3, 其次是海拔和坡向。 图1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_7.png<b>图1</b> 浙江瑞安公益林114个优势种的CCA排序^物种编号同<xref ref-type="table" rid="T1-1674-3466-52-4-496">表1</xref>。ELE: 高度; SL: 坡度; ASP: 方向; POS: 坡位<br/><b>Figure 1</b> CCA ordination of 114 dominant species in ecological service forest of Rui’an city in Zhejiang province^Species number see <xref ref-type="table" rid="T1-1674-3466-52-4-496">Table 1</xref>. ELE: Elevation; SL: Slope; ASP: Aspect; POS: Position Figure 1https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_7.png<b>图1</b> 浙江瑞安公益林114个优势种的CCA排序^物种编号同<xref ref-type="table" rid="T1-1674-3466-52-4-496">表1</xref>。ELE: 高度; SL: 坡度; ASP: 方向; POS: 坡位<br/><b>Figure 1</b> CCA ordination of 114 dominant species in ecological service forest of Rui’an city in Zhejiang province^Species number see <xref ref-type="table" rid="T1-1674-3466-52-4-496">Table 1</xref>. ELE: Elevation; SL: Slope; ASP: Aspect; POS: Position
图1 浙江瑞安公益林114个优势种的CCA排序^物种编号同表1。ELE: 高度; SL: 坡度; ASP: 方向; POS: 坡位 Figure 1 CCA ordination of 114 dominant species in ecological service forest of Rui’an city in Zhejiang province^Species number see Table 1. ELE: Elevation; SL: Slope; ASP: Aspect; POS: Position
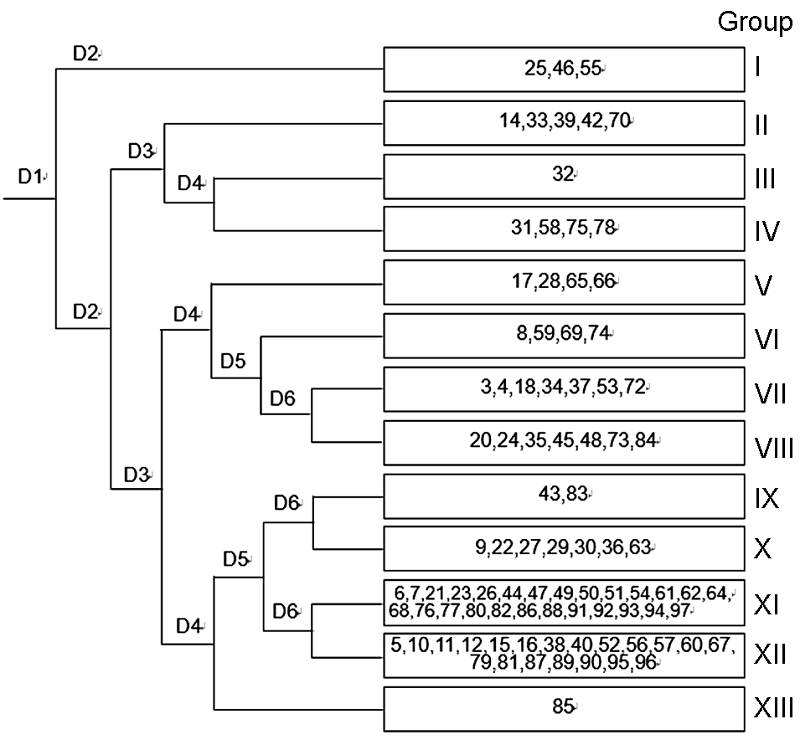
3.3 TWINSPAN分类我们对研究区92个样方进行TWINSPAN等级分类, 并将92个样方划分为13个群丛(图2)。 图2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_8.png<b>图2</b> 浙江瑞安公益林92个样方TWINSPAN分类等级^共调查97个样地, 但样地1、2、19、41和71数据缺失。<br/><b>Figure 2</b> Dendrogram of TWINSPAN classification of 92 plots in ecological service forest of Rui’an city in Zhejiang province^Ninety-seven plots were surveyed, and the data of the plot 1, 2, 19, 41, and 71 were missing. Figure 2https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_8.png<b>图2</b> 浙江瑞安公益林92个样方TWINSPAN分类等级^共调查97个样地, 但样地1、2、19、41和71数据缺失。<br/><b>Figure 2</b> Dendrogram of TWINSPAN classification of 92 plots in ecological service forest of Rui’an city in Zhejiang province^Ninety-seven plots were surveyed, and the data of the plot 1, 2, 19, 41, and 71 were missing.
图2 浙江瑞安公益林92个样方TWINSPAN分类等级^共调查97个样地, 但样地1、2、19、41和71数据缺失。 Figure 2 Dendrogram of TWINSPAN classification of 92 plots in ecological service forest of Rui’an city in Zhejiang province^Ninety-seven plots were surveyed, and the data of the plot 1, 2, 19, 41, and 71 were missing.
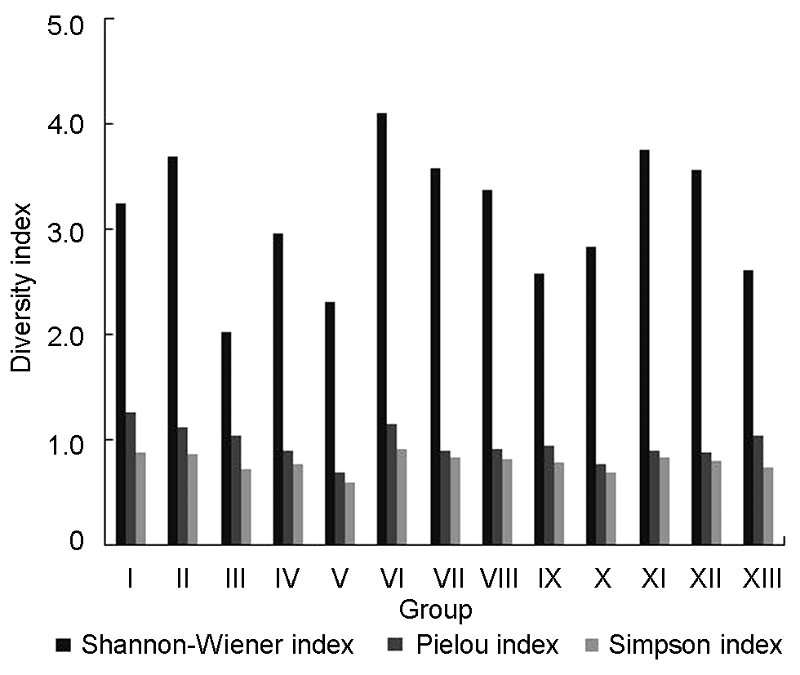
3.4 群落的物种多样性3.4.1 不同群落类型的物种多样性 将92个样地进行TWINSPAN等级分类得到13类群丛, 再分别进行物种多样性指数测算, 各群丛的群落物种多样性指数分布如图3所示。从图3可以看出, 这13个群丛的Shannon-Wiener指数(H)、Simpson指数(D)和Pielou均匀度指数(J)在各个群落中所呈现出的规律基本一致。群丛XI由于其涵盖的样地数最多而拥有最多的物种数(64种), 群丛III则因其只包含1个样地而物种数最少(7种)。群丛I的均匀度最高(1.2600), 群丛V的均匀度最低(0.6867); 群丛II、VI、VII和XI的多样性较高(这4个群丛的Simpson/Shannon-Wiener指数分别为4.100/0.912 1、3.755 1/0.841 0、3.688 1/0.872 3和3.578 0/0.841 0), 这4个群丛均分布在海拔200- 800 m的范围内, 所处地形为坡度缓和的上坡或中上坡, 拥有较好的水分、温度及光照条件, 利于植物生长。群丛III和V物种多样性指数最低(Simpson/Shan- non-Wiener指数分别为2.312 4/0.591 4和2.025 4/ 0.718 9)。群丛V是毛竹林, 林下灌木和草本数量少, 物种单一; 群丛III只涵盖1个样地, 分布海拔1 000 m, 虽然坡度缓和, 但高海拔和下坡导致其处于不良光热条件下, 物种多样性指数较低。 图3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_9.png<b>图3</b> 浙江瑞安公益林13个群丛的物种多样性指数<br/><b>Figure 3</b> 13 groups of species diversity index of communi- ties in ecological service forest of Rui’an city in Zhejiang pro- vince Figure 3https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_9.png<b>图3</b> 浙江瑞安公益林13个群丛的物种多样性指数<br/><b>Figure 3</b> 13 groups of species diversity index of communi- ties in ecological service forest of Rui’an city in Zhejiang pro- vince
图3 浙江瑞安公益林13个群丛的物种多样性指数 Figure 3 13 groups of species diversity index of communi- ties in ecological service forest of Rui’an city in Zhejiang pro- vince
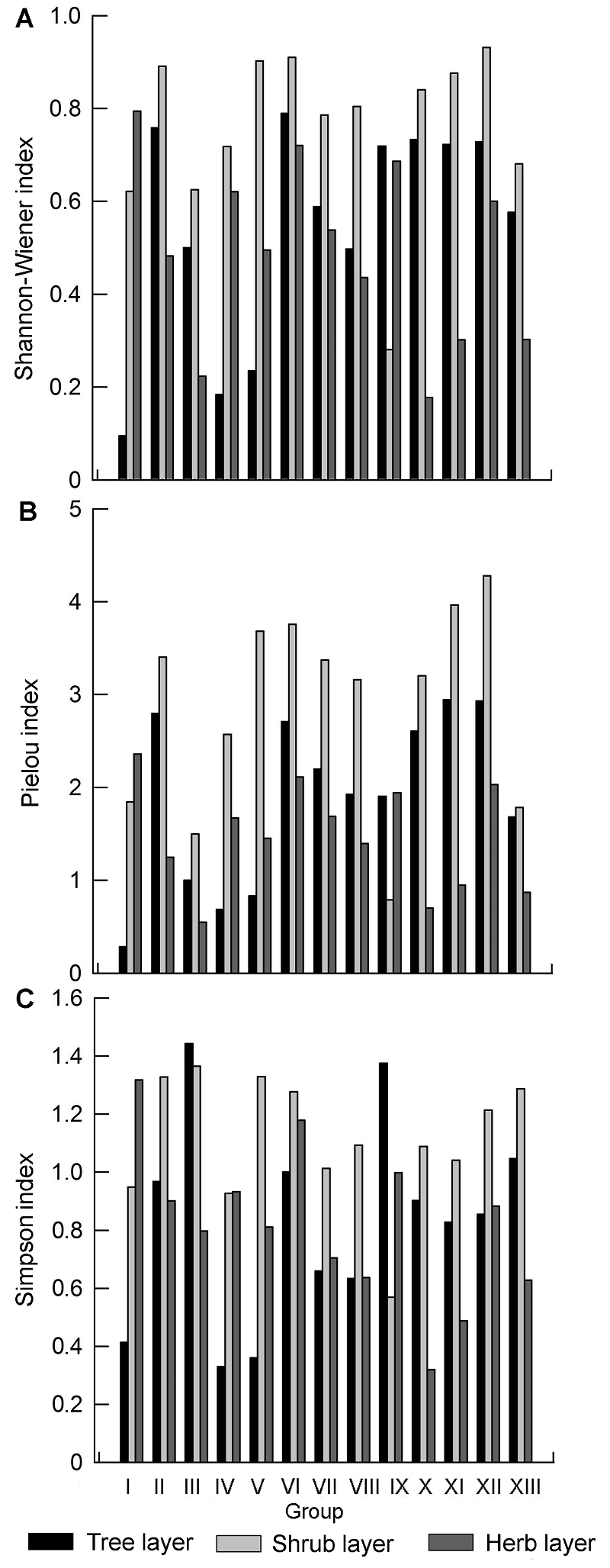
3.4.2 群落层次间物种多样性 通过对13类群丛的垂直层次之间的3个物种多样性指数进行差异性分析, 结果(图4; 表3)表明, 乔木层及灌木层的Simpson指数、Pielou均匀度指数和Shan- non-Wiener指数均存在显著差异(P=0.017, P=0.02, P=0.022, P值均在0.01-0.05之间); 灌木层及草本层的Simpson指数、Pielou均匀度指数和Shannon- Wiener指数均存在极显著差异(P=0.001, P=0.000, P=0.005, P值均小于0.01); 乔木层及草本层的Simp- son指数、Pielou均匀度指数和Shannon-Wiener指数差异均不显著(P=0.506, P=0.172, P=0.8, P值均大于0.05); 3个垂直层次的物种多样性总体规律为: 灌木层>乔木层>草本层。以上结果说明, 研究区内灌木层物种种类较多且盖度大, 阻碍了草本的生长、传播以及繁殖; 乔木层的林分组成比较单一, 多样性较低, 基本上是以马尾松、杉木和柳杉为主要优势种的针叶树种, 其在乔木层的竞争力较强, 但灌木层的物种数量和复杂性均优于乔木层, 故灌木层具有较高的生物多样性指数。 图4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_10.png<b>图4</b> 浙江瑞安公益林3个垂直层次的Shannon-Wiener (A)、Pielou (B)和Simpson (C)指数<br/><b>Figure 4</b> Shannon-Wiener (A), Pielou (B), and Simpson (C) index of the 3 vertical levels in ecological service forest of Rui’an city in Zhejiang province Figure 4https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_10.png<b>图4</b> 浙江瑞安公益林3个垂直层次的Shannon-Wiener (A)、Pielou (B)和Simpson (C)指数<br/><b>Figure 4</b> Shannon-Wiener (A), Pielou (B), and Simpson (C) index of the 3 vertical levels in ecological service forest of Rui’an city in Zhejiang province
图4 浙江瑞安公益林3个垂直层次的Shannon-Wiener (A)、Pielou (B)和Simpson (C)指数 Figure 4 Shannon-Wiener (A), Pielou (B), and Simpson (C) index of the 3 vertical levels in ecological service forest of Rui’an city in Zhejiang province
表3 Table 3 表3 表3 浙江瑞安公益林乔木层、灌木层和草本层的差异分析 Table 3 Difference analysis of tree layer, shrub layer and herb layer in ecological service forest of Rui’an city in Zhejiang pro- vince
Index
Shannon-Wiener
Simpson
Pielou
Layer
Tree
Shrub
Herb
Tree
Shrub
Herb
Tree
Shrub
Herb
Mean
1.885
2.871
1.460
0.548
0.759
0.491
0.832
1.114
0.815
SE
0.257
0.298
0.159
0.066
0.050
0.054
0.098
0.062
0.075
表3 浙江瑞安公益林乔木层、灌木层和草本层的差异分析 Table 3 Difference analysis of tree layer, shrub layer and herb layer in ecological service forest of Rui’an city in Zhejiang pro- vince
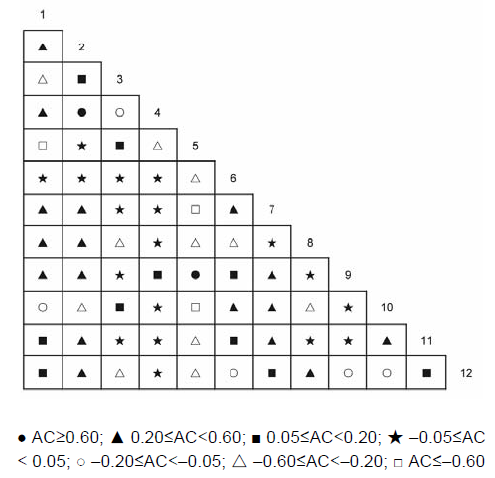
3.5 群落不同层次的种间联结分析经χ2检验得到乔木层植物优势种之间的关系(表4; 图5)。结果显示, 正关联种对有44对, 占总对数的66.7%; 负关联种对有22对, 占总对数的33.3%; 正负关联比为2; 显著率(包括极显著)为18.2%。显著正相关的种对有: 枫香树对苦槠, 枫香树对山矾, 苦槠对杨梅, 苦槠对青冈, 木荷对石栎, 木荷对青冈, 青冈对石栎, 青冈对甜槠, 青冈对杨梅, 山矾对杨梅, 石栎对甜槠。显著负相关的种对有: 马尾松对木荷。 表4 Table 4 表4 表4 浙江瑞安公益林13个生态关联乔木层的卡方检验及联结系数(AC)和共同出现百分率(PC) Table 4 Chi-square test, association coefficient (AC) and percentage co-occurrence (PC) in tree layer of 13 ecological associations in ecological service forest of Rui’an city in Zhejiang province
Test methods
Positive correlations
Negative correlations
None correlations
χ2
P<0.01
0.01<P<0.05
P>0.05
P<0.01
0.01<P<0.05
P>0.05
ad=bc
χ2-test
7
4
33
0
1
21
0
AC
AC≥0.60
0.20≤AC<0.60
0.05≤AC<0.20
AC≤-0.60
-0.60≤AC<-0.20
-0.20≤AC<-0.05
-0.05≤AC<0.05
AC values
2
17
10
3
11
5
18
PC
PC≥0.7
0.4≤PC<0.7
0.2≤PC<0.4
-
-
-
0≤PC<0.2
PC values
0
4
30
-
-
-
10
- 表示无数据。- indicate no data.
表4 浙江瑞安公益林13个生态关联乔木层的卡方检验及联结系数(AC)和共同出现百分率(PC) Table 4 Chi-square test, association coefficient (AC) and percentage co-occurrence (PC) in tree layer of 13 ecological associations in ecological service forest of Rui’an city in Zhejiang province
图5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_11.png<b>图5</b> 浙江瑞安公益林乔木层优势种的联结系数(AC)<br/><b>Figure 5</b> Association coefficient (AC) of dominant species in tree layer in ecological service forest of Rui’an city in Zhejiang province Figure 5https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_11.png<b>图5</b> 浙江瑞安公益林乔木层优势种的联结系数(AC)<br/><b>Figure 5</b> Association coefficient (AC) of dominant species in tree layer in ecological service forest of Rui’an city in Zhejiang province
图5 浙江瑞安公益林乔木层优势种的联结系数(AC) Figure 5 Association coefficient (AC) of dominant species in tree layer in ecological service forest of Rui’an city in Zhejiang province
灌木层植物优势种之间的关系(表5; 图6): 正关联种对有115对, 占总对数的41.7%; 负关联种对有141对, 占总对数的51.1%; 无关联种对3对; 正负关联比为0.816; 显著率(包括极显著)为3.99%。显著正相关的种对有: 茶对赤楠, 赤楠对乌饭树, 石栎对光叶菝葜(S. corbularia), 石栎对甜槠, 石栎对油茶, 檵木对油桐, 檵木对光叶菝葜, 山矾对乌饭树, 光叶菝葜对竹蕨, 杜鹃对柃木, 柃木对杉木。 表5 Table 5 表5 表5 浙江瑞安公益林13个生态关联灌木层的卡方检验及联结系数(AC)和共同出现百分率(PC) Table 5 Chi-square test, association coefficient (AC) and percentage co-occurrence (PC) in shrub layer of 13 ecological associations in ecological service forest of Rui’an city in Zhejiang province
Test methods
Positive correlations
Negative correlations
None correlations
χ2
P<0.01
0.01<P<0.05
P>0.05
P<0.01
0.01<P<0.05
P>0.05
ad=bc
χ2-test
4
4
104
0
0
141
20
AC
AC≥0.60
0.20≤AC<0.60
0.05≤AC<0.20
AC≤-0.60
-0.60≤AC<-0.20
-0.20≤AC<-0.05
-0.05≤AC<0.05
AC values
0
22
44
60
46
29
55
PC
PC≥0.7
0.4≤PC<0.7
0.2≤PC<0.4
-
-
-
0≤PC<0.2
PC values
0
1
22
-
-
-
92
- 表示无数据。- indicate no data.
表5 浙江瑞安公益林13个生态关联灌木层的卡方检验及联结系数(AC)和共同出现百分率(PC) Table 5 Chi-square test, association coefficient (AC) and percentage co-occurrence (PC) in shrub layer of 13 ecological associations in ecological service forest of Rui’an city in Zhejiang province
图6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_12.png<b>图6</b> 浙江瑞安公益林灌木层优势种的联结系数(AC)<br/><b>Figure 6</b> Association coefficient (AC) of dominant species in shrub layer in ecological service forest of Rui’an city in Zhejiang province Figure 6https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_12.png<b>图6</b> 浙江瑞安公益林灌木层优势种的联结系数(AC)<br/><b>Figure 6</b> Association coefficient (AC) of dominant species in shrub layer in ecological service forest of Rui’an city in Zhejiang province
图6 浙江瑞安公益林灌木层优势种的联结系数(AC) Figure 6 Association coefficient (AC) of dominant species in shrub layer in ecological service forest of Rui’an city in Zhejiang province
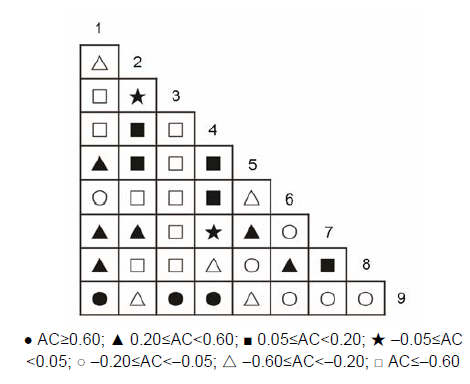
草本层植物优势种之间的关系(表6; 图7): 正关联种对有15对, 占总对数的41.7%; 负关联种对有21对, 占总对数的58.3%; 正负关联比为0.714; 显著率(包括极显著)为8.33%。显著正相关的种对有: 黑足鳞毛蕨对金发藓, 异型莎草对金发藓, 白茅对五节芒。 表6 Table 6 表6 表6 浙江瑞安公益林13个生态关联草本层的卡方检验及联结系数(AC)和共同出现百分率(PC) Table 6 Chi-square test, association coefficient (AC) and percentage co-occurrence (PC) in herb layer of 13 ecological associations in ecological service forest of Rui’an city in Zhejiang province
Test methods
Positive correlations
Negative correlations
None correlations
χ2
P<0.01
0.01<P<0.05
P>0.05
P<0.01
0.01<P<0.05
P>0.05
ad=bc
χ2-test
2
1
12
0
0
21
0
AC
AC≥0.60
0.20≤AC<0.60
0.05≤AC<0.20
AC≤-0.60
-0.60≤AC<-0.20
-0.20≤AC<-0.05
-0.05≤AC<0.05
AC values
3
6
5
8
4
6
2
PC
PC≥0.7
0.4≤PC<0.7
0.2≤PC<0.4
-
-
-
0≤PC<0.2
PC values
0
0
3
-
-
-
12
- 表示无数据。- indicate no data.
表6 浙江瑞安公益林13个生态关联草本层的卡方检验及联结系数(AC)和共同出现百分率(PC) Table 6 Chi-square test, association coefficient (AC) and percentage co-occurrence (PC) in herb layer of 13 ecological associations in ecological service forest of Rui’an city in Zhejiang province
图7https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_13.png<b>图7</b> 浙江瑞安公益林草本层优势种的联结系数(AC)<br/><b>Figure 7</b> Association coefficient (AC) of dominant species of herb layer in ecological service forest of Rui’an city in Zhejiang province Figure 7https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-4-496/img_13.png<b>图7</b> 浙江瑞安公益林草本层优势种的联结系数(AC)<br/><b>Figure 7</b> Association coefficient (AC) of dominant species of herb layer in ecological service forest of Rui’an city in Zhejiang province
图7 浙江瑞安公益林草本层优势种的联结系数(AC) Figure 7 Association coefficient (AC) of dominant species of herb layer in ecological service forest of Rui’an city in Zhejiang province
BrianJL, KasardaB (1977). Contemporary Urban Ecology. New York: Macmillan. pp. 95-113. [本文引用: 1]
[43]
ClarkDB, PalmerMW, ClarkDA (1999). Edaphic factors and the landscape—scale distributions of tropical rain forest trees.Ecology 80, 2662-2675. [本文引用: 1]
[44]
FormanRTT (1995). Land Mosaics: the Ecology of Lands- capes and Regions. Cambridge: Cambridge University Press. pp. 133-142. [本文引用: 1]
[45]
HenriquesRPB, HayJDV (1998). The plant communities of a foredune in southeastern Brazil.Can J Bot 76, 1323-1330. [本文引用: 1]
[46]
HurlbertSH (1969). A coefficient of interspecific association.Ecology 50, 1-9. [本文引用: 1]
[47]
PeetRK (1974). The measurement of species diversity. Annu Rev Ecol System 5, 285-307. [本文引用: 1]
[48]
PielouEC (1975). Ecology Diversity. New York: J. Wiley and Sons. pp. 62-82. [本文引用: 1]
[49]
RadJE, MantheyM, MatajiA (2009). Comparison of plant species diversity with different plant communities in deci- duous forests.Inter J Environ Sci Technol 6, 389-394. [本文引用: 1]
[50]
RoutledgeRD (1979). Diversity indices: which ones are ad- missible?J Theor Biol 76, 503-515. [本文引用: 1]
[51]
SwansonFJ, KratzTK, CaineN, WoodmanseeRG (1988). Landform effects on ecosystem patterns and processes: geomorphic features of the earth’s surface regulate the distribution of organisms and processes. BioScience 38(2), 92-98. [本文引用: 1]
[52]
WilseyBJ, PotvinC (2000). Biodiversity and ecosystem functioning: importance of species evenness in an old fie- ld.Ecology 81, 887-892. [本文引用: 1]
[53]
WoodwardFI, MckeeIF (1991). Vegetation and climate.Environ Inter 17, 535-546. [本文引用: 1]
四川龙门山西北部植被分布与地形因子的相关性 1 2006
... 环境因子影响植物的分布、生长以及群落的演替方向, 因此拥有不同环境条件的区域能够指示不同群落类型(商天其等, 2015).以往的研究表明, 在大尺度区域中, 气候条件是决定植物类型或生活型分布的主要因素(Woodward and Mckoo, 1991), 而对于群落这类中小尺度区域, 地形和土壤条件是影响物种分布的主要原因(Henriques and Hay, 1998; Clark et al., 1999; 沈泽昊和张新时, 2000).本研究利用研究区内92个固定监测样地的114个优势种(重要值>0.1)进行典范对应分析(CCA排序), 结果表明, 植物分布受到海拔、坡度和坡向这3个地形因子的影响.海拔是影响山地生境差异的主导因子(岳明等, 2002).如枫香树和赤楠等物种喜暖、腐殖质厚的土壤, 由于低海拔区域热量条件好, 水分较为充足, 故该种群大多分布在此区域; 而随着海拔升高, 环境温度逐渐降低, 更适合黄山松及柳杉等物种生长.而坡度是地形的另一项重要属性, 坡度的变化决定土壤持水量大小, 制约植物生长发育过程中对水分的需求(陈瑶等, 2006).同时, 土壤腐殖质的厚度也受到坡度变化的影响, 坡度越缓, 土壤腐殖质越深厚, 进而影响植物分布.如甜槠、苦槠和金发藓等物种喜湿润、深厚的土壤, 故在坡度较缓的环境中分布较多, 而地莓和水竹等灌木物种因其耐贫瘠、干旱而分布在坡度较陡处. ...

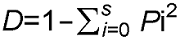
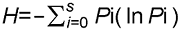
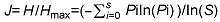
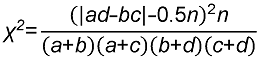
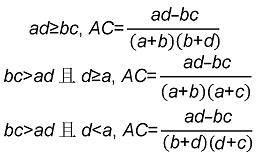
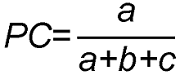


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}