Abstract: Plant survival and reproduction are closely related to light signal transduction. Both light intensity and light quality regulate plant growth and development. In natural or artificial ecosystems, the light received by an individual plant can be modified by its neighbors, and the photosynthetically active radiation and ratio of red light to far red light (R:FR) will significantly decrease, called shade. Shade stress has many effects during distinct stages of plant growth and development, including seed germination inhibition, seedling hypocotyl elongation and early flowering. Subsequently, shade decreases crop yield and quality; thus, it is unfavorable for agricultural production. Phytohormones are key endogenous factors in regulating plant growth and development. Numerous studies demonstrated that auxin, gibberellin and brassinosteroids are involved in the shade stress response. Under shade conditions, light signaling will be perceived and integrated by the plant, and the integrated signal can affect the biosynthesis and signal transduction of phytohormones. Different phytohormones have distinct pathways in response to shade stress. The crosstalk among phytohormone signaling pathways under shade stress are still unclear. In this review, we summarize the mechanisms of response of the main phytohormones (auxin, gibberellin, brassinosteroid) to shade stress, then the effects of shade on phytohormone synthesis and signal transduction, and the molecular mechanism of phytohormone regulation of growth of plants under shade stress. Finally, we propose future research directions.
2 植物感知荫蔽的分子机制当处于荫蔽胁迫时, 植物体首先需要对外界光信号进行感知, 那么在此过程中, 有哪些重要组分参与其中呢? 事实上, 针对不同波长的光, 植物进化出不同种类的光感受器来进行捕捉, 这其中包括感知红光与远红光的光敏色素(phytochromes, PHYs) (Meng and Chory, 2011), 感知蓝光的隐花色素(cryptochromes, CRYs) (Chaves et al., 2011)、向光素(phototropins, PHOTs) (Christie, 2007)以及ZTL (ZEITLUPE)/FKF (Flavin Binding, Kelch Repeat, F-box protein 1)/ LKP2 (LOV KELCH PROTEIN2) (Suetsugu and Wada, 2013) 3种蛋白, 还有感知紫外线(UV-B)的UVR8 (UV Resistance locus 8) (刘明雪等, 2012; Jenkins, 2014)。因此, 植物可以通过不同的光感受器感知外界光信号的变化, 进而判断自身周围的植株密度与荫蔽胁迫程度。 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008)。以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015)。Pfr主要感知远红光变化, 而Pr负责感知红光变化。随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975)。在5种光敏色素中, phyB功能最强, 对其研究也最为深入。phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003)。phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999)。当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008)。 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子。植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011)。在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014)。研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a)。而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013)。 除了R:FR降低外, 蓝光辐射量的降低同样会导致荫蔽胁迫。近年来, 针对低辐射量蓝光引起荫蔽胁迫的研究取得了突破性进展。研究表明, PHYB过表达植株35Spro::PHYB-CFP在低蓝光下, 其下胚轴并未显著伸长, 这表明phyB具有在低蓝光条件下抑制避荫反应的生物学功能(Pedmale et al., 2016)。进一步的研究表明, 作为光敏色素互作因子, PIF4和PIF5除了能与phyB互作外, 也能与隐花色素CRY1和CRY2直接互作, 且在低蓝光条件下, PIF5蛋白的表达量会逐渐升高(Pedmale et al., 2016)。为探明低蓝光与低R:FR引起的避荫反应的差异, 将植株同时用低蓝光与低R:FR处理, 结果显示, 野生型植株叶柄伸长量显著大于低R:FR单独处理。而pif4/pif5/pif7三突变体植株无论是单独或者共同用低蓝光和低R:FR处理, 结果都没有叶柄伸长的表型。这些证据表明低蓝光能增强低R:FR引起的避荫反应, 且低蓝光与低R:FR引起的避荫反应可能都依赖于PIFs (de Wit et al., 2016)。
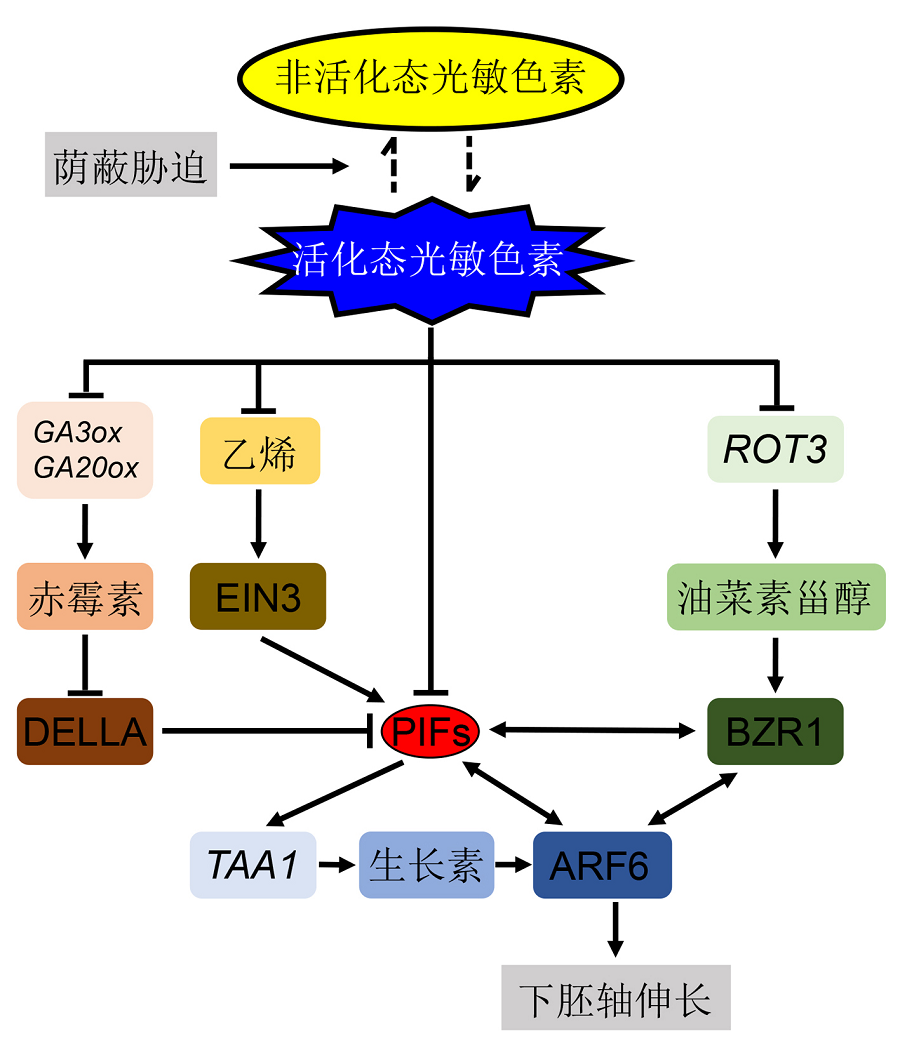
3 参与荫蔽胁迫响应的主要植物激素植物激素在调控植物生长发育过程中发挥关键作用。随着环境的变化, 激素自身含量和信号也会发生相应变化, 进而调控植物产生适应性反应。因此, 植物激素可使植物在变化的环境中保持稳定的生长状态。大量研究表明, 在参与植物荫蔽胁迫响应过程中, 植物激素扮演着关键角色, 这些激素通过调控自身含量及与光信号通路互作等方式实现对荫蔽胁迫的响应(图1)。 3.1 生长素生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013)。最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014)。以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA。在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016)。在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008)。这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000)。进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010)。这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要。 最近的一项研究较为详细地阐明了生长素在荫蔽胁迫下调控下胚轴伸长的分子机制(Procko et al., 2014)。在荫蔽胁迫下, 与对照相比, 切除子叶后的芜菁(Brassica rapa)下胚轴并未显著伸长(Procko et al., 2014)。当对其子叶与下胚轴分别进行荫蔽胁迫时, 发现只有当子叶受到胁迫时, 下胚轴才会显著伸长; 而当子叶接收正常光信号时, 无论下胚轴是否处于荫蔽胁迫, 都不会对下胚轴长度产生影响。在对荫蔽相关的标记基因表达进行分析后发现, 子叶会先于下胚轴对外界荫蔽胁迫作出响应(Procko et al., 2014)。进一步研究表明, 在荫蔽处理下, 下胚轴的生长素浓度自上而下呈递减趋势(Procko et al., 2014)。这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016)。因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长。 在荫蔽胁迫下, 生长素变化与光信号存在怎样的关系呢? 研究表明, 在植物合成下胚轴伸长所需的IAA时, PIF4、PIF5和PIF7均参与调控这一过程, 其中PIF4和PIF5能增强TAA1蛋白的稳定性, 而PIF7调节TAA1的磷酸化(Lorrain et al., 2008; Li et al., 2012a)。PIF4和PIF5还能直接促进YUCCA家族基因的表达, PIF7也能直接作用于YUC8和YUC9的启动子(Hor- nitschek et al., 2012; Li et al., 2012a)。因此, 光信号的改变能通过PIFs直接影响到IAA的含量。而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010)。除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012)。因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-139/img_1.png<b>图1</b> 几种重要植物激素响应荫蔽胁迫的信号通路网络<br/>荫蔽胁迫下, 光敏色素由活化态Pfr形式转变为非活化态Pr形式, 由此减弱对PIFs的抑制作用。不同植物激素对荫蔽胁迫的响应不同, 但最终都会与PIFs产生互作。在荫蔽胁迫下, 赤霉素(GA)合成基因<i>GA3ox</i>和<i>GA20ox</i>的表达量增加, 进而导致GA含量升高, GA含量的增加又会促进DELLA蛋白的降解, 而DELLA蛋白能抑制PIF4行使功能。油菜素甾醇(BR)合成基因<i>ROT3</i>在荫蔽胁迫下表达量也会增加, 进而促进BR含量的升高, BR的增加能促进BR转录因子基因<i>BZR1</i>的转录, 而BZR1能与PIF4蛋白产生结构上的互作。在荫蔽胁迫下, 乙烯(ETH)的含量也会增加, ETH的增加能促进ETH转录因子基因<i>EIN3</i>的转录, 而EIN3被证明能上调<i>PIF3</i>基因的表达。生长素(IAA)的合成和信号转导均受PIFs的影响, 其中PIF4、PIF5和PIF7对IAA的合成具有促进作用, 且PIF4和PIF5还促进IAA下游转录因子基因<i>ARF6</i>的表达, IAA也是促进下胚轴伸长的主要植物激素。荫蔽胁迫下, 不同信号通路之间存在互作关系。例如, 在调控下胚轴伸长时, PIF4、BZR1和ARF6被证明具有协同作用。<br/><b>Figure 1</b> The proposed plant shade response signaling networks mediated by several important phytohormones <br/>Phytochrome is transferred from active Pfr to inactive Pr statuses under shade conditions, and this further attenuates the inhibition effect on PIFs. Although the shade response is distinct among different phytohormones, finally the interaction between all phytohormones with PIFs is documented. Under shade conditions, the transcription level of GA biosynthesis genes <i>GA3ox</i> and <i>GA20ox</i> were increased, which further promotes degradation of DELLA and finally inhibits the functions of PIFs. The expression of BR biosynthesis gene <i>ROT3</i> is also increased under shade conditions, and further enhances BR biosynthesis and promotes <i>BZR1</i> transcription, and finally the interaction between BZR1 and PIF4 was detected. Similar to GA and BR, ethylene level was also increased under shade conditions. The increase of ethylene then induces <i>EIN3</i> transcription, and transcription factor EIN3 further enhances the transcription of <i>PIF3</i>. Both auxin biosynthesis and signaling pathways are regulated by PIFs; in detail, PIF4, PIF5 and PIF7 enhance IAA biogenesis, and PIF4 and PIF5 also promote <i>ARF6</i> expression. So far, auxin is the most important phytohormone mediates plant shade response. In a word, under shade conditions, the crosstalk networks among different phytohormones signaling pathways were documented. For example, the synergy effect of PIF4, BZR1 and ARF6 was detected under shade conditions, with regard to hypocotyl elongation. Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-139/img_1.png<b>图1</b> 几种重要植物激素响应荫蔽胁迫的信号通路网络<br/>荫蔽胁迫下, 光敏色素由活化态Pfr形式转变为非活化态Pr形式, 由此减弱对PIFs的抑制作用。不同植物激素对荫蔽胁迫的响应不同, 但最终都会与PIFs产生互作。在荫蔽胁迫下, 赤霉素(GA)合成基因<i>GA3ox</i>和<i>GA20ox</i>的表达量增加, 进而导致GA含量升高, GA含量的增加又会促进DELLA蛋白的降解, 而DELLA蛋白能抑制PIF4行使功能。油菜素甾醇(BR)合成基因<i>ROT3</i>在荫蔽胁迫下表达量也会增加, 进而促进BR含量的升高, BR的增加能促进BR转录因子基因<i>BZR1</i>的转录, 而BZR1能与PIF4蛋白产生结构上的互作。在荫蔽胁迫下, 乙烯(ETH)的含量也会增加, ETH的增加能促进ETH转录因子基因<i>EIN3</i>的转录, 而EIN3被证明能上调<i>PIF3</i>基因的表达。生长素(IAA)的合成和信号转导均受PIFs的影响, 其中PIF4、PIF5和PIF7对IAA的合成具有促进作用, 且PIF4和PIF5还促进IAA下游转录因子基因<i>ARF6</i>的表达, IAA也是促进下胚轴伸长的主要植物激素。荫蔽胁迫下, 不同信号通路之间存在互作关系。例如, 在调控下胚轴伸长时, PIF4、BZR1和ARF6被证明具有协同作用。<br/><b>Figure 1</b> The proposed plant shade response signaling networks mediated by several important phytohormones <br/>Phytochrome is transferred from active Pfr to inactive Pr statuses under shade conditions, and this further attenuates the inhibition effect on PIFs. Although the shade response is distinct among different phytohormones, finally the interaction between all phytohormones with PIFs is documented. Under shade conditions, the transcription level of GA biosynthesis genes <i>GA3ox</i> and <i>GA20ox</i> were increased, which further promotes degradation of DELLA and finally inhibits the functions of PIFs. The expression of BR biosynthesis gene <i>ROT3</i> is also increased under shade conditions, and further enhances BR biosynthesis and promotes <i>BZR1</i> transcription, and finally the interaction between BZR1 and PIF4 was detected. Similar to GA and BR, ethylene level was also increased under shade conditions. The increase of ethylene then induces <i>EIN3</i> transcription, and transcription factor EIN3 further enhances the transcription of <i>PIF3</i>. Both auxin biosynthesis and signaling pathways are regulated by PIFs; in detail, PIF4, PIF5 and PIF7 enhance IAA biogenesis, and PIF4 and PIF5 also promote <i>ARF6</i> expression. So far, auxin is the most important phytohormone mediates plant shade response. In a word, under shade conditions, the crosstalk networks among different phytohormones signaling pathways were documented. For example, the synergy effect of PIF4, BZR1 and ARF6 was detected under shade conditions, with regard to hypocotyl elongation.
图1 几种重要植物激素响应荫蔽胁迫的信号通路网络 荫蔽胁迫下, 光敏色素由活化态Pfr形式转变为非活化态Pr形式, 由此减弱对PIFs的抑制作用。不同植物激素对荫蔽胁迫的响应不同, 但最终都会与PIFs产生互作。在荫蔽胁迫下, 赤霉素(GA)合成基因GA3ox和GA20ox的表达量增加, 进而导致GA含量升高, GA含量的增加又会促进DELLA蛋白的降解, 而DELLA蛋白能抑制PIF4行使功能。油菜素甾醇(BR)合成基因ROT3在荫蔽胁迫下表达量也会增加, 进而促进BR含量的升高, BR的增加能促进BR转录因子基因BZR1的转录, 而BZR1能与PIF4蛋白产生结构上的互作。在荫蔽胁迫下, 乙烯(ETH)的含量也会增加, ETH的增加能促进ETH转录因子基因EIN3的转录, 而EIN3被证明能上调PIF3基因的表达。生长素(IAA)的合成和信号转导均受PIFs的影响, 其中PIF4、PIF5和PIF7对IAA的合成具有促进作用, 且PIF4和PIF5还促进IAA下游转录因子基因ARF6的表达, IAA也是促进下胚轴伸长的主要植物激素。荫蔽胁迫下, 不同信号通路之间存在互作关系。例如, 在调控下胚轴伸长时, PIF4、BZR1和ARF6被证明具有协同作用。 Figure 1 The proposed plant shade response signaling networks mediated by several important phytohormones Phytochrome is transferred from active Pfr to inactive Pr statuses under shade conditions, and this further attenuates the inhibition effect on PIFs. Although the shade response is distinct among different phytohormones, finally the interaction between all phytohormones with PIFs is documented. Under shade conditions, the transcription level of GA biosynthesis genes GA3ox and GA20ox were increased, which further promotes degradation of DELLA and finally inhibits the functions of PIFs. The expression of BR biosynthesis gene ROT3 is also increased under shade conditions, and further enhances BR biosynthesis and promotes BZR1 transcription, and finally the interaction between BZR1 and PIF4 was detected. Similar to GA and BR, ethylene level was also increased under shade conditions. The increase of ethylene then induces EIN3 transcription, and transcription factor EIN3 further enhances the transcription of PIF3. Both auxin biosynthesis and signaling pathways are regulated by PIFs; in detail, PIF4, PIF5 and PIF7 enhance IAA biogenesis, and PIF4 and PIF5 also promote ARF6 expression. So far, auxin is the most important phytohormone mediates plant shade response. In a word, under shade conditions, the crosstalk networks among different phytohormones signaling pathways were documented. For example, the synergy effect of PIF4, BZR1 and ARF6 was detected under shade conditions, with regard to hypocotyl elongation.
AlabadíD, GilJ, BlázquezMA, García-MmartínezJ (2004). Gibberellins repress photomorphogenesis in darkness. Pl- ant Physiol 134, 1050-1057. [本文引用: 1]
[6]
AlveyL, HarberdNP (2005). DELLA proteins: integrators of multiple plant growth regulatory inputs? Physiol Plantarum 123, 153-160. [本文引用: 1]
[7]
BoccalandroHE, PloschukEL, YanovskyMJ, SánchezRA, GatzC, CasalJJ (2003). Increased phytochrome B alleviates density effects on tuber yield of field potato crops. Plant Physiol 133, 1539-1546. [本文引用: 1]
[8]
BottoJF, SanchezRA, WhitelamGC, CasalJJ (1996). Phytochrome A mediates the promotion of seed germination by very low fluences of light and canopy shade light in Arabidopsis. Plant Physiol 110, 439-444. [本文引用: 1]
[9]
CagnolaJI, PloschukE, Benech-ArnoldT, FinlaysonSA, CasalJJ (2012). Stem transcriptome reveals mechanisms to reduce the energetic cost of shade-avoidance respon- ses in tomato. Plant Physiol 160, 1110-1119. [本文引用: 1]
[10]
CasalJJ (2012). Shade avoidance. Arabidopsis Book 10, e157. [本文引用: 1]
[11]
CasalJJ (2013). Photoreceptor signaling networks in plant responses to shade. Annu Rev Plant Biol 64, 403-427. [本文引用: 1]
[12]
ChavesI, PokornyR, ByrdinM, HoangN, RitzT, BrettelK, EssenLO, van Der HorstGT, BatschauerA, AhmadM (2011). The cryptochromes: blue light photoreceptors in plants and animals. Annu Rev Plant Biol 62, 335-364. [本文引用: 1]
DasD, St OngeKR, VoesenekLACJ, PierikR, SasidharanR (2016). Ethylene- and shade-induced hypocotyl elongation share transcriptome patterns and functional regulators. Plant Physiol 172, 718-733. [本文引用: 2]
[15]
De LucasM, DavièreJM, RodrÍguez-FalcónM, PontinM, Iglesias-PedrazJM, LorrainS, FankhauserC, BlázquezMA, TitarenkoE, PratS (2008). A molecular framework for light and gibberellin control of cell elongation. Nature 451, 480-484. [本文引用: 1]
[16]
de WitM, KeuskampDH, BongersFJ, HornitschekP, GommersCMM, ReinenE, Martínez-CerónC, FankhauserC, PierikR (2016). Integration of phytochrome and cryptochrome signals determines plant growth during competition for light.Curr Biol 26, 3320-3326. [本文引用: 1]
[17]
de WitM, LorrainS, FankhauserC (2014). Auxin-mediated plant architectural changes in response to shade and high temperature.Physiol Plantarum 151, 13-24. [本文引用: 1]
[18]
de WitM, SpoelSH, Sanchez-PerezGF, GommersCMM, PieterseCMJ, VoesenekLACJ, PierikR (2013). Perception of low red:far-red ratio compromises both salicylic acid- and jasmonic acid-dependent pathogen defences in Arabidopsis. Plant J 75, 90-103. [本文引用: 2]
[19]
DevlinPF, YanovskyMJ, KaySA (2003). A genomic analy- sis of the shade avoidance response in Arabidopsis. Plant Physiol 133, 1617-1629. [本文引用: 1]
[20]
Djakovic-PetrovicT, de WitM, VoesenekLACJ, PierikR (2007). DELLA protein function in growth responses to canopy signals. Plant J 51, 117-126. [本文引用: 1]
[21]
DuboisPG, OlsefskiGT, Flint-GarciaSA, SetterTL, HoekengaOA, BrutnellTP (2010). Physiological and genetic characterization of end-of-day far-red light response in maize seedlings. Plant Physiol 154, 173-186. [本文引用: 1]
[22]
EvansJR, PoorterH (2001). Photosynthetic acclimation of plants to growth irradiance: the relative importance of specific leaf area and nitrogen partitioning in maximizing carbon gain. Plant Cell Environ 24, 755-767. [本文引用: 1]
[23]
FengSH, MartinezC, GusmaroliG, WangY, ZhouJL, WangF, ChenLY, YuL, Iglesias-PedrazJM, KircherS, SchäferE, FuXD, FanLM, DengXW (2008). Coordinated regulation ofArabidopsis thaliana development by light and gibberellins. Nature 451, 475-479. [本文引用: 1]
[24]
FinlaysonSA, KrishnareddySR, KebromTH, CasalJJ (2010). Phytochrome regulation of branching in Arabidopsis. Plant Physiol 152, 1914-1927. [本文引用: 1]
[25]
FleetCM, SunTP (2005). A DELLAcate balance: the role of gibberellin in plant morphogenesis. Curr Opin Plant Biol 8, 77-85. [本文引用: 1]
FranklinKA, DavisSJ, StoddartWM, VierstraRD, WhitelamGC (2003). Mutant analyses define multiple roles for phytochrome C in Arabidopsis photomorphogenesis. Plant Cell 15, 1981-1989. [本文引用: 1]
[28]
GamageHK (2011). Phenotypic variation in heteroblastic woody species does not contribute to shade survival. AOB Plants doi:10.1093/aobpla/plr013. [本文引用: 1]
[29]
GongWZ, QiPF, DuJB, SunX, WuXL, SongC, LiuWG, WuYS, YuXB, YongTW, WangXC, YangF, YanYH, YangWY (2014). Transcriptome analysis of shade- induced inhibition on leaf size in relay intercropped soybean. PLoS One 9, e98465. [本文引用: 1]
[30]
HisamatsuT, KingRW, HelliwellCA, KoshiokaM (2005). The involvement of gibberellin 20-oxidase genes in phytochrome-regulated petiole elongation of Arabidopsis. Plant Physiol 138, 1106-1116. [本文引用: 1]
[31]
HolmesMG, SmithH (1975). The function of phytochrome in plants growing in the natural environment. Nature 254, 512-514. [本文引用: 1]
[32]
HornitschekP, KohnenMV, LorrainS, RougemontJ, LjungK, López-VidrieroI, Franco-ZorrillaJM, SolanoR, TrevisanM, PradervandS, XenariosI, FankhauserC (2012). Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. Plant J 71, 699-711. [本文引用: 3]
[33]
JenkinsGI (2014). The UV-B photoreceptor UVR8: from structure to physiology. Plant Cell 26, 21-37. [本文引用: 1]
[34]
KeuskampDH, PollmannS, VoesenekLACJ, PeetersAJM, PierikR (2010). Auxin transport through PIN- FORMED 3 (PIN3) controls shade avoidance and fitness during competition. Proc Natl Acad Sci USA 107, 22740-22744. [本文引用: 2]
KimGT, TsukayaH, UchimiyaH (1998). TheROTUNDIFOLIA3 gene of Arabidopsis thaliana encodes a new member of the cytochrome P-450 family that is required for the regulated polar elongation of leaf cells. Gene Dev 12, 2381-2391. [本文引用: 1]
[37]
KurepinLV, WaltonLJ, HaywardA, EmeryRJ, PharisRP, ReidDM (2012). Interactions between plant hormones and light quality signaling in regulating the shoot growth ofAra- bidopsis thaliana seedlings. Botany 90, 237-246. [本文引用: 1]
[38]
LeivarP, MonteE (2014). PIFs: systems integrators in plant development. Plant Cell 26, 56-78. [本文引用: 1]
[39]
LeivarP, QuailPH (2011). PIFs: pivotal components in a cellular signaling hub. Trends Plant Sci 16, 19-28. [本文引用: 1]
[40]
LiL, LjungK, BretonG, SchmitzRJ, Pruneda-PazJ, Cowing-ZitronC, ColeBJ, IvansLJ, PedmaleUV, JungHS, EckerJR, KaySA, ChoryJ (2012a). Linking photoreceptor excitation to changes in plant architecture. Gene Dev 26, 785-790. [本文引用: 3]
[41]
LiQF, WangC, JiangL, LiS, SunSS, HeJX (2012b).An interaction between BZR1 and DELLAs mediates direct signaling crosstalk between brassinosteroids and gibbe- rellins in Arabidopsis. Sci Signal 5, ra72. [本文引用: 1]
[42]
LiuX, ZhangH, ZhaoY, FengZ, LiQ, YangHQ, LuanS, LiJ, HeZH (2013). Auxin controls seed dormancy through stimulation of abscisic acid signaling by inducing ARF- mediated ABI3 activation in Arabidopsis.Proc Natl Acad Sci USA 110, 15485-15490. [本文引用: 1]
[43]
LocascioA, BlázquezMA, AlabadÍD (2013). Genomic analysis of DELLA protein activity. Plant Cell Physiol 54, 1229-1237. [本文引用: 1]
MuraseK, HiranoY, SunTP, HakoshimaT (2008). Gibberellin-induced DELLA recognition by the gibberellin receptor GID1. Nature 456, 459-463. [本文引用: 1]
[51]
NozueK, HarmerSL, MaloofJN (2011). Genomic analysis of circadian clock-, light-, and growth-correlated genes reveals PHYTOCHROME-INTERACTING FACTOR5 as a modulator of auxin signaling in Arabidopsis. Plant Physiol 156, 357-372. [本文引用: 1]
[52]
NozueK, TatAV, DevisettyUK, RobinsonM, MumbachMR, IchihashiY, LekkalaS, MaloofJN (2015). Shade avoidance components and pathways in adult plants revealed by phenotypic profiling. PLoS Genet 11, e1004953. [本文引用: 1]
[53]
OhE, ZhuJY, BaiMY, ArenhartRA, SunY, WangZY (2014). Cell elongation is regulated through a central circuit of interacting transcription factors in the Arabidopsis hypocotyl.eLife 3, e03031. [本文引用: 2]
[54]
OhE, ZhuJY, WangZY (2012). Interaction between BZR1 and PIF4 integrates brassinosteroid and environmental responses. Nat Cell Biol 14, 802-809. [本文引用: 2]
[55]
PedmaleUV, HuangSC, ZanderM, ColeBJ, HetzelJ, LjungK, ReisPAB, SrideviP, NitoK, NeryJR, EckerJR, ChoryJ (2016). Cryptochromes interact directly with PIFs to control plant growth in limiting blue light.Cell 164, 233-245. [本文引用: 2]
[56]
PierikR, Djakovic-PetrovicT, KeuskampDH, de WitM, VoesenekLACJ (2009). Auxin and ethylene regulate elongation responses to neighbor proximity signals independent of gibberellin and DELLA proteins in Arabidopsis. Plant Physiol 149, 1701-1712. [本文引用: 1]
[57]
PigliucciM, SchmittJ (1999). Genes affecting phenotypic plasticity in Arabidopsis: pleiotropic effects and reproductive fitness of photomorphogenic mutants. J Evol Biol 12, 551-562. [本文引用: 1]
[58]
ProckoC, BurkoY, JaillaisY, LjungK, LongJA, ChoryJ (2016). The epidermis coordinates auxin-induced stem growth in response to shade. Gene Dev 30, 1529-1541. [本文引用: 1]
ReddySK, HolaluSV, CasalJJ, FinlaysonSA (2013). Abscisic acid regulates axillary bud outgrowth responses to the ratio of red to far-red light. Plant Physiol 163, 1047-1058. [本文引用: 1]
[61]
ReedJW, FosterKR, MorganPW, ChoryJ (1996). Phytochrome B affects responsiveness to gibberellins in Arabidopsis. Plant Physiol 112, 337-342. [本文引用: 1]
[62]
ReedJW, NagpalP, PooleDS, FuruyaM, ChoryJ (1993). Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell 5, 147-157. [本文引用: 1]
[63]
ReineckeDM, WickramarathnaAD, OzgaJA, KurepinLV, JinAL, GoodAG, PharisRP (2013). Gibberellin 3-oxi- dase gene expression patterns influence gibberellin biosynthesis, growth, and development in pea. Plant Physiol 163, 929-945. [本文引用: 1]
[64]
SanchezSE, CagnolaJI, CrepyM, YanovskyMJ, CasalJJ (2011). Balancing forces in the photoperiodic control of flowering. Photochem Photobiol Sci 10, 451-460. [本文引用: 1]
[65]
SchwechheimerC (2008). Understanding gibberellic acid signaling—are we there yet? Curr Opin Plant Biol 11, 9-15. [本文引用: 1]
[66]
ShimadaA, Ueguchi-TanakaM, NakatsuT, NakajimaM, NaoeY, OhmiyaH, KatoH, MatsuokaM (2008). Structural basis for gibberellin recognition by its receptor GID1. Nature 456, 520-523. [本文引用: 1]
[67]
ShinomuraT, NagataniA, HanzawaH, KubotaM, Wata- nabeM, FuruyaM (1996). Action spectra for phytochrome A- and B-specific photoinduction of seed germination inArabidopsis thaliana. Proc Natl Acad Sci USA 93, 8129-8133. [本文引用: 1]
[68]
ShuK, ChenQ, WuYR, LiuRJ, ZhangHW, WangPF, LiYL, WangSF, TangSY, LiuCY, YangWY, CaoXF, SerinoG, XieQ (2016a). ABI4 mediates antagonistic effects of abscisic acid and gibberellins at transcript and protein levels. Plant J 85, 348-361. [本文引用: 1]
ShuK, ZhangHW, WangSF, ChenML, WuYR, TangSY, LiuCY, FengYQ, CaoXF, XieQ (2013). ABI4 regulates primary seed dormancy by regulating the biogenesis of abscisic acid and gibberellins in Arabidopsis. PLoS Genet 9, e1003577. [本文引用: 1]
[71]
SmalleJ, HaegmanM, KurepaJ, Van MontaguM, St- raetenDVD (1997). Ethylene can stimulate Arabidopsis hypocotyl elongation in the light. Proc Natl Acad Sci USA 94, 2756-2761. [本文引用: 1]
[72]
SteindlerC, MatteucciA, SessaG, WeimarT, OhgishiM, AoyamaT, MorelliG, RubertiI (1999). Shade avoidance responses are mediated by the ATHB-2 HD-zip protein, a negative regulator of gene expression. Development 126, 4235-4245. [本文引用: 2]
[73]
SuetsuguN, WadaM (2013). Evolution of three LOV blue light receptor families in green plants and photosynthetic stramenopiles: phototropin, ZTL/FKF1/LKP2 and aureochrome.Plant Cell Physiol 54, 8-23. [本文引用: 1]
[74]
TaoY, FerrerJL, LjungK, PojerF, HongF, LongJA, LiL, MorenoJE, BowmanME, IvansLJ, ChengYF, LimJ, ZhaoYD, BallaréCL, SandbergG, NoelJP, ChoryJ (2008). Rapid synthesis of auxin via a new tryptophan- dependent pathway is required for shade avoidance in plants. Cell 133, 164-176. [本文引用: 1]
VertG, WalcherCL, ChoryJ, NemhauserJL (2008). Integration of auxin and brassinosteroid pathways by Auxin Response Factor 2. Proc Natl Acad Sci USA 105, 9829-9834. [本文引用: 1]
[77]
WoodwardAW, BartelB (2005). Auxin: regulation, action, and interaction. Ann Bot 95, 707-735. [本文引用: 1]
[78]
YamaguchiR, NakamuraM, MochizukiN, KaySA, NagataniA (1999). Light-dependent translocation of a phytochrome B-GFP fusion protein to the nucleus in transgenic Arabidopsis. J Cell Biol 145, 437-445. [本文引用: 1]
[79]
ZhangY, MaybaO, PfeifferA, ShiH, TeppermanJM, SpeedTP, QuailPH (2013). A quartet of PIF bHLH factors provides a transcriptionally centered signaling hub that regulates seedling morphogenesis through differential expression-patterning of shared target genes in Arabidopsis. PLoS Genet 9, e1003244. [本文引用: 1]
[80]
ZhaoYD (2012). Auxin biosynthesis: a simple two-step pathway converts tryptophan to indole-3-acetic acid in plants. Mol Plant 5, 334-338. [本文引用: 1]
[81]
ZhongSW, ShiH, XueC, WangL, XiYP, LiJG, QuailPH, DengXW, GuoHW (2012). A molecular framework of light-controlled phytohormone action in Arabidopsis. Curr Biol 22, 1530-1535. [本文引用: 1]
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
光敏色素信号通路中磷酸化修饰研究进展 1 2015
... 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008).以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015).Pfr主要感知远红光变化, 而Pr负责感知红光变化.随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975).在5种光敏色素中, phyB功能最强, 对其研究也最为深入.phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003).phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999).当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008). ...
1 2007
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2004
... 赤霉素是一类调控植物生长发育的激素, 在具有生理活性的赤霉素中, GA1和GA4已被证明参与植物光形态建成, 与光信号关系紧密(Alabadi et al., 2004).植物体内具有生理活性的赤霉素主要由合成基因GA20ox和GA3ox以及分解基因GA2ox调控, 调控这些基因的表达可调节赤霉素的含量, 进而调控茎秆的伸长(Reinecke et al., 2013).荫蔽胁迫下, 赤霉素合成相关基因GA20ox与GA3ox表达量升高(Devlin et al., 2003), 进而导致植株体内GA1、GA19和GA20的含量显著增加, 且植株在荫蔽胁迫下对赤霉素的敏感性也会显著增强(Reed et al., 1996; Hisamatsu et al., 2005). ...
1 2005
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2003
... 在对作物的研究中发现, 荫蔽胁迫会导致大豆叶面积降低(Gong et al., 2014)、玉米茎秆过度伸长(Dubois et al., 2010)、小麦(Triticum aestivum)的分蘖数减少(Ugarte et al., 2010)以及马铃薯(Solanum tuberosum)块茎发育减缓(Boccalandro et al., 2003)等, 这些影响最终都导致作物产量降低.除了对作物形态的影响, 荫蔽胁迫还会降低作物对病原菌的抵抗能力, 增加作物感病的概率及生物量的损失(de Wit et al., 2013), 因此荫蔽胁迫对农业生产极为不利. ...
1 1996
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2012
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 2012
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
... 乙烯作为具有挥发性的植物激素, 不仅能调控植株的生长发育, 还能作为植株感知周围植物生长环境的直接信号.研究表明, 外源施加乙烯或者乙烯的合成前体物质ACC, 拟南芥幼苗的下胚轴比对照显著伸长(Smalle et al., 1997; Das et al., 2016).而在荫蔽胁迫下, 乙烯含量与下胚轴长度均显著增加, 乙烯的不敏感突变体ein2-1和ein3-1/eil1-3在荫蔽下则没有表现出下胚轴过度伸长的表型; 使用乙烯作用抑制剂1-MCP (1-methylcyclopropane)处理植株, 也同样没有出现下胚轴伸长的表型.这说明乙烯在荫蔽胁迫下起正调控作用(Pierik et al., 2009). ... ... 通过转录组分析, 研究人员发现乙烯和荫蔽胁迫在调控下胚轴伸长时具有类似的转录组分和功能调控因子(Das et al., 2016).乙烯信号通路中的转录因子EIN3被证明能结合在PIF3的启动子上, 进而促进PIF3的表达, 通过PIF3来调控下胚轴的长度(Zhong et al., 2012).然而在荫蔽胁迫下, 乙烯含量是如何升高的还有待进一步研究. ...
1 2008
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2016
... 除了R:FR降低外, 蓝光辐射量的降低同样会导致荫蔽胁迫.近年来, 针对低辐射量蓝光引起荫蔽胁迫的研究取得了突破性进展.研究表明, PHYB过表达植株35Spro::PHYB-CFP在低蓝光下, 其下胚轴并未显著伸长, 这表明phyB具有在低蓝光条件下抑制避荫反应的生物学功能(Pedmale et al., 2016).进一步的研究表明, 作为光敏色素互作因子, PIF4和PIF5除了能与phyB互作外, 也能与隐花色素CRY1和CRY2直接互作, 且在低蓝光条件下, PIF5蛋白的表达量会逐渐升高(Pedmale et al., 2016).为探明低蓝光与低R:FR引起的避荫反应的差异, 将植株同时用低蓝光与低R:FR处理, 结果显示, 野生型植株叶柄伸长量显著大于低R:FR单独处理.而pif4/pif5/pif7三突变体植株无论是单独或者共同用低蓝光和低R:FR处理, 结果都没有叶柄伸长的表型.这些证据表明低蓝光能增强低R:FR引起的避荫反应, 且低蓝光与低R:FR引起的避荫反应可能都依赖于PIFs (de Wit et al., 2016). ...
1 2014
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
2 2013
... 在对作物的研究中发现, 荫蔽胁迫会导致大豆叶面积降低(Gong et al., 2014)、玉米茎秆过度伸长(Dubois et al., 2010)、小麦(Triticum aestivum)的分蘖数减少(Ugarte et al., 2010)以及马铃薯(Solanum tuberosum)块茎发育减缓(Boccalandro et al., 2003)等, 这些影响最终都导致作物产量降低.除了对作物形态的影响, 荫蔽胁迫还会降低作物对病原菌的抵抗能力, 增加作物感病的概率及生物量的损失(de Wit et al., 2013), 因此荫蔽胁迫对农业生产极为不利. ... ... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 2003
... 赤霉素是一类调控植物生长发育的激素, 在具有生理活性的赤霉素中, GA1和GA4已被证明参与植物光形态建成, 与光信号关系紧密(Alabadi et al., 2004).植物体内具有生理活性的赤霉素主要由合成基因GA20ox和GA3ox以及分解基因GA2ox调控, 调控这些基因的表达可调节赤霉素的含量, 进而调控茎秆的伸长(Reinecke et al., 2013).荫蔽胁迫下, 赤霉素合成相关基因GA20ox与GA3ox表达量升高(Devlin et al., 2003), 进而导致植株体内GA1、GA19和GA20的含量显著增加, 且植株在荫蔽胁迫下对赤霉素的敏感性也会显著增强(Reed et al., 1996; Hisamatsu et al., 2005). ...
1 2007
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2010
... 在对作物的研究中发现, 荫蔽胁迫会导致大豆叶面积降低(Gong et al., 2014)、玉米茎秆过度伸长(Dubois et al., 2010)、小麦(Triticum aestivum)的分蘖数减少(Ugarte et al., 2010)以及马铃薯(Solanum tuberosum)块茎发育减缓(Boccalandro et al., 2003)等, 这些影响最终都导致作物产量降低.除了对作物形态的影响, 荫蔽胁迫还会降低作物对病原菌的抵抗能力, 增加作物感病的概率及生物量的损失(de Wit et al., 2013), 因此荫蔽胁迫对农业生产极为不利. ...
1 2001
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2008
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2010
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2005
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
2 2008
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ... ... 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008).以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015).Pfr主要感知远红光变化, 而Pr负责感知红光变化.随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975).在5种光敏色素中, phyB功能最强, 对其研究也最为深入.phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003).phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999).当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008). ...
1 2003
... 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008).以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015).Pfr主要感知远红光变化, 而Pr负责感知红光变化.随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975).在5种光敏色素中, phyB功能最强, 对其研究也最为深入.phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003).phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999).当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008). ...
1 2011
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2014
... 在对作物的研究中发现, 荫蔽胁迫会导致大豆叶面积降低(Gong et al., 2014)、玉米茎秆过度伸长(Dubois et al., 2010)、小麦(Triticum aestivum)的分蘖数减少(Ugarte et al., 2010)以及马铃薯(Solanum tuberosum)块茎发育减缓(Boccalandro et al., 2003)等, 这些影响最终都导致作物产量降低.除了对作物形态的影响, 荫蔽胁迫还会降低作物对病原菌的抵抗能力, 增加作物感病的概率及生物量的损失(de Wit et al., 2013), 因此荫蔽胁迫对农业生产极为不利. ...
1 2005
... 赤霉素是一类调控植物生长发育的激素, 在具有生理活性的赤霉素中, GA1和GA4已被证明参与植物光形态建成, 与光信号关系紧密(Alabadi et al., 2004).植物体内具有生理活性的赤霉素主要由合成基因GA20ox和GA3ox以及分解基因GA2ox调控, 调控这些基因的表达可调节赤霉素的含量, 进而调控茎秆的伸长(Reinecke et al., 2013).荫蔽胁迫下, 赤霉素合成相关基因GA20ox与GA3ox表达量升高(Devlin et al., 2003), 进而导致植株体内GA1、GA19和GA20的含量显著增加, 且植株在荫蔽胁迫下对赤霉素的敏感性也会显著增强(Reed et al., 1996; Hisamatsu et al., 2005). ...
1 1975
... 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008).以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015).Pfr主要感知远红光变化, 而Pr负责感知红光变化.随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975).在5种光敏色素中, phyB功能最强, 对其研究也最为深入.phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003).phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999).当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008). ...
3 2012
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ... ... 在荫蔽胁迫下, 生长素变化与光信号存在怎样的关系呢? 研究表明, 在植物合成下胚轴伸长所需的IAA时, PIF4、PIF5和PIF7均参与调控这一过程, 其中PIF4和PIF5能增强TAA1蛋白的稳定性, 而PIF7调节TAA1的磷酸化(Lorrain et al., 2008; Li et al., 2012a).PIF4和PIF5还能直接促进YUCCA家族基因的表达, PIF7也能直接作用于YUC8和YUC9的启动子(Hor- nitschek et al., 2012; Li et al., 2012a).因此, 光信号的改变能通过PIFs直接影响到IAA的含量.而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010).除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ... ... ; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ...
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ... ... 在荫蔽胁迫下, 生长素变化与光信号存在怎样的关系呢? 研究表明, 在植物合成下胚轴伸长所需的IAA时, PIF4、PIF5和PIF7均参与调控这一过程, 其中PIF4和PIF5能增强TAA1蛋白的稳定性, 而PIF7调节TAA1的磷酸化(Lorrain et al., 2008; Li et al., 2012a).PIF4和PIF5还能直接促进YUCCA家族基因的表达, PIF7也能直接作用于YUC8和YUC9的启动子(Hor- nitschek et al., 2012; Li et al., 2012a).因此, 光信号的改变能通过PIFs直接影响到IAA的含量.而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010).除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ...
3 2011
... 油菜素甾醇(BR)是目前已知的唯一一种固醇类植物激素, 其对下胚轴的伸长同样具有至关重要的作用.例如, 在荫蔽胁迫下, BR的合成突变体diminuto/ dwarf1未出现下胚轴过度伸长的表型(Luccioni et al., 2002).而在低蓝光引起的荫蔽胁迫下, BR的另一合成突变体rot3与受体突变体bri1-1的下胚轴伸长均受到显著抑制(Kim et al., 1998; Keuskamp et al., 2011).对野生型植株外源施加BR的合成抑制剂芸苔素唑(brassinazole)同样也能观察到下胚轴伸长被抑制的现象(Keuskamp et al., 2011).然而在对BR及其转录因子BZR1 (Brassinazole Resistant 1)的研究中发现, 无论是BR的含量还是BZR1的表达量都不会随着光信号的变化而变化, 这表明在荫蔽胁迫下, 低R:FR促进植株对BR的敏感性(Luo et al., 2010). ... ... ).对野生型植株外源施加BR的合成抑制剂芸苔素唑(brassinazole)同样也能观察到下胚轴伸长被抑制的现象(Keuskamp et al., 2011).然而在对BR及其转录因子BZR1 (Brassinazole Resistant 1)的研究中发现, 无论是BR的含量还是BZR1的表达量都不会随着光信号的变化而变化, 这表明在荫蔽胁迫下, 低R:FR促进植株对BR的敏感性(Luo et al., 2010). ... ... 在进一步对BR转录因子BZR1的结构研究中发现, BZR1与PIF4能在N端产生结构上的互作(Oh et al., 2012).在后续的研究中, 人们发现在调控下胚轴生长时BZR1与PIF彼此依赖, 无论是PIFs的降解或BZR1活性的降低, 都会减弱它们对下胚轴生长的调控(Oh et al., 2012).BR除了与PIF有联系, 还可与生长素相互作用.例如, 在激素互作的研究中, 发现同时施用生长素和BR对促进下胚轴生长有叠加作用(Keuskamp et al., 2011).分子层面的研究更是揭示了BR与生长素的关系.BR信号途径中的GSK3型激酶BIN2 (BRASSINOSTEROID-INSENSITIVE 2)能通过氧化磷酸化作用降低生长素的转录因子阻遏物ARF2对DNA的结合能力和转录抑制能力, 从而使活化型ARFs能调控下游基因的表达(Vert et al., 2008); 且生长素的另一活化型转录因子ARF6也被证明与BZR1具有共同的靶标基因(Oh et al., 2014).除了与生长素相关, BR与GA同样有关联.通过荧光蛋白标记, 证明BZR1与DELLA蛋白中的RGA互相作用, 且通过抑制对方的活性调控其目的基因的表达(Oh et al., 2014).BR调控下胚轴伸长的方式较为复杂, 主要通过与多种信号通路, 尤其是生长素信号通路互作而实现调控. ...
1 1998
... 油菜素甾醇(BR)是目前已知的唯一一种固醇类植物激素, 其对下胚轴的伸长同样具有至关重要的作用.例如, 在荫蔽胁迫下, BR的合成突变体diminuto/ dwarf1未出现下胚轴过度伸长的表型(Luccioni et al., 2002).而在低蓝光引起的荫蔽胁迫下, BR的另一合成突变体rot3与受体突变体bri1-1的下胚轴伸长均受到显著抑制(Kim et al., 1998; Keuskamp et al., 2011).对野生型植株外源施加BR的合成抑制剂芸苔素唑(brassinazole)同样也能观察到下胚轴伸长被抑制的现象(Keuskamp et al., 2011).然而在对BR及其转录因子BZR1 (Brassinazole Resistant 1)的研究中发现, 无论是BR的含量还是BZR1的表达量都不会随着光信号的变化而变化, 这表明在荫蔽胁迫下, 低R:FR促进植株对BR的敏感性(Luo et al., 2010). ...
1 2012
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 2014
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ...
1 2011
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ...
3 2012
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ... ... 在荫蔽胁迫下, 生长素变化与光信号存在怎样的关系呢? 研究表明, 在植物合成下胚轴伸长所需的IAA时, PIF4、PIF5和PIF7均参与调控这一过程, 其中PIF4和PIF5能增强TAA1蛋白的稳定性, 而PIF7调节TAA1的磷酸化(Lorrain et al., 2008; Li et al., 2012a).PIF4和PIF5还能直接促进YUCCA家族基因的表达, PIF7也能直接作用于YUC8和YUC9的启动子(Hor- nitschek et al., 2012; Li et al., 2012a).因此, 光信号的改变能通过PIFs直接影响到IAA的含量.而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010).除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ... ... ; Li et al., 2012a).因此, 光信号的改变能通过PIFs直接影响到IAA的含量.而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010).除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ...
1 2012
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ...
1 2013
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
1 2013
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
3 2008
... 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008).以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015).Pfr主要感知远红光变化, 而Pr负责感知红光变化.随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975).在5种光敏色素中, phyB功能最强, 对其研究也最为深入.phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003).phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999).当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008). ... ... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ... ... 在荫蔽胁迫下, 生长素变化与光信号存在怎样的关系呢? 研究表明, 在植物合成下胚轴伸长所需的IAA时, PIF4、PIF5和PIF7均参与调控这一过程, 其中PIF4和PIF5能增强TAA1蛋白的稳定性, 而PIF7调节TAA1的磷酸化(Lorrain et al., 2008; Li et al., 2012a).PIF4和PIF5还能直接促进YUCCA家族基因的表达, PIF7也能直接作用于YUC8和YUC9的启动子(Hor- nitschek et al., 2012; Li et al., 2012a).因此, 光信号的改变能通过PIFs直接影响到IAA的含量.而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010).除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ...
1 2002
... 油菜素甾醇(BR)是目前已知的唯一一种固醇类植物激素, 其对下胚轴的伸长同样具有至关重要的作用.例如, 在荫蔽胁迫下, BR的合成突变体diminuto/ dwarf1未出现下胚轴过度伸长的表型(Luccioni et al., 2002).而在低蓝光引起的荫蔽胁迫下, BR的另一合成突变体rot3与受体突变体bri1-1的下胚轴伸长均受到显著抑制(Kim et al., 1998; Keuskamp et al., 2011).对野生型植株外源施加BR的合成抑制剂芸苔素唑(brassinazole)同样也能观察到下胚轴伸长被抑制的现象(Keuskamp et al., 2011).然而在对BR及其转录因子BZR1 (Brassinazole Resistant 1)的研究中发现, 无论是BR的含量还是BZR1的表达量都不会随着光信号的变化而变化, 这表明在荫蔽胁迫下, 低R:FR促进植株对BR的敏感性(Luo et al., 2010). ...
1 2010
... 油菜素甾醇(BR)是目前已知的唯一一种固醇类植物激素, 其对下胚轴的伸长同样具有至关重要的作用.例如, 在荫蔽胁迫下, BR的合成突变体diminuto/ dwarf1未出现下胚轴过度伸长的表型(Luccioni et al., 2002).而在低蓝光引起的荫蔽胁迫下, BR的另一合成突变体rot3与受体突变体bri1-1的下胚轴伸长均受到显著抑制(Kim et al., 1998; Keuskamp et al., 2011).对野生型植株外源施加BR的合成抑制剂芸苔素唑(brassinazole)同样也能观察到下胚轴伸长被抑制的现象(Keuskamp et al., 2011).然而在对BR及其转录因子BZR1 (Brassinazole Resistant 1)的研究中发现, 无论是BR的含量还是BZR1的表达量都不会随着光信号的变化而变化, 这表明在荫蔽胁迫下, 低R:FR促进植株对BR的敏感性(Luo et al., 2010). ...
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 2000
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
1 2008
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2011
... 在荫蔽胁迫下, 生长素变化与光信号存在怎样的关系呢? 研究表明, 在植物合成下胚轴伸长所需的IAA时, PIF4、PIF5和PIF7均参与调控这一过程, 其中PIF4和PIF5能增强TAA1蛋白的稳定性, 而PIF7调节TAA1的磷酸化(Lorrain et al., 2008; Li et al., 2012a).PIF4和PIF5还能直接促进YUCCA家族基因的表达, PIF7也能直接作用于YUC8和YUC9的启动子(Hor- nitschek et al., 2012; Li et al., 2012a).因此, 光信号的改变能通过PIFs直接影响到IAA的含量.而对于生长素的转运, 因荫蔽胁迫而产生的过量生长素会上调PIN3的表达, 从而使生长素被更快地运送到下胚轴(Keuskamp et al., 2010).除了对生长素含量和分配的影响, 在低光合有效辐射条件下, PIF4和PIF5还能增加植物对生长素的敏感性(Nozue et al., 2011; Hornitschek et al., 2012).因此, 荫蔽胁迫下光信号的改变对生长素的含量、转运以及植物对其敏感度均有影响, 生长素也被认为是调控下胚轴伸长最关键的植物激素. ...
1 2015
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
2 2014
... 在进一步对BR转录因子BZR1的结构研究中发现, BZR1与PIF4能在N端产生结构上的互作(Oh et al., 2012).在后续的研究中, 人们发现在调控下胚轴生长时BZR1与PIF彼此依赖, 无论是PIFs的降解或BZR1活性的降低, 都会减弱它们对下胚轴生长的调控(Oh et al., 2012).BR除了与PIF有联系, 还可与生长素相互作用.例如, 在激素互作的研究中, 发现同时施用生长素和BR对促进下胚轴生长有叠加作用(Keuskamp et al., 2011).分子层面的研究更是揭示了BR与生长素的关系.BR信号途径中的GSK3型激酶BIN2 (BRASSINOSTEROID-INSENSITIVE 2)能通过氧化磷酸化作用降低生长素的转录因子阻遏物ARF2对DNA的结合能力和转录抑制能力, 从而使活化型ARFs能调控下游基因的表达(Vert et al., 2008); 且生长素的另一活化型转录因子ARF6也被证明与BZR1具有共同的靶标基因(Oh et al., 2014).除了与生长素相关, BR与GA同样有关联.通过荧光蛋白标记, 证明BZR1与DELLA蛋白中的RGA互相作用, 且通过抑制对方的活性调控其目的基因的表达(Oh et al., 2014).BR调控下胚轴伸长的方式较为复杂, 主要通过与多种信号通路, 尤其是生长素信号通路互作而实现调控. ... ... ).除了与生长素相关, BR与GA同样有关联.通过荧光蛋白标记, 证明BZR1与DELLA蛋白中的RGA互相作用, 且通过抑制对方的活性调控其目的基因的表达(Oh et al., 2014).BR调控下胚轴伸长的方式较为复杂, 主要通过与多种信号通路, 尤其是生长素信号通路互作而实现调控. ...
2 2012
... 在进一步对BR转录因子BZR1的结构研究中发现, BZR1与PIF4能在N端产生结构上的互作(Oh et al., 2012).在后续的研究中, 人们发现在调控下胚轴生长时BZR1与PIF彼此依赖, 无论是PIFs的降解或BZR1活性的降低, 都会减弱它们对下胚轴生长的调控(Oh et al., 2012).BR除了与PIF有联系, 还可与生长素相互作用.例如, 在激素互作的研究中, 发现同时施用生长素和BR对促进下胚轴生长有叠加作用(Keuskamp et al., 2011).分子层面的研究更是揭示了BR与生长素的关系.BR信号途径中的GSK3型激酶BIN2 (BRASSINOSTEROID-INSENSITIVE 2)能通过氧化磷酸化作用降低生长素的转录因子阻遏物ARF2对DNA的结合能力和转录抑制能力, 从而使活化型ARFs能调控下游基因的表达(Vert et al., 2008); 且生长素的另一活化型转录因子ARF6也被证明与BZR1具有共同的靶标基因(Oh et al., 2014).除了与生长素相关, BR与GA同样有关联.通过荧光蛋白标记, 证明BZR1与DELLA蛋白中的RGA互相作用, 且通过抑制对方的活性调控其目的基因的表达(Oh et al., 2014).BR调控下胚轴伸长的方式较为复杂, 主要通过与多种信号通路, 尤其是生长素信号通路互作而实现调控. ... ... ).在后续的研究中, 人们发现在调控下胚轴生长时BZR1与PIF彼此依赖, 无论是PIFs的降解或BZR1活性的降低, 都会减弱它们对下胚轴生长的调控(Oh et al., 2012).BR除了与PIF有联系, 还可与生长素相互作用.例如, 在激素互作的研究中, 发现同时施用生长素和BR对促进下胚轴生长有叠加作用(Keuskamp et al., 2011).分子层面的研究更是揭示了BR与生长素的关系.BR信号途径中的GSK3型激酶BIN2 (BRASSINOSTEROID-INSENSITIVE 2)能通过氧化磷酸化作用降低生长素的转录因子阻遏物ARF2对DNA的结合能力和转录抑制能力, 从而使活化型ARFs能调控下游基因的表达(Vert et al., 2008); 且生长素的另一活化型转录因子ARF6也被证明与BZR1具有共同的靶标基因(Oh et al., 2014).除了与生长素相关, BR与GA同样有关联.通过荧光蛋白标记, 证明BZR1与DELLA蛋白中的RGA互相作用, 且通过抑制对方的活性调控其目的基因的表达(Oh et al., 2014).BR调控下胚轴伸长的方式较为复杂, 主要通过与多种信号通路, 尤其是生长素信号通路互作而实现调控. ...
2 2016
... 除了R:FR降低外, 蓝光辐射量的降低同样会导致荫蔽胁迫.近年来, 针对低辐射量蓝光引起荫蔽胁迫的研究取得了突破性进展.研究表明, PHYB过表达植株35Spro::PHYB-CFP在低蓝光下, 其下胚轴并未显著伸长, 这表明phyB具有在低蓝光条件下抑制避荫反应的生物学功能(Pedmale et al., 2016).进一步的研究表明, 作为光敏色素互作因子, PIF4和PIF5除了能与phyB互作外, 也能与隐花色素CRY1和CRY2直接互作, 且在低蓝光条件下, PIF5蛋白的表达量会逐渐升高(Pedmale et al., 2016).为探明低蓝光与低R:FR引起的避荫反应的差异, 将植株同时用低蓝光与低R:FR处理, 结果显示, 野生型植株叶柄伸长量显著大于低R:FR单独处理.而pif4/pif5/pif7三突变体植株无论是单独或者共同用低蓝光和低R:FR处理, 结果都没有叶柄伸长的表型.这些证据表明低蓝光能增强低R:FR引起的避荫反应, 且低蓝光与低R:FR引起的避荫反应可能都依赖于PIFs (de Wit et al., 2016). ... ... ).进一步的研究表明, 作为光敏色素互作因子, PIF4和PIF5除了能与phyB互作外, 也能与隐花色素CRY1和CRY2直接互作, 且在低蓝光条件下, PIF5蛋白的表达量会逐渐升高(Pedmale et al., 2016).为探明低蓝光与低R:FR引起的避荫反应的差异, 将植株同时用低蓝光与低R:FR处理, 结果显示, 野生型植株叶柄伸长量显著大于低R:FR单独处理.而pif4/pif5/pif7三突变体植株无论是单独或者共同用低蓝光和低R:FR处理, 结果都没有叶柄伸长的表型.这些证据表明低蓝光能增强低R:FR引起的避荫反应, 且低蓝光与低R:FR引起的避荫反应可能都依赖于PIFs (de Wit et al., 2016). ...
1 2009
... 乙烯作为具有挥发性的植物激素, 不仅能调控植株的生长发育, 还能作为植株感知周围植物生长环境的直接信号.研究表明, 外源施加乙烯或者乙烯的合成前体物质ACC, 拟南芥幼苗的下胚轴比对照显著伸长(Smalle et al., 1997; Das et al., 2016).而在荫蔽胁迫下, 乙烯含量与下胚轴长度均显著增加, 乙烯的不敏感突变体ein2-1和ein3-1/eil1-3在荫蔽下则没有表现出下胚轴过度伸长的表型; 使用乙烯作用抑制剂1-MCP (1-methylcyclopropane)处理植株, 也同样没有出现下胚轴伸长的表型.这说明乙烯在荫蔽胁迫下起正调控作用(Pierik et al., 2009). ...
1 1999
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2016
... 最近的一项研究较为详细地阐明了生长素在荫蔽胁迫下调控下胚轴伸长的分子机制(Procko et al., 2014).在荫蔽胁迫下, 与对照相比, 切除子叶后的芜菁(Brassica rapa)下胚轴并未显著伸长(Procko et al., 2014).当对其子叶与下胚轴分别进行荫蔽胁迫时, 发现只有当子叶受到胁迫时, 下胚轴才会显著伸长; 而当子叶接收正常光信号时, 无论下胚轴是否处于荫蔽胁迫, 都不会对下胚轴长度产生影响.在对荫蔽相关的标记基因表达进行分析后发现, 子叶会先于下胚轴对外界荫蔽胁迫作出响应(Procko et al., 2014).进一步研究表明, 在荫蔽处理下, 下胚轴的生长素浓度自上而下呈递减趋势(Procko et al., 2014).这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016).因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长. ...
6 2014
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ... ... 最近的一项研究较为详细地阐明了生长素在荫蔽胁迫下调控下胚轴伸长的分子机制(Procko et al., 2014).在荫蔽胁迫下, 与对照相比, 切除子叶后的芜菁(Brassica rapa)下胚轴并未显著伸长(Procko et al., 2014).当对其子叶与下胚轴分别进行荫蔽胁迫时, 发现只有当子叶受到胁迫时, 下胚轴才会显著伸长; 而当子叶接收正常光信号时, 无论下胚轴是否处于荫蔽胁迫, 都不会对下胚轴长度产生影响.在对荫蔽相关的标记基因表达进行分析后发现, 子叶会先于下胚轴对外界荫蔽胁迫作出响应(Procko et al., 2014).进一步研究表明, 在荫蔽处理下, 下胚轴的生长素浓度自上而下呈递减趋势(Procko et al., 2014).这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016).因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长. ... ... )下胚轴并未显著伸长(Procko et al., 2014).当对其子叶与下胚轴分别进行荫蔽胁迫时, 发现只有当子叶受到胁迫时, 下胚轴才会显著伸长; 而当子叶接收正常光信号时, 无论下胚轴是否处于荫蔽胁迫, 都不会对下胚轴长度产生影响.在对荫蔽相关的标记基因表达进行分析后发现, 子叶会先于下胚轴对外界荫蔽胁迫作出响应(Procko et al., 2014).进一步研究表明, 在荫蔽处理下, 下胚轴的生长素浓度自上而下呈递减趋势(Procko et al., 2014).这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016).因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长. ... ... ).当对其子叶与下胚轴分别进行荫蔽胁迫时, 发现只有当子叶受到胁迫时, 下胚轴才会显著伸长; 而当子叶接收正常光信号时, 无论下胚轴是否处于荫蔽胁迫, 都不会对下胚轴长度产生影响.在对荫蔽相关的标记基因表达进行分析后发现, 子叶会先于下胚轴对外界荫蔽胁迫作出响应(Procko et al., 2014).进一步研究表明, 在荫蔽处理下, 下胚轴的生长素浓度自上而下呈递减趋势(Procko et al., 2014).这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016).因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长. ... ... ).进一步研究表明, 在荫蔽处理下, 下胚轴的生长素浓度自上而下呈递减趋势(Procko et al., 2014).这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016).因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长. ... ... ).这表明当生长素从子叶被运输到下胚轴后, 其首先促进下胚轴表皮细胞的伸长, 在表皮细胞伸长后, 内部组织的生长才能不被表皮细胞所限制, 进而导致下胚轴整体伸长(Procko et al., 2014, 2016).因此, 在荫蔽胁迫下, 植株子叶中合成大量生长素, 然后被运输到下胚轴并作用于表皮细胞, 进而促进下胚轴过度伸长. ...
1 2013
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 1996
... 赤霉素是一类调控植物生长发育的激素, 在具有生理活性的赤霉素中, GA1和GA4已被证明参与植物光形态建成, 与光信号关系紧密(Alabadi et al., 2004).植物体内具有生理活性的赤霉素主要由合成基因GA20ox和GA3ox以及分解基因GA2ox调控, 调控这些基因的表达可调节赤霉素的含量, 进而调控茎秆的伸长(Reinecke et al., 2013).荫蔽胁迫下, 赤霉素合成相关基因GA20ox与GA3ox表达量升高(Devlin et al., 2003), 进而导致植株体内GA1、GA19和GA20的含量显著增加, 且植株在荫蔽胁迫下对赤霉素的敏感性也会显著增强(Reed et al., 1996; Hisamatsu et al., 2005). ...
1 1993
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2013
... 赤霉素是一类调控植物生长发育的激素, 在具有生理活性的赤霉素中, GA1和GA4已被证明参与植物光形态建成, 与光信号关系紧密(Alabadi et al., 2004).植物体内具有生理活性的赤霉素主要由合成基因GA20ox和GA3ox以及分解基因GA2ox调控, 调控这些基因的表达可调节赤霉素的含量, 进而调控茎秆的伸长(Reinecke et al., 2013).荫蔽胁迫下, 赤霉素合成相关基因GA20ox与GA3ox表达量升高(Devlin et al., 2003), 进而导致植株体内GA1、GA19和GA20的含量显著增加, 且植株在荫蔽胁迫下对赤霉素的敏感性也会显著增强(Reed et al., 1996; Hisamatsu et al., 2005). ...
1 2011
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2008
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 2008
... 在赤霉素信号转导通路中, 起关键作用的是下游的DELLA家族蛋白.DELLA蛋白负调控赤霉素下游响应基因的表达, 进而抑制赤霉素调控的植物生长发育, 是一类生长抑制蛋白(Alvey and Harberd, 2005; Achard et al., 2007).例如, 拟南芥体内共有5种DEL- LA蛋白, 分别是GAI、RGA、RGL1、RGL2和RGL3 (Fleet and Sun, 2005).当植物体内GA含量升高时, 赤霉素受体GID1 (GIBBERELLIN INSENSITIVE DWARF1)通过识别GA信号分子进而改变自身构型, 再与DELLA蛋白相结合形成复合体(Murase et al., 2008; Shimada et al., 2008), 该复合体再通过与SCFSLYE3泛素连接酶互作, 进而将DELLA蛋白降解, 从而解除其对下游基因转录的抑制(Schwechheimer, 2008).生化分析表明, DELLA蛋白能与PIFs结合, 从而阻碍其调控下游基因的表达(Locascio et al., 2013).深入研究后发现, DELLA蛋白可直接与PIF4结合, 进而使PIF4完全失去功能(de-Lucas et al., 2008; Feng et al., 2008).而通过绿色荧光蛋白标记分析, 发现在荫蔽胁迫下, 细胞核内的RGA1几乎完全消失, 这也证明荫蔽胁迫时植物下胚轴的伸长与DELLA蛋白的降解相关(Djakovic-Petrovic et al., 2007).综上所述, 荫蔽胁迫下光信号的改变主要是促进GA含量的升高, 进而减弱DELLA蛋白对生长的抑制, 从而促进下胚轴伸长. ...
1 1996
... 荫蔽对植物的整个生命周期均有不同程度的影响.当荫蔽程度较弱时, 植物对荫蔽的响应主要为耐荫反应(shade tolerance).不同植物的耐荫性各不相同.研究表明, 比叶面积(specific leaf area, SLA)与植物的耐荫性密切相关(Evans and Poorter, 2001).高比叶面积的植物对荫蔽更为敏感, 耐荫性也更低(Gam- age, 2011).而随着荫蔽程度加剧, 当超过植物的耐受性时, 植物便会产生相应的避荫反应(shade av- oidance).为使幼苗不在荫蔽胁迫下生长(Casal, 2012), 在种子萌发阶段, 荫蔽胁迫下的种子便会显著延迟萌发, 只有当外界荫蔽信号解除, 种子才能回复到正常萌发模式(Botto et al., 1996; Shinomura et al., 1996).在萌发后的生长阶段, 荫蔽胁迫会影响植物整体株型的变化, 最显著的表现是下胚轴、叶柄以及茎秆的过度伸长(Franklin, 2008), 这将有利于植物在胁迫下获得更多光照.而正是由于植物将更多的能量供给下胚轴、叶柄以及茎秆生长, 荫蔽胁迫下植物的叶片面积和分枝数均会显著降低(Reed et al., 1993; Finlayson et al., 2010).到了生殖生长阶段, 为了繁衍后代, 荫蔽胁迫下的植物花期将会提前(Pigli- ucci and Schmitt, 1999; Sanchez et al., 2011), 且最终因单株荚数的减少而降低产量(Procko et al., 2014).植物适应荫蔽胁迫的这些现象, 称作避荫综合征(shade avoidance syndrome, SAS). ...
1 2016
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 2016
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 2013
... 植物激素作为调控植物生长发育的关键因子, 它们在荫蔽胁迫下发挥重要作用.除了上述提到的4种植物激素, 还有其它植物激素参与荫蔽胁迫响应.例如, 脱落酸(ABA)在调控种子休眠与萌发(Shu et al., 2013)、幼苗生长(Shu et al., 2016a)及开花时间(Shu et al., 2016b)等方面具有重要作用; 此外, 脱落酸还被证明是植物响应非生物胁迫最关键的植物激素(Mittler and Blumwald, 2015), 而荫蔽胁迫正是非生物胁迫之一.研究表明, 在荫蔽胁迫下, 脱落酸能抑制植物腋芽的生长(Reddy et al., 2013), 且叶片中脱落酸的含量会显著上升(Cagnola et al., 2012).但是, 作为调控非生物胁迫的关键植物激素, 脱落酸响应荫蔽胁迫的分子机理还未得到深入研究.而其它植物激素如细胞分裂素(Kurepin et al., 2012)、水杨酸及茉莉酸(de Wit et al., 2013)等, 在荫蔽胁迫下, 植物对它们的敏感性也都会有相应的变化.然而, 目前的研究同样无法证明它们受光信号调节, 或是由于其它植物激素的改变而间接调节.总之, 这些植物激素是否及如何参与植物荫蔽胁迫响应, 值得深入研究. ...
1 1997
... 乙烯作为具有挥发性的植物激素, 不仅能调控植株的生长发育, 还能作为植株感知周围植物生长环境的直接信号.研究表明, 外源施加乙烯或者乙烯的合成前体物质ACC, 拟南芥幼苗的下胚轴比对照显著伸长(Smalle et al., 1997; Das et al., 2016).而在荫蔽胁迫下, 乙烯含量与下胚轴长度均显著增加, 乙烯的不敏感突变体ein2-1和ein3-1/eil1-3在荫蔽下则没有表现出下胚轴过度伸长的表型; 使用乙烯作用抑制剂1-MCP (1-methylcyclopropane)处理植株, 也同样没有出现下胚轴伸长的表型.这说明乙烯在荫蔽胁迫下起正调控作用(Pierik et al., 2009). ...
2 1999
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ... ... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
1 2010
... 在对作物的研究中发现, 荫蔽胁迫会导致大豆叶面积降低(Gong et al., 2014)、玉米茎秆过度伸长(Dubois et al., 2010)、小麦(Triticum aestivum)的分蘖数减少(Ugarte et al., 2010)以及马铃薯(Solanum tuberosum)块茎发育减缓(Boccalandro et al., 2003)等, 这些影响最终都导致作物产量降低.除了对作物形态的影响, 荫蔽胁迫还会降低作物对病原菌的抵抗能力, 增加作物感病的概率及生物量的损失(de Wit et al., 2013), 因此荫蔽胁迫对农业生产极为不利. ...
1 2008
... 在进一步对BR转录因子BZR1的结构研究中发现, BZR1与PIF4能在N端产生结构上的互作(Oh et al., 2012).在后续的研究中, 人们发现在调控下胚轴生长时BZR1与PIF彼此依赖, 无论是PIFs的降解或BZR1活性的降低, 都会减弱它们对下胚轴生长的调控(Oh et al., 2012).BR除了与PIF有联系, 还可与生长素相互作用.例如, 在激素互作的研究中, 发现同时施用生长素和BR对促进下胚轴生长有叠加作用(Keuskamp et al., 2011).分子层面的研究更是揭示了BR与生长素的关系.BR信号途径中的GSK3型激酶BIN2 (BRASSINOSTEROID-INSENSITIVE 2)能通过氧化磷酸化作用降低生长素的转录因子阻遏物ARF2对DNA的结合能力和转录抑制能力, 从而使活化型ARFs能调控下游基因的表达(Vert et al., 2008); 且生长素的另一活化型转录因子ARF6也被证明与BZR1具有共同的靶标基因(Oh et al., 2014).除了与生长素相关, BR与GA同样有关联.通过荧光蛋白标记, 证明BZR1与DELLA蛋白中的RGA互相作用, 且通过抑制对方的活性调控其目的基因的表达(Oh et al., 2014).BR调控下胚轴伸长的方式较为复杂, 主要通过与多种信号通路, 尤其是生长素信号通路互作而实现调控. ...
1 2005
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
1 1999
... 由于荫蔽胁迫下R:FR显著降低, 因此在感知荫蔽胁迫时, 光敏色素发挥最主要的作用(Franklin, 2008).以拟南芥为例, 其体内共有5种光敏色素(phyA-phyE)用于感知R:FR的变化, 不同的光敏色素又以是否具有生理功能而将其分为活化(Pfr)与非活化(Pr) 2种状态(岳晶等, 2015).Pfr主要感知远红光变化, 而Pr负责感知红光变化.随着外界光环境的变化, 光敏色素会在Pfr与Pr之间相互转化(Holmes and Smith, 1975).在5种光敏色素中, phyB功能最强, 对其研究也最为深入.phyB在感知荫蔽下光信号变化过程中占主导地位(Franklin et al., 2003).phyB首先在细胞质中合成, 当R:FR增高时, phyB便从Pr形式转化为Pfr形式, 并进入细胞核中行使功能; 当R:FR降低时, 活化状态的Pfr将被运回细胞质中以Pr形式贮存(Yamaguchi et al., 1999).当PHYB基因突变后, 植物无法感知外界R:FR的变化, 与野生型相比phyB突变体下胚轴极显著伸长(Lorrain et al., 2008). ...
1 2013
... 光敏色素互作因子(Phytochrome Interacting Factors, PIFs)是处于光敏色素下游的一类与光敏色素发生直接相互作用的转录因子.植物体内共有7种不同构型的PIFs (PIF1, PIF3-8)能与PhyB互作, PIFs通过结合下游目的基因的启动子进而调控其表达(Leivar and Quail, 2011).在细胞核中, 活化态的Pfr既可对PIFs进行磷酸化, 导致其降解, 也可抑制PIFs与目的基因启动子的结合, 因此phyB通过PIFs间接调控植物基因的表达(Li et al., 2012b; Leivar and Monte, 2014).研究表明, PIFs家族中的PIF4、PIF5和PIF7参与植物荫蔽胁迫响应, 其中pif4/pif5双突变体与pif7突变体在荫蔽胁迫时下胚轴伸长均受到显著抑制(Lorrain et al., 2008; Li et al., 2012a).而在PIFs调控的下游基因中, 也包含了编码细胞壁修饰酶的ATHB4 (ARABIDOPSIS THALIANA HOMEOBOX PROTEIN4)以及合成植物激素的相关基因, 因此得以调控下胚轴的伸长(Steindler et al., 1999; Hornits- chek et al., 2012; Zhang et al., 2013). ...
1 2012
... 生长素是调控植物生长发育的关键激素之一, 对细胞分裂与扩张及种子休眠等具有显著调节作用(Wood- ward and Bartel, 2005; Liu et al., 2013).最新研究证明, 生长素在调节植物株型中起关键作用(de Wit et al., 2014).以拟南芥为例, 植物体内的生长素(3-吲哚乙酸, IAA)需先由色氨酸(Trp)经TAA1 (TRYPTO- PHAN AMINOTRANSFERASE OF ARABIDOPSIS1)合成3-吲哚丙酮酸盐(3-IPA), 3-IPA再经YUC (YUCCA)家族蛋白的作用生成IAA.在此过程中YUC家族蛋白是IAA合成的关键限速酶(Zhao, 2012; 帅海威等, 2016).在荫蔽胁迫下, 生长素合成四突变体yuc2/yuc5/yuc8/yuc9下胚轴长度无显著变化(Nozue et al., 2015); 生长素不敏感突变体axr1-12下胚轴伸长会受到抑制; 使用生长素转运抑制剂萘草胺(nap- talam, NPA)处理野生型幼苗, 同样能显著抑制下胚轴生长(Steindler et al., 1999; Tao et al., 2008).这些研究表明, 生长素可能是调控植物株型的关键激素(Morelli and Ruberti, 2000).进一步研究表明, 生长素转运突变体pin3-3在荫蔽胁迫时下胚轴伸长受到抑制, 并且发现在荫蔽胁迫下PIN3蛋白含量升高(Ke- uskamp et al., 2010).这进一步证实生长素的合成、运输及信号转导对于植物荫蔽胁迫响应非常重要. ...
1 2012
... 通过转录组分析, 研究人员发现乙烯和荫蔽胁迫在调控下胚轴伸长时具有类似的转录组分和功能调控因子(Das et al., 2016).乙烯信号通路中的转录因子EIN3被证明能结合在PIF3的启动子上, 进而促进PIF3的表达, 通过PIF3来调控下胚轴的长度(Zhong et al., 2012).然而在荫蔽胁迫下, 乙烯含量是如何升高的还有待进一步研究. ...
 ,
, 

{kind=link}
{kind=link}