杨艳 1 , 2 ,
张海琴 1 ,
凡星 1 ,
沙莉娜 1 ,
康厚扬 1 ,
王益 1 ,
周永红 1 , * , 1 四川农业大学小麦研究所, 成都 6111302 西华师范大学环境科学与工程学院, 南充 637009Yang Yan 1 , 2 ,
Zhang Haiqin 1 ,
Fan Xing 1 ,
Sha Lina 1 ,
Kang Houyang 1 ,
Wang Yi 1 ,
Zhou Yonghong 1 , * , 1 Triticeae Research Institute, Sichuan Agricultural University, Chengdu 611130, China2 College of Environmental Science and Engineering, China West Normal University, Nanchong 637009, China引用本文 杨艳 ,
张海琴 ,
凡星 ,
沙莉娜 ,
康厚扬 ,
王益 ,
周永红 . 偃麦草属植物醇溶蛋白和谷蛋白多态性及系统学研究. , 2017, 52(5): 579-589
贡献者
* 通讯作者。E-mail: zhouyh@sicau.edu.cn 基金资助
国家自然科学基金(No.31270243) ;
接受日期:2016-07-31接受日期:2016-11-11网络出版日期:2017-09-1
-->Copyright
2017《植物学报》编辑部
Contributors
* Author for correspondence. E-mail: zhouyh@sicau.edu.cn History
Received :
Accepted :
Online :
摘要: 运用酸性聚丙烯酰胺凝胶电泳(A-PAGE)和十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)对偃麦草属(Elytrigia ) 24个物种的醇溶蛋白和谷蛋白进行了研究。以中国春为参照, 按醇溶蛋白和谷蛋白电泳图谱条带迁移距离大小和条带多态性对供试材料进行聚类分析。结果显示, 24份供试材料均呈现不同的醇溶蛋白和谷蛋白电泳图谱, 共分离出醇溶蛋白带纹83条和谷蛋白带纹53条, 多态性均达100%, 并且在相同染色体组组成的物种中, 染色体数目越多, 其蛋白带纹越多; 偃麦草属24个物种的醇溶蛋白和谷蛋白图谱均有明显差异, 蛋白图谱可作为鉴定偃麦草属物种的指纹图谱。聚类分析结果显示, 在材料间的醇溶蛋白遗传相似性系数为0.66时, 24份材料被划分为6个主要类群, 含E和St基因组的物种具有较近的亲缘关系; 在谷蛋白遗传相似性系数为0.62时, 24份材料被聚为4大类。聚类结果表明, 染色体组成未知的物种Et. pachynera 可能为异源多倍体物种。 关键词 :
偃麦草属 ;
醇溶蛋白 ;
谷蛋白 ;
多态性 Abstract :
The polymorphism of gliadin and glutelin in 24 species in Elytrigia was studied by using A-PAGE and SDS-PAGE. With reference to Chinese Spring, cluster analysis was applied in all the tested materials based on the stripe migration distance and stripe polymorphism on electrophoretography. Different electrophoresis patterns of gliadin and glutelin appeared in all the 24 tested materials, and 83 gliadin and 53 glutelin bands were extracted, which indicated 100% polymorphism. The Elytrigia species could be an excellent germplasm source for improving the wheat proteins. Furthermore, the number of bands increased with increasing chromosome number within the same genome. The gliadin and glutelin patterns from the 24 Elytrigia species showed differences, which could be used as a fingerprint for identifying different species from Elytrigia . The 24 species were divided into six groups with 0.66 genetic similarity coefficient for gliadin, and species carrying E and St genes had closer phylogenetic relationship. However, the 24 species were divided into 4 groups with 0.62 genetic similarity coefficient for glutelin. Although the genome composition of Et . pachynera is unknown, the cluster results implied that it is an allopolyploid species. Key words :
Elytrigia ;
gliadin ;
glutelin ;
polymorphism 偃麦草属(
Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(
Tzvelev, 1976 )。偃麦草属植物是小麦(
Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性。该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(
陈默君和贾慎修, 2002 ;
吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(
马渐新等, 1999 ;
王洪刚等, 2000 ;
吕伟东等, 2007 )。
Desvaux (1810) 以
Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括E
e 、E
b 、St、ESt和EStP等染色体组组成。根据
Dewey (1984) 和
Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种
Et. repens 应划归披碱草属(
Elymus ) (
De- wey, 1980 ;
Assadi and Runemark, 1995 )。由此可见, 偃麦草属的分类和系统地位仍然存在很大争议。
醇溶蛋白和谷蛋白是小麦及其亲缘植物的主要种子贮藏蛋白, 发育时期生理因素和外界环境均不影响其组成, 其电泳带纹特征完全受基因控制, 因此基因表达的差异能够通过电泳得到真实的体现(
Draper, 1987 )。醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(
Kawaura et al., 2005 )。普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(
Metakovsky et al., 1986 )。谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS)。高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(
Jiang and Dong, 2000 )。目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(
Nakamura, 2000 ;
高梅等, 2007 ;
Asfaw, 2008 ;
郑青焕等, 2016 ); 长穗偃麦草(
Elytrigia elongata (Host) Nev- ski.)的分子遗传学(
王际睿等, 2004 ;
吴卫东等, 2010 )和细胞学(
Lawrence and Shepherd, 1981 ;
唐朝晖等, 2004 )特性。关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道。本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据。
1 材料与方法1.1 供试材料选取偃麦草属(
Elytrigia L.)植物24种(24份)作为供试材料。供试材料的种名、基因组组成、编号及来源见
表1 。其中, 含有PI编号的材料来源于美国国家资源库, Y编号的来源于四川农业大学小麦研究所。为确保取材的准确性, 在蕾期进行了套袋处理, 取材后通过细胞学方法确定其倍性, 并随机选取5粒种子, 分别用于A-PAGE和SDS-PAGE实验。以中国春(Chinese Spring, Cs)作为醇溶蛋白和谷蛋白电泳的对照材料。
表1 Table 1 表1 表1 供试偃麦草属材料
Table 1 The materials of
Elytrigia used in this study
No. Species Genome 2n Accession No. Origin 1 Elytrigia bessarabica (Savul & Rayss) DubovikE 2x PI531711 Estonia 2 Et. caespitosa ssp. nodosa (Nevski) TzvelESt 4x PI531734 Estonia 3 Et. caespitosa (C. Koch) NevskiESt 4x PI547311 Leningrad Russian Federation 4 Et. elongata (Host) NevskiE 2x PI578682 Nebraska United States 5 Et. farcta (Viv.) HolubEEE 6x PI531727 France 6 Et. geniculata (Nevski) TzvelevStSt 4x PI639753 Russian Federation 7 Et. geniculata ssp. pruinifera (Nevski) Tzvelev- 6x PI547374 Russian Federation 8 Et. intermedia (Host) NevskiEESt 6x PI223668 Iran 9 Et. intermedia ssp. intermedia (Host) NevskiEESt 6x PI229917 Iran 10 Et. kosaninii (Nábélek) DubovikSt 2x PI237636 Turkey 11 Et. libanotica (Hackel) HolubSt 2x PI229583 Iran 12 Et. pachynera Prokudin- 6x PI440059 Former Soviet Union 13 Et. podpera (Nábělek) HolubEEE 6x PI228387 Iran 14 Et. pontica (Podp.) HolubEEEEE 10x PI636523 Argentina 15 Et. pungens (Pers.) TutinEStStP 8x PI547268 Leningrad Russian Federation 16 Et. pycnantha (Godr.) Á. LöveEStP 6x PI531741 France 17 Et. rechingeri (Runemark) HulubEE 4x PI531745 Greece 18 Et. repens (L.) NevskiStStH 6x Y0814 China 19 Et. scirpea (K. Presl) HolubEE 4x PI531749 Italy 20 Et. scythica (Nevski) NevskiESt 4x PI283272 Former Soviet Union 21 Et. stipifolia (Czern. ex Nevski) NevskiSt 2x PI313960 Former Soviet Union 22 Et. strigosa (M. Bieb.) NevskiStSt 4x PI531752 Estonia 23 Et. tauri (Boiss. & Bal.) TzvelSt 2x PI401330 Iran 24 Et. varnensis (Velen.) HolubEEEEStSt 12x PI281863 Germany
- 表示该物种染色体组成未知。- indicate the genome composition is unknown. 表1
供试偃麦草属材料
Table 1
The materials of
Elytrigia used in this study
1.2 方法1.2.1 醇溶蛋白的提取及电泳
将单粒种子碾碎后, 加入1.5 mL离心管中, 添加70 μL样品提取液(2-氯乙醇25%、甲基绿0.05%和丙三醇5%), 振荡1小时, 室温下静置过夜。1 500 ×
g 离心5分钟, 取上清液, 置于4˚C冰箱保存备用。电泳方法采用酸性聚丙烯酰胺凝胶电泳(A-PAGE), 电泳仪器为Bio-Rad公司的PROTEAN
® II Xiell型电泳槽, 上样量为5 µL, 稳压电泳。200 V定压电泳30分钟后, 400 V定压继续电泳2.5小时。室温保持在25-28˚C, 通过冷水循环保持电泳体系内部温度为4˚C。根据
Kawaura等(2005) 的分区方法对醇溶蛋白电泳谱带进行分区。
1.2.2 谷蛋白的提取及电泳
取1粒完整、无病害种子, 研磨至粉末状后倒入1.5 mL离心管中, 加入70 μL提取液(0.062 5 mol·L
-1 的Tris-HCl (pH6.8)、SDS 2%、丙三醇10%、β-巯基乙醇3%和溴酚蓝0.002%), 振荡1小时, 100˚C水浴5分钟, 期间轻轻颠倒混合2-3次。冷却至室温, 1 500 ×
g 离心5分钟, 将上清移至1.5 mL离心管中, 4˚C保存备用。采用十二烷基硫酸钠-聚丙烯酰胺凝胶电泳法(SDS-PAGE)进行电泳, 电泳仪器为北京六一仪器厂的DYY-12型电泳仪, 上样量为2 µL, 电流90 mA, 室温下进行单向一步电泳, 获得并观察谷蛋白谱带。
1.2.3 数据处理
电泳图谱按条带有(记为1)无(记为0)的方法分别赋值。按照Jaccard (1908)的方法计算材料间的Jaccard遗传相似系数(
GS ), 即
GS =2
N ij /(
N i +
N j ), 其中,
N i 代表材料i的条带数目,
N j 代表材料j的条带数目,
N ij 表示材料i和j中均出现的条带数目。利用
GS 按不加权成 对群算术平均法(UPGMA)进行聚类, 用NTSYSPC 2.10e软件进行数据统计分析。
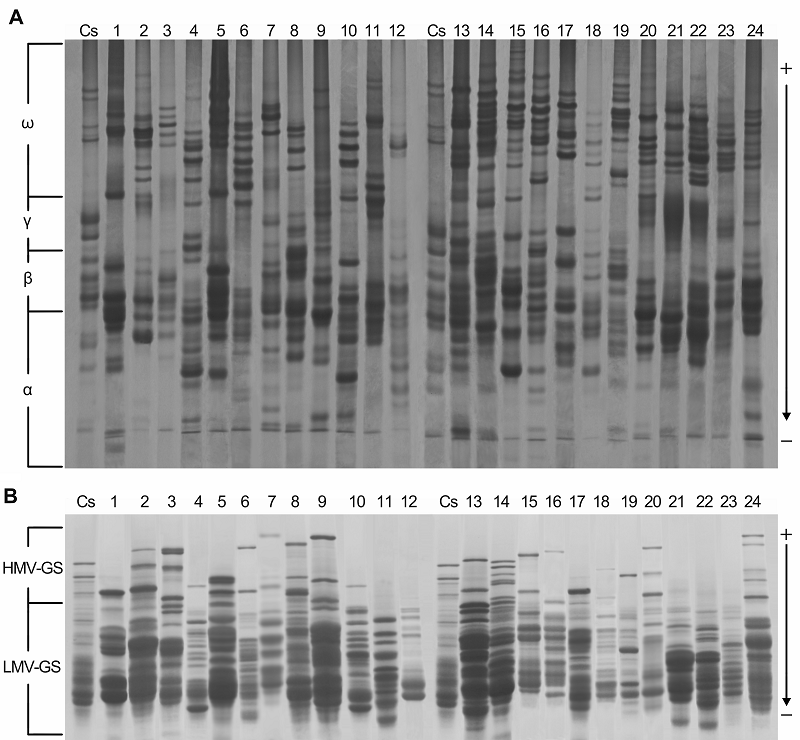
2 结果与讨论2.1 醇溶蛋白和谷蛋白的多态性及图谱差异比较以中国春为对照, 24份偃麦草属植物材料的醇溶蛋白和谷蛋白电泳图谱见
图1 , 条带数见
表2 。由
图1 可知, 供试的24份材料间具有数量和染色深浅不同的醇溶蛋白和谷蛋白带纹, 它们分别出现了24种不同的醇溶蛋白和谷蛋白电泳图谱, 均表现出较大变异, 易于区分, 即两种蛋白均具有丰富的遗传多样性, 这是蛋白相应编码基因位点多态性的表现特征。因此蛋白图谱能够作为鉴定偃麦草属植物的指纹图谱, 用于区分和鉴定不同来源的偃麦草属物种。
表2 Table 2 表2 表2 偃麦草属24份材料的醇溶蛋白和谷蛋白带纹数
Table 2 The number of gliadin bands and glutelin bands in 24 accessions of
Elytrigia No. The num- The num- No. The number of The number of 1 14 14 13 22 20 2 19 14 14 31 22 3 14 14 15 25 15 4 23 16 16 30 10 5 15 18 17 16 16 6 20 20 18 19 17 7 22 17 19 22 16 8 24 19 20 28 17 9 22 14 21 12 16 10 20 15 22 19 19 11 16 13 23 17 15 12 16 11 24 22 15
The accession number see Table 1 . 材料序号同表1 。 表2
偃麦草属24份材料的醇溶蛋白和谷蛋白带纹数
Table 2
The number of gliadin bands and glutelin bands in 24 accessions of
Elytrigia 图1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-579/img_1.png <b>图1</b> 偃麦草属24份材料的醇溶蛋白A-PAGE电泳图谱(A)和谷蛋白SDS-PAGE电泳图谱(B)<br/>Cs: 中国春小麦。其余材料序号同<xref ref-type="table" rid="T1-1674-3466-52-5-579">表1</xref>。<br/><b>Figure 1</b> A-PAGE gliadin electyophoregram result (A) and SDS-PAGE glutelin electyophoregram result (B) of 24 accessions of <i>Elytrigia<br/></i>Cs: China Spring. The accession number see <xref ref-type="table" rid="T1-1674-3466-52-5-579">Table 1</xref>. Figure 1 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-579/img_1.png <b>图1</b> 偃麦草属24份材料的醇溶蛋白A-PAGE电泳图谱(A)和谷蛋白SDS-PAGE电泳图谱(B)<br/>Cs: 中国春小麦。其余材料序号同<xref ref-type="table" rid="T1-1674-3466-52-5-579">表1</xref>。<br/><b>Figure 1</b> A-PAGE gliadin electyophoregram result (A) and SDS-PAGE glutelin electyophoregram result (B) of 24 accessions of <i>Elytrigia<br/></i>Cs: China Spring. The accession number see <xref ref-type="table" rid="T1-1674-3466-52-5-579">Table 1</xref>. 在新窗口打开 下载 下载原图ZIP 生成PPT 图1
偃麦草属24份材料的醇溶蛋白A-PAGE电泳图谱(A)和谷蛋白SDS-PAGE电泳图谱(B)
Cs: 中国春小麦。其余材料序号同
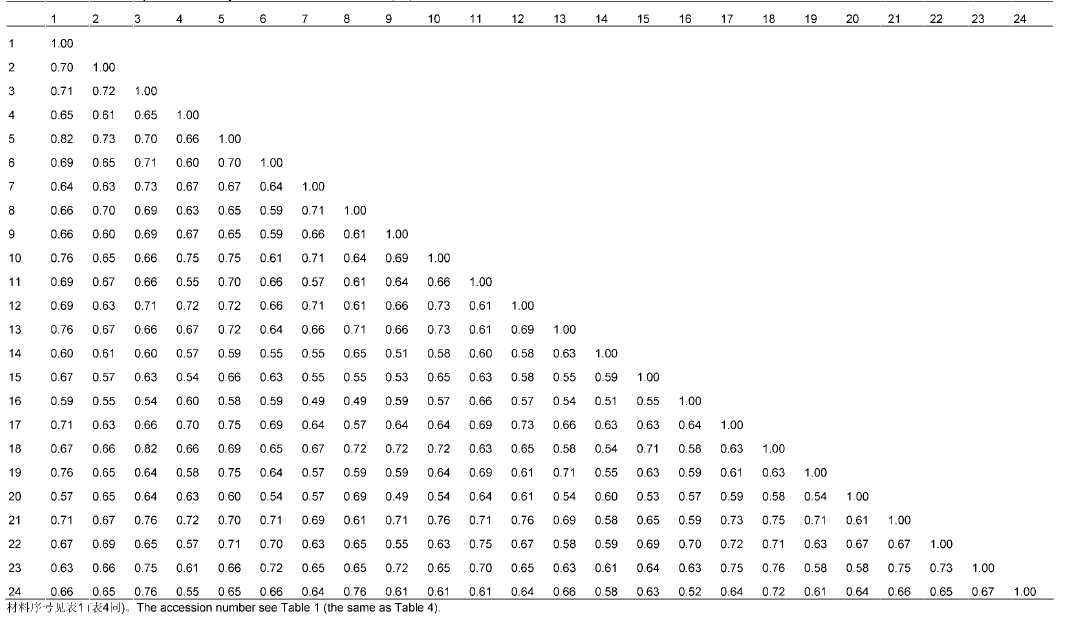
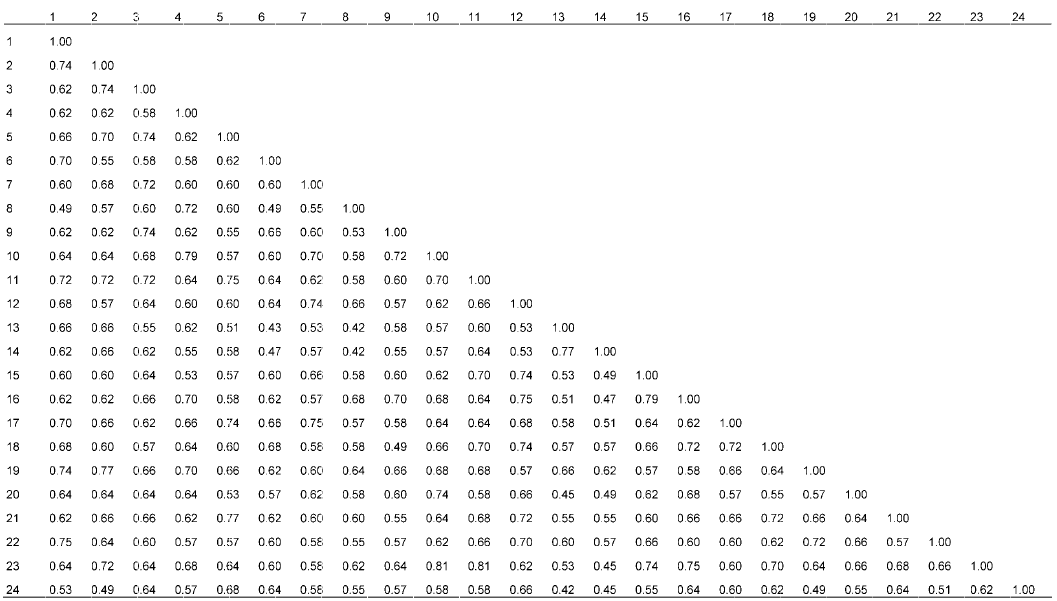
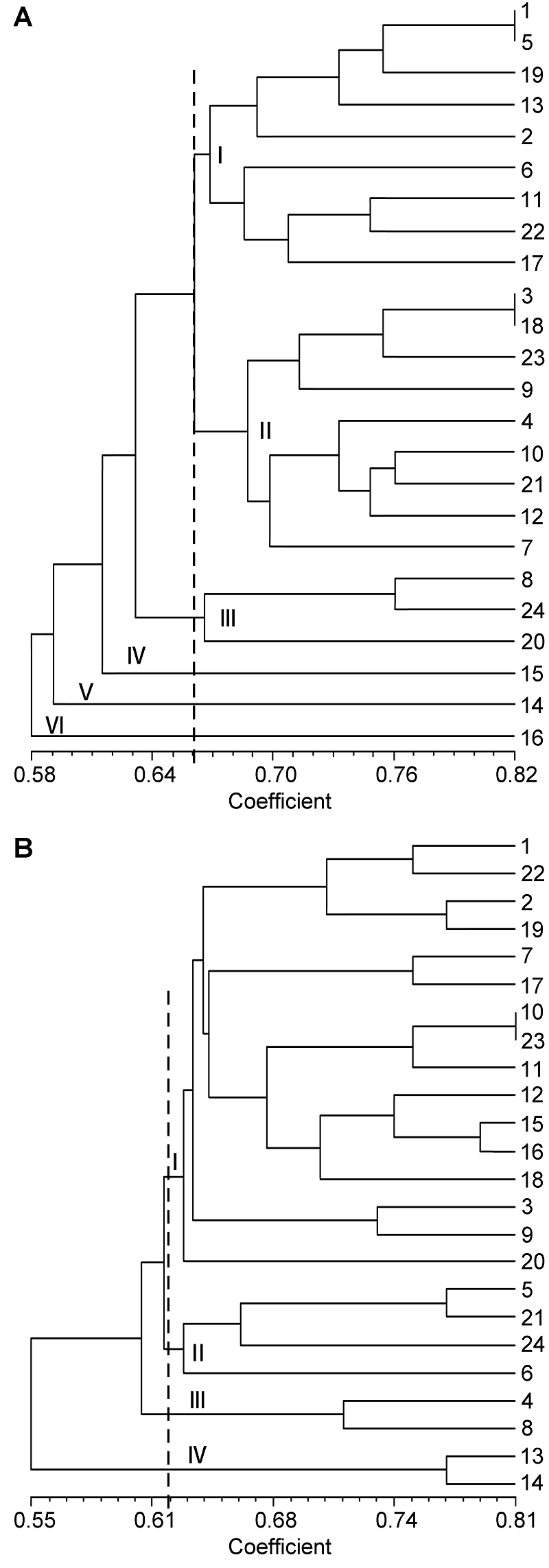
表1。Elytrigia Cs: China Spring. The accession number see Table 1.图1A 可知, 醇溶蛋白带纹主要分布在ω和β区, α区也存在较多带纹, γ区的带纹则相对较少。在83条迁移率不同的带纹中, 无任何1条带纹为24份材料所共有, 即多态性达100%, 表明该属植物的醇溶蛋白多态性极高, 其醇溶蛋白遗传差异较大。同时, 从图1A 中也可看出, 含有St基因组的物种, 其醇溶蛋白带纹主要分布在ω和β区, γ和α区的带纹相对较少; 含E基因组的物种, 其醇溶蛋白带纹主要分布在ω和α区, β区也存在较多的带纹, 而γ区的带纹则相对较少; 含有ESt和EStP染色体组的异源多倍体物种中, 醇溶蛋白带纹主要分布于ω和β区, α区具有较多带纹, 而γ区的带纹相对较少。但也有少数例外, 如Et. intermedia ssp. intermedia 的醇溶蛋白带纹在α、β和γ区分布较多, 而在ω区的高分子量醇溶蛋白却相对较少; 以及Et. pachynera 的蛋白带纹在α区分布较多, 在β、γ和ω区却非常少。图1B 显示, 谷蛋白主要是LMW-GS, 而HMW-GS较少。在53条迁移率不同的谷蛋白带纹中, 无任何1条带纹为24份材料所共有, 即多态性为100%, 表明偃麦草属植物的谷蛋白多态性极高, 其谷蛋白遗传差异较大。同时, 图1B 显示, 含有E和EStP基因组的物种谷蛋白带纹主要是LMW- GS, HMW-GS几乎没有; 含有St和ESt基因组的物种, 其谷蛋白主要为LMW-GS, HMW-GS较少, 但Et. elongata 、Et. bessarabica 、Et. rechingeri 和Et. scirpea 几乎没有高分子量谷蛋白。表3 )和谷蛋白(表4 )遗传相似性系数(Jaccard系数)分析结果显示, 供试材料的醇溶蛋白遗传相似性系数变异范围在0.49- 0.82之间, 平均为0.65; 谷蛋白遗传相似性系数变异范围在0.42-0.81之间, 平均为0.62。以上结果进一步表明偃麦草属植物具有较丰富的蛋白质遗传多样性。表3 Table 3 表3 表3 24份偃麦草属植物醇溶蛋白的遗传和相似性系数 Table 3 Genetic similarity coefficients of gliadin form 24 accessions of Elytnigia表4 Table 4 表4 表4 24份偃麦草属植物谷蛋白的遗传相似性系数 Table 4 Genetic similarity coefficients of glutein from 24 accessions of ElytnigiaEt. intermedia ssp. intermedia 和Et. scythica 、Et. intermedia 和Et. pycnantha 以及Et. geniculata ssp. pruinifera 和Et. pycnantha 之间的相似性系数均为0.49, 表明它们的醇溶蛋白差异均较大。Et. bessarabica 与Et. farcta 、Et. caespitosa 与Et. repens 之间的相似性系数均高达0.82, 表明其醇溶蛋白遗传差异较小。从谷蛋白来看, Et. podpera 和Et. intermedia 、Et. pontica 和Et. intermedia , 以及Et. podpera 和Et. varnensis 之间的相似性系数均为0.42, 表明它们之间的谷蛋白遗传差异均较大。Et. libanotica 与Et. tauri 、Et. tauri 与Et. kosaninii 之间的相似性系数最大(达0.81)。 Et. kosaninii 与Et. elongata 、Et. pycnantha 与Et. pungens 之间的谷蛋白差异也较小, 相似性系数为0.79。杨瑞武等, 2004 ), 结果(图2A )显示, 醇溶蛋白遗传相似性系数为0.66时, 所有材料被聚为6大类。第I类包括Et. bessarabica 、Et. caespitosa ssp. nodosa 、Et. farcta 、Et. geniculata 、Et. libano- tica 、Et. podpera 、Et. rechingeri 、Et. scirpea 和Et. strigosa ; 第II类包括Et. caespitosa 、Et. elongata 、Et. geniculata ssp. pruinifera 、Et. intermedia ssp. intermedia 、Et. kosaninii 、Et. pachynera 、Et. repens 、Et. stipifolia 和Et. tauri ; 第III类包括Et. intermedia 、Et. scythica 和Et. varnensis ; Et. pungens 单独聚为第IV类; Et. pontica 单独聚为第V类; Et. pycnantha 单独聚为第VI类。在第I类中, Et. geniculate 、Et. libano- tica 、Et. rechingeri 和Et. strigosa 先聚在一起, 再与Et. bessarabica 、Et. casepitosa ssp. nodosa 、Et. 图2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-579/img_4.png <b>图2</b> 24份偃麦草属材料的醇溶蛋白(A)和谷蛋白(B)聚类图<br/>材料序号同<xref ref-type="table" rid="T1-1674-3466-52-5-579">表1</xref>。<br/><b>Figure 2</b> The dendrogram of gliadin (A) and glutelin (B) from 24 accessions of <i>Elytrigia<br/></i>The accession number see <xref ref-type="table" rid="T1-1674-3466-52-5-579">Table 1</xref>. Figure 2 https://www.chinbullbotany.com/article/2017/1674-3466/1674-3466-52-5-579/img_4.png <b>图2</b> 24份偃麦草属材料的醇溶蛋白(A)和谷蛋白(B)聚类图<br/>材料序号同<xref ref-type="table" rid="T1-1674-3466-52-5-579">表1</xref>。<br/><b>Figure 2</b> The dendrogram of gliadin (A) and glutelin (B) from 24 accessions of <i>Elytrigia<br/></i>The accession number see <xref ref-type="table" rid="T1-1674-3466-52-5-579">Table 1</xref>. 在新窗口打开 下载 下载原图ZIP 生成PPT 表1。Elytrigia The accession number see Table 1.farcta 、Et. podpera 和Et. scirpea 聚类。在第II类中, Et. caespitosa 、Et. intermedia ssp. intermedia 、Et. repens 和Et. tauri 先聚在一起, 再与Et. elongata 、Et. geniculata ssp. pruinifera 、Et. kosaninii 、Et. pachy- nera 和Et. stipifolia 聚在一起。杨瑞武等, 2004 ), 结果(图2B )显示, 谷蛋白遗传相似性系数为0.62时, 所有材料被聚为4大类: 第I类包括Et. bessarabica 、Et. caespitosa ssp. nodosa 、Et. cae- spitosa 、Et. elongata 、Et. geniculata ssp. pruinifera 、Et. intermedia ssp. intermedia 、Et. kosaninii 、Et. libanotica 、Et. pachynera 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 、Et. strigosa 和Et. tauri ; 第II类包括Et. farcta 、Et. geniculata 、Et. stipifolia 和Et. varnensis ; Et. intermedia 单独聚为第III类; 第IV类包括Et. podpera 和Et. pontica 。在第I类中, Et. elongata 、Et. kos- aninii 、Et. libanotica 、Et. pachynera 、Et. pungens 、Et. pycnantha 、Et. repens 、Et. scythica 和Et. tauri 先聚在一起, 然后与Et. bessarabica 、Et. caespitosa ssp. nodosa 、Et. scirpea 和Et. strigosa 聚类, 再与Et. geniculata ssp. pruinifera 及Et. rechingeri 聚类, 最后与Et. caespitosa 及Et. intermedia ssp. intermedia 聚在一起。张学勇等, 1995 ; 董超华等, 2003 )。本研究分析了取自偃麦草属24个种24份材料的蛋白电泳图谱, 结果表明, 醇溶蛋白和谷蛋白电泳图谱在物种间差异明显, 表明它们能够作为偃麦草属植物鉴定的指纹图谱, 用于区分和鉴定偃麦草属物种。Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究。杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系。张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位。王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据。翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱。杨瑞武等(2000) 通过研究披碱草属植物的醇溶蛋白, 发现小麦族植物的醇溶蛋白带纹多少与染色体数目有关。本研究(表2 )表明, 偃麦草属含有相同染色体组的物种中, 染色体数目越多, 其醇溶蛋白和谷蛋白带纹也越多; 而含有不同染色体组的物种之间, 其染色体数目与醇溶蛋白和谷蛋白带纹数目无关。因此, 我们认为仅按照形态学标准划分传统偃麦草属的范围是不合理的。图2 )来看, 含有相同基因组的物种基本先聚在一起, 然后与亲缘关系较近的其它基因组的物种聚类。就醇溶蛋白聚类结果(图2 A)而言, 第I类中, 含有St基因组的物种(Et. libanotica 、Et. geniculata 和Et. strigosa )与含E基因组的物种(Et. bessarabica 、Et. farcta 、Et. podpera 、Et. rechingeri 和Et. scirpea )聚在一起, 说明在偃麦草属中含有E和St基因组物种的醇溶蛋白的遗传相似度很高, 也表明这些物种之间亲缘关系较近, 这与前人的细胞生物学(于海清, 2007 )和分子系统学(Mason-Gamer et al., 2002 ; Seberg and Frederiksen, 2008 ; Kim et al., 2010 )研究结果一致。第II类中, St基因组的物种有Et. tauri 、Et. kosaninii 和Et. stipifolia , 说明具有St基因组的物种分别包含在第I和II类中, 表明含有St基因组 的不同物种间存在一定程度的分化(Stebbins and Pun, 1953 ), 从而导致醇溶蛋白带纹的差异性较大。我们发现含ESt基因组的物种Et. caespitosa ssp. nodosa 在醇溶蛋白图谱中与含E基因组的物种Et. podpera 聚类在一起, 相似性系数为0.67 (表3 ); 在谷蛋白电泳图谱中, 与含E基因组的物种Et. scirpea 聚在一起, 相似性系数为0.77 (表4 ), 表明Et. caespitosa ssp. nodosa 与含有E基因组的物种的亲缘关系较密切。Et. repens (StH)、Et. pungens (EStStP)和Et. pycnantha (EStP), 醇溶蛋白电泳图谱对其系统地位分类不明确。而在谷蛋白图谱(图2B )中, Et. repens 、Et. pun- gens 和Et. pycnantha 聚类在一起, 能较好地反映它们的系统关系, 由此推测编码谷蛋白的基因可能存在于St基因组上。同时, 对于未知染色体组成的物种Et. pachynera , 我们推测其为异源多倍体物种, 而不属于含E染色体组的Lophopyrum Á. Löve。因此, 醇溶蛋白和谷蛋白的相关研究结果能在一定程度上反映偃麦草属植物的系统分类关系, 可用于该属植物种间遗传差异及亲缘关系方面的研究, 但要尽量综合考虑形态学、细胞学和分子生物学等方面的资料才能更全面地揭示其真实的系统关系。Dewey, 1984 ; Li et al., 2008 ; Li and Wang, 2009 ; Mao et al., 2010 )。本研究表明, 偃麦草属植物的蛋白变异类型较为丰富, 可能存在大量能用于麦类作物蛋白质品质改良的基因资源。孙善澄(1981) 利用中间偃麦草(Elytrigia re- pens (L.) Nevski)与普通小麦杂交, 育成了小偃1号、龙麦1号和新曙光6号等小麦新品种。通过远缘杂交的常规方法就能将Et. elongata 、Et. pontica 、Et. intermedia 、Et. scythica 、Et. pycnantha 和Et. pungens 等偃麦草属植物丰富的醇溶蛋白基因转移到小麦中, 以期改良栽培小麦的蛋白质品质。Dvořák等(1986) 利用长穗偃麦草1E染色体分别代换普通小麦的1A、1B和1D染色体, 得到了一套代换系材料。朱振东和贾继增(2003) 及王际睿等(2004) 对长穗偃麦草中HMW-GS基因进行了分离和鉴定。李玉京等(1998) 对源于长穗偃麦草的高分子量谷蛋白特异表达启动子区域序列进行了研究。这些研究促使更多的偃麦草属优良基因可以转移到麦类作物中。本研究中, 从偃麦草属植物醇溶蛋白亚基的组成来看, Et. geniculata 、Et. kosaninii 、Et. libanotica 、Et. stipifolia 、Et. strigosa 和Et. tauri 的醇溶蛋白亚基主要是分布于ω区的高分子量亚基和β区的低分子量亚基; Et. bessarabica 、Et. caespitosa 、Et. caespitosa ssp. nodosa 、Et. elongata 、Et. farcta 、Et. geniculata ssp. pruinifera 、Et. intermedia 、Et. podpera 、Et. pontica 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 和Et. varnensis 的蛋白带纹主要在ω、β和α区, γ区几乎没有; Et. intermedia ssp. intermedia 的醇溶蛋白亚基主要是分布于α和β区的低分子量亚基和γ区的高分子量亚基, 在ω区的高分子量亚基分布相对较少; Et. pachynera 的醇溶蛋白亚基主要是分布在α区的低分子量亚基。因此, 在利用偃麦草属植物醇溶蛋白亚基进行基因克隆时, 应根据不同的蛋白质功能和需求选择相应的材料, 才会使麦类作物的品质改良更加行之有效。The authors have declared that no competing interests exist. 参考文献 文献选项 原文顺序 文献年度倒序 文中引用次数倒序 被引期刊影响因子 [1] 陈默君 , 贾慎修 (2002 ). 中国饲用植物 . 北京: 中国农业出版社. pp. 136 -140 .[2] 董超华 , 徐如宏 , 张庆勤 (2003 ). 小麦醇溶蛋白和谷蛋白研究进展. 山地农业生物学报 22, 164 -168 .[3] 高梅 , 张国权 , 魏益民 , 张继澍 , 党娇 , 王秋娟 (2007 ). 小麦醇溶蛋白A-PAGE电泳鉴定技术的分析. 西北农林科技大学学报(自然科学版) 35(9 ), 53 -57 .[4] 兰秀锦 , 魏育明 , 王志容 , 郑有良 (1999 ). 中国节节麦与中东节节麦的醇溶蛋白遗传多样性比较研究. 四川农业大学学报 17, 245 -248 .[5] 李守明 , 梁维 , 魏凌基 , 齐军仓 , 王金玲 (2011 ). 利用两种电泳技术分析大麦品种的醇溶蛋白差异及亲缘关系. 石河子大学学报(自然科学版) 29, 15 -19 .[6] 李玉京 , 李滨 , 刘建中 , 李继云 , 李振声 , 姚树江 (1998 ). 低磷营养胁迫对小麦-长穗偃麦草附加系酸性磷酸酶同工酶的影响. 中国农业科学 31(4 ), 26 -31 .[7] 吕伟东 , 徐鹏彬 , 蒲训 (2007 ). 偃麦草属种质资源在普通小麦育种中的应用现状简介. 草业学报 16, 136 -140 .[8] 马渐新 , 周荣华 , 董玉琛 , 贾继增 (1999 ). 来自长穗偃麦草的抗小麦条锈病基因的定位. 科学通报 44, 65 -69 .[9] 孙善澄 (1981 ). 小偃麦新品种与中间类型的选育途径、程序和方法. 作物学报 7, 51 -58 .[10] 唐慧慧 , 丁毅 , 胡耀军 (2002 ). 中国近缘野生大麦醇溶蛋白的遗传多态性研究. 武汉植物学研究 20, 251 -257 .[11] 唐朝晖 , 刘守斌 , 刘少翔 , 孙玉 , 张兰萍 , 逯成芳 , 孙善澄 , 刘广田 (2004 ). 二倍体长穗偃麦草高分子量谷蛋白亚基多态性研究. 华北农学报 19, 34 -36 .[12] 王红 , 王瑞 , 陈新宏 , 赵继新 , 钟刚 (2008 ). 国内外普通小麦醇溶蛋白的遗传差异. 西北农业学报 17(6 ), 67 -72 .[13] 王洪刚 , 刘树兵 , 亓增军 , 孔凡晶 , 高居荣 (2000 ). 中间偃麦草在小麦遗传改良中的应用研究. 山东农业大学学报(自然科学版) 31, 333 -336 .[14] 王际睿 , 颜泽洪 , 魏育明 , 郑有良 (2004 ). 长穗偃麦草y型高分子量谷蛋白基因的鉴定与分子克隆. 农业生物技术学报 12, 143 -146 .[15] 吴卫东 , 陈梦竹 , 陈凡国 , 夏光敏 (2010 ). 长穗偃麦草与老芒麦中α-醇溶蛋白基因的分离与鉴定. 分子植物育种 8, 20 -28 .[16] 杨瑞武 , 魏秀华 , 周永红 , 郑有良 (2004 ). 赖草属植物醇溶蛋白的遗传多态性. 云南植物研究 26, 103 -110 .[17] 杨瑞武 , 周永红 , 郑有良 (2000 ). 披碱草属的醇溶蛋白研究. 四川农业大学学报 18, 11 -14 .[18] 杨瑞武 , 周永红 , 郑有良 , 胡超 (2001 ). 小麦族四个属模式种的醇溶蛋白分析. 广西植物 21, 239 -242 .[19] 于海清 (2007 ). 拟鹅观草属四倍体物种的分子细胞遗传学研究. 博士论文. 雅安: 四川农业大学 . pp. 12 -42 .[20] 翟旭光 , 潘志芬 , 商闯 , 邓光兵 , 余懋群 (2009 ). 燕麦麦谷蛋白SDS-PAGE电泳分析. 西南农业学报 22, 24 -28 .[21] 张学勇 , 杨欣明 , 董玉琛 (1995 ). 醇溶蛋白电泳在小麦种质资源遗传分析中的应用. 中国农业科学 28(4 ), 25 -32 .[22] 张颖 , 张利 , 张海琴 , 丁春邦 , 周永红 (2006 ). 鹅观草属、披碱草属、猬草属和仲彬草属物种的醇溶蛋白分析. 四川农业大学学报 24, 256 -262 .[23] 郑青焕 , 李晓萍 , 郭超 , 昝凯 , 陈真真 , 白宇浩 , 刘洋 , 赵继新 , 武军 , 王中华 , 陈新宏 (2016 ). 21份印度小麦高分子谷蛋白亚基、醇溶蛋白及品质分析. 麦类作物学报 36, 62 -68 .[24] 朱振东 , 贾继增 (2003 ). 小麦SSR标记的发展及应用. 遗传 25, 355 -360 .[25] Asfaw Z 2008 ). Variation in hordein polypeptide pattern within Ethiopian barley, Hordeum vulgare L. (Poaceae).Hereditas 110, 185 -191 .[26] Assadi M Runemark H 1995 ). Hybridisation, genomic constitution and generic delimitation in Elymus s.l. (Poa- ceae: Triticeae).Plant Syst Evol 194, 189 -205 .[27] Desvaux (1810 ). Eustachys. Paris: Nouv Bull Sci Soc Phi- lom . pp. 190 .[28] Dewey DR 1980 ). Morphological, cytological and taxonomic relationships between Agropyron repens and Agropyron elongatiforme (Gramineae).Syst Bot 51, 61 -70 .[29] Dewey DR 1984 ). The genomic system of classification as a guide to intergeneric hybridization with the perennial Triticeae. In: Gustafson JP, ed. Gene Manipulation in Plant Improvement. New York: Plenum Press. pp. 209 -279 .[30] Draper SR 1987 ). ISTA variety committee report of the working group for biochemical tests for cultivar identification 1983-1986.Seed Sci Technol 15, 431 -434 .[31] Dvořák J Kasarda DD Dietler MD Lew EJL Anderson OD Litts JC Shewry PR 1986 ). Chromosomal location of seed storage protein genes in the genome of Elytrigia elongata.Can J Genet Cytol 28, 818 -830 .[32] Jaccard P 1908 ). Nouvelles researches sur la distribution florale.Bull Soc Vaud Sci Nat 44, 223 -270 .[33] Jiang GM Dong M 2000 ). A comparative study on photosynthesis and water use efficiency between clonal and non-clonal plant species along the Northeast China Transect (NECT).Acta Bot Sin 42, 855 -863 .[34] Kawaura K Mochida K Ogihara Y 2005 ). Expression profile of two storage-protein gene families in hexaploid wheat revealed by large-scale analysis of expressed sequence tags.Plant Physiol 139, 1870 -1880 .[35] Kim TW Kim JC Fedak G Son JH Park KC Kim NS 2010 ). Sequence variation in ITS spacers and 5.8S rDNA and relationship of E, St, P, Ns, Xm, and H genomes in the genera of Agropyron, Elytrigia, Leymus, Pascopyrum, Psathyrostachys, and Hordeum. Genes Genomics 32, 477 -485 .[36] Lawrence GJ Shepherd KW 1981 ). Chromosomal location of genes controlling seed proteins in species related to wheat.Theor Appl Genet 59, 25 -31 .[37] Li HJ Wang XM 2009 ). Thinopyrum ponticum and Th. intermedium: the promising source of resistance to fungal and viral diseases of wheat.J Genet Genomics 36, 557 -565 .[38] Li ZS Li B Tong YP 2008 ). The contribution of distant hybridization with decaploid Agropyron elongatum to wheat improvement in China.J Genet Genomics 35, 451 -456 .[39] Löve Á 1984 ). Conspectus of the Triticeae.Feddes Rep 95, 425 -521 .[40] Mao PS Huang Y Wang XG Meng L Mao PC Zhang GF 2010 ). Cytological evaluation and karyotype analysis in plant germplasms of Elytrigia Desv.Agric Sci China 9, 1553 -1560 .[41] Mason-Gamer RJ Orme NL Anderson CM 2002 ). Phylogenetic analysis of North American Elymus and the mono- genomic Triticeae (Poaceae) using three chloroplast DNA data sets.Genome 45, 991 -1002 .[42] Metakovsky EV Akhmedov MG Sozinov AA 1986 ). Genetic analysis of gliadin-encoding genes reveals gene clusters as well as single remote genes.Theor Appl Ge- net 73, 278 -285 .[43] Nakamura H 2000 ). Allelic variation at high-molecular- weight glutenin subunit loci, Glu-A1, Glu-B1 and Glu-D1 in Japanese and Chinese hexaploid wheats.Euphytica 112, 187 -193 .[44] Seberg O Frederiksen S 2008 ). A phylogenetic analysis of the monogenomic Triticeae (Poaceae) based on morpho- logy.Bot J Linn Soc 136, 75 -97 .[45] Stebbins Jr GL Pun FT 1953 ). Artificial and natural hybrids in the Gramineae, tribe Hordeae. V. Diploid hybrids of Agropyron.Am J Bot 40, 444 -449 .[46] Tzvelev NN 1976 ). Triticeae Dum Poaceae URSS. Leningrad: Nauka Publishers Leningrad Section. pp. 181 -203 .
Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...张学勇等, 1995 ; 董超华等, 2003 ).本研究分析了取自偃麦草属24个种24份材料的蛋白电泳图谱, 结果表明, 醇溶蛋白和谷蛋白电泳图谱在物种间差异明显, 表明它们能够作为偃麦草属植物鉴定的指纹图谱, 用于区分和鉴定偃麦草属物种. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...Dvořák等(1986) 利用长穗偃麦草1E染色体分别代换普通小麦的1A、1B和1D染色体, 得到了一套代换系材料.朱振东和贾继增(2003) 及王际睿等(2004) 对长穗偃麦草中HMW-GS基因进行了分离和鉴定.李玉京等(1998) 对源于长穗偃麦草的高分子量谷蛋白特异表达启动子区域序列进行了研究.这些研究促使更多的偃麦草属优良基因可以转移到麦类作物中.本研究中, 从偃麦草属植物醇溶蛋白亚基的组成来看, Et. geniculata 、Et. kosaninii 、Et. libanotica 、Et. stipifolia 、Et. strigosa 和Et. tauri 的醇溶蛋白亚基主要是分布于ω区的高分子量亚基和β区的低分子量亚基; Et. bessarabica 、Et. caespitosa 、Et. caespitosa ssp. nodosa 、Et. elongata 、Et. farcta 、Et. geniculata ssp. pruinifera 、Et. intermedia 、Et. podpera 、Et. pontica 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 和Et. varnensis 的蛋白带纹主要在ω、β和α区, γ区几乎没有; Et. intermedia ssp. intermedia 的醇溶蛋白亚基主要是分布于α和β区的低分子量亚基和γ区的高分子量亚基, 在ω区的高分子量亚基分布相对较少; Et. pachynera 的醇溶蛋白亚基主要是分布在α区的低分子量亚基.因此, 在利用偃麦草属植物醇溶蛋白亚基进行基因克隆时, 应根据不同的蛋白质功能和需求选择相应的材料, 才会使麦类作物的品质改良更加行之有效. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...孙善澄(1981) 利用中间偃麦草(Elytrigia re- pens (L.) Nevski)与普通小麦杂交, 育成了小偃1号、龙麦1号和新曙光6号等小麦新品种.通过远缘杂交的常规方法就能将Et. elongata 、Et. pontica 、Et. intermedia 、Et. scythica 、Et. pycnantha 和Et. pungens 等偃麦草属植物丰富的醇溶蛋白基因转移到小麦中, 以期改良栽培小麦的蛋白质品质. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...g 离心5分钟, 取上清液, 置于4˚C冰箱保存备用.电泳方法采用酸性聚丙烯酰胺凝胶电泳(A-PAGE), 电泳仪器为Bio-Rad公司的PROTEAN® II Xiell型电泳槽, 上样量为5 µL, 稳压电泳.200 V定压电泳30分钟后, 400 V定压继续电泳2.5小时.室温保持在25-28˚C, 通过冷水循环保持电泳体系内部温度为4˚C.根据Kawaura等(2005) 的分区方法对醇溶蛋白电泳谱带进行分区. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Dvořák等(1986) 利用长穗偃麦草1E染色体分别代换普通小麦的1A、1B和1D染色体, 得到了一套代换系材料.朱振东和贾继增(2003) 及王际睿等(2004) 对长穗偃麦草中HMW-GS基因进行了分离和鉴定.李玉京等(1998) 对源于长穗偃麦草的高分子量谷蛋白特异表达启动子区域序列进行了研究.这些研究促使更多的偃麦草属优良基因可以转移到麦类作物中.本研究中, 从偃麦草属植物醇溶蛋白亚基的组成来看, Et. geniculata 、Et. kosaninii 、Et. libanotica 、Et. stipifolia 、Et. strigosa 和Et. tauri 的醇溶蛋白亚基主要是分布于ω区的高分子量亚基和β区的低分子量亚基; Et. bessarabica 、Et. caespitosa 、Et. caespitosa ssp. nodosa 、Et. elongata 、Et. farcta 、Et. geniculata ssp. pruinifera 、Et. intermedia 、Et. podpera 、Et. pontica 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 和Et. varnensis 的蛋白带纹主要在ω、β和α区, γ区几乎没有; Et. intermedia ssp. intermedia 的醇溶蛋白亚基主要是分布于α和β区的低分子量亚基和γ区的高分子量亚基, 在ω区的高分子量亚基分布相对较少; Et. pachynera 的醇溶蛋白亚基主要是分布在α区的低分子量亚基.因此, 在利用偃麦草属植物醇溶蛋白亚基进行基因克隆时, 应根据不同的蛋白质功能和需求选择相应的材料, 才会使麦类作物的品质改良更加行之有效. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...杨瑞武等, 2004 ), 结果(图2A )显示, 醇溶蛋白遗传相似性系数为0.66时, 所有材料被聚为6大类.第I类包括Et. bessarabica 、Et. caespitosa ssp. nodosa 、Et. farcta 、Et. geniculata 、Et. libano- tica 、Et. podpera 、Et. rechingeri 、Et. scirpea 和Et. strigosa ; 第II类包括Et. caespitosa 、Et. elongata 、Et. geniculata ssp. pruinifera 、Et. intermedia ssp. intermedia 、Et. kosaninii 、Et. pachynera 、Et. repens 、Et. stipifolia 和Et. tauri ; 第III类包括Et. intermedia 、Et. scythica 和Et. varnensis ; Et. pungens 单独聚为第IV类; Et. pontica 单独聚为第V类; Et. pycnantha 单独聚为第VI类.在第I类中, Et. geniculate 、Et. libano- tica 、Et. rechingeri 和Et. strigosa 先聚在一起, 再与Et. bessarabica 、Et. casepitosa ssp. nodosa 、Et. ...杨瑞武等, 2004 ), 结果(图2B )显示, 谷蛋白遗传相似性系数为0.62时, 所有材料被聚为4大类: 第I类包括Et. bessarabica 、Et. caespitosa ssp. nodosa 、Et. cae- spitosa 、Et. elongata 、Et. geniculata ssp. pruinifera 、Et. intermedia ssp. intermedia 、Et. kosaninii 、Et. libanotica 、Et. pachynera 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 、Et. strigosa 和Et. tauri ; 第II类包括Et. farcta 、Et. geniculata 、Et. stipifolia 和Et. varnensis ; Et. intermedia 单独聚为第III类; 第IV类包括Et. podpera 和Et. pontica .在第I类中, Et. elongata 、Et. kos- aninii 、Et. libanotica 、Et. pachynera 、Et. pungens 、Et. pycnantha 、Et. repens 、Et. scythica 和Et. tauri 先聚在一起, 然后与Et. bessarabica 、Et. caespitosa ssp. nodosa 、Et. scirpea 和Et. strigosa 聚类, 再与Et. geniculata ssp. pruinifera 及Et. rechingeri 聚类, 最后与Et. caespitosa 及Et. intermedia ssp. intermedia 聚在一起. ...杨瑞武等(2000) 通过研究披碱草属植物的醇溶蛋白, 发现小麦族植物的醇溶蛋白带纹多少与染色体数目有关.本研究(表2 )表明, 偃麦草属含有相同染色体组的物种中, 染色体数目越多, 其醇溶蛋白和谷蛋白带纹也越多; 而含有不同染色体组的物种之间, 其染色体数目与醇溶蛋白和谷蛋白带纹数目无关.因此, 我们认为仅按照形态学标准划分传统偃麦草属的范围是不合理的. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...图2 )来看, 含有相同基因组的物种基本先聚在一起, 然后与亲缘关系较近的其它基因组的物种聚类.就醇溶蛋白聚类结果(图2 A)而言, 第I类中, 含有St基因组的物种(Et. libanotica 、Et. geniculata 和Et. strigosa )与含E基因组的物种(Et. bessarabica 、Et. farcta 、Et. podpera 、Et. rechingeri 和Et. scirpea )聚在一起, 说明在偃麦草属中含有E和St基因组物种的醇溶蛋白的遗传相似度很高, 也表明这些物种之间亲缘关系较近, 这与前人的细胞生物学(于海清, 2007 )和分子系统学(Mason-Gamer et al., 2002 ; Seberg and Frederiksen, 2008 ; Kim et al., 2010 )研究结果一致.第II类中, St基因组的物种有Et. tauri 、Et. kosaninii 和Et. stipifolia , 说明具有St基因组的物种分别包含在第I和II类中, 表明含有St基因组 的不同物种间存在一定程度的分化(Stebbins and Pun, 1953 ), 从而导致醇溶蛋白带纹的差异性较大.我们发现含ESt基因组的物种Et. caespitosa ssp. nodosa 在醇溶蛋白图谱中与含E基因组的物种Et. podpera 聚类在一起, 相似性系数为0.67 (表3 ); 在谷蛋白电泳图谱中, 与含E基因组的物种Et. scirpea 聚在一起, 相似性系数为0.77 (表4 ), 表明Et. caespitosa ssp. nodosa 与含有E基因组的物种的亲缘关系较密切. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...张学勇等, 1995 ; 董超华等, 2003 ).本研究分析了取自偃麦草属24个种24份材料的蛋白电泳图谱, 结果表明, 醇溶蛋白和谷蛋白电泳图谱在物种间差异明显, 表明它们能够作为偃麦草属植物鉴定的指纹图谱, 用于区分和鉴定偃麦草属物种. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...Aegilops tauschii ) (张学勇等, 1995 ; 兰秀锦等, 1999 )和大麦(Hordeum vulgare ) (唐慧慧等, 2002 ; 李守明等, 2011 )进行了起源与进化研究.杨瑞武等(2001) 运用醇溶蛋白图谱差异探讨了小麦族4个属模式种的亲缘关系, 表明其能够反映一定的系统关系.张颖等(2006) 对鹅观草属(Roegneria )、披碱草属、猬草属(Hystrix )和仲彬草属(Kengyilia )物种的醇溶蛋白进行了分析, 并揭示了一定的物种系统地位.王红等(2008) 利用A-PAGE技术分析了小麦种子醇溶蛋白图谱的差异, 认为蛋白电泳图谱能够作为种间亲缘关系远近的一项指标, 并为品种改良提供了依据.翟旭光等(2009) 采用SDS-PAGE电泳方法对71份不同染色体组型的燕麦(Avena sativa )材料的麦谷蛋白进行了电泳分析, 发现燕麦麦谷蛋白SDS-PAGE电泳图谱可以作为燕麦指纹图谱. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Dvořák等(1986) 利用长穗偃麦草1E染色体分别代换普通小麦的1A、1B和1D染色体, 得到了一套代换系材料.朱振东和贾继增(2003) 及王际睿等(2004) 对长穗偃麦草中HMW-GS基因进行了分离和鉴定.李玉京等(1998) 对源于长穗偃麦草的高分子量谷蛋白特异表达启动子区域序列进行了研究.这些研究促使更多的偃麦草属优良基因可以转移到麦类作物中.本研究中, 从偃麦草属植物醇溶蛋白亚基的组成来看, Et. geniculata 、Et. kosaninii 、Et. libanotica 、Et. stipifolia 、Et. strigosa 和Et. tauri 的醇溶蛋白亚基主要是分布于ω区的高分子量亚基和β区的低分子量亚基; Et. bessarabica 、Et. caespitosa 、Et. caespitosa ssp. nodosa 、Et. elongata 、Et. farcta 、Et. geniculata ssp. pruinifera 、Et. intermedia 、Et. podpera 、Et. pontica 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 和Et. varnensis 的蛋白带纹主要在ω、β和α区, γ区几乎没有; Et. intermedia ssp. intermedia 的醇溶蛋白亚基主要是分布于α和β区的低分子量亚基和γ区的高分子量亚基, 在ω区的高分子量亚基分布相对较少; Et. pachynera 的醇溶蛋白亚基主要是分布在α区的低分子量亚基.因此, 在利用偃麦草属植物醇溶蛋白亚基进行基因克隆时, 应根据不同的蛋白质功能和需求选择相应的材料, 才会使麦类作物的品质改良更加行之有效. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Dewey, 1984 ; Li et al., 2008 ; Li and Wang, 2009 ; Mao et al., 2010 ).本研究表明, 偃麦草属植物的蛋白变异类型较为丰富, 可能存在大量能用于麦类作物蛋白质品质改良的基因资源. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Dvořák等(1986) 利用长穗偃麦草1E染色体分别代换普通小麦的1A、1B和1D染色体, 得到了一套代换系材料.朱振东和贾继增(2003) 及王际睿等(2004) 对长穗偃麦草中HMW-GS基因进行了分离和鉴定.李玉京等(1998) 对源于长穗偃麦草的高分子量谷蛋白特异表达启动子区域序列进行了研究.这些研究促使更多的偃麦草属优良基因可以转移到麦类作物中.本研究中, 从偃麦草属植物醇溶蛋白亚基的组成来看, Et. geniculata 、Et. kosaninii 、Et. libanotica 、Et. stipifolia 、Et. strigosa 和Et. tauri 的醇溶蛋白亚基主要是分布于ω区的高分子量亚基和β区的低分子量亚基; Et. bessarabica 、Et. caespitosa 、Et. caespitosa ssp. nodosa 、Et. elongata 、Et. farcta 、Et. geniculata ssp. pruinifera 、Et. intermedia 、Et. podpera 、Et. pontica 、Et. pungens 、Et. pycnantha 、Et. rechingeri 、Et. repens 、Et. scirpea 、Et. scythica 和Et. varnensis 的蛋白带纹主要在ω、β和α区, γ区几乎没有; Et. intermedia ssp. intermedia 的醇溶蛋白亚基主要是分布于α和β区的低分子量亚基和γ区的高分子量亚基, 在ω区的高分子量亚基分布相对较少; Et. pachynera 的醇溶蛋白亚基主要是分布在α区的低分子量亚基.因此, 在利用偃麦草属植物醇溶蛋白亚基进行基因克隆时, 应根据不同的蛋白质功能和需求选择相应的材料, 才会使麦类作物的品质改良更加行之有效. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...图2 )来看, 含有相同基因组的物种基本先聚在一起, 然后与亲缘关系较近的其它基因组的物种聚类.就醇溶蛋白聚类结果(图2 A)而言, 第I类中, 含有St基因组的物种(Et. libanotica 、Et. geniculata 和Et. strigosa )与含E基因组的物种(Et. bessarabica 、Et. farcta 、Et. podpera 、Et. rechingeri 和Et. scirpea )聚在一起, 说明在偃麦草属中含有E和St基因组物种的醇溶蛋白的遗传相似度很高, 也表明这些物种之间亲缘关系较近, 这与前人的细胞生物学(于海清, 2007 )和分子系统学(Mason-Gamer et al., 2002 ; Seberg and Frederiksen, 2008 ; Kim et al., 2010 )研究结果一致.第II类中, St基因组的物种有Et. tauri 、Et. kosaninii 和Et. stipifolia , 说明具有St基因组的物种分别包含在第I和II类中, 表明含有St基因组 的不同物种间存在一定程度的分化(Stebbins and Pun, 1953 ), 从而导致醇溶蛋白带纹的差异性较大.我们发现含ESt基因组的物种Et. caespitosa ssp. nodosa 在醇溶蛋白图谱中与含E基因组的物种Et. podpera 聚类在一起, 相似性系数为0.67 (表3 ); 在谷蛋白电泳图谱中, 与含E基因组的物种Et. scirpea 聚在一起, 相似性系数为0.77 (表4 ), 表明Et. caespitosa ssp. nodosa 与含有E基因组的物种的亲缘关系较密切. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Dewey, 1984 ; Li et al., 2008 ; Li and Wang, 2009 ; Mao et al., 2010 ).本研究表明, 偃麦草属植物的蛋白变异类型较为丰富, 可能存在大量能用于麦类作物蛋白质品质改良的基因资源. ...Dewey, 1984 ; Li et al., 2008 ; Li and Wang, 2009 ; Mao et al., 2010 ).本研究表明, 偃麦草属植物的蛋白变异类型较为丰富, 可能存在大量能用于麦类作物蛋白质品质改良的基因资源. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ...Dewey, 1984 ; Li et al., 2008 ; Li and Wang, 2009 ; Mao et al., 2010 ).本研究表明, 偃麦草属植物的蛋白变异类型较为丰富, 可能存在大量能用于麦类作物蛋白质品质改良的基因资源. ...图2 )来看, 含有相同基因组的物种基本先聚在一起, 然后与亲缘关系较近的其它基因组的物种聚类.就醇溶蛋白聚类结果(图2 A)而言, 第I类中, 含有St基因组的物种(Et. libanotica 、Et. geniculata 和Et. strigosa )与含E基因组的物种(Et. bessarabica 、Et. farcta 、Et. podpera 、Et. rechingeri 和Et. scirpea )聚在一起, 说明在偃麦草属中含有E和St基因组物种的醇溶蛋白的遗传相似度很高, 也表明这些物种之间亲缘关系较近, 这与前人的细胞生物学(于海清, 2007 )和分子系统学(Mason-Gamer et al., 2002 ; Seberg and Frederiksen, 2008 ; Kim et al., 2010 )研究结果一致.第II类中, St基因组的物种有Et. tauri 、Et. kosaninii 和Et. stipifolia , 说明具有St基因组的物种分别包含在第I和II类中, 表明含有St基因组 的不同物种间存在一定程度的分化(Stebbins and Pun, 1953 ), 从而导致醇溶蛋白带纹的差异性较大.我们发现含ESt基因组的物种Et. caespitosa ssp. nodosa 在醇溶蛋白图谱中与含E基因组的物种Et. podpera 聚类在一起, 相似性系数为0.67 (表3 ); 在谷蛋白电泳图谱中, 与含E基因组的物种Et. scirpea 聚在一起, 相似性系数为0.77 (表4 ), 表明Et. caespitosa ssp. nodosa 与含有E基因组的物种的亲缘关系较密切. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...Draper, 1987 ).醇溶蛋白是单亚基结构, 在酸性聚丙烯酰胺凝胶电泳(acid polyacrylamide gel electrophoresis, A-PAGE)中, 根据分子量大小和迁移率的不同, 其电泳图谱可划分为α、β、γ和ω 4个区(Kawaura et al., 2005 ).普通小麦及其近缘属物种中含有丰富的醇溶蛋白等位变异(Metakovsky et al., 1986 ).谷蛋白在十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS polyacry- lamide gel electrophoresis, SDS-PAGE)图谱上, 被分为A、B、C三个区域, 即A区为高分子量谷蛋白亚基(HMW-GS), B和C区是低分子量谷蛋白亚基(LMW- GS).高分子量谷蛋白是小麦及其近缘物种的重要种子贮藏蛋白, 与种子的生理功能紧密相关, 能在种子发芽过程中提供能量(Jiang and Dong, 2000 ).目前, 国内外****运用醇溶蛋白和谷蛋白电泳图谱对小麦族植物进行的研究主要集中在以下两个方面: 麦类作 物及其近缘属物种的多态性和品质分析(Nakamura, 2000 ; 高梅等, 2007 ; Asfaw, 2008 ; 郑青焕等, 2016 ); 长穗偃麦草(Elytrigia elongata (Host) Nev- ski.)的分子遗传学(王际睿等, 2004 ; 吴卫东等, 2010 )和细胞学(Lawrence and Shepherd, 1981 ; 唐朝晖等, 2004 )特性.关于偃麦草属物种的醇溶蛋白和谷蛋白的多态性与系统学意义的研究还未见报道.本研究分别利用A-PAGE和SDS-PAGE技术, 分析了24种偃麦草属植物的醇溶蛋白和谷蛋白电泳图谱, 旨在构建该属植物醇溶蛋白和谷蛋白的“指纹图谱”, 探讨其种间醇溶蛋白和谷蛋白多态性和指纹图谱的系统学意义, 以期为揭示其系统关系和麦类作物遗传改良提供理论依据. ...图2 )来看, 含有相同基因组的物种基本先聚在一起, 然后与亲缘关系较近的其它基因组的物种聚类.就醇溶蛋白聚类结果(图2 A)而言, 第I类中, 含有St基因组的物种(Et. libanotica 、Et. geniculata 和Et. strigosa )与含E基因组的物种(Et. bessarabica 、Et. farcta 、Et. podpera 、Et. rechingeri 和Et. scirpea )聚在一起, 说明在偃麦草属中含有E和St基因组物种的醇溶蛋白的遗传相似度很高, 也表明这些物种之间亲缘关系较近, 这与前人的细胞生物学(于海清, 2007 )和分子系统学(Mason-Gamer et al., 2002 ; Seberg and Frederiksen, 2008 ; Kim et al., 2010 )研究结果一致.第II类中, St基因组的物种有Et. tauri 、Et. kosaninii 和Et. stipifolia , 说明具有St基因组的物种分别包含在第I和II类中, 表明含有St基因组 的不同物种间存在一定程度的分化(Stebbins and Pun, 1953 ), 从而导致醇溶蛋白带纹的差异性较大.我们发现含ESt基因组的物种Et. caespitosa ssp. nodosa 在醇溶蛋白图谱中与含E基因组的物种Et. podpera 聚类在一起, 相似性系数为0.67 (表3 ); 在谷蛋白电泳图谱中, 与含E基因组的物种Et. scirpea 聚在一起, 相似性系数为0.77 (表4 ), 表明Et. caespitosa ssp. nodosa 与含有E基因组的物种的亲缘关系较密切. ...图2 )来看, 含有相同基因组的物种基本先聚在一起, 然后与亲缘关系较近的其它基因组的物种聚类.就醇溶蛋白聚类结果(图2 A)而言, 第I类中, 含有St基因组的物种(Et. libanotica 、Et. geniculata 和Et. strigosa )与含E基因组的物种(Et. bessarabica 、Et. farcta 、Et. podpera 、Et. rechingeri 和Et. scirpea )聚在一起, 说明在偃麦草属中含有E和St基因组物种的醇溶蛋白的遗传相似度很高, 也表明这些物种之间亲缘关系较近, 这与前人的细胞生物学(于海清, 2007 )和分子系统学(Mason-Gamer et al., 2002 ; Seberg and Frederiksen, 2008 ; Kim et al., 2010 )研究结果一致.第II类中, St基因组的物种有Et. tauri 、Et. kosaninii 和Et. stipifolia , 说明具有St基因组的物种分别包含在第I和II类中, 表明含有St基因组 的不同物种间存在一定程度的分化(Stebbins and Pun, 1953 ), 从而导致醇溶蛋白带纹的差异性较大.我们发现含ESt基因组的物种Et. caespitosa ssp. nodosa 在醇溶蛋白图谱中与含E基因组的物种Et. podpera 聚类在一起, 相似性系数为0.67 (表3 ); 在谷蛋白电泳图谱中, 与含E基因组的物种Et. scirpea 聚在一起, 相似性系数为0.77 (表4 ), 表明Et. caespitosa ssp. nodosa 与含有E基因组的物种的亲缘关系较密切. ...Elytrigia )是禾本科小麦族的一个多倍体多年生属, 全球约有40个种, 主要分布在南北半球的亚热带与暖温带地区(Tzvelev, 1976 ).偃麦草属植物是小麦(Triticum aestivum )的野生近缘种, 具有小麦缺少的优良遗传特性.该属物种是良好的多年生草料, 且具有抗病、抗旱、抗寒、耐盐碱、大穗、多花和质优等优异性状, 生态适应性和繁殖能力强, 具有十分重要的应用价值(陈默君和贾慎修, 2002 ; 吕伟东等, 2007 ), 是小麦改良不可缺少的野生基因库(马渐新等, 1999 ; 王洪刚等, 2000 ; 吕伟东等, 2007 ).Desvaux (1810) 以Elytrigia repens (L.) Nevski (St- StH)为模式种建立偃麦草属, 其物种包括Ee 、Eb 、St、ESt和EStP等染色体组组成.根据Dewey (1984) 和Löve (1984) 提出的按照染色体组组成的分类法, 发现其模式种Et. repens 应划归披碱草属(Elymus ) (De- wey, 1980 ; Assadi and Runemark, 1995 ).由此可见, 偃麦草属的分类和系统地位仍然存在很大争议. ... 

{kind=link}
{kind=link}
{kind=link}
{kind=link}