李萍 1 , 2 ,
许言 1 ,
张晓丽 1 , 3 ,
李炯 4 ,
赵喜亭 1 , 3 ,
李明军 1 , 3 , * , 1 河南师范大学生命科学学院, 新乡 4530072 郑州澍青医学高等专科学校, 郑州 4500643 河南省高校道地中药材保育及利用工程技术研究中心/绿色药材生物技术河南省工程实验室, 新乡 4530074 河南省农业厅中药材生产技术服务中心, 郑州 450008Li Ping 1 , 2 ,
Xu Yan 1 ,
Zhang Xiaoli 1 , 3 ,
Li Jiong 4 ,
Zhao Xiting 1 , 3 ,
Li Mingjun 1 , 3 , * , 1 College of Life Sciences, Henan Normal University, Xinxiang 453007, China2 Zhengzhou Shuqing Medical College, Zhengzhou 450064, China3 Engineering Technology Research Center of Nursing and Utilization of Genuine Chinese Crude Drugs, University of Henan Province, Henan Province Engineering Laboratory of Green Medicinal Plant Biotechnology, Xinxiang 453007, China4 Chinese Medical Materials Manufacturing Technology Center, Department of Agriculture of Henan Province, Zhengzhou 450008, China引用本文 李萍 ,
许言 ,
张晓丽 ,
李炯 ,
赵喜亭 ,
李明军 . 不同诱导子对怀牛膝细胞生长及多糖含量的影响. , 2017, 52(5): 615-621
贡献者
* 通讯作者。E-mail: limingjun2002@263.net 基金资助
国家中医药行业科研专项(No.201407005-08)、河南省创新型科技人才队伍建设工程(No.C20130037)和新乡市科技创新平台建设项目(No.CP1401) ;
接受日期:2016-11-14接受日期:2017-03-6网络出版日期:2017-09-1
-->Copyright
2017《植物学报》编辑部
Contributors
* Author for correspondence. E-mail: limingjun2002@263.net History
Received :
Accepted :
Online :
摘要: 为提高怀牛膝(Achyranthes bidentata )悬浮培养细胞中牛膝多糖的含量, 以酵母提取物、榆黄蘑(Pleurotus citrinopileatus )及水杨酸作为诱导子, 分别在同一时期以不同浓度或在不同时期以相同浓度添加至细胞培养物中, 收获时测定细胞生长量和牛膝多糖含量。结果显示, 在培养12天时添加2.5% (v/v)酵母诱导子, 细胞干重最大, 可达46.75 g·L-1 , 多糖含量为5.76 mg·g-1 ; 在培养9天时添加5 mgGE·L-1 榆黄蘑诱导子, 收获时细胞中多糖含量可达6.56 mg·g-1 , 细胞干重达28.3 g·L-1 ; 1 mg·g-1 水杨酸对牛膝多糖的诱导效果不如以上2种诱导子明显。 关键词 :
怀牛膝 ;
细胞悬浮培养 ;
诱导子 ;
多糖 Abstract :
To enhance the polysaccharides content of Achyranthes bidentata suspension cell, yeast extract, Pleurotus citrinopileatus and salicylic acid were added at different concentrations or at different times as the elicitor. With 2.5% (v/v) yeast elicitor added at 12 days, the cell dry weight was the greatest, reaching 46.75 g·L-1 ; polysaccharides content was less, 5.76 mg·g-1 . With 5 mgGE·L-1 fungal elicitor added after culture for 9 days, polysaccharides content could reach 6.56 mg·g-1 ; with cell dry weight 28.3 g·L-1 . With 1 mg·g-1 SA elicitation, the effect was a little inferior to the above two effects. Key words :
Achyranthes bidentata ;
cell suspension culture ;
elicitor ;
polysaccharides 怀牛膝(
Achyranthes bidentata )隶属苋科牛膝属, 其干燥根具有逐瘀通经、补肝肾、强筋骨、利尿通淋和引血下行之功效(
国家药典委员会, 2015 )。牛膝含有多种药用成分, 其中重要的一类即植物中普遍存在的多糖类化合物, 具有多种生物学活性和功能(
石若夫, 2002 ;
崔倩倩等, 2015 )。药理研究表明, 牛膝多糖(
Achyranthes bidentata polysaccharides, ABPS)在抗肿瘤、抗衰老、免疫调节、降血糖和抗凝血等方面功效显著, 且具有分子量小和水溶性好等优点, 因此具有广阔的开发应用前景(
时春娟等, 2006 )。但由于多糖具有结构复杂、质量标准难以控制、结构测定及人工合成困难、天然含量低和不易分离等特点, 给多糖研究和应用带来了诸多限制(
李明军等, 2008a )。
通过植物细胞培养生产多糖可以解决以上问题。目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 )。利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 )。诱导子可分为生物诱导子与非生物诱导子两种。在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础。
1 材料与方法1.1 实验材料实验材料为怀牛膝(
Achyranthes bidentata Bl.)悬浮培养细胞。
1.2 实验方法1.2.1 诱导子的制备
酵母诱导子的制备: 采用
Hahn和Albersheim (1978) 的方法醇沉提取。将10 g酵母提取物溶解于100 mL双蒸水中, 加入无水乙醇至80% (v/v), 于4°C下静置3天, 沉淀。弃上清, 沉淀再溶解于100 mL双蒸水中, 121°C高压灭菌20分钟。
榆黄蘑(
Pleurotus citrinopileatus Sing.)诱导子的制备: 将榆黄蘑菌种在PDA固体斜面培养基上25°C恒温培养10天, 培养基组成为马铃薯20%, 蔗糖2%, 琼脂8%; 后转接到PDY液体培养基中置于25°C恒温振荡培养7天, 培养基组成为马铃薯20%, 蔗糖2%, 酵母提取物0.2%; 将菌球过滤, 称取3 g鲜重, 加15 mL双蒸水研磨, 8 000 ×
g 离心, 取上清液, 121°C高压灭菌20分钟(
Komaraiah et al., 2002 )。
水杨酸(salicylic acid, SA)母液质量浓度为0.5 mg·mL
-1 。
1.2.2 诱导子的添加
以50 g·L
-1 的接种量将怀牛膝细胞接种于添加30 g·L
-1 葡萄糖的B
5 培养基中, 在振荡培养箱中120 ×
g 悬浮培养, 培养温度为(25±2)°C。结合怀牛膝细胞生长曲线(
李萍等, 2015 )及诱导子添加时间(
Chen and Chen, 2000 )的研究结果, 实验初步选定在怀牛膝细胞接种12天时分别加入不同浓度的3种诱导子(酵母、榆黄蘑和水杨酸), 接种18天时收获细胞, 测定细胞干重和多糖含量, 以研究不同浓度的各种诱导子对细胞生长和多糖含量的影响。
酵母诱导子的添加浓度分别为0、0.5%、1.0%、1.5%、2.0%、2.5%、5.0%和10.0% (v/v)。榆黄蘑诱导子的浓度以单位mgGE·L
-1 (glucose equivalent, 葡萄糖当量)表示, 采用蒽酮硫酸法(
张纯等, 2003 )测定提取液的糖含量, 其添加浓度分别为0、5、20、50和100 mgGE·L
-1 。水杨酸的添加浓度分别为0、0.01、0.05、0.1、1和5 mg·L
-1 。
将上述实验获得的最适浓度的各种诱导子分别于细胞接种0、3、6、9、12和15天时添加入培养基中, 接种18天时收获细胞, 以研究不同诱导子的添加时间对细胞生长和多糖含量的影响。
1.2.3 细胞干重的测定
细胞干重测定参照
李明军等(2008a) 的方法进行。取1 mL摇匀的细胞悬液, 置于底部带3 mm深裂缝的Ep-pendorf管中, 2 325 ×
g 离心10分钟, 将得到的细胞置于60°C烘箱干燥至恒重, 称量得细胞干重(g·L
-1 )。
1.2.4 多糖含量的测定
取经40 μm筛过滤的细胞用重蒸水冲洗3次, 2 325 ×
g 离心10分钟, 称取0.2 g加水研磨至匀浆状, 加入1 mL 2 mol·L
-1 NaOH溶液于75-80°C水浴30分钟, 离心取上清液加入2倍体积的80%乙醇溶液沉淀过夜, 离心得沉淀, 稀释后采用蒽酮-硫酸比色法(
张纯等, 2003 )测定多糖含量。
采用Excel和SPSS软件对实验数据进行统计分析。采用LSD法进行多重比较检验。
2 结果与讨论2.1 酵母诱导子对怀牛膝细胞生长和多糖含量的影响2.1.1 酵母诱导子添加浓度
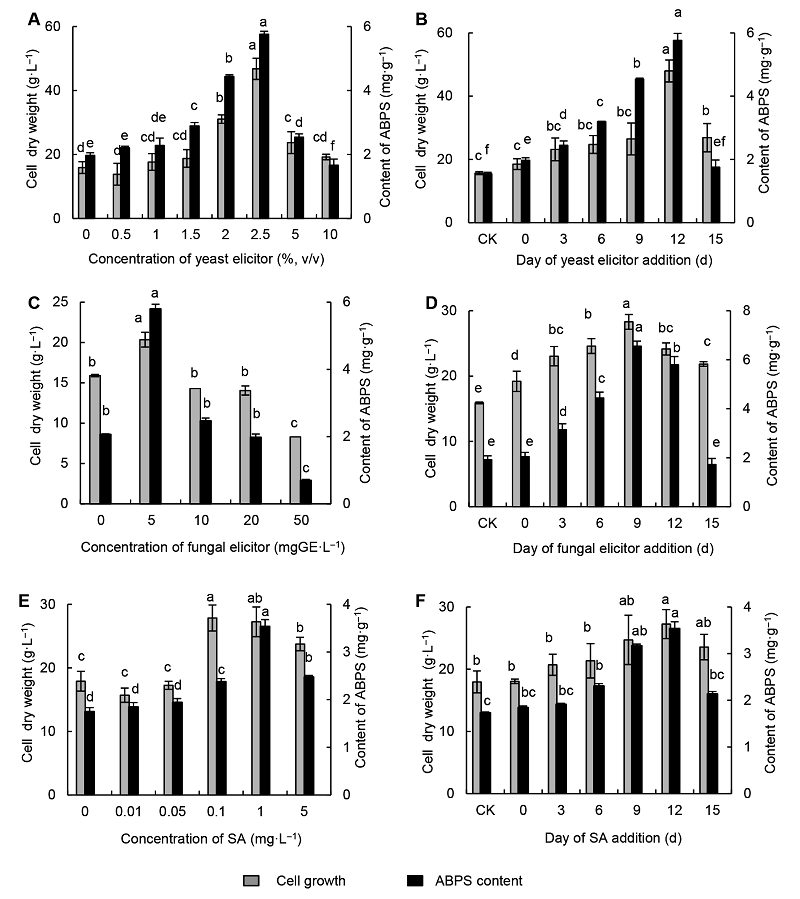
不同浓度(0、0.5%、1.0%、1.5%、2.0%、2.5%、5.0%和10.0% (v/v))酵母诱导子作用下, 怀牛膝细胞干重和多糖含量的测定结果见
图1A 。由
图1A 可知, 在较低的酵母诱导子浓度范围内, 细胞干重和多糖含量随其浓度的增加而有所升高, 在浓度为2.5%时均达到最高, 分别为46.75 g·L
-1 和5.76 mg·g
-1 , 与其它浓度相比差异显著; 酵母诱导子浓度超过2.5%, 细胞均产生不同程度的褐化, 细胞干重和多糖含量随之下降。因此, 酵母诱导子的最适浓度为2.5%。
图1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-615/img_1.png <b>图1</b> 不同诱导子对怀牛膝细胞生长和多糖含量的影响<br/>(A) 酵母诱导子浓度; (B) 酵母诱导子添加时间; (C) 真菌诱导子浓度; (D) 真菌诱导子添加时间; (E) 水杨酸浓度; (F) 水杨酸添加时间。不同小写字母表示差异显著(<i>P</i><0.05)。ABPS: 牛膝多糖; SA: 水杨酸<br/><b>Figure 1</b> Effect of different elicitors on the cell growth and polysaccharides contents of <i>Achyranthes bidentata</i><br/>(A) Concentration of yeast elicitor; (B) Addition day of yeast elicitor; (C) Concentration of fungal elicitor; (D) Addition day of fungal elicitor; (E) Concentration of salicylic acid; (F) Addition day of salicylic acid. Different lowercase letters indicate significant differences at <i>P</i><0.05. ABPS: <i>Achyranthes bidentata</i> polysaccharides; SA: Salicylic acid Figure 1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-615/img_1.png <b>图1</b> 不同诱导子对怀牛膝细胞生长和多糖含量的影响<br/>(A) 酵母诱导子浓度; (B) 酵母诱导子添加时间; (C) 真菌诱导子浓度; (D) 真菌诱导子添加时间; (E) 水杨酸浓度; (F) 水杨酸添加时间。不同小写字母表示差异显著(<i>P</i><0.05)。ABPS: 牛膝多糖; SA: 水杨酸<br/><b>Figure 1</b> Effect of different elicitors on the cell growth and polysaccharides contents of <i>Achyranthes bidentata</i><br/>(A) Concentration of yeast elicitor; (B) Addition day of yeast elicitor; (C) Concentration of fungal elicitor; (D) Addition day of fungal elicitor; (E) Concentration of salicylic acid; (F) Addition day of salicylic acid. Different lowercase letters indicate significant differences at <i>P</i><0.05. ABPS: <i>Achyranthes bidentata</i> polysaccharides; SA: Salicylic acid 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
不同诱导子对怀牛膝细胞生长和多糖含量的影响
(A) 酵母诱导子浓度; (B) 酵母诱导子添加时间; (C) 真菌诱导子浓度; (D) 真菌诱导子添加时间; (E) 水杨酸浓度; (F) 水杨酸添加时间。不同小写字母表示差异显著(
P <0.05)。ABPS: 牛膝多糖; SA: 水杨酸
Figure 1
Effect of different elicitors on the cell growth and polysaccharides contents of
Achyranthes bidentata (A) Concentration of yeast elicitor; (B) Addition day of yeast elicitor; (C) Concentration of fungal elicitor; (D) Addition day of fungal elicitor; (E) Concentration of salicylic acid; (F) Addition day of salicylic acid. Different lowercase letters indicate significant differences at
P <0.05. ABPS:
Achyranthes bidentata polysaccharides; SA: Salicylic acid
2.1.2 酵母诱导子添加时间
将2.5% (v/v)酵母诱导子在不同时间(接种0、3、6、9、12和15天)添加入培养基中, 接种18天怀牛膝细胞干重和多糖含量的测定结果见
图1B 。由
图1B 可知, 在延迟期(0和3天)和指数生长早期(6天)添加酵母诱导子, 细胞干重与对照相比无显著差异, 多糖含量稍有提高; 在指数生长中、后期(9、12天)添加, 多糖含量明显升高, 细胞干重也有较大提高; 接种12天添加, 细胞干重和多糖含量均达到最大值, 与其它添加时间相比差异显著; 在缓慢生长期早期(15天)添加, 细胞干重与对照相比有一定提高, 多糖含量与对照相比无显著差异。因此, 酵母诱导子的最适添加时间为接种后12天。
2.2 榆黄蘑真菌诱导子对怀牛膝细胞生长和多糖含量的影响2.2.1 榆黄蘑真菌诱导子添加浓度
将不同浓度(0、5、20、50和100 mgGE·L
-1 )的榆黄蘑诱导子添加于培养基中。接种3天后, 20和50 mgGE·
L
-1 浓度下的细胞即呈现不同程度的褐化, 以50 mg GE·L
-1 处理最为严重。接种18天细胞干重及多糖含量的测定结果见
图1C 。
由
图1C 可知, 榆黄蘑诱导子浓度为5 mgGE·L
-1 时, 细胞干重及多糖含量均比其它浓度及对照高, 且差异达显著水平, 分别为20.35 g·L
-1 和5.80 mg·g
-1 ; 浓度为10和20 mgGE·L
-1 时与对照相比无显著差异; 浓度为50 mgGE·L
-1 时细胞褐化严重, 细胞干重及多糖含量均较低。因此, 榆黄蘑诱导子的最适浓度为5 mgGE·L
-1 。
2.2.2 榆黄蘑真菌诱导子添加时间
将浓度为5 mgGE·L
-1 榆黄蘑诱导子在不同时间(接种0、3、6、9、12和15天)添加入培养基中, 接种18天细胞干重和多糖含量的测定结果见
图1D 。
由
图1D 可知, 在接种9天添加榆黄蘑诱导子, 细胞干重及多糖含量均达到最大值, 分别为28.3 g·L
-1 和6.56 mg·g
-1 , 与其它添加时间相比均达到显著差异; 在接种3、6和12天添加, 细胞干重和多糖含量与对照比均有显著提高; 在接种0和15天添加, 多糖含量与对照相比均无显著差异。因此, 榆黄蘑诱导子的最适添加时间为接种后9天。
2.3 水杨酸对怀牛膝细胞生长和多糖含量的影响2.3.1 水杨酸添加浓度
用不同浓度(0、0.01、0.05、0.1、1和5 mg·L
-1 )水杨酸处理后, 怀牛膝细胞干重和多糖含量的测定结果见
图1E 。
由
图1E 可知, 水杨酸浓度较低时, 对细胞干重和多糖含量影响不大, 与对照相比无显著差异; 浓度为0.1 mg·g
-1 时, 细胞干重达到最大, 为27.85 g·L
-1 , 与浓度为1 mg·g
-1 时无显著差异, 但与其它浓度相比均有显著差异; 水杨酸处理浓度为1 mg·g
-1 时, 细胞中多糖含量最大, 达3.54 mg·g
-1 ; 水杨酸浓度过高时, 两者均下降。因此, 水杨酸的最适浓度为1 mg·g
-1 。
2.3.2 水杨酸添加时间
将1 mg·g
-1 水杨酸在不同时间(接种0、3、6、9、12和15天)添加于培养基中, 接种18天细胞干重和多糖含量的测定结果见
图1F 。
图1F 显示, 在接种0、3和6天添加水杨酸, 细胞干重和多糖含量均没有太大的变化, 与对照相比无显著差异; 接种12天添加, 细胞干重最大, 但与9天及15天差异不显著, 多糖含量也最大, 与9天相比差异不显著, 但与其它浓度及对照相比均差异显著。因此, 水杨酸的最适添加时间为接种后12天。
2.4 讨论本研究首次探讨了不同诱导子对怀牛膝细胞生长和多糖含量的影响。结果表明, 酵母提取物、榆黄蘑及水杨酸均对怀牛膝细胞生长和多糖含量有明显影响。相比较而言, 榆黄蘑真菌诱导子可显著提高多糖含量, 但细胞生长量不及酵母诱导子。水杨酸对牛膝多糖的诱导效果不如以上2种诱导子明显。诱导子可诱导植物细胞的胁迫反应, 直接影响代谢产物的含量(
谢腾等, 2015 )。在短叶红豆杉(
Taxus brevifolia )细胞悬浮培养过程中加入各种诱导子, 可促使紫杉醇从细胞中分泌出来, 并可以进行连续培养(
张玲华和郭勇, 2006 )。但诱导子通常会影响细胞的正常生长。有****在红豆杉(
Taxus chinensis )细胞培养过程中首先添加壳聚糖诱导子, 使其产生适应, 这样就保证了细胞的生存力和渗透性; 然后加入Ag
+ 和茉莉酸甲酯2种诱导子, 结果表明已经适应壳聚糖的细胞中紫杉醇含量比未经过这一适应过程的细胞中紫杉醇含量高4.6倍, 释放到胞外的能力也提高3.2倍(
Zhang et al., 2007 )。
酵母诱导子的有效成分是糖类化合物, 可提高多种药用植物代谢产物的含量(
Funk et al., 1987 )。如酵母提取物浓度为2 g∙L
-1 时, 雷公藤(
Tripterygium wilfordii )甲素含量为对照的1.48倍(
李琰等, 2015 )。利用100 mg·L
-1 酵母提取物与50 g·L
-1 山梨糖醇共同诱导, 可使丹参(
Salvia miltiorrhiza )毛状根总丹参酮含量提高10倍, 丹参酮含量提高9倍(
Shi et al., 2007 )。酵母诱导子的添加浓度和添加时间对肉苁蓉(
Cistanche deserticola )细胞的生长和苯乙醇苷合成都有很大影响, 以在培养18天、添加浓度为0.8 μg·mL
-1 诱导效果最好(
魏涛等, 2005 )。本研究表明, 在怀牛膝细胞培养12天时添加2.5% (v/v)的酵母诱导子, 细胞生长和多糖含量均有显著提高, 分别可达46.75 g·L
-1 和5.76 mg·g
-1 。
真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 )。欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 )。魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 )。东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 )。黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 )。铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 )。以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性。本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证。本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 。综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子。另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究。
水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 )。添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 )。SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 )。加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 )。我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量。在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 )。
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究。
本研究就诱导子的添加时间对代谢产物含量的影响也进行了探讨, 结果表明, 在指数生长期的中后期(9-12天)添加各种诱导子, 怀牛膝细胞生长量和多糖含量均较高。这与对长春花(
Catharanthurs rose- us )、肉苁蓉和喜树(
Camptotheca acuminata )等的研究结果一致(
姜发军, 2004 ;
魏涛等, 2005 ;
于放, 2005 )。其原因可能是, 在细胞生长迟缓期及指数生长期早期, 还没有足够的前体可用于合成代谢需要的各种酶; 而在指数生长期的中后期, 各种代谢活动旺盛, 对诱导子比较敏感, 因此代谢产物含量明显升高, 细胞生长量也出现较大增长; 在缓慢生长期早期, 细胞的各种代谢水平都有不同程度的下降, 即使加入诱导子, 也仅能维持细胞的正常生长, 对代谢产物的合成影响很小。
The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 陈红艳 , 刘连成 , 董娟娥 , 夏广东 (2012 ). H2 O2 参与水杨酸诱导丹参培养细胞中丹酚酸B合成的信号转导. 生物工程学报 28, 834 -846 .[2] 初旸 (2008 ). 刺五加细胞培养体系的建立与有效成分积累的调控研究. 硕士论文. 沈阳: 沈阳药科大学 . pp. 65 -73 .[3] 崔倩倩 , 周志勇 , 刘朝奇 , 袁丁 (2015 ). 植物多糖作为免疫佐剂的研究进展. 时珍国医国药 26, 970 -972 .[4] 房慧勇 , 朱虹 , 丁海麦 , 韩华锐 , 刘晓兰 , 郝丽静 , 李旻辉 (2014 ). 影响愈伤组织中次生代谢产物含量的因素研究进展. 中国中药杂志 39, 2846 -2850 .[5] 古绍彬 , 龚慧 , 杨彬 , 卜美玲 (2013 ). 真菌诱导子在发酵工业中的应用现状及展望. 生物工程学报 29, 1558 -1572 .[6] 国家药典委员会 (2015 ). 中华人民共和国药典(2015年版) . 北京: 中国医药科技出版社. pp. 38 .[7] 姜发军 (2004 ). 黑曲霉提取物与水杨酸对长春花悬浮细胞生长及生物碱代谢作用的研究. 硕士论文. 长沙: 湖南农业大学 . pp. 4 .[8] 李明军 , 李萍 , 张晓丽 , 赵喜亭 , 王凤娟 , 张楠 , 邓丽 (2008 a). 氮源和碳源对怀牛膝悬浮培养细胞生长及多糖含量的影响. 见: 中华中医药学会2008年药用植物化学与中药有效成分分析研讨会论文集(上). 深圳: 中华中医药学会 . pp. 448 -451 .[9] 李明军 , 李萍 , 赵喜亭 , 张晓丽 , 周娜 , 王凤娟 (2008 b). 怀牛膝细胞悬浮培养及多糖含量变化的研究. 西北植物学报 28, 494 -500 .[10] 李萍 , 赵喜亭 , 李明军 , 周彩云 (2015 ). 怀牛膝细胞悬浮培养生理生化特性研究. 河南农业科学 44, 106 -110 .[11] 李文渊 , 高伟 , 邵爱娟 , 何云飞 , 黄璐琦 (2011 ). 诱导子对丹参有效成分次生代谢的诱导与调控. 中国中药杂志 36, 258 -262 .[12] 李琰 , 赵磊 , 崔蕾 , 雷嘉敏 , 张兴 (2015 ). 诱导子对雷公藤不定根生长和次生代谢产物含量的影响. 生物工程学报 31, 734 -743 .[13] 李永成 , 陶文沂 (2008 ). 内生真菌培养液对东北红豆杉细胞生长及紫杉醇合成的影响. 植物学通报 25, 552 -558 .[14] 石若夫 (2002 ). 栀子组织和悬浮细胞培养及其多糖的分离纯化和生物学活性的研究. 博士论文. 大连: 大连理工大学. pp . 12 .[15] 时春娟 , 周永达 , 张剑波 , 田庚元 (2006 ). 牛膝多糖研究进展. 中国新药杂志 15, 1330 -1334 .[16] 孙晶 , 高珂 , 王玲 , 吴素瑞 , 杨洪一 , 隋春 (2015 ). 不同培养基、外源激素和真菌诱导子对北柴胡毛状根生长及柴胡皂苷含量的影响. 生物技术通讯 26, 546 -550 .[17] 魏涛 , 成喜雨 , 杨秋婷 , 刘春朝 (2005 ). 酵母诱导子对肉苁蓉细胞悬浮培养影响的研究. 北京联合大学学报(自然科学版) 19, 69 -73 .[18] 谢腾 , 莫歌 , 张山山 , 王升 , 康利平 , 郭兰萍 (2015 ). 不同诱导子对植物次生代谢的影响. 中国实验方剂学杂志 21, 210 -215 .[19] 杨慧 , 陈晓梅 , 郭顺星 (2009 ). 真菌诱导子对铁皮石斛原球茎多糖含量的影响. 世界科学技术中医药现代化 11, 719 -722 .[20] 杨静 , 孙皓 (2014 ). 不同诱导子对黄芪悬浮培养细胞中黄芪多糖积累的影响. 安徽农业科学 42, 8954 -8956, 8959 .[21] 于放 (2005 ). 喜树细胞培养体系的建立与喜树异碱A和B的生物合成及过程调控. 博士论文. 大连: 大连理工大学. pp . 13 .[22] 张春荣 , 李玲 (2003 ). 水杨酸、茉莉酸甲酯和乙烯利对野葛细胞悬浮培养生产葛根素的影响. 植物资源与环境学报 12, 56 -57 .[23] 张纯 , 郭文彪 , 王雯佶 , 陈爱华 , 郭澄 (2003 ). 改良硫酸蒽酮比色法测定牛膝多糖的含量. 中华临床医药 4, 17 -18 .[24] 张玲华 , 郭勇 (2006 ). 药用植物细胞的选育与培养生产次级代谢产物. 生物技术通讯 17, 105 -108 .[25] 张顺仓 , 刘岩 , 沈双 , 梁宗锁 , 杨东风 (2011 ). 诱导子对丹参毛状根酚酸类和丹参酮类成分积累的影响. 中国中药杂志 36, 1269 -1274 .[26] Bulgakov VP Tchernoded GK Mischenko NP Khodakovskaya MV Glazunov VP Radchenko SV Zvereva EV Fedoreyev SA Zhuravlev YN 2002 ). Effect of salicylic acid, methyl jasmonate, ethephon and cantharidin on anthraquinone production by Rubia cordifolia callus cultures transformed with the rolB and rolC genes. J Biotechnol 97, 213 -221 .[27] Chen H Chen F 2000 ). Effects of yeast elicitor on the growth and secondary metabolism of a high-tanshinone- producing line of the Ti transformed Salvia miltiorrhiza cells in suspension culture.Process Biochem 35, 837 -840 .[28] Funk C Gugler K Brodelius P 1987 ). Increased secondary product formation in plant cell suspension cultures after treatment with a yeast carbohydrate preparation (eli- citor).Phytochemistry 26, 401 -405 .[29] Hahn MG Albersheim P 1978 ). Host-pathogen interactions: XIV. isolation and partial characterization of an elicitor from yeast extract.Plant Physiol 62, 107 -111 .[30] Han J Zhong JJ 2003 ). Effects of oxygen partial pressure on cell growth and ginsenoside and polysaccharide production in high density cell cultures of Panax notoginseng.Enzyme Microb Technol 32, 498 -503 .[31] Ibrahim AK Khalifa S Khafagi I Youssef D Khan I Mesbah M 2007 ). Stimulation of oleandrin production by combined Agrobacterium tumefaciens mediated transformation and fungal elicitation in Nerium oleander cell cultures.Enzyme Microb Technol 41, 331 -336 .[32] Komaraiah P Amrutha RN Kishor PBK Ramakrishna SV 2002 ). Elicitor enhanced production of plumbagin in suspension cultures of Plumbagorosea L. Enzyme Microb Technol 31, 634 -639 .[33] Liu S Zhong JJ 1997 ). Simultaneous production of ginseng saponin and polysaccharide by suspension cultures of Panax ginseng: nitrogen effects.Enzyme Microb Tech- nol 21, 518 -524 .[34] Shi M Kwok KW Wu JY 2007 ). Enhancement of tanshinone production in Salvia miltiorrhiza Bunge (red or Chinese sage) hairy-root culture by hyperosmotic stress and yeast elicitor.Biotechnol Appl Biochem 46, 191 -196 .[35] Vardar-Sukan F Sukan SS 1992 ). Recent Advances in Biotechnology. Berlin: Springer. pp . 209 .[36] Wang YD Yuan YJ Wu JC 2004 ). Induction studies of methyl jasmonate and salicylic acid on taxane production in suspension cultures of Taxus chinensis var. mairei.Bio- chem Eng J 19, 259 -265 .[37] Zhang CH Fevereiro PS He GY Chen ZJ 2007 ). Enhanced paclitaxel productivity and release capacity of Taxus chinensis cell suspension cultures adapted to chitosan.Plant Sci 172, 158 -163 .
H
2 O
2 参与水杨酸诱导丹参培养细胞中丹酚酸B合成的信号转导
1
2012
... 水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 ).添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 ).SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 ).加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 ).我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量.在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 ).
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究. ...
刺五加细胞培养体系的建立与有效成分积累的调控研究
1
2008
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
植物多糖作为免疫佐剂的研究进展
1
2015
... 怀牛膝(
Achyranthes bidentata )隶属苋科牛膝属, 其干燥根具有逐瘀通经、补肝肾、强筋骨、利尿通淋和引血下行之功效(
国家药典委员会, 2015 ).牛膝含有多种药用成分, 其中重要的一类即植物中普遍存在的多糖类化合物, 具有多种生物学活性和功能(
石若夫, 2002 ;
崔倩倩等, 2015 ).药理研究表明, 牛膝多糖(
Achyranthes bidentata polysaccharides, ABPS)在抗肿瘤、抗衰老、免疫调节、降血糖和抗凝血等方面功效显著, 且具有分子量小和水溶性好等优点, 因此具有广阔的开发应用前景(
时春娟等, 2006 ).但由于多糖具有结构复杂、质量标准难以控制、结构测定及人工合成困难、天然含量低和不易分离等特点, 给多糖研究和应用带来了诸多限制(
李明军等, 2008a ). ...
影响愈伤组织中次生代谢产物含量的因素研究进展
1
2014
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
真菌诱导子在发酵工业中的应用现状及展望
1
2013
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
1
2015
... 怀牛膝(
Achyranthes bidentata )隶属苋科牛膝属, 其干燥根具有逐瘀通经、补肝肾、强筋骨、利尿通淋和引血下行之功效(
国家药典委员会, 2015 ).牛膝含有多种药用成分, 其中重要的一类即植物中普遍存在的多糖类化合物, 具有多种生物学活性和功能(
石若夫, 2002 ;
崔倩倩等, 2015 ).药理研究表明, 牛膝多糖(
Achyranthes bidentata polysaccharides, ABPS)在抗肿瘤、抗衰老、免疫调节、降血糖和抗凝血等方面功效显著, 且具有分子量小和水溶性好等优点, 因此具有广阔的开发应用前景(
时春娟等, 2006 ).但由于多糖具有结构复杂、质量标准难以控制、结构测定及人工合成困难、天然含量低和不易分离等特点, 给多糖研究和应用带来了诸多限制(
李明军等, 2008a ). ...
黑曲霉提取物与水杨酸对长春花悬浮细胞生长及生物碱代谢作用的研究
2
2004
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
... 本研究就诱导子的添加时间对代谢产物含量的影响也进行了探讨, 结果表明, 在指数生长期的中后期(9-12天)添加各种诱导子, 怀牛膝细胞生长量和多糖含量均较高.这与对长春花(
Catharanthurs rose- us )、肉苁蓉和喜树(
Camptotheca acuminata )等的研究结果一致(
姜发军, 2004 ;
魏涛等, 2005 ;
于放, 2005 ).其原因可能是, 在细胞生长迟缓期及指数生长期早期, 还没有足够的前体可用于合成代谢需要的各种酶; 而在指数生长期的中后期, 各种代谢活动旺盛, 对诱导子比较敏感, 因此代谢产物含量明显升高, 细胞生长量也出现较大增长; 在缓慢生长期早期, 细胞的各种代谢水平都有不同程度的下降, 即使加入诱导子, 也仅能维持细胞的正常生长, 对代谢产物的合成影响很小. ...
氮源和碳源对怀牛膝悬浮培养细胞生长及多糖含量的影响
3
2008
... 怀牛膝(
Achyranthes bidentata )隶属苋科牛膝属, 其干燥根具有逐瘀通经、补肝肾、强筋骨、利尿通淋和引血下行之功效(
国家药典委员会, 2015 ).牛膝含有多种药用成分, 其中重要的一类即植物中普遍存在的多糖类化合物, 具有多种生物学活性和功能(
石若夫, 2002 ;
崔倩倩等, 2015 ).药理研究表明, 牛膝多糖(
Achyranthes bidentata polysaccharides, ABPS)在抗肿瘤、抗衰老、免疫调节、降血糖和抗凝血等方面功效显著, 且具有分子量小和水溶性好等优点, 因此具有广阔的开发应用前景(
时春娟等, 2006 ).但由于多糖具有结构复杂、质量标准难以控制、结构测定及人工合成困难、天然含量低和不易分离等特点, 给多糖研究和应用带来了诸多限制(
李明军等, 2008a ). ...
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
... 细胞干重测定参照
李明军等(2008a) 的方法进行.取1 mL摇匀的细胞悬液, 置于底部带3 mm深裂缝的Ep-pendorf管中, 2 325 ×
g 离心10分钟, 将得到的细胞置于60°C烘箱干燥至恒重, 称量得细胞干重(g·L
-1 ). ...
怀牛膝细胞悬浮培养及多糖含量变化的研究
1
2008
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
怀牛膝细胞悬浮培养生理生化特性研究
2
2015
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
... 以50 g·L
-1 的接种量将怀牛膝细胞接种于添加30 g·L
-1 葡萄糖的B
5 培养基中, 在振荡培养箱中120 ×
g 悬浮培养, 培养温度为(25±2)°C.结合怀牛膝细胞生长曲线(
李萍等, 2015 )及诱导子添加时间(
Chen and Chen, 2000 )的研究结果, 实验初步选定在怀牛膝细胞接种12天时分别加入不同浓度的3种诱导子(酵母、榆黄蘑和水杨酸), 接种18天时收获细胞, 测定细胞干重和多糖含量, 以研究不同浓度的各种诱导子对细胞生长和多糖含量的影响. ...
诱导子对丹参有效成分次生代谢的诱导与调控
1
2011
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
诱导子对雷公藤不定根生长和次生代谢产物含量的影响
1
2015
... 酵母诱导子的有效成分是糖类化合物, 可提高多种药用植物代谢产物的含量(
Funk et al., 1987 ).如酵母提取物浓度为2 g∙L
-1 时, 雷公藤(
Tripterygium wilfordii )甲素含量为对照的1.48倍(
李琰等, 2015 ).利用100 mg·L
-1 酵母提取物与50 g·L
-1 山梨糖醇共同诱导, 可使丹参(
Salvia miltiorrhiza )毛状根总丹参酮含量提高10倍, 丹参酮含量提高9倍(
Shi et al., 2007 ).酵母诱导子的添加浓度和添加时间对肉苁蓉(
Cistanche deserticola )细胞的生长和苯乙醇苷合成都有很大影响, 以在培养18天、添加浓度为0.8 μg·mL
-1 诱导效果最好(
魏涛等, 2005 ).本研究表明, 在怀牛膝细胞培养12天时添加2.5% (v/v)的酵母诱导子, 细胞生长和多糖含量均有显著提高, 分别可达46.75 g·L
-1 和5.76 mg·g
-1 . ...
内生真菌培养液对东北红豆杉细胞生长及紫杉醇合成的影响
1
2008
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
栀子组织和悬浮细胞培养及其多糖的分离纯化和生物学活性的研究
1
2002
... 怀牛膝(
Achyranthes bidentata )隶属苋科牛膝属, 其干燥根具有逐瘀通经、补肝肾、强筋骨、利尿通淋和引血下行之功效(
国家药典委员会, 2015 ).牛膝含有多种药用成分, 其中重要的一类即植物中普遍存在的多糖类化合物, 具有多种生物学活性和功能(
石若夫, 2002 ;
崔倩倩等, 2015 ).药理研究表明, 牛膝多糖(
Achyranthes bidentata polysaccharides, ABPS)在抗肿瘤、抗衰老、免疫调节、降血糖和抗凝血等方面功效显著, 且具有分子量小和水溶性好等优点, 因此具有广阔的开发应用前景(
时春娟等, 2006 ).但由于多糖具有结构复杂、质量标准难以控制、结构测定及人工合成困难、天然含量低和不易分离等特点, 给多糖研究和应用带来了诸多限制(
李明军等, 2008a ). ...
牛膝多糖研究进展
1
2006
... 怀牛膝(
Achyranthes bidentata )隶属苋科牛膝属, 其干燥根具有逐瘀通经、补肝肾、强筋骨、利尿通淋和引血下行之功效(
国家药典委员会, 2015 ).牛膝含有多种药用成分, 其中重要的一类即植物中普遍存在的多糖类化合物, 具有多种生物学活性和功能(
石若夫, 2002 ;
崔倩倩等, 2015 ).药理研究表明, 牛膝多糖(
Achyranthes bidentata polysaccharides, ABPS)在抗肿瘤、抗衰老、免疫调节、降血糖和抗凝血等方面功效显著, 且具有分子量小和水溶性好等优点, 因此具有广阔的开发应用前景(
时春娟等, 2006 ).但由于多糖具有结构复杂、质量标准难以控制、结构测定及人工合成困难、天然含量低和不易分离等特点, 给多糖研究和应用带来了诸多限制(
李明军等, 2008a ). ...
不同培养基、外源激素和真菌诱导子对北柴胡毛状根生长及柴胡皂苷含量的影响
1
2015
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
酵母诱导子对肉苁蓉细胞悬浮培养影响的研究
2
2005
... 酵母诱导子的有效成分是糖类化合物, 可提高多种药用植物代谢产物的含量(
Funk et al., 1987 ).如酵母提取物浓度为2 g∙L
-1 时, 雷公藤(
Tripterygium wilfordii )甲素含量为对照的1.48倍(
李琰等, 2015 ).利用100 mg·L
-1 酵母提取物与50 g·L
-1 山梨糖醇共同诱导, 可使丹参(
Salvia miltiorrhiza )毛状根总丹参酮含量提高10倍, 丹参酮含量提高9倍(
Shi et al., 2007 ).酵母诱导子的添加浓度和添加时间对肉苁蓉(
Cistanche deserticola )细胞的生长和苯乙醇苷合成都有很大影响, 以在培养18天、添加浓度为0.8 μg·mL
-1 诱导效果最好(
魏涛等, 2005 ).本研究表明, 在怀牛膝细胞培养12天时添加2.5% (v/v)的酵母诱导子, 细胞生长和多糖含量均有显著提高, 分别可达46.75 g·L
-1 和5.76 mg·g
-1 . ...
... 本研究就诱导子的添加时间对代谢产物含量的影响也进行了探讨, 结果表明, 在指数生长期的中后期(9-12天)添加各种诱导子, 怀牛膝细胞生长量和多糖含量均较高.这与对长春花(
Catharanthurs rose- us )、肉苁蓉和喜树(
Camptotheca acuminata )等的研究结果一致(
姜发军, 2004 ;
魏涛等, 2005 ;
于放, 2005 ).其原因可能是, 在细胞生长迟缓期及指数生长期早期, 还没有足够的前体可用于合成代谢需要的各种酶; 而在指数生长期的中后期, 各种代谢活动旺盛, 对诱导子比较敏感, 因此代谢产物含量明显升高, 细胞生长量也出现较大增长; 在缓慢生长期早期, 细胞的各种代谢水平都有不同程度的下降, 即使加入诱导子, 也仅能维持细胞的正常生长, 对代谢产物的合成影响很小. ...
不同诱导子对植物次生代谢的影响
2
2015
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
... 本研究首次探讨了不同诱导子对怀牛膝细胞生长和多糖含量的影响.结果表明, 酵母提取物、榆黄蘑及水杨酸均对怀牛膝细胞生长和多糖含量有明显影响.相比较而言, 榆黄蘑真菌诱导子可显著提高多糖含量, 但细胞生长量不及酵母诱导子.水杨酸对牛膝多糖的诱导效果不如以上2种诱导子明显.诱导子可诱导植物细胞的胁迫反应, 直接影响代谢产物的含量(
谢腾等, 2015 ).在短叶红豆杉(
Taxus brevifolia )细胞悬浮培养过程中加入各种诱导子, 可促使紫杉醇从细胞中分泌出来, 并可以进行连续培养(
张玲华和郭勇, 2006 ).但诱导子通常会影响细胞的正常生长.有****在红豆杉(
Taxus chinensis )细胞培养过程中首先添加壳聚糖诱导子, 使其产生适应, 这样就保证了细胞的生存力和渗透性; 然后加入Ag
+ 和茉莉酸甲酯2种诱导子, 结果表明已经适应壳聚糖的细胞中紫杉醇含量比未经过这一适应过程的细胞中紫杉醇含量高4.6倍, 释放到胞外的能力也提高3.2倍(
Zhang et al., 2007 ). ...
真菌诱导子对铁皮石斛原球茎多糖含量的影响
2
2009
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
不同诱导子对黄芪悬浮培养细胞中黄芪多糖积累的影响
2
2014
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
... 水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 ).添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 ).SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 ).加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 ).我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量.在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 ).
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究. ...
喜树细胞培养体系的建立与喜树异碱A和B的生物合成及过程调控
2
2005
... 水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 ).添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 ).SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 ).加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 ).我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量.在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 ).
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究. ...
... 本研究就诱导子的添加时间对代谢产物含量的影响也进行了探讨, 结果表明, 在指数生长期的中后期(9-12天)添加各种诱导子, 怀牛膝细胞生长量和多糖含量均较高.这与对长春花(
Catharanthurs rose- us )、肉苁蓉和喜树(
Camptotheca acuminata )等的研究结果一致(
姜发军, 2004 ;
魏涛等, 2005 ;
于放, 2005 ).其原因可能是, 在细胞生长迟缓期及指数生长期早期, 还没有足够的前体可用于合成代谢需要的各种酶; 而在指数生长期的中后期, 各种代谢活动旺盛, 对诱导子比较敏感, 因此代谢产物含量明显升高, 细胞生长量也出现较大增长; 在缓慢生长期早期, 细胞的各种代谢水平都有不同程度的下降, 即使加入诱导子, 也仅能维持细胞的正常生长, 对代谢产物的合成影响很小. ...
水杨酸、茉莉酸甲酯和乙烯利对野葛细胞悬浮培养生产葛根素的影响
1
2003
... 水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 ).添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 ).SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 ).加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 ).我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量.在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 ).
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究. ...
改良硫酸蒽酮比色法测定牛膝多糖的含量
2
2003
... 酵母诱导子的添加浓度分别为0、0.5%、1.0%、1.5%、2.0%、2.5%、5.0%和10.0% (v/v).榆黄蘑诱导子的浓度以单位mgGE·L
-1 (glucose equivalent, 葡萄糖当量)表示, 采用蒽酮硫酸法(
张纯等, 2003 )测定提取液的糖含量, 其添加浓度分别为0、5、20、50和100 mgGE·L
-1 .水杨酸的添加浓度分别为0、0.01、0.05、0.1、1和5 mg·L
-1 . ...
... 取经40 μm筛过滤的细胞用重蒸水冲洗3次, 2 325 ×
g 离心10分钟, 称取0.2 g加水研磨至匀浆状, 加入1 mL 2 mol·L
-1 NaOH溶液于75-80°C水浴30分钟, 离心取上清液加入2倍体积的80%乙醇溶液沉淀过夜, 离心得沉淀, 稀释后采用蒽酮-硫酸比色法(
张纯等, 2003 )测定多糖含量. ...
药用植物细胞的选育与培养生产次级代谢产物
1
2006
... 本研究首次探讨了不同诱导子对怀牛膝细胞生长和多糖含量的影响.结果表明, 酵母提取物、榆黄蘑及水杨酸均对怀牛膝细胞生长和多糖含量有明显影响.相比较而言, 榆黄蘑真菌诱导子可显著提高多糖含量, 但细胞生长量不及酵母诱导子.水杨酸对牛膝多糖的诱导效果不如以上2种诱导子明显.诱导子可诱导植物细胞的胁迫反应, 直接影响代谢产物的含量(
谢腾等, 2015 ).在短叶红豆杉(
Taxus brevifolia )细胞悬浮培养过程中加入各种诱导子, 可促使紫杉醇从细胞中分泌出来, 并可以进行连续培养(
张玲华和郭勇, 2006 ).但诱导子通常会影响细胞的正常生长.有****在红豆杉(
Taxus chinensis )细胞培养过程中首先添加壳聚糖诱导子, 使其产生适应, 这样就保证了细胞的生存力和渗透性; 然后加入Ag
+ 和茉莉酸甲酯2种诱导子, 结果表明已经适应壳聚糖的细胞中紫杉醇含量比未经过这一适应过程的细胞中紫杉醇含量高4.6倍, 释放到胞外的能力也提高3.2倍(
Zhang et al., 2007 ). ...
诱导子对丹参毛状根酚酸类和丹参酮类成分积累的影响
1
2011
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
1
2002
... 水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 ).添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 ).SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 ).加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 ).我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量.在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 ).
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究. ...
1
2000
... 以50 g·L
-1 的接种量将怀牛膝细胞接种于添加30 g·L
-1 葡萄糖的B
5 培养基中, 在振荡培养箱中120 ×
g 悬浮培养, 培养温度为(25±2)°C.结合怀牛膝细胞生长曲线(
李萍等, 2015 )及诱导子添加时间(
Chen and Chen, 2000 )的研究结果, 实验初步选定在怀牛膝细胞接种12天时分别加入不同浓度的3种诱导子(酵母、榆黄蘑和水杨酸), 接种18天时收获细胞, 测定细胞干重和多糖含量, 以研究不同浓度的各种诱导子对细胞生长和多糖含量的影响. ...
1
1987
... 酵母诱导子的有效成分是糖类化合物, 可提高多种药用植物代谢产物的含量(
Funk et al., 1987 ).如酵母提取物浓度为2 g∙L
-1 时, 雷公藤(
Tripterygium wilfordii )甲素含量为对照的1.48倍(
李琰等, 2015 ).利用100 mg·L
-1 酵母提取物与50 g·L
-1 山梨糖醇共同诱导, 可使丹参(
Salvia miltiorrhiza )毛状根总丹参酮含量提高10倍, 丹参酮含量提高9倍(
Shi et al., 2007 ).酵母诱导子的添加浓度和添加时间对肉苁蓉(
Cistanche deserticola )细胞的生长和苯乙醇苷合成都有很大影响, 以在培养18天、添加浓度为0.8 μg·mL
-1 诱导效果最好(
魏涛等, 2005 ).本研究表明, 在怀牛膝细胞培养12天时添加2.5% (v/v)的酵母诱导子, 细胞生长和多糖含量均有显著提高, 分别可达46.75 g·L
-1 和5.76 mg·g
-1 . ...
1
1978
... 酵母诱导子的制备: 采用
Hahn和Albersheim (1978) 的方法醇沉提取.将10 g酵母提取物溶解于100 mL双蒸水中, 加入无水乙醇至80% (v/v), 于4°C下静置3天, 沉淀.弃上清, 沉淀再溶解于100 mL双蒸水中, 121°C高压灭菌20分钟. ...
1
2003
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
1
2007
... 真菌诱导子的有效成分为其细胞壁酶解后的多糖类物质, 其可刺激细胞代谢产物的形成(
姜发军, 2004 ).欧洲夹竹桃(
Nerium oleander )细胞培养过程中, 采用10 g·L
-1 曲霉菌诱导, 欧夹竹桃甙含量可达到对照的8.8倍(
Ibrahim et al., 2007 ).魔芋(
Amo- rphophallus rivieri )内生菌菌丝提取物抑制了丹参毛状根的生长和酚酸类成分的积累, 促进了丹参酮的积累(
张顺仓等, 2011 ).东北红豆杉(
Taxus cuspidata )内生真菌
Fusarium mairei 可明显提高细胞紫杉醇产量和释放率, 对细胞生长短暂抑制后又可使其恢复至正常水平(
李永成和陶文沂, 2008 ).黑曲霉诱导子对北柴胡(
Bupleurum chinese )毛状根生长有抑制作用, 对柴胡皂苷合成则有一定的促进作用(
孙晶等, 2015 ).铁皮石斛在培养14天时加入浓度为250 mL·L
-1 的小菇属MF24真菌诱导子, 培养2天后, 多糖含量最高(5.92%), 与平均值(4.59%)相比提高了29.0% (
杨慧等, 2009 ).以上研究表明, 真菌诱导子的选择还处于探索阶段, 并无很强的针对性.本研究选择榆黄蘑作为诱导子, 一是考虑到其细胞壁水解产物主要成分为多糖, 对怀牛膝细胞中多糖的合成可能起到诱导作用; 二是其药食兼用, 比霉菌等致病菌作为诱导子在安全性上有保证.本研究结果显示, 榆黄蘑真菌诱导子对怀牛膝多糖含量影响比较显著, 以5 mgGE·L
-1 浓度在接种9天添加, 收获时多糖含量可达6.56 mg·g
-1 , 但此时细胞干重仅为28.3 g·L
-1 .综合两方面考虑, 酵母诱导子的诱导效果好于榆黄蘑真菌诱导子.另外, 真菌诱导子在细胞培养中的应用, 除了其在生物技术上的用途, 还可以用来揭示微生物与植物之间的相互作用及植物防御反应的信号传递途径等(
房慧勇等, 2014 ), 但还有待于进行深入研究. ...
1
2002
... 榆黄蘑(
Pleurotus citrinopileatus Sing.)诱导子的制备: 将榆黄蘑菌种在PDA固体斜面培养基上25°C恒温培养10天, 培养基组成为马铃薯20%, 蔗糖2%, 琼脂8%; 后转接到PDY液体培养基中置于25°C恒温振荡培养7天, 培养基组成为马铃薯20%, 蔗糖2%, 酵母提取物0.2%; 将菌球过滤, 称取3 g鲜重, 加15 mL双蒸水研磨, 8 000 ×
g 离心, 取上清液, 121°C高压灭菌20分钟(
Komaraiah et al., 2002 ). ...
1
1997
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
1
2007
... 酵母诱导子的有效成分是糖类化合物, 可提高多种药用植物代谢产物的含量(
Funk et al., 1987 ).如酵母提取物浓度为2 g∙L
-1 时, 雷公藤(
Tripterygium wilfordii )甲素含量为对照的1.48倍(
李琰等, 2015 ).利用100 mg·L
-1 酵母提取物与50 g·L
-1 山梨糖醇共同诱导, 可使丹参(
Salvia miltiorrhiza )毛状根总丹参酮含量提高10倍, 丹参酮含量提高9倍(
Shi et al., 2007 ).酵母诱导子的添加浓度和添加时间对肉苁蓉(
Cistanche deserticola )细胞的生长和苯乙醇苷合成都有很大影响, 以在培养18天、添加浓度为0.8 μg·mL
-1 诱导效果最好(
魏涛等, 2005 ).本研究表明, 在怀牛膝细胞培养12天时添加2.5% (v/v)的酵母诱导子, 细胞生长和多糖含量均有显著提高, 分别可达46.75 g·L
-1 和5.76 mg·g
-1 . ...
1
1992
... 通过植物细胞培养生产多糖可以解决以上问题.目前已在人参(
Panax ginseng )、三七(
Panax notogin- seng )、黄芪(
Astragalus membranaceus )、刺五加(
Acanthopanax senticosus )和铁皮石斛(
Dendro- bium candidum )等多种植物中进行了相关研究(
Liu and Zhong, 1997 ;
Han and Zhong, 2003 ;
初旸, 2008 ;
杨慧等, 2009 ;
杨静和孙皓, 2014 ); 并且利用生物反应器培养紫松果菊(
Echinacea purpurea )细胞生产免疫活性多糖已经获得成功(
Vardar-Sukan and Sukan, 1992 ).利用诱导子来提高代谢物的产量, 是目前植物细胞培养中常用的方法(
李文渊等, 2011 ;
古绍彬等, 2013 ;
谢腾等, 2015 ).诱导子可分为生物诱导子与非生物诱导子两种.在怀牛膝细胞培养及多糖相关研究(
李明军等, 2008a ,
2008b ;
李萍等, 2015 )的基础上, 我们进一步探究了诱导子对怀牛膝悬浮培养细胞生长和多糖含量的影响, 以期提高怀牛膝细胞生长量和多糖含量, 为通过细胞培养途径工厂化生产牛膝多糖奠定技术基础. ...
1
2004
... 水杨酸是一种广泛存在于植物体内的酚类物质, 参与许多生理过程, 同时在植物抗病反应中发挥重要作用(
于放, 2005 ).添加0.1 mg·L
-1 SA, 可使野葛(
Pueraria lobata )细胞葛根素产量提高12.61% (
张春荣和李玲, 2003 ).SA还可以显著提高茜草(
Rubia cordifolia )细胞中蒽醌的产量(
Bulgakov et al., 2002 ).加入10.0 mg·L
-1 水杨酸可使黄芪多糖的含量达到对照组的2.04倍(
杨静和孙皓, 2014 ).我们在怀牛膝细胞培养的12天添加1 mg·g
-1 的SA可以显著提高细胞的生长量和多糖含量.在SA诱导丹参细胞Sal B合成的研究中, 发现H
2 O
2 是介导Sal B合成积累的信号分子, SA处理可有效诱导细胞中H
2 O
2 产生及PAL和TAT活性升高(
陈红艳等, 2012 ).
Wang等(2004) 对SA作用机理的研究做了大胆的设想, 假设有SA受体存在, 建立了一种可以推断诱导子诱导效应的模型, 且模型推断出的最佳浓度与实验所得结果一致, 只是该模型还有一定的局限性, 更加完善的模型还有待进一步研究. ...
1
2007
... 本研究首次探讨了不同诱导子对怀牛膝细胞生长和多糖含量的影响.结果表明, 酵母提取物、榆黄蘑及水杨酸均对怀牛膝细胞生长和多糖含量有明显影响.相比较而言, 榆黄蘑真菌诱导子可显著提高多糖含量, 但细胞生长量不及酵母诱导子.水杨酸对牛膝多糖的诱导效果不如以上2种诱导子明显.诱导子可诱导植物细胞的胁迫反应, 直接影响代谢产物的含量(
谢腾等, 2015 ).在短叶红豆杉(
Taxus brevifolia )细胞悬浮培养过程中加入各种诱导子, 可促使紫杉醇从细胞中分泌出来, 并可以进行连续培养(
张玲华和郭勇, 2006 ).但诱导子通常会影响细胞的正常生长.有****在红豆杉(
Taxus chinensis )细胞培养过程中首先添加壳聚糖诱导子, 使其产生适应, 这样就保证了细胞的生存力和渗透性; 然后加入Ag
+ 和茉莉酸甲酯2种诱导子, 结果表明已经适应壳聚糖的细胞中紫杉醇含量比未经过这一适应过程的细胞中紫杉醇含量高4.6倍, 释放到胞外的能力也提高3.2倍(
Zhang et al., 2007 ). ...


{kind=link}
{kind=link}