Abstract: Histone regulator (HIRA) is a specific chaperone for histone H3.3 and plays an important role in the incorporation of histone H3.3 into chromatin. HIRA is indispensable in mammalian embryo development and DNA damage repair process, but we have few studies of the function of an HIRA homolog in plants. Here, we studied the function of Arabidopsis thaliana AtHIRA in somatic homologous recombination (HR) and meiotic homologous recombination. We used the somatic HR system and the meiotic homologous recombination system in wild type and the hira-1 mutant, a loss- of-function mutant of AtHIRA. Both intramolecular and intermolecular HR frequency was lower in hira-1 than the wild type under normal growth conditions and under bleomycin or UV-C treatment, with no significant difference in the frequency of meiotic recombination of microsporocytes between the wild type and hira-1 mutant under normal growth conditions. As well, under normal growth conditions or bleomycin treatment, loss-of-function of AtHIRA in Arabidopsis did not affect the DNA damage level. On qRT-PCR, the expression of RAD51 and RAD54, two DNA repair-related genes, was higher in hira-1 than the wild type. In addition, hira-1 had a salt-sensitive phenotype as compared with the wild type under salt stress. AtHIRA may play a role in somatic HR and the salinity response in Arabidopsis.
Key words:HIRA ; homologous recombination ; DNA damage repair ; meiotic ; salt stress
动植物细胞无时无刻不在累积外源和内源因素导致的DNA损伤, 因此DNA损伤修复机制对于动植物细胞的正常运转十分重要。在多种DNA损伤类型中, DNA双链断裂是最为严重的类型之一, 如果不及时修复, 对细胞的生存会产生极大的威胁(Xu and Price, 2011)。同源重组修复是体细胞DNA双链断裂损伤修复的重要通路之一(Heyer et al., 2010), 它依靠损伤位点的同源模板对DNA断裂处进行精确修复, 从而维持染色质的完整性和稳定性。此外, 在减数分裂过程中, 配子细胞通过同源重组过程进行染色体上同源片段的重组交换, 因此同源重组过程对配子细胞基因组多样性的形成也具有重要意义(Wijeratne and Ma, 2007)。目前, 对同源重组的发生过程已有较为清晰和深入的了解(Schuermann et al., 2005), 该过程中的许多重要因子已被发现并报道。例如, 在同源重组中发挥重要作用的RAD51蛋白可以与损伤处的DNA末端结合, 形成DNA-纤维结构, 从而完成后续的同源配对等过程(Krejci et al., 2012)。
HIR/HIRA (HISTONE REGULATOR A)是一类在进化上较为保守的H3.3特异性分子伴侣。HIRA蛋白最早在酿酒酵母(Saccharomyces cerevisiae)中被分离和鉴定, 当时发现HIRA对组蛋白基因的表达有负调节作用(Sherwood and Osley, 1991)。随后人们发现, 在许多真核生物中均存在HIRA的同源蛋白, 且以异源多聚物的形式存在(Amin et al., 2012)。HIRA蛋白的N端是一段串联的WD40结构域, C端有1个重要的功能结构域即HIRA。蛋白中间区段的结构域被命名为HIRA-B, 该结构域对于HIRA蛋白与其它蛋白(如ASF1)的结合有重要意义(Tang et al., 2006)。目前, 在动物中关于HIRA的功能研究较为深入。例如, 实验表明, HIRA在果蝇(Drosophila melanogaster)精细胞核解凝过程中对组蛋白掺入染色质有重要意义(Loppin et al., 2005)。对非洲爪蟾(Xenopus laevis)的研究 表明, 下调HIRA的表达会引发原肠胚的生理缺陷(Szenker et al., 2012)。在小鼠(Mus musculus)中的研究表明, 敲除HIRA后会导致胚胎致死现象(Rob- erts et al., 2002)。
相比于动物, 在植物中HIRA功能的研究尚不够深入。以拟南芥(Arabidopsis thaliana)为例, AtHIRA在拟南芥中为单拷贝基因, 其序列与高等动物中HIRA基因有较高的同源性。之前, 关于拟南芥中AtHIRA基因功能缺失突变体hira-1的研究结果表明, AtHIRA功能缺失的拟南芥仅表现出短根和叶片下垂等表型, 而植物整体的生长发育及育性等方面均没有明显缺陷(Ingouff et al., 2010; Nie et al., 2014)。这与大部分动物中HIRA缺失会导致明显的生理缺陷存在较大差异, 表明在动植物中HIRA发挥的功能可能有所不同。
1.2 实验方法1.2.1 植物组织染色 研究采用的GUS基因报告系统为已报道的1445和IC9C (Fritsch et al., 2004; Molinier et al., 2004; Sch- uermann et al., 2009)。GUS基因报告株系与hira-1突变体杂交后, 筛选后代获得纯合株系。随后参照相关报道, 对bleomycin和UV-C处理前后的拟南芥进行组织化学染色分析(Gao et al., 2012)。将拟南芥在37°C黑暗条件下放入GUS染液染色2小时, 依次用50%、75%和100%乙醇脱色至植物材料褪绿, 并保存于100%的乙醇中。在解剖显微镜(M205 FA, Leica, 德国)下观察并统计GUS点的数目。不同处理材料的统计数目均在60株以上, 3次重复。根据统计结果计算平均值和标准差(SD), 用Student’s t test计算差异显著性。 bleomycin处理: 将培养14天的拟南芥幼苗在含有10 µmol·L-1 bleomycin的MS液体培养基中培养1天, 随后转移至正常MS液体培养基中培养3天。紫外UV-C处理: 将培养14天的拟南芥幼苗置于4 000 J·m-2的UV-C紫外光下照射, 之后立刻转移回正常培养条件。 1.2.2 彗星实验 将培养14天的拟南芥幼苗置于含有2 µmol·L-1 bleomycin的MS液体培养基中, 处理6小时后收集, 参照Zhu等(2006)的方法进行彗星实验。制备铺有1%正常熔点琼脂糖溶液的载玻片, 随后切割拟南芥幼苗制备细胞核悬浮液, 并与1%低熔点琼脂糖混合, 将混合液铺在预制的载玻片上。经高盐缓冲液(2.5 mol·L-1 NaCl, 10 mmol·L-1 Tris-HCl, pH7.5, 100 mmol·L-1 EDTA)裂解后, 将载玻片放入电泳槽中, 在40V电压下电泳, 之后经过中和、干燥和染色, 将载玻片置于Imager A2显微镜(Zeiss, Jena, 德国)下检测细胞核拖尾长度。使用CometScore软件(http://autocomet. com)对图像进行定量分析和误差分析。根据结果计算出平均值和标准差, 并利用Student’s t test计算差异显著性。 1.2.3 荧光定量RT-PCR 使用TRIzol试剂盒(Invitrogen, Carlsbad, CA, 美国)提取植物材料中的总RNA, 随后利用Improm-II反转录酶(Promega, Madison, WI, 美国)体系进行反转录实验。选取ACTIN2作为qRT-PCR的内参基因, 实验中检测的基因为GUS、RAD51、RAD54、PARP1、PARP2、BRCA1和ATM。所用引物序列见表1。 1.2.4 荧光四分体分析 本研究使用在拟南芥突变体qrt1-2的2号(I2)及5号染色体(I5)上插入了dsRED2、eYFP和CFP荧光基因的同源重组报告株系(Berchowitz et al., 2007), 通过遗传杂交将hira-1的突变位点导入I2和I5荧光报告系统, 并在F4群体中筛选荧光信号(dsRED2、eYFP和CFP)杂合、而hira-1突变体纯合的株系, 使用Imager A2显微镜(Zeiss, Jena, 德国)观察并统计四分体荧光信号。 表1 Table 1 表1 表1 本研究所用引物 Table 1 Primers used in this study
Primer name
Primer sequence (5′-3′)
ACTIN2-F
GGCGATGAAGCTCAATCCAAA
ACTIN2-R
GGTCACGACCAGCAAGATCAAG
GUS-F
AAGTGGATTGATGTGATATCTC
GUS-R
TTCGCGCTGATACCAGACG
ATM-F
TGCAGCTGCGTCTCTGCATGA
ATM-R
CTTCATGCCGCCCTTGGGCA
BRCA1-F
TGCTCAGGGCTCACAGTTGAAGA
BRCA1-R
TGCAGGCTCCGTTTTCATTGATTG
PARP1-F
TGCTCGCGCGAACTCACTTCT
PARP1-R
AGCCTCTCCACCAGAACGGCT
PARP2-F
AGCCTGAAGGCCCGGGTAACA
PARP2-R
GCTGTCTCAGTTTTGGCTGCCG
RAD51-F
CGCCATTTCCCTCCACTCTCAAGC
RAD51-R
ACCTGCTGCCTGAAGCTGTTCG
RAD54-F
TGAGAGACAGGTGGGCACTCC
RAD54-R
ACGTCACCTCGTCACCTGCTGA
表1 本研究所用引物 Table 1 Primers used in this study
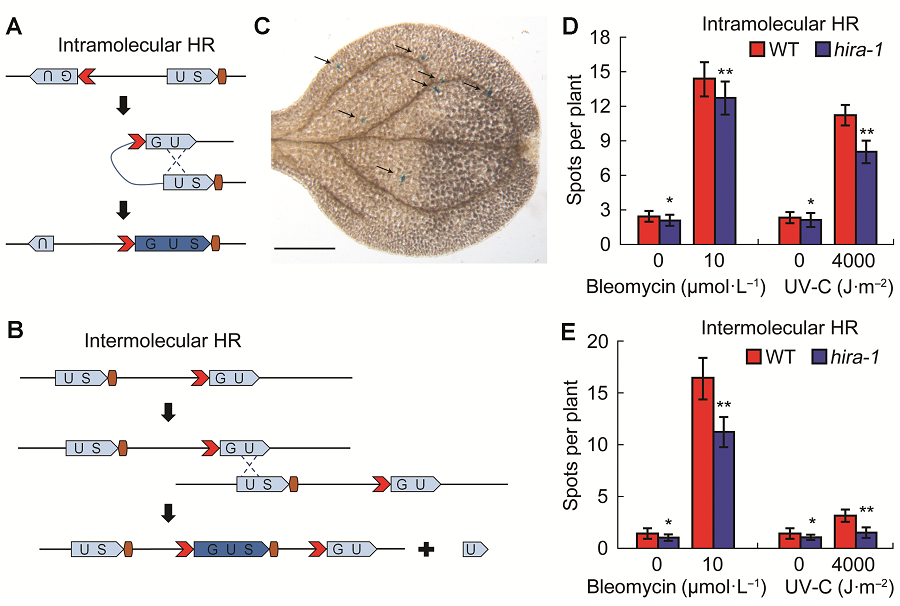
2 结果与讨论2.1 AtHIRA基因功能失活对体细胞同源重组频率的影响为分析AtHIRA在体细胞同源重组中的功能, 我们分别选择分子内同源重组(intramolecular homologous recombination)报告株系1445以及分子间同源重组(intermolecular homologous recombination)报告株系IC9C。每种报告株系中均含有两段不完整且分离的GUS (β-glucuronidase)基因。这两段基因中存在着一段相同的序列(Gherbi et al., 2001; Schuermann et al., 2009)。当体细胞同源重组事件发生后, 这两段基因能够重新组合并且形成一个有功能的GUS基因(Schuermann et al., 2009), 如图1A和B所示。当GUS基因正常表达后, 利用组织化学染色法可在植物组织中观察到蓝色的染色点, 如图1C所示。 分子内同源重组株系1445的统计结果表明, 在正常培养条件下, hira-1突变体的同源重组频率低于野生型(图1D)。基因毒试剂bleomycin或UV-C处理后, hira-1突变体和野生型的同源重组频率均明显升高, 但相比野生型, hira-1突变体体细胞的同源重组水平明显下降(图1D)。分子间同源重组株系IC9C的统计结果表明, 无论在正常培养条件下还是bleomycin处理或UV-C处理后, hira-1突变体的分子间重组频率也均低于野生型(图1E)。上述结果表明, hira-1突变体的体细胞同源重组频率低于野生型, 且DNA损伤处理后尤为明显, 可见AtHIRA在体细胞同源重组中发挥了一定作用。 图1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_1.png<b>图1</b> 拟南芥<i>hira-1</i>突变体的体细胞同源重组水平低于野生型<br/>(A) 1445株系中的分子内同源重组事件示意, 在经过1次同源重组事件后, <i>GUS</i>基因的两个分离片段可以重新组成1个有功能的<i>GUS</i>基因; (B) IC9C株系中的分子间同源重组事件示意, 不同分子间的片段经过同源重组事件重新形成有功能的<i>GUS</i>基因; (C) 组织化学染色后出现在拟南芥叶片中的每个箭头指示的蓝点代<xref ref-type="table" rid="T1-1674-3466-53-1-42">表1</xref>个有功能的<i>GUS</i>基因, 表明发生了1次同源重组事件(Bar=500 μm); (D) 野生型和<i>hira-1</i>突变体的分子内同源重组水平比较; (E) 野生型和<i>hira-1</i>突变体的分子间同源重组水平比较。WT: 野生型; HR: 同源重组。* <i>P</i><0.05; ** <i>P</i><0.01<br/><b>Figure 1</b> <i> hira1 </i>mutant shows reduced homologus recombination frequency compared with Col-0 of Arabidopsis<br/>(A) Scheme of homologus recombination (HR) event in intramolecular line 1445. The two fragments of <i>GUS</i> gene can recombine to form a function<i> GUS</i> gene after a HR event; (B) Scheme of HR event in intermolecular line IC9C. The recombination of separated <i>GUS</i> fragments require intermolecular interaction to restore a functional <i>GUS</i> gene after a HR event; (C) Arabidopsis leaf with arrow labeled blue spots/sectors represent a functional <i>GUS</i> gene, which indicate an independent HR event (Bar=500 μm); (D) Comparison of intramolecular recombination frequency between Col-0 and <i>hira-1</i>; (E) Comparison of intermolecular recombination frequency between Col-0 and <i>hira-1. </i>WT: Wild type; HR: Homologus recombination. * <i>P</i><0.05; ** <i>P</i><0.01 Figure 1https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_1.png<b>图1</b> 拟南芥<i>hira-1</i>突变体的体细胞同源重组水平低于野生型<br/>(A) 1445株系中的分子内同源重组事件示意, 在经过1次同源重组事件后, <i>GUS</i>基因的两个分离片段可以重新组成1个有功能的<i>GUS</i>基因; (B) IC9C株系中的分子间同源重组事件示意, 不同分子间的片段经过同源重组事件重新形成有功能的<i>GUS</i>基因; (C) 组织化学染色后出现在拟南芥叶片中的每个箭头指示的蓝点代<xref ref-type="table" rid="T1-1674-3466-53-1-42">表1</xref>个有功能的<i>GUS</i>基因, 表明发生了1次同源重组事件(Bar=500 μm); (D) 野生型和<i>hira-1</i>突变体的分子内同源重组水平比较; (E) 野生型和<i>hira-1</i>突变体的分子间同源重组水平比较。WT: 野生型; HR: 同源重组。* <i>P</i><0.05; ** <i>P</i><0.01<br/><b>Figure 1</b> <i> hira1 </i>mutant shows reduced homologus recombination frequency compared with Col-0 of Arabidopsis<br/>(A) Scheme of homologus recombination (HR) event in intramolecular line 1445. The two fragments of <i>GUS</i> gene can recombine to form a function<i> GUS</i> gene after a HR event; (B) Scheme of HR event in intermolecular line IC9C. The recombination of separated <i>GUS</i> fragments require intermolecular interaction to restore a functional <i>GUS</i> gene after a HR event; (C) Arabidopsis leaf with arrow labeled blue spots/sectors represent a functional <i>GUS</i> gene, which indicate an independent HR event (Bar=500 μm); (D) Comparison of intramolecular recombination frequency between Col-0 and <i>hira-1</i>; (E) Comparison of intermolecular recombination frequency between Col-0 and <i>hira-1. </i>WT: Wild type; HR: Homologus recombination. * <i>P</i><0.05; ** <i>P</i><0.01
图1 拟南芥hira-1突变体的体细胞同源重组水平低于野生型 (A) 1445株系中的分子内同源重组事件示意, 在经过1次同源重组事件后, GUS基因的两个分离片段可以重新组成1个有功能的GUS基因; (B) IC9C株系中的分子间同源重组事件示意, 不同分子间的片段经过同源重组事件重新形成有功能的GUS基因; (C) 组织化学染色后出现在拟南芥叶片中的每个箭头指示的蓝点代表1个有功能的GUS基因, 表明发生了1次同源重组事件(Bar=500 μm); (D) 野生型和hira-1突变体的分子内同源重组水平比较; (E) 野生型和hira-1突变体的分子间同源重组水平比较。WT: 野生型; HR: 同源重组。* P<0.05; ** P<0.01 Figure 1 hira1 mutant shows reduced homologus recombination frequency compared with Col-0 of Arabidopsis (A) Scheme of homologus recombination (HR) event in intramolecular line 1445. The two fragments of GUS gene can recombine to form a function GUS gene after a HR event; (B) Scheme of HR event in intermolecular line IC9C. The recombination of separated GUS fragments require intermolecular interaction to restore a functional GUS gene after a HR event; (C) Arabidopsis leaf with arrow labeled blue spots/sectors represent a functional GUS gene, which indicate an independent HR event (Bar=500 μm); (D) Comparison of intramolecular recombination frequency between Col-0 and hira-1; (E) Comparison of intermolecular recombination frequency between Col-0 and hira-1. WT: Wild type; HR: Homologus recombination. * P<0.05; ** P<0.01
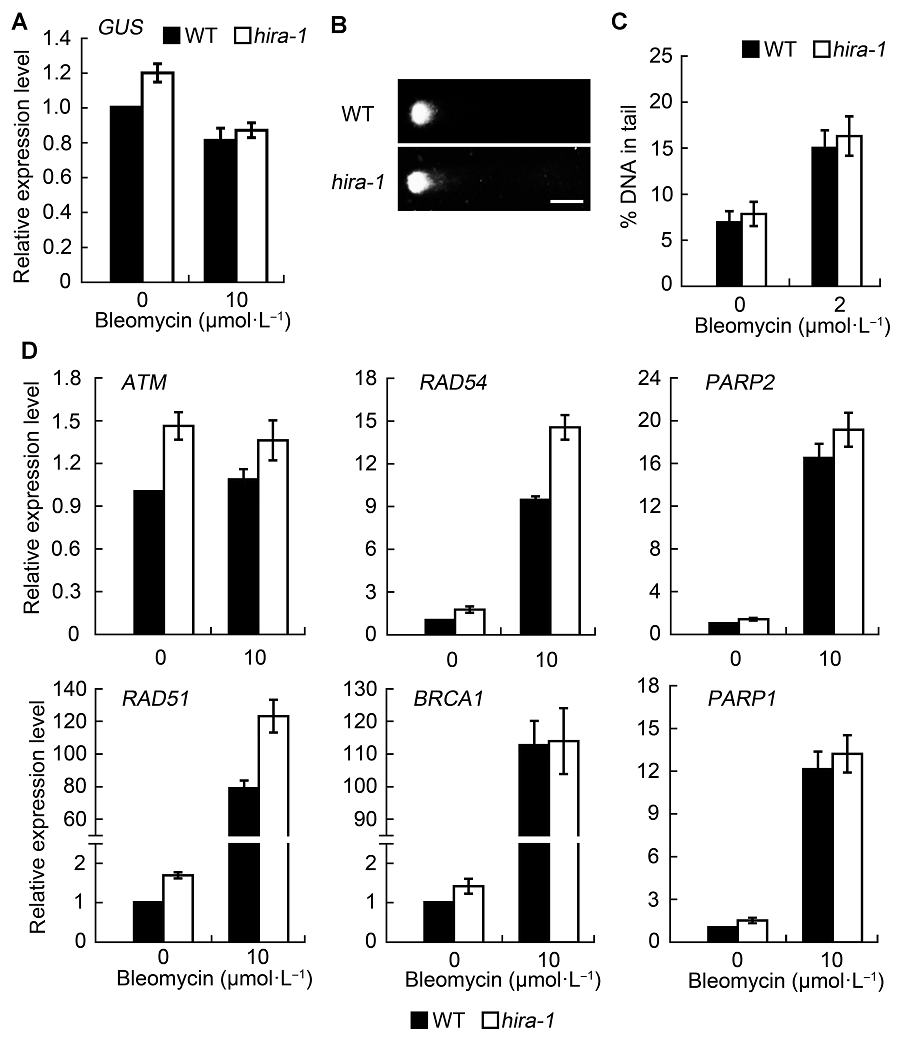
2.2 AtHIRA基因功能失活对DNA损伤程度的影响为在细胞层面探明hira-1突变体体细胞同源重组水平下降的原因, 我们首先检测了GUS基因在不同背景下的表达水平, 发现在hira-1突变体及野生型植株中, GUS基因的表达水平无明显差异(图2A)。随后我们通过彗星实验分别对正常生长条件下及bleomycin处理后的幼苗中的DNA损伤程度进行检测(图2B)。结果表明, 处理前后野生型和hira-1突变体的DNA损伤水平均比较接近(图2C)。同时, 我们检测了一些同源重组通路中重要基因的表达水平, 包括RAD51、RAD54、PARP1、PARP2、BRCA1及ATM。结果显示, bleo- mycin处理导致这些基因普遍上调表达。经bleomycin处理后, 除RAD51和RAD54在hira-1突变体中表达水平的上调明显高于野生型外, 其余基因在野生型和hira-1突变体中的表达水平并无明显差异(图2D)。上述结果表明, 经相同浓度的bleomycin处理后, 野生型与hira-1突变体中的DNA损伤程度无明显差异。与体细胞同源重组水平相反, 经bleomycin处理后, hira-1突变体的部分同源重组通路相关基因表达水平明显高于野生型。此结果暗示, AtHIRA可能还通过其它途径影响体细胞的同源重组水平。 图2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_2.png<b>图2</b> <i>AtHIRA</i>功能缺失对DNA损伤水平的影响及DNA损伤修复相关基因的表达水平检测(平均值±标准差)<br/>(A) 在正常条件和bleomycin处理下, 拟南芥野生型和<i>hira-1</i>突变体中<i>GUS</i>基因的表达水平; (B) 损伤处理后, 野生型与<i>hira-1</i>突变体中细胞核彗星示意图(Bar=10 μm); (C) 野生型和<i>hira-1</i>突变体的细胞核彗尾中DNA百分比统计。统计结果均从超过100个细胞核中得出; (D) 实时荧光定量PCR检测野生型和<i>hira-1</i>突变体的DNA损伤修复相关基因的表达水平。实验经3次生物学重复。<br/><b>Figure 2</b> Effect of <i>AtHIRA</i> mutation on DNA damage level and expression level of DNA repair genes (means±SD)<br/>(A) Expression level of <i>GUS</i> gene in wild type and <i>hira-1 </i>mutant of Arabidopsis under normal conditions and bleomycin treatment; (B) Representative comet images of wild-type and <i>hira-1</i> nuclei after bleomycin treatment (Bar=10 μm); (C) The average percentage of DNA in comet tails of wild type and <i>hira-1</i> mutant. More than 100 individual nuclei were recorded and calculated; (D) Relative expression level of DNA repair genes by RT-qPCR between wild type and <i>hira-1.</i> Three biological repeats were analyzed. Figure 2https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_2.png<b>图2</b> <i>AtHIRA</i>功能缺失对DNA损伤水平的影响及DNA损伤修复相关基因的表达水平检测(平均值±标准差)<br/>(A) 在正常条件和bleomycin处理下, 拟南芥野生型和<i>hira-1</i>突变体中<i>GUS</i>基因的表达水平; (B) 损伤处理后, 野生型与<i>hira-1</i>突变体中细胞核彗星示意图(Bar=10 μm); (C) 野生型和<i>hira-1</i>突变体的细胞核彗尾中DNA百分比统计。统计结果均从超过100个细胞核中得出; (D) 实时荧光定量PCR检测野生型和<i>hira-1</i>突变体的DNA损伤修复相关基因的表达水平。实验经3次生物学重复。<br/><b>Figure 2</b> Effect of <i>AtHIRA</i> mutation on DNA damage level and expression level of DNA repair genes (means±SD)<br/>(A) Expression level of <i>GUS</i> gene in wild type and <i>hira-1 </i>mutant of Arabidopsis under normal conditions and bleomycin treatment; (B) Representative comet images of wild-type and <i>hira-1</i> nuclei after bleomycin treatment (Bar=10 μm); (C) The average percentage of DNA in comet tails of wild type and <i>hira-1</i> mutant. More than 100 individual nuclei were recorded and calculated; (D) Relative expression level of DNA repair genes by RT-qPCR between wild type and <i>hira-1.</i> Three biological repeats were analyzed.
图2 AtHIRA功能缺失对DNA损伤水平的影响及DNA损伤修复相关基因的表达水平检测(平均值±标准差) (A) 在正常条件和bleomycin处理下, 拟南芥野生型和hira-1突变体中GUS基因的表达水平; (B) 损伤处理后, 野生型与hira-1突变体中细胞核彗星示意图(Bar=10 μm); (C) 野生型和hira-1突变体的细胞核彗尾中DNA百分比统计。统计结果均从超过100个细胞核中得出; (D) 实时荧光定量PCR检测野生型和hira-1突变体的DNA损伤修复相关基因的表达水平。实验经3次生物学重复。 Figure 2 Effect of AtHIRA mutation on DNA damage level and expression level of DNA repair genes (means±SD) (A) Expression level of GUS gene in wild type and hira-1 mutant of Arabidopsis under normal conditions and bleomycin treatment; (B) Representative comet images of wild-type and hira-1 nuclei after bleomycin treatment (Bar=10 μm); (C) The average percentage of DNA in comet tails of wild type and hira-1 mutant. More than 100 individual nuclei were recorded and calculated; (D) Relative expression level of DNA repair genes by RT-qPCR between wild type and hira-1. Three biological repeats were analyzed.
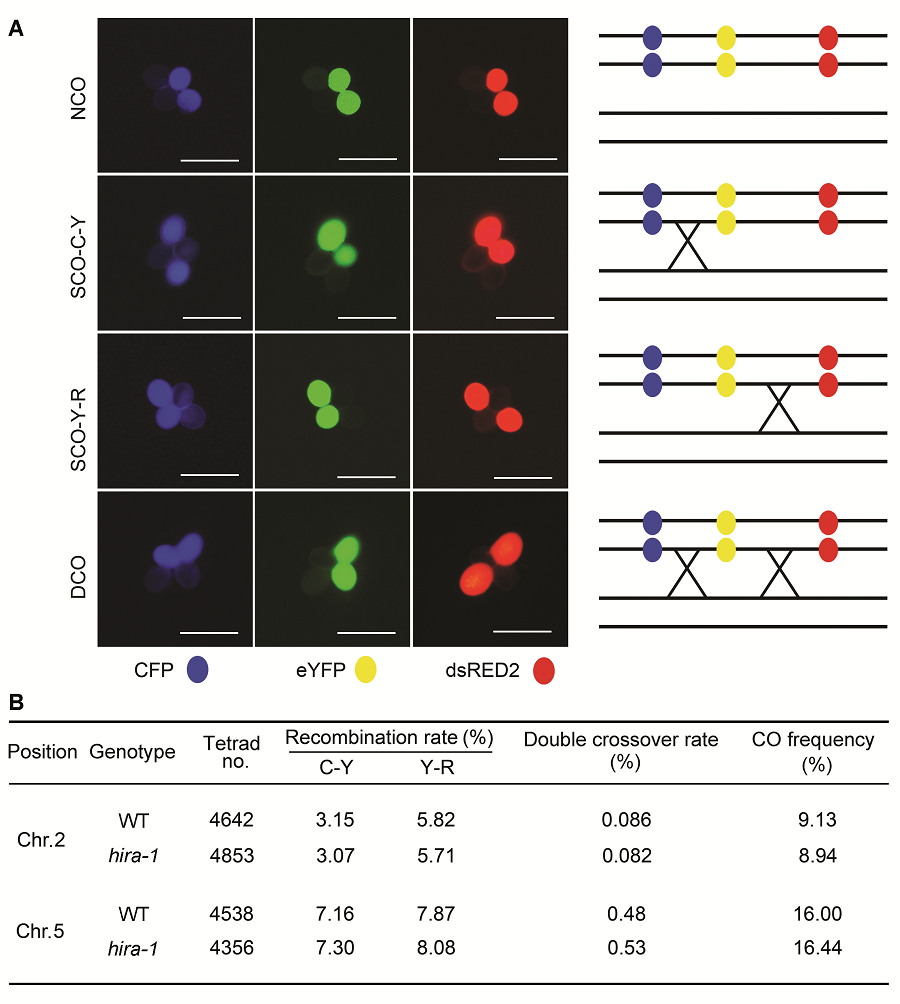
2.3 AtHIRA基因功能失活对减数分裂同源重组频率的影响利用花粉四分体荧光标记系统(Francis et al., 2007), 我们还检测了AtHIRA对减数分裂同源重组的影响。花粉四分体荧光系统是在花粉四分体黏连不分离(即qrt1-2纯合突变背景)的条件下, 观察花粉四分体中荧光蛋白交换情况的可视化实验手段。通过对大量四分体进行观察, 分别统计不交换(NCO)、单交换(SCO- C-Y和SCO-Y-R)以及双交换(DCO)的四分体数目(图3A)。计算交换频率(交换频率=((SCO-C-Y+SCO-Y-R+ 2DCO)/total tetrads)×100%)后发现, 在2号染色体上, 野生型和hira-1突变体的花粉交换频率分别为9.13%和8.94% (图3B); 在5号染色体上, 二者的花粉交换频率分别为16.00%和16.44% (图3B)。结果表明, 野生型和hira-1突变体间的减数分裂同源重组频率没有明显差异, AtHIRA功能失活不能显著改变减数分裂过程中染色体的交换频率, 这一结果表明AtHIRA功能失活对减数分裂同源重组过程的影响不明显。 图3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_3.png<b>图3</b> 利用荧光标记花粉四分体系统比较拟南芥野生型(WT)和<i>hira-1</i>突变体的减数分裂同源重组频率<br/>(A) 荧光四分体类型示意图: 不交换(NCO)、2种单交换(SCO-C-Y和SCO-Y-R)和1种双交换(DCO)。右侧是每种荧光类型对应的交换示意图(Bars=10 μm); (B) 在野生型和<i>hira-1</i>突变体中观察到的各类型荧光四分体数目。<br/><b>Figure 3</b> Comparison of meiotic recombination frequency between wild type (WT) and <i>hira-1</i> mutant of Arabidopsis using the fluorescent tagged line tetrad analysis system<br/>(A) Examples of tetrad fluorescent patterns including no cross over (NCO), two types of single cross overs (SCO-C-Y and SCO- Y-R) and one type of double cross over (DCO). The schematic representation of corresponding CO events is shown at right of each tetrad class (Bars=10 μm); (B) Number of each tetrad fluorescent patterns observed in wild type and <i>hira-1.</i> Figure 3https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_3.png<b>图3</b> 利用荧光标记花粉四分体系统比较拟南芥野生型(WT)和<i>hira-1</i>突变体的减数分裂同源重组频率<br/>(A) 荧光四分体类型示意图: 不交换(NCO)、2种单交换(SCO-C-Y和SCO-Y-R)和1种双交换(DCO)。右侧是每种荧光类型对应的交换示意图(Bars=10 μm); (B) 在野生型和<i>hira-1</i>突变体中观察到的各类型荧光四分体数目。<br/><b>Figure 3</b> Comparison of meiotic recombination frequency between wild type (WT) and <i>hira-1</i> mutant of Arabidopsis using the fluorescent tagged line tetrad analysis system<br/>(A) Examples of tetrad fluorescent patterns including no cross over (NCO), two types of single cross overs (SCO-C-Y and SCO- Y-R) and one type of double cross over (DCO). The schematic representation of corresponding CO events is shown at right of each tetrad class (Bars=10 μm); (B) Number of each tetrad fluorescent patterns observed in wild type and <i>hira-1.</i>
图3 利用荧光标记花粉四分体系统比较拟南芥野生型(WT)和hira-1突变体的减数分裂同源重组频率 (A) 荧光四分体类型示意图: 不交换(NCO)、2种单交换(SCO-C-Y和SCO-Y-R)和1种双交换(DCO)。右侧是每种荧光类型对应的交换示意图(Bars=10 μm); (B) 在野生型和hira-1突变体中观察到的各类型荧光四分体数目。 Figure 3 Comparison of meiotic recombination frequency between wild type (WT) and hira-1 mutant of Arabidopsis using the fluorescent tagged line tetrad analysis system (A) Examples of tetrad fluorescent patterns including no cross over (NCO), two types of single cross overs (SCO-C-Y and SCO- Y-R) and one type of double cross over (DCO). The schematic representation of corresponding CO events is shown at right of each tetrad class (Bars=10 μm); (B) Number of each tetrad fluorescent patterns observed in wild type and hira-1.
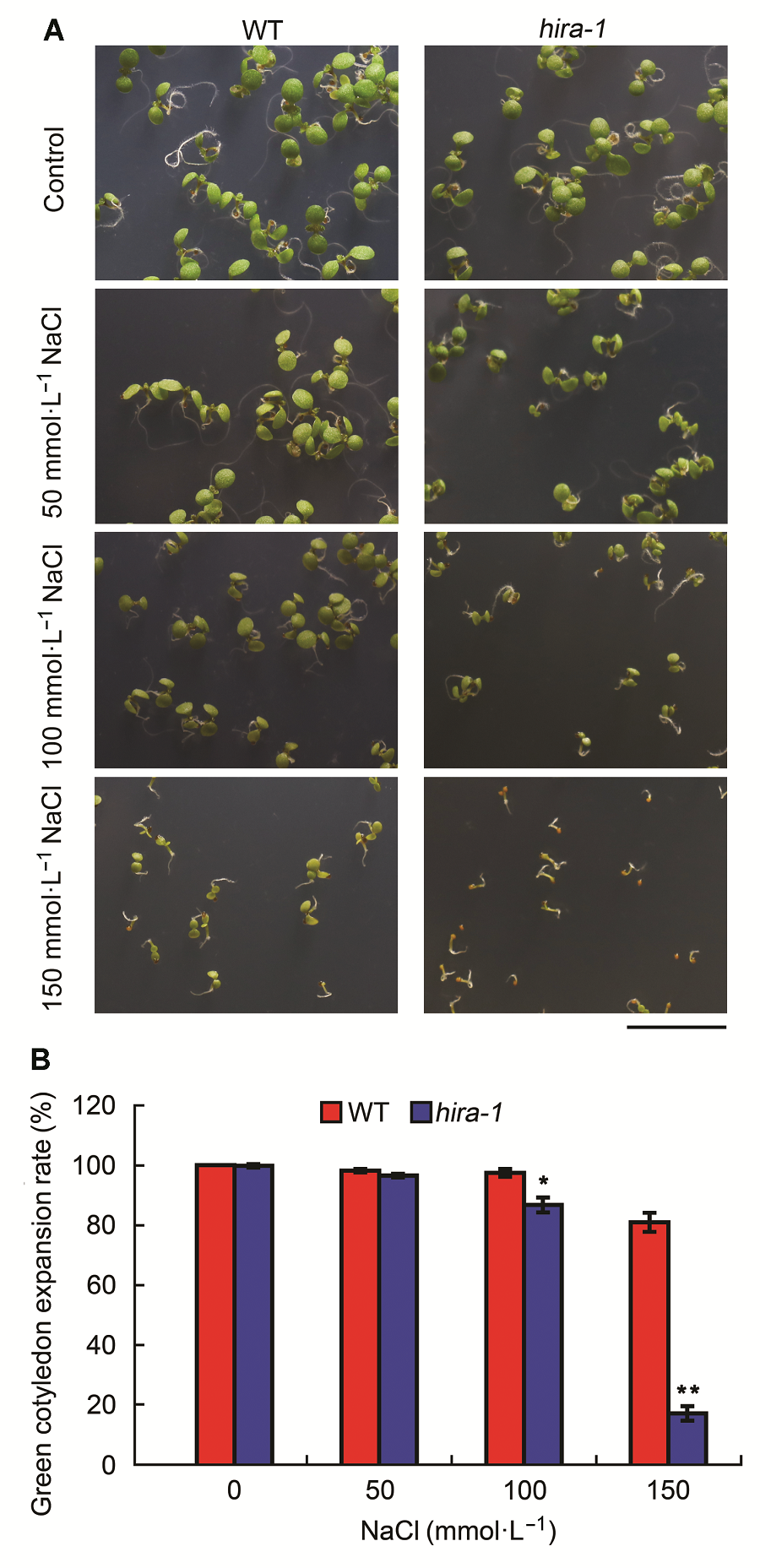
2.4 AtHIRA基因功能失活对盐胁迫的响应为了研究AtHIRA基因在拟南芥盐胁迫响应过程中的作用, 我们将野生型和hira-1突变体种子播种在添加了50、100和150 mmol·L-1 NaCl的MS固体培养基上。植物正常培养7天后进行表型对比观察和子叶展开率统计。结果表明, 在NaCl浓度为100和150 mmol·L-1时, hira-1突变体幼苗受到的生长抑制比野生型更加明显(图4A)。同时, 随着盐浓度的升高(超过100 mmol·L-1 NaCl), hira-1的子叶展开率显著低于野生型, 如图4B所示。上述实验结果表明, AtHIRA基因功能缺失使得植株对高浓度盐胁迫更加敏感。 图4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_4.png<b>图4</b> 拟南芥<i>hira-1</i>突变体对盐胁迫敏感(平均值±标准差)<br/>(A) 在含0、50、100和500 mmol·L<sup>-1</sup> NaCl培养基上生长7天的野生型(WT)与<i>hira-1</i>突变体幼苗的表型(Bar=1 cm); (B) 生长7天的野生型与<i>hira-1</i>突变体幼苗绿色子叶展开率。每种基因型统计数目超过100株。实验经3次生物学重复。<br/><b>Figure 4</b> Arabidopsis mutant<i> hira-1 </i>exhibited hypersensitivi- ty to salt stress (means±SD)<br/>(A) Phenotypes of 7-day-old seedlings of wild type (WT) and <i>hira-1 </i>grown on media containing 0, 50, 100, and 150 mmol·L<sup>-1</sup> NaCl (Bar=1 cm); (B) Comparison of green cotyledon expansion rates of 7-day-old seedlings between wild type and <i>hira-1</i>. Over 100 seeds each genotype were recorded. Three biological repeats were analyzed. Figure 4https://www.chinbullbotany.com/article/2018/1674-3466/1674-3466-53-1-42/img_4.png<b>图4</b> 拟南芥<i>hira-1</i>突变体对盐胁迫敏感(平均值±标准差)<br/>(A) 在含0、50、100和500 mmol·L<sup>-1</sup> NaCl培养基上生长7天的野生型(WT)与<i>hira-1</i>突变体幼苗的表型(Bar=1 cm); (B) 生长7天的野生型与<i>hira-1</i>突变体幼苗绿色子叶展开率。每种基因型统计数目超过100株。实验经3次生物学重复。<br/><b>Figure 4</b> Arabidopsis mutant<i> hira-1 </i>exhibited hypersensitivi- ty to salt stress (means±SD)<br/>(A) Phenotypes of 7-day-old seedlings of wild type (WT) and <i>hira-1 </i>grown on media containing 0, 50, 100, and 150 mmol·L<sup>-1</sup> NaCl (Bar=1 cm); (B) Comparison of green cotyledon expansion rates of 7-day-old seedlings between wild type and <i>hira-1</i>. Over 100 seeds each genotype were recorded. Three biological repeats were analyzed.
图4 拟南芥hira-1突变体对盐胁迫敏感(平均值±标准差) (A) 在含0、50、100和500 mmol·L-1 NaCl培养基上生长7天的野生型(WT)与hira-1突变体幼苗的表型(Bar=1 cm); (B) 生长7天的野生型与hira-1突变体幼苗绿色子叶展开率。每种基因型统计数目超过100株。实验经3次生物学重复。 Figure 4 Arabidopsis mutant hira-1 exhibited hypersensitivi- ty to salt stress (means±SD) (A) Phenotypes of 7-day-old seedlings of wild type (WT) and hira-1 grown on media containing 0, 50, 100, and 150 mmol·L-1 NaCl (Bar=1 cm); (B) Comparison of green cotyledon expansion rates of 7-day-old seedlings between wild type and hira-1. Over 100 seeds each genotype were recorded. Three biological repeats were analyzed.
AminAD, VishnoiN, ProchassonP (2012). A global requirement for the HIR complex in the assembly of chromatin.Biochim Biophys Acta 1819, 264-276. [本文引用: 1]
[3]
AndersenSL, SekelskyJ (2010). Meiotic versus mitotic recombination: two different routes for double-strand b- reak repair: the different functions of meiotic versus mi- totic DSB repair are reflected in different pathway usage and different outcomes. Bioessays 32, 1058-1066. [本文引用: 1]
[4]
FrancisKE, LamSY, HarrisonBD, BeyAL, BerchowitzLE, CopenhaverGP (2007). Pollen tetrad-based visual assay for meiotic recombination in Arabidopsis. Proc Natl Acad Sci USA 104, 3913-3918. [本文引用: 2]
[5]
FritschO, BenvenutoG, BowlerC, MolinierJ, HohnB (2004). The INO80 protein controls homologous recombination in Arabidopsis thaliana. Mol Cell 16, 479-485. [本文引用: 1]
[6]
GaoJ, ZhuY, ZhouWB, MolinierJ, DongAW, ShenWH (2012). NAP1 family histone chaperones are required for somatic homologous recombination in Arabidopsis.Plant Cell 24, 1437-1447. [本文引用: 1]
[7]
GherbiH, GallegoME, JalutN, LuchtJM, HohnB, WhiteCI (2001). Homologous recombination in planta is stimulated in the absence of Rad50.EMBO Rep 2, 287-291. [本文引用: 1]
[8]
GirautL, FalqueM, DrouaudJ, PereiraL, MartinOC, MézardC (2011). Genome-wide crossover distribution in Arabidopsis thaliana meiosis reveals sex-specific patterns along chromosomes. PLoS Genet 7, e1002354. [本文引用: 1]
[9]
HeyerWD, EhmsenKT, LiuJ (2010). Regulation of homologous recombination in eukaryotes.Annu Rev Genet 44, 113-139. [本文引用: 1]
[10]
IngouffM, RademacherS, HolecS, ŠoljićL, XinN, ReadshawA, FooSH, LahouzeB, SprunckS, BergerF (2010). Zygotic resetting of the HISTONE 3 variant re- pertoire participates in epigenetic reprogramming in Arabi- dopsis.Curr Biol 20, 2137-2143. [本文引用: 1]
[11]
KrejciL, AltmannovaV, SpirekM, ZhaoXL (2012). Homologous recombination and its regulation.Nucleic Acids Res 40, 5795-5818. [本文引用: 1]
[12]
LoppinB, BonnefoyE, AnselmeC, LaurenconA, KarrTL, CoubleP (2005). The histone H3.3 chaperone HIRA is essential for chromatin assembly in the male pronucleus.Nature 437, 1386-1390. [本文引用: 1]
[13]
Melamed-BessudoC, LevyAA (2012). Deficiency in DNA methylation increases meiotic crossover rates in euchro- matic but not in heterochromatic regions in Arabidopsis.Proc Natl Acad Sci USA 109, E981-E988. [本文引用: 1]
[14]
MolinierJ, RiesG, BonhoefferS, HohnB (2004). Interchromatid and interhomolog recombination in Arabidopsis thaliana. Plant Cell 16, 342-352. [本文引用: 1]
[15]
NieX, WangHF, LiJ, HolecS, BergerF (2014). The HIRA complex that deposits the histone H3.3 is conserved in Arabidopsis and facilitates transcriptional dynamics.Biol Open 3, 794-802. [本文引用: 1]
[16]
OkadaT, EndoM, SinghMB, BhallaPL (2005). Analysis of the histoneH3 gene family in Arabidopsis and identification of the male-gamete-specific variant AtMGH3. Plant J 44, 557-568. [本文引用: 1]
[17]
OteroS, DesvoyesB, GutierrezC (2014). Histone H3 dynamics in plant cell cycle and development.Cytogenet Genome Res 143, 114-124. [本文引用: 1]
[18]
RobertsC, SutherlandHF, FarmerH, KimberW, HalfordS, CareyA, BrickmanJM, Wynshaw-BorisA, ScamblerPJ (2002). Targeted mutagenesis of theHira gene results in gastrulation defects and patterning abnormalities of mesoendodermal derivatives prior to early embryo- nic lethality. Mol Cell Biol 22, 2318-2328. [本文引用: 1]
[19]
SchuermannD, FritschO, LuchtJM, HohnB (2009). Replication stress leads to genome instabilities in Arabidopsis DNA polymerase δ mutants.Plant Cell 21, 2700-2714. [本文引用: 3]
[20]
SchuermannD, MolinierJ, FritschO, HohnB (2005). The dual nature of homologous recombination in plants.Tren- ds Genet 21, 172-181. [本文引用: 1]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}