Abstract: Rice is a staple food for more than 3 billion people in the world, and it is one of the most important food crops. As one of the types of cultivated rice, fragrant rice is favoured worldwide by consumers because of its agreeable scent. In recent years, with the rapid development of rice functional genomics and sequencing technology, great progress has been made in understanding the fragrant gene in rice, and a series of functional markers has been developed for screening the fragrant gene and breeding new rice varieties. This paper mainly reviews the progress in the genetic basis, gene function and regulation of the fragrant gene and the development and application of gene functional markers in rice. It provides useful references for the cultivation of new varieties of fragrant rice.
图1 水稻2-乙酰-1-吡咯啉(2-AP)可能的合成途径 Figure 1 Possible synthesis pathway of 2-acetyl-1-pyrroline (2-AP) in rice
通过对水稻香味基因的测序结果进行比对分析, 大多数研究者认为, 由于Badh2基因的第2或第7外显子片段存在缺失, 导致2-AP的大量积累(Kovach et al., 2009)而产生香味。但实际上, 有些水稻品种的2-AP含量较高, 却没有Badh2的等位基因, 这表明可能还存在其它能够引起香味的非Badh2的等位基因(Amarawathi et al., 2008)。多数研究者对香稻香味基因的研究仅注重香稻与非香稻外显子之间的差异, 但对真核生物而言, 除了基因的外显子, 内含子在基因的表达和调控中也发挥着重要作用(Fu et al., 1995; Jeon et al., 2000)。徐小龙等(2011)通过对水稻香味基因Badh2进行测序, 结果显示香稻的编码链并无变异发生, 既不属于第7外显子突变类型, 也不属于第2外显子突变类型, 而在Badh2基因的第2和4内含子区域的核苷酸序列却发生了突变。张江丽等(2015)通过鉴定323份水稻种质资源, 发现具香味的水稻品种共86份, 其中有80份属于第7外显子变异的突变类型, 进一步对另6份香稻种质的Badh2基因进行克隆并测序分析, 结果显示这6份香稻品种的Badh2基因序列完整。因此, 水稻的香味性状还可能受到其它基因或内含子的调控。
3 水稻香味基因功能标记的开发自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育。Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015)。Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点。同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点。针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析。随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008)。Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点。在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014)。根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015)。上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程。 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015)。针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015)。在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能。在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015)。其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I。因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段。 表1 Table 1 表1 表1 水稻香味基因Badh2功能标记的开发(He and Park, 2015) Table 1 Development of functional markers for Badh2 gene in rice (He and Park, 2015)
4 香味基因在水稻遗传育种中的应用目前, 主要的水稻育种技术仍然是常规杂交育种, 然而, 利用传统育种方法来培育香稻新品种费时费力 (Shan et al., 2015)。由于香味基因主要由隐性基因控制, 传统育种先要通过杂交和回交的方法, 将Badh2突变的基因导入现有的优良品种中, 然后在后代群体中进行筛选, 可是这一过程在苗期很难快速鉴定。然而, 我国水稻育种学家在优质、高产及多抗香稻新品种的培育方面还是做了大量研究, 并筛选到一系列香稻的恢复系、不育系以及新的杂交组合(黄庭旭等, 2006; 况浩池等, 2007; 刘光春等, 2008; 江青山等, 2008)。随着分子标记尤其是Badh2基因功能标记的开发, 分子标记辅助选择已经广泛应用于水稻遗传育种, 大大加快了香稻新品种的筛选与培育进程, 并且许多香稻品种已通过审定, 在生产中得到大面积的推广应用。 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003)。随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009)。此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012)。利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选。同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题。 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014)。在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现。于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015)。因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程。
5 展望香味是水稻重要的品质性状之一, 优质香米的消费需求将持续增加。因此, 香稻新品种选育以及香味基因研究受到越来越多水稻遗传育种学家和植物科学家的重视。随着水稻功能基因组研究的快速发展, 关于水稻中香味基因的遗传基础、功能及其调控规律等将会越来越明朗。最近十几年, 基因测序技术突飞猛进, 大量不同来源的水稻品种和香稻种质资源完成测序分析, 发现了一系列香味基因的变异位点及其等位基因(He and Park, 2015, He et al., 2015)。针对这些变异位点, 香味基因功能标记也被不断开发并已经广泛应用于香味基因筛选和杂交香稻骨干亲本培育。尽管香味基因功能标记已经用于香稻恢复系的选育(杜雪树等, 2009), 而且很多香稻品种已通过审定并在生产中得到较大面积的推广应用, 但是目前香稻遗传育种面临的主要问题依然是香型优良的恢复系极少, 减缓了香稻的遗传育种进程。 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种。例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011)。同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013)。高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011)。香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用。 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014)。此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略。利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015)。因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育。
The authors have declared that no competing interests exist.
徐小龙, 赵国超, 李建粤 (2011). 24种香稻品种甜菜碱醛脱氢酶2基因突变位点的分析及分子标记开发. 植物分类与资源学报 33, 667-673. DOI:10.3724/SP.J.1143.2011.11019URLFunctional molecular markers M7 and M2 have been developed based on the DNA sequence differences of badh2 between fragrant rice varieties and nonfragrant varieties in intron2, intron 4, exon7 and exon 2 respectively. PCR analyses on genome DNA of exon7mutated fragrant rice Wxiang 99075, exon2mutated fragrant rice Wuxiang14,nonfragrant rice 261S and the F1 plants by M7 and M2 showed that M7 and M2 could be absolutely used to the molecular markerassisted rice breeding experiments when exon7mutated and exon2mutated fragrant rice varieties are used as parents. The design of M7 primers took mutations both in exons and intrones into account. Moreover, taking 261S,Wxiang 99075 and Wuxiang14 as controls, the mutation sites of badh2 in 22 fragrant rice varieties were analyzed, it was showed that fragrant rice varieties could be classified into 3 types: exon 2mutated fragrant rice, exon 7mutated fragrant rice and nonexonmutated fragrant rice. At the same time, the mutation sites of badh2 in the main fragrant rice varieties which are grown in Shanghai and the surrounding areas have been verified. This research laid an important foundation for molecular markerassisted selection for novel fragrant rice.
闫影, 诸光明, 张丽霞, 万常照, 曹黎明, 赵志鹏, 吴书俊 (2015). 水稻香味基因分子标记的开发及应用. 西北植物学报 35, 269-274. DOI:10.7606/j.issn.1000-4025.2015.02.0269URLAccording to the mutation in exon 7 of 2 (Betaine aldehyde dehydrogenasegene) in fragrant rice,we designed a functional marker YY5-YY8,which can be used to check rice materials with this mutation or not.Other two reported markers for exon 2-mutated and exon 4 or 5-mutated types of 2,InDel-E2 and FMbadh2-E4-5,were also used to analyze the mutations of 2 in 80 fragrant rice materials from different ecological regions.It showed that 26 fragrant rice materials were of the exon 7-mutated type,37 fragrant rice materials were of exon 2-mutated type and none of them was found to be of exon 4 or 5-mutated type.This study constructed a new molecular marker YY5-YY8 for checking the mutation of 2 and identified the mutation types of these fragrant rice varieties for breeding high quality fragrant rice. [本文引用: 5]
AhnSN, BollichCN, TanksleySD (1992). RFLP tagging of a gene for aroma in rice.Theor Appl Genet 84, 825-828. DOI:10.1007/BF00227391PMID:24201481URLWe report here the identification of a DNA marker closely linked to a gene for aroma in rice. The DNA marker was identified by testing 126 mapped rice genomic, cDNA , and oat cDNA , clones as hybridization probes against Southern blots, consisting of DNA from a pair of nearly isogenic lines (NILs) with or without the aroma gene. Chromosomal segments introgressed from the donor genome were distinguished by RFLPs between the NILs. Linkage association of the clone with the gene was verified using an F 3 segregating for aroma. Cosegregation of the scented phenotype and donor-derived allele indicated the presence of linkage between the DNA marker and the gene. RFLP analysis showed that the gene is linked to a single-copy DNA clone, RG28 , on chromosome 8, at a distance of 4.5 cM. The availability of a linked DNA marker may facilitate early selection for the aroma gene in rice breeding programs.
[22]
AmarawathiY, SinghR, SinghAK, SinghVP, MohapatraT, SharmaTR, SinghNK (2008). Mapping of quantitative trait loci for basmati quality traits in rice ( Oryza sativa L.). Mol Breed 21, 49-65. DOI:10.1007/s11032-007-9108-8URLTraditional basmati rice varieties are very low yielding due to their poor harvest index, tendency to lodging and increasing susceptibility to foliar diseases; hence there is a need to develop new varieties combining the grain quality attributes of basmati with high yield potential to fill the demand gap. Genetic control of basmati grain and cooking quality traits is quite complex, but breeding work can be greatly facilitated by use of molecular markers tightly linked to these traits. A set of 209 recombinant inbred lines (RILs) developed from a cross between basmati quality variety Pusa 1121 and a contrasting quality breeding line Pusa 1342, were used to map the quantitative trait loci (QTLs) for seven important quality traits namely grain length (GL), grain breadth (GB), grain length to breadth ratio (LBR), cooked kernel elongation ratio (ELR), amylose content (AC), alkali spreading value (ASV) and aroma. A framework molecular linkage map was constructed using 110 polymorphic simple sequence repeat (SSR) markers distributed over the 12 rice chromosomes. A number of QTLs, including three for GL, two for GB, two for LBR, three for aroma and one each for ELR, AC and ASV were mapped on seven different chromosomes. While location of majority of these QTLs was consistent with the previous reports, one QTL for GL on chromosomes 1, and one QTL each for ELR and aroma on chromosomes 11 and 3, respectively, are being reported here for the first time. Contrary to the earlier reports of monogenic recessive inheritance, the aroma in Pusa 1121 is controlled by at least three genes located on chromosomes 3, 4 and 8, and similar to the reported association of badh2 gene with aroma QTL on chromosome 8, we identified location of badh1 gene in the aroma QTL interval on chromosome 4. A discontinuous 5 + 3 bp deletion in the seventh exon of badh2 gene, though present in all the RILs with high aroma, was not sufficient to impart this trait to the rice grains as many of the RILs possessing this deletion showed only mild or no aroma expression. [本文引用: 5]
[23]
ArikitS, YoshihashiT, WanchanaS, UyenTT, HuongNTT, WongpornchaiS, VanavichitA (2011). Deficiency in the amino aldehyde dehydrogenase encoded byGm- AMADH2, the homologue of rice Os2AP, enhances 2- acetyl-1-pyrroline biosynthesis in soybeans(Glycine max L.). Plant Biotechnol J 9, 75-87. [本文引用: 2]
[24]
BirlaDS, MalikK, SaingerM, ChaudharyD, JaiwalR, JaiwalPK (2017). Progress and challenges in improving the nutritional quality of rice ( Oryza sativa L.). Crit Rev Food Sci Nutr 57, 2455-2481. DOI:10.1080/10408398.2015.1084992PMID:26513164URLRice is a staple food for more than 3 billion people in more than 100 countries of the world but ironically it is deficient in many bioavailable vitamins, minerals, essential amino- and fatty-acids and phytochemicals that prevent chronic diseases like type 2 diabetes, heart disease, cancers, and obesity. To enhance the nutritional and other quality aspects of rice, a better understanding of the regulation of the processes involved in the synthesis, uptake, transport, and metabolism of macro-(starch, seed storage protein and lipid) and micronutrients (vitamins, minerals and phytochemicals) is required. With the publication of high quality genomic sequence of rice, significant progress has been made in identification, isolation, and characterization of novel genes and their regulation for the nutritional and quality enhancement of rice. During the last decade, numerous efforts have been made to refine the nutritional and other quality traits either by using the traditional breeding with high through put technologies such as marker assisted selection and breeding, or by adopting the transgenic approach. A significant improvement in vitamins (A, folate, and E), mineral (iron), essential amino acid (lysine), and flavonoids levels has been achieved in the edible part of rice, i.e., endosperm (biofortification) to meet the daily dietary allowance. However, studies on bioavailability and allergenicity on biofortified rice are still required. Despite the numerous efforts, the commercialization of biofortified rice has not yet been achieved. The present review summarizes the progress and challenges of genetic engineering and/or metabolic engineering technologies to improve rice grain quality, and presents the future prospects in developing nutrient dense rice to save the everincreasing population, that depends solely on rice as the staple food, from widespread nutritional deficiencies. [本文引用: 2]
[25]
BradburyLMT, FitzgeraldTL, HenryRJ, JinQS, WatersDLE (2005a). The gene for fragrance in rice.Plant Biotechnol J 3, 363-370. DOI:10.1111/j.1467-7652.2005.00131.xPMID:17129318URLThe flavour or fragrance of basmati and jasmine rice is associated with the presence of 2-acetyl-1-pyrroline. A recessive gene ( fgr ) on chromosome 8 of rice has been linked to this important trait. Here, we show that a gene with homology to the gene that encodes betaine aldehyde dehydrogenase (BAD) has significant polymorphisms in the coding region of fragrant genotypes relative to non-fragrant genotypes. The accumulation of 2-acetyl-1-pyrroline in fragrant rice genotypes may be explained by the presence of mutations resulting in a loss of function of the fgr gene product. The allele in fragrant genotypes has a mutation introducing a stop codon upstream of key amino acid sequences conserved in other BADs. The fgr gene corresponds to the gene encoding BAD2 in rice, while BAD1 is encoded by a gene on chromosome 4. BAD has been linked to stress tolerance in plants. However, the apparent loss of function of BAD2 does not seem to limit the growth of fragrant rice genotypes. Fragrance in domesticated rice has apparently originated from a common ancestor and may have evolved in a genetically isolated population, or may be the outcome of a separate domestication event. This is an example of effective human selection for a recessive trait during domestication. [本文引用: 8]
[26]
BradburyLMT, GilliesSA, BrushettDJ, WatersDLE, HenryRJ (2008). Inactivation of an aminoaldehyde dehydrogenase is responsible for fragrance in rice.Plant Mol Biol 68, 439-449. DOI:10.1007/s11103-008-9381-xPMID:18704694URLRice ( Oryza sativa ) has two betaine aldehyde dehydrogenase homologs, BAD1 and BAD2, encoded on chromosome four and chromosome eight respectively. BAD2 is responsible for the characteristic aroma of fragrant rice. Complementary DNA clones of both BAD1 and BAD2 were isolated and expressed in E. coli . BAD2 had optimum activity at pH 10, little to no affinity towards N-acetyl-γ-aminobutyraldehyde (NAGABald) with a Km of approximately 1002mM and moderate affinity towards γ-guanidinobutyraldehyde (GGBald) and betaine aldehyde (bet-ald) with Km values of approximately 26002μM and 6302μM respectively. A lower Km of approximately 902μM was observed with γ-aminobutyraldehyde (GABald), suggesting BAD2 has a higher affinity towards this substate in02vivo. The enzyme encoded on chromosome four, BAD1, had optimum activity at pH 9.5, showed little to no affinity towards bet-ald with a Km of 302mM and had moderate affinity towards GGBald, NAGABald and GABald with Km values of approximately 545, 420 and 49702μM respectively. BAD1 had a half life roughly double that of BAD2. We discuss the implications of these findings on the pathway of fragrance generation in Basmati and Jasmine rice and the potential of rice to accumulate the osmoprotectant glycine betaine. [本文引用: 1]
[27]
BradburyLMT, HenryRJ, JinQS, ReinkeRF, WatersDLE (2005b). A perfect marker for fragrance genotyping in rice.Mol Breed 16, 279-283. DOI:10.1007/s11032-005-0776-yURLAllele specific amplification (ASA) is a low-cost, robust technique that can be utilised to discriminate between alleles that differ by SNP's, insertions or deletions, within a single PCR tube. Fragrance in rice, a recessive trait, has been shown to be due to an eight02bp deletion and three SNP's in a gene on chromosome 8 which encodes a putative betaine aldehyde dehydrogenase 2 (BAD2). Here we report a single tube ASA assay which allows discrimination between fragrant and non-fragrant rice varieties and identifies homozygous fragrant, homozygous non-fragrant and heterozygous non-fragrant individuals in a population segregating for fragrance. External primers generate a fragment of approximately 58002bp as a positive control for each sample. Internal and corresponding external primers produce a 35502bp fragment from a non-fragrant allele and a 25702bp fragment from a fragrant allele, allowing simple analysis on agarose gels. [本文引用: 3]
[28]
ChenML, WeiXJ, ShaoGN, TangSQ, LuoJ, HuPS (2012). Fragrance of the rice grain achieved via artificial microRNA-induced down-regulation of OsBADH2. Plant Breed 131, 584-590. DOI:10.1111/j.1439-0523.2012.01989.xURL2-acetyl-1-pyrroline (2AP) is the principal compound responsible for grain fragrance in rice. In fragrant rice cultivars, BADH2 (betaine-aldehyde dehydrogenase 2) is inactivated. Here, we describe the effect of amiRNA (artificial microRNA) transgenesis targeted at BADH2 in rice. BADH2 (but not BADH1) expression was down-regulated in transgenic lines where the amiRNA was driven by the maize ubiquitin promoter, and in these lines, grain 2AP content was also significantly elevated; meanwhile, 2AP could not be detected in the grain of wild-type lines. The leaf proline content in the transgenics was increased through the simultaneous down-regulation of PRODH and up-regulation of P5CS. In transgenic lines where the same amiRNA was driven by the endosperm-specific promoter GluC, no effect on BADH2 expression was detectable, and the content of grain 2AP did not differ from that present in wild-type grains. [本文引用: 3]
[29]
ChenSH, WuJ, YangY, ShiWW, XuML (2006). The fgr gene responsible for rice fragrance was restricted within 69 kb. Plant Sci 171, 505-514. DOI:10.1016/j.plantsci.2006.05.013URLFrom more than 100 volatile flavor components, 2-acetyl-1-pyrroline (2AP) stands out as the key component of aroma in both basmati- and jasmine-style fragrant rice. It has been reported that a single recessive gene (fgr) on chromosome 8 is responsible for the production of 2AP. Our initial mapping efforts with SSR markers confirmed the previous mapping result, and placed the fgr locus between RM8264 and RM3459 with the physical distance of 800kb. The fgr region was then saturated with high-density markers developed by exploiting sequence diversities between indica and japonica rice subspecies. After mapping with segregant populations consisting of totally 2891 individuals, the fgr locus was restricted to an interval of 69kb flanked by the left marker L02 and the right marker L06. Furthermore, the fgr locus was confirmed by simultaneous investigation of both genotypes and 2AP levels for the key recombinants and their offspring. Sequence analysis of the fgr region revealed three candidate genes encoding respective eukaryotic-type carbonic anhydrase, 3-methylcrotonyl-CoA carboxylase beta chain, and betaine aldehyde dehydrogenase. The result presented here would pave the way for both cloning and marker-assisted selection of the fgr gene. [本文引用: 2]
[30]
ChenSH, YangY, ShiWW, JiQ, HeF, ZhangZD, ChengZK, LiuXN, XuML (2008). Badh2, encoding betaine aldehyde dehydrogenase, inhibits the biosynthesis of 2-ace- tyl-1-pyrroline, a major component in rice fragrance. Plant Cell 20, 1850-1861. [本文引用: 12]
[31]
CordeiroGM, ChristopherMJ, HenryRJ, ReinkeRF (2002). Identification of microsatellite markers for fragr- ance in rice by analysis of the rice genome sequence.Mol Breed 9, 245-250. DOI:10.1023/A:1020350725667URL [本文引用: 1]
[32]
DaygonVD, PrakashS, CalingacionM, RiedelA, OvendenB, SnellP, MitchellJ, FitzgeraldM (2016). Understanding the jasmine phenotype of rice through metabolite profiling and sensory evaluation.Metabolomics 12, 63. DOI:10.1007/s11306-016-0989-6URLIntroduction Aromatic rices are culturally and economically important for many countries in Asia. Investigation of the volatile compounds emitted by rice during cooking is the key to understanding the... [本文引用: 1]
[33]
DongYJ, TsuzukiE, TeraoH, YosimuraA, YasuiH (2001a). Inheritance of aroma and identification of RELP markers linked to aroma genes in two rice cultivars ( Oryza sativa L.). Bull Fac Agric 48, 59-65. [本文引用: 1]
[34]
DongYJ, TsuzukiE, TeraoH (2001b). Trisomic genetic analysis of aroma in three Japanese native rice varieties ( Oryza sativa L.). Euphytica 117, 191-196. DOI:10.1023/A:1026502115436URLInformation on the genetics of aroma in rice facilitates breeding and selection of new aromatic varieties with high yield and good quality. Objective of the present study was to make clear the number of genes controlling aroma, and the allelism of aroma genes and the location of aroma gene(s) on the chromosome in three Japanese native aromatic rice varieties (Kabashiko, Shiroikichi and Henroyori). Lack of leaf aroma in all F 1 plants of non-aromatic/aromatic crosses indicated the recessive nature of aroma, and the segregation ratios (3:1) of non-aromatic to aromatic plants in its F 2 populations from Nipponbare/aromatic varieties crosses revealed that each of the three aromatic varieties contains a single recessive gene for aroma. Through trisomic analysis, the segregation of non-aromatic and aromatic plants in all F 2 populations from the crosses between trisomics lines NT8, with an extra chromosome 8, and aromatic varieties deviated significantly from disomic segregation of 3:1 ratios, and fitted to trisomic segregation, however, in other F 2 populations derived from other 7 types of trisomic F 1 plants, the segregation ratios of non-aromatic to aromatic were 3:1, indicating that the single recessive aroma gene was located on chromosome 8 in three aromatic varieties. [本文引用: 1]
[35]
FitzgeraldMA, Sackville HamiltonNR, CalingacionMN, VerhoevenHA, ButardoVM (2008). Is there a second fragrance gene in rice?Plant Biotechnol J 6, 416-423. DOI:10.1111/pbi.2008.6.issue-4URL [本文引用: 1]
[36]
FuHY, KimSY, ParkWD (1995). High-level tuber expression and sucrose inducibility of a potato Sus4 sucrose syn- thase gene require 5' and 3' flanking sequences and the leader intron.Plant Cell 7, 1387-1394. [本文引用: 1]
[37]
GarlandS, LewinL, BlakeneyA, ReinkeR, HenryR (2000). PCR-based molecular markers for the fragrance gene in rice ( Oryza sativa L.). Theor Appl Genet 101, 364-371. [本文引用: 1]
[38]
GoufoP, FalcoV, BritesC, WesselDF, KratzS, RosaEAS, CarrancaC, TrindadeH (2014). Effect of elevated carbon dioxide concentration on rice quality: nutritive val- ue, color, milling, cooking, and eating qualities.Cereal Chem J 91, 513-521. DOI:10.1094/CCHEM-12-13-0256-RURLIncrease in the atmospheric carbon dioxide concentration ([CO2]) enhanced the concentration of carbohydrates in rice grains as per results of a previous study. However, its impact on other quality traits is little known. To investigate the effect of CO2 levels (375 [本文引用: 1]
[39]
HalfordNG, CurtisTY, ChenZW, HuangJH (2014). Effects of abiotic stress and crop management on cereal grain composition: implications for food quality and safety.J Exp Bot 66, 1145-1156. DOI:10.1093/jxb/eru473PMID:25428997URLThe effects of abiotic stresses and crop management on cereal grain composition are reviewed, focusing on phytochemicals, vitamins, fibre, protein, free amino acids, , and oils. These effects are discussed in the context of nutritional and processing quality and the potential for formation of processing contaminants, such as , , hydroxymethylfurfuryl, and trans fatty acids. The implications of climate change for cereal grain quality and food safety are considered. It is concluded that the identification of specific environmental stresses that affect grain composition in ways that have implications for food quality and safety and how these stresses interact with genetic factors and will be affected by climate change needs more investigation. Plant researchers and breeders are encouraged to address the issue of processing contaminants or risk appearing out of touch with major end-users in the food industry, and not to overlook the effects of environmental stresses and crop management on crop composition, quality, and safety as they strive to increase yield. [本文引用: 1]
[40]
HaunW, CoffmanA, ClasenBM, DemorestZL, LowyA, RayE, RetterathA, StoddardT, JuilleratA, CedroneF, MathisL, VoytasDF, ZhangF (2014). Improved soybean oil quality by targeted mutagenesis of the fatty acid desaturase 2 gene family. Plant Biotechnol J 12, 934-940. DOI:10.1111/pbi.12201PMID:24851712URLSummary Soybean oil is high in polyunsaturated fats and is often partially hydrogenated to increase its shelf life and improve oxidative stability. The trans-fatty acids produced through hydrogenation pose a health threat. Soybean lines that are low in polyunsaturated fats were generated by introducing mutations in two fatty acid desaturase 2 genes ( FAD2-1A and FAD2-1B ), which in the seed convert the monounsaturated fat, oleic acid, to the polyunsaturated fat, linoleic acid. Transcription activator-like effector nucleases (TALENs) were engineered to recognize and cleave conserved DNA sequences in both genes. In four of 19 transgenic soybean lines expressing the TALENs, mutations in FAD2-1A and FAD2-1B were observed in DNA extracted from leaf tissue; three of the four lines transmitted heritable FAD2 -1 mutations to the next generation. The fatty acid profile of the seed was dramatically changed in plants homozygous for mutations in both FAD2-1A and FAD2-1B : oleic acid increased from 20% to 80% and linoleic acid decreased from 50% to under 4%. Further, mutant plants were identified that lacked the TALEN transgene and only carried the targeted mutations. The ability to create a valuable trait in a single generation through targeted modification of a gene family demonstrates the power of TALENs for genome engineering and crop improvement. [本文引用: 1]
[41]
HeQ, ParkYJ (2015). Discovery of a novel fragrant allele and development of functional markers for fragrance in rice.Mol Breed 35, 217. DOI:10.1007/s11032-015-0412-4URL [本文引用: 18]
[42]
HeQ, YuJ, KimTS, ChoYH, LeeYS, ParkYJ (2015). Resequencing reveals different domestication rate for BA- DH1 and BADH2 in rice(Oryza sativa). PLoS One 10, e0134801.
[43]
JeonJS, LeeS, JungKH, JunSH, KimC, AnG (2000). Tissue-preferential expression of a rice α-tubulin gene, Os- TubA1, mediated by the first intron. Plant Physiol 12, 1005-1014. [本文引用: 1]
[44]
JinQS, QinBQ, YanWC, LuoRB (1995). Tagging of a gene for aroma in rice by RAPD and RFLP(I).Acta Agric Zhejiangensis 7, 439-442. URLThe feasibility of identifying molecular markers linked to aromatic gene in rice was investigated utilizing random amplified polimorphic DNA (RAPD) technology. 300 random primers were screened against bulked DNA samples of genotyped individuals from the F 2 segregating population of CT9993/KDML105 for polymorphic DNA fragnment linked to the aromatic gene. One primer (generated by a 5'-GAACGGACTC-3' decamer) was idendified to amplified 1.5 kb polymorphic fragment only in non-aromatic lines and other non-aromatic cultivars, and it was shown to be closely linked to the aromatic locus. This type of analysis offers an effective means of identifying useful molecular markers and applying them to rice breeding selection strategies. [本文引用: 1]
[45]
JinQS, QinBQ, YanWC, LuoRB (1996). Tagging of a gene for aroma in rice by RAPD and RFLP(II).Acta Agric Zhe- jiangensis 8, 19-23. URLThe feasibility of identifying molecular markers linked to aromatic gene in rice was investigated utilizing random amplified polimorphic DNA (RAPD) technology. 300 random primers were screened against bulked DNA samples of genotyped individuals from the F 2 segregating population of CT9993/KDML105 for polymorphic DNA fragnment linked to the aromatic gene. One primer (generated by a 5'-GAACGGACTC-3' decamer) was idendified to amplified 1.5 kb polymorphic fragment only in non-aromatic lines and other non-aromatic cultivars, and it was shown to be closely linked to the aromatic locus. This type of analysis offers an effective means of identifying useful molecular markers and applying them to rice breeding selection strategies. [本文引用: 1]
[46]
JinQS, WatersD, CordeiroGM, HenryRJ, ReinkeRF (2003). A single nucleotide polymorphism (SNP) marker linked to the fragrance gene in rice ( Oryza sativa L.). Plant Sci 165, 359-364. DOI:10.1016/S0168-9452(03)00195-XURLThe whole rice genome sequence was used to assist in the identification of a single nucleotide polymorphism (SNP) marker linked to the fragrance gene ( fgr) in rice. Genes flanked by restriction fragment length polymorphism and microsatellite markers known to be linked to the fragrance gene were identified by DNA sequence alignment of EST sequences against BAC clones covering this region of chromosome eight. Re-sequencing and comparison of parts of these genes derived from a fragrant and a non-fragrant cultivar revealed only one SNP (a C/T transition) in more than 6 kbp of sequence from 14 genes. Ten of eleven fragrant genotypes and six of 14 non-fragrant genotypes tested carried the C allele. This approach indicated a generally low level of SNP polymorphism in cultivated rice suggesting that association of SNP with phenotypes should be an efficient path to gene discovery in cultivated rice. [本文引用: 1]
[47]
JuwattanasomranR, SomtaP, ChankaewS, ShimizuT, WongpornchaiS, KagaA, SrinivesP (2011). A SNP in GmBADH2 gene associates with fragrance in vegetable soybean variety ‘Kaori’ and SNAP marker development for the fragrance. Theor Appl Genet 122, 533-541. DOI:10.1007/s00122-010-1467-6PMID:21046066URLAbstract Fragrance in soybean is due to the presence of 2-acetyl-1-pyrroline (2AP). BADH2 gene coding for betaine aldehyde dehydrogenase has been identified as the candidate gene responsible for fragrance in rice (Oryza sativa L.). In this study, using the RIL population derived from fragrant soybean cultivar "Kaori" and non-fragrant soybean cultivar "Chiang Mai 60" (CM60), STS markers designed from BADH2 homolog were found associating with 2AP production. Genetic mapping demonstrated that QTL position of fragrance and 2AP production coincides with the position of GmBADH2 (Glycine max betaine aldehyde dehydrogenase 2). Sequence comparison of GmBADH2 between Kaori and non-fragrant soybeans revealed non-synonymous single-nucleotide polymorphism (SNP) in exon 10. Nucleotide substitution of G to A in the exon results in an amino acid change of glycine (GGC; G) to aspartic acid (GAC; D) in Kaori. The amino acid substitution changes the conserved EGCRLGPIVS motif of GmBADH2, which is essential for functional activity of GmBADH2 protein, to EGCRLDPIVS motif, suggesting that the SNP in GmBADH2 is responsible for the fragrance in Kaori. Five single nucleotide-amplified polymorphism (SNAP) markers which are PCR-based allele specific SNP markers were developed for fragrance based on the SNP in GmBADH2. Two markers specific to A allele produced a band in only Kaori, while three markers specific to G alleles produced a band in only CM60. The simple PCR-based allele specific SNAP markers developed in the present study are useful in marker-assisted breeding of fragrant soybean. [本文引用: 1]
[48]
JuwattanasomranR, SomtaP, KagaA, ChankaewS, ShimizuT, SorajjapinunW, SrinivesP (2012). Identification of a new fragrance allele in soybean and development of its functional marker.Mol Breed 29, 13-21. DOI:10.1007/s11032-010-9523-0URLAbstract122 population using Chamame as the fragrance donor revealed that the location of the fragrance QTL nearly coincided with that of the functional marker, confirming the association between GmBADH2 and fragrance in Chamame. [本文引用: 1]
[49]
KovachMJ, CalingacionMN, FitzgeraldMA, McCouchSR (2009). The origin and evolution of fragrance in rice ( Oryza sativa L.). Proc Natl Acad Sci USA 106, 14444-14449. DOI:10.1073/pnas.0904077106PMID:19706531URLFragrance in the grain is one of the most highly valued grain quality traits in rice, yet the origin and evolution of the betaine aldehyde dehydrogenase gene (BADH2) underlying this trait remains unclear. In this study, we identify eight putatively nonfunctional alleles of the BADH2 gene and show that these alleles have distinct geographic and genetic origins. Despite multiple origins of the fragrance trait a single allele, badh2.1, is the predominant allele in virtually all fragrant rice varieties today, including the widely recognized Basmati and Jasmine types. Haplotype analysis allowed us to establish a single origin of the badh2.1 allele within the Japonica varietal group and demonstrate the introgression of this allele from Japonica to Indica. Basmati-like accessions were nearly identical to the ancestral Japonica haplotype across a 5.3-Mb region flanking BADH2 regardless of their fragrance phenotype, demonstrating a close evolutionary relationship between Basmati varieties and the Japonica gene pool. These results clarify the relationships among fragrant rice varieties and challenge the traditional assumption that the fragrance trait arose in the Indica varietal group. [本文引用: 12]
[50]
KusanoM, YangZG, OkazakiY, NakabayashiR, FukushimaA, SaitoK (2015). Using metabolomic approaches to explore chemical diversity in rice.Mol Plant 8, 58-67. DOI:10.1016/j.molp.2014.11.010PMID:25578272URLRice (Oryza sativa) is an excellent resource to provide not only calories for life but also bioactive compounds for welfare. Chemical diversity of the rice metabolome can be estimated by metabolomic approaches such as mass spectrometry (MS) and/or nuclear magnetic resonance spectroscopy. Current knowledge on the primary and secondary metabolites in rice detected by MS, challenges in estimating the structure(s) of unknown metabolites, and the examples of metabolite profiling of rice are presented. [本文引用: 1]
[51]
LiT, LiuB, SpaldingMH, WeeksDP, YangB (2012). High-efficiency TALEN-based gene editing produces disease-resistant rice.Nat Biotechnol 30, 390-392. DOI:10.1038/nbt.2199PMID:22565958URLThe article focuses on a study which examined the use of transcriptor activator-like (TAL) effector nucleases (TALEN) in editing a specific S gene in rice to prevent the virulence strategy of Xanthomonas oryzae. For TALEN-based disruption, the rice bacterial blight susceptibility gene was targeted. Results of the study showed that bacterial infection assays were resistant to infection by pathogenic Xoo. [本文引用: 1]
[52]
LiangZ, ZhangK, ChenKL, GaoCX (2014). Targeted mutagenesis in Zea mays using TALENs and the CRISPR/ Cas system. J Genet Genom 41, 63-68. DOI:10.1016/j.jgg.2013.12.001PMID:24576457URLTranscription activator-like effector nucleases (TALENs) and clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated (Cas) systems have emerged as powerful tools for genome editing in a variety of species. Here, we report, for the first time, targeted mutagenesis in Zea mays using TALENs and the CRISPR/Cas system. We designed five TALENs targeting 4 genes, namely ZmPDS, ZmIPK1A, ZmIPK, ZmMRP4, and obtained targeting efficiencies of up to 23.1% in protoplasts, and about 13.3% to 39.1% of the transgenic plants were somatic mutations. Also, we constructed two gRNAs targeting the ZmIPK gene in maize protoplasts, at frequencies of 16.4% and 19.1%, respectively. In addition, the CRISPR/Cas system induced targeted mutations in Z. mays protoplasts with efficiencies (13.1%) similar to those obtained with TALENs (9.1%). Our results show that both TALENs and the CRISPR/Cas system can be used for genome modification in maize. [本文引用: 1]
[53]
LorieuxM, PetrovM, HuangN, GuiderdoniE, GhesquièreA (1996). Aroma in rice: genetic analysis of a quantitative trait.Theor Appl Genet 93, 1145-1151. DOI:10.1007/BF00230138PMID:24162494URLA new approach was developed which succeeded in tagging for the first time a major gene and two QTLs controlling grain aroma in rice. It involved a combination of two techniques, quantification of volatile compounds in the cooking water by gas chromatography, and molecular marker mapping. Four types of molecular marker were used (RFLPs, RAPDs, STSs, isozymes). Evaluation and mapping were performed on a doubled haploid line population which (1) conferred a precise character evaluation by enabling the analysis of large quantities of grains per genotype and (2) made possible the comparison of gas chromatography results and sensitive tests. The population size (135 lines) provided a good mapping precision. Several markers on chromosome 8 were found to be closely linked to a major gene controlling the presence of 2-acetyl-1-pyrroline (AcPy), the main compound of rice aroma. Moreover, our results showed that AcPy concentration in plants is regulated by at least two chromosomal regions. Estimations of recombination fractions on chromosome 8 were corrected for strong segregation distortion. This study confirms that AcPy is the major component of aroma. Use of the markers linked to AcPy major gene and QTLs for marker-assisted selection by successive backcrosses may be envisaged. [本文引用: 2]
[54]
MahattanataweeK, RouseffRL (2014). Comparison of aroma active and sulfur volatiles in three fragrant rice cultivars using GC-Olfactometry and GC-PFPD.Food Chem 154, 1-6. DOI:10.1016/j.foodchem.2013.12.105PMID:24518308URLAroma volatiles from three cooked fragrant rice types (Jasmine, Basmati and Jasmati) were characterised and identified using SPME GC–O, GC–PFPD and confirmed using GC–MS. A total of 26, 23, and 22 aroma active volatiles were observed in Jasmine, Basmati and Jasmati cooked rice samples. 2-Acetyl-1-pyrroline was aroma active in all three rice types, but the sulphur-based, cooked rice character impact volatile, 2-acetyl-2-thiazoline was aroma active only in Jasmine rice. Five additional sulphur volatiles were found to have aroma activity: dimethyl sulphide, 3-methyl-2-butene-1-thiol, 2-methyl-3-furanthiol, dimethyl trisulphide, and methional. Other newly-reported aroma active rice volatiles were geranyl acetate, β-damascone, β-damascenone, and ɑ-ionone, contributing nutty, sweet floral attributes to the aroma of cooked aromatic rice. The first two principal components from the principal component analysis of sulphur volatiles explained 60% of the variance. PC1 separated Basmati from the other two cultivars and PC2 completely separated Jasmine from Jasmati cultivars. [本文引用: 2]
[55]
MathureSV, JawaliN, ThenganeRJ, NadafAB (2014). Comparative quantitative analysis of headspace volatiles and their association with BADH2 marker in non-basmati scented, basmati and non-scented rice(Oryza sativa L.) cultivars of India. Food Chem 142, 383-391. DOI:10.1016/j.foodchem.2013.07.066PMID:24001856URLFor the first time, 91 Indian rice (Oryza sativa L.) cultivars, belonging to non-basmati scented (77), basmati (9) and non-scented (5) categories, were quantitatively analysed for 23 headspace volatiles. In addition, the BADH2 marker related to 2-acetyl-1-pyrroline (2AP) synthesis is validated among the cultivars representing these categories. The non-basmati scented cultivars (15) excelled in 2-acetyl-1-pyrroline (2AP) content compared to basmati cultivars. The major aroma volatile (2AP) correlated positively with 1-tetradecene and indole, and negatively with benzyl alcohol. On the basis of nonanal, octanal, decanal and 1-octen-3-ol contents, basmati cultivars were different from non-scented cultivars. Benzyl alcohol, 2-phenylethanol, 2-amino acetophenone, indole, 1-hexanol and nonanoic acid exhibited significant variation among rice categories under study. This study reports 16 non-basmati scented cultivars with variations in the BADH2 locus rendering a marker based on 8bp deletion in BADH2 as unsuitable for MAS in rice cultivars under study. [本文引用: 1]
[56]
MurtyDS, NicodemusKD, HouseLR (1982). Inheritance of basmati and dimpled seed in sorghum.Crop Sci 22, 1080-1082. DOI:10.2135/cropsci1982.0011183X002200050046xURLStudies on F, F, F, and backcross generations of three crosses and their reciprocals in sorghum, (L.) Moench, showed that the inheritance of (scented plant) and dimpled seed characters is controlled by two independent recessive genes. The plump seed character exhibited xenia. [本文引用: 1]
[57]
MyintKM, ArikitS, WanchanaS, YoshihashiT, Choo- wongkomonK, VanavichitA (2012). A PCR-based mar- ker for a locus conferring the aroma in Myanmar rice ( Oryza sativa L.). Theor Appl Genet 125, 887-896. DOI:10.1007/s00122-012-1880-0PMID:22576235URLAromatic rice is an important commodity for international trade, which has encouraged the interest of rice breeders to identify the genetic control of rice aroma. The recessive Os2AP gene, which is located on chromosome 8, has been reported to be associated with rice aroma. The 8-bp deletion in exon 7 is an aromatic allele that is present in most aromatic accessions, including the most popular aromatic rice varieties, Jasmine and Basmati. However, other mutations associated with aroma have been detected, but the other mutations are less frequent. In this study, we report an aromatic allele, a 3-bp insertion in exon 13 of Os2AP , as a major allele found in aromatic rice varieties from Myanmar. The insertion is in frame and causes an additional tyrosine (Y) in the amino acid sequence. However, the mutation does not affect the expression of the Os2AP gene. A functional marker for detecting this allele was developed and tested in an aroma-segregating F 2 population. The aroma phenotypes and genotypes showed perfect co-segregation of this population. The marker was also used for screening a collection of aromatic rice varieties collected from different geographical sites of Myanmar. Twice as many aromatic Myanmar rice varieties containing the 3-bp insertion allele were found as the varieties containing the 8-bp deletion allele, which suggested that the 3-bp insertion allele originated in regions of Myanmar. [本文引用: 1]
[58]
NiuXL, TangW, HuangWZ, RenGJ, WangQL, LuoD, XiaoYY, YangSM, WangF, LuBR, GaoFY, LuTG, LiuYS (2008). RNAi-directed downregulation of OsBADH2 results in aroma (2-acetyl-1-pyrroline) production in rice(Ory- za sativa L.). BMC Plant Biol 8, 100. DOI:10.1186/1471-2229-8-100PMID:2588449URLpAbstract/p pBackground/p pAromatic rice is popular worldwide because of its characteristic fragrance. Genetic studies and physical fine mapping reveal that a candidate gene (itfgr/it/itOsBADH2/it) homologous to itbetaine aldehyde dehydrogenase /itis responsible for aroma metabolism in fragrant rice varieties, but the direct evidence demonstrating the functions of itOsBADH2 /itis lacking. To elucidate the physiological roles of itOsBADH2/it, sequencing approach and RNA interference (RNAi) technique were employed to analyze allelic variation and functions of itOsBADH2 /itgene in aroma production. Semi-quantitative, real-time reverse transcription-polymerase chain reaction (RT-PCR), as well as gas chromatography-mass spectrometry (GC-MS) were conducted to determine the expression levels of itOsBADH2 /itand the fragrant compound in wild type and transgenic itOsBADH2/it-RNAi repression lines, respectively./p pResults/p pThe results showed that multiple mutations identical to itfgr /itallele occur in the 13 fragrant rice accessions across China; itOsBADH2 /itis expressed constitutively, with less expression abundance in mature roots; the disrupted itOsBADH2 /itby RNA interference leads to significantly increased 2-acetyl-1-pyrroline production./p pConclusion/p pWe have found that the altered expression levels of itOsBADH2 /itgene influence aroma accumulation, and the prevalent aromatic allele probably has a single evolutionary origin./p [本文引用: 3]
[59]
OotsukaK, TakahashiI, TanakaK, ItaniT, TabuchiH, YoshihashiT, TonouchiA, IshikawaR (2014). Genetic polymorphisms in Japanese fragrant landraces and novel fragrant allele domesticated in northern Japan.Breed Sci 64, 115-124. DOI:10.1270/jsbbs.64.115PMID:4065318URLABSTRACT Rice fragrance is an important characteristic for Southeast Asian consumers, and fragrant landraces from Japan were first recorded in the 17th century. Principal component analysis clearly showed that Japanese fragrant landraces were genetically different from non-Japanese fragrant landraces. Japanese fragrant landraces were composed of six clades, none of which carried the most common fragrance mutation, an 8-bp deletion in exon 7 of Badh2. Fragrant landraces comprised two major groups carrying different Badh2 mutations. One group carried a known SNP at exon13 and the other a SNP at the exon1-intron1 junction as splicing donor site. The latter was considered to be a potential splicing mutant group as a novel allele at Badh2. Heterozygosity (He) scores in the two fragrant groups were not significantly different from non-fragrant landraces and modern cultivars. However, lower He scores were found around the Badh2 locus in the two groups. The potential splicing mutant group showed a more extended haplotype than the E13 SNP group. A likely causal factor responsible for loss of function is a novel splicing mutation allele that may have been generated quite recently. The fragrance allele has dispersed as a result of out-crossing under local environmental conditions. [本文引用: 4]
[60]
PauleCM, PowersJJ (1989). Sensory and chemical examination of aromatic and nonaromatic rices.J Food Sci 54, 343-346. DOI:10.1111/j.1365-2621.1989.tb03076.xURLABSTRACTTwo different panels were trained to evaluate the character of four aromatic and two nonaromatic rices. The judges used terms with which they were familiar to describe the odor of aromatic rice: pandan for the orientals and popcorn for the nonorientals. The rice volatiles were separated by a gas chromatograph equipped with a packed column and a sniffing port. The peaks possessing the aromatic principle were identified by GC-MS and correlated with the desirable aroma terms. 2-Acetyl-l-pyrroline showed highly significant positive correlations with the descriptive terms; hexanol showed negative correlations. [本文引用: 1]
[61]
PengB, KongHL, LiYB, WangLQ, ZhongM, SunL, GaoGJ, ZhangQL, LuoLJ, WangGW, XieWB, ChenJX, YaoW, PengY, LeiL, LianXM, XiaoJH, XuCG, LiXH, HeYQ (2014a).OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat Commun 5, 4847. DOI:10.1038/ncomms5847PMID:4175581URLGrains from cereals contribute an important source of protein to human food, and grain protein content (GPC) is an important determinant of nutritional quality in cereals. Here we show that the quantitative trait locus (QTL) qPC1 in rice controls GPC by regulating the synthesis and accumulation of glutelins, prolamins, globulins, albumins and starch. qPC1 encodes a putative amino acid transporter OsAAP6, which functions as a positive regulator of GPC in rice, such that higher expression of OsAAP6 is correlated with higher GPC. OsAAP6 greatly enhances root absorption of a range of amino acids and has effects on the distribution of various amino acids. Two common variations in the potential cis-regulatory elements of the OsAAP6 5鈥-untranslated region seem to be associated with GPC diversity mainly in indica cultivars. Our results represent the first step toward unravelling the mechanism of regulation underlying natural variation of GPC in rice. Grain protein content (GPC) contributes to the nutritional quality of cereals. Here, the authors show that the OsAAP6 quantitative trait locus in rice controls GPC by regulating the synthesis and accumulation of several grain storage proteins and starch. [本文引用: 1]
[62]
PengB, WangLQ, FanCC, JiangGH, LuoLJ, LiYB, HeYQ (2014b). Comparative mapping of chalkiness components in rice using five populations across two environments.BMC Genet 15, 49. DOI:10.1186/1471-2156-15-49PMID:24766995URLBackground Chalkiness is a major constraint in rice production because it is one of the key factors determining grain quality (appearance, processing, milling, storing, eating, and cooking quality) and price. Its reduction is a major goal, and the primary purpose of this study was to dissect the genetic basis of grain chalkiness. Using five populations across two environments, we also sought to determine how many quantitative trait loci (QTL) can be consistently detected. We obtained an integrated genetic map using the data from five mapping populations and further confirmed the reliability of the identified QTL. Results A total of 79 QTL associated with six chalkiness traits (chalkiness rate, white core rate, white belly rate, chalkiness area, white core area, and white belly area) were mapped on 12 chromosomes using five populations (two doubled haploid lines and three recombinant inbred lines) across two environments (Hainan in 2004 and Wuhan in 2004). The final integrated map included 430 markers; 58.3% of the QTL clustered together (QTL clusters), 71.4% of the QTL clusters were identified in two or more populations, and 36.1% of the QTL were consistently detected in the two environments. The QTL could be detected again and showed dominance (qWBR1, qWBR8, qWBR12, and qCR5) or overdominance effects (qWCR7) for the rate of the white belly or white core, respectively, and all four QTL clusters derived from Zhenshan 97 controlling white belly rate were stably and reliably identified in an F2 population. Conclusions Our results identified 79 QTL associated with six chalkiness traits using five populations across two environments and yielded an integrated genetic map, indicating most of the QTL clustered together and could be detected in different backgrounds. The identified QTL were stable and reliable in the F2 population, and they may facilitate our understanding of the QTL related to chalkiness traits in different populations and various environments, the relationships among the various chalkiness QTL, and the genetic basis for chalkiness. Thus, our results may be immediately used for map-based cloning of important QTL and in marker-assisted breeding to improve grain quality in rice breeding. [本文引用: 1]
[63]
PrathephaP (2009). The fragrance ( >fgr) gene in natural populations of wild rice(Oryza rufipogon Griff.). Genet Re- sour Crop Evol 56, 13-18. [本文引用: 2]
[64]
SchieberleP (1990). The role of free amino acids present in yeast as precursors of the odorants 2-acetyl-1-pyrroline and 2-acetyltetrahydropyridine in wheat bread crust.Z Le- bensm Unters Forsch 191, 206-209. DOI:10.1007/BF01197621URLWir konnten kürzlich zeigen, da08 die B01ckerhefe eine entscheidende Quelle von Vorl01ufern zur Bildung der R02staromastoffe 2-Acetyl-1-pyrrolin (ACPY) und 2-Acetyltetrahydropyridin (ACTPY) in der Wei08brotkruste ist. Um die Rolle freier Aminos01uren der Hefe für die Bildung beider Geruchsstoffe zu untersuchen, wurden die Konzentrationen der in Hefe vorliegenden Aminos01uren bestimmt. Elf Aminos01uren, deren Konzentrationen 60 mg/100 g Trockenhefe überstiegen, wurden in Modellversuchen mit 2-Oxopropanal umgesetzt und die freigesetzten Mengen von ACPY und ACTPY über eine Isotopenverdünnungsanalyse bestimmt. ACPY wurde sowohl aus Prolin als auch aus Ornithin freigesetzt, w01hrend ACTPY ausschlie08lich aus Prolin entstand. Aus den übrigen Aminos01uren wurden die beiden Aromastoffe nicht gebildet. Weitere Versuche ergaben, da08 die Bildung von ACPY aus Ornithin über 4-Aminobutyraldehyd und 1-Pyrrolin als Intermediate erfolgt. Die Menge an freiem Ornithin in der Hefe war mehr als dreimal so gro08 wie die des freien Prolins. Weiterhin erh02hten Zus01tze von Prolin bzw. Ornithin zu Weizenteigen die Konzentrationen von ACPY in der Kruste um den Faktor 2 bzw. 4. Die Daten lie08en den Schlu08 zu, da08 Ornithin der wichtigste Vorl01ufer zur Bildung von ACPY beim Backen ist. [本文引用: 1]
[65]
ShanQW, WangYP, ChenKL, LiangZ, LiJ, ZhangY, ZhangK, LiuJX, VoytasDF, ZhengXL, ZhangY, GaoCX (2013). Rapid and efficient gene modification in rice and Brachypodium using TALENs. Mol Plant 6, 1365-1368. DOI:10.1093/mp/sss162PMID:23288864URL [本文引用: 4]
[66]
ShanQW, ZhangY, ChenKL, ZhangK, GaoCX (2015). Creation of fragrant rice by targeted knockout of the OsBADH2 gene using TALEN technology. Plant Biotechnol J 13, 791-800. DOI:10.1111/pbi.12312PMID:25599829URLSummary Fragrant rice is favoured worldwide because of its agreeable scent. The presence of a defective badh2 allele encoding betaine aldehyde dehydrogenase (BADH2) results in the synthesis of 2-acetyl-1-pyrroline (2AP), which is a major fragrance compound. Here, transcription activator-like effector nucleases (TALENs) were engineered to target and disrupt the OsBADH2 gene. Six heterozygous mutants (30%) were recovered from 20 transgenic hygromycin-resistant lines. Sanger sequencing confirmed that these lines had various indel mutations at the TALEN target site. All six transmitted the BADH2 mutations to the T1 generation; and four T1 mutant lines tested also efficiently transmitted the mutations to the T2 generation. Mutant plants carrying only the desired DNA sequence change but not the TALEN transgene were obtained by segregation in the T1 and T2 generations. The 2AP content of rice grains of the T1 lines with homozygous mutations increased from 0 to 0.35 0.75 mg/kg, which was similar to the content of a positive control variety harbouring the badh2-E7 mutation. We also simultaneously introduced three different pairs of TALENs targeting three separate rice genes into rice cells by bombardment and obtained lines with mutations in one, two and all three genes. These results indicate that targeted mutagenesis using TALENs is a useful approach to creating important agronomic traits. [本文引用: 13]
[67]
ShaoGN, TangA, TangSQ, LuoJ, JiaoGA, WuJL, HuPS (2011). A new deletion mutation of fragrant gene and the development of three molecular markers for fragrance in rice.Plant Breed 130, 172-176. DOI:10.1111/j.1439-0523.2009.01764.xURLFragrance is an important quality trait of rice, which is mainly controlled by loss-function of the betaine aldehyde dehydrogenase gene (badh2) on rice chromosome 8. Two deletion mutations of Badh2 have been reported to associate with fragrance in rice. Allelic test showed that the fragrant gene of ‘Zaimiaoxiangnuo’ is allelic to badh2-E7 (8 bp deletion in exon 7 of Badh2). Sequencing and BLAST analysis showed that the allele in ‘Zaimiaoxiangnuo’ is a new mutation, i.e. deletion of 803 bp between exons 4 and 5. A marker FMbadh2-E4-5 was developed for the detection of the new deletion site of fragrant gene. FMbadh2-E4-5 and the two previously reported functional markers were used to genotype the fragrant genes of four non-fragrant and 22 fragrant rice varieties. The deletions were absent in all non-fragrant rice varieties, but present in all fragrant rice varieties. Eight fragrant rice varieties were positive for the FMbadh2-E4-5 marker. These findings are useful in the breeding of fragrant rice varieties. [本文引用: 3]
[68]
ShaoGN, TangSQ, ChenML, WeiXJ, HeJW, LuoJ, JiaoGA, HuYC, XieLH, HuPS (2013). Haplotype variation at Badh2, the gene determining fragrance in rice. Genomics 101, 157-162. DOI:10.1016/j.ygeno.2012.11.010PMID:23220350URL78 Genetic diversity and structure have been done in this context. 78 Linkage disequilibrium in the Badh2 region has been elucidated. 78 Three new possible FNPs for Badh2 were obtained. 78 A new model for the evolution of Badh2 was proposed. [本文引用: 6]
[69]
ShiWW, YangY, ChenSH, XuML (2008). Discovery of a new fragrance allele and the development of functional markers for the breeding of fragrant rice varieties.Mol Bre- ed 22, 185-192. DOI:10.1007/s11032-008-9165-7URLThe recessive fgr gene on chromosome 8 is associated with rice fragrance. It has been reported that this gene is a non-functional badh2 allele and that the functional Badh2 allele encoding putative betaine aldehyde dehydrogenase (BADH2) could render rice non-fragrant. Here we report the discovery of a new badh2 allele and the development of functional markers for the badh2 locus. A total of 24 fragrant and ten non-fragrant rice varieties were studied and sequenced for their Badh2/badh2 loci. Of the 24 fragrant rice varieties, 12 were found to have the known badh2 allele ( badh2-E7 ), which has an 8-bp deletion and three single nucleotide polymorphisms ( SNPs ) in exon 7; the others had a novel null badh2 allele ( badh2-E2 ), which has a sequence identical to that of the Badh2 allele in exon 7, but with a 7-bp deletion in exon 2. Both null badh2 alleles are responsible for rice fragrance. Based on sequence divergence amongst the functional Badh2 and two null badh2 alleles, we developed functional markers which can be easily used to distinguish non-fragrant from fragrant rice and to differentiate between two kinds of fragrant rice. These functional markers will find their usefulness in breeding for fragrant rice varieties via marker-assisted selection. [本文引用: 7]
[70]
ShiYQ, ZhaoGC, XuXL, LiJY (2014). Discovery of a new fragrance allele and development of functional markers for identifying diverse fragrant genotypes in rice.Mol Breed 33, 701-708. DOI:10.1007/s11032-013-9986-xURLDiscovery of new fragrance alleles provides important genetic resources for breeding fragrant rice. In this study, a hybrid complementation test demonstrated the association of a new fragrance allele... [本文引用: 2]
[71]
StockerBD, RothR, JoosF, SpahniR, SteinacherM, ZaehleS, BouwmanL, RiX, PrenticeIC (2013). Multiple greenhouse-gas feedbacks from the land biosphere under future climate change scenarios.Nat Clim Change 3, 666-672. DOI:10.1038/nclimate1864URLAtmospheric concentrations of the three important greenhouse gases (GHGs) CO2, CH4 and N2O are mediated by processes in the terrestrial biosphere that are sensitive to climate and CO2. This leads to feedbacks between climate and land and has contributed to the sharp rise in atmospheric GHG concentrations since pre-industrial times. Here, we apply a process-based model to reproduce the historical atmospheric N2O and CH4 budgets within their uncertainties and apply future scenarios for climate, land-use change and reactive nitrogen (Nr) inputs to investigate future GHG emissions and their feedbacks with climate in a consistent and comprehensive framework1. Results suggest that in a business-as-usual scenario, terrestrial N2O and CH4 emissions increase by 80 and 45%, respectively, and the land becomes a net source of C by AD 2100. N2O and CH4 feedbacks imply an additional warming of 0.4–0.565°C by AD 2300; on top of 0.8–1.065°C caused by terrestrial carbon cycle and Albedo feedbacks. The land biosphere represents an increasingly positive feedback to anthropogenic climate change and amplifies equilibrium climate sensitivity by 22–27%. Strong mitigation limits the increase of terrestrial GHG emissions and prevents the land biosphere from acting as an increasingly strong amplifier to anthropogenic climate change. [本文引用: 1]
[72]
TianZX, QianQ, LiuQQ, YanMX, LiuXF, YanCJ, LiuGF, GaoZY, TangSZ, ZengDL, WangYH, YuJM, GuMH, LiJY (2009). Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities.Proc Natl Acad Sci USA 106, 21760-21765. DOI:10.1073/pnas.0912396106PMID:20018713URLMore than half of the world's population uses rice as a source of carbon intake every day. Improving grain quality is thus essential to rice consumers. The three main properties that determine rice eating and cooking quality—amylose content, gel consistency, and gelatinization temperature—correlate with one another, but the underlying mechanism of these properties remains unclear. Through an association analysis approach, we found that genes related to starch synthesis cooperate with each other to form a fine regulating network that controls the eating and cooking quality and defines the correlation among these three properties. Genetic transformation results verified the association findings and also suggested the possibility of developing elite cultivars through modification with selected major and/or minor starch synthesisrelated genes. [本文引用: 1]
[73]
TrossatC, RathinasabapathiB, HansonAD (1997). Trans- genically expressed betaine aldehyde dehydrogenase efficiently catalyzes oxidation of dimethylsulfoniopropionaldehyde and ω-aminoaldehydes.Plant Physiol 113, 1457-1461. DOI:10.1104/pp.113.4.1457PMID:12223684URLTobacco (Nicotianum tabacum L.) plants engineered to express a sugar beet (Beta vulgaris L.) betaine aldehyde dehydrogenase (BADH) cDNA acquired not only BADH activity, but also three other aldehyde dehydrogenase activities (those measured with 3-dimethylsulfoniopropionaldehyde, 3-aminopropionaldehyde, and 4-aminobutyraldehyde, all of which are natural products). This shows that BADH is not, as believed up to now, a substrate-specific enzyme and that its role may not be limited to glycine betaine synthesis. [本文引用: 2]
[74]
TsuzukiE, ShimokawaE (1990). Inheritance of aroma in rice.Euphytica 46, 157-159. DOI:10.1007/BF00022309URL [本文引用: 1]
[75]
VoytasDF, GaoCX (2014). Precision genome engineering and agriculture: opportunities and regulatory challenges.PLoS Biol 12, e1001877. DOI:10.1371/journal.pbio.1001877PMID:4051594URLPlant agriculture is poised at a technological inflection point. Recent advances in genome engineering make it possible to precisely alter DNA sequences in living cells, providing unprecedented control over a plant's genetic material. Potential future crops derived through genome engineering include those that better withstand pests, that have enhanced nutritional value, and that are able to grow on marginal lands. In many instances, crops with such traits will be created by altering only a few nucleotides among the billions that comprise plant genomes. As such, and with the appropriate regulatory structures in place, crops created through genome engineering might prove to be more acceptable to the public than plants that carry foreign DNA in their genomes. Public perception and the performance of the engineered crop varieties will determine the extent to which this powerful technology contributes towards securing the world's food supply. [本文引用: 1]
[76]
WanchanaS, KamolsukyunyongW, RuengphayakS, ToojindaT, TragoonrungS, VanavichitA (2005). A rapid construction of a physical contig across a 4.5 cM region for rice grain aroma facilitates marker enrichment for positional cloning.Sci Asia 31, 299-306. DOI:10.2306/scienceasia1513-1874.2005.31.299URL
[77]
WangYP, ChengX, ShanQW, ZhangY, LiuJX, GaoCX, QiuJL (2014). Simultaneous editing of three homoeoalleles in hexaploid bread wheat confers heritable resistance to powdery mildew.Nat Biotechnol 32, 947-951. DOI:10.1038/nbt.2969PMID:25038773URLSequence-specific nucleases have been applied to engineer targeted modifications in polyploid genomes, but simultaneous modification of multiple homoeoalleles has not been reported. Here we use transcription activator-like effector nuclease (TALEN) and clustered, regularly interspaced, short palindromic repeats (CRISPR)-Cas9 (refs. 4,5) technologies in hexaploid bread wheat to introduce targeted mutations in the three homoeoalleles that encode MILDEW-RESISTANCE LOCUS (MLO) proteins. Genetic redundancy has prevented evaluation of whether mutation of all three MLO alleles in bread wheat might confer resistance to powdery mildew, a trait not found in natural populations. We show that TALEN-induced mutation of all three TaMLO homoeologs in the same plant confers heritable broad-spectrum resistance to powdery mildew. We further use CRISPR-Cas9 technology to generate transgenic wheat plants that carry mutations in the TaMLO-A1 allele. We also demonstrate the feasibility of engineering targeted DNA insertion in bread wheat through nonhomologous end joining of the double-strand breaks caused by TALENs. Our findings provide a methodological framework to improve polyploid crops. [本文引用: 1]
[78]
WendtT, HolmP, StarkerC, ChristianM, VoytasD, Brinch-PedersenH, HolmeIB (2013). TAL effector nucleases induce mutations at a pre-selected location in the genome of primary barley transformants.Plant Mol Biol 83, 279-285. DOI:10.1007/s11103-013-0078-4URL [本文引用: 1]
[79]
YoshihashiT, HuongNTT, InatomiH (2002). Precursors of 2-acetyl-1-pyrroline, a potent flavor compound of an aromatic rice variety.J Agric Food Chem 50, 2001-2004. DOI:10.1021/jf011268sPMID:11902947URLAbstract The biological formation of a potent flavor compound, 2-acetyl-1-pyrroline, in the aromatic rice variety (Khao Dawk Mali 105) was studied in seedlings and callus of the rice. Concentrations of 2-acetyl-1-pyrroline were determined by GC-MS-SIM using an isotope dilution method. Increases in concentration occurred when proline, ornithine, and glutamate were present in solution, with proline increasing the concentration by more than 3-fold compared to that of the control. Results of tracer experiments using (15)N-proline, (15)N-glycine, and proline-1-(13)C indicated that the nitrogen source of 2-acetyl-1-pyrroline was proline, whereas the carbon source of the acetyl group was not the carboxyl group of proline. 2-acetyl-1-pyrroline was formed in the aromatic rice at temperatures below that of thermal generation in bread baking, and formed in the aerial part of aromatic rice from proline as the nitrogen precursor. [本文引用: 1]
[80]
YundaengC, SomtaP, TangphatsornruangS, WongpornchaiS, SrinivesP (2013). Gene discovery and functional marker development for fragrance in sorghum (Sor- ghum bicolor(L.) Moench). Theor Appl Genet 126, 2897-2906. DOI:10.1007/s00122-013-2180-zPMID:23975246URLAbstractKey messagebp deletion causes a premature stop codon inAbstract2-acetyl-1-pyrroline (2AP) is a potent volatile compound causing fragrance in several plants and foods. Seeds of some varieties of rice, sorghum and soybean possess fragrance. The genes responsible for fragrance in rice and soybean are orthologs that correspond to BADH2 population of 187 plants developed from a cross between KU630 (non-fragrant) and IS19912 (fragrant) was used. Leaves of F and F progenies were evaluated for fragrance by organoleptic test, while seeds of F plants were analyzed for 2AP. The tests consistently showed that the fragrance is controlled by a single recessive gene. Gene expression analysis of [本文引用: 1]
[81]
ZhaoXQ, FitzgeraldM (2013). Climate change: implications for the yield of edible rice.PLoS One 8, e66218. DOI:10.1371/journal.pone.0066218PMID:3680399URLAbstract Global warming affects not only rice yield but also grain quality. A better understanding of the effects of climate factors on rice quality provides information for new breeding strategies to develop varieties of rice adapted to a changing world. Chalkiness is a key trait of physical quality, and along with head rice yield, is used to determine the price of rice in all markets. In the present study, we show that for every 1% decrease in chalkiness, an increase of 1% in head rice yield follows, illustrating the dual impact of chalk on amount of marketable rice and its value. Previous studies in controlled growing conditions report that chalkiness is associated with high temperature. From 1980-2009 at IRRI, Los Ba os, the Philippines, annual minimum and mean temperatures, and diurnal variation changed significantly. The objective of this study was to determine how climate impacts chalkiness in field conditions over four wet and dry seasons. We show that low relative humidity and a high vapour pressure deficit in the dry season associate with low chalk and high head rice yield in spite of higher maximum temperature, but in the opposite conditions of the wet season, chalk is high and head rice yield is low. The data therefore suggest that transpirational cooling is a key factor affecting chalkiness and head rice yield, and global warming per se might not be the major factor that decreases the amount and quality of rice, but other climate factors in combination, that enable the crop to maintain a cool canopy. [本文引用: 1]
云南地方香稻与非香稻遗传多样性比较 1 2009
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
分子标记辅助选择选育香稻恢复系 3 2009
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ... ... 香味是水稻重要的品质性状之一, 优质香米的消费需求将持续增加.因此, 香稻新品种选育以及香味基因研究受到越来越多水稻遗传育种学家和植物科学家的重视.随着水稻功能基因组研究的快速发展, 关于水稻中香味基因的遗传基础、功能及其调控规律等将会越来越明朗.最近十几年, 基因测序技术突飞猛进, 大量不同来源的水稻品种和香稻种质资源完成测序分析, 发现了一系列香味基因的变异位点及其等位基因(He and Park, 2015, He et al., 2015).针对这些变异位点, 香味基因功能标记也被不断开发并已经广泛应用于香味基因筛选和杂交香稻骨干亲本培育.尽管香味基因功能标记已经用于香稻恢复系的选育(杜雪树等, 2009), 而且很多香稻品种已通过审定并在生产中得到较大面积的推广应用, 但是目前香稻遗传育种面临的主要问题依然是香型优良的恢复系极少, 减缓了香稻的遗传育种进程. ...
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ...
香稻胚乳的垩白性状研究及扫描电镜观察 1 2016
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
b). 水稻垩白性状的遗传研究进展 1 2016
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ...
水稻香味基因的研究进展 1 2009
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
一种水稻香味基因功能标记的开发 2 2008
... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
水稻香米基因标记的开发与应用 1 2008
... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ...
胚乳性状的质量-数量分析 1 1995
... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ...
24种香稻品种甜菜碱醛脱氢酶2基因突变位点的分析及分子标记开发 2011
两种筛选水稻badh2-E2类型香味基因分子标记的建立 1 2015
... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
水稻香味基因分子标记的开发及应用 5 2015
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ... ... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... ), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
不同来源水稻种质资源香味基因badh2位点的鉴定 2015
水稻香味基因的精细定位 2008
香稻研究进展 1 2009
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
1992
5 2008
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 为进一步解析水稻香味基因的遗传特点, 对巴斯马蒂香稻的香味基因进行全基因组关联分析, 发现在香稻的基因组内, 除了第8号染色体上的Badh2基因, 还存在2个控制稻米香味的数量性状位点(quantita- tive trait loci, QTL) , 它们分别位于水稻的第3和4号染色体上(Amarawathi et al., 2008).其中第4号染色体上的数量性状位点可能与Badh1基因有关, 而Badh1与Badh2属于同源基因, 并且水稻的Badh1基因与高粱(Sorghum bicolor)和大麦(Hordeum vulgare)基因组中的Badh1基因也高度同源(Bradbury et al., 2005a, 2008).近期, 有研究表明, Badh1基因与水稻在种子萌发阶段的耐盐性呈显著正相关(He et al., 2015).Badh1基因突变也与稻米香味相关, 但是效应远小于Badh2基因(Amarawathi et al., 2008).故水稻Badh1基因与香味之间的关系还需要进一步研究. ... ... 基因(Amarawathi et al., 2008).故水稻Badh1基因与香味之间的关系还需要进一步研究. ... ... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... 通过对水稻香味基因的测序结果进行比对分析, 大多数研究者认为, 由于Badh2基因的第2或第7外显子片段存在缺失, 导致2-AP的大量积累(Kovach et al., 2009)而产生香味.但实际上, 有些水稻品种的2-AP含量较高, 却没有Badh2的等位基因, 这表明可能还存在其它能够引起香味的非Badh2的等位基因(Amarawathi et al., 2008).多数研究者对香稻香味基因的研究仅注重香稻与非香稻外显子之间的差异, 但对真核生物而言, 除了基因的外显子, 内含子在基因的表达和调控中也发挥着重要作用(Fu et al., 1995; Jeon et al., 2000).徐小龙等(2011)通过对水稻香味基因Badh2进行测序, 结果显示香稻的编码链并无变异发生, 既不属于第7外显子突变类型, 也不属于第2外显子突变类型, 而在Badh2基因的第2和4内含子区域的核苷酸序列却发生了突变.张江丽等(2015)通过鉴定323份水稻种质资源, 发现具香味的水稻品种共86份, 其中有80份属于第7外显子变异的突变类型, 进一步对另6份香稻种质的Badh2基因进行克隆并测序分析, 结果显示这6份香稻品种的Badh2基因序列完整.因此, 水稻的香味性状还可能受到其它基因或内含子的调控. ...
2 2011
... 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种.例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011).同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013).高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ... ... ), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ...
2 2017
... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ... ... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
8 2005
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ... ... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 为进一步解析水稻香味基因的遗传特点, 对巴斯马蒂香稻的香味基因进行全基因组关联分析, 发现在香稻的基因组内, 除了第8号染色体上的Badh2基因, 还存在2个控制稻米香味的数量性状位点(quantita- tive trait loci, QTL) , 它们分别位于水稻的第3和4号染色体上(Amarawathi et al., 2008).其中第4号染色体上的数量性状位点可能与Badh1基因有关, 而Badh1与Badh2属于同源基因, 并且水稻的Badh1基因与高粱(Sorghum bicolor)和大麦(Hordeum vulgare)基因组中的Badh1基因也高度同源(Bradbury et al., 2005a, 2008).近期, 有研究表明, Badh1基因与水稻在种子萌发阶段的耐盐性呈显著正相关(He et al., 2015).Badh1基因突变也与稻米香味相关, 但是效应远小于Badh2基因(Amarawathi et al., 2008).故水稻Badh1基因与香味之间的关系还需要进一步研究. ... ... 针对2-AP生物合成途径及其调控网络, 不同的研究小组开展了大量的相关研究.在非香水稻中, 显性Badh2等位基因编码的甜菜碱醛脱氢酶的活性较高, 抑制2-AP的合成; 在香稻中, Badh2的2个隐性等位基因(Badh2-E2和Badh2-E7)不能产生有活性的甜菜碱醛脱氢酶, 因而可以诱导2-AP的形成(Chen et al., 2008).放射性同位素示踪实验结果表明, 脯氨酸是2-AP生物合成的前体物质之一, 它为2-AP提供氮源, 并且2-AP的形成与脯氨酸的积累呈显著正相关(Yoshihashi et al., 2002).也有研究结果显示, 先由脯氨酸和鸟氨酸间接合成2-AP的前体物质γ-氨基丁醛(γ-aminobutyraldehyde, GABald), 再经过中间体1-吡咯啉(Δ1-P)进而合成2-乙酰-1-吡咯啉(Schie- berle, 1990).甜菜碱醛脱氢酶不仅能够催化3-氨基丙醛(A-Pald)、二甲基硫丙醛(DMSPald)和甜菜醛(Betald), 还可以催化2-AP的前体物质γ-氨基丁醛的合成(Trossat et al., 1997; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ...
1 2008
... 为进一步解析水稻香味基因的遗传特点, 对巴斯马蒂香稻的香味基因进行全基因组关联分析, 发现在香稻的基因组内, 除了第8号染色体上的Badh2基因, 还存在2个控制稻米香味的数量性状位点(quantita- tive trait loci, QTL) , 它们分别位于水稻的第3和4号染色体上(Amarawathi et al., 2008).其中第4号染色体上的数量性状位点可能与Badh1基因有关, 而Badh1与Badh2属于同源基因, 并且水稻的Badh1基因与高粱(Sorghum bicolor)和大麦(Hordeum vulgare)基因组中的Badh1基因也高度同源(Bradbury et al., 2005a, 2008).近期, 有研究表明, Badh1基因与水稻在种子萌发阶段的耐盐性呈显著正相关(He et al., 2015).Badh1基因突变也与稻米香味相关, 但是效应远小于Badh2基因(Amarawathi et al., 2008).故水稻Badh1基因与香味之间的关系还需要进一步研究. ...
3 2005
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
3 2012
... 尽管稻米的香味由多种挥发物质组成, 但是目前普遍认为2-乙酰-1-吡咯啉(2-AP)是香稻芳香的主要化合物(Paule and Powers, 1989; Kovach et al., 2009), 并且2-AP具有较低的气味阈值, 易溶于乙醇和乙醚.市场上印度香米和泰国香米均属于籼型香稻品种, 2-AP为其提供独特的香味.前期大量的研究结果表明, 水稻第8号染色体上的Badh2基因突变能够使稻米产生香味, 而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ... ... 基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
2 2006
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ...
12 2008
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... 尽管稻米的香味由多种挥发物质组成, 但是目前普遍认为2-乙酰-1-吡咯啉(2-AP)是香稻芳香的主要化合物(Paule and Powers, 1989; Kovach et al., 2009), 并且2-AP具有较低的气味阈值, 易溶于乙醇和乙醚.市场上印度香米和泰国香米均属于籼型香稻品种, 2-AP为其提供独特的香味.前期大量的研究结果表明, 水稻第8号染色体上的Badh2基因突变能够使稻米产生香味, 而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... ), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 针对2-AP生物合成途径及其调控网络, 不同的研究小组开展了大量的相关研究.在非香水稻中, 显性Badh2等位基因编码的甜菜碱醛脱氢酶的活性较高, 抑制2-AP的合成; 在香稻中, Badh2的2个隐性等位基因(Badh2-E2和Badh2-E7)不能产生有活性的甜菜碱醛脱氢酶, 因而可以诱导2-AP的形成(Chen et al., 2008).放射性同位素示踪实验结果表明, 脯氨酸是2-AP生物合成的前体物质之一, 它为2-AP提供氮源, 并且2-AP的形成与脯氨酸的积累呈显著正相关(Yoshihashi et al., 2002).也有研究结果显示, 先由脯氨酸和鸟氨酸间接合成2-AP的前体物质γ-氨基丁醛(γ-aminobutyraldehyde, GABald), 再经过中间体1-吡咯啉(Δ1-P)进而合成2-乙酰-1-吡咯啉(Schie- berle, 1990).甜菜碱醛脱氢酶不仅能够催化3-氨基丙醛(A-Pald)、二甲基硫丙醛(DMSPald)和甜菜醛(Betald), 还可以催化2-AP的前体物质γ-氨基丁醛的合成(Trossat et al., 1997; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... ; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... ), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ...
1 2002
... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
1 2016
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
1 2001
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ...
1 2001
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ...
1 2008
... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ...
1 1995
... 通过对水稻香味基因的测序结果进行比对分析, 大多数研究者认为, 由于Badh2基因的第2或第7外显子片段存在缺失, 导致2-AP的大量积累(Kovach et al., 2009)而产生香味.但实际上, 有些水稻品种的2-AP含量较高, 却没有Badh2的等位基因, 这表明可能还存在其它能够引起香味的非Badh2的等位基因(Amarawathi et al., 2008).多数研究者对香稻香味基因的研究仅注重香稻与非香稻外显子之间的差异, 但对真核生物而言, 除了基因的外显子, 内含子在基因的表达和调控中也发挥着重要作用(Fu et al., 1995; Jeon et al., 2000).徐小龙等(2011)通过对水稻香味基因Badh2进行测序, 结果显示香稻的编码链并无变异发生, 既不属于第7外显子突变类型, 也不属于第2外显子突变类型, 而在Badh2基因的第2和4内含子区域的核苷酸序列却发生了突变.张江丽等(2015)通过鉴定323份水稻种质资源, 发现具香味的水稻品种共86份, 其中有80份属于第7外显子变异的突变类型, 进一步对另6份香稻种质的Badh2基因进行克隆并测序分析, 结果显示这6份香稻品种的Badh2基因序列完整.因此, 水稻的香味性状还可能受到其它基因或内含子的调控. ...
1 2000
... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
1 2014
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ...
1 2014
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ...
1 2014
... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
18 2015
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ... ... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 为进一步解析水稻香味基因的遗传特点, 对巴斯马蒂香稻的香味基因进行全基因组关联分析, 发现在香稻的基因组内, 除了第8号染色体上的Badh2基因, 还存在2个控制稻米香味的数量性状位点(quantita- tive trait loci, QTL) , 它们分别位于水稻的第3和4号染色体上(Amarawathi et al., 2008).其中第4号染色体上的数量性状位点可能与Badh1基因有关, 而Badh1与Badh2属于同源基因, 并且水稻的Badh1基因与高粱(Sorghum bicolor)和大麦(Hordeum vulgare)基因组中的Badh1基因也高度同源(Bradbury et al., 2005a, 2008).近期, 有研究表明, Badh1基因与水稻在种子萌发阶段的耐盐性呈显著正相关(He et al., 2015).Badh1基因突变也与稻米香味相关, 但是效应远小于Badh2基因(Amarawathi et al., 2008).故水稻Badh1基因与香味之间的关系还需要进一步研究. ... ... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... ; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... 尽管稻米的香味由多种挥发物质组成, 但是目前普遍认为2-乙酰-1-吡咯啉(2-AP)是香稻芳香的主要化合物(Paule and Powers, 1989; Kovach et al., 2009), 并且2-AP具有较低的气味阈值, 易溶于乙醇和乙醚.市场上印度香米和泰国香米均属于籼型香稻品种, 2-AP为其提供独特的香味.前期大量的研究结果表明, 水稻第8号染色体上的Badh2基因突变能够使稻米产生香味, 而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... , He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... 遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... ; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... ; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... 水稻香味基因Badh2功能标记的开发(He and Park, 2015) ... ... Development of functional markers for Badh2 gene in rice (He and Park, 2015) ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ... ... 香味是水稻重要的品质性状之一, 优质香米的消费需求将持续增加.因此, 香稻新品种选育以及香味基因研究受到越来越多水稻遗传育种学家和植物科学家的重视.随着水稻功能基因组研究的快速发展, 关于水稻中香味基因的遗传基础、功能及其调控规律等将会越来越明朗.最近十几年, 基因测序技术突飞猛进, 大量不同来源的水稻品种和香稻种质资源完成测序分析, 发现了一系列香味基因的变异位点及其等位基因(He and Park, 2015, He et al., 2015).针对这些变异位点, 香味基因功能标记也被不断开发并已经广泛应用于香味基因筛选和杂交香稻骨干亲本培育.尽管香味基因功能标记已经用于香稻恢复系的选育(杜雪树等, 2009), 而且很多香稻品种已通过审定并在生产中得到较大面积的推广应用, 但是目前香稻遗传育种面临的主要问题依然是香型优良的恢复系极少, 减缓了香稻的遗传育种进程. ... ... , He et al., 2015).针对这些变异位点, 香味基因功能标记也被不断开发并已经广泛应用于香味基因筛选和杂交香稻骨干亲本培育.尽管香味基因功能标记已经用于香稻恢复系的选育(杜雪树等, 2009), 而且很多香稻品种已通过审定并在生产中得到较大面积的推广应用, 但是目前香稻遗传育种面临的主要问题依然是香型优良的恢复系极少, 减缓了香稻的遗传育种进程. ... ... 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种.例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011).同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013).高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ...
2015
1 2000
... 通过对水稻香味基因的测序结果进行比对分析, 大多数研究者认为, 由于Badh2基因的第2或第7外显子片段存在缺失, 导致2-AP的大量积累(Kovach et al., 2009)而产生香味.但实际上, 有些水稻品种的2-AP含量较高, 却没有Badh2的等位基因, 这表明可能还存在其它能够引起香味的非Badh2的等位基因(Amarawathi et al., 2008).多数研究者对香稻香味基因的研究仅注重香稻与非香稻外显子之间的差异, 但对真核生物而言, 除了基因的外显子, 内含子在基因的表达和调控中也发挥着重要作用(Fu et al., 1995; Jeon et al., 2000).徐小龙等(2011)通过对水稻香味基因Badh2进行测序, 结果显示香稻的编码链并无变异发生, 既不属于第7外显子突变类型, 也不属于第2外显子突变类型, 而在Badh2基因的第2和4内含子区域的核苷酸序列却发生了突变.张江丽等(2015)通过鉴定323份水稻种质资源, 发现具香味的水稻品种共86份, 其中有80份属于第7外显子变异的突变类型, 进一步对另6份香稻种质的Badh2基因进行克隆并测序分析, 结果显示这6份香稻品种的Badh2基因序列完整.因此, 水稻的香味性状还可能受到其它基因或内含子的调控. ...
1 1995
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ...
1 1996
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ...
1 2003
... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
1 2011
... 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种.例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011).同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013).高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ...
1 2012
... 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种.例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011).同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013).高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ...
12 2009
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... ; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... ; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... ).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... 尽管稻米的香味由多种挥发物质组成, 但是目前普遍认为2-乙酰-1-吡咯啉(2-AP)是香稻芳香的主要化合物(Paule and Powers, 1989; Kovach et al., 2009), 并且2-AP具有较低的气味阈值, 易溶于乙醇和乙醚.市场上印度香米和泰国香米均属于籼型香稻品种, 2-AP为其提供独特的香味.前期大量的研究结果表明, 水稻第8号染色体上的Badh2基因突变能够使稻米产生香味, 而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 通过对水稻香味基因的测序结果进行比对分析, 大多数研究者认为, 由于Badh2基因的第2或第7外显子片段存在缺失, 导致2-AP的大量积累(Kovach et al., 2009)而产生香味.但实际上, 有些水稻品种的2-AP含量较高, 却没有Badh2的等位基因, 这表明可能还存在其它能够引起香味的非Badh2的等位基因(Amarawathi et al., 2008).多数研究者对香稻香味基因的研究仅注重香稻与非香稻外显子之间的差异, 但对真核生物而言, 除了基因的外显子, 内含子在基因的表达和调控中也发挥着重要作用(Fu et al., 1995; Jeon et al., 2000).徐小龙等(2011)通过对水稻香味基因Badh2进行测序, 结果显示香稻的编码链并无变异发生, 既不属于第7外显子突变类型, 也不属于第2外显子突变类型, 而在Badh2基因的第2和4内含子区域的核苷酸序列却发生了突变.张江丽等(2015)通过鉴定323份水稻种质资源, 发现具香味的水稻品种共86份, 其中有80份属于第7外显子变异的突变类型, 进一步对另6份香稻种质的Badh2基因进行克隆并测序分析, 结果显示这6份香稻品种的Badh2基因序列完整.因此, 水稻的香味性状还可能受到其它基因或内含子的调控. ... ... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... ; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ...
1 2015
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
1 2012
... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
1 2014
... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
2 1996
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ...
2 2014
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... ; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ...
1 2014
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ...
1 1982
... 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种.例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011).同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013).高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ...
1 2012
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
3 2008
... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ...
4 2014
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... ; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ...
1 1989
... 尽管稻米的香味由多种挥发物质组成, 但是目前普遍认为2-乙酰-1-吡咯啉(2-AP)是香稻芳香的主要化合物(Paule and Powers, 1989; Kovach et al., 2009), 并且2-AP具有较低的气味阈值, 易溶于乙醇和乙醚.市场上印度香米和泰国香米均属于籼型香稻品种, 2-AP为其提供独特的香味.前期大量的研究结果表明, 水稻第8号染色体上的Badh2基因突变能够使稻米产生香味, 而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ...
1 2014
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
1 2014
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
2 2009
... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... ; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ...
1 1990
... 针对2-AP生物合成途径及其调控网络, 不同的研究小组开展了大量的相关研究.在非香水稻中, 显性Badh2等位基因编码的甜菜碱醛脱氢酶的活性较高, 抑制2-AP的合成; 在香稻中, Badh2的2个隐性等位基因(Badh2-E2和Badh2-E7)不能产生有活性的甜菜碱醛脱氢酶, 因而可以诱导2-AP的形成(Chen et al., 2008).放射性同位素示踪实验结果表明, 脯氨酸是2-AP生物合成的前体物质之一, 它为2-AP提供氮源, 并且2-AP的形成与脯氨酸的积累呈显著正相关(Yoshihashi et al., 2002).也有研究结果显示, 先由脯氨酸和鸟氨酸间接合成2-AP的前体物质γ-氨基丁醛(γ-aminobutyraldehyde, GABald), 再经过中间体1-吡咯啉(Δ1-P)进而合成2-乙酰-1-吡咯啉(Schie- berle, 1990).甜菜碱醛脱氢酶不仅能够催化3-氨基丙醛(A-Pald)、二甲基硫丙醛(DMSPald)和甜菜醛(Betald), 还可以催化2-AP的前体物质γ-氨基丁醛的合成(Trossat et al., 1997; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ...
4 2013
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ... ... 水稻中有200多种挥发性物质(Mahattanatawee and Rouseff, 2014; He and Park, 2015), 其中2-乙酰-1-吡咯啉(2-acetyl-1-pyrroline, 2-AP)是香稻中主要的挥发性物质之一(Mathure et al., 2014; Mahattanatawee and Rouseff, 2014).目前, 已建立多种方法用于检测水稻材料中的香味, 其中咀嚼法和KOH法是传统育种过程中最为常用的方法(Bradbury et al., 2005b; 刘化龙等, 2014; 闫影等, 2015), 但是这2种方法主要依靠人的感官来进行判定香味, 准确性较差, 且难以大规模应用于香稻新品种的筛选与培育.因此, 如何简单、准确而又快速地鉴定香稻中的香味一直是水稻育种工作者亟须解决的难题之一(闫影等, 2015).香稻中香味的遗传基础比较复杂, 但是多数研究者认为香味是由单个隐性基因控制的.位于水稻第8号染色体上的隐性基因fgr是1个与香味密切相关的基因, 目前该基因已被分离克隆(Bradbury et al., 2005a; Chen et al., 2006, 2008).进一步研究显示, 基因fgr编码甜菜碱醛脱氢酶(Betaine aldehyde dehydrogenase homologue 2, Badh2).抑制fgr基因的表达或者敲除fgr基因, 均会使Badh2酶功能缺失, 造成2-AP前体物质增加, 进而积累2-AP使水稻产生香味(Niu et al., 2008; Shan et al., 2013).尽管有关水稻香味基因的生化代谢途径研究取得了重要进展, 但是关于香味基因的等位基因变异、控制香味的基因数目以及香味基因在水稻新品种培育中的应用等方面还需要深入研究.本文综述了近年来水稻香味基因研究取得的新进展, 以及香味基因功能标记的开发与利用, 以期为优质香稻新品种的培育提供借鉴与参考. ... ... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ... ... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
13 2015
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ... ... ).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... ; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 尽管稻米的香味由多种挥发物质组成, 但是目前普遍认为2-乙酰-1-吡咯啉(2-AP)是香稻芳香的主要化合物(Paule and Powers, 1989; Kovach et al., 2009), 并且2-AP具有较低的气味阈值, 易溶于乙醇和乙醚.市场上印度香米和泰国香米均属于籼型香稻品种, 2-AP为其提供独特的香味.前期大量的研究结果表明, 水稻第8号染色体上的Badh2基因突变能够使稻米产生香味, 而非香水稻中Badh2基因编码的Badh2蛋白具有甜菜碱脱氢酶的活性, 突变类型中甜菜碱脱氢酶活性丧失, 2-AP的积累使稻米产生香味(Kovach et al., 2009; He and Park, 2015; Shan et al., 2015).进一步研究发现, Badh2基因存在多个转录起始位点.在非香水稻中, 尽管Badh2基因具有3个不同长度的转录本, 但是只有全长为1 509 bp的Badh2基因转录水平较高, 进而显著影响2-AP的含量, 其余的等位基因表达水平较低(Chen et al., 2008).在香稻品种中, 主要的转录起始位点位于Badh2基因起始密码子的下游, 并且Badh2基因在香稻品种的转录被严重抑制(Chen et al., 2008).Badh2基因除了在水稻的根中不表达, 在水稻其余组织和器官中均有表达, 并且该基因编码的蛋白具有3个结构域, 定位于细胞质中(Chen et al., 2008).采用RNAi抑制或者互补遗传转化实验已经证实, Badh2基因功能的丧失或者基因表达量下调, 都会使稻米产生香味(Chen et al., 2008, 2012), 其显性等位基因则抑制2-AP在水稻体内的合成(Chen et al., 2008), 而2-AP正是从香稻米饭中挥发出来的1种芳香性的化学物质.目前为止, 虽然已经证实Badh2基因功能的丧失可以导致米香物质2-AP的产生, 但是Badh2蛋白如何调控2-AP的合成过程以及2-AP生物合成途径及其相关的调控网络和调控机制等尚不清楚. ... ... 针对2-AP生物合成途径及其调控网络, 不同的研究小组开展了大量的相关研究.在非香水稻中, 显性Badh2等位基因编码的甜菜碱醛脱氢酶的活性较高, 抑制2-AP的合成; 在香稻中, Badh2的2个隐性等位基因(Badh2-E2和Badh2-E7)不能产生有活性的甜菜碱醛脱氢酶, 因而可以诱导2-AP的形成(Chen et al., 2008).放射性同位素示踪实验结果表明, 脯氨酸是2-AP生物合成的前体物质之一, 它为2-AP提供氮源, 并且2-AP的形成与脯氨酸的积累呈显著正相关(Yoshihashi et al., 2002).也有研究结果显示, 先由脯氨酸和鸟氨酸间接合成2-AP的前体物质γ-氨基丁醛(γ-aminobutyraldehyde, GABald), 再经过中间体1-吡咯啉(Δ1-P)进而合成2-乙酰-1-吡咯啉(Schie- berle, 1990).甜菜碱醛脱氢酶不仅能够催化3-氨基丙醛(A-Pald)、二甲基硫丙醛(DMSPald)和甜菜醛(Betald), 还可以催化2-AP的前体物质γ-氨基丁醛的合成(Trossat et al., 1997; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... ).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... ).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... 目前, 主要的水稻育种技术仍然是常规杂交育种, 然而, 利用传统育种方法来培育香稻新品种费时费力 (Shan et al., 2015).由于香味基因主要由隐性基因控制, 传统育种先要通过杂交和回交的方法, 将Badh2突变的基因导入现有的优良品种中, 然后在后代群体中进行筛选, 可是这一过程在苗期很难快速鉴定.然而, 我国水稻育种学家在优质、高产及多抗香稻新品种的培育方面还是做了大量研究, 并筛选到一系列香稻的恢复系、不育系以及新的杂交组合(黄庭旭等, 2006; 况浩池等, 2007; 刘光春等, 2008; 江青山等, 2008).随着分子标记尤其是Badh2基因功能标记的开发, 分子标记辅助选择已经广泛应用于水稻遗传育种, 大大加快了香稻新品种的筛选与培育进程, 并且许多香稻品种已通过审定, 在生产中得到大面积的推广应用. ... ... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ... ... 基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ... ... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ... ... ), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
3 2011
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ...
6 2013
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... ; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... ).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ... ... 自从Badh2基因被分离克隆以来, 已经在该基因内发现了十几个变异位点(Kovach et al., 2009; Shao et al., 2013; He and Park, 2015, He et al., 2015), 并针对这些位点设计了一系列分子标记, 用于香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育.Bradbury等(2005b)利用Badh2基因在第7外显子有8 bp的缺失和3个单核苷酸多态性变异位点, 设计引物进行PCR扩增来鉴定Badh2等位基因的类型, 随后针对该位点设计了1个新的分子标记YY5-YY8, 用于稻米香味的快速检测分析(闫影等, 2015).Shi等(2008)对24个香稻品种和10个非香水稻品种进行测序, 发现有12个香稻品种在Badh2基因的第7外显子处有1个8 bp的缺失和3个单核苷酸多态性突变位点.同时他们还发现新的没有生物学功能的Badh2等位基因, 该等位基因在第7外显子处存在相同的变异, 但是在第2外显子处还发现1个7 bp缺失位点.针对这2个外显子的变异区域, Shi等(2008)设计了多对分子标记进行PCR检测分析.随后, 在Badh2基因的第7外显子和第2外显子处突变的检测中, 研究者简化了引物设计, 只需要采用1对引物即可进行水稻香味基因分子标记检测(王丰等, 2008; 王军等, 2008).Shao等(2011)通过基因测序和序列分析, 发现在Badh2基因的第4和5外显子之间可能存在803 bp的缺失, 并开发了1个标记FMbadh2-E4-5用于检测这个缺失的新位点.在研究香稻品种中发现, Badh2基因的第13外显子处存在1个单核苷酸突变, 导致水稻产生香味, 并且该突变位点正好位于内含子和外显子的剪接位点处, 由此设计了1个新的分子标记(Ootsuka et al., 2014).根据295个不同水稻品种的测序分析显示, 有部分香稻品种在Badh2基因的第12外显子处有1个3 bp的缺失, 在第13外显子处有1个3 bp的插入, 由此设计功能性分子标记FME12-3, 并利用2个F2遗传群体对功能标记FME12-3进行了验证(He and Park, 2015).上述一系列分子标记的开发, 为今后水稻香味基因的鉴定、不同香稻品种的筛选以及香稻新品种的培育带来了极大的便利, 加快了香稻遗传育种和推广应用进程. ...
7 2008
... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... ), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ... ... 目前, 在Badh2基因中至少存在17个变异位点, 这些位点分布于Badh2基因的5'UTR区、第1外显子、第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间以及第7、8、10、12、13和14外显子处(He and Park, 2015).针对这些变异位点, 已经在Badh2基因的第1外显子与第1内含子连接处、第2外显子、第4外显子与第5外显子之间、第7、12、13和14外显子处设计了多对分子标记, 用来鉴定Ba- dh2基因的突变类型(Bradbury et al., 2005a, 2005b; Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011; Ootsuka et al., 2014; He and Park, 2015).在Badh2基因的第1、8和10外显子处也存在多个变异位点, 但是它们与水稻的香味之间并没有直接的关联, 表明这些变异位点大多是同义突变, 或者突变的发生虽然改变了Badh2蛋白的序列, 但并不影响Badh2蛋白的功能.在上述分子标记的基础之上, 研究人员进一步开发了6个水稻香味基因的功能标记(表1), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... ), 并利用分离群体进行了验证(Shi et al., 2008; Kovach et al., 2009; Ootsuka et al., 2014; He and Park, 2015).其中, 功能标记FMU1-2不但可以检测Badh2基因在第7外显子处的8 bp的缺失突变, 而且能够检测Badh2基因的5'UTR区是否存在1个8 bp的缺失; 功能标记FME2-7、FME7、FME12-3、FME13和FME14分别用于鉴定香味基因在Badh2的第2、7、12、13和14外显子处的变异类型; 功能标记FME14属于CAPS (the cleaved amplified polymorphic sequence)标记, 故对其PCR产物的检测需要借助限制性内切酶Bsl I.因此, 这些水稻香味基因功能标记的开发, 为香稻分子育种提供了一种准确、快速且有效的手段. ... ... 早期研究发现, 利用与frg基因紧密连锁的分子标记(如RG28、SCU015RM和RSP04)能够区分香稻和非香水稻(Garland et al., 2000; Cordeiro et al., 2002; Jin et al., 2003).随着Badh2基因的分离克隆和功能的深入研究, 研究人员针对Badh2基因的内部设计了一系列特异引物(即功能性标记)用于香味基因的筛选(Bradbury et al., 2005b; Shi et al., 2008; 王丰等, 2008; He and Park, 2015; 闫影等, 2015; 许言福等, 2015)以及香稻恢复系的选育(杜雪树等, 2009).此外, 转基因技术也在香稻新品种的培育过程中应用并推广(Niu et al., 2008; Chen et al., 2012).利用RNAi介导的Badh2基因沉默, 也可以使非香水稻转变为香稻品种(Chen et al., 2012), 但是这种技术往往不能够完全抑制Badh2基因的表达, 并且需要在转基因后代中进行大量筛选.同时, 利用RNAi介导的转基因技术培育香稻新品种, 还面临着后期转基因风险评估与严格的监管, 在新品种推广与应用等方面还面临着诸多问题. ... ... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ...
2 2014
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ... ... 香稻由于其稻米具有独特的香味, 在国内外市场上受到广大消费者的青睐(Kovach et al., 2009; Shan et al., 2015).水稻香味基因的遗传基础研究也取得了重要进展.Ahn等(1992)发现控制香味的隐性基因位于水稻的第8号染色体上, 与分子标记RG28的遗传距离为4.5 cM.随后, 有多个研究小组利用不同的遗传群体和不同类型的分子标记在水稻第8号染色体的RG28附近区域检测到1个控制水稻香味的隐性基因(Jin et al., 1995, 1996; Lorieux et al., 1996; Dong et al., 2001a, 2001b).张涛等(2008)以粳型香稻和籼型非香水稻为实验材料, 通过精细定位将香味基因定位在水稻第8号染色体的20 175 367-20 386 172 bp之间大约252 kb的区域内.Wanchana等(2005)借助分子标记将控制香味的基因fgr定位在RM223-RM342之间.进一步研究发现, 香稻和非香水稻之间的差异在这2个分子标记之间的1个编码甜菜碱脱氢酶的基因(Badh2)上.通过图位克隆, 对fgr区域进行测序, 发现香稻和非香水稻在Badh2基因的序列上存在显著差异.香稻在Badh2基因的第7外显子区存在突变, 导致Badh2蛋白功能的丧失.因此, Badh2基因很有可能就是控制水稻香味的fgr基因(Bradbury et al., 2005a).为了验证Badh2基因的功能, 将fgr区域内的3个候选基因同时进行互补转化实验, 结果表明, Cah和Mccc2两个候选基因在转基因前后其稻米中的香味并未发生显著改变, 但是在Badh2基因的转基因单株中, 其香味发生明显变化; 同时, 利用RNAi技术抑制Badh2基因的表达, 也可以使非香水稻产生香味.以上结果证实了Badh2基因就是控制稻米中香味的隐性基因(Chen et al., 2006, 2008, 2012; Niu et al., 2008).Badh2基因全长1 509 bp, 包含15个外显子和14个内含子, 编码503个氨基酸(Chen et al., 2008; Shan et al., 2015).对香稻和非香水稻品种进行比较测序, 发现Badh2基因在不同的水稻品种中存在多种类型的突变.例如, 在香稻品种中, 常见的是Badh2基因的第7外显子有1个8 bp的缺失和3个单核苷酸多态性位点(Bradbury et al., 2005a); 在不同的香稻品种中, Badh2基因还在第2外显子有1个7 bp的缺失, 第4和第5外显子之间存在1个803 bp的缺失(Shi et al., 2008; Kovach et al., 2009; Shao et al., 2011), 并且在第1、10、13和14外显子均存在变异位点(Shi et al., 2008; Amarawathi et al., 2008; Kovach et al., 2009).进一步研究发现, 在Badh2基因的第1外显子和第1内含子的剪接位点、启动子区域及其5'UTR区也有插入、缺失或者单个核苷酸突变位点(Shao et al., 2011, 2013; Shi et al., 2014; Ootsuka et al., 2014).水稻的香味差异很大程度上是由Badh2基因的等位变异决定的, 与香味有关的水稻品种常常包括其第7外显子8 bp的缺失和3个单核苷酸多态性(SNPs), 或者第2外显子7 bp的缺失(Shi et al., 2008; Shao et al., 2013).因此, 当Badh2基因在编码区或者调控区域发生突变时, 能够产生无生物学活性的甜菜碱脱氢酶, 导致水稻产生香味. ...
1 2013
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ...
1 2009
... 水稻(Oryza sativa)是世界上最主要的粮食作物之一, 为全世界30多亿人口提供大约25%的能量(Tian et al., 2009; Peng et al., 2014a; 黎舒佳等, 2015).对东南亚地区的大部分居民而言, 稻米为其提供的能量达35%以上(Peng et al., 2014b; Kusano et al., 2015).作为栽培水稻类型之一的香稻, 其香味是极其重要的食味品质性状之一(唐傲等, 2009).香稻的稻米由于具有香气馥郁和米饭芬芳等特点, 其被视为水稻中的珍品(Bradbury et al., 2005a; 赵志鹏等, 2009; Myint et al., 2012; Daygon et al., 2016).传统的香稻品种地域性强、抗性较差且产量偏低, 难以大面积推广种植(白现广等, 2009; 杜雪树等, 2009), 故香米的市场价格较高, 但是人们对香米的需求近年来却显著增加(Shao et al., 2013; Shi et al., 2014; 彭波等, 2016a).因此, 香味基因研究及其在遗传育种中的应用已受到水稻遗传学家和育种学家的广泛关注. ...
2 1997
... 针对2-AP生物合成途径及其调控网络, 不同的研究小组开展了大量的相关研究.在非香水稻中, 显性Badh2等位基因编码的甜菜碱醛脱氢酶的活性较高, 抑制2-AP的合成; 在香稻中, Badh2的2个隐性等位基因(Badh2-E2和Badh2-E7)不能产生有活性的甜菜碱醛脱氢酶, 因而可以诱导2-AP的形成(Chen et al., 2008).放射性同位素示踪实验结果表明, 脯氨酸是2-AP生物合成的前体物质之一, 它为2-AP提供氮源, 并且2-AP的形成与脯氨酸的积累呈显著正相关(Yoshihashi et al., 2002).也有研究结果显示, 先由脯氨酸和鸟氨酸间接合成2-AP的前体物质γ-氨基丁醛(γ-aminobutyraldehyde, GABald), 再经过中间体1-吡咯啉(Δ1-P)进而合成2-乙酰-1-吡咯啉(Schie- berle, 1990).甜菜碱醛脱氢酶不仅能够催化3-氨基丙醛(A-Pald)、二甲基硫丙醛(DMSPald)和甜菜醛(Betald), 还可以催化2-AP的前体物质γ-氨基丁醛的合成(Trossat et al., 1997; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ... ... ); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ...
1 1990
... 在不同的水稻品种中, 并不是所有的香稻品种在Badh2基因的第7外显子都存在1个8 bp的缺失和3个单核苷酸多态性位点; 在非香水稻品种中也发现Badh2基因的第7外显子发生同样的突变(Fitzgerald et al., 2008).对来源于亚洲38个国家和地区的280份普通野生稻及242份水稻品种分析表明, Badh2基因存在8种无功能活性的单倍型.Badh2基因来源于类似粳稻遗传背景的水稻材料, 现有的籼型香稻中的香味基因是从粳型香稻中导入, 并不断经过人工选择(Kovach et al., 2009).针对普通野生稻的研究发现, 在亚洲普通野生稻中也存在Badh2基因第7外显子有缺失的类型, 而这些缺失或者突变类型的Badh2基因, 在野生稻中较少出现(Prathepha, 2009), 它们不能进行正常的翻译, 进而抑制水稻产生香味(Chen et al., 2008).前期研究表明, 现有栽培水稻品种中的香味基因最早起源于普通野生稻, 并经历了长期的人工选择与驯化过程(Kovach et al., 2009; Prathepha, 2009; Shao et al., 2013; He et al., 2015).在人工选择与驯化过程中, 香味基因很可能产生了多种变异, 导致不同的研究小组得出的结论存在较大差异.多数研究者认为香味遗传受单个隐性基因的控制(Shao et al., 2013; He et al., 2015); 也有研究者认为香稻香味的遗传是由多个隐性基因决定(Tsuzuki and Shimokawa, 1990); 还有****认为香味遗传受主效基因+微效基因控制(徐辰武和莫惠栋, 1995), 并存在多个QTLs控制稻米的香味(Lorieux et al., 1996; Amara- wathi et al., 2008).总之, 水稻香味基因的遗传基础比较复杂, 控制水稻香味的基因, 少则1对, 多则2-4对或更多, 其原因可能包括以下3点: (1) 不同香味类型(如爆米花型、茉莉花型、紫罗兰型和山核桃香型等)的香稻品种的遗传基础有所差异; (2) 香味基因与多种环境因素(光照、温度和土壤肥力等)之间存在相互作用; (3) 水稻香味成分具多样性, 但目前针对不同类型香味的鉴定方法和技术还不够完善. ...
1 2014
... 近年来, 基因组编辑技术(genome editing tech- nology)发展迅速, 包括转录激活样效应因子核酸酶(transcription activator-like effector nuclease, TAL- EN)技术、锌指核酸酶(Zinc-finger nuclease, ZFN)技术和成簇规律间隔短回文重复(clustered regularly interspaced short palindromic repeat, CRISPR)技术, 它们都已经应用于作物的遗传育种中(Voytas and Gao, 2014).在水稻中, 任何插入、缺失或者替换Badh2基因, 都将导致过早出现终止密码子或者编码的氨基酸改变甚至不编码对应的Badh2蛋白, 进而使非香水稻产生香味(Bradbury et al., 2005a; Shi et al., 2008; Kovach et al., 2009), 即丧失功能的Ba- dh2基因可以促进2-AP的合成与积累, 那么可以想象, 任何使Badh2基因功能丧失的突变, 都将导致1个新的香味基因出现.于是, 利用TALEN技术将Badh2基因敲除, 可以将1个非香水稻品种转变为香稻品种(Shan et al., 2013); 并且同样采用TALEN技术可以创建遗传纯合的突变型香稻单株(Shan et al., 2015).因此, 利用TALEN技术可以将非香水稻中的Badh2基因敲除(Shan et al., 2015; Birla et al., 2017), 快速产生对应的香稻, 大大加快优质、高产香稻新品种的培育进程. ...
2005
1 2014
... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
1 2013
... 随着基因组编辑技术的迅速发展, 序列特异性核酸酶(sequence-specific nucleases, SSNs)开始广泛用于多种植物的基因敲除, 包括重要的作物如水稻、玉米(Zea mays)、小麦(Triticum aestivum)、大麦和大豆(Li et al., 2012; Wendt et al., 2013; Haun et al., 2014; Liang et al., 2014; Wang et al., 2014).此外, TALEN技术也被应用于香稻的遗传育种(Shan et al., 2013, 2015; Birla et al., 2017), 这些新技术为水稻育种学家进行香稻品种的培育提供了一种全新的策略.利用SSNs (包括TALEN、ZFN和CRISPR)技术进行分子设计育种, 要比传统育种和转基因(RNAi或者基因工程)育种具有明显的优势: (1) 准确编辑目标基因; (2) 不需要杂交和回交过程, 省时方便; (3) 能够得到无筛选标记的单株(Shan et al., 2015).因此, 今后将会有越来越多的新技术, 如TALEN技术、ZFN技术和CRISPR技术应用于香稻的遗传育种, 进而加快优质、高产且多抗香稻新品种的培育. ...
1 2002
... 针对2-AP生物合成途径及其调控网络, 不同的研究小组开展了大量的相关研究.在非香水稻中, 显性Badh2等位基因编码的甜菜碱醛脱氢酶的活性较高, 抑制2-AP的合成; 在香稻中, Badh2的2个隐性等位基因(Badh2-E2和Badh2-E7)不能产生有活性的甜菜碱醛脱氢酶, 因而可以诱导2-AP的形成(Chen et al., 2008).放射性同位素示踪实验结果表明, 脯氨酸是2-AP生物合成的前体物质之一, 它为2-AP提供氮源, 并且2-AP的形成与脯氨酸的积累呈显著正相关(Yoshihashi et al., 2002).也有研究结果显示, 先由脯氨酸和鸟氨酸间接合成2-AP的前体物质γ-氨基丁醛(γ-aminobutyraldehyde, GABald), 再经过中间体1-吡咯啉(Δ1-P)进而合成2-乙酰-1-吡咯啉(Schie- berle, 1990).甜菜碱醛脱氢酶不仅能够催化3-氨基丙醛(A-Pald)、二甲基硫丙醛(DMSPald)和甜菜醛(Betald), 还可以催化2-AP的前体物质γ-氨基丁醛的合成(Trossat et al., 1997; Chen et al., 2008; Shan et al., 2015).在水稻中, Badh2蛋白能够将γ-氨基丁醛催化为γ-氨基丁酸(γ-aminobutyric acid, GABA) (Shan et al., 2015), 并且在非香水稻品种中GABA的含量要远高于香稻.针对甜菜碱醛脱氢酶的生物学功能, 不同的研究小组得出的研究结果存在较大的差异.有研究认为甜菜碱醛脱氢酶能够抑制2-乙酰-1-吡咯啉的合成(Chen et al., 2008); 也有研究结果显示, 甜菜碱醛脱氢酶参与2-乙酰-1-吡咯啉的分解(Trossat et al., 1997); 还有研究者认为, 甜菜碱醛脱氢酶与2-乙酰-1-吡咯啉合成的前体物质之间存在竞争性关系(Bradbury et al., 2005a).甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的生物合成过程中发挥重要作用, 但是甜菜碱醛脱氢酶在2-乙酰-1-吡咯啉的合成过程中是如何发挥作用的及是否还有其它物质参与调控等尚不清楚, 还有待深入研究.同时, 作为2-乙酰-1-吡咯啉合成的前体物质γ-氨基丁醛, 在2-乙酰-1-吡咯啉的生物合成过程中也可能起关键调控作用.香稻中Badh2蛋白功能的丧失可能导致γ-氨基丁醛在体内积累, 进而使γ-氨基丁醛转化为1-吡咯啉, 最终合成大量的2-乙酰-1-吡咯啉(Shan et al., 2015); 在非香水稻中, Badh2蛋白具有甜菜碱醛脱氢酶的催化活性, 可能将水稻体内的γ-氨基丁醛转化为γ-氨基丁酸, 同时抑制2-AP前体物质1-吡咯啉的合成, 最终导致不能合成2-AP (图1).水稻中, 鸟氨酸和脯氨酸都可以作为合成γ-氨基丁醛的前体物质, 这一合成过程的调控网络以及它们与甜菜碱醛脱氢酶之间的调控关系, 目前尚不清楚.鸟氨酸、脯氨酸、γ-氨基丁醛与1-吡咯啉之间复杂的调控关系仍需进一步研究. ...
1 2013
... 目前, 不仅在水稻中利用香味基因培育香稻新品种, 也在其它作物中发掘对应的香味基因并应用于作物的遗传育种.例如, 在大豆(Glycine max)中, 位于第6号染色体上GmBADH2基因第10外显子处发生2 bp的缺失, 将导致该基因提前终止翻译的过程, 进而产生香味(Juwattanasomran et al., 2011, 2012); 如果GmBADH2基因发生沉默, 将会促使2-AP在大豆中大量合成, 芳香型大豆随即产生(Arikit et al., 2011).同时, 在高粱中的研究发现, SbBADH2基因是控制高粱香味的隐性基因(Yundaeng et al., 2013).高粱、水稻和大豆的驯化历史并不相同, 这3种作物中的香味主要是由单个隐性基因控制的(Murty et al., 1982; He and Park, 2015), 其中香味基因发生突变与这些作物香味的产生关系密切, 并且3种作物中控制香味的隐性基因具有一定的同源性(Arikit et al., 2011).香味基因在水稻、高粱和大豆中功能的丧失不仅不影响其生存, 在干旱的条件下还会发挥积极作用. ...
1 2013
... 20世纪后半叶水稻的产量得到显著提高, 但是从20世纪80年代至今, 水稻产量一直徘徊不前.目前, 环境污染、水资源短缺、极端天气频发等因素不但影响水稻的生长和最终产量, 而且严重影响稻米品质(Stocker et al., 2013; Zhao and Fitzgerald, 2013; Goufo et al., 2014; Halford et al., 2014).随着世界人口的持续增长和生活水平的稳步提高, 传统的水稻育种已不能满足人们对优质高产水稻的强烈需求(彭波等, 2016b).寻找新的育种策略和方法(如分子标记辅助选择育种、全基因组育种、分子设计育种和转基因育种)将会弥补水稻传统育种的不足, 实现水稻产量突破和稻米品质的同步提高.经过十几年的努力, 水稻香味基因从初步定位到分离克隆以及功能分析, 再到现在应用于香稻的遗传育种, 已经取得了巨大进展(Shan et al., 2013, 2015; He and Park, 2015; 闫影等, 2015).同时, 新的技术和育种策略已经开始应用于香稻育种工作之中(Shan et al., 2015).明确香味基因的分子遗传机理, 采用分子标记辅助选择育种等新的育种策略和方法, 将会为香稻新品种培育提供重要参考. ...
 ,
, 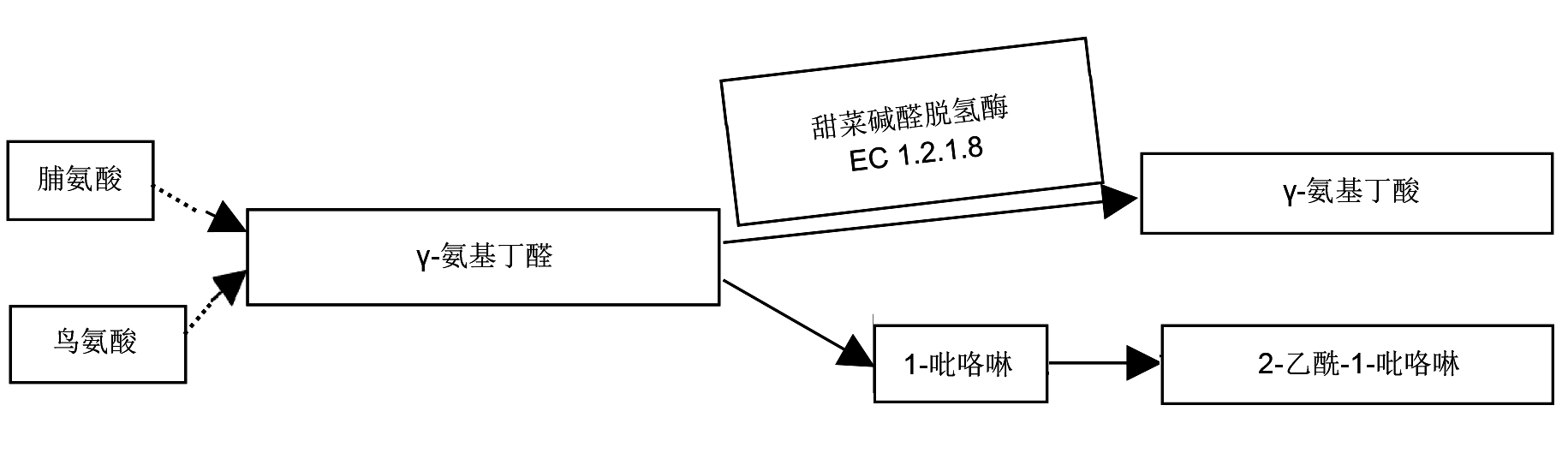

{kind=link}
{kind=link}