
西南大学生命科学学院, 教育部三峡库区生态环境重点实验室, 重庆 400715
引用本文
贡献者
基金资助
接受日期:2016-10-20接受日期:2017-03-18网络出版日期:2017-11-1
-->Copyright
2017《植物学报》编辑部
Contributors
History
Received:Accepted:Online:
摘要:
Abstract:
Key words:
亲免素(immunophilin)在生物体内广泛存在, 可与免疫抑制剂他克莫司(tacrolimus, FK506)、雷帕霉素(rapamycin)及环孢素A (cyclosporin A)结合, 是含量比较丰富的一类细胞受体蛋白。根据与亲免素结合的免疫抑制剂不同, 可将其分为3类: 能与FK506和雷帕霉素特异结合的FK506结合蛋白(FK506 binding proteins, FKBPs)、能与环孢素A结合的亲环素(cyclophilins, CYPs)以及能与5-羟基-1,4-萘醌结合的parvulin蛋白(Hennig et al., 1998; He et al., 2004)。亲免素具有分子伴侣(molecular chaperone)的功能, 能够指导含有脯氨酸残基的蛋白质正确折叠、组装和转运。亲免素的这种功能主要依赖于其具有肽基脯氨酰顺反异构酶(peptidyl-prolyl cis-trans isomerases, PPlase, EC 5.1.2.8)活性, 该酶能够催化肽基脯氨酸的顺反异构化, 对蛋白质的正确折叠、运输及活性调节具有重要作用(Barik et al., 2006)。亲免素在哺乳动物体内还能作为免疫抑制剂药物的受体, 亲免素与免疫抑制剂药物形成的复合体能够抑制钙调磷酸酶(calcineurin, CaN)或者蛋白激酶TOR (target of rapamycin)活性, 抑制下游信号传递或细胞周期的正常进行, 从而达到免疫抑制的作用(Geisler and Bailly, 2007; 于彦丽等, 2014)。
从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(Ahn et al., 2010; Gollan and Bhave, 2010)。同时, 越来越多的CYPs也已被成功鉴定, 如拟南芥有29个、水稻有27个、大豆(Glycine max)有62个、酵母有8个、人类有24个(Ahn et al., 2010; Trivedi et al., 2012; Mainali et al., 2014)。目前已发现的parvulin蛋白拟南芥有3个、苹果(Malus domestica)有1个、毛地黄(Digitalis lanata)有1个、百脉根(Lotus japonicus)有3个、棉花(Gossypium hirsutum)有1个、酵母有1个、人类有3个(Arevalo-Rodriguez et al., 2004; Wang et al., 2014; Pai? and Fulgosi, 2016; Dunyak and Gestwicki, 2016)。植物中的亲免素数量众多, 可能是植物在进化过程中适应不同环境的结果。最新研究表明, 亲免素参与生长素与油菜素内酯的运输和信号传递过程, 对植物的生长发育和逆境胁迫响应等都具有重要作用(Geisler et al., 2003; Zhao et al., 2016; Chaiwanon et al., 2016)。本文主要对植物亲免素结构和功能的最新研究进展进行综述, 以期为进一步研究该类蛋白质的功能和作用机制提供参考。
1 亲免素的结构高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (Barik, 2006; Geisler and Bailly, 2007)。有些植物的FKBPs含有3个FKBD, 例如拟南芥FKBP62和FKBP65, 只有第1个FKBD具有PPlase活性, 与人类的FKBP51和FKBP52相似, 只有前2个FKBD具有活性(表1) (Vasudevan et al., 2015)。TPR结构域通常形成1个α螺旋区域, 提供与HSP90相互作用位点。CaM-BDs通常在FKBPs的C末端与CaM结合, 尽管哺乳动物体内CaM与FKBP结合具有指导HSP90与TPRs相互作用, 以及促进底物与FKBD结合的功能, 但植物体内FKBPs与钙调蛋白结合的功能还不明确(Schiene-Fischer, 2015)。高等植物中亲环素也分为2类: 一类是单结构域CYPs, 只含有保守结构域CYPD (CYP domain), 具有PPlase活性; 另一类是具有多个结构域的CYPs, 除了具有PPlase活性, 还含有WD40 (WD40 domain)、TPR (tetratricopep- tide repeat)、U-box结构域(U-box domain)、锌指结构域(Zn-finger domain)、富含亮氨酸的结构域(Leu- rich)、RNA识别模体(RNA recognition motif, RRM)和螺旋束结构域(helical bundle domain) (He et al., 2004)。真核生物中的RRM能调节转录后基因的表达、蛋白-蛋白相互作用和形态建立, 人体中的RRM参与构成剪接体且能与mRNA结合(Maris et al., 2005; Wang et al., 2008; Schiene-Fischer, 2015)。植物体内CYPs参与调节基因的表达、热胁迫响应和叶绿体内复杂蛋白质的合成等(表1)。根据植物体内parvulin蛋白的底物特异性可将其分为依赖磷酸化(Pin1-type)和不依赖磷酸化(non-Pin1-type) 2类。目前发现parvulin蛋白参与植物的激素运输和胁迫响应(表1)。
表1
Table 1
表1
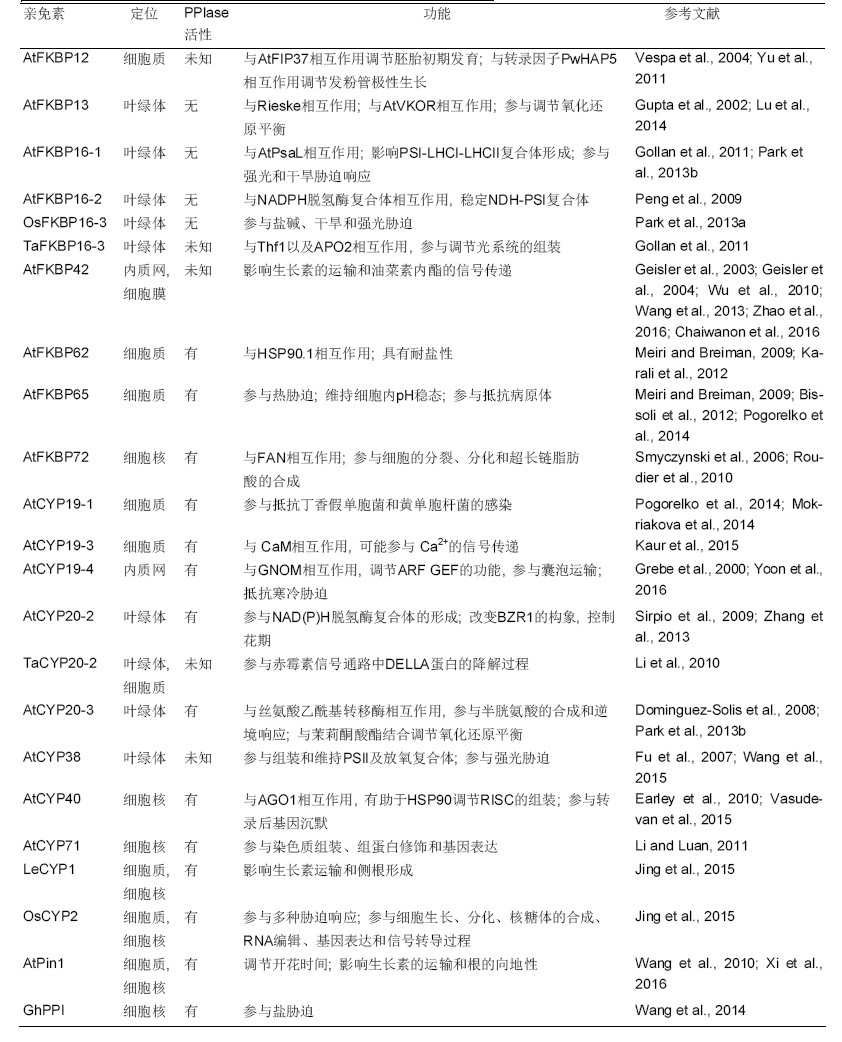 |
表1
部分植物亲免素在细胞内的定位及功能
Table 1
The localization and function of some immunophilins in plant cells
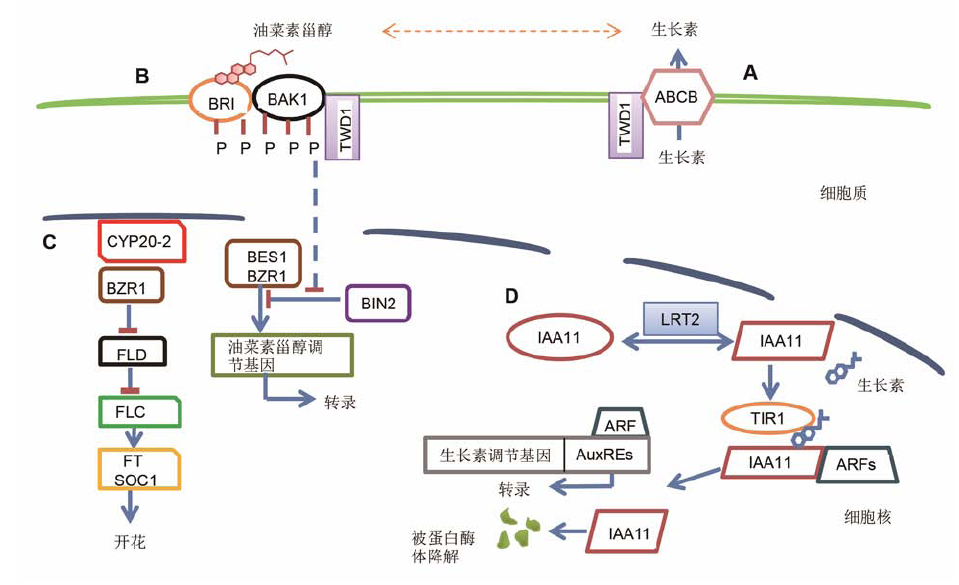
2 亲免素与植物生长发育2.1 FKBPs与植物生长发育植物的生长发育与激素密不可分。亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用。拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布。拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输。TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(图1) (Geisler et al., 2003; Wu et al., 2010; Wang et al., 2013)。目前报道TWD1定位在细胞膜、液泡和内质网, 其定位的多样性意味着功能的多样性(Kamphausen et al., 2002; Geisler et al., 2003; Wu et al., 2010)。近期研究表明, TWD1影响下胚轴表皮细胞内肌动蛋白束和肌动蛋白丝的动态平衡; ACTIN7是ABCB1-TWD1复合体的一部分, 能够调节ABCB、PIN和TWD1的表达和定位; 当TWD1突变后, 生长素运输和细胞骨架受到影响, 从而导致twd1突变体严重扭曲矮化的表型(Zhu and Geisler, 2015; Zhu et al., 2016)。
图1
Figure 1
下载原图ZIP
生成PPT
图1
亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(Geisler and Bailly, 2007; Wu et al., 2010); (B) 在BR的作用下, TWD1与BRI1和BAK1相互作用促进其磷酸化, 抑制BIN2的活性,影响BES1/BZR1的磷酸化, 从而调节BR响应基因的表达(Zhao et al., 2016; Chaiwanon et al., 2016); (C) AtCYP20-2与BZR1相互作用改变BZR1的构象, 调节FLD的表达, 改变开花时间(Zhang et al., 2013); (D) 在有生长素时, LRT2与OsIAA11相互作用, 使OsIAA11构象发生变化, 促进其与OsTIR1相互作用, 形成OsTIR1-OsIAA11复合体。在OsTIR1-IAA11复合体的帮助下泛素分子连接到OsIAA11上, 然后OsIAA11蛋白被26S蛋白酶体降解, ARFs形成有活性的形式, 从而激活生长素调节基因的表达(Jing et al.,2015)。
Figure 1
Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (Geisler and Bailly, 2007; Wu et al., 2010); (B) TWD1 physically interacts with BRI1 and BAK1 in a BR-independent manner, which may facilitate the phosphorylation of BRI1 and BAK1, inhibit BIN2 activity,and affect BES1/BZR1 phosphorylation, eventually regulate the BR response gene expression (Zhao et al., 2016; Chaiwanon et al., 2016); (C) A conformational change of BZR1 mediated by AtCYP20-2 causes altered flowering time through the modulation of FLD expression (Zhang et al., 2013); (D) In the presence of auxin, OsIAA11 conformation was changed by interacting with LRT2, and then formed the OsTIR1-OsIAA11 complex. OsTIR1-OsIAA11 complex facilitated OsIAA11 binding to 26S proteasome and caused OsIAA11 degradation; Finally, ARFs became active and activated auxin-regulated genes (Jing et al., 2015).
最新研究显示, 拟南芥TWD1也参与油菜素甾醇(brassinosteroid, BR)的信号传递。在根生长和下胚轴伸长过程中, twd1-2以及twd1-5对油菜素内酯(brassinolide, BL)的敏感程度降低; 在twd1-5中磷酸化和未磷酸化的BES1含量均降低。实验结果显示TWD1在植物体内能与BRI1和BAK1相互作用。以上表明TWD1与BRI1和BAK1相互作用影响了BR信号转导(图1) (Zhao et al., 2016; Chaiwanon et al., 2016)。
在哺乳动物中, FKBPs与转录因子相互作用较为常见, 在植物体内也发现FKBPs与转录因子相互作用调节细胞的分裂与分化。拟南芥突变体pas1 (pasticcion1)的细胞分裂异常、分化紊乱, 对野生型植株施加外源性细胞分裂素后出现与pas1相似的表型。研究发现, PAS1 (即FKBP72)的C末端与FAN转录因子(FKBP-associated NAC)相互作用, 促进了FAN对细胞分裂的抑制(Harrar et al., 2003; Smyczynski et al., 2006)。另一方面, PAS1参与超长链脂肪酸(very- long-chain fatty acids, VLCFAs)的合成。PAS1的功能缺失后, VLCFAs在拟南芥体内的含量下降, 影响鞘脂类物质合成, 使细胞不能正常分裂分化(Roudier et al., 2010)。
2.2 CYPs与植物生长发育拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育。早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(Jackson and S?ll, 1999)。最近的研究显示, CYP19-4s (AtCYP19-4和OsCYP19-4)能与 AtPP2A相互作用, OsCYP19-4过量表达植株与PIN1 RNAi植株表型相似, 说明Os- CYP19-4可能影响生长素的极性运输和PIN的定位(Yoon et al., 2016)。水稻lrt2 (lateral rootless 2, 即cyp2)突变体的表型与生长素突变体表型相似, 侧根发育不良、分蘖数和植株高度减少、花序发育受损, 表明LRT2参与生长素的信号通路并且在调节侧根发育方面具有重要作用(Kang et al., 2013; Zheng et al., 2013)。另有研究表明, 在生长素存在时, LRT2使OsIAA11构象发生变化, 促进其与TIR1 (transport inhibitor response 1)相互作用形成OsTIR1-IAA11复合体, 在OsTIR1-IAA11复合体的帮助下, 泛素分子连接到OsIAA11上, 然后OsIAA11蛋白被26S蛋白酶体降解, ARFs (auxin response element)形成有活性的形式, 从而激活生长素调节基因的表达(图1) (Jing et al., 2015)。最近的研究表明, 番茄(Lycopersicon esculentum)的DGT (LeCYP1)可以通过筛管运输, 将野生型嫁接到dgt突变体, 可使突变体的表型得到一定程度的恢复; LeCYP1具有提高植株对生长素的响应能力和调节根冠比的作用(Spiegelman et al., 2015)。另外, DGT能够影响根部和茎部生长素的运输, 控制侧根的形成; 在酵母和烟草(Nicotiana benthamiana)系统中, DGT通过影响生长素转运蛋白PIN在细胞膜上的定位调节生长素的输出(Ivanchenko et al., 2015)。CYPs主要通过调节生长素转运蛋白的定位和生长素调节基因的表达, 影响生长素的运输和生长素信号通路, 进而调控植物的生长发育。
在拟南芥开花信号途径中, FLC (FLOWERING LOCUS C)通过抑制成花信号FT (FLOWERING LO- CUS T)和转录因子基因SOC1 (SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1)的表达抑制开花。研究发现, 拟南芥AtCYP20-2与BZR1相互作用改变BZR1的构象, 调节FLD (FLOWERING LOCUS D)的表达; 过量表达CYP20-2使FLD表达量增加, 而FLD能抑制FLC染色质的甲基化和乙酰化, 从而导致开花提前(图1); 而过量表达小麦CYP20-2后开花延迟, 其原因可能是TaCYP20-2参与赤霉素信号通路中DELLA蛋白的降解过程(Li et al., 2010; Zhang et al., 2013)。
拟南芥ROC1 (AtCYP18-3)与光敏色素、隐花色素共同参与幼苗的去黄化过程。突变体roc1和roc1-D能够改变转录因子BES1的磷酸化模式, 但ROC1不能与BES1相互作用; CYP18-3的作用机制可能与TWD1相似, 通过与BRI1和BAK1相互作用影响BES1的磷酸化(Trupkin et al., 2012)。有趣的是, 在茎伸长过程中, 赤霉素与ROC1具有拮抗作用, At- CYP18-3也许不仅参与BR的信号通路, 还可能间接参与赤霉素的信号传递过程(Ma et al., 2013)。
2.3 Parvulins与植物生长发育目前, 关于parvulin蛋白的研究相对较少。研究表明, parvulin蛋白能调节植物的生长发育和抗逆性。拟南芥依赖磷酸化的AtPin1有1个PPlase结构域, 能够改变PIN1的构象, 使其极性定位发生改变, 进而影响生长素的运输和根的向地性; 另外, Pin1At与AGL24 (AGAMOUS-LIKE 24)和SOC1相互作用改变其构象, 调节开花时间(Wang et al., 2010; Xi et al., 2016)。棉花GhPPI在盐胁迫下表达量升高, 该蛋白是目前已报道的唯一与盐胁迫相关的parvulin蛋白(Wang et al., 2014)。
2.4 叶绿体内亲免素的功能拟南芥叶绿体定位的亲免素多达17种, 包括11种FKBPs、5种CYPs和1种parvulin蛋白。定位在叶绿体的亲免素中, 只有AtFKBP13和AtCYP20-2具有PPlase活性, 但是AtFKBP13和AtCYP20-2的双突变体无论是在正常条件下、寒冷胁迫下还是强光胁迫下与野生型相比都没有太大差异(Ingelsson et al., 2009)。由于其它定位在叶绿体的亲免素都没有PPl- ase活性, 因此定位在叶绿体内的亲免素的功能可能与PPlase活性无关(Ingelsson et al., 2009)。叶绿体是光合作用的场所, 定位在叶绿体的亲免素在植物光合作用中具有重要作用。
小麦TaFKBP16-1和TaFKBP16-3在光系统I (PSI)的组装过程中扮演着重要角色。TaFKBP16-1与PSI的1个亚基PsaL (PSI subunit L)相互作用, TaFK- BP16-3与Thf1 (Thylakoid Formation 1)和APO2 (Os02g50010)存在相互作用, 因此TaFKBP16-1和TaFKBP16-3可能调控PSI的组装(Gollan et al., 2011)。最近研究显示, AtFKBP16-1能够稳定PSI的亚基PsaL在细胞内的含量, 并且AtFKBP16-1影响LH- CII转移到PSI-LHCI上形成PSI-LHCI-LHCII复合体的过程, 说明FKBPs与PSI的组装和功能有着密切关系(Seok et al., 2014)。
拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用。拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(Lima et al., 2006)。拟南芥类囊体膜的氧化还原酶AtVKOR-DsbA定位于类囊体腔, 其参与氧化还原反应、催化二硫键的形成、维持细胞内ROS的平衡以及PSII的组装。酵母双杂交实验显示, AtFKBP20-2与AtVKOR存在相互作用, 暗示AtFKBP20-2可能参与PSII的组装(Lu et al., 2014)。Atcyp38突变体植株发育不良, 对光敏感, 不能正确组装PSII超级复合体; 并且AtCYP38也参与放氧复合体的正确组装(Fu et al., 2007; Sirpio et al., 2008; Vasudevande et al., 2012)。研究表明, 菠菜(Spin- acia oleracea) TLP40是拟南芥AtCYP38的同源蛋白, 能够调节PSII系统中磷酸蛋白酶的活性。不过, TLP40具有PPlase活性, 而AtCYP38不具有PPlase活性, 具体原因需要进一步研究(Rokka et al., 2000)。
NADPH脱氢酶NDH复合体能够调节PSI的循环和叶绿体内的电子传递, NDH与PSI形成NDH-PSI复合体, AtFKBP16-2能与NADPH脱氢酶复合体相互作用, 从而稳定NDH-PSI复合体(Peng et al., 2009)。AtCYP20-2是定位于类囊体腔且具有PPlase活性的亲环素, 其基因表达受光调控。最初报道AtCYP20-2与PSII的组装有关, 但是cyp20-2在有光和无光条件下均能生长, 且其PSII均无异常。后续研究表明, At- CYP20-2与NAD(P)H复合体的形成有关, 其在NDH疏水结构域的形成中起作用, 并且在NAD(P)H脱氢酶复合体的形成过程中作为重要的分子伴侣发挥作用(Sirpio et al., 2009)。
2.5 亲免素调节基因表达基因沉默、表观遗传学调控和转录水平的调控对植物的生长发育产生重要影响。拟南芥cyp40突变体叶片数量减少和形态改变与microRNA活性减弱有关。成熟的microRNA能够与RNA诱导沉默复合体(RNA- induced silencing complex, RISC)结合, 形成具有切割活性的RNA沉默复合物, 该复合物与靶基因互补配对使靶基因沉默或抑制靶基因的翻译。AGO1 (ARGONAUTE1)是RISC的重要组成部分(Voinnet, 2009; Earley et al., 2010)。AtCYP40与AGO1相互作用促进或维持AGO1的活性, 从而有助于HSP90调节RISC的组装, 影响基因沉默或基因的翻译(Earley et al., 2010; Vasudevan et al., 2015)。
拟南芥突变体cyp71茎尖分生组织活性下降, 侧生器官发育异常, 花的形态发生改变, 根的生长受抑制(Li et al., 2007)。AtCYP71定位于细胞核, 其N末端有4个重复结构域WD40, CYPD结构域在C末端, AtCYP71的重复结构域WD40与组蛋白H3相互作用, 影响H3K27的甲基化水平, 从而导致cyp71出现异常表型(Li et al., 2007)。进一步研究证明, AtCYP71能与FAS1和LHP1相互作用, 参与染色质组装和组蛋白修饰(Li and Luan, 2011)。其它细胞核定位的亲免素, 如AtFKBP43和AtFKBP53, 具有多个结构域, FK- BPD在C末端, N末端结构域具有高度可变性, 可能与核酸或者蛋白质相互作用(He et al., 2004)。At- FKBP53被证明在植物体内作为组蛋白的分子伴侣, 与组蛋白H3通过N末端的酸性结构域相互作用。有趣的是, PPlase活性对AtFKBP53的分子伴侣功能不重要, AtFKBP53能够抑制核糖体RNA基因的表达, 具有使染色质重塑和调节转录的功能(Li and Luan, 2010)。
3 亲免素对非生物及生物胁迫的响应3.1 亲免素对非生物胁迫的响应FKBPs对热胁迫、盐胁迫、干旱及强光胁迫都有响应。拟南芥中2个高度同源的基因FKBP62 (AtROF1)与FKBP65 (AtROF2)参与植物耐热性调控。但是在热胁迫下, 2个基因的表达量和调节方式不同, AtROF1促进热激蛋白的表达而AtROF2抑制热激蛋白的表达(Aviezer-Hagai et al., 2007; Meiri and Breiman, 2009)。AtROF2二级结构主要是β折叠、无规则卷曲及转角等, 仅含少量的α螺旋; 不同温度下AtROF2的二级结构呈2种形式, 当温度达到65°C时, AtROF2结构发生轻微变化以适应高温, 直到温度达到97°C该结构仍然稳定。AtROF2在高温下结构稳定的特性, 可能在研究植物耐热性方面具有重要作用(Lighezan et al., 2013)。在盐胁迫下, 过量表达AtROF1的拟南芥种子萌发率提高, AtROF1的FKBD可以与作为第二信使传递信号的磷脂酰肌醇-3-磷酸和磷脂酰肌醇- 3,5-二磷酸相互作用。因此, AtROF1可能通过与其相互作用影响细胞内的信号传递, 激活植物对逆境的响应。另外, ROF2还能调节细胞内的酸碱性, 植物体过量表达ROF2能提高对有毒阳离子的耐受性(Karali et al., 2012; Bissoli et al., 2012)。
在盐碱、干旱和强光等逆境胁迫下, 水稻Os- FKBP16-3的表达量增加, 将OsFKBP16-3转化拟南芥, 转基因植株对盐碱、干旱和强光的耐受性提高(Park et al., 2013a)。在极端环境下, 植物体内会产生活性氧簇(ROS)。FKBP16-3定位在叶绿体内, 其含有一段保守的氧化还原位点CxxxC, 能被ROS氧化形成二硫键; FKBP16-3可能通过修饰CxxxC调节氧化还原过程, 以减少逆境对植物的伤害(Park et al., 2013a)。另外, ROS在环境胁迫或者生物胁迫下可以作为信号分子通过诱导特殊的激酶激活MAPK通路。因此, 亲免素也可能通过调控ROS的产生激活MAPK通路参与植物对胁迫的响应(Pogorelko et al., 2014)。
CYPs在植物抵抗盐碱、干旱、寒冷和强光胁迫方面具有重要作用, 并且能够增加水稻分蘖, 从而提高产量。过量表达OsCYP2可以提高水稻种子的耐盐性, 并且OsCYP2可能通过调节抗氧化酶活性控制细胞内ROS的含量, 从而提高植株的耐盐性(Ruan et al., 2011)。OsCYP18-2与OsSKIP (ski-interacting protein)相互作用, 使OsCYP18-2的定位从细胞质转移到细胞核, OsCYP18-2-OsSKIP复合体可能参与mRNA的转录和mRNA前体剪接或稳定性等; 并且过量表达OsCYP18-2能提高水稻的耐旱性, 原因可能是OsCYP18-2-OsSKIP复合体在细胞核中调节与抗逆相关的基因表达(Lee et al., 2015)。内质网定位的OsCYP19-4具有PPlase活性, 在水稻中过量表达OsCYP19-4能够增强其耐寒性, 并且显著提高水稻的分蘖和穗的数量, 从而使产量提高(Yoon et al., 2016)。Atcyp38突变体光合作用受到一定程度的抑制。研究显示, CYP38参与强光胁迫响应, 突变体cyp38在强光下PsbO2表达量增加, 导致D1蛋白降解, ROS产量增多; 野生型在强光下细胞质内Ca2+和CaM3含量增多以促进CYP38的表达, CYP38能够抑制PsbO2的活性, 减少D1蛋白的降解, 从而降低ROS以及强光对植物体的伤害(Wang et al., 2015)。
叶绿体内半胱氨酸合成过程中的重要酶(丝氨酸乙酰基转移酶(SAT1))能与AtCYP20-3相互作用, 指导SAT1的正确折叠, 保证半胱氨酸合成通路, 以适应各种逆境(Dominguez-Solis et al., 2008)。植物激素中茉莉酮酸酯家族对植物的生长发育和胁迫响应具有重要作用。在胁迫响应过程中, AtCYP20-3能与植物激素茉莉酮酸酯结合调节细胞内氧化还原平衡(Park et al., 2013b)。研究表明, AtCYP19-3具有PP- lase活性, 并且对 Cu2+敏感, 而 Cu2+能与巯基反应, 这意味着PPlase可能参与调节氧化还原反应(Kaur et al., 2015)。无论体内还是体外, AtCYP19-3的N末端都能与CaM相互作用, 并且这种相互作用依赖Ca2+, 但是CaM不影响AtCYP19-3的PPlase活性, 这可能意味着AtCYP19-3参与 Ca2+的信号传递过程(Kaur et al., 2015)。作为第二信使的 Ca2+对植物响应生物和非生物胁迫具有重要作用, 而AtCYP19-3能与CaM相互作用影响Ca2+, 说明AtCYP19-3可能从上游调节植物对各种胁迫的响应。
3.2 亲免素对生物胁迫的响应生物胁迫对作物生产具有严重危害。研究发现, 病原体入侵时, AtCYP19、AtCYP57和AtFKBP65的表达量显著提高。AtCYP19、AtCYP57和AtFKBP65基因的缺失突变体更易感染丁香假单胞菌和黄单胞杆菌, 突变导致对其病原体表面保守分子特征(pathogen associated molecular patterns, PAMPs)的识别有缺陷; 过量表达AtCYP19、AtCYP57和AtFKBP65的拟南芥对丁香假单胞菌的抵抗力增强; 并且AtCYP19、AtCYP57和AtFKBP65能够激活病原体相关分子模式触发免疫(PAMPs triggered immunity, PTI), 使胼胝质沉淀和过氧化氢积累以抵抗病原体(Pogorelko et al., 2014; Mokriakova et al., 2014)。
然而, 最近研究发现, AtCYP18-3 (ROC1)在植物免疫系统中具有负调控作用, ROC1能与效应子触发免疫(effector triggered immunity, ETI)的受体RIN4相互作用, 减少RIN4的磷酸化, 使抗性基因RPM1不能够识别效应因子, 从而不能激活ETI途径; 而大豆CYP1能激活疫霉属效应因子Avr3b的酶活性。这说明亲免素参与生物胁迫的方式较多, 其作用机制有待进一步研究(Li et al., 2014; Kong et al., 2015)。
4 展望近年来的研究表明, FKBPs与多种激素关系密切。拟南芥中调节生长素运输的TWD1与人类FKBP38结构相似, 人类FKBP38的PPlase活性能够促进囊性纤维化跨膜传导调节蛋白(CFTR)的翻译后折叠, 并且其TPR结构域与Hsp90相互作用能够抑制PPlase活性, 对CFTR的合成进行负反馈调节(Banasavadi-Sid- degowda et al., 2011)。TWD1调节ABCBs的方式是否与人类的FKBP38相似, 即作为分子伴侣指导ABCBs转运蛋白的折叠和运输还有待证明。动物的FKBP51和FKBP52作为甾醇类激素受体的分子伴侣, 能够与HSP90形成复合体, 帮助受体的正确定位以及激素与受体的结合(Sivils et al., 2011)。同样, 拟南芥TWD1参与油菜素甾醇的信号传递途径, 但不同的是, TWD1通过与磷酸激酶BRI1和BAK1相互作用调节对BR的响应(Zhao et al., 2016; Chaiwanon et al., 2016)。这在一定程度上说明, 拟南芥TWD1与动物FKBP52在功能上有相似性但在执行功能的方式上有所不同。另外, TWD1与BRI1和BAK1的哪个结构域相互作用, 是否与PPlase活性有关, 也需要进一步验证。
CYPs参与生长素、油菜素内酯、赤霉素和茉莉酸等激素的信号转导途径, 其中生长素、赤霉素和茉莉酸均以蛋白酶体降解抑制子的方式激活信号通路(Jing et al., 2015)。有趣的是, TaCYP20-2与赤霉素的信号抑制子Rht/DELLA蛋白的降解有关; OsLRT2/ CYP2参与生长素抑制蛋白Aux/IAA的降解; 表明亲环素在调节激素信号通路过程中具有重要作用(Li et al., 2015; Jing et al., 2015)。
在哺乳动物中, 亲免素的主要功能是与小的配体结合(如免疫抑制剂), 或者作为分子伴侣调节蛋白质的结构、活性和稳定性。在高等植物中, 亲免素通过与相关蛋白质相互作用调控生长发育; 并且FBKPs对免疫抑制剂不敏感, PPlase活性也不保守; 而亲环素对免疫抑制剂敏感, PPlase活性也较为保守。例如, AtROC1、LeCYP1和OsCYP2都具有PPlase活性(表1)。高等植物中亲免素对免疫抑制剂的不同敏感性, 可能是由于在进化中丢失了PPIase活性所致。迄今为止, 关于parvulin蛋白的研究相对较少。有研究将parvulin蛋白归类于不同于亲免素的PPlase, 主要是由于它不能被FK506、雷帕霉素和环孢素A等免疫抑制剂抑制(Rahfeld et al., 1994; He et al., 2004; Dunyak and Gestwicki, 2016)。最新研究发现, 拟南芥parvulin蛋白Pin1At能够改变PIN1的构象, 影响PIN1的极性定位, 进而影响生长素的运输和根的向地性; LeDGT/CYP1具有PPlase活性, 也影响PIN在细胞膜上的定位和生长素的运输; 但PIN蛋白的正确定位与PPlase活性的关系还需要进一步验证(Ivan- chenko et al., 2015; Xi et al., 2016)。
植物体内的亲免素能够对环境胁迫做出响应。例如, 它通过与热休克蛋白、磷脂酰肌醇、钙调蛋白的相互作用, 激活某些信号通路、参与氨基酸(如半胱氨酸)合成等响应环境胁迫。叶绿体内的亲免素与光合膜的组装和氧化还原过程等有关, 但是亲免素在叶绿体内的功能对胁迫响应有何影响需进一步探明。亲免素在抵抗外界胁迫及在叶绿体中的功能已在农作物中进行了相关研究, 并且在水稻耐盐增产和小麦抗旱等方面取得了一定的进展。因此, 亲免素在农作物改良和分子育种等方面有着广阔的应用前景。
参考文献
文献选项
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | [本文引用: 1] |
| [2] | [本文引用: 2] |
| [3] | DOI:10.2741/1405PMID:15353296URLFront Biosci. 2004 Sep 1;9:2420-46. Research Support, N.I.H., Extramural; Research Support, Non-U.S. Gov't; Research Support, U.S. Gov't, P.H.S.; Review [本文引用: 1] |
| [4] | DOI:10.1007/s11103-006-9085-zURL [本文引用: 1] |
| [5] | DOI:10.1074/jbc.M111.269993PMID:3234856URLFK506-binding protein 38 (FKBP38), a membrane-anchored, tetratricopeptide repeat (TPR)-containing immunophilin, associates with nascent plasma membrane ion channels in the endoplasmic reticulum (ER). It promotes the maturation of the human ether- -go-go-related gene (HERG) potassium channel and maintains the steady state level of the cystic fibrosis transmembrane conductance regulator (CFTR), but the underlying mechanisms remain unclear. Using a combination of steady state and pulse-chase analyses, we show that FKBP38 knockdown increases protein synthesis but inhibits the post-translational folding of CFTR, leading to reduced steady state levels of CFTR in the ER, decreased processing, and impaired cell surface functional expression in Calu-3 human airway epithelial cells. The membrane anchorage of FKBP38 is necessary for the inhibition of protein synthesis but not for CFTR post-translational folding. In contrast, the peptidylprolyl cis/trans isomerase active site is utilized to promote CFTR post-translational folding but is not important for regulation of protein synthesis. Uncoupling FKBP38 from Hsp90 by substituting a conserved lysine in the TPR domain modestly enhances CFTR maturation and further reduces its synthesis. Removing the N-terminal glutamate-rich domain (ERD) slightly enhances CFTR synthesis but reduces its maturation, suggesting that the ERD contributes to FKBP38 biological activities. Our data support a dual role for FKBP38 in regulating CFTR synthesis and post-translational folding. In contrast to earlier prediction but consistent with in vitro enzymological studies, FKBP38 peptidylprolyl cis/trans isomerase plays an important role in membrane protein biogenesis on the cytoplasmic side of the ER membrane, whose activity is negatively regulated by Hsp90 through the TPR domain. [本文引用: 1] |
| [6] | DOI:10.1007/s00018-006-6215-3PMID:17075696URLAbstract Immunophilins are chaperones that may also exhibit peptidylprolyl isomerase (PPIase) activity. This review summarizes our knowledge of the two largest families of immunophilins, namely cyclophilin and FK506-binding protein, and a novel chimeric dual-family immunophilin, named FK506- and cyclosporin-binding protein (FCBP). The larger members of each family are modular in nature, consisting of multiple PPIase and/or protein-protein interaction domains. Despite the apparent difference in their sequence and three-dimensional structure, the three families encode similar enzymatic and biological functions. Recent studies have revealed that many immunophilins possess a chaperone function independent of PPIase activity. Knockout animal studies have confirmed multiple essential roles of immunophilins in physiology and development. An immunophilin is indeed a natural 'protein-philin' (Greek 'philin' = friend) that interacts with proteins to guide their proper folding and assembly. [本文引用: 2] |
| [7] | [本文引用: 1] |
| [8] | DOI:10.1016/j.molp.2016.01.008PMID:26808213URLMutation of the immunophilin-like FK506-binding protein TWISTED DWARF1 (FKBP42/TWD1) causes dwarf and twisted-organ phenotypes inArabidopsis. However, the function of FKBP42 is not fully understood at the molecular level. Using genetic, physiological and immunological experiments, we show here that FKBP42 is necessary for brassinosteroid (BR) signal transduction. Thetwd1mutant showed reduced BR sensitivity in growth responses and activation of the BZR1 transcription factor. However,twd1showed normal responses to an inhibitor of BIN2/GSK3, suggesting thattwd1has defect upstream of BIN2 in the BR signaling pathway.In vitroandin vivoassays showed that TWD1 interacts physically with the kinase domains of the BR receptor kinases BRI1 and BAK1. TWD1 is not required for normal localization of BRI1-GFP to the plasma membrane or for activation of the flagellin receptor kinase FLS2. Our results suggest that FKBP42/TWD1 plays a specific role in the activation of BRI1 receptor kinase. [本文引用: 5] |
| [9] | DOI:10.1073/pnas.0808204105URLCyclophilins belong to a large family of enzymes called "peptidyl prolyl isomerases" that assist protein folding and assembly. The cyclophilin CYP20-3 (also known as "ROC4") is the only member of this group located in the stroma (soluble phase) of chloroplasts. In the present study we isolated mutant Arabidopsis plants defective in the CYP20-3 gene and found them to be hypersensitive to oxidative stress conditions created by high light levels, rose bengal, high salt levels, and osmotic shock. Chloroplast serine acetyltransferase (SAT1), a rate-limiting enzyme in cysteine biosynthesis, was identified as an interacting partner for CYP20-3 by protein interaction analyses. In the present experiments, SAT1 activity increased significantly under conditions of light and oxidative stress in concert with total thiols in wild-type plants. By contrast, these parameters changed only marginally in experiments with the cyp20-3 mutant, suggesting that CYP20-3 links light and stress to SAT1 activity and cysteine biosynthesis. In further support of this conclusion, our analyses showed that the salt-hypersensitive phenotype of the mutant developed under illumination and not in the dark. Together with the earlier report that CYP20-3 foldase activity is enhanced by thioredoxin-mediated reduction, our findings suggest that CYP20-3 links photosynthetic electron transport and redox regulation to the folding of SAT1, thereby enabling the cysteine-based thiol biosynthesis pathway to adjust to light and stress conditions. [本文引用: 1] |
| [10] | DOI:10.1021/acs.jmedchem.6b00411PMID:27409354URLPeptidyl-proline isomerases (PPIases) are a chaperone superfamily comprising the FK506-binding proteins (FKBPs), cyclophilins and parvulins. PPIases catalyze the cis/trans isomerization of proline, acting as a regulatory switch during folding, activation and/or degradation of many proteins. These “clients” include proteins with key roles in cancer, neurodegeneration and psychiatric disorders, suggesting that PPIase inhibitors could be important therapeutics. However, the active site of PPIases is shallow, solvent-exposed and well conserved between family members, making selective inhibitor design challenging. Despite these hurdles, macrocyclic natural products, including FK506, rapamycin and cyclosporin, bind PPIases with nanomolar or better affinity. De novo attempts to derive new classes of inhibitors have been somewhat less successful, often showcasing the “undruggable” features of PPIases. Interestingly, the most potent of these next-generation molecules tend to integrate features of the natural produ... [本文引用: 2] |
| [11] | [本文引用: 2] |
| [12] | DOI:10.1073/pnas.0707851104PMID:17909185URLPhotosynthetic light reactions rely on the proper function of large protein complexes (including photosystems I and II) that reside in the thylakoid membrane. Although their composition, structure, and function are known, the repertoire of assembly and maintenance factors is still being determined. Here we show that an immunophilin of the cyclophilin type, CYP38, plays a critical role in the assembly and maintenance of photosystem II (PSII) supercomplexes (SCs) in Arabidopsis. Mutant plants with the CYP38 gene interrupted by T-DNA insertion showed stunted growth and were hypersensitive to high light. Leaf chlorophyll fluorescence analysis and thylakoid membrane composition indicated that cyp38 mutant plants had defects in PSII SCs. Sucrose supplementation enabled the rescue of the mutant phenotype under low-light conditions, but failed to mitigate hypersensitivity to high-light stress. Protein radiolabeling assays showed that, although individual thylakoid proteins were synthesized equally in mutant and wild type, the assembly of the PSII SC was impaired in the mutant. In addition, the D1 and D2 components of the mutant PSII had a short half-life under high-light stress. The results provide evidence that CYP38 is necessary for the assembly and stabilization of PSII. [本文引用: 1] |
| [13] | DOI:10.1016/j.tplants.2007.08.015URL [本文引用: 4] |
| [14] | DOI:10.1091/mbc.E03-11-0831PMID:15133126URLAbstract Previously, the immunophilin-like protein TWD1 from Arabidopsis has been demonstrated to interact with the ABC transporters AtPGP1 and its closest homologue, AtPGP19. Physiological and biochemical investigation of pgp1/pgp19 and of twd1 plants suggested a regulatory role of TWD1 on AtPGP1/AtPGP19 transport activities. To further understand the dramatic pleiotropic phenotype that is caused by loss-of-function mutation of the TWD1 gene, we were interested in other TWD1 interacting proteins. AtMRP1, a multidrug resistance-associated (MRP/ABCC)-like ABC transporter, has been isolated in a yeast two-hybrid screen. We demonstrate molecular interaction between TWD1 and ABC transporters AtMRP1 and its closest homologue, AtMRP2. Unlike AtPGP1, AtMRP1 binds to the C-terminal tetratricopeptide repeat domain of TWD1, which is well known to mediate protein-protein interactions. Domain mapping proved that TWD1 binds to a motif of AtMRP1 that resembles calmodulin-binding motifs; and calmodulin binding to the C-terminus of MRP1 was verified. By membrane fractionation and GFP-tagging, we localized AtMRP1 to the central vacuolar membrane and the TWD1-AtMRP1 complex was verified in vivo by coimmunoprecipitation. We were able to demonstrate that TWD1 binds to isolated vacuoles and has a significant impact on the uptake of metolachlor-GS and estradiol-beta-glucuronide, well-known substrates of vacuolar transporters AtMRP1 and AtMRP2. |
| [15] | DOI:10.1091/mbc.e02-10-0698URL [本文引用: 3] |
| [16] | DOI:10.1007/s11103-009-9547-1PMID:19768557URLThe FK506-binding proteins (FKBPs) are a class of peptidyl-prolyl cis / trans isomerase enzymes, some of which can also operate as molecular chaperones. FKBPs comprise a large ubiquitous family, found in virtually every part of the cell and involved in diverse processes from protein folding to stress response. Higher plant genomes typically encode about 20 FKBPs, half of these found in the chloroplast thylakoid lumen. Several FKBPs in plants are regulators of hormone signalling pathways, with important roles in seed germination, plant growth and stress response. Some FKBP isoforms exists as homologous duplicates operating in finely tuned mechanisms to cope with abiotic stress. In order to understand the roles of the plant FKBPs, especially in view of the warming environment, we have identified and analysed the gene families encoding these proteins in rice using computational approaches. The work has led to identification of all FKBPs from the rice genome, including novel high molecular weight forms. The rice FKBP family appears to have evolved by duplications of FKBP genes, which may be a strategy for increased stress tolerance. [本文引用: 1] |
| [17] | DOI:10.1111/j.1399-3054.2011.01503.xPMID:21848652URLFK506-binding proteins (FKBPs) and cyclophilins, collectively called immunophilins, conserve peptidyl-prolyl cis/trans isomerase (PPIase) active sites, although many lack PPIase activity. The chloroplast thylakoid contains a large proportion of the plant immunophilin family, but their functions within this compartment are unclear. Some lumenal immunophilins are important for assembly of photosynthetic complexes, implicating them in the maintenance and turnover of the photosynthetic apparatus during acclimation processes. In this investigation into the functions of three FKBPs localized to the thylakoid of Triticum aestivum (wheat), we present the first evidence of PPIase activity in the thylakoid of a cereal plant, and also show that PPIase activity is not conserved in all lumenal FKBPs. Using yeast two-hybrid analysis we found that the PPIase-active FKBP13 interacts with the globular domain of the wheat Rieske protein, with potential impact on photosynthetic electron transfer. Specific interaction partners for PPIase-deficient FKBP16-1 and FKBP16-3 link these isoforms to photosystem assembly. [本文引用: 1] |
| [18] | DOI:10.2307/3870940PMID:10715321URLThe Arabidopsis GNOM protein, a guanine nucleotide exchange factor (GEF) that acts on ADP ribosylation factor (ARF)-type G proteins, is required for coordination of cell polarity along the apical-basal embryo axis. Interallelic complementation of gnom mutants suggested that dimerization is involved in GNOM function. Here, direct interaction between GNOM molecules is demonstrated in vitro and by using a yeast two-hybrid system. Interaction was confined to an N-terminal domain conserved within a subgroup of large ARF GEFs. The same domain mediated in vitro binding to cyclophilin 5 (Cyp5), which was identified as a GNOM interactor in two-hybrid screening. Cyp5 displayed peptidylprolyl cis/trans-isomerase and protein refolding activities that were sensitive to cyclosporin A. Cyp5 protein accumulated in several plant organs and, like GNOM, was partitioned between cytosolic and membrane fractions. Cyp5 protein was also expressed in the developing embryo. Our results suggest that Cyp5 may regulate the ARF GEF function of the GNOM protein during embryogenesis. |
| [19] | DOI:10.1073/pnas.222550399PMID:12424338URLImmunophilins are intracellular receptors of the immunosuppressants cyclosporin A, FK506, and rapamycin. Although all immunophilins possess peptidyl-prolyl isomerase activity and are identified from a wide range of organisms, little is known about their cellular functions. We report the characterization and functional analysis of an FK506 and rapamycin-binding protein (AtFKBP13) from Arabidopsis. The AtFKBP13 protein is synthesized as a precursor that is imported into chloroplasts and processed to the mature form located in the thylakoid lumen, as shown by chloroplast import assays and Western blot analysis. Experiments show that AtFKBP13 is translocated across the thylakoid membrane by the 螖pH-dependent pathway. Yeast two-hybrid screening identified Rieske FeS protein, a subunit of the cytochrome bf complex in the photosynthetic electron transport chain, as an interacting partner for AtFKBP13. Both yeast two-hybrid and in vitro protein-protein interaction assays showed that the precursor, but not the mature form, of AtFKBP13 interacted with Rieske protein, suggesting that interaction between the two proteins occurs along the import pathway. When AtFKBP13 expression was suppressed by RNA interference method, the level of Rieske protein was significantly increased in the transgenic plants. |
| [20] | DOI:10.1104/pp.102.019026URLPASTICCINO (PAS) genes are required for coordinated cell division and differentiation during plant development. In loss-of-function pas mutants, plant aerial tissues showed ectopic cell division that was specifically enhanced by cytokinins, leading to disorganized tumor-like tissue. To determine the role of the PAS genes in controlling cell proliferation, we first analyzed the expression profiles of several genes involved in cell division and meristem function. Differentiated and meristematic cells of the pas mutants were more competent for cell division as illustrated by the ectopic and enlarged expression profiles of CYCLIN-DEPENDENT KINASE A and CYCLIN B1. The expression of meristematic homeobox genes KNOTTED-LIKE IN ARABIDOPSIS (KNAT2, KNAT6), and SHOOT MERISTEMLESS was also increased in pas mutants. Moreover, the loss of meristem function caused by shoot meristemless mutation can be suppressed by pas2. The KNAT2 expression pattern defines an enlarged meristematic zone in pas mutants that can be mimicked in wild type by cytokinin treatment. Cytokinin induction of the primary cytokinin response markers, ARABIDOPSIS RESPONSE REGULATOR (ARR5 and ARR6), was enhanced and lasted longer in pas mutants, suggesting that PAS genes in wild type repress cytokinin responses. The expression of the cytokinin-regulated cyclin D, cyclin D3.1, was nonetheless not modified in pas mutants. However, primary auxin response genes were down-regulated in pas mutants, as shown by a lower auxin induction of IAA4 and IAA1 genes, demonstrating that the auxin response was also modified. Altogether, our results suggest that PAS genes are involved in the hormonal control of cell division and differentiation. [本文引用: 1] |
| [21] | DOI:10.1104/pp.103.031005PMID:15047905URLImmunophilins are defined as receptors for immunosuppressive drugs including cyclosporin A, FK506, and rapamycin. The cyclosporin A receptors are referred to as cyclophilins (CYPs) and FK506- and rapamycin-binding proteins are abbreviated as FKBPs. These two groups of proteins (collectively called immunophilins) share little sequence homology, but both have peptidyl prolyl cis/trans isomerase (PPIase) activity that is involved in protein folding processes. Studies have identified immunophilins in all organisms examined including bacteria, fungi, animals, and plants. Nevertheless, the physiological function of immunophilins is poorly understood in any organism. In this study, we have surveyed the genes encoding immunophilins in Arabidopsis genome. A total of 52 genes have been found to encode putative immunophilins, among which 23 are putative FKBPs and 29 are putative CYPs. This is by far the largest immunophilin family identified in any organism. Both FKBPs and CYPs can be classified into single domain and multiple domain members. The single domain members contain a basic catalytic domain and some of them have signal sequences for targeting to a specific organelle. The multiple domain members contain not only the catalytic domain but also defined modules that are involved in protein-protein interaction or other functions. A striking feature of immunophilins in Arabidopsis is that a large fraction of FKBPs and CYPs are localized in the chloroplast, a possible explanation for why plants have a larger immunophilin family than animals. Parvulins represent another family of PPIases that are unrelated to immunophilins in protein sequences and drug binding properties. Three parvulin genes were found in Arabidopsis genome. The expression of many immunophilin and parvulin genes is ubiquitous except for those encoding chloroplast members that are often detected only in the green tissues. The large number of genes and diversity of structure domains and cellular localization make PPIases a versatile superfamily of proteins that clearly function in many cellular processes in plants. [本文引用: 4] |
| [22] | DOI:10.1021/bi973162pPMID:9558330URLAbstract In contrast to FK506 binding proteins and cyclophilins, the parvulin family of peptidyl-prolyl cis/trans isomerases (PPIases; E.C. 5.2.1.8) cannot be inhibited by either FK506 or cyclosporin A. We have found that juglone, 5-hydroxy-1,4-naphthoquinone, irreversibly inhibits the enzymatic activity of several parvulins, like the E. coli parvulin, the yeast Ess1/Ptf1, and human Pin1, in a specific manner, thus allowing selective inactivation of these enzymes in the presence of other PPIases. The mode of action was studied by analyzing the inactivation kinetics and the nature of products of the reaction of E. coli parvulin and its Cys69Ala variant with juglone. For all parvulins investigated, complete inactivation was obtained by a slow process that is characterized by pseudo-first-order rate constants in the range of 5.3 x 10(-)4 to 4. 5 x 10(-)3 s-1. The inactivated parvulin contains two juglone molecules that are covalently bound to the side chains of Cys41 and Cys69 because of a Michael addition of the thiol groups to juglone. Redox reactions did not contribute to the inactivation process. Because thiol group modification was shown to proceed 5-fold faster than the rate of enzyme inactivation, it was considered as a necessary but not sufficient condition for inactivation. When measured by far-UV circular dichroism (CD), the rate of structural alterations following thiol group modification parallels exactly the rate of inactivation. Thus, partial unfolding of the active site of the parvulins was thought to be the cause of the deterioration of PPIase activity. [本文引用: 1] |
| [23] | [本文引用: 2] |
| [24] | DOI:10.1242/dev.113225PMID:25617431URLCyclophilin A is a conserved peptidyl-prolyl cis-trans isomerase (PPIase) best known as the cellular receptor of the immunosuppressant cyclosporine A. Despite significant effort, evidence of developmental functions of cyclophilin A in non-plant systems has remained obscure. Mutations in a tomato (Solanum lycopersicum) cyclophilin A ortholog, DIAGEOTROPICA (DGT), have been shown to abolish the organogenesis of lateral roots; however, a mechanistic explanation of the phenotype is lacking. Here, we show that the dgt mutant lacks auxin maxima relevant to priming and specification of lateral root founder cells. DGT is expressed in shoot and root, and localizes to both the nucleus and cytoplasm during lateral root organogenesis. Mutation of ENTIRE/IAA9, a member of the auxin-responsive Aux/IAA protein family of transcriptional repressors, partially restores the inability of dgt to initiate lateral root primordia but not the primordia outgrowth. By comparison, grafting of a wild-type scion restores the process of lateral root formation, consistent with participation of a mobile signal. Antibodies do not detect movement of the DGT protein into the dgt rootstock; however, experiments with radiolabeled auxin and an auxin-specific microelectrode demonstrate abnormal auxin fluxes. Functional studies of DGT in heterologous yeast and tobacco-leaf auxin-transport systems demonstrate that DGT negatively regulates PIN-FORMED (PIN) auxin efflux transporters by affecting their plasma membrane localization. Studies in tomato support complex effects of the dgt mutation on PIN expression level, expression domain and plasma membrane localization. Our data demonstrate that DGT regulates auxin transport in lateral root formation. [本文引用: 2] |
| [25] | DOI:10.1007/s004380051147PMID:10628867URLThe heterotrimeric protein phosphatase 2A ( PP2A ) is a component of multiple signaling pathways in eukaryotes. Disruption of PP2A activity in Arabidopsis is known to alter auxin transport and growth response pathways. We demonstrated that the regulatory subunit A of an Arabidopsis PP2A interacts with a novel cyclophilin, ROC7. The gene for this cyclophilin encodes a protein that contains a unique 30-amino acid extension at the N-terminus, which distinguishes the gene product from all previously identified Arabidopsis cyclophilins. Altered forms of ROC7 cyclophilin with mutations in the conserved DENFKL domain did not bind to PP2A . Unlike protein phosphatase 2B, PP2A activity in Arabidopsis extracts was not affected by the presence of the cyclophilin-binding molecule cyclosporin. The ROC7 transcript was expressed to high levels in all tissues tested. Expression of an ROC7 antisense transcript gave rise to increased root growth. These results indicate that cyclophilin may have a role in regulating PP2A activity, by a mechanism that differs from that employed for cyclophilin regulation of PP2B . [本文引用: 1] |
| [26] | DOI:10.1038/ncomms8395URL [本文引用: 5] |
| [27] | [本文引用: 1] |
| [28] | DOI:10.1111/tpj.2013.74.issue-1URL [本文引用: 1] |
| [29] | DOI:10.1371/journal.pone.0048241PMID:23133621URLA direct interaction of theArabidopsis thalianaimmunophilin ROF1 with phosphatidylinositol-3-phosphate and phosphatidylinositol-3,5-bisphosphate was identified using a phosphatidylinositol-phosphate affinity chromatography of cell suspension extracts, combined with a mass spectrometry (nano LC ESI-MS/MS) analysis. The first FK506 binding domain was shown sufficient to bind to both phosphatidylinositol-phosphate stereoisomers. GFP-tagged ROF1 under the control of a 35S promoter was localised in the cytoplasm and the cell periphery ofNicotiana tabacumleaf explants. Immunofluorescence microscopy ofArabidopsis thalianaroot tips verified its cytoplasmic localization and membrane association and showed ROF1 localization in the elongation zone which was expanded to the meristematic zone in plants grown on high salt media. Endogenous ROF1 was shown to accumulate in response to high salt treatment inArabidopsis thalianayoung leaves as well as in seedlings germinated on high salt media (0.15 and 0.2 M NaCl) at both an mRNA and protein level. Plants over-expressing ROF1, (WSROF1OE), exhibited enhanced germination under salinity stress which was significantly reduced in therof161knock out mutants and abolished in the double mutants of ROF1 and of its interacting homologue ROF2 (WSrof161/261). Our results show that ROF1 plays an important role in the osmotic/salt stress responses of germinatingArabidopsis thalianaseedlings and suggest its involvement in salinity stress responses through a phosphatidylinositol-phosphate related protein quality control pathway. [本文引用: 1] |
| [30] | [本文引用: 2] |
| [31] | DOI:10.1371/journal.ppat.1005139PMID:26317500URLAbstract Plant pathogens secrete an arsenal of effector proteins to impair host immunity. Some effectors possess enzymatic activities that can modify their host targets. Previously, we demonstrated that a Phytophthora sojae RXLR effector Avr3b acts as a Nudix hydrolase when expressed in planta; and this enzymatic activity is required for full virulence of P. sojae strain P6497 in soybean (Glycine max). Interestingly, recombinant Avr3b produced by E. coli does not have the hydrolase activity unless it was incubated with plant protein extracts. Here, we report the activation of Avr3b by a prolyl-peptidyl isomerase (PPIase), cyclophilin, in plant cells. Avr3b directly interacts with soybean cyclophilin GmCYP1, which activates the hydrolase activity of Avr3b in a PPIase activity-dependent manner. Avr3b contains a putative Glycine-Proline (GP) motif; which is known to confer cyclophilin-binding in other protein substrates. Substitution of the Proline (P132) in the putative GP motif impaired the interaction of Avr3b with GmCYP1; as a result, the mutant Avr3bP132A can no longer be activated by GmCYP1, and is also unable to promote Phytophthora infection. Avr3b elicits hypersensitive response (HR) in soybean cultivars producing the resistance protein Rps3b, but Avr3bP132A lost its ability to trigger HR. Furthermore, silencing of GmCYP1 rendered reduced cell death triggered by Avr3b, suggesting that GmCYP1-mediated Avr3b maturation is also required for Rps3b recognition. Finally, cyclophilins of Nicotiana benthamiana can also interact with Avr3b and activate its enzymatic activity. Overall, our results demonstrate that cyclophilin is a "helper" that activates the enzymatic activity of Avr3b after it is delivered into plant cells; as such, cyclophilin is required for the avirulence and virulence functions of Avr3b. [本文引用: 1] |
| [32] | DOI:10.1111/pce.12531PMID:25847193URLAbstract Cyclophilin 18-2 ( CYP18-2 ) genes, homologues of human peptidyl-prolyl isomerase-like 1 ( PPiL1 ), are conserved across multicellular organisms and S chizosaccharomyces pombe . Although PPiL1 is known to interact with ski-interacting protein (SKIP), a transcriptional co-regulator and spliceosomal component, there have been no functional analyses of PPiL1 homologues in plants. Rice cyclophilin 18-2 (OsCYP18-2) bound directly to amino acids 56鈥95 of OsSKIP and its binding was independent of cyclosporin A, a cyclophilin-binding drug. Moreover, OsCYP18-2 exhibited PPIase activity regardless of its interaction with OsSKIP. Therefore, the binding site for OsCYP18-2's interaction with SKIP was distinct from the PPIase active site. OsCYP18-2's interaction with SKIP full-length protein enabled OsCYP18-2's translocation from the cytoplasm into the nucleus and AtSKIP interacted in planta with both AtCYP18-2 and OsCYP18-2. Drought and salt stress induced similar expression of OsCYP 18-2 and OsSKIP . Overexpression of OsCYP 18-2 in transgenic rice and A rabidopsis thaliana plants enhanced drought tolerance and altered expression and pre-mRNA splicing patterns of stress-related genes in Arabidopsis under drought conditions. Furthermore, OsCYP18-2 caused transcriptional activation with/without OsSKIP in the GAL4 system of yeast; thus the OsSKIP-OsCYP18-2 interaction has an important role in the transcriptional and post-transcriptional regulation of stress-related genes and increases tolerance to drought stress. [本文引用: 1] |
| [33] | DOI:10.1021/pr100560vPMID:20527814URLAbstract Dwarfism with a "Green Revolution" phenotype is a desirable agronomic trait for crop cultivators as associated with increased yield, improved lodging resistance and higher fertility. Few dwarf mutants of hexaploid wheat (Triticum aestivum), except for Rht-B1 and Rht-D1, have been identified. Here, we report on a novel dwarf natural wheat mutant, which is identified as a gibberellic acid (GA)-insensitive dwarf (gaid) mutant for its semidominant blocking GA signaling pathway. Physiological and morphological investigations showed that the shoot elongation of gaid mutant plants is insensitive to exogenous GA(3) treatment. Expression of TaGA20ox1 in the gaid mutant was enhanced after GA(3) treatment. The short stem of gaid resulted from reduced cell elongation. The transcript expression of Rht, encoding a DELLA protein negatively regulating GA signaling in wheat, displayed similar patterns between gaid and wild type. Contrarily, the degradation of Rht induced by GA(3) treatment was suppressed in the mutant. 2-DE screening assay showed that the expression patterns of the mutant, as well as their responses to GA(3), were changed as compared with the wild type. In the mutant, one of enriched proteins was identified as TaCYP20-2 by Q-TOF MS approach and immunoblotting. TaCYP20-2 was localized in the chloroplast and cell plasma membrane. The transcript of TaCYP20-2 was higher in gaid than that in wild type. Molecular genetic data showed that overexpression of TaCYP20-2 in wheat resulted in a dwarfism similar to that of gaid. It suggests that TaCYP20-2 is a new member that regulates wheat stem development mediated by DELLA protein degradation of GA signaling pathway. [本文引用: 1] |
| [34] | DOI:10.1105/tpc.107.053579PMID:17704213URLChromatin-based silencing provides a crucial mechanism for the regulation of gene expression. We have identified a WD40 domain cyclophilin, CYCLOPHILIN71 (CYP71), which functions in gene repression and organogenesis in Arabidopsis thaliana. Disruption of CYP71 resulted in ectopic activation of homeotic genes that regulate meristem development. The cyp71 mutant plants displayed dramatic defects, including reduced apical meristem activity, delayed and abnormal lateral organ formation, and arrested root growth. CYP71 was associated with the chromatin of target gene loci and physically interacted with histone H3. The cyp71 mutant showed reduced methylation of H3K27 at target loci, consistent with the derepression of these genes in the mutant. As CYP71 has close homologs in eukaryotes ranging from fission yeast to human, we propose that it serves as a highly conserved histone remodeling factor involved in chromatin-based gene silencing in eukaryotic organisms. [本文引用: 2] |
| [35] | DOI:10.1038/cr.2010.22URL [本文引用: 1] |
| [36] | DOI:10.1093/mp/ssr036PMID:21596687URLChromatin is the primary carrier of epigenetic information in higher eukaryotes. AtCYP71 contains both cyclophilin domain and WD40 repeats. Loss of AtCYP71 function causes drastic pleiotropic phenotypic defects. Here, we show that AtCYP71 physically interacts with FAS1 and LHP1, respectively, to modulate their distribution on chromatin. The lhp1 cyp71 double mutant showed more severe phenotypes than the single mutants, suggesting that AtCYP71 and LHP1 synergistically control plant development. Such synergism was in part illustrated by the observation that LHP1 association with its specific target loci requires AtCYP71 function. We also demonstrate that AtCYP71 physically interacts with FAS1 and is indispensable for FAS1 targeting to the KNAT1 locus. Together, our data suggest that AtCYP71 is involved in fundamental processes of chromatin assembly and histone modification in plants. [本文引用: 1] |
| [37] | DOI:10.1016/j.chom.2014.09.007PMID:4768788URLIn plants, immune receptors such as RPM1 indirectly recognize bacterial effectors to activate immune responses. Li et al. show that a cyclophilin modulates RPM1 responses via proline isomerization of the RPM1-interacting protein RIN4, suggesting that conformation changes in receptor interacting proteins can serve as a molecular switch for immune activation. [本文引用: 2] |
| [38] | [本文引用: 1] |
| [39] | DOI:10.1073/pnas.0605452103PMID:16894144URLPhotosystem II (PSII) catalyzes the first of two photosynthetic reactions that convert sunlight into chemical energy. Native PSII is a supercomplex consisting of core and light-harvesting chlorophyll proteins. Although the structure of PSII has been resolved by x-ray crystallography, the mechanism underlying its assembly is poorly understood. Here, we report that an immunophilin of the chloroplast thylakoid lumen is required for accumulation of the PSII supercomplex in Arabidopsis thaliana. The immunophilin, FKBP20-2, belongs to the FK-506 binding protein (FKBP) subfamily that functions as peptidyl-prolyl isomerases (PPlases) in protein folding. FKBP20-2 has a unique pair of cysteines at the C terminus and was found to be reduced by thioredoxin (Trx) (itself reduced by NADPH by means of NADP-Trx reductase). The FKBP20-2 protein, which contains only two of the five amino acids required for catalysis, showed a low level of PPlase activity that was unaffected on reduction by Trx. Genetic disruption of the FKBP20-2 gene resulted in reduced plant growth, consistent with the observed lower rate of PSII activity determined by fluorescence (using leaves) and oxygen evolution (using isolated chloroplasts). Analysis of isolated thylakoid membranes with blue native gels and immunoblots showed that accumulation of the PSII supercomplex was compromised in mutant plants, whereas the levels of monomer and dimer building blocks were elevated compared with WT. The results provide evidence that FKBP20-2 participates specifically in the accumulation of the PSII supercomplex in the chloroplast thylakoid lumen by means of a mechanism that has yet to be determined. [本文引用: 1] |
| [40] | DOI:10.2174/0929866521666141121153138PMID:254128991URLThe Arabidopsis thylakoid membrane bimodular oxidoreductase, AtVKOR, could catalyze disulfide bond formation, and its direct functional domain (thioredoxin-like domain) is located in the thylakoid lumen according to the topological structure. Many proteins have one or several disulfide bonds in the thylakoid lumen, including photosynthetic chain components. A yeast two-hybrid assay was used to identify potential targets for the AtVKOR, and a Trx-like domain was constructed into a BD vector as bait. Twenty-two thylakoid lumenal proteins with disulfides were selected. The cDNAs encoding these proteins were constructed into an AD vector. Eight proteins were identified from the hybrid results to interact with AtVKOR, including HCF164, cytochrome c6A, violaxanthin deepoxidase, embryo sac development arrest 3 protein (EDA3), two members pentapeptide repeat proteins (TL17 and TL20.3), and two FK-506 binding proteins (FKBP13 and FKBP20-2). The BIACORE system was used to demonstrate that the recombinant HCF164 and Trx-like domain of AtVKOR could interact directly in vitro. The KD value for binding HCF164 to AtVKOR was calculated as 2.5 10-6 M. These results suggest that AtVKOR can interact with partial thylakoid lumenal proteins and indicates AtVKOR plays an important role in regulating the thylakoid lumen redox. [本文引用: 1] |
| [41] | [本文引用: 1] |
| [42] | [本文引用: 1] |
| [43] | DOI:10.1111/j.1742-4658.2005.04653.xPMID:2015853797URLThe RNA recognition motif (RRM), also known as RNA-binding domain (RBD) or ribonucleoprotein domain (RNP) is one of the most abundant protein domains in eukaryotes. Based on the comparison of more than 40 structures including 15 complexes (RRM090009RNA or RRM090009protein), we reviewed the structure090009function relationships of this domain. We identified and classified the different structural elements of the RRM that are important for binding a multitude of RNA sequences and proteins. Common structural aspects were extracted that allowed us to define a structural leitmotif of the RRM090009nucleic acid interface with its variations. Outside of the two conserved RNP motifs that lie in the center of the RRM 0205-sheet, the two external 0205-strands, the loops, the C- and N-termini, or even a second RRM domain allow high RNA-binding affinity and specific recognition. Protein090009RRM interactions that have been found in several structures reinforce the notion of an extreme structural versatility of this domain supporting the numerous biological functions of the RRM-containing proteins. [本文引用: 1] |
| [44] | DOI:10.1111/j.1365-313X.2009.03878.xPMID:19366428URLArabidopsis ROF1 (AtFKBP62) is a peptidyl prolyl cis / trans isomerase and a member of the FKBP (FK506 binding protein) family. ROF1 expression is induced by heat stress and developmentally regulated. In this study, we show that ROF1 binds heat shock proteins HSP90.1 via its tetratricopeptide repeat domain, and localizes in the cytoplasm under normal conditions. Exposure to heat stress induces nuclear localization of the ROF1–HSP90.1 complex, which is dependent upon the presence of the transcription factor HsfA2, which interacts with HSP90.1 but not with ROF1. Nuclear localization of ROF1 was not detected in Arabidopsis HSP90.1 and HsfA2 knockout mutants. The rof1 knockout plants exhibited collapse when 24–48 h passed between acclimation at 37°C and exposure to 45°C. Transgenic ROF1 over-expressors showed better survival in response to exposure to 45°C than wild-type plants did. In rof1 knockout mutants, the level of expression of small HSPs regulated by HsfA2 was dramatically reduced after exposure to 37°C and recovery for 24–48 h, and correlates well with the mutant phenotype. We suggest a role for ROF1 in prolongation of thermotolerance by sustaining the levels of small HSPs that are essential for survival at high temperatures. [本文引用: 1] |
| [45] | [本文引用: 1] |
| [46] | DOI:10.1007/s00709-015-0828-zURL [本文引用: 1] |
| [47] | [本文引用: 2] |
| [48] | DOI:10.1073/pnas.1218872110URL [本文引用: 1] |
| [49] | DOI:10.1105/tpc.109.068791PMID:19903870URLIn higher plants, the chloroplast NAD(P) H dehydrogenase (NDH) complex mediates photosystem I (PSI) cyclic and chlororespiratory electron transport. We reported previously that NDH interacts with the PSI complex to form a supercomplex (NDH-PSI). In this study, NDH18 and FKBP16-2 (FK506 Binding Protein 16-2), detected in the NDH-PSI supercomplex by mass spectrometry, were shown to be NDH subunits by the analysis of their knockdown lines. On the basis of extensive mutant characterization, we propose a structural model for chloroplast NDH, whereby NDH is divided into four subcomplexes. The subcomplex A and membrane subcomplex are conserved in cyanobacteria, but the subcomplex B and lumen subcomplex are specific to chloroplasts. Two minor light-harvesting complex I proteins, Lhca5 and Lhca6, were required for the full-size NDH-PSI supercomplex formation. Similar to err pgr5 double mutants that completely lack cyclic electron flow activity around PSI, the Ihca6 pgr5 double mutant exhibited a severe defect in growth. Consistent with the impaired NDH activity, photosynthesis was also severely affected in mature leaves of Ihca6 pgr5. We conclude that chloroplast NDH became equipped with the novel subcomplexes and became associated with PSI during the evolution of land plants, and this process may have facilitated the efficient operation of NDH. [本文引用: 1] |
| [50] | [本文引用: 2] |
| [51] | [本文引用: 1] |
| [52] | DOI:10.1104/pp.123.4.1525URL [本文引用: 1] |
| [53] | DOI:10.1105/tpc.109.071209PMID:20145257URLVery-long-chain fatty acids (VLCFAs) are essential for many aspects of plant development and necessary for the synthesis of seed storage triacylglycerols, epicuticular waxes, and sphingolipids. Identification of the acetyl-CoA carboxylase PASTICCINO3 and the 3-hydroxy acyl-CoA dehydratase PASTICCINO2 revealed that VLCFAs are important for cell proliferation and tissue patterning. Here, we show that the immunophilin PASTICCINO1 (PAS1) is also required for VLCFA synthesis. Impairment of PAS1 function results in reduction of VLCFA levels that particularly affects the composition of sphingolipids, known to be important for cell polarity in animals. Moreover, PAS1 associates with several enzymes of the VLCFA elongase complex in the endoplasmic reticulum. The pas1 mutants are deficient in lateral root formation and are characterized by an abnormal patterning of the embryo apex, which leads to defective cotyledon organogenesis. Our data indicate that in both tissues, defective organogenesis is associated with the mistargeting of the auxin efflux carrier PIN FORMED1 in specific cells, resulting in local alteration of polar auxin distribution. Furthermore, we show that exogenous VLCFAs rescue lateral root organogenesis and polar auxin distribution, indicating their direct involvement in these processes. Based on these data, we propose that PAS1 acts as a molecular scaffold for the fatty acid elongase complex in the endoplasmic reticulum and that the resulting VLCFAs are required for polar auxin transport and tissue patterning during plant development. [本文引用: 1] |
| [54] | DOI:10.1186/1471-2229-11-34PMID:3050798URLpAbstract/p pBackground/p pHigh Salinity is a major environmental stress influencing growth and development of rice. Comparative proteomic analysis of hybrid rice shoot proteins from Shanyou 10 seedlings, a salt-tolerant hybrid variety, and Liangyoupeijiu seedlings, a salt-sensitive hybrid variety, was performed to identify new components involved in salt-stress signaling./p pResults/p pPhenotypic analysis of one protein that was upregulated during salt-induced stress, cyclophilin 2 (OsCYP2), indicated that itOsCYP2 /ittransgenic rice seedlings had better tolerance to salt stress than did wild-type seedlings. Interestingly, wild-type seedlings exhibited a marked reduction in maximal photochemical efficiency under salt stress, whereas no such change was observed for itOsCYP2/it-transgenic seedlings. itOsCYP2/it-transgenic seedlings had lower levels of lipid peroxidation products and higher activities of antioxidant enzymes than wild-type seedlings. Spatiotemporal expression analysis of itOsCYP2 /itshowed that it could be induced by salt stress in both Shanyou 10 and Liangyoupeijiu seedlings, but Shanyou 10 seedlings showed higher itOsCYP2 /itexpression levels. Moreover, circadian rhythm expression of itOsCYP2 /itin Shanyou 10 seedlings occurred earlier than in Liangyoupeijiu seedlings. Treatment with PEG, heat, or ABA induced itOsCYP2 /itexpression in Shanyou 10 seedlings but inhibited its expression in Liangyoupeijiu seedlings. Cold stress inhibited itOsCYP2 /itexpression in Shanyou 10 and Liangyoupeijiu seedlings. In addition, OsCYP2 was strongly expressed in shoots but rarely in roots in two rice hybrid varieties./p pConclusions/p pTogether, these data suggest that OsCYP2 may act as a key regulator that controls ROS level by modulating activities of antioxidant enzymes at translation level. OsCYP2 expression is not only induced by salt stress, but also regulated by circadian rhythm. Moreover, OsCYP2 is also likely to act as a key component that is involved in signal pathways of other types of stresses-PEG, heat, cold, or ABA./p [本文引用: 1] |
| [55] | DOI:10.1016/j.bbagen.2014.11.012PMID:25445709URLAccessory domains covalently linked to a PPIase domain supply an additional layer of control to the catalysis of prolyl isomerization in specific client proteins. Understanding these control mechanisms will provide new insights into the physiological mode of action of the multidomain PPIases and their ability to form therapeutic targets. This article is part of a Special Issue entitled Proline-directed Foldases: Cell Signaling Catalysts and Drug Targets. [本文引用: 2] |
| [56] | DOI:10.1111/ppl.12116PMID:24124981URLArabidopsis contains 16 putative chloroplast lumen-targeted immunophilins (IMMs). Proteomic analysis has enabled the subcellular localization of IMMs experimentally, but the exact biological and physiological roles of most luminal IMMs remain to be discovered. FK506-binding protein (FKBP) 16-1, one of the lumenal IMMs containing poorly conserved amino acid residues for peptidyl-prolyl isomerase (PPIase) activity, was shown to play a possible role in chloroplast biogenesis in Arabidopsis, and was also found to interact with PsaL in wheat. In this study, further evidence is provided for the notion that Arabidopsis FKBP16-1 (AtFKBP16-1) is transcriptionally and post-transcriptionally regulated by environmental stresses including high light (HL) intensity, and that overexpression of AtFKBP16-1 plants exhibited increased photosynthetic stress tolerance. A blue native-polyacrylamide gel electrophoresis/two-dimensional (BN-PAGE/2-D) analysis revealed that the increase of AtFKBP16-1 affected the levels of photosystem I (PSI)-light harvesting complex I (LHCI) and PSI-LHCI-light harvesting complex II (LHCII) supercomplex, and consequently enhanced tolerance under conditions of HL stress. In addition, plants overexpressing AtFKBP16-1 showed increased accumulation of PsaL protein and enhanced drought tolerance. Using a protease protection assay, AtFKBP16-1 protein was found to have a role in PsaL stability. The AtPsaL levels also responded to abiotic stresses derived from drought, and from methyl viologen stresses in wild-type plants. Taken together, these results suggest that AtFKBP16-1 plays a role in the acclimation of plants under photosynthetic stress conditions, probably by regulating PsaL stability. [本文引用: 1] |
| [57] | DOI:10.1016/j.febslet.2009.06.031PMID:19549520URLAtCYP20-2 is one of 16 immunophilins in thylakoid lumen. The presence of the isomerase domain in AtCYP20-2, an enrichment of AtCYP20-2 in the stroma membranes and it co-migration with NAD(P)H dehydrogenase (NDH) in native gels provide evidence that AtCYP20-2 is an auxiliary protein of NDH. When different NDH mutants were studied, AtCYP20-2 was found to be strongly reduced especially in mutants deficient in the membrane domain of NDH, thus suggesting a role in the assembly of NDH hydrophobic domain. Lack of AtCYP20-2, however, did not lead to severe malfunction of NDH, indicating redundancy in the function of lumenal immunophilins. [本文引用: 1] |
| [58] | DOI:10.1111/j.1365-313X.2008.03532.xPMID:18445132URLAtCYP38 is a thylakoid lumen protein comprising the immunophilin domain and the phosphatase inhibitor module. Here we show the association of AtCYP38 with the photosystem II (PSII) monomer complex and address its functional role using AtCYP38-deficient mutants. The dynamic greening process of etiolated leaves failed in the absence of AtCYP38, due to specific problems in the biogenesis of PSII complexes. Also the development of leaves under short-day conditions was severely disturbed. Detailed biophysical and biochemical analysis of mature AtCYP38-deficient plants from favorable growth conditions (long photoperiod) revealed: (i) intrinsic malfunction of PSII, which (ii) occurred on the donor side of PSII and (iii) was dependent on growing light intensity. AtCYP38 mutant plants also showed decreased accumulation of PSII, which was shown not to originate from impaired D1 synthesis or assembly of PSII monomers, dimers and supercomplexes as such but rather from the incorrect fine-tuning of the oxygen-evolving side of PSII. This, in turn, rendered PSII centers extremely susceptible to photoinhibition. AtCYP38 deficiency also drastically decreased the in vivo phosphorylation of PSII core proteins, probably related to the absence of the AtCYP38 phosphatase inhibitor domain. It is proposed that during PSII assembly AtCYP38 protein guides the proper folding of D1 (and CP43) into PSII, thereby enabling the correct assembly of the water-splitting Mn 4 a cluster even with high turnover of PSII. [本文引用: 1] |
| [59] | DOI:10.1016/j.coph.2011.03.010PMID:21511531URLThe large FK506-binding protein FKBP52 has been characterized as an important positive regulator of androgen, glucocorticoid and progesterone receptor signaling pathways. FKBP52 associates with receptor sp90 complexes and is proposed to have roles in both receptor hormone binding and receptor subcellular localization. Data from biochemical and cellular studies have been corroborated in whole animal models as fkbp52-deficient male and female mice display characteristics of androgen, glucocorticoid and/or progesterone insensitivity. FKBP52 receptor specificity and the specific phenotypes displayed by the fkbp52-deficient mice have firmly established FKBP52 as a promising target for the treatment of a variety of hormone-dependent diseases. Recent studies demonstrated that the FKBP52 FK1 domain and the proline-rich loop within this domain are functionally important for FKBP52 regulation of receptor function. Based on these data, efforts are currently underway to target the FKBP52 FK1 domain and the proline-rich loop with small molecule inhibitors. [本文引用: 1] |
| [60] | DOI:10.1074/jbc.M601815200PMID:16803883URLPASTICCINO1 (PAS1) is a high molecular weight FK506-binding protein (FKBP) involved in the control of cell proliferation and differentiation during plant development. Mutations in the C-terminal region of PAS1 result in severe developmental defects. We show here that the C-terminal domain of PAS1 controls the subcellular distribution of this protein. We also demonstrated in vitro and in vivo, by Forster resonance energy transfer, that this C-terminal region is required for interaction with FAN (FKBP-associated NAC), a new member of the plant-specific family of NAC transcription factors. PAS1 and FAN are translocated into the nucleus upon auxin treatment in plant seedlings. The nuclear translocation of PAS1 is dependent on the presence of the C terminus of the protein. Finally, we showed that FAN is involved in PAS1-regulated processes because FAN overproduction partly complemented the pas1 phenotype. We suggest that PAS1 regulates the function of this NAC-like transcription factor by controlling its targeting to the nucleus upon plant cell division. [本文引用: 1] |
| [61] | DOI:10.1111/tpj.12932PMID:26173789URLSummary The plant vascular system serves as a conduit for delivery of both nutrients and signaling molecules to various distantly located organs. The anucleate sieve tube system of the angiosperm phloem delivers sugars and amino acids to developing organs, and has recently been shown to contain a unique population of RNA and proteins. Grafting studies have established that a number of these macromolecules are capable of moving long distances between tissues, thus providing support for operation of a phloem-mediated inter-organ communication network. Currently, our knowledge of the roles played by such phloem-borne macromolecules is in its infancy. Here, we show that, in tomato, translocation of a phloem-mobile cyclophilin, SlCyp1, from a wild-type scion into a mutant rootstock results in restoration of vascular development and lateral root initiation. This process occurs through reactivation of auxin response pathways and reprogramming of the root transcriptome. Moreover, we show that long-distance trafficking of SlCyp1 is associated with regulation of the shoot-to-root ratio in response to changing light intensities, by modulating root growth. We conclude that long-distance trafficking of SlCyp1 acts as a rheostat to control the shoot-to-root ratio, by mediating root development to integrate photosynthesis and light intensity with requirements for access to water and mineral nutrients. [本文引用: 1] |
| [62] | [本文引用: 1] |
| [63] | DOI:10.1111/j.1365-313X.2012.05013.xPMID:22463079URLAlthough multiple photoreceptors converge to control common aspects of seedling de-etiolation, we are relatively ignorant of the genes acting at or downstream of their signalling convergence. To address this issue we screened for mutants under a mixture of blue plus far-red light and identified roc1-1D. The roc1-1D mutant, showing elevated expression of the ROTAMASE CYCLOPHILIN 1 (ROC1/AtCYP18-3) gene, and partial loss-of function roc1 alleles, has defects in phytochrome A (phyA)-, cryptochrome 1 (cry1)- and phytochrome B (phyB)-mediated de-etiolation, including long hypocotyls under blue or far-red light. These mutants show elevated sensitivity to brassinosteroids in the light but not in the dark. Mutations at brassinosteroid signalling genes and the application of a brassinosteroid synthesis inhibitor eliminated the roc1 and roc1-D phenotypes. The roc1 and roc1-D mutants show altered patterns of phosphorylation of the transcription factor BES1, a known point of control of sensitivity to brassinosteroids, which correlate with the expression levels of genes directly targeted by BES1. We propose a model where perception of light by phyA, cry1 or phyB activates ROC1 (at least in part by enhancing its expression). This in turn reduces the intensity of brassinosteroid signalling and fine-tunes seedling de-etiolation. [本文引用: 1] |
| [64] | DOI:10.1105/tpc.111.093781PMID:22706283URLCyclophilin38 (CYP38) is one of the highly divergent cyclophilins from Arabidopsis thaliana. Here, we report the crystal structure of the At-CYP38 protein (residues 83 to 437 of 437 amino acids) at 2.39-03 resolution. The structure reveals two distinct domains: an N-terminal helical bundle and a C-terminal cyclophilin β-barrel, connected by an acidic loop. Two N-terminal β-strands become part of the C-terminal cyclophilin β-barrel, thereby making a previously undiscovered domain organization. This study shows that CYP38 does not possess peptidyl-prolyl cis/trans isomerase activity and identifies a possible interaction of CYP38 with the E-loop of chlorophyll protein47 (CP47), a component of photosystem II. The interaction of CYP38 with the E-loop of CP47 is mediated through its cyclophilin domain. The N-terminal helical domain is closely packed together with the putative C-terminal cyclophilin domain and establishes a strong intramolecular interaction, thereby preventing the access of the cyclophilin domain to other proteins. This was further verified by protein—protein interaction assays using the yeast two-hybrid system. Furthermore, the non-Leucine zipper N-terminal helical bundle contains several new elements for protein—protein interaction that may be of functional significance. Together, this study provides the structure of a plant cyclophilin and explains a possible mechanism for autoinhibition of its function through an intramolecular interaction. [本文引用: 1] |
| [65] | DOI:10.1016/j.bbagen.2014.12.017PMID:25529299URLThis article is part of a Special Issue entitled Proline-directed Foldases: Cell Signaling Catalysts and Drug Targets. [本文引用: 2] |
| [66] | DOI:10.1104/pp.103.028050URL |
| [67] | DOI:10.1016/j.cell.2009.01.046PMID:19239888URLMicroRNAs (miRNAs) are key posttranscriptional regulators of eukaryotic gene expression. Plants use highly conserved as well as more recently evolved, species-specific miRNAs to control a vast array of biological processes. This Review discusses current advances in our understanding of the origin, biogenesis, and mode of action of plant miRNAs and draws comparisons with their metazoan counterparts. [本文引用: 1] |
| [68] | DOI:10.1105/tpc.112.105999PMID:23321285URLPlant architecture is influenced by the polar, cell-to-cell transport of auxin that is primarily provided and regulated by plasma membrane efflux catalysts of the PIN-FORMED and B family of ABC transporter (ABCB) classes. The latter were shown to require the functionality of the FK506 binding protein42 TWISTED DWARF1 (TWD1), although underlying mechanisms are unclear. By genetic manipulation of TWD1 expression, we show here that TWD1 affects shootward root auxin reflux and, thus, downstream developmental traits, such as epidermal twisting and gravitropism of the root. Using immunological assays, we demonstrate a predominant lateral, mainly outward-facing, plasma membrane location for TWD1 in the root epidermis characterized by the lateral marker ABC transporter G36/PLEIOTROPIC DRUG-RESISTANCE8/PENETRATION3. At these epidermal plasma membrane domains, TWD1 colocalizes with nonpolar ABCB1. In planta bioluminescence resonance energy transfer analysis was used to verify specific ABC transporter B1 (ABCB1)-TWD1 interaction. Our data support a model in which TWD1 promotes lateral ABCB-mediated auxin efflux via protein-protein interaction at the plasma membrane, minimizing reflux from the root apoplast into the cytoplasm. [本文引用: 1] |
| [69] | DOI:10.1016/j.plaphy.2013.12.020PMID:24468661URL61The GhPPI mRNA level is upregulated under NaCl stress.61GhPPI is located in the nucleus.61GhPPI exhibits PPIase activity and suppresses aggregation of creatine kinase during refolding.61Key amino acids were predicted using homology modeling and molecular docking.61PPIase activity of GhPPI mutants was decreased compared to the wide type. [本文引用: 2] |
| [70] | DOI:10.1016/j.febslet.2008.01.055PMID:18258190URLHuman nuclear cyclophilin 33 (hCyP33) was the first protein which was found to contain an RNA-binding motif and a PPIase domain. It was not known what cellular and physiological roles are played by the RNA-binding activity as well as the PPIase activity of hCyP33. In this paper, we investigated the binding specificity of hCyP33 to different cellular RNA using ion-exchange chromatography and affinity adsorption. Furthermore, the influence of different cellular RNAs to the PPIase activity of hCyP33 was investigated using a protease-coupled method. The results show that hCyP33 binds specifically to mRNA, namely poly(A) +RNA, and that binding stimulates the PPIase activity of hCyP33. [本文引用: 1] |
| [71] | [本文引用: 1] |
| [72] | DOI:10.3389/fpls.2015.00777PMID:4586435URLAbstract As a member of the Immunophilin family, cyclophilin38 (CYP38) is discovered to be localized in the thylakoid lumen, and is reported to be a participant in the function regulation of thylakoid membrane protein. However, the molecule mechanisms remain unclear. We found that, CYP38 plays an important role in the process of regulating and protecting the plant to resist high light (HL) stress. Under HL condition, the gene expression of CYP38 is enhanced, and if CYP38 gene is deficient, photochemistry efficiency, and chlorophyll content falls distinctly, and excessive reactive oxygen species synthesis occurs in the chloroplast. Western blot results showed that the D1 degradation rate of cyp38 mutant plants is faster than that of wide type plants. Interestingly, both gene expression and activity of PsbO2 were drastically enhanced in cyp38 mutant plants and less changed when the deleted gene of CYP38 was restored under HL treatment. This indicates that CYP38 may impose a negative regulation effect on PsbO2, which exerts a positive regulation effect in facilitating the dephosphorylation and subsequent degradation of D1. It is also found that, under HL condition, the cytoplasmic calcium ([Ca(2+)]cyt) concentration and the gene expression level of calmodulin 3 (CaM3) arose markedly, which occurs upstream of CYP38 gene expression. In conclusion, our results indicate that CYP38 plays an important role in plant strengthening HL resistibility, which provides a new insight in the research of mechanisms of CYP38 protein in plants. [本文引用: 1] |
| [73] | DOI:10.1105/tpc.110.078360PMID:20971896URLMultidrug resistance ABC transporters in plants are required for polar transport of the hormone auxin (indole-3-acetic acid). They are studied in animals primarily because their overexpression confers resistance to anticancer agents. Immunophilins are studied in both plants and animals for their roles in folding and trafficking of proteins, particularly those with signal transducing functions and susceptibility to immunosuppressant drugs. Previous genetic and molecular studies in Arabidopsis thaliana established a physical and functional interaction between some ABCB transporters and the TWISTED DWARF1 (TWD1) immunophilin. In this work, confocal microscopy of fluorescently tagged TWD1 shows it to reside at the endoplasmic reticulum (ER). Mutations in TWD1 caused mislocalization of ABCB1, ABCB4, and ABCB19 to the ER instead of the plasma membrane as shown by confocal microscopy of fluorescently tagged fusion proteins and transmission electron microscopy of immunogold-labeled samples in the case of ABCB19. Localization of the unrelated PIN-FORMED2 auxin transporter or plasma membrane marker proteins was not affected by loss of TWD1. Abnormal spread of auxin signaling into the elongation zone of twd1 roots, attributable to mislocalized ABCB transporters and suppressed by an auxin transport inhibitor, appeared to cause the twisted cell files characteristic of twd1 roots. [本文引用: 4] |
| [74] | DOI:10.1038/ncomms10430PMID:26791759URLRoot gravitropism allows plants to establish root systems and its regulation depends on polar auxin transport mediated by PIN-FORMED (PIN) auxin transporters. PINOID (PID) and PROTEIN PHOSPHATASE 2A (PP2A) act antagonistically on reversible phosphorylation of PINs. This regulates polar PIN distribution and auxin transport. Here we show that a peptidyl-prolylcis/transisomerase Pin1At regulates root gravitropism. Downregulation ofPin1Atsuppresses root agravitropic phenotypes ofpp2aaand35S:PID, while overexpression ofPin1Ataffects root gravitropic responses and enhances thepp2aaagravitropic phenotype. Pin1At also affects auxin transport and polar localization of PIN1 in stele cells, which is mediated by PID and PP2A. Furthermore, Pin1At catalyses the conformational change of the phosphorylated Ser/Thr-Pro motifs of PIN1. Thus, Pin1At mediates the conformational dynamics of PIN1 and affects PID- and PP2A-mediated regulation of PIN1 polar localization, which correlates with the regulation of root gravitropism. Asymmetric transport of auxin allows plants to orient growth of their roots towards gravity. Here, the authors show that the peptidyl-prolylcis/transisomerase Pin1At contributes to root gravitropism and propose that it acts by altering the polarity of the PIN1 auxin transport protein. [本文引用: 2] |
| [75] | [本文引用: 2] |
| [76] | DOI:10.1093/jxb/err120PMID:3192995URLThe HAP complex occurs in many eukaryotic organisms and is involved in multiple physiological processes. Here it was found that in Picea wilsonii, HAP5 (PwHAP5), a putative CCAAT-binding transcription factor gene, is involved in pollen tube development and control of tube orientation. Quantitative real-time reverse transcription-PCR showed that PwHAP5 transcripts were expressed strongly in germinating pollen and could be induced by Ca2+. Overexpression of PwHAP5 in pollen altered pollen tube orientation, whereas the tube with PwHAP5RNAi showed normal growth without diminishing pollen tube growth. Furthermore, PwFKBP12, which encodes an FK506-binding protein (FKBP) was screened and a bimolecular fluorescence complementation assay performed to confirm the interaction of PwHAP5 and PwFKBP12 in vivo. Transient expression of PwFKBP12 in pollen showed normal pollen tube growth, whereas the tube with PwFKBP12RNAi bent. The phenotype of overexpression of HAP5 on pollen tube was restored by FKBP12. Altogether, our study supported the role of HAP5 in pollen tube development and orientation regulation and identified FKBP12 as a novel partner to interact with HAP5 involved in the process. |
| [77] | DOI:10.1105/tpc.113.110296PMID:23897924URLBrassinosteroids (BRs) regulate many physiological processes during plant development, including flowering. However, little is known about the components of BR signaling that mediate flowering. Here, we report that BRASSINAZOLE-RESISTANT1 (BZR1), the conformation of which is altered by a cyclophilin (CYP20-2), binds cis-elements in the FLOWERING LOCUS D (FLD) promoter to regulate flowering. Both bzr1-1D and fld-4 showed delayed flowering. Electrophoretic mobility shift assay and chromatin immunoprecipitation revealed that BZR1 bound to a putative BR response cis-element and suppressed the expression of FLD. Overexpression of FLD partially rescued the late flowering of pBZR1:mBZR1 Pro234-Leu -CFP (mx3). Yeast two-hybrid and pull-down assays demonstrated that BZR1 interacts with CYP20-2. Arabidopsis thaliana CYP20-2 had greater peptidyl-prolyl cis-trans isomerase activity than did wheat (Triticum aestivum) CYP20-2. Fourier transform infrared spectroscopy revealed conformation changes in BZR1, dependent on interaction with CYP20-2. Due to differences in activity and substrate preference between CYP20-2 proteins from wheat and Arabidopsis, At-CYP20-2 verexpressing lines showed earlier flowering, whereas Ta CYP20-2 lines flowered later. Immunoblot and chromatin immunoprecipitation assays showed that histone H3 trimethyl Lys4 and H3 acetylation levels were negatively correlated with the transcription of FLD (a putative histone demethylase) in various lines. Therefore, a conformational change of BZR1 mediated by CYP20-2 causes altered flowering through modulation of FLD expression. [本文引用: 3] |
| [78] | DOI:10.1016/j.molp.2016.01.007PMID:26802250URLThese studies demonstrated that TWD1 associates with the brassinosteroid receptor, BRI1, and regulates the early events of BR signaling pathway. [本文引用: 5] |
| [79] | DOI:10.1093/mp/sst052PMID:23501875URL [本文引用: 1] |
| [80] | DOI:10.1105/tpc.15.00726PMID:27053424URLPlant growth and architecture is regulated by the polar distribution of the hormone auxin. Polarity and flexibility of this process is provided by constant cycling of auxin transporter vesicles along actin filaments, coordinated by a positive auxin-actin feedback loop. Both polar auxin transport and vesicle cycling are inhibited by synthetic auxin transport inhibitors, such as 1-N-naphthylphthalamic acid (NPA), counteracting the effect of auxin; however, underlying targets and mechanisms are unclear. Using NMR, we map the NPA binding surface on the Arabidopsis thaliana ABCB chaperone TWISTED DWARF1 (TWD1). We identify ACTIN7 as a relevant, although likely indirect, TWD1 interactor, and show TWD1-dependent regulation of actin filament organization and dynamics and that TWD1 is required for NPA-mediated actin cytoskeleton remodeling. The TWD1-ACTIN7 axis controls plasma membrane presence of efflux transporters, and as a consequence act7 and twd1 share developmental and physiological phenotypes indicative of defects in auxin transport. These can be phenocopied by NPA treatment or by chemical actin (de)stabilization. We provide evidence that TWD1 determines downstream locations of auxin efflux transporters by adjusting actin filament debundling and dynamizing processes and mediating NPA action on the latter. This function appears to be evolutionary conserved since TWD1 expression in budding yeast alters actin polarization and cell polarity and provides NPA sensitivity. [本文引用: 1] |
| [81] | DOI:10.1093/jxb/erv308PMID:26085676URLAbstract Polar auxin transport and the action of the actin cytoskeleton are tightly interconnected, which is documented by the finding that auxin transporters reach their final destination by active movement of secretory vesicles along F-actin tracks. Moreover, auxin transporter polarity and flexibility is thought to depend on transporter cycling that requires endocytosis and exocytosis of vesicles. In this context, we have reviewed the current literature on an involvement of the actin cytoskeleton in polar auxin transport and identify known similarities and differences in its structure, function and dynamics in comparison to non-plant organisms. By describing how auxin modulates actin expression and actin organization and how actin and its stability affects auxin-transporter endocytosis and recycling, we discuss the current knowledge on regulatory auxin-actin feedback loops. We focus on known effects of auxin and of auxin transport inhibitors on the stability and organization of actin and examine the functionality of auxin and/or auxin transport inhibitor-binding proteins with respect to their suitability to integrate auxin/auxin transport inhibitor action. Finally, we indicate current difficulties in the interpretation of organ, time and concentration-dependent auxin/auxin transport inhibitor treatments and formulate simple future experimental guidelines. The Author 2015. Published by Oxford University Press on behalf of the Society for Experimental Biology. All rights reserved. For permissions, please email: journals.permissions@oup.com. [本文引用: 1] |
植物FKBP基因家族的结构及生物学功能
1
2014
... 亲免素(immunophilin)在生物体内广泛存在, 可与免疫抑制剂他克莫司(tacrolimus, FK506)、雷帕霉素(rapamycin)及环孢素A (cyclosporin A)结合, 是含量比较丰富的一类细胞受体蛋白.根据与亲免素结合的免疫抑制剂不同, 可将其分为3类: 能与FK506和雷帕霉素特异结合的FK506结合蛋白(FK506 binding proteins, FKBPs)、能与环孢素A结合的亲环素(cyclophilins, CYPs)以及能与5-羟基-1,4-萘醌结合的parvulin蛋白(
2
2010
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
... )有62个、酵母有8个、人类有24个(
1
2004
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
1
2007
... FKBPs对热胁迫、盐胁迫、干旱及强光胁迫都有响应.拟南芥中2个高度同源的基因FKBP62 (AtROF1)与FKBP65 (AtROF2)参与植物耐热性调控.但是在热胁迫下, 2个基因的表达量和调节方式不同, AtROF1促进热激蛋白的表达而AtROF2抑制热激蛋白的表达(
1
2011
... 近年来的研究表明, FKBPs与多种激素关系密切.拟南芥中调节生长素运输的TWD1与人类FKBP38结构相似, 人类FKBP38的PPlase活性能够促进囊性纤维化跨膜传导调节蛋白(CFTR)的翻译后折叠, 并且其TPR结构域与Hsp90相互作用能够抑制PPlase活性, 对CFTR的合成进行负反馈调节(
2
2006
... 亲免素(immunophilin)在生物体内广泛存在, 可与免疫抑制剂他克莫司(tacrolimus, FK506)、雷帕霉素(rapamycin)及环孢素A (cyclosporin A)结合, 是含量比较丰富的一类细胞受体蛋白.根据与亲免素结合的免疫抑制剂不同, 可将其分为3类: 能与FK506和雷帕霉素特异结合的FK506结合蛋白(FK506 binding proteins, FKBPs)、能与环孢素A结合的亲环素(cyclophilins, CYPs)以及能与5-羟基-1,4-萘醌结合的parvulin蛋白(
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
1
2012
... FKBPs对热胁迫、盐胁迫、干旱及强光胁迫都有响应.拟南芥中2个高度同源的基因FKBP62 (AtROF1)与FKBP65 (AtROF2)参与植物耐热性调控.但是在热胁迫下, 2个基因的表达量和调节方式不同, AtROF1促进热激蛋白的表达而AtROF2抑制热激蛋白的表达(
5
2016
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
... 亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(
... Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (
... 最新研究显示, 拟南芥TWD1也参与油菜素甾醇(brassinosteroid, BR)的信号传递.在根生长和下胚轴伸长过程中, twd1-2以及twd1-5对油菜素内酯(brassinolide, BL)的敏感程度降低; 在twd1-5中磷酸化和未磷酸化的BES1含量均降低.实验结果显示TWD1在植物体内能与BRI1和BAK1相互作用.以上表明TWD1与BRI1和BAK1相互作用影响了BR信号转导(
... 近年来的研究表明, FKBPs与多种激素关系密切.拟南芥中调节生长素运输的TWD1与人类FKBP38结构相似, 人类FKBP38的PPlase活性能够促进囊性纤维化跨膜传导调节蛋白(CFTR)的翻译后折叠, 并且其TPR结构域与Hsp90相互作用能够抑制PPlase活性, 对CFTR的合成进行负反馈调节(
1
2008
... 叶绿体内半胱氨酸合成过程中的重要酶(丝氨酸乙酰基转移酶(SAT1))能与AtCYP20-3相互作用, 指导SAT1的正确折叠, 保证半胱氨酸合成通路, 以适应各种逆境(
2
2016
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
... 在哺乳动物中, 亲免素的主要功能是与小的配体结合(如免疫抑制剂), 或者作为分子伴侣调节蛋白质的结构、活性和稳定性.在高等植物中, 亲免素通过与相关蛋白质相互作用调控生长发育; 并且FBKPs对免疫抑制剂不敏感, PPlase活性也不保守; 而亲环素对免疫抑制剂敏感, PPlase活性也较为保守.例如, AtROC1、LeCYP1和OsCYP2都具有PPlase活性(
2
2010
... 基因沉默、表观遗传学调控和转录水平的调控对植物的生长发育产生重要影响.拟南芥cyp40突变体叶片数量减少和形态改变与microRNA活性减弱有关.成熟的microRNA能够与RNA诱导沉默复合体(RNA- induced silencing complex, RISC)结合, 形成具有切割活性的RNA沉默复合物, 该复合物与靶基因互补配对使靶基因沉默或抑制靶基因的翻译.AGO1 (ARGONAUTE1)是RISC的重要组成部分(
... ).AtCYP40与AGO1相互作用促进或维持AGO1的活性, 从而有助于HSP90调节RISC的组装, 影响基因沉默或基因的翻译(
1
2007
... 拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用.拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(
4
2007
... 亲免素(immunophilin)在生物体内广泛存在, 可与免疫抑制剂他克莫司(tacrolimus, FK506)、雷帕霉素(rapamycin)及环孢素A (cyclosporin A)结合, 是含量比较丰富的一类细胞受体蛋白.根据与亲免素结合的免疫抑制剂不同, 可将其分为3类: 能与FK506和雷帕霉素特异结合的FK506结合蛋白(FK506 binding proteins, FKBPs)、能与环孢素A结合的亲环素(cyclophilins, CYPs)以及能与5-羟基-1,4-萘醌结合的parvulin蛋白(
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
... 亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(
... Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (
2004
3
2003
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
... 植物的生长发育与激素密不可分.亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用.拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布.拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输.TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(
... ;
1
2010
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
1
2011
... 小麦TaFKBP16-1和TaFKBP16-3在光系统I (PSI)的组装过程中扮演着重要角色.TaFKBP16-1与PSI的1个亚基PsaL (PSI subunit L)相互作用, TaFK- BP16-3与Thf1 (Thylakoid Formation 1)和APO2 (Os02g50010)存在相互作用, 因此TaFKBP16-1和TaFKBP16-3可能调控PSI的组装(
2000
2002
1
2003
... 在哺乳动物中, FKBPs与转录因子相互作用较为常见, 在植物体内也发现FKBPs与转录因子相互作用调节细胞的分裂与分化.拟南芥突变体pas1 (pasticcion1)的细胞分裂异常、分化紊乱, 对野生型植株施加外源性细胞分裂素后出现与pas1相似的表型.研究发现, PAS1 (即FKBP72)的C末端与FAN转录因子(FKBP-associated NAC)相互作用, 促进了FAN对细胞分裂的抑制(
4
2004
... 亲免素(immunophilin)在生物体内广泛存在, 可与免疫抑制剂他克莫司(tacrolimus, FK506)、雷帕霉素(rapamycin)及环孢素A (cyclosporin A)结合, 是含量比较丰富的一类细胞受体蛋白.根据与亲免素结合的免疫抑制剂不同, 可将其分为3类: 能与FK506和雷帕霉素特异结合的FK506结合蛋白(FK506 binding proteins, FKBPs)、能与环孢素A结合的亲环素(cyclophilins, CYPs)以及能与5-羟基-1,4-萘醌结合的parvulin蛋白(
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
... 拟南芥突变体cyp71茎尖分生组织活性下降, 侧生器官发育异常, 花的形态发生改变, 根的生长受抑制(
... 在哺乳动物中, 亲免素的主要功能是与小的配体结合(如免疫抑制剂), 或者作为分子伴侣调节蛋白质的结构、活性和稳定性.在高等植物中, 亲免素通过与相关蛋白质相互作用调控生长发育; 并且FBKPs对免疫抑制剂不敏感, PPlase活性也不保守; 而亲环素对免疫抑制剂敏感, PPlase活性也较为保守.例如, AtROC1、LeCYP1和OsCYP2都具有PPlase活性(
1
1998
... 亲免素(immunophilin)在生物体内广泛存在, 可与免疫抑制剂他克莫司(tacrolimus, FK506)、雷帕霉素(rapamycin)及环孢素A (cyclosporin A)结合, 是含量比较丰富的一类细胞受体蛋白.根据与亲免素结合的免疫抑制剂不同, 可将其分为3类: 能与FK506和雷帕霉素特异结合的FK506结合蛋白(FK506 binding proteins, FKBPs)、能与环孢素A结合的亲环素(cyclophilins, CYPs)以及能与5-羟基-1,4-萘醌结合的parvulin蛋白(
2
2009
... 拟南芥叶绿体定位的亲免素多达17种, 包括11种FKBPs、5种CYPs和1种parvulin蛋白.定位在叶绿体的亲免素中, 只有AtFKBP13和AtCYP20-2具有PPlase活性, 但是AtFKBP13和AtCYP20-2的双突变体无论是在正常条件下、寒冷胁迫下还是强光胁迫下与野生型相比都没有太大差异(
... ).由于其它定位在叶绿体的亲免素都没有PPl- ase活性, 因此定位在叶绿体内的亲免素的功能可能与PPlase活性无关(
2
2015
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
... 在哺乳动物中, 亲免素的主要功能是与小的配体结合(如免疫抑制剂), 或者作为分子伴侣调节蛋白质的结构、活性和稳定性.在高等植物中, 亲免素通过与相关蛋白质相互作用调控生长发育; 并且FBKPs对免疫抑制剂不敏感, PPlase活性也不保守; 而亲环素对免疫抑制剂敏感, PPlase活性也较为保守.例如, AtROC1、LeCYP1和OsCYP2都具有PPlase活性(
1
1999
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
5
2015
... 亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(
... Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
... CYPs参与生长素、油菜素内酯、赤霉素和茉莉酸等激素的信号转导途径, 其中生长素、赤霉素和茉莉酸均以蛋白酶体降解抑制子的方式激活信号通路(
... ;
1
2002
... 植物的生长发育与激素密不可分.亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用.拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布.拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输.TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(
1
2013
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
1
2012
... FKBPs对热胁迫、盐胁迫、干旱及强光胁迫都有响应.拟南芥中2个高度同源的基因FKBP62 (AtROF1)与FKBP65 (AtROF2)参与植物耐热性调控.但是在热胁迫下, 2个基因的表达量和调节方式不同, AtROF1促进热激蛋白的表达而AtROF2抑制热激蛋白的表达(
2
2015
... 叶绿体内半胱氨酸合成过程中的重要酶(丝氨酸乙酰基转移酶(SAT1))能与AtCYP20-3相互作用, 指导SAT1的正确折叠, 保证半胱氨酸合成通路, 以适应各种逆境(
... 的信号传递过程(
1
2015
... 然而, 最近研究发现, AtCYP18-3 (ROC1)在植物免疫系统中具有负调控作用, ROC1能与效应子触发免疫(effector triggered immunity, ETI)的受体RIN4相互作用, 减少RIN4的磷酸化, 使抗性基因RPM1不能够识别效应因子, 从而不能激活ETI途径; 而大豆CYP1能激活疫霉属效应因子Avr3b的酶活性.这说明亲免素参与生物胁迫的方式较多, 其作用机制有待进一步研究(
1
2015
... CYPs在植物抵抗盐碱、干旱、寒冷和强光胁迫方面具有重要作用, 并且能够增加水稻分蘖, 从而提高产量.过量表达OsCYP2可以提高水稻种子的耐盐性, 并且OsCYP2可能通过调节抗氧化酶活性控制细胞内ROS的含量, 从而提高植株的耐盐性(
1
2010
... 在拟南芥开花信号途径中, FLC (FLOWERING LOCUS C)通过抑制成花信号FT (FLOWERING LO- CUS T)和转录因子基因SOC1 (SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1)的表达抑制开花.研究发现, 拟南芥AtCYP20-2与BZR1相互作用改变BZR1的构象, 调节FLD (FLOWERING LOCUS D)的表达; 过量表达CYP20-2使FLD表达量增加, 而FLD能抑制FLC染色质的甲基化和乙酰化, 从而导致开花提前(
2
2007
... 拟南芥突变体cyp71茎尖分生组织活性下降, 侧生器官发育异常, 花的形态发生改变, 根的生长受抑制(
... 出现异常表型(
1
2010
... 拟南芥突变体cyp71茎尖分生组织活性下降, 侧生器官发育异常, 花的形态发生改变, 根的生长受抑制(
1
2011
... 拟南芥突变体cyp71茎尖分生组织活性下降, 侧生器官发育异常, 花的形态发生改变, 根的生长受抑制(
2
2014
... 然而, 最近研究发现, AtCYP18-3 (ROC1)在植物免疫系统中具有负调控作用, ROC1能与效应子触发免疫(effector triggered immunity, ETI)的受体RIN4相互作用, 减少RIN4的磷酸化, 使抗性基因RPM1不能够识别效应因子, 从而不能激活ETI途径; 而大豆CYP1能激活疫霉属效应因子Avr3b的酶活性.这说明亲免素参与生物胁迫的方式较多, 其作用机制有待进一步研究(
... CYPs参与生长素、油菜素内酯、赤霉素和茉莉酸等激素的信号转导途径, 其中生长素、赤霉素和茉莉酸均以蛋白酶体降解抑制子的方式激活信号通路(
1
2013
... FKBPs对热胁迫、盐胁迫、干旱及强光胁迫都有响应.拟南芥中2个高度同源的基因FKBP62 (AtROF1)与FKBP65 (AtROF2)参与植物耐热性调控.但是在热胁迫下, 2个基因的表达量和调节方式不同, AtROF1促进热激蛋白的表达而AtROF2抑制热激蛋白的表达(
1
2006
... 拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用.拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(
1
2014
... 拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用.拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(
1
2013
... 拟南芥ROC1 (AtCYP18-3)与光敏色素、隐花色素共同参与幼苗的去黄化过程.突变体roc1和roc1-D能够改变转录因子BES1的磷酸化模式, 但ROC1不能与BES1相互作用; CYP18-3的作用机制可能与TWD1相似, 通过与BRI1和BAK1相互作用影响BES1的磷酸化(
1
2014
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
1
2005
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
1
2009
... FKBPs对热胁迫、盐胁迫、干旱及强光胁迫都有响应.拟南芥中2个高度同源的基因FKBP62 (AtROF1)与FKBP65 (AtROF2)参与植物耐热性调控.但是在热胁迫下, 2个基因的表达量和调节方式不同, AtROF1促进热激蛋白的表达而AtROF2抑制热激蛋白的表达(
1
2014
... 生物胁迫对作物生产具有严重危害.研究发现, 病原体入侵时, AtCYP19、AtCYP57和AtFKBP65的表达量显著提高.AtCYP19、AtCYP57和AtFKBP65基因的缺失突变体更易感染丁香假单胞菌和黄单胞杆菌, 突变导致对其病原体表面保守分子特征(pathogen associated molecular patterns, PAMPs)的识别有缺陷; 过量表达AtCYP19、AtCYP57和AtFKBP65的拟南芥对丁香假单胞菌的抵抗力增强; 并且AtCYP19、AtCYP57和AtFKBP65能够激活病原体相关分子模式触发免疫(PAMPs triggered immunity, PTI), 使胼胝质沉淀和过氧化氢积累以抵抗病原体(
1
2016
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
2
2013
... 在盐碱、干旱和强光等逆境胁迫下, 水稻Os- FKBP16-3的表达量增加, 将OsFKBP16-3转化拟南芥, 转基因植株对盐碱、干旱和强光的耐受性提高(
... ).在极端环境下, 植物体内会产生活性氧簇(ROS).FKBP16-3定位在叶绿体内, 其含有一段保守的氧化还原位点CxxxC, 能被ROS氧化形成二硫键; FKBP16-3可能通过修饰CxxxC调节氧化还原过程, 以减少逆境对植物的伤害(
1
2013
... 叶绿体内半胱氨酸合成过程中的重要酶(丝氨酸乙酰基转移酶(SAT1))能与AtCYP20-3相互作用, 指导SAT1的正确折叠, 保证半胱氨酸合成通路, 以适应各种逆境(
1
2009
... NADPH脱氢酶NDH复合体能够调节PSI的循环和叶绿体内的电子传递, NDH与PSI形成NDH-PSI复合体, AtFKBP16-2能与NADPH脱氢酶复合体相互作用, 从而稳定NDH-PSI复合体(
2
2014
... 在盐碱、干旱和强光等逆境胁迫下, 水稻Os- FKBP16-3的表达量增加, 将OsFKBP16-3转化拟南芥, 转基因植株对盐碱、干旱和强光的耐受性提高(
... 生物胁迫对作物生产具有严重危害.研究发现, 病原体入侵时, AtCYP19、AtCYP57和AtFKBP65的表达量显著提高.AtCYP19、AtCYP57和AtFKBP65基因的缺失突变体更易感染丁香假单胞菌和黄单胞杆菌, 突变导致对其病原体表面保守分子特征(pathogen associated molecular patterns, PAMPs)的识别有缺陷; 过量表达AtCYP19、AtCYP57和AtFKBP65的拟南芥对丁香假单胞菌的抵抗力增强; 并且AtCYP19、AtCYP57和AtFKBP65能够激活病原体相关分子模式触发免疫(PAMPs triggered immunity, PTI), 使胼胝质沉淀和过氧化氢积累以抵抗病原体(
1
1994
... 在哺乳动物中, 亲免素的主要功能是与小的配体结合(如免疫抑制剂), 或者作为分子伴侣调节蛋白质的结构、活性和稳定性.在高等植物中, 亲免素通过与相关蛋白质相互作用调控生长发育; 并且FBKPs对免疫抑制剂不敏感, PPlase活性也不保守; 而亲环素对免疫抑制剂敏感, PPlase活性也较为保守.例如, AtROC1、LeCYP1和OsCYP2都具有PPlase活性(
1
2000
... 拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用.拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(
1
2010
... 在哺乳动物中, FKBPs与转录因子相互作用较为常见, 在植物体内也发现FKBPs与转录因子相互作用调节细胞的分裂与分化.拟南芥突变体pas1 (pasticcion1)的细胞分裂异常、分化紊乱, 对野生型植株施加外源性细胞分裂素后出现与pas1相似的表型.研究发现, PAS1 (即FKBP72)的C末端与FAN转录因子(FKBP-associated NAC)相互作用, 促进了FAN对细胞分裂的抑制(
1
2011
... CYPs在植物抵抗盐碱、干旱、寒冷和强光胁迫方面具有重要作用, 并且能够增加水稻分蘖, 从而提高产量.过量表达OsCYP2可以提高水稻种子的耐盐性, 并且OsCYP2可能通过调节抗氧化酶活性控制细胞内ROS的含量, 从而提高植株的耐盐性(
2
2015
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
... ;
1
2014
... 小麦TaFKBP16-1和TaFKBP16-3在光系统I (PSI)的组装过程中扮演着重要角色.TaFKBP16-1与PSI的1个亚基PsaL (PSI subunit L)相互作用, TaFK- BP16-3与Thf1 (Thylakoid Formation 1)和APO2 (Os02g50010)存在相互作用, 因此TaFKBP16-1和TaFKBP16-3可能调控PSI的组装(
1
2009
... NADPH脱氢酶NDH复合体能够调节PSI的循环和叶绿体内的电子传递, NDH与PSI形成NDH-PSI复合体, AtFKBP16-2能与NADPH脱氢酶复合体相互作用, 从而稳定NDH-PSI复合体(
1
2008
... 拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用.拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(
1
2011
... 近年来的研究表明, FKBPs与多种激素关系密切.拟南芥中调节生长素运输的TWD1与人类FKBP38结构相似, 人类FKBP38的PPlase活性能够促进囊性纤维化跨膜传导调节蛋白(CFTR)的翻译后折叠, 并且其TPR结构域与Hsp90相互作用能够抑制PPlase活性, 对CFTR的合成进行负反馈调节(
1
2006
... 在哺乳动物中, FKBPs与转录因子相互作用较为常见, 在植物体内也发现FKBPs与转录因子相互作用调节细胞的分裂与分化.拟南芥突变体pas1 (pasticcion1)的细胞分裂异常、分化紊乱, 对野生型植株施加外源性细胞分裂素后出现与pas1相似的表型.研究发现, PAS1 (即FKBP72)的C末端与FAN转录因子(FKBP-associated NAC)相互作用, 促进了FAN对细胞分裂的抑制(
1
2015
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
1
2012
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
1
2012
... 拟南芥ROC1 (AtCYP18-3)与光敏色素、隐花色素共同参与幼苗的去黄化过程.突变体roc1和roc1-D能够改变转录因子BES1的磷酸化模式, 但ROC1不能与BES1相互作用; CYP18-3的作用机制可能与TWD1相似, 通过与BRI1和BAK1相互作用影响BES1的磷酸化(
1
2012
... 拟南芥AtFKBP20-2和AtCYP38在PSII的组装和维持中发挥重要作用.拟南芥Atfkbp20-2突变体生长缓慢, 光系统II (PSII)复合体含量下降, PSII的单体和二聚体比野生型含量多, 说明FKBP20-2对PSII的组装和稳定有一定的影响(
2
2015
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
... 基因沉默、表观遗传学调控和转录水平的调控对植物的生长发育产生重要影响.拟南芥cyp40突变体叶片数量减少和形态改变与microRNA活性减弱有关.成熟的microRNA能够与RNA诱导沉默复合体(RNA- induced silencing complex, RISC)结合, 形成具有切割活性的RNA沉默复合物, 该复合物与靶基因互补配对使靶基因沉默或抑制靶基因的翻译.AGO1 (ARGONAUTE1)是RISC的重要组成部分(
2004
1
2009
... 基因沉默、表观遗传学调控和转录水平的调控对植物的生长发育产生重要影响.拟南芥cyp40突变体叶片数量减少和形态改变与microRNA活性减弱有关.成熟的microRNA能够与RNA诱导沉默复合体(RNA- induced silencing complex, RISC)结合, 形成具有切割活性的RNA沉默复合物, 该复合物与靶基因互补配对使靶基因沉默或抑制靶基因的翻译.AGO1 (ARGONAUTE1)是RISC的重要组成部分(
1
2013
... 植物的生长发育与激素密不可分.亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用.拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布.拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输.TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(
2
2014
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
... 目前, 关于parvulin蛋白的研究相对较少.研究表明, parvulin蛋白能调节植物的生长发育和抗逆性.拟南芥依赖磷酸化的AtPin1有1个PPlase结构域, 能够改变PIN1的构象, 使其极性定位发生改变, 进而影响生长素的运输和根的向地性; 另外, Pin1At与AGL24 (AGAMOUS-LIKE 24)和SOC1相互作用改变其构象, 调节开花时间(
1
2008
... 高等植物体内FKBPs数量众多, 根据结构可将其分为2类: 一类是单结构域成员, 这类FKBPs仅含有1个FKB结构域(FKBD), 具有PPIase活性; 另一类是具有多个结构域的FKBPs, 一般包括3个FKBD、TPR结构域(tetratricopeptide repeat domains)和1个C末端的钙调蛋白结构域(calmodulin-binding domains, CaM- BDs) (
1
2010
... 目前, 关于parvulin蛋白的研究相对较少.研究表明, parvulin蛋白能调节植物的生长发育和抗逆性.拟南芥依赖磷酸化的AtPin1有1个PPlase结构域, 能够改变PIN1的构象, 使其极性定位发生改变, 进而影响生长素的运输和根的向地性; 另外, Pin1At与AGL24 (AGAMOUS-LIKE 24)和SOC1相互作用改变其构象, 调节开花时间(
1
2015
... CYPs在植物抵抗盐碱、干旱、寒冷和强光胁迫方面具有重要作用, 并且能够增加水稻分蘖, 从而提高产量.过量表达OsCYP2可以提高水稻种子的耐盐性, 并且OsCYP2可能通过调节抗氧化酶活性控制细胞内ROS的含量, 从而提高植株的耐盐性(
4
2010
... 植物的生长发育与激素密不可分.亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用.拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布.拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输.TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(
... ;
... 亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(
... Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (
2
2016
... 目前, 关于parvulin蛋白的研究相对较少.研究表明, parvulin蛋白能调节植物的生长发育和抗逆性.拟南芥依赖磷酸化的AtPin1有1个PPlase结构域, 能够改变PIN1的构象, 使其极性定位发生改变, 进而影响生长素的运输和根的向地性; 另外, Pin1At与AGL24 (AGAMOUS-LIKE 24)和SOC1相互作用改变其构象, 调节开花时间(
... 在哺乳动物中, 亲免素的主要功能是与小的配体结合(如免疫抑制剂), 或者作为分子伴侣调节蛋白质的结构、活性和稳定性.在高等植物中, 亲免素通过与相关蛋白质相互作用调控生长发育; 并且FBKPs对免疫抑制剂不敏感, PPlase活性也不保守; 而亲环素对免疫抑制剂敏感, PPlase活性也较为保守.例如, AtROC1、LeCYP1和OsCYP2都具有PPlase活性(
2
2016
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
... CYPs在植物抵抗盐碱、干旱、寒冷和强光胁迫方面具有重要作用, 并且能够增加水稻分蘖, 从而提高产量.过量表达OsCYP2可以提高水稻种子的耐盐性, 并且OsCYP2可能通过调节抗氧化酶活性控制细胞内ROS的含量, 从而提高植株的耐盐性(
2011
3
2013
... 亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(
... Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (
... 在拟南芥开花信号途径中, FLC (FLOWERING LOCUS C)通过抑制成花信号FT (FLOWERING LO- CUS T)和转录因子基因SOC1 (SUPPRESSOR OF OVEREXPRESSION OF CONSTANS1)的表达抑制开花.研究发现, 拟南芥AtCYP20-2与BZR1相互作用改变BZR1的构象, 调节FLD (FLOWERING LOCUS D)的表达; 过量表达CYP20-2使FLD表达量增加, 而FLD能抑制FLC染色质的甲基化和乙酰化, 从而导致开花提前(
5
2016
... 从20世纪80年代初发现亲免素以来, 已有23个拟南芥(Arabidopsis thaliana)、29个水稻(Oryza sativa)、4个酵母(Saccharomyces cerevisiae)、18个人类(Homo sapiens) FKBPs被鉴定出来(
... 亲免素与相关蛋白质相互作用调节下游信号的传递
(A) ABCB蛋白在TWD1的帮助下正确定位于细胞膜, 将细胞质内的生长素运出; 这个过程可能受BR (brassinosteroid)的影响(
... Immunophilins regulate downstream signaling by interacting with related proteins
(A) TWD1 helps the correct secretion of ABCB transporters to the plasma membrane, thus regulating ABCB-mediated auxin transport; Brassinosteroid might affect this process (
... 最新研究显示, 拟南芥TWD1也参与油菜素甾醇(brassinosteroid, BR)的信号传递.在根生长和下胚轴伸长过程中, twd1-2以及twd1-5对油菜素内酯(brassinolide, BL)的敏感程度降低; 在twd1-5中磷酸化和未磷酸化的BES1含量均降低.实验结果显示TWD1在植物体内能与BRI1和BAK1相互作用.以上表明TWD1与BRI1和BAK1相互作用影响了BR信号转导(
... 近年来的研究表明, FKBPs与多种激素关系密切.拟南芥中调节生长素运输的TWD1与人类FKBP38结构相似, 人类FKBP38的PPlase活性能够促进囊性纤维化跨膜传导调节蛋白(CFTR)的翻译后折叠, 并且其TPR结构域与Hsp90相互作用能够抑制PPlase活性, 对CFTR的合成进行负反馈调节(
1
2013
... 拟南芥CYPs能调节生长素的运输和信号转导, 从而影响植物的生长和组织器官的发育.早期的研究表明, 拟南芥PP2A (heterotrimeric protein phosphatase 2A)的1个亚基A能够与AtCYP20-1相互作用调节PP2A的活性, 影响生长素转运蛋白的正确定位, 从而影响生长素的运输和植物生长(
1
2016
... 植物的生长发育与激素密不可分.亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用.拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布.拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输.TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(
1
2015
... 植物的生长发育与激素密不可分.亲免素与生长素、油菜素内酯、细胞分裂素等都具有复杂而密切的联系, 在植物的生长发育过程中具有重要作用.拟南芥AtFKBP42 (TWISTED DWARF1, TWD1)参与多种激素的运输与分布.拟南芥ABC (ATP binding cassette)家族中的ABCB1、ABCB4和ABCB19等成员参与生长素运输.TWD1的N末端PPlase-like结构域与ABCB1和ABCB19的C末端相互作用, 揭示了TWD1可以调控生长素的运输和分布(

{kind=link}
{kind=link}